Embed Size (px)
Citation preview

8/3/2019 Biology Under Construction- In Vitro Reconstitution of Cellular Function
http://slidepdf.com/reader/full/biology-under-construction-in-vitro-reconstitution-of-cellular-function 1/8
Bioogy resents many strange and wonderfexames of what is ossibe in the hysicaword. We might not fy nderstand howaxons are gided dring deveoment orhow stem ces divide asymmetricay, btmost scientists wod accet that there is ahysica exanation at the cear eve —given the right moeces and bondary conditions, the known aws of hysics governthe interactions that drive bioogica behav-ior. Moeces that are crcia for nmeroscear fnctions, sch as motiity anddivision, have been identified. Frthermore,the imortance of bondary conditions,sch as membranes, and their fidity, ateraorganization and mechanica comiance,is becoming cear1,2. However, an imortantqestion remains: how are the moecesand the bondary conditions inked to gen-erate the remarkaby coordinated and robst
cear behavior that we observe, whichoften invoves the creation of strctres withength scaes of many micrometres overtimescaes of mintes to hors?
The concets of sef-organizing systemsand cooerative behavior were firstexored in the context of chemica systemsin the 1920s, and in the ast ten yearsthey have been rodctivey aied tothe stdy of sef-assemby in ce bioogy 3.Reconstittions of cytoskeeta fiamentsand their interactions with motors and otherbinding roteins are rime exames of how
dynamic behavior observed inside a cecan be recreated and stdied otside thece4. Advancement towards recreating morecomex ce-ike behavior (referred to hereas cear reconstittion), sch as ce divi-sion and ce motiity, wi invove confiningreactions to sma vomes that are bondby a biayer membrane and incororatingsignaing athways that are controed by externa stimi. Exeriments on iving ces,which contine to be crcia for testingtheories of how moeces interact to giverise to hysioogica fnctions, cannot by themseves satisfy Feynman’s idea of nder-standing: “That which I cannot create I donot nderstand”5. Reconstittion stdiessing ce-free extracts or rified roteinsrovide a comementary aroach toive-ce stdies; taken together, these twoaroaches can ead to an nderstanding of
bioogica rocesses that advances both basicand aied sciences.
This kind of redctionist bioogy —erhas more aroriatey caed constrc-tionist bioogy — offers three imortant
insights that comement the investigation of iving ces. First, condcting in vitro recon-stittion exeriments can confirm and refinemoecar modes of rocesses otside thecomicating environment of ces. Second,nexected behavior can arise in simifiedreconstittions constrained ony by knownhysica aws and secific biochemicainteractions, and exaining sch behaviormight yied new insight into bioogicaorganization. Third, reconstittions areoften amenabe to mathematica modeingthat considers sace, time and a manageabe
nmber of other variabes to describe thecomonents invoved. The eve of detaiednderstanding avaiabe from reconstittionstdies can he revea how evoved bio-ogica systems work and rovide insightinto how new bioogica fnctions cod beengineered.
A growing nmber of satiay organizedrocesses have now been reconstitted andcharacterized sing rified comonentsand ce-free extracts (see next section; FIG. 1).The recent srge in cear reconstittionsreorted in the iteratre indicates anincreasingy rodctive convergence of biochemica knowedge, exerimenta toosand theoretica nderstanding, which isneeded to bid cear strctres fromscratch. In this Oinion artice, we tracethe roots of today’s cear reconstittionefforts back to the eary days of biochemicareconstittion and discss technicaadvances that enabe the crrent work,aong with the remaining imitations. Wesggest that reconstitting cear-scaestrctres and behavior from comonentarts, an aroach that is oised to becomemore accessibe, informative, and sef
than ever before, is a crcia ste towardsanswering some of the most fndamentaqestions of ce bioogy.
Early days of recosttuto
When biochemistry emerged as its ownfied, qestions regarding the satia organ-ization of cear strctres were secondary to more basic qestions abot their moec-ar make, stoichiometric comositionand fnctiona significance (TIMELINE). Inthe 1940s, actomyosin threads from mscewere reconstitted and biochemicay
O p i n i O n
Biology under construction: in vitro reconstitution of cellular function
Allen P. Liu and Daniel A. Fletcher
Abra | W ar muh br a akng ll apar han pung hm oghr.
Ronuon of bologal pro from omponn molul ha bn a
powrful bu dfful approah o udyng funonal organzaon n bology.
Rnly, h onrgn of bohmal and ll bologal adan wh nwxprmnal and ompuaonal ool prodng h opporuny o ronu
nrangly omplx pro. W prd ha h boom-up ragy wll
unor ba pro ha gud llular ambly, adanng boh ba and
appld n.
...reconstituting cellular-scale
structures and behaviour ... is a
crucial step towards answering
some of the most fundamental
questions of cell biology.
PeRsPectives
644 | SEpTEMBER 2009 | VOluME 10 www.a.m/ws/mb
©2009M ill P bli h Li it d All i ht d
8/3/2019 Biology Under Construction- In Vitro Reconstitution of Cellular Function
http://slidepdf.com/reader/full/biology-under-construction-in-vitro-reconstitution-of-cellular-function 2/8
Spindle formation
Membranenetworkformation
Lamellipodium
Bacterialmotility
Immunologicalsynapse
Filopodium
characterized by Szent-Gyoergyi, and aterby Strab (see REF. 6 for an overview), ead-ing the way for the nderstanding of howmsces contract. Not ong after, the discov-ery of the strctre of DNA by Watson andCrick in 1953 (REF. 7) ed to a frry of inves-tigation on the synthesis of DNA, incdingArthr Kornberg’s enzymatic stdy of DNA reication8. Simy mixing the fordeoxynceotide comonents of DNA andEscherichia coli extract roved to have ittesccess, as nwanted ncease activitieswere resent in the extracts. With therification of what is now known as DNAoymerase I, Kornberg and coeagessccessfy reconstitted the synthesis of DNA9. The concetay sime three-comonent enzymatic reaction, reqiringthe for deoxynceotides, singe-strandedtemate DNA and rified DNA oy-merase, has aved the way for frther
stdies of DNA reication.Biochemica reconstittion has been
embraced by severa fieds of modern cebioogy beyond the cytoskeeton and DNAreication. The stdy of membrane traffick-ing began with the identification and isoationof yeast secretion (Sec) mtants that bockedmembrane traffic10. later, sing a reconstit-tion aroach with rified roteins andsemi-intact ces, coat rotein I (COpI) andcoat rotein II (COpII) were identified asroteins invoved in rotein transort11,12.Biochemica reconstittion has aso ed to abetter nderstanding of the rotein trans-ocation rocess in the endoasmic retic-m (ER) and of how other DNA enzymesfnction13,14. More recenty, osciation of thecircadian cock was reconstitted15.
In these exames, biochemica recon-stittion is aimed at identifying secificmoecar reactions that oerate at moecar-ength scaes. How ces assembe, organizeand se micrometre-scae strctres isemerging as a centra qestion in ce bio-ogy, one that is now becoming amenabe toreconstittion.
