Embed Size (px)
Citation preview

BIOTIC AND ABIOTIC INTERACTIONS IN CENTRAL TEXAS
STREAMS WITH SPECIAL REFERENCE
TO NAUCORIDAE (HEMIPTERA)
by
DAVID PATRICK HERRMANN, B.S.
A THESIS
IN
ENTOMOLOGY
Submitted to the Graduate Faculty of Texas Tech University in
Partial Fulfillment of the Requirements for
the Degree of
MASTER OF SCIENCE
Approved
Accepted
May, 1992

ACKNOWLEDGMENTS
I am indebted to my major advisors, Robert Sites and Michael Willig
for support throughout this project. Under their guidance and supervision,
numerous doors of opportunity have been opened for which I will always be
grateful. I also thank Harlan Thorvilson who served on my committee and
provided critical reviews of my work. I gratefully acknowledge the
following people for their instrumental field and laboratory assistance:
Jeffery Back, Leslie Baker, Daniel Brooks, Gerardo Camilo, Steven Cox,
Brian Croyle, Clara Langner, Richard Stevens and Judy York. Only through
their patience and diligence was this project completed successfully.
Anthony Adamo, James Herrmann, Becky Nichols, and James Suh provided
technical assistance for which I am most thankful. I also extend thanks to
Brant Baugh, John Cary, Thomas Doederlein, Dou Dou Faye, Doug Paxton,
Sergio Sanchez-Pena, and Clinton Smith for their friendships and assistance
with various chapters of this manuscript. Timothy Elkner and Dary 1
Moorhead critically reviewed portions of this manuscript. The staff of the
Texas Tech University Center at Junction (C. Len Ainsworth, Bill Durbin,
Martha Richardson, and Betty Street) provided logistic support during the
long summer months of field work. This project was supported by
Advanced Research Program grant 003644-081, State of Texas Higher
Education Coordinating Board.
11

TABLE OF CONTENTS
ACKNOWLEOOMENTS ....................................................................... 11
LIST OF TABLES .................................................................................. IV
LI~T OF ~<:J~E~ ················································································"i
I. IN'TRODUCTION ...................................................................... 1
o"~I1Vi~~ ............................................................................. 1
~tllcl)f Sit~s ........................................................................... ~
Stlldy Organisms ................................................................. 2
Literatllre Cited .................................................................... 4
II. A LABORATORY FLOW TANK WITH VARIABLE CURRENT AND DEPTHS FOR REPLICATIN<:J RIFFLES AND SHALLOW STREAMS ................................... 5
Abstract ................................................................................ 5
Introduction ......................................................................... 5
Materials and Methods ........................................................ 7
D. . ISCUS Sl on ......................................................................... .
Literature Cited ................................................................. .
ill. INFLUENCE OF C~RENT VELOCITY ON SUBSTRATUM SELECTION BY NAUCORIDAE (HEMIPTERA): AN EXPERIMENTAL APPROACH VIA
13
15
STRE-AM SIMULATION ......................................................... 16
Abstract .............................................................................. 16
Introduction ....................................................................... 17
Materials and Methods ...................................................... 19
Results ............................................................................... 21
D. . lSCUSSIOll •••••••••••••••••••••••••••••••••••••••••••••••••••••••••.•.•.•••••••••••• 26
Litemtllre Cited .................................................................. 31
lll

IV. DENSITY-DEPENDENT HABITAT SELECTION AND COMPETITION AMONG THREE SYMPATRIC NAUCORIDS ........................................................................... 35
Abstract .............................................................................. 35
In.troduction ....................................................................... 35
Materials an.d Methods ...................................................... 39
~~Slll~ •••••••••.••..•••••••••••••.••••••••••••••••••••.•••••••••••••••••••••••••••••• ~3
Discussion .......................................................................... 52
Litera.ture Cited .................................................................. 59
V. EFFECfS OF SUBSffiATUM SIZE AND HETE~OGENEITY ON MAC~OBENTHIC DIVE~SITY AND Cc:>~c:>~~c:>l'l .............................................................. ()~
Abstract .............................................................................. 63
In.troduction ....................................................................... 64
Materials an.d Methods ...................................................... 66
~esults an.d Discussion ...................................................... 68
Literature Cited .................................................................. 85
IV

