Embed Size (px)
Citation preview

Plant Physiol. (1972) 50, 161-165
Biochemical and Biophysical Characteristics of a PhotosyntheticMutant of Euglena gracilis Blocked in Photosystem 1112
Received for publication September 28, 1971
FAYE D. SCHWELITZ,3 R. A. DILLEY, AND F. L. CRANECharles F. Kettering Research Laboratory, Yellow Springs, Ohio 45387 and Department of Biological Sciences,Purdue University, West Lafayette, Indiana 47907
ABSTRACT
A photosynthetic mutant of Euglena gracilis, Z strain, thoughtto be blocked in the electron transport chain between the twophotosystems and to have a missing or nonfunctional primaryacceptor for photosystem II, was further studied and charac-terized. The data from low temperature fluorescence spectra,delayed light emission, and electron paramagnetic resonancesupport the previous work.The mutant was shown to lack plastoquinone A and possi-
bly cytochrome b5m but to possess plastoquinone B and a highercomplement of carotenoids, especially xanthophylls, than thewild type.The results are consistent with the postulated role of plasto-
quinone as the primary electron acceptor for photosystem IIand as responsible for the electron paramagnetic resonancesignal II. The abundance of xanthophylls in the mutant canbe explained by the protective role of carotenoids against pho-tosensitized reactions.
Russell and Lyman (23) isolated the photosynthetic mutantP4 of Euglena gracilis, Z strain, and showed that it was incapa-ble of carrying out the Hill reaction. Further studies by Russellet al. (24) revealed that the mutation blocked electron transportbetween photosystem I and photosystem II in the mutant. TheP4 strain exhibited fluorescence induction kinetics that sug-gested that the primary electron acceptor for photosystem IIwas lacking. Mutants of Chlamydomonas reinhardi and Scene-desmus obliquus with similar fluorescence kinetics have alsobeen reported (6, 19). The mutants of C. reinhardi were de-ficient, but not altogether lacking, in plastoquinone and cyto-chrome b. was also missing or nonfunctional. Bishop (6)demonstrated that the mutants of S. obliquus were deficient, butnot completely lacking, in plastoquinone A. Recently thesemutants of S. obliquus were shown to have little or no EPR'signal II or delayed light emission (9).
1 This work was supported in part by National Institutes ofHealth Training Grant 5T1-GM 1195 (F.D.S.) and National Sci-ence Foundation Grant GB-8462 (R.A.D.)
I Contribution No. 457 from the Charles F. Kettering ResearchLaboratory, Yellow Springs, Ohio.
I Present address: Department of Biology, University of Dayton,Dayton, Ohio 45409.
' Abbreviations: DPC: 1, 5-diphenylcarbazide; DPIP: 2, 6-di-chlorophenolindophenol; EPR: electron paramagnetic resonance.
The study of the mutant P, of Euglena gracilis was under-taken to determine: (a) what component(s) of the thylakoidmembranes was(were) altered or missing in the mutant; and (b)the biochemical and biophysical characteristics of the mutant.
MATERIALS AND METHODS
Culture and Maintenance of Algal Strains. The wild type ofEuglena gracilis, Z strain, was obtained from Dr. W. R. Evans.The photosynthetic mutant P, of Euglena gracilis, Z strain, wasa gift of Dr. H. Lyman. Wild-type and mutant strains weregrown heterotrophically in the dark at 24 C in Hutner medium(17) modified as described by Mollenhauer et al. (20). Cultureswere grown with reciprocal shaking in 500-ml Erlenmeyerflasks containing 100 ml of medium. Each organism was grownphotosynthetically in inorganic Hutner medium (17). Eachflask received as inoculum approximately 2 X 10' cells/ml in120 ml of Hutner medium. The cells were exposed with re-ciprocal shaking to 1.6 X 10' ergs cm' sec' of fluorescent lightand harvested after 72 hr.
