Embed Size (px)
Citation preview

BB44CH08-Schneider ARI 28 May 2015 15:47
Biophysics ofChannelrhodopsinFranziska Schneider,∗,# Christiane Grimm,and Peter HegemannExperimental Biophysics, Institute of Biology, Humboldt-Universitat zu Berlin, 10115 Berlin,Germany; email: [email protected], [email protected], [email protected]
Annu. Rev. Biophys. 2015. 44:167–86
The Annual Review of Biophysics is online atbiophys.annualreviews.org
This article’s doi:10.1146/annurev-biophys-060414-034014
Copyright c© 2015 by Annual Reviews.All rights reserved
∗Corresponding author
#Current address: Cardiac Biophysics and SystemsBiology Group, National Heart and LungInstitute, Imperial College London, UB9 6JH,United Kingdom
Keywords
ion selectivity, photocycle, voltage dependence, color-tuning,channelrhodopsin-activation model
Abstract
Channelrhodopsins (ChRs) are directly light-gated ion channels that func-tion as sensory photoreceptors in flagellated green algae, allowing these algaeto identify optimal light conditions for growth. In neuroscience, ChRs con-stitute the most versatile tools for the light-induced activation of selectedcells or cell types with unprecedented precision in time and space. In recentyears, many ChR variants have been discovered or engineered, and countlesselectrical and spectroscopic studies of these ChRs have been carried out, bothin host cells and on purified recombinant proteins. With significant supportfrom a high-resolution 3D structure and from molecular dynamics calcu-lations, scientists are now able to develop models that conclusively explainChR activation and ion conductance on the basis of chromophore isomeriza-tion, structural changes, proton transfer reactions, and water rearrangementon timescales ranging from femtoseconds to minutes.
167
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44CH08-Schneider ARI 28 May 2015 15:47
Photocurrents:electric currentsconducted by ChRsupon light activation
Contents
INTRODUCTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 168Channelrhodopsins in Their Native Environment . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 168Channelrhodopsins in Optogenetics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 168Channelrhodopsin Variants . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 169
CHANNELRHODOPSIN STRUCTURE . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 169Retinal-Binding Pocket and Active Site Complex . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 169Ion Permeation Pathway . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 171
CHANNELRHODOPSIN PHOTOACTIVATION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 172Photocycle Models and Retinal Isomerizations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 172Color Tuning in Channelrhodopsins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 174
CHANNELRHODOPSIN PHOTOCURRENT PROPERTIES. . . . . . . . . . . . . . . . . . . 176Electrophysiological Reaction Schemes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 176Ion Selectivity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 177Putative Selectivity Filter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 177Voltage Dependence . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 179
THE HELIX 2 (H2)-TILT MODEL FOR CHANNELRHODOPSINACTIVATION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 180
APPLICATIONS OF CHANNELRHODOPSINS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 181
INTRODUCTION
Channelrhodopsins in Their Native Environment
Channelrhodopsins (ChRs) are light-gated cation channels that serve as sensory photoreceptors inmotile green algae (47, 48, 68, 72). In these algae, ChRs are located at the surface of a specializedcellular compartment called the eyespot. This organelle consists of (a) an optical device comprisingtwo or more layers of pigmented granules that function as quarter-wave stack antennae and providedirectional sensitivity and (b) a specialized overlying plasma membrane that contains the ChRs andother proteins of the sensory machinery (15, 22). Upon photon absorption, ChRs mediate fast,proton- and cation-driven inward currents, resulting in plasma membrane depolarization (47, 48).At low light intensities, the electrical signal is most probably amplified by secondary voltage-gatedchannels, such as voltage-gated Ca2+ channels, whereas at high light intensities, the photocurrentis mainly mediated by ChRs directly (30). The local increase in cytosolic Ca2+ concentration istransmitted to the flagella base (basal bodies) by an unknown signaling cascade. The direction-modulated signal allows the alga to adjust the plane, frequency, and three-dimensional pattern ofits flagellar beating, thereby enabling the alga to exhibit phototactic and photophobic responses(22, 24).
Channelrhodopsins in Optogenetics
After the initial characterization of ChRs as directly light-gated ion channels in 2002 and 2003(47, 48), the unique combination of both photosensor and channel within a single, relatively smallprotein has drawn attention from not only algae researchers but also a larger scientific communityof cell biologists and neuroscientists. Since their initial functional expression in human embryonic
168 Schneider · Grimm · Hegemann
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44CH08-Schneider ARI 28 May 2015 15:47
kidney (HEK) cells (48), ChRs have been successfully expressed in a large variety of vertebratecells to depolarize the plasma membrane following light application. In neurons, ChR-mediatedcation currents were shown to be sufficient for eliciting action potentials in a spatiotemporallydefined manner (10, 27). This finding was another milestone in the rapidly developing field ofoptogenetics. Thus far, ChRs have been applied to induce synaptic plasticity (84), unravel neuronalconnectivity (54), and manipulate behavior in ChR-expressing animals including worms, flies,chickens, zebrafish, and rodents (1, 11, 37, 46, 65). Potential clinical applications of ChRs includerestoration of vision (9), repair of hearing impairment (66), and treatment of neural disorders suchas Parkinson’s disease (19).
Channelrhodopsin Variants
The first identified ChRs were two isoforms from Chlamydomonas reinhardtii (ChR1 and ChR2)(47, 48, 68, 72) and the two corresponding ChRs from the colonial alga Volvox carteri (VChR1and VChR2) (14, 32, 82). Owing to its superior expression in host cells, ChR2 is currently notonly the best-studied ChR but also the most commonly used optogenetic tool for membranedepolarization. Subsequently, a number of ChR genes were discovered in related chlorophyceanalgae, including Mesostigma viride (MChR), Chlamydomonas augustae (CaChR1), Chlamydomonasyellowstonensis, Dunaliella salina (DChR), Pleodorina starii, Pyramimonas gelidicola, and Platymonassubcordiformis (PsChR) (17, 18, 25, 83). Very recently, 61 additional ChR homologs were identifiedfrom a large genomic screen and were tested for photocurrent properties in selected cell sys-tems (33). The two ChRs originating from Stigeoclonium helveticum (ShChR) and Chlamydomonasnoctigama (CnChR1)—referred to as Chronos and Chrimson, respectively—exhibit highly distinctcharacteristics (see the sections titled “Color Tuning in Channelrhodopsins” and “Voltage Depen-dence” for details), rendering them potent candidates for optogenetic applications (33). Today,sequence comparison in combination with structural modeling facilitates our understanding ofthe structural and functional dynamics of the different ChRs and enables the design of new ChRsadapted to specific research questions.
CHANNELRHODOPSIN STRUCTURE
ChRs consist of an N-terminal membrane-spanning domain followed by a C-terminal intracellulardomain that has been proposed to be relevant for ChR targeting and protein–protein interactionsin the alga (43). The N-terminal module comprises seven transmembrane helices (H1–H7) anda retinal chromophore, and is sufficient for light-gated channel activity (47, 48). As is alreadyapparent in the low-resolution (6 A) structure from electron microscopy, ChR2 forms dimers inwhich H3 and H4 are located at the dimer interface (44). High-resolution X-ray crystallographyon a ChR1/ChR2 chimera (C1C2) unveiled protomer linkage via three disulfide bridges at theN-terminal extracellular end (31). This linkage, however, has little or no relevance for ion channelfunctionality (62).
Retinal-Binding Pocket and Active Site Complex
As in all microbial rhodopsins, the retinal chromophore is bound to a conserved lysine residue inH7, the corresponding residue to K2571 in ChR2, via a protonated retinal Schiff base (RSBH+)
1Unless otherwise indicated, we use the ChR2 numbering of amino acids throughout this article.
www.annualreviews.org • Biophysics of Channelrhodopsin 169
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44CH08-Schneider ARI 28 May 2015 15:47
H6H7
C1C2ChR2
H5
H4H3
H1
H2
a
ccc
Extracellular side
Cytosol
Structural model of C1C2
ee S102S63S102S63
Y109Y70Y109Y70
H304H265H304H265
R307R268R307R268
H173H134H173H134
E2E2
E1E1
N297N258N297N258
E3E3
Site of Na+ accumulation
Inner gate
Central gate
Q95Q56Q95Q56
ff
dd
ee
A91S52A91S52
E4E4
E5E5
dd
ff
bb
T198T159T198T159
S194S155S194S155
D195D156D195D156C167
C128C167C128
Ion permeation pathway
Retinal binding pocket
E292E253 = Ci2E292E253 = Ci2
E162E123 = Ci1E162E123 = Ci1
H2
E129E90 = E3E129E90 = E3
E122E83 = E2E122E83 = E2
E121E82 = E1E121E82 = E1
E136E97 = E4E136E97 = E4
E140E101 = E5E140E101 = E5
K132K93 = K1K132K93 = K1
bb
cc
K296K257K296K257
G220G181G220G181
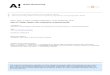
Figure 1Important structural elements of channelrhodopsins (ChRs). The structural model depicts the C1C2 chimera and is based on its crystalstructure solved by Kato et al. (31) (PDB ID: 3UG9). The model reflects dark-adapted C1C2, and thus a closed channel conformation.Helices H1–H5 originate from ChR1, and H6 and H7 originate from ChR2. The retinal is shown in lime green. (a) Overall proteinstructure with framed regions of interest; (b) retinal binding pocket; (c) ion permeation pathway; (d ) inner gate; (e) central gate; ( f ) siteof Na+ accumulation. The gray wireframes in panels c and f depict cavities within the protein in which water molecules and cations mayreside.