LIST OFT ABLES
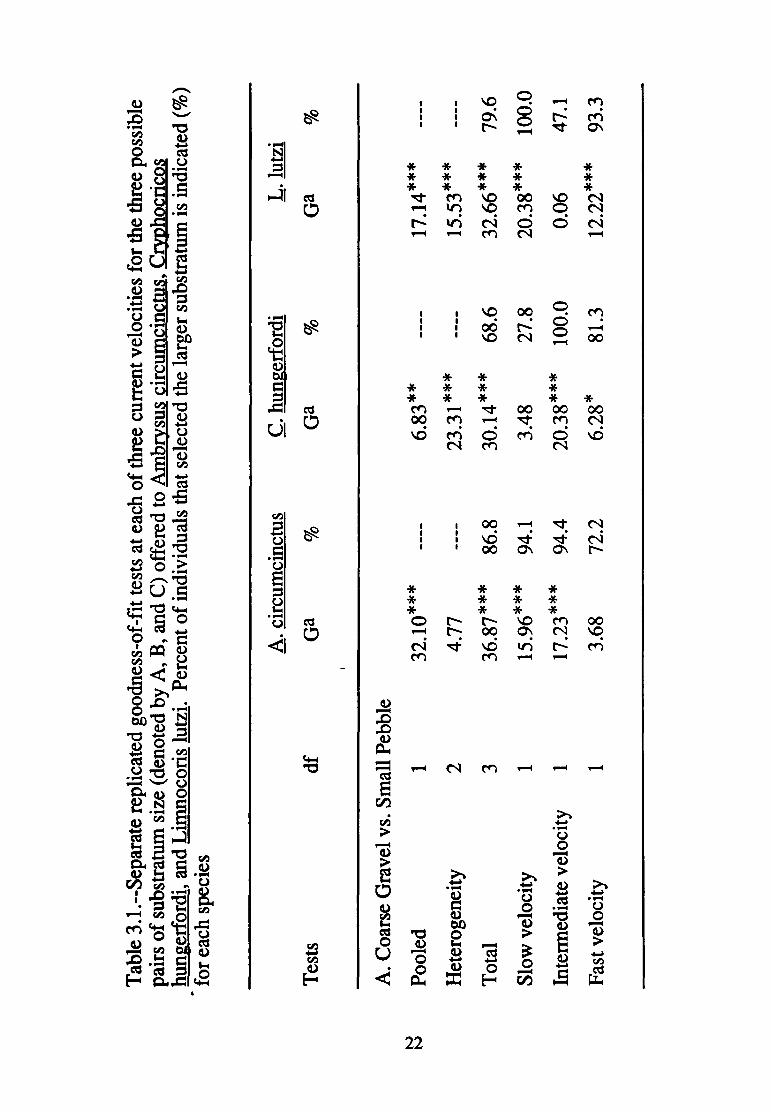
3.1--Separate replicated goodness of fit tests at each of three current velocities for the three possible pairs of substratum size (denoted by A, B, and C) offered to Ambrysus circumcinctus, Cryphocricos hun~erfordi, and Limnocoris lutzi .......................... 22
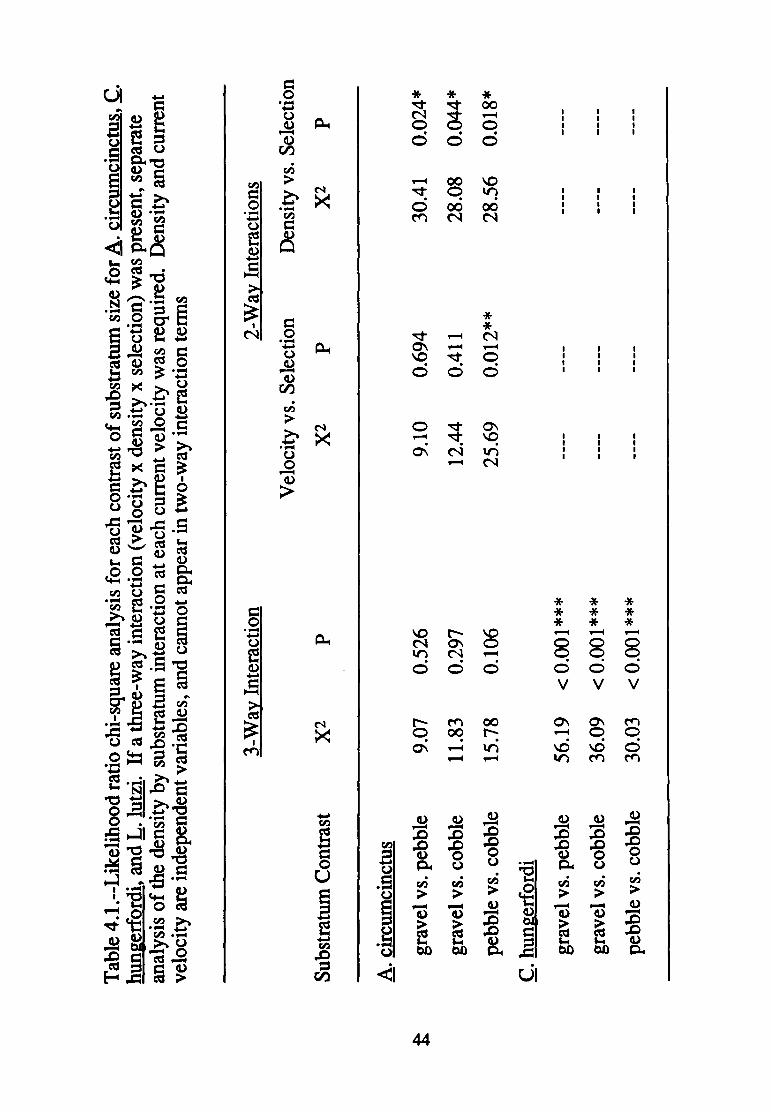
4.1--Likelihood ratio chi-square analyses for each contrast of substratum size for A. circumcinctus, .C. hun~erfordi, and L. l11~i ........................................................................................... ~~
4.2--Likelihood ratio chi-square analyses for the two-way interaction of density and substratum selection at each of three current velocities for experiments in which a significant three-way interaction (velocity x density x substratum) was detected ......................................................................................... 46
4.4--A priori comparisons of substratum selection for different density treatments based upon Z-scores (SPSS Inc., 1988) ........... 4 7
5.1--Abiotic conditions (mean+ S.D.) at South and North Llano River sites (N=25) ............................................................... 69
5.2--Mean density(+ S.E.) of macro invertebrate taxa (per 0.09 m2 found in the South and North Llano Rivers according to substrate size (CG, coarse gravel; SP, small pebble; SC, small cobble; HM, heterogeneous mix) (n=75) ............................ 70
5.3--Comparisons of species diversity between all possible pairs of four substratum types in each of two different rivers South, (SLR) and North, (NLR) Llano River ............................... 80
v

LIST OF FIGURES
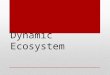
2.1--Side view depicting overall format of the artificial stream. Water is cycled through the pump within the lower reservoir into the upper reservoir .......................................... 8
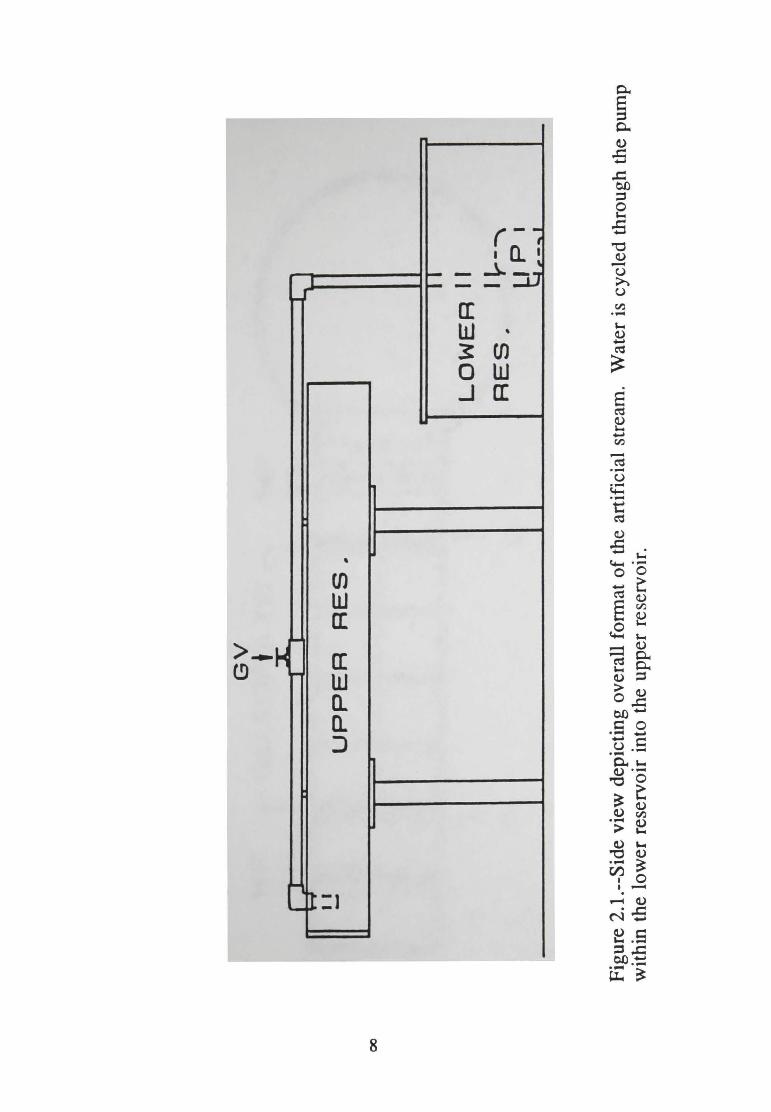
2.2--Plan view showing components (quadrats, weir, and screen frames) within the upper reservoir. Abbreviations are: GV= gate valve, WF=weir frame, S=screen, P=pump ............................. 9
2.2--Frontal and rear views of the upper reservoir showing weir used to raise the water level and screen for retaining organisms in the upper reservoir ..................................................................... 11
2.3--The relationship between the lower weir height and current velocity .............................................................................. 14
V1