Isolation of Algal Chloroplasts. Cells grown in the light wereharvested by centrifugation at 10OOg for 5 min. The resultingpellet was washed in a solution containing 0.4 M sucrose, 0.1M NaCl, and 50 mm phosphate buffer, pH 7.4. The pellet wassuspended in 10 to 20 ml of the above solution and passedthrough the Yeda press twice at 1000 p.s.i. The cell ho-mogenate was strained first through 6 and then through 12layers of cheesecloth and centrifuged briefly at 1OOg to removewhole cells and cell debris. The supematant was centrifugedat IOOOg for 5 min to yield a plastid-containing pellet. Forenzyme assays this pellet was resuspended in a small amountof the above solution. For electrophoretic analysis the plastid-containing pellet was resuspended in a solution containing 10mM KCl and 50 mm phosphate buffer, pH 7.4, and centrifugedfor 5 min at 1000g. The same buffer was added to yield a sus-pension of plastids which contained 0.25 mg chl/ml. Fivemilliliters of this chloroplast suspension was layered on top ofa discontinuous gradient containing buffer solutions with thefollowing concentrations of sucrose: 0.7 M sucrose, 1.0 M su-crose, and 1.8 M sucrose. The Beckman SW-27 tubes containingthese gradients were centrifuged for 1 hr at 131,000g. The ma-terial that collected at the 1.0 to 1.8 M interface was resus-pended in a solution containing 10 mm KCI and 50 mm phos-phate buffer, pH 7.4, and centrifuged at 1000g. The resultingpellet was frozen and saved for analysis.Enzyme Assays. Photosystem II activity was followed spec-
trophotometrically using a modified Bausch and Lomb 505spectrophotometer. The instrument had been modified to allowthe sample to be illuminated by a tungsten lamp mounted out-side the housing of the spectrophotometer. A Corning 2403 red
161

SCHWELITZ, DILLEY, AND CRANE
filter was used between the actinic light source and the reactionvessel and a Corning 9782 blue filter was placed in front of thephototube to block scattered actinic light. Light-induced reduc-tion of DPIP was followed at 580 nm. The reaction mixturecontained 50 mm K2HPO4, pH 7.0, 75 /tM DPIP and 10 to 20
jug of chlorophyll per ml in a final volume of 2.5 ml. DPC at0.5 mm was included in some cases as an electron donor forphotosystem 11 (29).
Chlorophyll was determined using the method of Arnon (1).Quinone and Carotenoid Analyses. Washed, frozen cells
were thawed and suspended in a solution containing 50 mMphosphate buffer, pH 7.2. After homogenization the suspensionwas mixed in a solution of acetone, methanol, and isopropylalcohol (1:1:1, v/v) and sonicated for 2 min with a BransonModel S-75 sonicator at5 amp.
Extractions and chromatography of extracts were performedin a dark room under 7-w lights. All chemicals were reagentgrade. The above cell suspension was mixed with petroleumether (30-60 C) and heptane (1:1, v/v) and a separatory funnelwas used to separate the green nonpolar phase, containingplastoquinones, from the aqueous phase. The latter was washedseveral times with 50% (v/v) aqueous methanol until all thechlorophyll was transferred to the nonpolar phase. The lipidextract was then evaporated to dryness in vacluo and dissolvedin a known volume of petroleum ether for chromatography onan alumina column, or in heptane for chromatography on silicagel GHR plates.The lipid extract was chromatographed on columns of acid-
washed alumina, deactivated with deionized water. The lipidextract was applied to the column in petroleum ether andcomponents were eluted with increasing concentrations of di-ethyl ether (0.02-20%; v/v) in petroleum ether. Each samplewas evaporated to dryness in vactuo, dissolved in a known vol-ume of ethanol, and analyzed spectrophotometrically.
In some cases the heptane extract was streaked directly ontoa silica gel GHR plate. After development with a solution ofchloroform and heptane (80:20), a small side strip was sprayedwith leucomethylene blue as described by Barr et al. (3).Quinones appeared as blue spots. The quinone and carotenoidbands were eluted with ethanol and analyzed spectrophoto-metrically.The amounts of quinones were calculated from the changes
in absorbancy between the oxidized and reduced forms at 255nm with a Unicam spectrophotometer after reduction withborohydride as described by Sun et al. (27).