linkage (Figure 1a,b). The charge of the RSBH+ is stabilized by a counterion complex comprisingtwo glutamate residues, E123 and E253 [termed counterion-1 (Ci1) and counterion-2 (Ci2) inthe remainder of this article] (Figure 1b). In the C1C2 structure, distances between the RSBnitrogen and the Ci1 and Ci2 (in this case E162 and D292) carboxyl groups are almost equal (3.4and 3.0 A, respectively) (31). Moreover, theoretical calculations of pKa values propose that, in thedark, Ci1 is protonated and Ci2 is deprotonated, suggesting that Ci2 functions as a proton acceptorin C1C2 (31). The nearest water molecule is more distal (4.4 A) than the respective glutamatecarboxyl groups are, proposing a direct proton transfer from the RSBH+ to Ci2 (31), in clear
170 Schneider · Grimm · Hegemann
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44CH08-Schneider ARI 28 May 2015 15:47
Channelrhodopsin-ET-accelerated(ChETA): ChRvariants that havemutated Ci1 and thatexhibit fast,voltage-insensitivephotocycle kinetics
Photocycle:cyclic series ofphotointermediatesthat appear afterphotoexcitation of theprotein-boundchromophore
DC pair: homologousresidues to ChR2C128 and D156 thatare criticaldeterminants of thekinetics of channelopening and closing
Step-functionrhodopsins (SFRs):slow-cycling ChRmutants for whichboth on- andoff-switching aretriggered by light ofdifferent wavelengths
Retinal bindingpocket: a cavitywithin the protein inwhich the retinal isfixed by interactionwith selected aminoacids and by covalentlinkage to K257
Access channel:cavity within the C1C2structure framed bypolar residues; in MDsimulations watermolecules penetratefrom the extracellularbulk phase
contrast to the case of bacteriorhodopsin (BR), in which Ci1 and Ci2 are connected to the RSBnitrogen via water 402 (42). Accordingly, mutation of Ci2 to alanine almost completely abolishesphotocurrents, whereas mutation of Ci1 to threonine, alanine, or glutamine in ChR2 results infunctional ChRs that are widely used for fast action potential firing in neurons, owing to theiraccelerated photocycling. These ChR mutants are commonly referred to as channelrhodopsin-ET-accelerated (ChETA) mutants (7, 21). Recent spectroscopic findings and molecular dynamics(MD) simulations suggest that ChR2 E90 undergoes a side flip in the ChETA mutants, therebyat least partially replacing the Ci1 charge (36).
Another pair of residues that are highly relevant for gating and consecutive photocycle kineticscomprises C128 and D156 of ChR2; these residues are referred to as the DC pair or DC gate(Figure 1b). Individual mutation of either of these residues causes up to a ten thousand–folddeceleration in channel opening and closing [step-function rhodopsins (SFRs)] (3, 8), which inturn results in current saturation at much lower light intensities during prolonged illumination andthereby imparts increased operational light sensitivity to host cells (8). Mutations in both residuescompletely arrest the photocycle in the conducting state: Channel closing has not been observedfor the double mutant (81). The corresponding residues in BR, T90 and D115, interact withthe retinal chromophore and control H3–H4 dynamics during the photocycle (29, 42). Althoughevidence for different arrangements of the two residues in ChRs exists, the role of the DC gate inthe coupling of retinal isomeric changes to channel gating remains to be elucidated (78).
ChR2 T159 constitutes another important amino acid of the retinal binding pocket, that isin direct contact with D156 (Figure 1b). Replacement of this amino acid (T159) with cysteinecauses improved retinal binding and a threefold to tenfold increased stationary photocurrent,depending on the host system (7, 57, 74). In the presence of C159, step-function mutants show onlymoderately decelerated photocycle kinetics. Accordingly, DC pair mutations do not generate slowstep-function rhodopsins in VChR1 and MChR, as both feature a cysteine at the correspondingposition (VChR1 C154 and MChR C178, respectively) (57). T159 operates in conjunction withD156 and S155 to form an OH-cluster (hydrogen-bonding network) along H4 that is crucial forprotein dynamics following light excitation of the chromophore (Figure 1b).
Ion Permeation Pathway
The high-resolution structure of the C1C2 chimera reveals a hydrophilic pore between H1,H2, H3, and H7 that was already previously assumed to serve as the cation permeation pathwayin ChRs (31). Polar and charged residues of H2 that appear with a roughly seven–amino acidperiodicity and face toward H3 and H7 are key elements of the pore (77). In ChR2, these polarresidues include E82 (E1), E83 (E2), E90 (E3), K93 (K1), E97 (E4), and E101 (E5) (Figure 1c).Individual substitutions of E1, E2, E4, or E5 to alanine or glutamine reduce photocurrents bytwofold to sixfold, whereas multiple substitutions result in more dramatic current reduction(71, 77). Consistent with the mutant data, MD simulations detect two preferential sites for Na+
accumulation; the first site is located between E4, E5, S52, and Q56 (Figure 1f ), and the secondone is near the hydrophilic cluster composed of E1, E2, H134, H265, and R268 on the cytosolicside (Figure 1d ). Moreover, E1 and E3 are highly conserved among ChRs and constitute keydeterminants for cation selectivity (see the section titled “Putative Selectivity Filter”).
In MD simulations, the hydrophilic pore attracts water from the extracellular bulk phase; thisso-called access channel with a diameter of 8 A is framed by side chains of polar residues includingthe essential R120 and the polar E4 and E5 (55, 78). Near the RSBH+, the water distributionis discontinuous, and the channel is blocked by S63, E3, and N258, which are interconnectedby several hydrogen bonds (referred to as the central gate, see Figure 1e) (31, 78). A second
www.annualreviews.org • Biophysics of Channelrhodopsin 171
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44CH08-Schneider ARI 28 May 2015 15:47
Central gate:hydrogen-bondedChR residues thatblock the cation-permeation pathwayfrom the extracellularside; channel openingrequires reorientationof these residues
Inner gate:hydrophilic amino acidcluster that blocks thecation-permeationpathway from theintracellular side;reorientation of thiscluster might be finalstep in ChR gating
Infrared (IR)differencespectroscopy:measurement ofchanges in IR lightabsorption betweenphotocycleintermediates and darkstates that representdipole changes ofmolecule vibrations
restriction site (referred to as the inner gate, see Figure 1d ) is given by Y70 in combination withthe hydrophilic cluster of E1 and E2 and their hydrogen-bonding partners H134 and R268, whichare located on H3 and H7, respectively. The hydrogen-bonding network of both gates keeps thechannel internally closed, preventing water influx from both the cytoplasmic and extracellularbulk phases. Channel opening and the resulting cation conductance thus require conformationalrearrangements at the two gates (gating).
CHANNELRHODOPSIN PHOTOACTIVATION
Photocycle Models and Retinal Isomerizations
In microbial rhodopsins, photon absorption triggers retinal isomerization and inititates a sequenceof thermal conformational changes that finally reestablish the original dark state (this cyclic pro-cess is known as the photocycle; see Figure 2a for ChR2). The electronically excited state of thechromophore is vibrationally inactivated within 150 fs after ChR2 photoexcitation (75). Follow-ing conical intersection and conversion to the electronic ground state (which occur on a 400-fstimescale), vibrational inactivation continues in conjunction with further retinal restructuring, andthe early K-like intermediate is reached after 2.7 ps (64, 75). This early K-product seems either tobe identical to or to smoothly fade to (without color shift) the early P500 intermediate previouslyidentified by flash photolysis experiments (15). Major rearrangements of the protein backbonealready occur during the formation of the P500 intermediate (40, 48, 60, 75) and correspond tomore red-shifted photointermediates in VChR1 (P600) and CaChR (K-like, ∼P580) (15, 32, 67).The RSB is then deprotonated, yielding the blue-shifted P390 state in ChR2 (4, 15, 32, 60, 67).The primary proton acceptor is Ci1 or Ci2, depending on the ChR variant and the external condi-tions that determine the steric configuration and protonation states of the respective residues (31,36, 40, 78). The P390 state is in equilibrium with the P520 state, exhibiting a reprotonated RSB(4, 15, 60). Residue D156 was proposed to function as the proton donor (40), but this assumptionis challenged by the fact that (a) D156 is not conserved in all ChRs, and (b) the kinetics of the D156deprotonation does not parallel RSB reprotonation (36). Whereas there is general consensus aboutP520 being a conductive state, P390 is formed prior to the opening of the channel but equilibrateswith P520, and the involvement of P390 in ion conduction is still under debate (4, 40, 69). It isconceivable that the retinal conformation is changed again during P390 formation, triggering therearrangement of the inner gate and final channel gating. Thus, one could define an early P390state and a late P390′ state for ChR; these would be analogous to the M and M′ states of BR (26).The late P390′ state in equilibrium with P520 could be the first ion conducting state that ap-pears during the ChR photocycle. In ChRs with red-shifted dark states such as VChR1, C1V1, orReaChR (see the section titled “Color-Tuning in Channelrhodopsins”), the absorption spectra ofthe dark state and the conducting state (corresponding to P520) almost completely overlap. Pho-toactivation of these ChRs causes apparent chromophore bleaching without an absorption shiftduring time-resolved spectroscopy due to lower extinction coefficients of the cis-chromophores(32, 38, 67, 81). In ChR2, channel closure is accompanied by the transition from the conductingP520 state to the nonconducting P480 and P480′ states that biphasically revert to the dark state(4, 59). Major conformational reorientations occur during dark state recovery on a timescale ofseveral seconds, even in wild-type ChR2 (60).