CHAPTER I
INTRODUCTION
Overview
Ecology focuses on the factors which influence the structure and
function of ecosystems. In freshwater systems, many species exhibit a
benthic mode of existence (Ward, 1992). Therefore, it is not surprising
that the substratum has been considered a major determinant of the
distribution and abundance of macroinvertebrates (Minshall, 1984). Recent
studies have questioned the importance of the substratum (Statzner et al.,
1988), in tum focusing on hydraulic attributes as primary factors that
influence community function and structure. Nonetheless, current and
substratum are correlated in natural systems (Reice, 1980) and may have
independent effects on species microdistributions. Only rigorous
experimentation and hypotheses testing can elucidate the importance of
each factor separately, thereby facilitating a fundamental understanding of
complex interactions occurring in lotic systems.
This thesis comprises four manuscripts, each of which deals directly
or indirectly with microhabitat selection of stream invertebrates with
particular reference to creeping water bugs (Naucoridae). The first
manuscript (Chapter II) is methodological and concerns the simulation of
lotic environments in laboratory situations. The paper briefly reviews
problems associated with riffle simulation (including current limitations),
introduces potential solutions, and describes a simple yet flexible design for
use in ecological studies. Subsequent experiments within the thesis use this
methodology, at least in part. In the second manuscript (Chapter ill),
several experiments examined the influence of current velocity on
1

substratum selection by three riffle-inhabiting predators (Hemiptera:
Naucoridae) of central Texas streams. The experiments were aimed at
testing the hypotheses that current velocity mediates macrobenthic
microdistributions. The third paper (Chapter IV) builds upon the results
obtained in Chapter m, and describes additional laboratory experiments
that evaluated intra- and interspecific competition for space among
naucorids. Finally, the fourth manuscript (Chapter V) expands the domain
of the thesis to include entire macro benthic assemblages, and evaluates the
importance of substratum size and heterogeneity on species diversity and
taxonomic composition. This thesis illustrates the complexity of biotic and
abiotic interactions occurring in stream communities and exemplifies the
usefulness of a quantitative approach in lotic ecology.
Study Sites
The South and North Llano Rivers are located on the Edwards
Plateau in central Texas. Naucorids for stream simulation were collected
from the former; whereas, both are considered and compared in the
chapter dealing with substratum effects on community organization.
Appropriate details of the study sites are included in the materials and
methods of each chapter.
Study Or&anisms
Creeping water bugs (Hemiptera: Naucoridae) are predacious
aquatic insects common in lentic and lotic habitats of the southwestern U.S.
and tropical regions of the world. Although these insects are considered
keystone consumers in many aquatic systems (Sites and Willig, 1991 ), little
is known of their basic biologies (Gonsoulin, 1973; Constantz, 1974; Stout,
2

1978; Venk.atesan and Cloarec, 1988). As a consequence, the role of biotic
and abiotic interactions are evaluated on substratum selection by the three
common riffle inhabiting species, Ambr.ysus circumcinctus (Montandon,
Cryphocricos hun&erfordi (Usinger}, and Limnocoris lutzi (La Rivers},
and entire macroinvertebrate assemblages.
3

Literature Cited
Constantz, G. D. 1974. The mating behavior of a creeping water bug, Ambr.ysus occidenta}is (Hemiptera: Naucoridae ). Am. Midi. Nat. 92:234-239.
Gonsoulin, G. J. 1973. Seven families of aquatic and semiaquatic Hemiptera in Louisiana. Entomol. News 84:83-88.
Minshall, G. W. 1984. Aquatic insect-substratum relationships, pp. 358-400. In V. H. Resh and D. M. Rosenberg (eds.). The ecology of aquatic insects. Praeger, New York.
Reice, S. R. 1980. The role of substratum in benthic macroinvertebrate microdistribution and litter decomposition in a woodland stream. Ecology 61:580-590.
Sites, R. W. and M. R. Willig. 1991. Microhabitat associations of three sympatric species of Naucoridae (Insecta: Hemiptera). Environ. Entomol. 20:127-134.
Statzner, B., J. A. Gore and V. H. Resh. 1988. Hydraulic stream ecology: observed patterns and potential applications. J. N. Am. Benthol. Soc. 7:307-360.
Stout, R. J. 1978. Migration of the aquatic hemipteran Limnocoris insularis (Naucoridae) in a tropical lowland stream (Costa Rica, Central America). Brenesia 14:1-11.
Venkatesan, P. and A. Cloarec. 1988. Density dependent prey selection in llyocoris (Naucoridae). Aquat. Insec. 10:105-116.
Ward, J. V. 1992. Aquatic insect ecology. 1. Biology and habitat. Wiley and Sons, Inc. New York.
4