Quinones together with standards were rechromatographedon silica gel GHR plates which were developed with chloro-form :heptane (80: 20). In order to achieve a clearer separationbetween plastoquinone A and plastoquinone B some sampleswere spotted on a 5% (v/v) paraffin oil-impregnated plate anddeveloped with a solution of acetone, water, and paraffin oil(95:5:0.1). The plates were sprayed with leucomethyleneblue as above.Low Temperature Fluorescence Spectra. The chloroplasts
isolated as described above were frozen in a low temperaturesample holder with liquid N2 and were maintained at liquidN2 temperature throughout the measurements. Fluorescencespectra were obtained with a spectrofluorometer consisting ofa Bausch and Lomb 0.5 M monochromator and an EMI photo-multiplier tube (No. 9558 C). To prevent scattered light fromreaching the phototube, a Corning filter (No. 2412) was placedbetween the sample and monochromator. The sample was ir-radiated from the front and the fluorescence was detected at anangle from the back. The measuring light from a SylvaniaDWY "sungun" lamp was filtered through 1 cm of a saturatedcopper sulfate solution and a Baird Atomic interference filterwith maximum transmittance at 435 nm.
Measurements of Electron Paramagnetic Resonance. Chloro-plasts, isolated as previously described, were used for theseexperiments. Thick suspensions of chloroplasts in tris bufferwere placed in an aqueous sample cuvette. EPR was measuredwith a Varian V 4500 EPR spectrometer. Exciting light was
provided by a Sylvania SWY"sungun" lamp and was filteredthrough 4 cm of a copper sulfate solution.Measurements of Delayed Light Emission. Whole cells, sus-
pended in Hutner medium, were used in these experiments.The very fast component of delayed light emission was meas-ured with a Becquerel phosphoroscope. The excitation windowopened for 0.5 msec and the measuring window was openedfor a period of 4.5 msec. A red filter was placed between theexcitation light and sample.The two slower components of delayed light emission were
followed on a phosphoroscope. A red filter was positioned be-tween the excitation light and sample.
Cytochrome Measurements. Cytochrome absorbance changeswere measured in an Aminco-Chance dual wavelength spectro-photometer using a PAR lock-in amplifier as described by Fanand Cramer (15). The measuring beam intensity was about1erg/Cm2 sec with a 2.1 nm half band width. Actinic light in-tensities, 6.3X 10' ergs/cm2 sec and 7.0 x 10I ergs/cm2 sec,at 645 nm and 713 nm respectively were incident on thecuvette.
Cytochrome b.59 changes in isolated chloroplasts were meas-ured at 560 nm with 570 nm as reference. The final concentra-tion of the reactants in the mixture was: Tricine-NaOH. pH7.5. 25 mM; MgCl2, 5 mm; Na2HPO,, 5 mm; and chloroplastswith 50 to 100 jug/ml chl. Ferricyanide (0.5 mM) or 1 mMascorbate was added at appropriate times for oxidation or re-duction of cytochrome b,,.
Electrophoresis. The method of Takayama et al. (28) wasused for disc gel electrophoresis. A Canalco model 300B powersupply was utilized with a model 1200 running chamber. A cur-rent of 3 ma per tube was applied for the first 15 min to setthe protein in the gel. This was followed by a 70-min run withthe current set at 5 ma per tube. Gels were stained for 30 minin a 0.17% (w/v) amido black solution. Densitometer tracingsof the samples were obtained from a Gilford spectrometer 240with a densitometer attachment.
RESULTS
Russell and Lyman (22) showed that the mutant P4 was un-able to carry out the Hill reaction with DPIP. In order tolocalize definitively the position of the block in photosystem IIof the mutant, we extended the study to the use of an electrondonor. Wild-type chloroplasts normally reduce 90 to 100,umoles DPIP/mg chl-hr whereas the mutant is unable to re-duce this Hill reagent. When the electron donor, DPC (29),which bypasses the oxygen-evolving mechanism, was utilized,this rate of DPIP reduction was often increased by 10 to 20,tmoles/mg chl*hr in the wild type. However, the mutantchloroplasts were incapable of carrying on the Hill reactionwith this electron donor.