Retinal extraction and Raman measurements indicate a mixture of retinal isoforms, even fordark-adapted (DAapp) ChR2 or C128T; the latter exhibits 22–40% 13-cis retinal and 60%–78%all-trans retinal (58, 59, 70). During continuous illumination, the ratio of 13-cis retinal to all-trans retinal increases, and minor amounts of 11-cis and 9-cis isoforms appear. Accordingly, it has
172 Schneider · Grimm · Hegemann
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44CH08-Schneider ARI 28 May 2015 15:47
I p/ I
p m
ax
Wavelength λ (nm)
TsChR*
PsChR*
ChR2
C1C2
ChR1
VChR1
ReachR
C1V1
Chrimson
*Data: Jonas Wietek
400 450 500 550 600 650
0.0
0.5
1.0
D480trans, anti
P500
P390P520
cis, anti
P480
D470cis, syn
P500'
P390'P520'trans, syn
P480'
ChR2
a Photocycle model of ChR2
b Spectral variety of ChR activation
Figure 2Channelrhodopsin (ChR) photoactivation. (a) Complex photocycle model for ChR2 implyingphotoactivation of two different dark states (D480 and D470) with distinct retinal configurations (4, 59, 60).Transition between the two cycles occurs in the presence of late P480 photointermediates. Light activationis indicated by blue arrows. (b) Palette of color-tuned ChRs. Normalized action spectra of selected ChRswere recorded in human embryonic kidney (HEK) 293 cells. The reader is referred to Table 1 for therespective action maxima of each ChR and for relevant references. Abbreviations: C1C2/ReachR/C1V1,chimeric ChRs; ChR1/ChR2, Chlamydomonas reinhardtii ChRs; Chrimson, Chlamydomonas noctigama ChR;PsChR, Platymonias subcordiformus ChR; TsChR, Tetraselmis striata ChR; VChR1, Volvox carteri ChR.
been concluded that multiple retinal isomerizations occur in parallel in ChRs. Both all-trans,15-anti retinal and 13-cis,15-syn retinal stabilize a salt bridge between the RSB and the counterioncomplex, thereby favoring closed channel conformations, but both 13-cis,15-anti retinal and all-trans,15-syn retinal may evoke formation of the conducting states (53, 59, 70). The correspondingphotocycles imply the photoconversion of all-trans,15-anti retinal to 13-cis,15-anti retinal, as wellas that of 13-cis,15-syn retinal to all-trans,15-syn retinal, whereas the transition between the twocycles was proposed to occur during population of late photocycle intermediate states (P480);however, this hypothesis has never been experimentally verified (Figure 2a) (59). The two retinalconformations of the open channel states might be linked to the two conductive states withdifferent ion selectivities, as observed in electrical measurements (see Figure 3c,d and the sectiontitled “Electrophysiological Reaction Schemes”) (6, 63).
www.annualreviews.org • Biophysics of Channelrhodopsin 173
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44CH08-Schneider ARI 28 May 2015 15:47
C
O
D C
O
D
C1 C2
O1 O2
200 ms
50 pA 70 mM Mg2+ pHe 9.0
a Current–voltage relationship of ChR2 b Peak recovery kinetics of ChR2
c Electrophysiological reaction schemes d Different reversal potentials of I0 and Is
C D
O1 O2
Ip
Is
I0
I0
470 nm
500 pA
100 ms
–60 –30 30
–1,000
–500
E (mV)
I s (p
A)
I p (I
max
)
+40 mV
–60 mV2 s
0.2 Imax
470 nm
–60 mV
Δt (s)0 5 10 15
0.0
0.5
1.0
Erev
–20 mV
0 mV
470 nm
–20
0
20
470 nm
E rev
(mV
)
IsI0Is
Is
Ip
Ip
Figure 3Electrophysiological properties of channelrhodopsins (ChRs). (a,b,d ) Photocurrents of ChR2 measured under voltage-clamp conditionsin human embryonic kidney (HEK) 293 cells. (a) ChR2 photocurrents at different holding potentials during continuous, high-intensityillumination. Currents exhibit peak levels (Ip) that inactivate to stationary levels of reduced amplitude (Is). The initial current (I0) isobtained by linear extrapolation to time zero of illumination. The corresponding current–voltage relationship of Is is given as an inset.The blue arrow indicates the reversal potential (Erev). (b) Recovery of ChR2 Ip depends on the time interval between two consecutiveilluminations. Is is unaffected by the dark interval. (c) Reaction schemes to explain the electrophysiological properties of ChRs. Theupper schemes depict three-state models; C indicates a closed state, O indicates an open state, and D indicates a desensitized state (15,48, 50). The lower schemes depicts four-state models consisting of two closed states (D and C; C1 and C2) and two open states (O1 andO2) (4, 23, 51). The bottom right scheme is consistent with the photocycle model depicted in Figure 2a. Light-induced transitions arehighlighted by light blue arrows. (d ) ChR2 photocurrents measured at conditions close to the respective reversal potentials (Erev)[internal solution: N-methyl-D-glucamine (NMG), pH 9.0]. Peak currents (Ip) are inwardly directed, whereas stationary photocurrents(Is) are outwardly directed. Accordingly, Erev of Ip/I0 is positively shifted compared with the reversal potential of the corresponding Is.
Color Tuning in Channelrhodopsins
The naturally occurring ChRs studied thus far exhibit action spectrum maxima between 436 nm(ChR from Tetraselmis striata) and 590 nm (Chrimson from Chlamydomonas noctigama) (33). Thepalette of color-tuned ChRs is supplemented by chimeric ChRs that combine helices from differentChRs, as summarized in Figure 2b and Table 1.
174 Schneider · Grimm · Hegemann
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44CH08-Schneider ARI 28 May 2015 15:47
Table 1 Overview of color-tuned channelrhodopsin (ChR) variants
ChR variant OriginMaxima of action spectrum at
pH 7.2a Reference(s)TsChR Tetraselmis striata 436 nm (33)PsChR Platymonas subcordiformus 444 nm (17)ChR2 Chlamydomonas reinhardtii 460 nm (48)VChR2 Volvox carteri 465 nm (32)ChR1 Chlamydomonas reinhardtii 487 nm (47)C1C2 = ChEF Chlamydomonas reinhardtii 458 nm (39, 73, 76)VChR1 Volvox carteri 536 nm (57, 82)ReachR Chlamydomonas reinhardtii/Volvox carteri 531 nm (38)MChR Mesostigma viride 531 nm (18)C1V1 Chlamydomonas reinhardtii/Volvox carteri 536 nm (57, 81)Chrimson Chlamydomonas noctigama 587 nm (33)
aAction spectra were recorded in human embryonic kidney (HEK) 293 cells, and maxima were determined by fitting to a five-parameter Weibull function.
In general, retinal absorption depends on the conformation of the retinal (e.g., nonplanarity),electrical interactions of the RSBH+ with the counterion complex, and other electrostatic interac-tions that either stabilize or destabilize the ground state or excited states (2, 41). Characterizationof different ChR chimera showed that H7 is a major determinant for the wavelength sensitivityof ChR activation (57, 76) because the exact localization of the RSB lysine (K257 of ChR2) de-fines both the direct intrahelical interactions with Ci2 and interhelical interactions of the RSBH+
with Ci1. Moreover, H7 may determine the number and position of water molecules along thechannel pore, thereby indirectly influencing counterion arrangement. Most obvious attempts tocolor-shift ChR absorption included exchanging the counterion residues to vary the distanceand interaction strength between the counterions and the RSBH+. In ChR2 and PsChR, ex-changes of Ci1 by glutamine (E123Q and E106Q) lead to bathochromically shifted absorptionby 23 and 30 nm, whereas replacements of Ci2 by asparagine (D253N and D236N) inducedsmaller bathochromic shifts (16 and 14 nm, respectively) (17). In contrast, mutation of Ci1 inC1V1 (E162T) lead to a hypsochromic shift most likely caused by water invasion into the ac-tive site complex in the mutant (57, 81). Apart from counterion residues, the polarity of aminoacids directly interacting with the retinal polyene chain, and with the β-ionone ring in partic-ular, influences retinal geometry and electrostatics. A prominent example is that of the residueat position 181 in ChR2 (Figure 1b). While all blue-light absorbing ChRs (and MChR) pos-sess a nonpolar glycine residue, the red-shifted ChRs VChR1 (and its derivatives) and Chrim-son have a polar serine residue at the corresponding position. Accordingly, C1C2 G220S is redshifted by 13 nm (F. Schneider, unpublished data), and C1V1 S220G shows a 12-nm blue shift(57). Although the retinal-binding pocket is the most important contributor to color tuning inChRs, long-range interactions within the protein can also modulate the absorption spectrum,most likely by indirectly influencing the geometry of the counterion complex (the active site).A glutamate residue found in ChR1 (E87), PsChR1, VChR1, DChR1, and related ChRs causesbathochromic shifts upon acidification of the extracellular solution, whereas a comparable spectralshift is not present in ChR2 (17, 73). Mutation of this residue to an uncharged glutamine stabilizesthe red-shifted form in ChR1-like ChRs. Because C1C2 E87 is more than 20 A away from theRSB nitrogen, this effect can only be explained by secondary conformational changes in the protein.