CHAPTER IT
A LABORATORY FLOW TANK WITH VARIABLE
CURRENT AND DEPTHS FOR REPLICATING
~ESANDSHALLOWSTREAMS
Abstract
Laboratory streams have been used for ecological studies of lotic
organisms, providing a means to determine the relative importances of
physical factors in structuring communities. Herein, an inexpensive
( <$500) artificial stream is described that has several advantages over other
designs; it offers: (1) accurate current velocities of 0 - 45.5 em/sec, (2) a
large surface area for experimentation, (3) both spatial and temporal
current homogeneity, and ( 4) riffle simulation. This artificial stream
requires approximately 1.8 m2 of floor space and can be used indoors.
Introduction
Studying the behavior of benthic animals is aided by laboratory
studies (Cummins, 1962). For example, the association between behavior
and abiotic factors can be determined by individually varying factors that
are not independent in natural situations. Such experiments are
fundamental to studies of interspecific interaction (competition or
predation) or habitat selection.
Artificial streams provide a means to experimentally modify a
number of parameters which are correlated in nature or would require
extensive modification of natural systems. The options in artificial stream
design include open or closed systems (Gee and Bartnik, 1969). Open
systems require constant input of new water and are useful in observing the
5

spawning behavior of fish (Gee and Bartnik, 1969); however, it is difficult
to suppress environmental influences and this design includes many natural
constraints. The closed system, in which water is internally circulated, is
often used in aquatic insect studies and can be used to determine which
factors affect behavior and distribution of macrobenthos. Closed systems
have been used to rear insect larvae (Sudia, 1951; Mason and Lewis, 1970),
determine the effects of siltation on the microdistributions of stream insects
(Cummins and Lauff, 1969), evaluate the effects of substratum type on
prey vulnerability (Fuller and Rand, 1990), and determine the effects of
population density on the emigration rates of aquatic larvae (Wiley, 1981 ).
Proper simulation in closed systems requires an appropriate pump
mechanism. For example, air-powered (Mackay, 1981) and propeller
powered designs (Hartman, 1965; Vogel and LaBarbera, 1978) have been
used; however, spatially uniform currents were difficult to obtain.
Techniques can be used to minimize heterogeneous currents and
turbulence. Lauff and Cummins (1964) developed an excellent design that
produced uniform currents; however, velocities were apparently limited
(see Cummins and Lauff, 1969). Thus, design problems are two-fold:
either spatial uniformity is not achieved, or the potential for a realistic
range of current velocities is compromised by the design.
In contrast, a water-fall system provides a swift, homogeneous
current in which water travels along an upper tank, falls into a reservoir,
and is recycled into the upper tank. There has been reluctance to use
water-fall systems because the pumps required for such designs generally
heat the water and produce appreciable turbulence (Vogel and LaBarbera,
1978). The water-heating problem can be minimized by modifying the
design to function as a radiator. This is accomplished by exposing the
6

water within the upper reservoir to a large surface area, in tum, allowing
for the loss of excessive heat. Turbulence can be minimized by the
addition of a weir located near the site of water introduction in the upper
tank.
Materials and Methods
The artificial stream comprises three main components: ( 1) a
rectangular plywood trough (upper reservoir) supported by four wooden
posts, (2) a cylindrical galvanized tin tank (lower reservoir), and (3) a
submersible sewage pump with plumbing. Water is lifted from the lower
reservoir to the upstream end of the upper reservoir, where it then empties
and travels through the upper reservoir. The water then falls into the
lower reservoir and is recycled. Current velocities are manipulated with a
gate valve and stream depths with a lower weir.
Upper reservoir
The upper reservoir (243.8 x 60.1 x 30.5 em) was constructed from
1.9 em (3/4 in.) plywood (Figure 2.1). Galvanized nails and wood glue
were used to secure the sides of the upper reservoir to its base. Silicon
putty prevented leakage from joints and nail holes. The entire upper
resetvoir was coated with a potable epoxy. Fiberglass may be substituted
for the epoxy. If a potable epoxy is used, the upper reservoir might
require a grade-4 plastic lining to prevent possible leakage. The upper
reservoir stream path was divided into eight quadrats (30.5 x 30.5 em)
with 2.5 em (1 in.) comer molding used as a partition (Figure 2.2). Two
7
00
-G
V
1 r
I I
I I
""""
~
I I -
UP
PE
R
RE
S.
I I
LO
WE
R I
I
RE
S.
I~-,
~~-:
Fig
ure
2.1.
--S
ide
view
dep
icti
ng o
vera
ll fo
rmat
of
the
arti
fici
al s
trea
m.
Wat
er is
cyc
led
thro
ugh
the
pum
p w
ithi
n th
e lo
wer
res
ervo
ir i
nto
the
uppe
r re
serv
oir.
\0
WF n
~ Q
UA
DR
AT
Sl
t s t s
WF
n II
II
" II " " \\ \\
Fig
ure
2.2.
--P
lan
view
sho
win
g co
mpo
nent
s (q
uadr
ats,
wei
r, a
nd s
cree
n fr
ames
) w
ithi
n th
e up
per
rese
rvoi
r.
Abb
revi
atio
ns a
re:
GV
= g
ate
valv
e, W
F =
wei
r fr
ame,
S =
scre
en,
P =
pu
mp
.

61.0 x 30.5 em fiberglass screens (1 mm2 mesh) were used to retain
organisms within the quadrats, collect debris, and prevent experimental
materials from entering the lower reservoir (Figure 2.2). Screens were
located 30.5 em from each end of the upper reservoir, and were attached
vertically to frames constructed from 2.5 em (1 in.) corner molding
(Figure 2.2) to provide stabilization.
Water level manipulations and the production of homogeneous
currents were accomplished by the addition of two weirs. The upstream
weir was located 29 em from the farthest upstream position, and the second
was located immediately downstream from the downstream screen frame.
The weir at the upper end (61.0 x 1.9 x 10.2 em) reduced turbulence
caused by the introduction of water at the upstream position; whereas, the
second weir (61.0 x 1.9 x 5.1 em) controlled water depth. Variation in
current depth was controlled by using lower weirs of different heights.
Weirs were stabilized by two wooden frames (60.1 x 5.1 x 2.5 em)
positioned 2.5 em apart. Wooden posts (Figure 2.3) (10.2 x 10.2 x 75.0
em) attached beneath the upper reservoir provided support. The slope of
the stream path could be manipulated by changing the heights of these
supports.
Lower reservoir
The lower reservoir consisted of a galvanized tin tank ( 119.4 em
diam, 61.0 em height) with a volume capacity of 2.73 m3. This reservoir
was the ideal size for the pump described herein; however, more powerful
pumps may be used to increase the current velocity, and in such cases the
10

- -
fi
I
Fig
ure
2.3.
--F
ront
al a
nd r
ear
view
s o
f up
per
rese
rvoi
r sh
owin
g w
eir
used
to
rais
e th
e w
ater
lev
el a
nd
scre
en f
or r
etai
ning
org
anis
ms
in t
he u
pper
res
ervo
ir.
The
site
of
wat
er i
ntro
duct
ion
into
the
upp
er
rese
rvoi
r is
clo
sed
at t
he u
pstr
eam
pos
itio
n, t
here
by f
orci
ng w
ater
to t
rave
l ov
er th
e ex
peri
men
tal
area
to
war
d th
e lo
wer
res
ervo
ir w
here
the
wat
er is
rec
ycle
d.