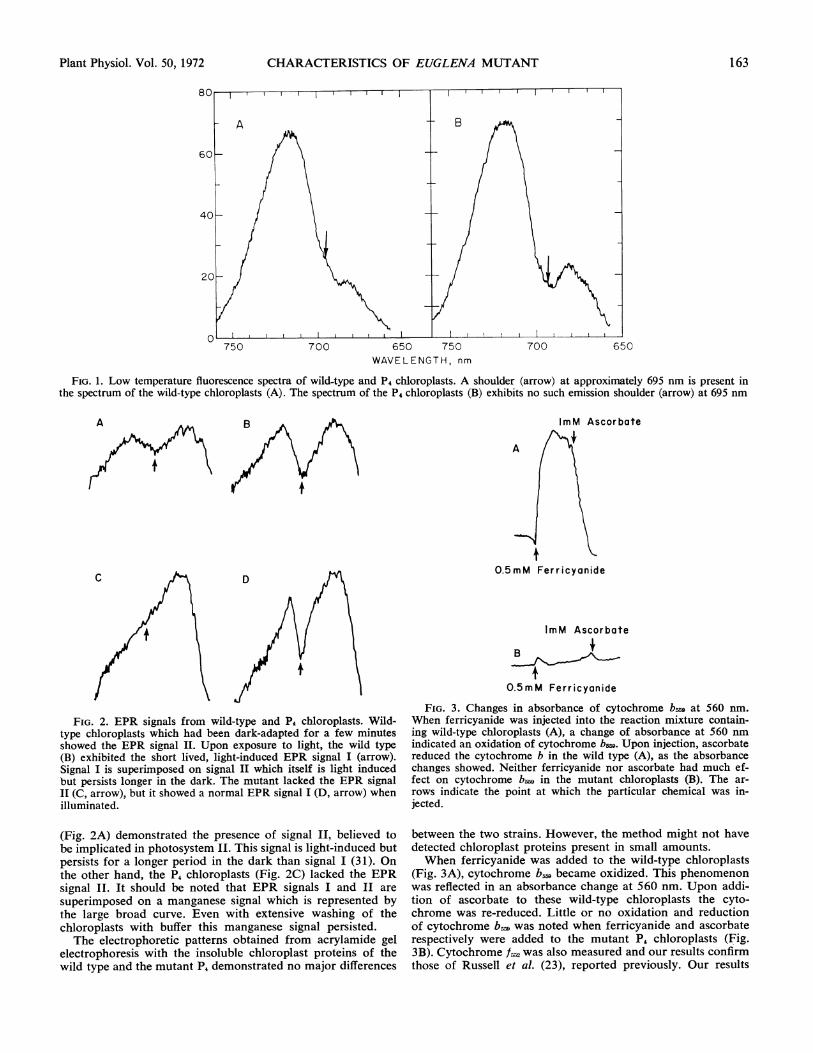
Figure 1 shows the results of the low temperature fluores-cence experiments. The emission spectra of the wild-type (A)and mutant (B) chloroplasts appear similar with the exceptionof a shoulder at 695 nm which is present in the wild-type (A)spectrum but lacking in that of the mutant (B).The wild-type cells exhibited three components of delayed
light emission: 5 msec, 30 msec, and 10 sec components. TheP4 cells lacked all three components of delayed light emission.
Both wild-type (Fig. 2B) and mutant P4 (Fig. 2D) exhibitedthe light-induced, short-lived EPR signal I, thought to be as-sociated with photosystem I (31). The wild-type chloroplasts
Plant Physiol. Vol. 50, 1972162
CHARACTERISTICS OF EUGLENA MUTANT
750 700 650 750 700 650WAVELENGTH, nm
FIG. 1. Low temperature fluorescence spectra of wild-type and P4 chloroplasts. A shoulder (arrow) at approximately 695 nm is present inthe spectrum of the wild-type chloroplasts (A). The spectrum of the P4 chloroplasts (B) exhibits no such emission shoulder (arrow) at 695 nm
A B
C D
FIG. 2. EPR signals from wild-type and P4 chloroplasts. Wild-type chloroplasts which had been dark-adapted for a few minutesshowed the EPR signal II. Upon exposure to light, the wild type(B) exhibited the short lived, light-induced EPR signal I (arrow).Signal I is superimposed on signal II which itself is light inducedbut persists longer in the dark. The mutant lacked the EPR signalII (C, arrow), but it showed a normal EPR signal I (D, arrow) whenilluminated.