www.annualreviews.org • Biophysics of Channelrhodopsin 175
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44CH08-Schneider ARI 28 May 2015 15:47
CHANNELRHODOPSIN PHOTOCURRENT PROPERTIES
ChR photocurrents have been intensively characterized by electrical recordings from ChR-expressing Xenopus oocytes and HEK cells, mainly under voltage-clamp conditions. At negativevoltages (−60 mV), cells expressing ChR exhibit inward-directed photocurrents carried by protonsand cations (Figure 3a). Excitation of ChR1 and ChR2 with bright light pulses evokes photocur-rent rise with time constants of approximately 200 μs, and the maximum of the early peak currentsIp is reached within 1–2 ms (4, 73). Ip decreases to a stationary current Is in a process referredto as inactivation. This transition is caused by channel desensitization through equilibration oftwo open states (see the section titled “Electrophysiological Reaction Schemes” below) and bythe accumulation of late nonconducting photocycle intermediates (P480 and P480′) (6, 48). Afterlight application, currents decline biexponentially to baseline, emphasizing the presence of at leasttwo conducting states that contribute to Is (23, 51). In ChR2, channel closure shows effective timeconstants of 10 to 20 ms. Repetitive ChR activation starts from partially dark-adapted ChRs, re-sulting in reduced peak currents, and the initial peak amplitude is only regained after a dark periodof several seconds (23, 48) (Figure 3b). All kinetic parameters strongly depend on experimentalconditions such as voltage, pH, and the quality of the actinic light (23, 48).
Electrophysiological Reaction Schemes
Analogous to photocycle models based on spectroscopic findings, electrophysiological reactionschemes were developed to quantitatively describe photocurrent properties. The challenge wasto develop consistent models that account for (a) the fast closure of the channel after either asingle flash or a longer light pulse, (b) the slow recovery of the transient peak current in therange of seconds, and (c) the existence of two conducting states with different selectivities (seebelow). Three-state models comprising a closed state (C), an open state (O), and a desensitizedstate (D) were employed to explain channel inactivation and the recovery kinetics of the peakcurrent (Figure 3c, upper row) (15, 48, 50). These models imply a second light-dependent stepin the reaction scheme, such that light either induces photoactivation of the D state with reducedefficiency or accelerates the reverse reaction from D to C (15, 50). However, all three-state modelsfailed to reproduce both the biexponential off-kinetics of the channel current and alterations ofselectivity between early and late photocurrents. Moreover, a simple four-state model implyingtwo consecutive open states, O1 and O2 (Figure 3c, bottom left), could not explain both fastchannel closure and slow recovery kinetics of the peak current. Only by employing a four-statemodel with two closed and two open states (C1, C2, O1, and O2), could the channel kinetics anddark recovery of Ip be properly modeled (Figure 3c, bottom right) (23, 51). Dark-adapted ChRmolecules reside in the C1 state, which is photoconverted to O1 upon light excitation. Duringprolonged illumination, O1 and O2 equilibrate within milliseconds and eventually convert totheir respective closed states, C1 and C2, whereas the C2-to-C1 back conversion needs two to tenseconds of darkness2 (51). The C1-to-O1 photoactivation exhibits high quantum efficiency, andO1 shows high conductivity but is rather short-lived (51). Inversely, the C2-to-O2 conversion isproposed to be less effective, and O2 is less conductive but is long lived (51). Furthermore, onestudy of the ChR2 C128T mutant showed that the different dark-adapted states and their respectiveopen states are populated depending on the color and duration of the preceding illumination (59).Thus, the conductivity of ChRs is adapted according to the color, duration, and intensity of theapplied light (23, 51).
2In reality, full dark adaptation is more complex, as seen from the photocycle in Figure 2a.
176 Schneider · Grimm · Hegemann
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44CH08-Schneider ARI 28 May 2015 15:47
Ion Selectivity
Although all ChRs primarily conduct protons, they also conduct monovalent cations, and to someextent divalent cations, especially under physiological conditions of low proton concentration. Thefollowing relative conductivities have been determined for ChR1, ChR2, VChR1, and VChR2:p(H+) � p(Li+) > p(Na+) > p(K+) > p(Rb+) > p(Cs+) ≈ p(Ca2+) > p(Mg2+) (6, 15, 39, 48,63, 73, 82). Given the reversal potentials of ChR2, the relative proton conductance has beenestimated to be 105–106 times higher than the Na+ conductivity, whereas the Ca2+ conductancewas calculated to be ∼12% of the Na+ conductance (39, 48). In a solution containing differentcations (e.g., a standard extracellular solution containing H+, Na+, Ca2+, and Mg2+), all cationspresent compete for binding and transport in a voltage- and pH-dependent fashion. Thus, forexample, Na+ currents are larger at pH 9.0 than they are in the presence of competing H+ at pH7.0 (20, 63).
Studies have shown that when recording photocurrents close to the reversal potential, the earlyphotocurrent I0 and the stationary current Is can exhibit different directionalities of cation flow(e.g., inward-directed I0 and outward-directed Is) (Figure 4d ). These differences in directionalitymost impressively demonstrate the different reversal voltages and distinct cation selectivities of I0
and Is (6, 63).An enzyme kinetics model was used to quantitatively describe ChR-mediated cation transport
via two open states (20, 63). The initial current, I0, reflects conductance via only the first openstate, O1, whereas the stationary photocurrent, Is, is fed by the conductances of both open states,O1 and O2 (63). The model further implies competition between four different cationic substratesfor binding to an empty binding site of the protein (63). After loading, the binding site switchesfrom an external exposure to an internal exposure, thereby transporting cations to the other sideof the membrane. Subsequently, the cation is released and the empty binding site can reorient.After independent determination of the binding affinities for H+ and for all relevant cations,photocurrents can be dissected to understand the contributions of competing cations at a givenexperimental condition. The peculiar difficulty is that the competition between ions significantlydepends on the applied voltage; therefore, the relative conductances cannot be determined fromreversal voltages as formalized by the Goldmann–Hodgkin–Katz equation. For ChR2 at a highNa+ concentration, pHe 7.2, and −60 mV, the initial current, I0, is driven mainly by protons, witha minor contribution from Na+ (33%), whereas the stationary current, Is, shows higher relativeNa+ currents (48%) (63). Other ChR variants, such as C1V1 and CatCh+ (ChR2 L132C-T159C),show generally higher Na+ currents than ChR2 (63). In most ChRs, considerable Ca2+ currentsare only observed at high, nonphysiological Ca2+ concentrations above 10 mM (63). In contrast,Mg2+ is bound more tightly, resulting in small Mg2+ currents even at concentrations as low as afew millimolar. Only in CatCh+ are Mg2+ currents larger; these currents contribute to 15% of Is
under standard conditions (63).
Putative Selectivity Filter
Highly selective channels, such as K+ channels or Na+ channels, contain so-called selectivity filtersthat determine the kind of transported ions. These selectivity filters represent the narrowest part ofthe ion-conducting pathway and adopt conserved symmetric structures that specifically bind onlyone ion species, for example, by mimicking the hydration shell of the respective ion (12). AlthoughChRs naturally occur as dimeric proteins, the ion-conducting pathway is located between H1, H2,H3, and H7 of each monomer (for details, see the section titled “Ion Permeation Pathway”), so nosymmetric selectivity filter is found (31). Moreover, ChRs are relatively unselective for different
www.annualreviews.org • Biophysics of Channelrhodopsin 177
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44CH08-Schneider ARI 28 May 2015 15:47
Cytosol
Extracellularside
Extracellular sideExtracellular side
CytosolCytosolNa+
H6H6H7H7
H3H3
H1H1
H2H2
H4H4H134H134
R268R268
N258N258
Ci2Ci2
Ci1Ci1E3E3
E2E2
E1E1
K257K257
ChR2ChR2
DC pairDC pair
hν
H2 K1K1
1
2
3
1
2
3
Figure 4Helix 2 (H2)-tilt (HT) model for ChR2 activation. The numbered panels on the left depict the three stepsrequired for ChR2 pore formation: � photon absorption and retinal isomerization; � disruption of theE3–N258 hydrogen bond and deprotonation and downflip of E3; and � outward movement of H2,disconnection of hydrogen bonds at the inner gate, and continuous pore formation. Blue spheres depictwater molecules, white spheres depict Na+ ions, and dark blue bars reflect closed gates or barriers in theprotein. The inset on the right highlights important residues involved in pore formation. The DC pair iscomposed of C128 and D156 at the active site; however, its role in channel activation remains to beinvestigated. The ChR2 structure was gained by homology modeling using the C1C2 crystal structure as atemplate (31). The gray wireframe depicts cavities in which water molecules and cations may reside indark-adapted ChR2. Hydrogen bonds depicted in this panel are drawn according to Reference 36.
cations with various atomic radii (e.g., Li+ versus Cs+), arguing against a defined binding site fora specific cation. Most likely, ChRs mediate cation flux via a water-filled pore without completedehydration of the transported ions.