lower reseiVoir size should change correspondingly.
A 0.5 Hp submersible sewage pump (Goulds, model, 3887, Seneca
Falls, N.Y.) was positioned within the lower reseiVoir (Figure 2.1). A 5.1
em diam (2 in.) PVC pipe, including two elbow fittings (Figure 2.1),
carried water from the pump to the upstream position of the upper
reseiVoir. A brass gate valve was inserted along the horizontal length of
the pipe, and regulated current velocity. All pipe joints were sealed with
PVC pipe cement.
Performance
To obtain accurate current velocities, the lower reseiVoir was placed
behind the upper reseiVoir (opposite the normal position), filled with water,
and depth recorded. Water was then pumped into the upper reseiVoir until
the water level in the lower reseiVoir fell immediately below the intake
level of the pump. Depth was measured (while the pump was running) in
the upper reseiVoir at the position in the stream path for which a known
current velocity was required. The current velocity was then calculated as:
V = m2~hfta
where V =current velocity, r =radius of the lower reseiVoir, ~=change
in the water depth of the lower reseiVoir, t = time the pump was on, and a
= cross-sectional area of the current. After six trials this system produced
an average maximum velocity (+ SD) of 45.45 + 2.73 em/sec . With a 5.1
em lower weir, an average maximum (+ SD) of 13.7 + 0.5 em/sec current
velocity was obtained. This velocity could then be further reduced to 0.0
em/sec without harm to the pump by closing the gate valve.
12

Discussion
This system is inexpensive ($460.00 in 1990) and can be constructed
by one person in two days. The system can be used indoors or outdoors,
but indoor usage minimizes temperature fluctuations. Water temperature
should be monitored with a maximum minimum thermometer. The
average water temperature after 32 hrs of continuous use was 29.1 °C
(constant room temperature of 25°C). The 4.1 oc temperature difference is
attributable to electrical resistance within the pump. This temperature may
not be suitable for some research; however, cooling devices are available
and can be placed within the lower reservoir. Minimal temperature
elevation is obtained by maintaining the lower reservoir at maximum
capacity.
For studies in toxicology or those in which sensitive stream
organisms are used, it is important that the upper reservoir be coated with
a non-toxic (e.g., fiberglass) substance to prevent intoxication of the biota.
In addition, experiments should be designed that simulate depths and
current velocities that correspond to the natural conditions encountered by
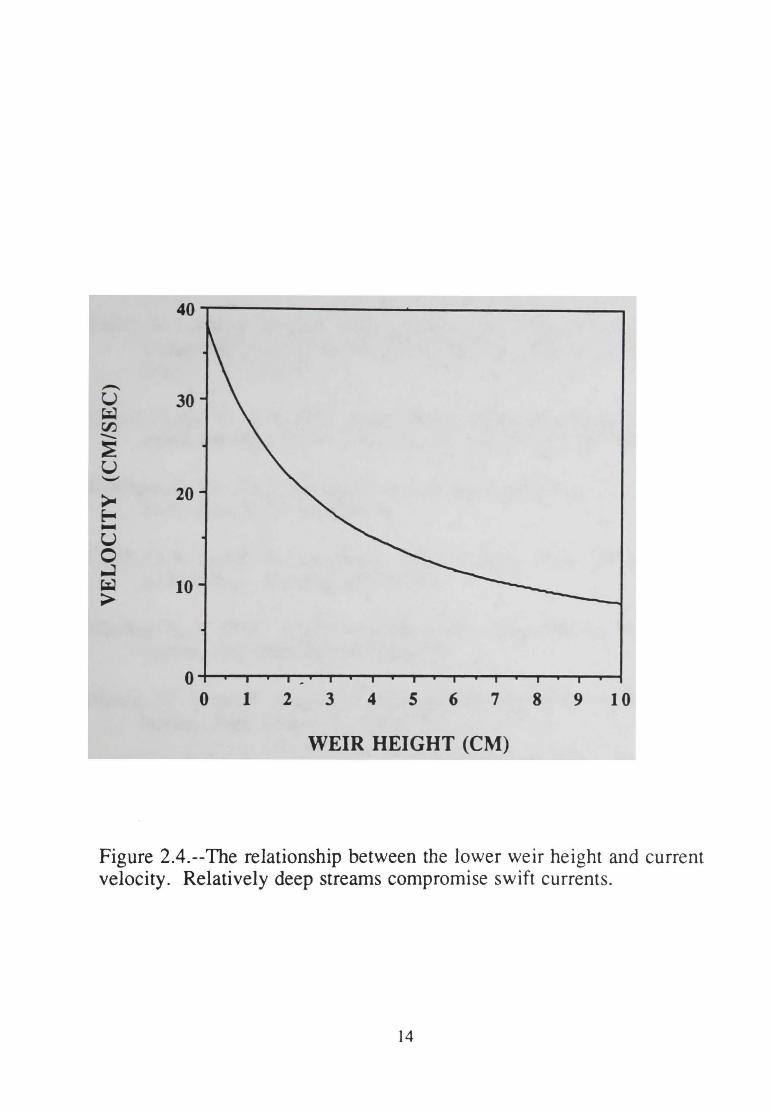
the study organism(s). Because current velocity is inversely proportional
to stream depth, relatively high lower weirs decrease the potential for swift
currents (Figure 2.4 ).
13
-u 30 ~ VJ -~ u ->- 20 E-..... u 0 .J ~ 10 >
0+---~~---r--~~~~---r--~~--~
0 1 2 3 4 s 6 7 8 9 10
WEIR HEIGHT (CM)
Figure 2.4.--The relationship between the lower weir height and current velocity. Relatively deep streams compromise swift currents.
14

Literature Cited
Cummins, K. W. 1962. An evaluation of some techniques for the collection and analysis of benthic samples with special emphasis on lotic samples. Am. Midi. Nat. 67:477-504.
Cummins, K. W. and G. H. Lauff. 1969. The influence of substrate particle size on the microdistribution of stream macrobenthos. Hydrobiologia 34:145-181.
Fuller, R. L. and P. S. Rand. 1990. Influence of substrate type on vulnerability of prey to predacious aquatic insects. J. N. Am. Benthol. Soc. 9:1-8.
Gee, J. H. and V. G. Bartnik. 1969. Simple stream tank simulating a rapids environment. J. Fish. Res. Bd. Canada 26:2227-2230.
Hartman, G. F. 1965. An aquarium with simulated stream flow. Am. Fish. Soc. Trans. 94:274-276.
Lauff, G. H. and K. W. Cummins. 1964. A model stream for studies in lotic ecology. Ecology 45: 188-191.
Mackay, R. J. 1981. A miniature laboratory stream powered by air bubbles. Hydrobiologia 83:383-385.
Mason, W. T. and P. A. Lewis. 1970. Rearing devices for stream insect larvae. Prog. Fish-Cult. 32:61-62.
Sudia, W. D. 1951. A device for rearing animals requiring a flowing water environment. Ohio J. Sci. 51:197-202.
Vogel, S. and M. LaBarbera. 1978. Simple flow tanks for research and teaching. Bioscience 28:638-643.
Wiley, M. J. 1981. Interacting influences of density and preference on the emigration rates of some lotic chironomid larvae (Diptera: Chironomidae). Ecology 62:426-438.
15