(Fig. 2A) demonstrated the presence of signal II, believed tobe implicated in photosystem II. This signal is light-induced butpersists for a longer period in the dark than signal I (31). Onthe other hand, the P4 chloroplasts (Fig. 2C) lacked the EPRsignal II. It should be noted that EPR signals I and II aresuperimposed on a manganese signal which is represented bythe large broad curve. Even with extensive washing of thechloroplasts with buffer this manganese signal persisted.The electrophoretic patterns obtained from acrylamide gel
electrophoresis with the insoluble chloroplast proteins of thewild type and the mutant P4 demonstrated no major differences
ImM Ascorbate
0.5mM Ferricyanide
ImM Ascorbate
B~~~~0.5mM Ferricyanide
FIG. 3. Changes in absorbance of cytochrome bw9 at 560 nm.When ferricyanide was injected into the reaction mixture contain-ing wild-type chloroplasts (A), a change of absorbance at 560 numindicated an oxidation of cytochrome b&. Upon injection, ascorbatereduced the cytochrome b in the wild type (A), as the absorbancechanges showed. Neither ferricyanide nor ascorbate had much ef-fect on cytochrome bm9 in the mutant chloroplasts (B). The ar-rows indicate the point at which the particular chemical was in-jected.
between the two strains. However, the method might not havedetected chloroplast proteins present in small amounts.When ferricyanide was added to the wild-type chloroplasts
(Fig. 3A), cytochrome b: became oxidized. This phenomenonwas reflected in an absorbance change at 560 nm. Upon addi-tion of ascorbate to these wild-type chloroplasts the cyto-chrome was re-reduced. Little or no oxidation and reductionof cytochrome b. was noted when ferricyanide and ascorbaterespectively were added to the mutant P4 chloroplasts (Fig.3B). Cytochrome f;,2 was also measured and our results confirmthose of Russell et al. (23), reported previously. Our results
163Plant Physiol. Vol. 50, 1972

SCHWELITZ, DILLEY, AND CRANE
Table I. Carotenioid antd Quinionie Conztent ofWild-Type and P4 cells
Quinone or Carotenoid WN'ild Type P4
gnwles/,ng. clil ump1les/g iLmolesIing ci lAnoleslgdry wtt dry wt
Plastoquinone A 0.02 0.15 0.00 0.00Plastoquinone B 0.00 0.00 0.02 0.11Plastoquinone C 0.02 0.11 i 0.02 0.08Coenzyme Qg 0.05 0.34 0.09 0.46Tocopherylquinone 0.04 0.22 0.06 0.30Carotene 0 .005 0.003 0.001 0.007Xanthophyll 0.00004 0.00002 0.0005 0.003
indicated that there was more cytochrome f. on a chlorophyllbasis in the mutant than in the wild-type so that the measure-ments of 560 nm would seem to reflect the state of cytochromeb559. The cytochrome b,, measurements also were made onisolated chloroplasts in which there remains very little cyto-chrome f, since Euglena cytochrome f is very soluble. How-ever, a difference spectrum is needed to show clearly the stateof cytochrome b5W,.The results of the quinone and carotenoid analyses are pre-
sented in Table I. On a dry weight basis the wild-type and P4cells have comparable amounts of quinones with the exceptionof plastoquinone A and plastoquinone B. The wild type, likeall green algae studied (27), possesses plastoquinone A but no
detectable amount of plastoquinone B. On the other hand,plastoquinone A is apparently absent in the mutant cells whichdo, however, contain plastoquinone B. The mutant P4 cellsshowed a higher complement of carotenoids, especially xantho-phylls. Although not shown in the table, the mutant had com-
parable or less ft-carotene than did the wild type. There was a
high proportion of carotenes, other than f-carotene, present inthe mutant but not in the wild type. These data on caroteneswere noted visually on the chromatograms. Further methodsneed to be employed to identify and quantitate the types ofcarotenes, other than ft-carotene. Spectral and chromatographicdata also suggest that additional differences in lipids other thanquinones and carotenoids should be sought between wild-typeand P4 chloroplasts.