Several key elements have been identified to regulate cation selectivity in ChRs. Most impor-tantly, ChR2 S63, E90, and N258 of the central gate (Figure 1e) are critical determinants. Theintroduction of a second negative charge into the central gate region, as, for example, in ChR2S63D or N258D, favors the transport of divalent cations (31, 55). Mutations of ChR2 E90 resultin different ion selectivities depending on the character of the introduced residue. In contrast toChR2 E90Q and E90A, which show strongly reduced proton selectivity with only minor shifts in
178 Schneider · Grimm · Hegemann
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44CH08-Schneider ARI 28 May 2015 15:47
Electrophysiologicalcharacterization:time-resolvedmeasurement ofmembrane current orvoltage in ChR-expressing cells
reversal potentials between pH 4.0 and pH 9.0, ChR2 E90H exhibits no detectable Na+ conduc-tivity but shows pH-dependent proton currents (13, 20, 61). The ion selectivity of ChR2 E90Khas long been a mystery. At acidic pH, the mutant shows strongly enhanced inward currents;however, outward currents are also stimulated, and the reversal voltage does not change betweenpH 4.0 and pH 9.0 (13, 61). In fact, the introduction of a lysine or arginine residue with a netcharge of +1 into the central gate inverts the ChR2 ion selectivity to select monovalent anionswith high Cl− conductance when this ion is available as a substrate (ChloC) (79). However, aCl−-conducting channel has also been achieved by sequentially modeling the conducting pore,which has a negatively charged surface, into one that has a more positive surface (iC1C2) (5).Both of the Cl−-conducting ChRs, ChloC and iC1C2, exhibit only minor H+ conductance andnegligible cation conductance.
The counterion residues Ci1 and Ci2 indirectly affect ion selectivity, as they are located close tothe central gate but oriented toward the RSB. ChR2 E123A conducts mainly protons, consistentwith increased H+/Na+ ratios in homologous C1C2 E162A and D292A (20, 31). Interestingly,DChR naturally contains an alanine at the Ci1 position (A178) and is itself proton selective (83).ChR2 K93 (K1) was proposed to be directly hydrogen-bonded to Ci1 from the extracellular site(Figure 4, right) and to form a hydrogen bond with Ci2 via an interjacent water molecule (28).Substitution of K1 with alanine in C1C2 increases the ratio of H+/Na+ conductance (31), possiblyby favoring protonation of the counterion residues or by reorienting central gate residues such asE3.
MD simulations for ChR2 visualize two preferential Na+ occupancies (see the section titled“Ion Permeation Pathway”) (78). One occupancy site is located extracellular to the central gate,implying the involvement of S52, Q56, and E4 (Figure 1f ). But, this assignment is questionablebecause S52 is not conserved, and only replacement of Q56 results in enhanced selectivity for H+
and K+ (31), whereas E4 exchange to alanine does not alter cation selectivity (71). A second Na+
binding site is formed by E1, E2, H134, H265, and R268, residues that are close to or part ofthe inner gate (78). Of these residues, only H134 is crucial for selectivity (20). H134R, H134S,and H134N mutants show photocurrents carried by Na+ that are strongly inhibited by bothintracellular and extracellular H+. This inhibition is explained by inefficient proton release on thecytosolic side. The nearby residue ChR2 L132 faces away from the inner gate, but its replacementin ChR2 L132C, also referred to as CatCh, indirectly influences cation binding, resulting in a1.6-fold increase in Ca2+ selectivity, enlarged Mg2+ conductance, and high stationary currents(34, 57, 63).
Voltage Dependence
The electrophysiological characterization of ChRs is usually performed under voltage-clamp con-ditions; however, ChR activation in green algae or neurons rapidly depolarizes the membrane.This depolarization changes the properties of the channel because ChRs are highly voltage sensi-tive and exhibit inward rectification, voltage-dependent cation selectivity, and voltage-modulatedphotocycle kinetics (80).
ChRs display asymmetric current–voltage relations in which inward currents are stronger thanoutward currents (inward rectification) (Figure 3a). In inward-rectifying K+ channels, cytosolicMg2+ causes inward rectification by blocking channel activity in a voltage-dependent manner. Incontrast, variations in the internal Mg2+ concentration in ChRs do not affect current sizes, reversalpotentials, or the shapes of current–voltage relationships, so Mg2+ can be excluded as a potentialsource of inward rectification (20). On the basis of our previous kinetic analysis, we suggested thatChR rectification is caused by fast, voltage-dependent cation binding in cases in which protons
www.annualreviews.org • Biophysics of Channelrhodopsin 179
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44CH08-Schneider ARI 28 May 2015 15:47
Voltage dependence:membrane voltagedependence ofphotocycle kineticsand cation selectivity
and cations compete for one predominant external binding site (20). Accordingly, rectification issmall in ChR mutants that preferentially conduct either H+, Na+, or Cl−, such as the H+-selectiveE90H mutant or the Cl−-selective ChloC (20, 79). Interestingly, inward rectification is reducedupon mutating selected inner gate residues (ChR2 Y70, E1, and E2) ( J. Wietek, unpublishedresults). Thus, charged and polar inner gate residues might be part of the inherent voltage sensorof ChR that reorients in a manner dependent on the applied membrane voltage.
The current transport model implies voltage-sensitive rate constants, and the apparent voltageis determined using an elastic voltage-divider (20). In fact, ChR cation selectivity and thus the ratioof the conducted ions depend on the membrane voltage (63). At negative voltages (e.g., −60 mV),the stationary currents of selected ChRs (ChR2, C1V1, CatCh+) are mediated primarily by Na+
transport, whereas the initial current, I0, is driven mainly by protons (63). In contrast, whenconsidering reversal potentials, stationary currents show higher proton selectivity than do theirrespective initial currents (6, 63). Together, these findings demonstrate that ChR cation selectivitydiffers between conditions that are close to thermodynamic equilibrium and conditions in whichhigh electrochemical gradients produce strong unidirectional driving forces. The voltage-sensitivecation selectivity of early and late photocurrents may also explain the voltage-dependent degreeof photocurrent inactivation.
Whereas voltage-dependent steps in the ChR photocycle are difficult to analyze by spectro-scopic means, the voltage dependence of channel opening, closing, and recovery kinetics has beendetermined in electrophysiological measurements. Channel opening occurs within ∼200 μs as de-scribed for ChR1, ChR2, and a Volvox ChR chimera, and it shows little to no voltage dependence(4, 7, 15, 73). In contrast, ChR2 and ChR2 mutants (ChR2 H134R, ChR2 T159C) exhibit channelclosing rates that are accelerated twofold to threefold at negative membrane voltages (−100 mV)compared with positive voltages (+50 or +80 mV) (4, 7). Similarly, in ChR2 and C1V1, the ki-netics of peak recovery were shown to be threefold to fourfold faster at negative voltages than atpositive voltages (48, 57). Mutation of ChR2 Ci1 to threonine results in fast, voltage-independentchannel closing and in remarkably fast recovery kinetics (7, 21). Therefore, the counterion residueand its interaction partners, including ChR2 E3 and K1, which are involved in voltage-dependentprotonation reactions, reorganization of the hydrogen network, or conformational changes, mod-ulate channel closure and recovery of the dark-adapted state. Chronos, a ChR from Stigeocloniumhelveticum, displays very fast photocycle kinetics (33). Most interestingly, Chronos exhibits inverseoff-kinetics voltage-dependence: It displays faster channel closing at positive voltages (33). In theChronos primary sequence, the residues corresponding to ChR2 Ci1 and T159 are replaced bymethionine (M140) and asparagine (N176), respectively. To fully understand voltage sensing inChRs, however, further structural information on photointermediates, and on the conductingstates specifically, is urgently needed.
THE HELIX 2 (H2)-TILT MODEL FOR CHANNELRHODOPSINACTIVATION
Structural information, detailed electrical measurements, and time-resolved Fourier transforminfrared (FTIR) and electron paramagnetic resonance (EPR) measurements, in combination withMD calculations, resulted in the following model for ChR activation (Figure 4). All-trans to13-cis isomerization induces the movement of N258, which is next to the retinal-binding residueK257 (36). This movement is in agreement with early backbone conformational changes observedafter a femtosecond flash on a picosecond timescale (49). The next early event is the disruptionof one of the two hydrogen bonds between N258 and E90 (E3); this disruption causes a specialrearrangement of H2 and H7 and induces an outward flip, as well as deprotonation of E3 itself
180 Schneider · Grimm · Hegemann
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44CH08-Schneider ARI 28 May 2015 15:47
(36). The tilt of H2 in the MD simulations agrees with the light-induced movement of spin-labeled cysteines on H2 and H7, as measured by EPR of selected spin-labeled cysteines (35,62). The outward orientation of E3 opens up a small pore, and water molecules invade fromthe extracellular site into the vestibule between the inner gate and the outer gate, resulting inthe formation of the preopen state (36). Subsequently, only small conformational changes andproton-transfer reactions at the inner gate appear to be important for the transition from thepreopen state to the fully open state (59). These conformational changes might involve separationof the salt bridges E1/R268 and E2/H134 (Figure 1d and Figure 4). As both glutamic acids aredeprotonated before and after separation, however, reorientation at the inner gate is expectedto be almost infrared silent, and has not been seen in time-resolved FTIR measurements thusfar. Notably, a very recent electron microscopy study confirmed light-induced rearrangementsof H2, H6, and H7 and suggested that H2 and H7 become more flexible after illumination (47).This observation is consistent with disruption of the interconnecting hydrogen bonds of both thecentral gate and the inner gate (E3/N258, E1/R268 and E2/H134).