CHAPTER ill
INFLUENCE OF CURRENT VELOCITY ON SUBSTRATUM
SELECITON BY NAUCORIDAE (HEMIPTERA): AN
EXPERIMENTAL APPROACH VIA
STREAM SIMULATION
Abstract
Microhabitat preferences of three sympatric naucorids
(Ambrysus circumcinctus Montandon, Limnocoris lutzi La Rivers,
and Cryphocricos hungerfordi Usinger) from the South Llano River
of central Texas were evaluated in laboratory experiments. Each
species was individually subjected to all paired contrasts produced
from three substratum size classes (coarse gravel, small pebble, and
small cobble). Within each contrast, three trials were conducted,
each involving a different current velocity (slow, 6.6 + 0.1 em/sec;
intermediate, 9.8 + 0.1 em/sec; and fast, 13.7 + 0.5 em/sec). The
gravel substratum was never preferred; moreover, preference in the
pebble versus cobble contrast, when present, was only shown for the
cobble substratum. Overall, A. circumcinctus exhibited consistent
substratum preferences (cobble > pebble > gravel) independent of
current velocity. Cryphocricos hungerfordi exhibited selectivity in
which cobble was preferred over either gravel or pebble substrata;
however, a synergistic effect between current velocity and
substratum size was detected in the gravel versus pebble contrast.
Finally, b lutzi exhibited the highest degree of velocity-dependent
substratum selection. The laboratory analyses corroborate field
investigations by Sites and Willig (1991) and offer insight into the
16

influence of interacting abiotic variables on the microdistribution of
Naucoridae.
Introduction
The theory of erosion-deposition (Moon, 1939) suggests that
both the substratum and current are directly correlated within the
physical stream environment. Therefore, it is difficult to assess the
individual effects of either current or substratum on a stream fauna
under natural conditions (Minshall, 1984). Nonetheless, both are
important abiotic factors that influence the microdistribution of
stream invertebrates in a variety of habitats (Cummins and Lauff,
1969; Mackay and Kalff, 1969; Harman, 1972; Allan, 1975; de
March, 197 6; Minshall and Minshall, 1977; Lamberti and Resh,
1979; Williams, 1980; Erman and Erman, 1984; Statzner et al.,
1988). Because of the close relationship between substratum type
and current velocity, any interpretation of the effect of one of these
abiotic factors which excludes the other would be unrealistic (Reice,
1980). Substratum characteristics and flow attributes should be
simultaneously controlled to assess the relative role of each in
affecting site selection and population density. Works by Cummins
and Lauff (1969), Wiley (1981), and Fuller and Rand (1990)
produced lotic environments within laboratory settings to observe
the manner in which abiotic variables influence stream biota. These
"laboratory streams" permit the control and manipulation of
important abiotic and biotic parameters while simultaneously
reducing confounding effects. Such simulation is ideal for studies of
habitat selection.
17

Creeping water bugs (Hemiptera: Naucoridae) are predacious
aquatic insects common in lentic and lotic habitats of the
southwestern U.S. and tropical regions of the world. Although these
insects are considered keystone consumers in many aquatic systems
(Sites and Willig, 1991 ), little is known of their biologies
(Gonsoulin, 1973; Constantz, 1974; Venkatesan and Cloarec; 1988).
Most literature concerning naucorids addresses taxonomic (e.g.,
Usinger, 1947; La Rivers, 1971; 1974; 1976; Polhemus and
Polhemus 1988) or biogeographic questions (e.g., La Rivers, 1951;
Davis, 1986). The U.S. contains five naucorid genera (Ambrysus,
Cryphocricos, Limnocoris, Pelocoris, and Usingerina) which
comprise 21 species. The South Llano River in central Texas
harbors eight species (Sites and Willig, 1991 ), the most species rich
assemblage known to occur in temperate regions.
Ambrysus is the most speciose naucorid genus within the U.S.
Ambrysus circumcinctus Montandon is the most abundant naucorid
in the South Llano River where it easily can be found under large
rocks in swift waters (Sites and Willig, 1991). Cryphocricos is
represented in the U.S. by only ,C. hungerfordi Usinger (Polhemus
and Polhemus, 1988); the South Llano River demarks the
northeastern-most known boundary (Sites, 1990). Cuticular
evidence suggests plastral respiration within this genus (Parsons and
Hewson, 197 4 ). This species is morphologically hydrodynamic, and
exploits swift, deep, benthic habitats which contain large stones
(Parsons and Hewson, 1974; Sites and Willig, 1991). In the U.S.,
Limnocoris is represented by only one species, L. lutzi La Rivers.
This species is considered a substratum generalist (Sites and Willig,
18

1991) and occurs in riffle habitats. Although little is known of the
ecology of this species, its abundance is correlated with stream depth
(Sites and Willig, 1991). Limnocoris lutzi is the smallest (adult size)
naucorid inhabiting the South Llano River. The basic microhabitat
associations of each of these three species were described by Sites
and Willig (1991) who stressed the importance of substratum size
and current velocity; however, they could not uncouple the two. The
purpose of this study was to evaluate the influence of current
velocity on substratum selectivity for each of these three naucorid
species.
Materials and Methods
An artificial stream (Figure 2.1 ), described in detail elsewhere
(Herrmann et al., 1992), was used to evaluate substratum preferences
of the three most common riffle-dwelling naucorids (A.
circumcinctus, !:. hungerfordi, and L. lutzi) of the South Llano
River (Sites and Willig, 1991). Each species (n=18) was offered all
paired contrasts of three substratum size classes: (1) coarse gravel
(8-16 mm rocks), (2) small pebbles (16-32 mm rocks), and (3) small
cobble (64-128 mm rocks) [Wentworth (1922) classification scheme
after Minshall (1984)]. Each of these contrasts(= experiments) was
replicated under three current velocities (mean+ S.D.): slow (6.6 +
0.1 em/sec), intermediate (9.8 + 0.1 em/sec), and fast (13.7 + 0.5
em/sec), which each represent different trials. In all cases, water
depth was held constant with a 5.1 em lower weir. Four identically
designed streams were used, and each trial was conducted in a
randomly assigned stream.
19