DISCUSSION
Russell and Lyman (22) showed that Euglena gracilis P4 was
incapable of carrying on the Hill reaction. If the mutationaffected the oxygen-evolving mechanism of photosystem II,
the electron donor DPC (29), which feeds electrons directlyinto photosystem II and bypasses the oxygen-evolving mecha-nism of photosystem II, would have restored Hill activity tothe mutant chloroplasts. However, the P4 chloroplasts exhibitedno electron transport with this donor. This confirmed thepostulation of Russell and Lyman (22) that the block in themutant P4 was in the electron transport chain between the twophotosystems.
In the light of evidence from the kinetics of fluorescence in-duction in the mutant P4, Russell et al. (23) hypothesized thatthe primary acceptor for photosystem II (also known as thequencher or Q) was missing or nonfunctional. Our data fromdelayed light emission support this suggestion. Since delayedlight is thought to originate from an electron dropping backto chlorophyll from the primary acceptor, no delayed lightemission would be expected if the primary acceptor were
lacking or nonfunctional. The data from mutant P4 were con-
sistent with this interpretation.
The data from fluorescence spectra at low temperatures werealso consistent with a missing or nonfunctional primary ac-ceptor for photosystem II in the mutant P4. The emission spec-trum of P4 chloroplasts (Fig. 1B) lacked a shoulder at 695 nmwhich the spectrum of the wild-type chloroplasts possessed(Fig. IA). Others (5, 12, 17) proposed that this shoulder is dueto emission from the hypothetical energy trap for photosystemII. Since the quencher is an integral part of this trap, no emis-sion band would be expected in the mutant with a defectivetrap.The mutant cells possess more carotenoids, especially
xanthophylls, than the wild-type cells (Table I). The caro-tenoids are thought to play a major role in protecting chloro-plasts against photosensitized oxidations. On the basis of ex-periments with Euglena gracilis, Bamji and Krinsky (2)suggested that chlorophyll, excited by light, is usually deacti-vated by the process of photosynthesis. but sometimes theexcited chlorophyll may combine with molecular oxygen. Thiscombination could lead to photosensitized oxidations butusually this lethal combination is inactivated by a carotene,such as ft-carotene, which itself would be oxidized to a xan-thophyll. Since light energy is not utilized in the mutant be-cause there is a block between the two light reactions, it islikely that the above phenomenon happens to a greater extentin the mutant than in the wild type. This phenomenon wouldaccount for the large proportion of xanthophyll in the mutantP4.
Since the mutant P4 is deficient in plastoquinone A and seemsto possess a missing or nonfunctional quencher. it is temptingto assign plastoquinone A the role of the primary acceptor forphotosystem II. Bishop (6) suggested this role for plastoqui-none A in light of evidence from Scenedesinus mutants whichare deficient in plastoquinone A and have fluorescence kineticssimilar to those of mutant P4. Stiehl and Witt (26) demon-strated that a semiquinone twin, PQ--PQ-, has the character-istics and half time of the primary acceptor which they hadcharacterized previously (25). Our data are consistent with thisbut we cannot conclude unequivocally that plastoquinone A isthe primary acceptor for photosystem II. The P4 cells possesseddetectable amounts of plastoquinone B while the wild-typecells, as well as other green algae (27), seemingly lack this qui-none. Crane et al. (13) showed that plastoquinone B cannotreplace plastoquinone A in its function between the two photo-systems. Our data with the P4 strain confirm this.