To fully understand ChR gating, we need to accommodate the DC pair C128/D156. Modifi-cations in these residues dramatically slow down the photocycle from milliseconds to minutes (3,8, 81). The connection between H3 and H4 is important, as shown for the homologs T90/D115in BR (16); however, the detailed functionality of the DC pair for ChR activation is not yet clear.
APPLICATIONS OF CHANNELRHODOPSINS
ChRs are most commonly used to depolarize the plasma membranes of excitable cells, therebyeither directly activating cellular activity such as action potential firing or reducing the thresholdfor such events (10, 27). In fact, different ChR variants should be considered for the activationof different target cell types; for example, a fast ChR with voltage-independent off-kinetics, suchas a ChETA mutant, would be most suited to eliciting action potentials in fast-spiking interneu-rons (7, 21). In contrast, a ChR exhibiting moderate photocycling kinetics and moderate currentinactivation, as seen in ChR2 H134R, might be the preferred option to optogenetically mimicthe action potential of a cardiomyocyte (80). Moreover, bistable SFRs are the best choice for theprolonged induction of subthreshold depolarization without continuous illumination (3, 8).
ChRs might also be used to alter cation distributions along both plasma membranes andintracellular membranes. Although ChRs in general are not highly selective for a single cationspecies, the ionic photocurrent composition differs between different ChRs and should be takeninto account when choosing an appropriate ChR variant (63). For example, the Catch+ mutantis probably the best-suited variant for triggering Ca2+ influx into the cytosol from intracellularstores because its Ca2+ conductance is high at high Ca2+ concentrations (57, 63). Two recentlydeveloped ChRs, ChloC and iC1C2, represent anion-selective ChRs that can be used to clampthe voltage to the reversal potential for Cl−, thereby suppressing action potential firing (5, 79).
Color-tuned ChRs enable independent activation of distinct cellular populations and allowChRs to be used in combination with fluorescent sensor proteins. Furthermore, the improvedensemble of orange- and red-light activated ChRs, including ReachR and Chrimson, allows forChR activation in intact tissues and organs, owing to reduced absorption and scattering of red lightin living tissue (33, 38). Alternatively, ChRs that exhibit an elevated two-photon absorption cross-section, such as C1V1, can be employed for two-photon activation with near-infrared light (56).
ChR properties to consider include unitary conductance, retinal binding affinity, and proteinstability. Moreover, properties that strongly depend on the target cells, such as expression leveland membrane targeting, are of serious and equal importance, and these should be individuallytested for each ChR variant and cell type, respectively. Finally, to help users choose a ChR variant
www.annualreviews.org • Biophysics of Channelrhodopsin 181
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44CH08-Schneider ARI 28 May 2015 15:47
Table 2 Overview of channelrhodopsin (ChR) variants relevant for optogenetic applications
ChR variant Use(s) and special properties Reference(s)ChR2 H134R Widely used ChR, increased Na+ conductivity, improved retinal binding in
Caenorhabditis elegans(30, 39)
CatCh (ChR2 L132C) Enhanced Mg2+ and Ca2+ selectivity, large photocurrents, low inactivation (34, 57)ChR2 T159C Large photocurrents, improved retinal binding affinity (7, 74)CatCh+ (ChR2 L132C-T159C) Enhanced Mg2+ and Ca2+ selectivity, large photocurrents, low inactivation,
provides high light sensitivity to host cells, more stable expression than CatCh(52, 57, 63)
ChETA (ChR2 E123T-T159C) Fast photocycle at the expense of reduced transported charge per absorbedphoton, reduced voltage dependency of channel closing kinetics
(7, 21)
Chronos Very fast photocycle, large photocurrents, inverse voltage dependency of offkinetics, provides high light sensitivity to host cells
(33)
Step-function rhodopsins(ChR2 C128A/S/T; ChR2D156A/C/N)
Very slow photocycle kinetics and extended open-state lifetimes, provides veryhigh light sensitivity to cells, UV light– and green light–induced channelclosure (bistable rhodopsins)
(3, 8)
ChIEF Fast photocycle, low inactivation (39)C1V1 (E122T-E162T) Green light–induced activation, high two-photon cross-section (57, 81)ReachR Green light–induced activation, high expression level, and large photocurrents
in mammalian cells(38)
PsChR Violet light–induced activation, high unitary conductance and high Na+
conductivity, low inactivation, fast recovery kinetics(17)
Cs-Chrimson Orange light–induced activation, improved membrane targeting by use of theCsChR (ChR87) N-terminus
(33)
Slow ChloC(ChR2 E90R-D156N T159C)
Cl− selectivity, slow photocycle, improved photocurrents and shifted reversalpotential, used for voltage clamping to the Cl− reversal potential
(79)
iC1C2 Cl− selectivity, used for voltage clamping to the Cl− reversal potential (5)
with appropriate biophysical properties, Table 2 summarizes the most useful and most commonlyapplied ChR variants for optogenetic applications.
FUTURE ISSUES
1. 3D structures of different states as they occur during the cyclic reaction pathway (pho-tocycle) and structures of the open state(s) are most urgently needed.
2. Visualization of proton dynamics including structural changes and proton transfer reac-tions using nuclear magnetic resonance (NMR).
3. Extension of the activation model by stepwise inclusion of structural changes, waterdynamics, and proton-transfer steps.
4. Further improvement of the chloride-conducting channels and the possible generationof Na+ and K+-selective ChRs.
DISCLOSURE STATEMENT
The authors are not aware of any affiliations, memberships, funding, or financial holdings thatmight be perceived as affecting the objectivity of this review.
182 Schneider · Grimm · Hegemann
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44CH08-Schneider ARI 28 May 2015 15:47
ACKNOWLEDGMENTS
We thank Johannes Vierock for stimulating discussions during the preparation of the manuscript,and we thank Jonas Wietek for providing action spectra of selected ChR variants. The workwas supported by the Louis-Jeantet Foundation and the German Research Foundation, DFG(SFB1078 B2).
LITERATURE CITED
1. Arenkiel BR, Peca J, Davison IG, Feliciano C, Deisseroth K, et al. 2007. In vivo light-induced activationof neural circuitry in transgenic mice expressing channelrhodopsin-2. Neuron 54(2):205–18
2. Babitzki G, Denschlag R, Tavan P. 2009. Polarization effects stabilize bacteriorhodopsin’s chromophorebinding pocket: a molecular dynamics study. J. Phys. Chem. B 113(30):10483–95
3. Bamann C, Gueta R, Kleinlogel S, Nagel G, Bamberg E. 2010. Structural guidance of the photocycle ofchannelrhodopsin-2 by an interhelical hydrogen bond. Biochemistry 49(2):267–78
4. Bamann C, Kirsch T, Nagel G, Bamberg E. 2008. Spectral characteristics of the photocycle ofchannelrhodopsin-2 and its implication for channel function. J. Mol. Biol. 375(3):686–94
5. Berndt A, Lee SY, Ramakrishnan C, Deisseroth K. 2014. Structure-guided transformation of channel-rhodopsin into a light-activated chloride channel. Science 344:420–24
6. Berndt A, Prigge M, Gradmann D, Hegemann P. 2010. Two open states with progressive proton selec-tivities in the branched channelrhodopsin-2 photocycle. Biophys. J. 98(5):753–61
7. Berndt A, Schoenenberger P, Mattis J, Tye KM, Deisseroth K, et al. 2011. High-efficiency channel-rhodopsins for fast neuronal stimulation at low light levels. PNAS 108(18):7595–600
8. Berndt A, Yizhar O, Gunaydin LA, Hegemann P, Deisseroth K. 2009. Bi-stable neural state switches.Nat. Neurosci. 12(2):229–34
9. Bi A, Cui J, Ma Y-P, Olshevskaya E, Pu M, et al. 2006. Ectopic expression of a microbial-type rhodopsinrestores visual responses in mice with photoreceptor degeneration. Neuron 50(1):23–33
10. Boyden ES, Zhang F, Bamberg E, Nagel G, Deisseroth K. 2005. Millisecond-timescale, genetically tar-geted optical control of neural activity. Nat. Neurosci. 8(9):1263–68
11. Douglass AD, Kraves S, Deisseroth K, Schier AF, Engert F. 2008. Escape behavior elicited by single,channelrhodopsin-2-evoked spikes in zebrafish somatosensory neurons. Curr. Biol. 18(15):1133–37
12. Doyle DA. 1998. The structure of the potassium channel: molecular basis of K+ conduction and selectivity.Science 280(5360):69–77
13. Eisenhauer K, Kuhne J, Ritter E, Berndt A, Wolf S, et al. 2012. In channelrhodopsin-2 glu-90 is crucialfor ion selectivity and is deprotonated during the photocycle. J. Biol. Chem. 287(9):6904–11
14. Ernst OP, Sanchez Murcia PA, Daldrop P, Tsunoda SP, Kateriya S, Hegemann P. 2008. Photoactivationof channelrhodopsin. J. Biol. Chem. 283(3):1637–43
15. Foster KW, Smyth RD. 1980. Light antennas in phototactic algae. Microbiol. Rev. 44(4):572–63016. Garczarek F, Gerwert K. 2006. Polarized FTIR spectroscopy in conjunction with in situ H/D exchange
reveals the orientation of protein internal carboxylic acids. J. Am. Chem. Soc. 128(1):28–2917. Govorunova EG, Sineshchekov OA, Li H, Janz R, Spudich JL. 2013. Characterization of a highly efficient
blue-shifted channelrhodopsin from the marine alga Platymonas subcordiformis. J. Biol. Chem. 288:29911–2218. Govorunova EG, Spudich EN, Lane CE, Sineshchekov OA, Spudich JL. 2011. New channelrhodopsin
with a red-shifted spectrum and rapid kinetics from Mesostigma viride. mBio 2(3):e00115–1119. Gradinaru V, Mogri M, Thompson KR, Henderson JM, Deisseroth K. 2009. Optical deconstruction of
Parkinsonian neural circuitry. Science 324(5925):354–5920. Gradmann D, Berndt A, Schneider F, Hegemann P. 2011. Rectification of the channelrhodopsin early
conductance. Biophys. J. 101(5):1057–6821. Gunaydin LA, Yizhar O, Berndt A, Sohal VS, Deisseroth K, Hegemann P. 2010. Ultrafast optogenetic
control. Nat. Neurosci. 13(3):387–9222. Hegemann P. 2008. Algal sensory photoreceptors. Annu. Rev. Plant Biol. 59:167–8923. Hegemann P, Ehlenbeck S, Gradmann D. 2005. Multiple photocycles of channelrhodopsin. Biophys. J.