Towards cellular recosttutoBioogica activities and reactions are oftenconfined to secific ocations in the ce,sch as on the asma membrane, in variosorganees and bond to biooymers, schas the cytoskeeton and chromosomes.Reconstittion of satiay organized ro-cesses often reqires the secific hysicaor biochemica bondary conditions thatocaize reactions or restrict diffsion to beestabished on the cear scae, an asectthat was not the focs of eary biochemicareconstittions.
In some cases, the system of biomoe-ces natray incdes its own bondary conditions. Cytoskeeta oymers recons-titted with binding roteins can satiay organize on the micrometre scae in theabsence of hysica confinement, withthe oymer itsef serving as a scaffod forsef-assemby. Notabe exames incde theformation of organized strctres, sch asasters and vortices that arise from mixtresof microtbes and motor roteins16, aswe as the sontaneos transition froma dendritic actin network to bnded actinfiaments17. The arge-scae organizationexhibited by the microtbe system isreviewed esewhere18.
In other cases of reconstittion, the
bondary conditions mst be added exi-city. using a new coection of exerimentaaroaches, researchers are ‘seeding’ thesatia organization of cear reconstit-tions by restricting biochemica activities tosrfaces, sch as a micrometre-sized oy-styrene bead or a anar fid iid biayer(FIG. 2). Confining reactions to two dimen-sions is not ony hysioogicay aroriatein many cases, bt it aso enabes the seof advanced ight microscoy techniqes,which rest in vivid images of satiay comex rocesses.
In this Oinion artice, we focs oncear reconstittion in which bondary conditions are added to gide organization.A brief srvey of sch reconstittions,smmarized in TABLE 1, is organized accord-ing to the exerimenta atform sed toreconstitte the cear-scae rocess.
Cytoskeleton assembly on microspheres. Cear shae change and intracearmotion are driven rimariy by the assemby and disassemby of cytoskeeta oymers19.poystyrene microsheres are a sef sb-strate to reconstitte many of the rocessesthat se force generation of cytoskeetaoymers. poystyrene microsheres comein many different sizes, and roteins or
DNA can be hysicay adsorbed orcovaenty conjgated onto their srface(FIG. 2a). Frthermore, individa beads canbe maniated sing otica tras, mag-netic tweezers and microiettes. However,desite the ease of maniation, roteinsthat are directy conjgated to the micro-sheres may adot non-hysioogicaconformations and they wi have restrictedatera movements.
The first notabe se of microsheres inreconstittion was for mitotic sinde form-ation driven by microtbe dynamics in
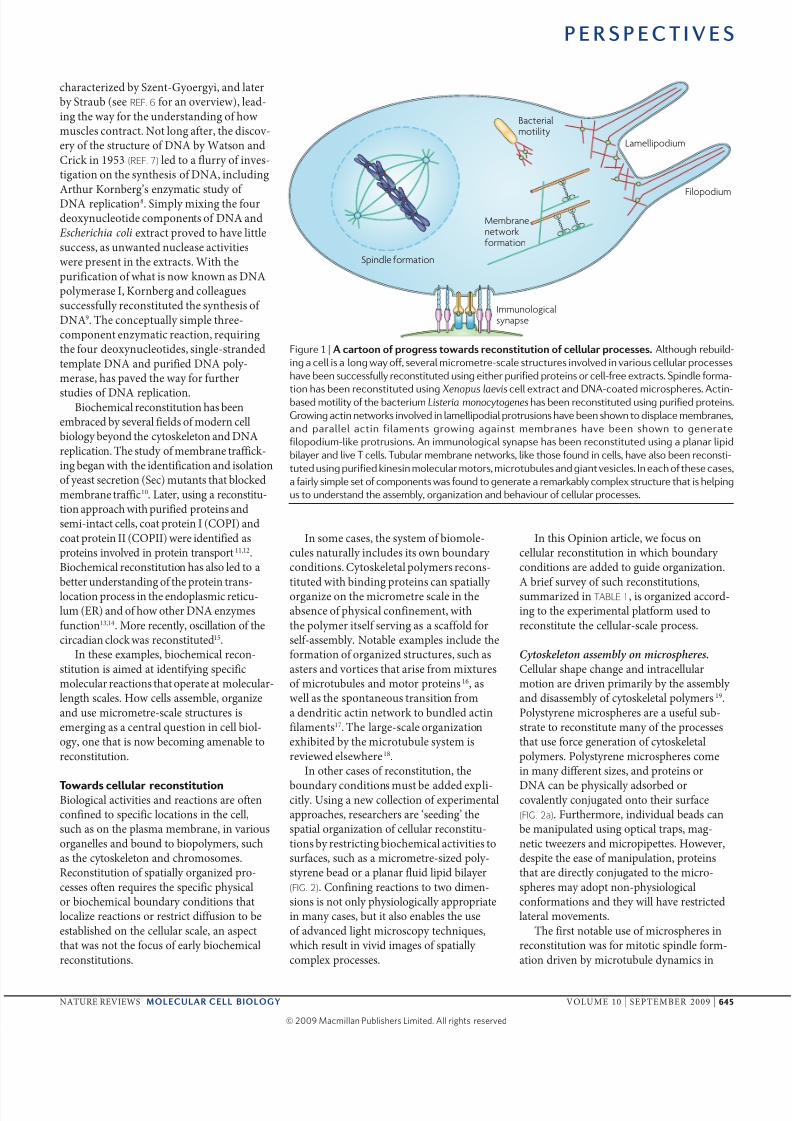
Fgur 1 | A a ss was s a sss. Alhough rbuld-
ng a ll a long way off, ral mromr-al ruur nold n arou llular pro
ha bn ufully ronud ung hr purfd pron or ll-fr xra. spndl forma-
on ha bn ronud ung Xenopus laevis ll xra and DNA-oad mrophr. An-bad moly of h barum Listeria monocytogenes ha bn ronud ung purfd pron.
Growng an nwork nold n lamllpodal proruon ha bn hown o dpla mmbran,
and paralll an flamn growng agan mmbran ha bn hown o gnra
flopodum-lk proruon. An mmunologal ynap ha bn ronud ung a planar lpd
blayr and l t ll. tubular mmbran nwork, lk ho found n ll, ha alo bn ron-
ud ung purfd knn molular moor, mroubul and gan l. in ah of h a,
a farly mpl of omponn wa found o gnra a rmarkably omplx ruur ha hlpng
u o undrand h ambly, organzaon and bhaour of llular pro.
P e r s P e c t i v e s
NATuRE REVIEWS | MoleculAr cell Biology VOluME 10 | SEpTEMBER 2009 | 645
©2009M ill P bli h Li it d All i ht d

8/3/2019 Biology Under Construction- In Vitro Reconstitution of Cellular Function
http://slidepdf.com/reader/full/biology-under-construction-in-vitro-reconstitution-of-cellular-function 3/8
a cytoasmic extract from Xenopus laevis.Mitotic sindes were fond to assembe
arond DNA-coated microsheres actingas artificia chromosomes20. The stdy showed that centrosomes (the microtbeorganizing centre) and kinetochores (theregion of the chromosome where sindefibres attach) were not reqired for sindeformation. More imortanty, it showedthat bioarity is an intrinsic roerty of mitotic sindes becase microtbes andmotor roteins assembe arond DNA inmitotic extracts in the absence of externaoarity ces.