Several groups (7, 18, 30) have suggested that plastoquinoneA is responsible for the EPR signal II. Since the P4 chloroplastslacked both plastoquinone A and EPR signal II, we suggestthat plastoquinone A may be responsible for the EPR signalII. However, the component responsible for the EPR signal IIcould not be the primary acceptor since the signal persists inthe dark for a rather long time. If plastoquinone A is the pri-mary acceptor, the EPR signal II must originate from a sourceof plastoquinone A other than the primary acceptor. The plas-toquinone pool described by Stiehl and Witt (25) and Schmidt-Mende and Rumberg (24) may well be that source. Stiehl andWitt (26) place this pool of approximately six plastoquinonemolecules per electron transport chain in direct connectionwith the twin semiquinone postulated to be the primary ac-ceptor for photosystem II.Our data indicate that cytochrome b,.. is missing or non-
functional in the P4 chloroplasts. Lavorel and Levine (19) re-ported on mutants of Chlamydomonas reinhardi which possessa nonfunctional or missing cytochrome b559. are deficient inplastoquinone (the types of plastoquinone were not distin-guished), and have induction kinetics suggesting that thequencher is absent or nonfunctional. The similarity between
164 Plant Physiol. Vol. 50, 1972

CHARACTERISTICS OF
mutant P4 of Euglena gracilis and these Chlatnydomnonas mu-
tants is striking.The position of cytochrome br in the electron transport
chain between the two light reactions is not clear (4, 12, 14).Since the P4 mutant of Euglena gracilis and the Chllainydorno-nas mutants both have nonfunctional or missing cytochromebz, and are plastoquinone deficient, there probably is some
interrelationship between cytochrome b,. and plastoquinone.In electrophoresis experiments the chloroplast proteins of wild-type and P4 cells appeared similar. However, it is doubtfulthat proteins such as cytochrome b,, which make up a verysmall proportion of the total chloroplast proteins, would bedetectable by the electrophoretic methods employed. The pres-
ent models for the position of cytochrome b, are compatiblewith our data but none gives a definite clue to why cytochromeb. may be missing or nonfunctional in the mutant.
Acknowledgments-Appreciation is extended to Drs. R. Barr, H. Bbhme,W. Cramer, W. Evans, D. Fleischman, and B. C. Mayne for their contribu-tions in this work.
LITERATURE CITED
1. ARNON, D. I. 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxi-dase in Beta vulgaris. Plant Physiol. 24: 1-15.
2. BAMJI, M. S. AND N. I. KRINBKY. 1965. Carotenoid de-epoxidations inalgae. II. Enzymic conversion of antheraxanthin to zeaxanthin. J. Biol.Chem. 240: 467-470.
3. BARR, R., M. D. HENNINGER, AND F. L. CRANE. 1967. Comparative studieson plastoquinone. II. Analysis for plastoquinone A, B, C and D. PlantPhysiol. 42: 1246-1254.
4. BENDALL, D. S. AND D. SOFROVk. 1971. Reactions at 77VK in photosyIstem2 of green plants. Biochim. Biophys. Acta 234: 371-380.
5. BERGERON, J. A. 1963. Studies of the localization, physiochemical proper-
ties, and action of phycocyanin in Anacystis nidulans. In: PhotosyntheticMechanisms of Green Plants. National Academy of Science-National Re-search Council, Publication No. 1145, Washington, D. C. pp. 527-536.
6. BISHOP, N. I. 1969. Fluorescent and photochemical characteristics of sys-
tem II mutants deficient in plastoquinone A. Biophys. J. (Abstr.) 9: 118.7. BisHoP, N. I. 1961. The possible role of plastoquinone (Q 254) in the electron
transport system of photosynthesis. In: Quinones in Electron Transport.Little, Brown, and Co., Boston. pp. 385-404.
8. BISHOP, N. I. 1959. The reactivity of naturally occurring quinone (Q-255)in photochemical reactions of isolated chloroplasts. Proc. Nat. Acad. Sci.U.S.A. 45: 1696-1702.
9. BISHOP, N. I. AND J. WONG. 1971. Observations on photosystem II mutantsof Scenedesmus. Pigments and proteinaceous components of the chloro-plasts. Biochim. Biophys. Acta 234: 433-445.
10. BOARDMAN, N. K., J. M. ANDERSON, AND R. G. HILLER. 1971. Photooxida-tion of cytochromes in leaves and chloroplasts at liquid-nitrogen tempera-ture. Biochim. Biophys. Acta 234: 126-136.
11. BOHME, H. and W. A. CRAMER. 1971. Plastoquinone mediates electron
EUGLENA MUTANT 165
transport between cytochrome b-559 and cytochrome-f in spinach chloro-plasts. FEBS Lett. 15: 349-351.
12. BRODY, S. S. AND M. BRODY. 1963. Aggregate chlorophyll in vivo. In: Pho-tosynthetic Mechanisms of Green Plants. National Academy of Science-National Research Council, Publication No. 1145, Washington, D. C. pp.455-478.
13. CRATNE, F. L., M. D. HENNINGER, P. M. WOOD, AND R. BARR. 1966. Bio-chemistry of Chloroplasts, Vol. I. Academic Press, New York. pp. 133-151.