89(6):3911–18
www.annualreviews.org • Biophysics of Channelrhodopsin 183
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44CH08-Schneider ARI 28 May 2015 15:47
24. Holland EM, Braun FJ, Nonnengasser C, Harz H, Hegemann P. 1996. The nature of rhodopsin-triggeredphotocurrents in Chlamydomonas. I. Kinetics and influence of divalent ions. Biophys. J. 70(2):924–31
25. Hou S-Y, Govorunova EG, Ntefidou M, Lane CE, Spudich EN, et al. 2011. Diversity of Chlamydomonaschannelrhodopsins. Photochem. Photobiol. 88(1):119–28
26. Hu JG, Sun BQ, Bizounok M, Hatcher ME, Lansing JC, et al. 1998. Early and late M intermediates inthe bacteriorhodopsin photocycle: a solid-state NMR study. Biochemistry 37(22):8088–96
27. Ishizuka T, Kakuda M, Araki R, Yawo H. 2006. Kinetic evaluation of photosensitivity in geneticallyengineered neurons expressing green algae light-gated channels. Neurosci. Res. 54(2):85–94
28. Ito S, Kato HE, Taniguchi R, Iwata T, Nureki O, Kandori H. 2014. Water-containing hydrogen-bondingnetwork in the active center of channelrhodopsin. J. Am. Chem. Soc. 136:3475–82
29. Joh NH, Min A, Faham S, Whitelegge JP, Yang D, et al. 2008. Modest stabilization by most hydrogen-bonded side-chain interactions in membrane proteins. Nature 453(7199):1266–70
30. Kateriya S. 2004. “Vision” in single-celled algae. News Physiol. Sci. 19(3):133–3731. Kato HE, Zhang F, Yizhar O, Ramakrishnan C, Nishizawa T, et al. 2012. Crystal structure of the chan-
nelrhodopsin light-gated cation channel. Nature 482(7385):369–7432. Kianianmomeni A, Stehfest K, Nematollahi G, Hegemann P, Hallmann A. 2009. Channelrhodopsins of
Volvox carteri are photochromic proteins that are specifically expressed in somatic cells under control oflight, temperature, and the sex inducer. Plant Physiol. 151(1):347–66
33. Klapoetke NC, Murata Y, Kim SS, Pulver SR, Birdsey-Benson A, et al. 2014. Independent optical exci-tation of distinct neural populations. Nat. Methods 11:338–46
34. Kleinlogel S, Feldbauer K, Dempski RE, Fotis H, Wood PG, et al. 2011. Ultra light-sensitive and fastneuronal activation with the Ca2+-permeable channelrhodopsin CatCh. Nat. Neurosci. 14(4):513–18
35. Krause N, Engelhard C, Heberle J, Schlesinger R, Bittl R. 2013. Structural differences between the closedand open states of channelrhodopsin-2 as observed by EPR spectroscopy. FEBS Lett. 587(20):3309–13
36. Kuhne J, Eisenhauer K, Ritter E, Hegemann P, Gerwert K, Bartl F. 2014. Early formation of the ion-conducting pore in channelrhodopsin-2. Angew. Chemie Int. Ed. 54:4953–57
37. Li X, Gutierrez DV, Hanson MG, Han J, Mark MD, et al. 2005. Fast noninvasive activation and inhi-bition of neural and network activity by vertebrate rhodopsin and green algae channelrhodopsin. PNAS102(49):17816–21
38. Lin JY, Knutsen PM, Muller A, Kleinfeld D, Tsien RY. 2013. ReaChR: a red-shifted variant of channel-rhodopsin enables deep transcranial optogenetic excitation. Nat. Neurosci. 16:1499–508
39. Lin JY, Lin MZ, Steinbach P, Tsien RY. 2009. Characterization of engineered channelrhodopsin variantswith improved properties and kinetics. Biophys. J. 96(5):1803–14
40. Lorenz-Fonfrıa VA, Resler T, Krause N, Nack M, Gossing M, et al. 2013. Transient protonation changesin channelrhodopsin-2 and their relevance to channel gating. PNAS 110(14):E1273–81
41. Luecke H, Schobert B, Lanyi JK, Spudich EN, Spudich JL. 2001. Crystal structure of sensory rhodopsinII at 2.4 angstroms: insights into color tuning and transducer interaction. Science 293(5534):1499–503
42. Luecke H, Schobert B, Richter HT, Cartailler JP, Lanyi JK. 1999. Structure of bacteriorhodopsin at1.55 A resolution. J. Mol. Biol. 291(4):899–911
43. Mittelmeier TM, Boyd JS, Lamb MR, Dieckmann CL. 2011. Asymmetric properties of the Chlamydomonasreinhardtii cytoskeleton direct rhodopsin photoreceptor localization. J. Cell Biol. 193(4):741–53
44. Muller M, Bamann C, Bamberg E, Kuhlbrandt W. 2011. Projection structure of channelrhodopsin-2 at6 A resolution by electron crystallography. J. Mol. Biol. 414(1):86–95
45. Muller M, Bamann C, Bamberg E, Kuhlbrandt W. 2014. Light-induced helix movements inchannelrhodopsin-2. J. Mol. Biol. 427(2):341–49
46. Nagel G, Brauner M, Liewald JF, Adeishvili N, Bamberg E, Gottschalk A. 2005. Light activation ofchannelrhodopsin-2 in excitable cells of Caenorhabditis elegans triggers rapid behavioral responses. Curr.Biol. 15(24):2279–84
47. Nagel G, Ollig D, Fuhrmann M, Kateriya S, Musti AM, et al. 2002. Channelrhodopsin-1: a light-gatedproton channel in green algae. Science 296(5577):2395–98
48. Nagel G, Szellas T, Huhn W, Kateriya S, Adeishvili N, et al. 2003. Channelrhodopsin-2, a directlylight-gated cation-selective membrane channel. PNAS 100(24):13940–45
184 Schneider · Grimm · Hegemann
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44CH08-Schneider ARI 28 May 2015 15:47
49. Neumann-Verhoefen M-K, Neumann K, Bamann C, Radu I, Heberle J, et al. 2013. Ultrafast infraredspectroscopy on channelrhodopsin-2 reveals efficient energy transfer from the retinal chromophore tothe protein. J. Am. Chem. Soc. 135(18):6968–76
50. Nikolic K, Degenaar P, Toumazou C. 2006. Modeling and engineering aspects of channelrhodopsin2system for neural photostimulation. Proc. Int. Conf. IEEE Eng. Med. Biol. Soc., 28th, New York, pp. 1626–29. Piscataway, NJ: IEEE
51. Nikolic K, Grossman N, Grubb MS, Burrone J, Toumazou C, Degenaar P. 2009. Photocycles ofchannelrhodopsin-2. Photochem. Photobiol. 85(1):400–11
52. Pan Z-H, Ganjawala TH, Lu Q, Ivanova E, Zhang Z. 2014. ChR2 mutants at L132 and T159 withimproved operational light sensitivity for vision restoration. PLOS ONE 9(6):e98924
53. Patzelt H, Simon B, terLaak A, Kessler B, Kuhne R, et al. 2002. The structures of the active center indark-adapted bacteriorhodopsin by solution-state NMR spectroscopy. PNAS 99(15):9765–70
54. Petreanu L, Huber D, Sobczyk A, Svoboda K. 2007. Channelrhodopsin-2-assisted circuit mapping oflong-range callosal projections. Nat. Neurosci. 10(5):663–68
55. Plazzo AP, De Franceschi N, Da Broi F, Zonta F, Sanasi MF, et al. 2012. Bioinformatic and mutationalanalysis of channelrhodopsin-2 protein cation-conducting pathway. J. Biol. Chem. 287(7):4818–25
56. Prakash R, Yizhar O, Grewe B, Ramakrishnan C, Wang N, et al. 2012. Two-photon optogenetic toolboxfor fast inhibition, excitation and bistable modulation. Nat. Methods 9:1171–79
57. Prigge M, Schneider F, Tsunoda SP, Shilyansky C, Wietek J, et al. 2012. Color-tuned channelrhodopsinsfor multiwavelength optogenetics. J. Biol. Chem. 287:31804–12
58. Radu I, Bamann C, Nack M, Nagel G, Bamberg E, Heberle J. 2009. Conformational changes ofchannelrhodopsin-2. J. Am. Chem. Soc. 131(21):7313–19
59. Ritter E, Piwowarski P, Hegemann P, Bartl FJ. 2013. Light-dark adaptation of channelrhodopsin C128Tmutant. J. Biol. Chem. 288(15):10451–58
60. Ritter E, Stehfest K, Berndt A, Hegemann P, Bartl FJ. 2008. Monitoring light-induced structuralchanges of Channelrhodopsin-2 by UV-visible and Fourier transform infrared spectroscopy. J. Biol. Chem.283(50):35033–41
61. Ruffert K, Himmel B, Lall D, Bamann C, Bamberg E, et al. 2011. Glutamate residue 90 in the predictedtransmembrane domain 2 is crucial for cation flux through channelrhodopsin 2. Biochem. Biophys. Res.Commun. 410(4):737–43