Each stream contained eight quadrats which were dusted with
a fme layer of washed sand (ca. 1 mm deep) before each trial. The
left upstream-most quadrat was packed loosely with the larger rock
class and the smaller rock class placed in the right upstream-most
quadrat. Thereafter, the two substratum size classes alternated in a
checkerboard pattern. This arrangement reduced the potential of
biased selection related to complex hydraulic features (see Statzner et
al., 1988).
Water from the South Llano River was used in all
experiments, changed biweekly, and replenished periodically to
compensate for evaporation. Before each trial, naucorids were
collected from a riffle in the South Llano River and placed in
holding containers for no more than 1 hr before use. Stream flow
was initiated at least 30 min before each trial to stabilize water
temperature. Eighteen individuals of a given species were released
into each stream and left undisturbed for 32 hrs, after which the
pumps were turned off and the lower weir removed. This allowed
water drainage and minimized naucorid movement. All insects were
then removed and their locations recorded. On average, 4.1% of all
individuals were found on the upper or lower retaining screens, but
these individuals were not included in any subsequent analyses (Lauff
and Cummins, 1964).
A replicated goodness-of-fit test based upon G-Statistics (Sokal
and Rohlf, 1981) was used to evaluate the hypothesis that each
species would occupy the two presented substrata equally in each
trial. For each paired contrast of substratum size, the three different
velocities constituted the replicates. A significant pooled effect in the
20

absence of significant heterogeneity indicated consistent selection of
one substratum size, regardless of current velocity. Significant
heterogeneity indicated substratum selection that was velocity
dependent. Examination of the significance of selection at each
velocity revealed whether the interaction involved only differences
in the magnitude of selection or changes in the direction of selection
(e.g., preference for the large substratum at the slow velocity, no
preference at the intermediate velocity, and preference for the
smaller substratum at the fast velocity). The absence of both
significant heterogeneity and a pooled effect indicated no substratum
preference at any current velocity.
Results
Ambrysus circumcinctus
The larger substratum was consistently preferred, regardless
of current velocity in all paired contrasts (Table 3.1) involving A.
circumcinctus. In particular, 86.8% of individuals preferred small
pebble to coarse gravel; 77.4% preferred small cobble to coarse
gravel and 70.4% of individuals preferred small cobble over small
pebble. Current velocity did not affect the magnitude or direction of
substratum selection in any case.
Cryphocricos hungerfordi
Substratum selection was complex for ~- hungerfordi. In the
gravel versus pebble contrast, significant heterogeneity occurred in
the presence of a significant pooled effect (Table 3.1A); this
21
N
N
Tab
le 3
.1.-
-Sep
arat
e re
plic
ated
goo
dnes
s-of
-fit
test
s at
eac
h o
f thr
ee c
urre
nt v
eloc
itie
s fo
r th
e th
ree
poss
ible
pa
irs
of s
ubst
ratu
m s
ize
(den
oted
by
A, B
, and
C)
offe
red
to A
mbr
.ysu
s ci
rcum
cinc
tus,
Czy
phoc
rico
s hu
nger
ford
i, a
nd L
imno
cori
s lu
tzi.
Per
cent
of
indi
vidu
als
that
sel
ecte
d th
e la
rger
sub
stra
tum
is i
ndic
ated
(%
) · f
or e
ach
spec
ies
A.
circ
umci
nctu
s
Tes
ts
df
oa
o/o
A. C
oars
e G
rave
l vs.
Sm
all P
ebbl
e
Poo
led
1 32
.10*
**
----
Het
erog
enei
ty
2 4.
77
----
Tot
al
3 36
.87*
**
86.8
Slo
w v
eloc
ity
1 15
.96*
**
94.1
Inte
rmed
iate
vel
ocit
y 1
17.2
3***
94
.4
Fas
t vel
ocit
y 1
3.68
72
.2
C. h
unge
rfor
di
oa
%
6.83
**
----
23.3
1 **
* ---
-30
.14
***
68.6
3.48
27
.8
20.3
8***
10
0.0
6.28
* 81
.3
L.
lutz
i
oa
17.1
4***
15.5
3***
32.6
6***
20.3
8***
0.06
12.2
2***
%
79.6
100.
0
47.1
93.3
Tab
le 3
.1
cont
inue
d
A.
circ
umci
nctu
s
Tes
ts
df
oa
%
B.
Coa
rse
Gra
vel v
s. S
mal
l Cob
ble
Poo
led
1 16
.77*
**
----
Het
erog
enei
ty
2 2.
40
----
N
Tot
al
3 19
.15*
**
77.4
U
l
Slo
w v
eloc
ity
1 5.
88*
77.8
Inte
rmed
iate
vel
ocit
y 1
2.04
66
.7
Fas
t vel
ocit
y 1
11.2
5***
. 88
.2
C. h
unge
rfor
di
oa
%
29.9
0***
---
-4.
99
----
34.9
0***
86
.3
17.2
3***
94
.4
14.7
0***
93
.8
2.97
70
.6
L.l
utzi
oa
1.05
0.28
1.33
0.25
1.01
0.07
%
57.4
53.6
62.5
53.3
Tab
le 3
.1
cont
inue
d
A. c
ircu
mci
nctu
s C
. hun
gerf
ordi
Tes
ts
df
a a
%
a a
%
C.
Sm
all P
ebbl
e vs
. Sm
all C
obbl
e
Poo
led
1 9.
23**
---
-11
.05*
**
----
Het
erog
enei
ty
2 5.
39
----
3.79
---
-N
T
otal
3
14.6
2**
70.4
14
.84
**
72.2
~
Slo
w v
eloc
ity
1 8.
73**
83
.3
0.22
55
.6
Inte
rmed
iate
vel
ocit
y 1
0.00
50
.0
5.88
* 77
.8
Fas
t vel
ocit
y 1
5.88
* 77
.8
8.73
**
83.3
asig
nifi
canc
e o
f G:
, P >
0.0
5; *
, 0.0
5 >
P >
0.0
1; *
*, 0
.01
> P
> 0
.001
; **
*, P
< 0
.001
L.l
utzi
a a
2.01
7.32
*
9.33
*
8.73
**
0.07
0.53
%
60.0
83.3
53.3
41.2