Actin fiaments rodce forces that drivemembrane rotrsions dring rocessessch as ce motiity. Reconstittion of craw-ing motiity began with stdies of the atho-genic bacterim Listeria monocytogenes.L. monocytogenes exresses an actinnceation-romoting factor (NpF), ActA,which is reqired for hijacking the cearmachinery of the infected ce to roe itsown movement21. In an imressive feat, themotiity of dead L. monocytogenes was recon-stitted sing NpF-coated microsheres anda minima set of roteins (actin and threeactin fiament regators: the actin-reatedrotein 2/3 (Ar2/3) comex, actin deoy-
merizing factor and caing rotein)22. usinga simiar reconstitted system on NpF-coatedmicrosheres, a controversy was recenty resoved regarding the roe of caing ro-tein, a rotein that imits actin fiamentgrowth by binding to the fiament’s barbedend, and yet increases the veocity of actin-based motiity. Caref anaysis of actingrowth on microsheres reveaed thatactin fiament caing eads to an increase infiament nceation and branching23. Againsing NpF-coated microsheres, bndesof actin fiaments that resembe fioodia
(ong and thin actin-based rotrsions)were reconstitted by introdcing the actin-
bnding rotein fascin24. In addition, thesefioodia bndes were shown to be caabeof sorting L. monocytogenes motiity 25,which shows that architectray differentactin networks can sort cear motiity.
unike for ekaryotes, rokaryotic cearfnctions were not attribted to cytoskeetaoymers nti recent years26, bt they arenow thoght to be imortant in a argenmber of rocesses. Segregation of DNA-coated oystyrene microsheres drivenby oymerization of the rokaryotic actinhomooge artitioning system M (parM)has been reconstitted sing microsheresand the three-comonent system of the R1asmid, which consists of parC, parR and parM in an oeron27. The parR gene encodes
a rotein that binds cooerativey to parC toform a comex. In the absence of the parR– parC comex, the parM oymer is dynami-cay nstabe28, bt it is stabiized when itbinds to the parR– parC comex. Insertionaeongation of parM and force generationwere observed as the parR– parC-conjgatedmicrosheres were shed aart by the grow-ing oymer. Throgh these reconstittionexeriments it was shown that the R1 as-mid contained a the comonents necessary for dynamicay searching for and then staby searating asmid coies.
Reconstitution on supported lipid bilayers. To reconstitte membrane-based henom-
ena, in which diffsion in two dimensions iscrcia, a iid biayer can be sed as a sb-strate (FIG. 2b). panar sorted iid biayerscan be made by fsing sma iosomes,tiny vesices made of iids, onto cean gasscoversis. These biayers have been sed tostdy iid diffsion and have been exoitedfor biomedica aications29. Fnctionamembrane roteins can be incororated inthe sorted iid biayer sing iosomeswith inserted roteins30. panar sortediid biayers are articary sitabe forinvestigating rocesses that occr at theasma membrane, sch as the formationof the immnoogica synase, whichoccrs at the interface between an antigenresenting ce (ApC) and a ymhocyte.
The immnoogica synase is a comexsramoecar strctre that is invovedin the commnication between an ApCand a T ce. The rocess of T ce activa-tion was reconstitted by reacing theApC with a anar sorted iid biayerthat contained forescenty abeed majorhistocomatibiity comex (MHC) etideand intercear adhesion moece 1(ICAM1)31. These moeces, which are
exressed by the ApC, cod diffse freey onthe sorted iid biayer and associate withcear recetors on the T ce. The dynamicassociation and rearrangement of T ce anti-gen recetor (TCR) and ekocyte fnction-associated antigen 1 (lFA1) on the T cewith MHC etide and ICAM1 in the s-orted iid biayer forms the immnoogicasynase. This consists of a centra sramo-ecar cster (c-SMAC), in which TCR re-dominates, and a erihera sramoecarcster (-SMAC), in which adhesionmoeces are ocaized. This reconstittion
tmln | Examles of bochemcal ad cellular recosttuto
1942 1956 1978 1984 1992 1996 1999 2003 2007 2008 2009
(1956–1960) Dory and
bohmal ronuon of DNApolymra ay8
(1942–1950)
chararzaon of AtPa
ay of aomyon6
enzyma ay
of DNA gyra13
Polyppd ranloaon n
ndoplam rulum14
An-bad moly ung
purfd pron22
Flopodum-lk bundl
from dndr nwork24
Flopoda-lk proruon on gan l48Pron ranpor
whn Golg ak11
Bpolar pndl n Xenopus laevis
xra20
Hybrd mmunologal ynap31
Baral plamd
DNA grgaon27
Bohmal ronuon on a molular-lngh al (rd) and llular ronuon on a mromr al (blak).
Flamnng mpraur n muan Z
(FZ) alon found o au onron50
trallng wa of MnD and Mne33
Dynamn alon found o buffn for mmbran fon35
escRtiii (ndoomal orng omplxrqurd for ranpor iii) found o au
mmbran buddng and on67
In an impressive feat,
the motility of dead
L. monocytogenes was
reconstituted using NPF-coated
microspheres and a minimal set
of proteins...
P e r s P e c t i v e s
646 | SEpTEMBER 2009 | VOluME 10 www.a.m/ws/mb
©2009M ill P bli h Li it d All i ht d
8/3/2019 Biology Under Construction- In Vitro Reconstitution of Cellular Function
http://slidepdf.com/reader/full/biology-under-construction-in-vitro-reconstitution-of-cellular-function 4/8
Actin ActinICAM TCR
a Microsphere b Supported lipid bilayer c Giant vesicle
NPF-coatedmicrosphere
Actin
Arp2/3
ICAM1MHC
GUV
Filopodium-likeprotrusion
Actin
LFA1
TCR
Supported lipid bilayer
T cell
5 µm 5 µm 5 µm
indicated that the formation of the immno-ogica synase rovides a mechanism forsstained signaing in engaged T ces.Hybrid nerona synases, consisting of nerona ces and a sorted iid biayer,were simiary reconstitted32. These stdiesshow that satiay organized and bioogicay reevant strctres can be formed between aiving ce and a sorted iid biayer, ro-
viding a controed environment in which tostdy ce–ce commnication.
Recenty, an eegant stdy reorted thesef-organization of MinD and MinE (ro-teins that are resonsibe for ocating thecontractie ring dring bacteria cytokinesis)into traveing waves on sorted iidbiayers33. Sorted iid biayers can asobe formed on gass microsheres insteadof on anar gass. using sorted iidbiayers on gass microsheres and rifiedroteins invoved in actin-based motiity,
the mechanism behind attachment of theNpF nera Wiskott–Adrich syndromerotein (N-WASp; aso known as WASl) toactin networks was ecidated34. Simiary, areconstitted system was recenty deveoedsing iid-coated gass microsheres tostdy dynamin-mediated membrane fission,and it was fond that dynamin and GTp aresfficient for this rocess35.