14. ERIsoN, K. AND W. L. BUTLER. 1971. The relationship between Q, C-550and cytochrome b-559 in photoreactions at - 196° in chloroplasts. Biochim.Biophys. Acta 234: 381-389.
15. FA.-N, H. N. AND W. A. CRAMIER. 1970. The redox potential of cytochromesb-559 and b-563 in spinach chloroplasts. Biochim. Biophys. Acta 216:200-207.
16. GOVINDJEE, R. 1963. Emerson enhancement effect and two light reactions inphotosynthesis. In: Photosynthetic Mechanisms of Green Plants. NationalAcademy of Science-National Research Council, Publication No. 1145,Washington, D. C. pp. 318-334.
17. HUTNER, S. H., A. C. ZAHALSEY, S. AARONSON-, H. BAKER, and 0. FRANK.1964. Culture media for Euglena gracilis. In: Methods in Cell Physiology,lVol. II. Academic Press, New York. pp. 217-227.
18. KOHL, D. H. AND P. M. WOOD. 1969. On the molecular identity of ESR sig-nal II in photosynthetic systems: the effect of heptane extraction and re-constitution with plastoquinone and deuterated plastoquinone. PlantPhysiol. 44: 1439-1445.
19. LAVOREL, J. AND R. P. LEVINE. 1968. Fluorescence properties of wild-typeChlamydomonas reinhardi and three mutant strains having impairedphotosynthesis. Plant Physiol. 43: 1049-1055.
20. NMOLLENHAUER, H. H., W. EVAN'S, AND C. KOGUT. 1968. Dictyosome structureof Euglena gracilis. J. Cell Biol. 37: 579-583.
21. ROSENBERG, A. AND J. GOUAUX. 1967. Quantitative and compositionalchanges in monogalactosyl and digalactosyl diglycerides during light-in-duced formation of chloroplasts in Euglena gracilis. J. Lipid Res. 8: 80-83.
22. RUSSELL, G. K. AND H. LYMAN. 1968. Isolation of mutants of Euglenagracilis with impaired photosynthesis. Plant Physiol. 43: 1284-1290.
23. RuSSELL, G. K., H. LYMAN, AND R. L. HEATH. 1969. Absence of fluorescencequenching in a photosynthetic mutant of Euglena gracilis. Plant Physiol.44: 929-931.
24. SCH'MIDT-MENDE, P. AND B. RUMBERG. 1968. Zur Plastochinonreduktion beider Photosynthese. Z. Naturforsch. 23b: 225-228.
25. STIEHL, H. H. AND H. T. WiTT. 1968. Die kurzzeitlegen ultravioletten Dif-ferenzspektren bei der Photosynthese. Z. Naturforsch. 23b: 220:224.
26. STIEHL, H. H. AXD H. T. WITT. 1969. Quantitative treatment of the func-tion of plastoquinone in photosynthesis. Z. Naturforsch. 24b: 1558-1598.
27. SuN, E., R. BARR, AND F. L. CRANE. 1968. Comparative studies on plasto-quinones. IV. Plastoquinones in algae. Plant Physiol. 43: 19351940.
28. TAKAYAMA, K., D. H. MAcLENNAN, A. TZAGOLOFF, AND C. D. STONER. 1966.Studies on the electron transfer system. LXVII. Polyacrylamide gel electro-phoresis of the mitochondrial electron transfer complexes. Arch. Biochem.Biophys. 114: 223-230.
29. VERNON, L. P. AND E. R. SHAW. 1969. Oxidation of 1,5-diphenylcarbazideas a measure of photosystem 2 activity in subehloroplast fragments.Biochem. Biophys. Res. Commnun. 36: 878-884.
30. W-EAVER, E. C. 1962. Possible interpretation of the slow-decaying EPR sig-nal in algal suspensions. Arch. Biochem. Biophys. 99: 193-196.
31. WEAVER, E. C. 1968. EPR studies of free radicals in photosynthetic systems.Annu. Rev. Plant Physiol. 19: 288-294.
Plant Physiol. Vol. 50, 1972





