62. Sattig T, Rickert C, Bamberg E, Steinhoff H-J, Bamann C. 2013. Light-induced movement of the trans-membrane helix b in channelrhodopsin-2. Angew. Chem. Int. Ed. Engl. 52(37):9705–8
63. Schneider F, Gradmann D, Hegemann P. 2013. Ion selectivity and competition in channelrhodopsins.Biophys. J. 105(1):91–100
64. Scholz F, Bamberg E, Bamann C, Wachtveitl J. 2012. Tuning the primary reaction of channelrhodopsin-2by imidazole, pH, and site-specific mutations. Biophys. J. 102(11):2649–57
65. Schroll C, Riemensperger T, Bucher D, Ehmer J, Voller T, et al. 2006. Light-induced activation of distinctmodulatory neurons triggers appetitive or aversive learning in Drosophila larvae. Curr. Biol. 16(17):1741–47
66. Shimano T, Fyk-Kolodziej B, Mirza N, Asako M, Tomoda K, et al. 2013. Assessment of the AAV-mediatedexpression of channelrhodopsin-2 and halorhodopsin in brainstem neurons mediating auditory signaling.Brain Res. 1511:138–52
67. Sineshchekov OA, Govorunova EG, Wang J, Li H, Spudich JL. 2013. Intramolecular proton transfer inchannelrhodopsins. Biophys. J. 104(4):807–17
68. Sineshchekov OA, Jung K-H, Spudich JL. 2002. Two rhodopsins mediate phototaxis to low- and high-intensity light in Chlamydomonas reinhardtii. PNAS 99(13):8689–94
69. Stehfest K, Hegemann P. 2010. Evolution of the channelrhodopsin photocycle model. ChemPhysChem11(6):1120–26
70. Stehfest K, Ritter E, Berndt A, Bartl F, Hegemann P. 2010. The branched photocycle of the slow-cyclingchannelrhodopsin-2 mutant C128T. J. Mol. Biol. 398(5):690–702
71. Sugiyama Y, Wang H, Hikima T, Sato M, Kuroda J, et al. 2009. Photocurrent attenuation by a singlepolar-to-nonpolar point mutation of channelrhodopsin-2. Photochem. Photobiol. Sci. 8(3):328–36
72. Suzuki T, Yamasaki K, Fujita S, Oda K, Iseki M, et al. 2003. Archaeal-type rhodopsins in Chlamydomonas:model structure and intracellular localization. Biochem. Biophys. Res. Commun. 301(3):711–17
www.annualreviews.org • Biophysics of Channelrhodopsin 185
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44CH08-Schneider ARI 28 May 2015 15:47
73. Tsunoda SP, Hegemann P. 2009. Glu 87 of channelrhodopsin-1 causes pH-dependent color tuning andfast photocurrent inactivation. Photochem. Photobiol. 85(2):564–69
74. Ullrich S, Gueta R, Nagel G. 2012. Degradation of channelopsin-2 in the absence of retinal and degra-dation resistance in certain mutants. Biol. Chem. 394:271–80
75. Verhoefen M-K, Bamann C, Blocher R, Forster U, Bamberg E, Wachtveitl J. 2010. The photocy-cle of channelrhodopsin-2: ultrafast reaction dynamics and subsequent reaction steps. ChemPhysChem11(14):3113–22
76. Wang H, Sugiyama Y, Hikima T, Sugano E, Tomita H, et al. 2009. Molecular determinants differentiatingphotocurrent properties of two channelrhodopsins from Chlamydomonas. J. Biol. Chem. 284(9):5685–96
77. Watanabe HC, Welke K, Schneider F, Tsunoda S, Zhang F, et al. 2012. Structural model of channel-rhodopsin. J. Biol. Chem. 287(10):7456–66
78. Watanabe HC, Welke K, Sindhikara DJ, Hegemann P, Elstner M. 2013. Towards an understanding ofchannelrhodopsin function: simulations lead to novel insights of the channel mechanism. J. Mol. Biol.425:1795–814
79. Wietek J, Wiegert JS, Adeishvili N, Schneider F, Watanabe H, et al. 2014. Conversion of channel-rhodopsin into a light-gated chloride channel. Science 344:409–12
80. Williams JC, Xu J, Lu Z, Klimas A, Chen X, et al. 2013. Computational optogenetics: empirically-derivedvoltage- and light-sensitive channelrhodopsin-2 model. PLOS Comput. Biol. 9:e1003220
81. Yizhar O, Fenno LE, Prigge M, Schneider F, Davidson TJ, et al. 2011. Neocortical excitation/inhibitionbalance in information processing and social dysfunction. Nature 477(7363):171–78
82. Zhang F, Prigge M, Beyriere F, Tsunoda SP, Mattis J, et al. 2008. Red-shifted optogenetic excitation: atool for fast neural control derived from Volvox carteri. Nat. Neurosci. 11(6):631–33
83. Zhang F, Vierock J, Yizhar O, Fenno LE, Tsunoda S, et al. 2011. The microbial opsin family of optogenetictools. Cell 147(7):1446–57
84. Zhang Y-P, Oertner TG. 2007. Optical induction of synaptic plasticity using a light-sensitive channel.Nat. Methods 4(2):139–41
186 Schneider · Grimm · Hegemann
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44-FrontMatter ARI 4 June 2015 10:35
Annual Review ofBiophysics
Volume 44, 2015Contents
Modeling Active Mechanosensing in Cell–Matrix InteractionsBin Chen, Baohua Ji, and Huajian Gao � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 1
Biostructural Science Inspired by Next-Generation X-Ray SourcesSol M. Gruner and Eaton E. Lattman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �33
Contemporary NMR Studies of Protein ElectrostaticsMathias A.S. Hass and Frans A.A. Mulder � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �53
Anatomy of Nanoscale PropulsionVinita Yadav, Wentao Duan, Peter J. Butler, and Ayusman Sen � � � � � � � � � � � � � � � � � � � � � � � � �77
Mechanisms of AutophagyNobuo N. Noda and Fuyuhiko Inagaki � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 101
Single-Cell PhysiologySattar Taheri-Araghi, Steven D. Brown, John T. Sauls, Dustin B. McIntosh,
and Suckjoon Jun � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 123
Roles for Synonymous Codon Usage in Protein BiogenesisJulie L. Chaney and Patricia L. Clark � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 143
Biophysics of ChannelrhodopsinFranziska Schneider, Christiane Grimm, and Peter Hegemann � � � � � � � � � � � � � � � � � � � � � � � � � 167
Structure and Mechanism of RNA Mimics of GreenFluorescent ProteinMingxu You and Samie R. Jaffrey � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 187
Regulation of Rad6/Rad18 Activity During DNA Damage ToleranceMark Hedglin and Stephen J. Benkovic � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 207
Structure Principles of CRISPR-Cas Surveillanceand Effector ComplexesTsz Kin Martin Tsui and Hong Li � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 229
Structural Biology of the Major Facilitator Superfamily TransportersNieng Yan � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 257
v
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.

BB44-FrontMatter ARI 4 June 2015 10:35
Specification of Architecture and Function of Actin Structuresby Actin Nucleation FactorsColleen T. Skau and Clare M. Waterman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 285
Structural Symmetry in Membrane ProteinsLucy R. Forrest � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 311
The Synaptic Vesicle Release MachineryJosep Rizo and Junjie Xu � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 339
Index
Cumulative Index of Contributing Authors, Volumes 40–44 � � � � � � � � � � � � � � � � � � � � � � � � � � � 369
Errata
An online log of corrections to Annual Review of Biophysics articles may be found athttp://www.annualreviews.org/errata/biophys
vi Contents
Ann
u. R
ev. B
ioph
ys. 2
015.
44:1
67-1
86. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
Acc
ess
prov
ided
by
WIB
6014
- H
umbo
ldt U
nive
rsita
et z
u B
erlin
- H
U B
erlin
on
04/2
0/16
. For
per
sona
l use
onl
y.