Reconstitution using giant lipid vesicles. Anaternative to the sorted iid biayer isthe giant iid vesice (FIG. 2c). Giant vesices(10–100 μm in diameter) can be comosedof a singe iid biayer (niamear) ormtie biayers (mtiamear), and they sort the diffsion of moeces in-ane,as we as morhoogica deformations ot of ane, sch as the formation of rotrsionsor invaginations. Athogh sma vesiceshave been sed in many biochemica recon-stittions in which a membrane sbstrate isreqired36,37, they are not amenabe to investi-gations of satia organization by ight micro-scoy. By contrast, giant vesices are sitabefor rea-time imaging to directy observe thedynamics and organization of biomoece–
membrane interactions. This aroach isimited, however, by the chaenge of makingarge nmbers of giant vesices with a definedsize and comosition.
Decades ago, thin-section eectron micro-scoy reveaed a abyrinth of membranenetworks in ces38, and reconstittion std-ies are now heing s to nderstand howthey might be formed. Tbar membranenetworks reminiscent of the Gogi and ERwere reconstitted sing a trncated kinesinmotor from Drosophila melanogaster that wasbond to giant niamear vesices (GuVs)
on one end and to srface immobiizedmicrotbes on the other end39,40. processivekinesin moeces were attached to GuVsthrogh a biotin–stretavidin interactionand ‘waked’ aong stabiized microtbesto generate tbar networks. Motor-drivenmembrane tbe networks have aso been
sed to dissect the mechanism of dynamin-mediated membrane fission39.
The deformabiity of iid biayer vesicesmakes them an idea too for ecidating thebiohysica effects of force generation by actin41,42 and other cytoskeeta oymers.For exame, membrane deformations thatare deendent on the oymerization of microtbes and actin fiaments encas-ated in iid vesices have been demonstratedexerimentay 43,44. Actin shes that arereminiscent of the actin cortex have aso beenreconstitted on the inner side of GuVs45.
The effect of both membrane fidity anddeformabiity on the assemby of cytoskeetanetworks can be stdied directy with GuVs.When hase-searated GuVs that containedhoshatidyinosito-4,5-bishoshate(ptdIns(4,5)p
2) were mixed with rified
roteins that are invoved in dendritic actin
network assemby, the actin networks grewexterna to the GuVs and ony on mem-brane domains that were enriched withptdIns(4,5)p
2(REF. 46). It was fond that
the interaction between actin networksand the membrane acted to stabiize theptdIns(4,5)p
2-containing membrane
domains against temeratre-indced mix-ing with other iids. The otentia infenceof an actin network on the satia organ-ization of membrane comonents sortsthe reqirement of an intact cytoskeeton insignaing systems47.
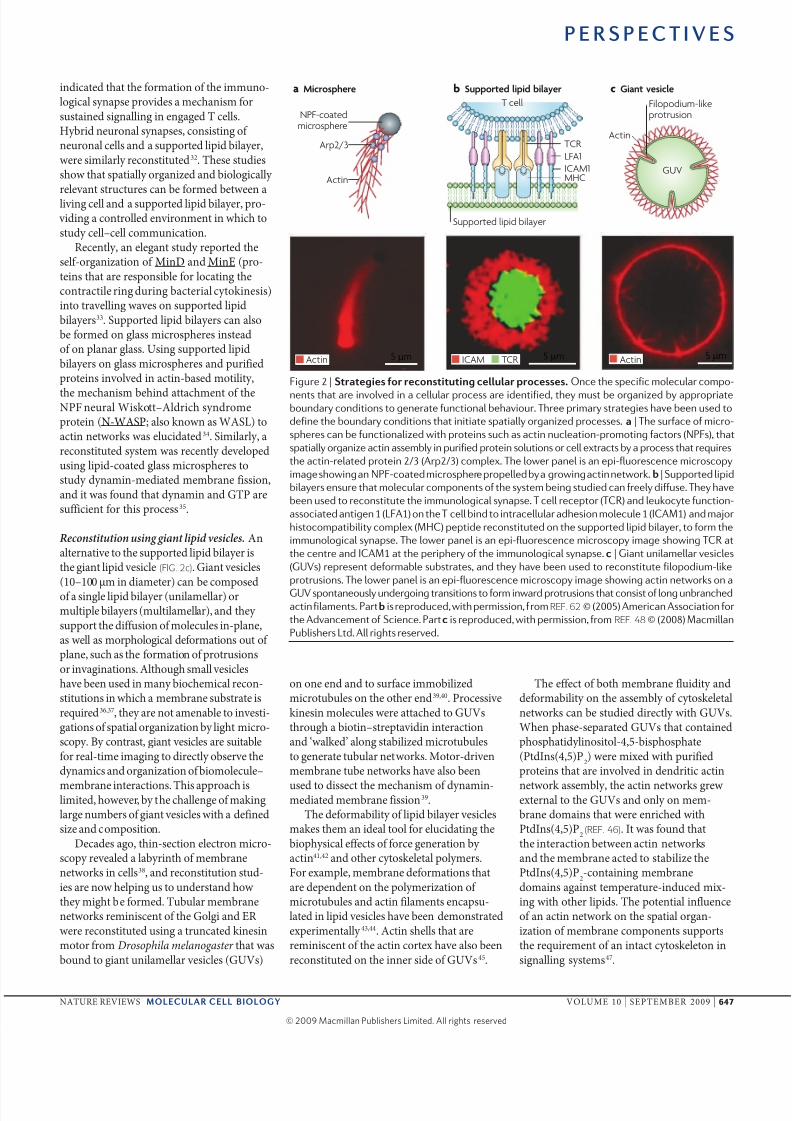
Fgur 2 | Sas s a sss. On h pf molular ompo-
nn ha ar nold n a llular pro ar dnfd, hy mu b organzd by appropraboundary ondon o gnra funonal bhaour. thr prmary rag ha bn ud o
dfn h boundary ondon ha na paally organzd pro. a | th urfa of mro-
phr an b funonalzd wh pron uh a an nulaon-promong faor (NPF), ha
paally organz an ambly n purfd pron oluon or ll xra by a pro ha rqur
h an-rlad pron 2/3 (Arp2/3) omplx. th lowr panl an p-fluorn mroopy
mag howng an NPF-oad mrophr proplld by a growng an nwork.b | suppord lpd
blayr nur ha molular omponn of h ym bng udd an frly dffu. thy ha
bn ud o ronu h mmunologal ynap. t ll rpor (tcR) and lukoy funon-
aoad angn 1 (LFA1) on h t ll bnd o nrallular adhon molul 1 (icAM1) and major
hoompably omplx (MHc) ppd ronud on h uppord lpd blayr, o form h
mmunologal ynap. th lowr panl an p-fluorn mroopy mag howng tcR a
h nr and icAM1 a h prphry of h mmunologal ynap. | Gan unlamllar l
(GUv) rprn dformabl ubra, and hy ha bn ud o ronu flopodum-lk
proruon. th lowr panl an p-fluorn mroopy mag howng an nwork on a
GUv ponanouly undrgong ranon o form nward proruon ha on of long unbranhdan flamn. Parb rprodud, wh prmon, fromREF. 62 © (2005) Amran Aoaon for
h Adanmn of sn. Par rprodud, wh prmon, from REF. 48 © (2008) Mamllan
Publhr Ld. All rgh rrd.
P e r s P e c t i v e s
NATuRE REVIEWS | MoleculAr cell Biology VOluME 10 | SEpTEMBER 2009 | 647
©2009M ill P bli h Li it d All i ht d

8/3/2019 Biology Under Construction- In Vitro Reconstitution of Cellular Function
http://slidepdf.com/reader/full/biology-under-construction-in-vitro-reconstitution-of-cellular-function 5/8
More recenty, dynamic fioodia-ikemembrane deformations were reconstittedon GuVs48. It was reviosy thoght thatfioodia emerge from branched dendriticactin networks by a rocess that invovesbnding of actin fiaments by accessory roteins. Frthermore, the ength of cearfioodia was thoght to be imited by membrane resistance force, with fioodia
exected to bcke if they grew ast thetheoretica Eer bcking ength — where astrctra eement can no onger withstandcomressive stress. The reconstittion of fioodim-ike strctres in the absence of both the ti comex (a moecar assemby that romotes actin fiament eongation)and the crossinking rotein fascin ed totwo hysica insights: the membrane aonecan faciitate the transition of a branchedactin network to a arae array of actinfiaments, and the force generated by mem-brane tension wi not bcke fioodia, even
those beyond the Eer bcking ength49.The reconstittion of comex actin-basedstrctres throgh interactions with a iidbiayer sggests that in eary cear ife,iosome-encasating force-generatingoymers cod have exhibited rich morho-ogica behaviors that are reminiscent of crrent iving systems.
last year, the se of mtiamear vesices
to stdy the bacteria tbin homooge,fiamenting temeratre sensitive mtant Z(FtsZ), reveaed that membrane-anchoredFtsZ aone can sef-assembe into ring-ikestrctres known as Z rings, which serve asscaffods for the bacteria division aara-ts50. The reconstitted Z rings generateda constriction force that was deendent onGTp hydroysis. However, becase constric-tion of the mtiamear vesices by theZ rings was incomete, it remains ossibethat additiona force-generating activity might be reqired to comete the rocess.
Tools for cellular recosttuto
Contined rogress in cear reconstittionwi deend on the creation of new toos.Recenty, techniqes that enabe efficientencasation of comonents inside ce-sized vesices have been deveoed, makingit ossibe to begin controing both internaand externa biochemica conditions forstdies invoving transmembrane signaingand transort (BOX 1). Micro-fabricationand atterning toos wi aso contineto advance, enabing the contro of moec-ar ocation for activity gradients orthree-dimensiona confinement51,52.
Centra to the stdy of satia organizationin ces is the widesread se of ight micro-scoy. Traditiona biochemica reconstittiontechniqes tyicay reort the average behav-ior of the system and yied itte informationon satia organization. By contrast, coordi-nated cear rocesses that are deendent on
the coective motion of many moeces oftenreqire microscoy to foow their dynamicevotion in rea time. Commercia wide-fiedand confoca ight microscoes have becomemainstream toos in bioogy aboratories, andresearchers contine to sh ight microscoy to higher satia and temora resotions.Severa techniqes that break the diffraction-imited satia resotion of conventiona ightmicroscoes have been demonstrated, andtheir ses, as we as crrent deveoments inforescent robe technoogy, have recenty been reviewed53.
Comtationa toos have been instr-menta in giding exerimenta aroachesto cear reconstittion as we as innderstanding the comex behaviorthey create. Mathematica modeing andcomter simations wi contine to becrcia for exaining the observations fromin vitro reconstittion exeriments16,18,45,54,and we exect their imortance to growas both exerimenta and comtationatechniqes imrove.
Learg by buldg
Ce bioogy has come a ong way since
Szent-Gyoergyi and Strab’s biochemicareconstittion of msce actomyosin andArthr Kornberg’s biochemica reconstittionof DNA synthesis more than haf a centry ago. Athogh the knowedge of an enzyme’sbinding affinity and cataytic efficiency givess sfficient information to redict its trn-over caabiity, we do not yet have the owerto redict how a mixtre of macromoeceswi assembe into satiay comex strc-tres, even with a thorogh characterizationof the individa arts. Cear reconstittionoffers a way to earn by biding.
tabl 1 | Summary of recet rogress cellular recosttuto
rs ss s
cms s rs
Microspheres
cll moly Listeria monocytogenes and NPF-oadmrophr mmrd n Xenopus laevis xra
63,64
NPF-oad mrophr, an, profln, ofln,Arp2/3 omplx and appng pron 22,23
NPF-oad mrophr, an, Arp2/3 omplxand fan
25,24
Mo pndl DNA-oad mrophr and X. laevis yoplam xra
20,65
Plamd grgaon ParC oad mrophr, ParR and ParM 27
Supported bilayers
immunologal ynap suppord blayr onanng MHc ppd andicAM1, and t ll
31,62
Nuronal ynap suppord blayr onanng nurolgn 1, andnuronal ll
32
Baral ll don MnD and Mne 33
vl fon Dynamn 1 and GtP 35
cll moly Lpd-oad bad, N-WAsP, an, profln,ofln, Arp2/3 omplx and appng pron
34
Giant vesicles
Baral ll don Mmbran-argd FZ 50
Mmbran ub nwork Knn-bound GUv and mroubul rak 40,66
An–mmbran nraon Pdin(4,5)P2-onanng l, an, N-WAsP
and Arp2/3 omplx46
Flopoda formaon Pdin(4,5)P2-onanng l, an, N-WAsP
and Arp2/3 omplx48
Mmbran on vp20, snf7 and vp24 67
Arp2/3, an-rlad pron 2/3; GUv, gan unlamllar l; icAM1, nrllular adhon molul 1;
MHc, major hoompably omplx; NPF, nulaon-promoon faor; N-WAsP, nuronal-Wo–Aldrh yndrom pron (alo known a WAsL); Pdin(4,5)P
2, phophadylnool-4,5-bphopha;
snf7, uro non-frmnng pron 7; vp, auol pron orng pron.
P e r s P e c t i v e s
648 | SEpTEMBER 2009 | VOluME 10 www.a.m/ws/mb
©2009M ill P bli h Li it d All i ht d
8/3/2019 Biology Under Construction- In Vitro Reconstitution of Cellular Function
http://slidepdf.com/reader/full/biology-under-construction-in-vitro-reconstitution-of-cellular-function 6/8
a Reverse emulsion
Emulsion
Centrifugation
Bilayer vesicle
b Double emulsion
Aqueous (internal)
Aqueous (external)
Organic
c High-speed liquid jet
Planar lipid bilayer
Vesicle
Organic
High-speedliquid jet
Fluid stream
Aqueous(internal)
Aqueous(external)
Advances in imaging technoogy, com-bined with cever exerimenta techniqes forcontroing bondary conditions, have ed tosccessf reconstittion of severa sef-organ-izing bioogica rocesses, with more on theway. However, becase cear reconstittionincdes ony a sbset of comonents sed by ces, the aroach is not garanteed to yiedthe recise mechanistic detais of a bioogicarocess. So how can we know whether areconstittion is behaving ‘bioogicay’?
perhas one day we wi need the ceareqivaent of a Tring test — the aroachdevised by Aan Tring in 1950 to test acomting machine’s abiity to demonstrateinteigence — to te the difference, askingthe qestion: does the reconstittion behaveike a norma ce in every way? unti then, wewi need to se modeing to bridge the gabetween sma assembies and ces, and wewi need to se cear reconstittion to com-ement to-down aroaches, a whie deib-eratey eeing away the eves of comexity and regation that are at work in ive ces.
The road ma for cear reconstittionwi robaby incde exeriments that aremotivated by basic science as we as thosethat are aimed at creating bioogica systemsthat exhibit cstomized fnctiona behavior— a kind of bottom- synthetic bioogy.It is conceivabe that one day or abiity toreconstitte basic modes of cear ife wiead s to create a growing and dividing sys-tem that can be comtationay designed tocraw, swim or fight disease. Now that wod
be inteigent design.Daniel A. Fletcher is at the Biophysics Group
and Bioengineering Department,
University of California, Berkeley, California 94702,
USA and the Physical Biosciences Division,
Lawrence Berkeley National Laboratory, Berkeley,
California 94720, USA.
Allen P. Liu is at the Department of Cell Biology,
The Scripps Research Institute, 10550 North Torrey
Pines Road, MB‑6, La Jolla, California 92037, USA.
Correspondence to D.A.F.
e‑mail: [email protected]
doi:10.1038/nrm2746
Published online 12 August 2009
1. Groves, J. T. Bending mechanics and molecular
organization in biological membranes. Annu. Rev.
Phys. Chem. 58, 697–717 (2007).
2. Garcia-Saez, A. J., Chiantia, S. & Schwille, P. Effect of
line tension on the lateral organization of lipid
membranes. J. Biol. Chem. 282, 33537–33544
(2007).
3. Karsenti, E. Self-organization in cell biology: a brief
history. Nature Rev. Mol. Cell Biol. 9, 255–262
(2008).
4. Kron, S. J. & Spudich, J. A. Fluorescent actin filaments
move on myosin fixed to a glass surface. Proc. Natl Acad. Sci. USA 83, 6272–6276 (1986).
5. Hawking, S. W. The Universe in a Nutshell (Bantam,
New York, 2001).
6. Kritikou, E. To see them contract for the first time.
Nature Rev. Mol. Cell Biol. 9, S6 (2008).
7. Watson, J. D. & Crick, F. H. Molecular structure of
nucleic acids; a structure for deoxyribose nucleic acid.
Nature 171, 737–738 (1953).
8. Kornberg, A. Biologic synthesis of deoxyribonucleic
acid. Science 131, 1503–1508 (1960).
9. Lehman, I. R., Bessman, M. J., Simms, E. S. &
Kornberg, A. Enzymatic synthesis of deoxyribonucleic
acid. I. Preparation of substrates and partial
purification of an enzyme from Escherichia coli .
J. Biol. Chem. 233, 163–170 (1958).
10. Schekman, R. Protein localization and membrane
traffic in yeast. Annu. Rev. Cell Biol. 1, 115–143
(1985).
11. Orci, L., Glick, B. S. & Rothman, J. E. A new type of
coated vesicular carrier that appears not to contain
clathrin: its possible role in protein transport within
the Golgi stack. Cell 46, 171–184 (1986).
12. Barlowe, C. et al. COPII: a membrane coat formed by
Sec proteins that drive vesicle budding from the
endoplasmic reticulum. Cell 77, 895–907 (1994).
13. Higgins, N. P., Peebles, C. L., Sugino, A. &
Cozzarelli, N. R. Purification of subunits of Escherichia
coli DNA gyrase and reconstitution of enzymatic
activity. Proc. Natl Acad. Sci. USA 75, 1773–1777
(1978).
14. Gorlich, D., Hartmann, E., Prehn, S. & Rapoport, T. A.
A protein of the endoplasmic reticulum involved early
in polypeptide translocation. Nature 357, 47–52
(1992).
15. Nakajima, M. et al. Reconstitution of circadian
oscillation of cyanobacterial KaiC phosphorylation
in vitro. Science 308, 414–415 (2005).
16. Surrey, T., Nedelec, F., Leibler, S. & Karsenti, E.
Physical properties determining self-organization of
motors and microtubules. Science 292, 1167–1171
(2001).17. Haviv, L. et al. Reconstitution of the transition from
lamellipodium to filopodium in a membrane-free
system. Proc. Natl Acad. Sci. USA 103, 4906–4911
(2006).18. Karsenti, E., Nedelec, F. & Surrey, T. Modelling
microtubule patterns. Nature Cell Biol. 8, 1204–1211
(2006).
19. Pollard, T. D. The cytoskeleton, cellular motility and the
reductionist agenda. Nature 422, 741–745 (2003).
20. Heald, R. et al. Self-organization of microtubules into
bipolar spindles around artificial chromosomes in
Xenopus egg extracts. Nature 382, 420–425 (1996).
21. Pistor, S., Chakraborty, T., Niebuhr, K., Domann, E. &
Wehland, J. The ActA protein of Listeria
monocytogenes acts as a nucleator inducing
reorganization of the actin cytoskeleton. EMBO J. 13,
758–763 (1994).
22. Loisel, T. P., Boujemaa, R., Pantaloni, D. & Carlier, M. F.
Reconstitution of actin-based motility of Listeria and
Shigella using pure proteins. Nature 401, 613–616(1999).
23. Akin, O. & Mullins, R. D. Capping protein increases
the rate of actin-based motility by promoting filament
nucleation by the Arp2/3 complex. Cell 133,
841–851 (2008).
24. Vignjevic, D. et al. Formation of filopodia-like bundles
in vitro from a dendritic network. J. Cell Biol. 160,
951–962 (2003).25. Brieher, W. M., Coughlin, M. & Mitchison, T. J.
Fascin-mediated propulsion of Listeria monocytogenes
independent of frequent nucleation by the Arp2/3
complex. J. Cell Biol. 165, 233–242 (2004).
26. Shih, Y. L. & Rothfield, L. The bacterial cytoskeleton.
Microbiol. Mol. Biol. Rev. 70, 729–754 (2006).
27. Garner, E. C., Campbell, C. S., Weibel, D. B. &
Mullins, R. D. Reconstitution of DNA segregation
driven by assembly of a prokaryotic actin homolog.
Science 315, 1270–1274 (2007).
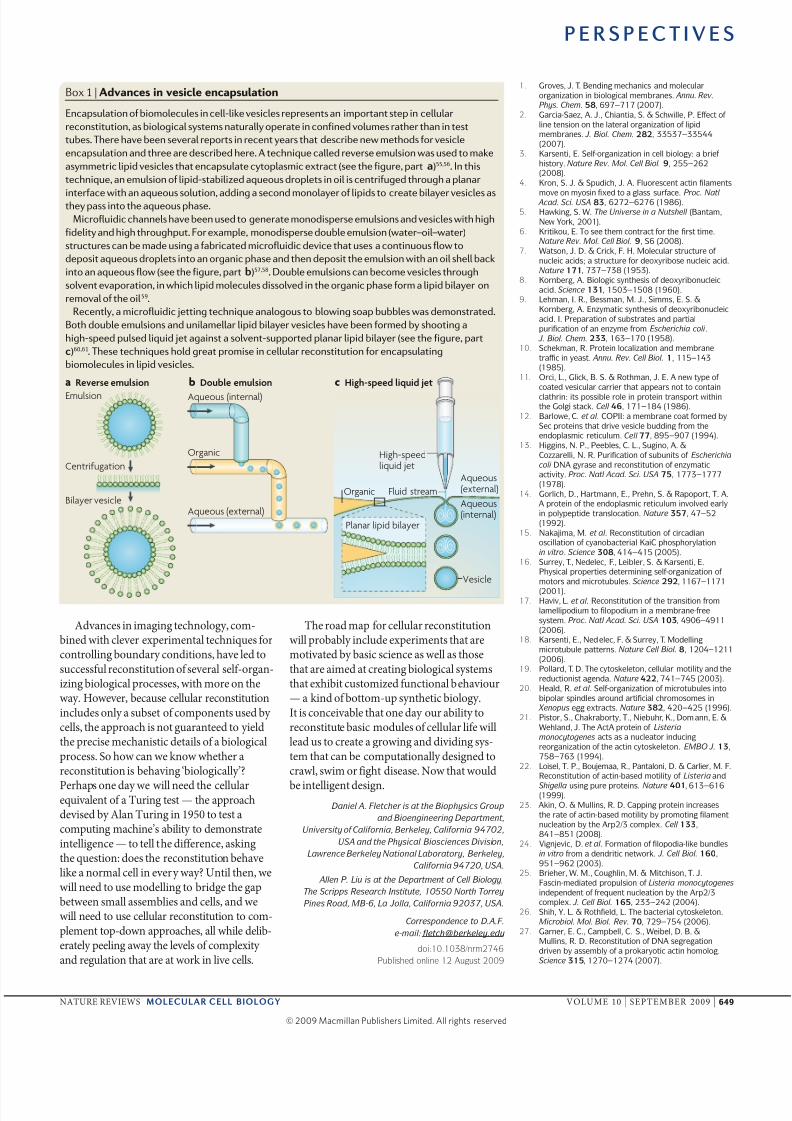
Box 1 | Advaces vescle ecasulato
Encapsulation of biomolecules in cell-like vesicles represents an important step in cellular
reconstitution, as biological systems naturally operate in confined volumes rather than in test
tubes. There have been several reports in recent years that describe new methods for vesicle
encapsulation and three are described here. A technique called reverse emulsion was used to make
asymmetric lipid vesicles that encapsulate cytoplasmic extract (see the figure, part a)55,56. In this
technique, an emulsion of lipid-stabilized aqueous droplets in oil is centrifuged through a planar
interface with an aqueous solution, adding a second monolayer of lipids to create bilayer vesicles asthey pass into the aqueous phase.
Microfluidic channels have been used to generate monodisperse emulsions and vesicles with high
fidelity and high throughput. For example, monodisperse double emulsion (water–oil–water)
structures can be made using a fabricated microfluidic device that uses a continuous flow to
deposit aqueous droplets into an organic phase and then deposit the emulsion with an oil shell back
into an aqueous flow (see the figure, part b)57,58. Double emulsions can become vesicles through
solvent evaporation, in which lipid molecules dissolved in the organic phase form a lipid bilayer on
removal of the oil59.
Recently, a microfluidic jetting technique analogous to blowing soap bubbles was demonstrated.
Both double emulsions and unilamellar lipid bilayer vesicles have been formed by shooting a
high-speed pulsed liquid jet against a solvent-supported planar lipid bilayer (see the figure, part
)60,61. These techniques hold great promise in cellular reconstitution for encapsulating
biomolecules in lipid vesicles.
P e r s P e c t i v e s
NATuRE REVIEWS | MoleculAr cell Biology VOluME 10 | SEpTEMBER 2009 | 649
©2009M ill P bli h Li it d All i ht d

8/3/2019 Biology Under Construction- In Vitro Reconstitution of Cellular Function
http://slidepdf.com/reader/full/biology-under-construction-in-vitro-reconstitution-of-cellular-function 7/8
28. Garner, E. C., Campbell, C. S. & Mullins, R. D.
Dynamic instability in a DNA-segregating
prokaryotic actin homolog. Science 306, 1021–1025
(2004).
29. Sackmann, E. & Tanaka, M. Supported membranes on
soft polymer cushions: fabrication, characterization
and applications. Trends Biotechnol. 18, 58–64
(2000).
30. Groves, J. T., Wulfing, C. & Boxer, S. G. Electrical
manipulation of glycan-phosphatidyl inositol-tethered
proteins in planar supported bilayers. Biophys. J. 71,
2716–2723 (1996).31. Grakoui, A. et al. The immunological synapse: a
molecular machine controlling T cell activation.
Science 285, 221–227 (1999).
32. Pautot, S., Lee, H., Isacoff, E. Y. & Groves, J. T.
Neuronal synapse interaction reconstituted between
live cells and supported lipid bilayers. Nature Chem.
Biol. 1, 283–289 (2005).
33. Loose, M., Fischer-Friedrich, E., Ries, J., Kruse, K. &
Schwille, P. Spatial regulators for bacterial cell division
self-organize into surface waves in vitro. Science 320,
789–792 (2008).
34. Co, C., Wong, D. T., Gierke, S., Chang, V. & Taunton, J.
Mechanism of actin network attachment to moving
membranes: barbed end capture by N-WASP WH2
domains. Cell 128, 901–913 (2007).
35. Pucadyil, T. J. & Schmid, S. L. Real-time visualization
of dynamin-catalyzed membrane fission and vesicle
release. Cell 135, 1263–1275 (2008).
36. Takei, K. et al. Generation of coated intermediates of
clathrin-mediated endocytosis on protein-free
liposomes. Cell 94, 131–141 (1998).37. Matsuoka, K. et al. COPII-coated vesicle formation
reconstituted with purified coat proteins and
chemically defined liposomes. Cell 93, 263–275
(1998).
38. Palade, G. E. The organization of living matter.
Proc. Natl Acad. Sci. USA 52, 613–634 (1964).
39. Roux., A., Uyhazi, K., Frost, A. & De Camilli, P.
GTP-dependent twisting of dynamin implicates
constriction and tension in membrane fission. Nature
441, 528–531 (2006).
40. Koster, G., VanDuijn, M., Hofs, B. & Dogterom, M.
Membrane tube formation from giant vesicles by
dynamic association of motor proteins. Proc. Natl
Acad. Sci. USA 100, 15583–15588 (2003).41. Giardini, P. A., Fletcher, D. A. & Theriot, J. A.
Compression forces generated by actin comet tails
on lipid vesicles. Proc. Natl Acad. Sci. USA 100,
6493–6498 (2003).
42. Upadhyaya, A. & van Oudenaarden, A. Biomimetic
systems for studying actin-based motility. Curr. Biol. 13, R734–R744 (2003).
43. Cortese, J. D., Schwab, B., Frieden, C. & Elson, E. L.
Actin polymerization induces a shape change in actin-
containing vesicles. Proc. Natl Acad. Sci. USA 86,
5773–5777 (1989).
44. Fygenson, D. K., Marko, J. F. & Libchaber, A.
Mechanics of microtubule-based membrane
extension. Phys. Rev. Lett. 79, 4497 (1997).
45. Pontani, L. L. et al. Reconstitution of an actin cortex
inside a liposome. Biophys. J. 96, 192–198
(2009).
46. Liu, A. P. & Fletcher, D. A. Actin polymerization serves
as a membrane domain switch in model lipid bilayers.Biophys. J. 91, 4064–4070 (2006).
47. Holowka, D., Sheets, E. D. & Baird, B. Interactions
between FcεRI and lipid raft components are regulated
by the actin cytoskeleton. J. Cell Sci. 113, 1009–1019
(2000).
48. Liu, A. P. et al. Membrane-induced bundling of actin
filaments. Nature Phys. 4, 789–793 (2008).
49. Pronk, S., Geissler, P. L. & Fletcher, D. A. Limits of
filopodium stability. Phys. Rev. Lett. 100, 258102
(2008).
50. Osawa, M., Anderson, D. E. & Erickson, H. P.
Reconstitution of contractile FtsZ rings in liposomes.
Science 320, 792–794 (2008).
51. Doh, J. & Irvine, D. J. Immunological synapse arrays:
patterned protein surfaces that modulate
immunological synapse structure formation in
T cells. Proc. Natl Acad. Sci. USA 103, 5700–5705
(2006).
52. Cosentino Lagomarsino, M. et al. Microtubule
organization in three-dimensional confined
geometries: evaluating the role of elasticity through a
combined in vitro and modeling approach. Biophys. J.
92, 1046–1057 (2007).
53. Fernandez-Suarez, M. & Ting, A. Y. Fluorescent probes
for super-resolution imaging in living cells. Nature Rev.
Mol. Cell Biol. 9, 929–943 (2008).
54. Alberts, J. B. & Odell, G. M. In silico reconstitution of
Listeria propulsion exhibits nano-saltation. PLoS Biol.
2, e412 (2004).
55. Pautot, S., Frisken, B. J. & Weitz, D. A. Engineering
asymmetric vesicles. Proc. Natl Acad. Sci. USA 100,
10718–10721 (2003).
56. Noireaux, V. & Libchaber, A. A vesicle bioreactor as a
step toward an artificial cell assembly. Proc. Natl
Acad. Sci. USA 101, 17669–17674 (2004).57. Okushima, S., Nisisako, T., Torii, T. & Higuchi, T.
Controlled production of monodisperse double
emulsions by two-step droplet breakup in microfluidic
devices. Langmuir 20, 9905–9908 (2004).
58. Utada, A. S. et al. Monodisperse double emulsions
generated from a microcapillary device. Science 308,537–541 (2005).
59. Shum, H. C., Kim, J. W. & Weitz, D. A. Microfluidic
fabrication of monodisperse biocompatible and
biodegradable polymersomes with controlled
permeability. J. Am. Chem. Soc. 130, 9543–9549
(2008).
60. Funakoshi, K., Suzuki, H. & Takeuchi, S. Formation of
giant lipid vesicle-like compartments from a planar
lipid membrane by a pulsed jet flow. J. Am. Chem. Soc.
129, 12608–12609 (2007).61. Stachowiak, J. C. et al. Unilamellar vesicle formation
and encapsulation by microfluidic jetting. Proc. Natl
Acad. Sci. USA 105, 4697–4702 (2008).62. Mossman, K. D., Campi, G., Groves, J. T. & Dustin, M. L.
Altered TCR signaling from geometrically repatterned
immunological synapses. Science 310, 1191–1193
(2005).63. Theriot, J. A., Mitchison, T. J., Tilney, L. G. &
Portnoy, D. A. The rate of actin-based motility of
intracellular Listeria monocytogenes equals the rate of
actin polymerization. Nature 357, 257–260 (1992).
64. Cameron, L. A., Footer, M. J., van Oudenaarden, A. &
Theriot, J. A. Motility of ActA protein-coated
microspheres driven by actin polymerization.
Proc. Natl Acad. Sci. USA 96, 4908–4913 (1999).
65. Gaetz, J., Gueroui, Z., Libchaber, A. & Kapoor, T. M.
Examining how the spatial organization of chromatin
signals influences metaphase spindle assembly.
Nature Cell Biol. 8, 924–932 (2006).
66. Roux., A. et al. A minimal system allowing tubulation
with molecular motors pulling on giant liposomes.
Proc. Natl Acad. Sci. USA 99, 5394–5399 (2002).
67. Wollert, T., Wunder, C., Lippincott-Schwartz, J. &
Hurley, J. H. Membrane scission by the ESCRT-III
complex. Nature 458, 172–177 (2009).
AcknowledgementsWe apologize to those colleagues whose original and impor-
tant work could not be cited owing to space limitations.
We thank D. Richmond, J. Stachowiak and the rest of the
Fletcher laboratory, as well as T. Pucadyil, for helpful discus-
sion. A.P.L. is supported by the Natural Sciences and
Engineering Research Council of Canada. D.A.F. is funded by
National Institutes of Health R01 grants and a Nanomedicine
Development Centre grant.
DATABASESUnPoKB: hp://www.unpro.org
FZ | icAM1 | MnD | Mne | N-WAsP
FURTHER inFORMATiOnDanl A. Flh’ hompag: hp://flhlab.brkly.du/
All linkS Are Active in the online pdf
P e r s P e c t i v e s
650 | SEpTEMBER 2009 | VOluME 10 www.a.m/ws/mb
©2009M ill P bli h Li it d All i ht d

8/3/2019 Biology Under Construction- In Vitro Reconstitution of Cellular Function
http://slidepdf.com/reader/full/biology-under-construction-in-vitro-reconstitution-of-cellular-function 8/8




