suggested that substratum selection depended upon current velocity.
Moreover, the direction of preference differed depending on current
velocity; in particular, the smaller substratum (coarse gravel) was
preferred at the slow velocity; whereas, the larger substratum (small
pebble) was preferred at the intermediate and fast velocities. In the
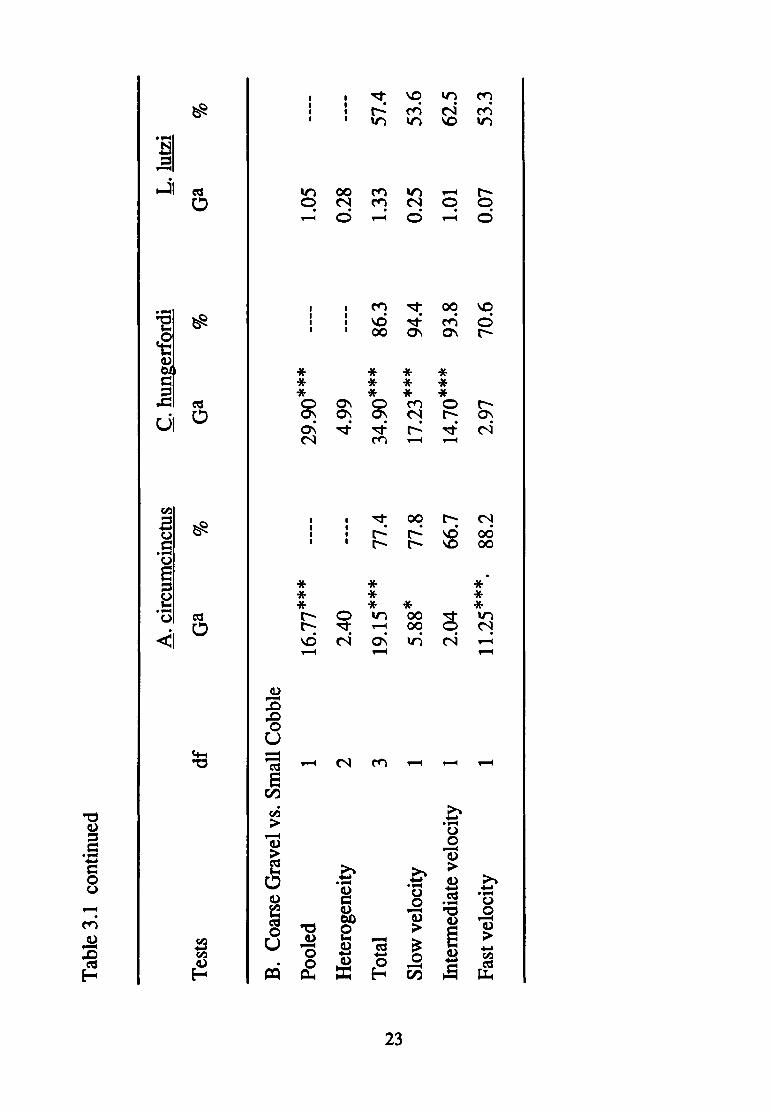
gravel versus cobble contrast {Table 3.1B), ~. hungerfordi exhibited
a consistent preference for the larger substratum (86.3% of all
individuals occurred in the cobble) over the smaller (gravel).
Finally, C. hungerfordi selected cobble over pebble (72.2% of
individuals) regardless of current velocity (Table 3.1C). Current
affected the direction of substratum selection for C. hungerfordi, but
only in the gravel versus pebble contrast. In the other contrasts,
neither the direction nor magnitude of substratum selection was
affected by current: preference in these latter two experiments was
consistently in the direction of the larger available substratum.
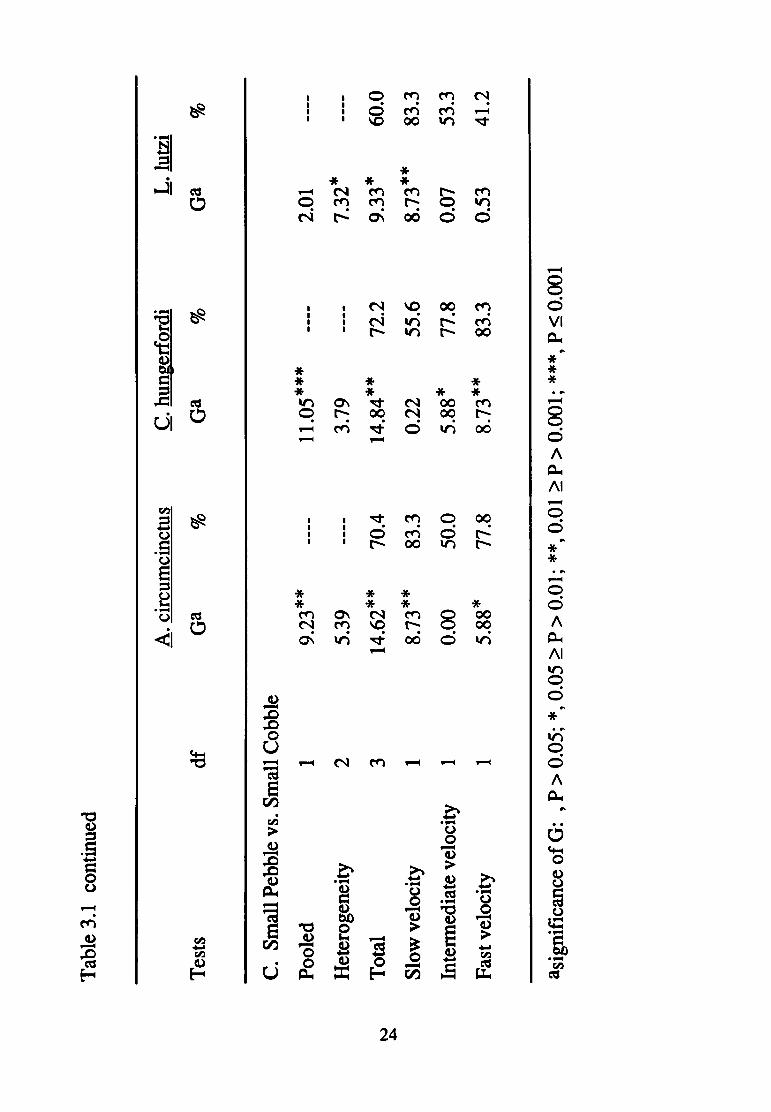
Limnocoris lutzi
Substratum selection varied with current velocity for L. lutzi
in the gravel versus pebble contrast {Table 3.1A). The larger
substratum (small pebble) was preferred at the slow and fast
velocities; whereas, no significant selection was apparent at the
intermediate current velocity. The significant pooled effect was
attributable to strong selection for the pebble substratum only during
two (slow and fast) of the three current trials, (>90%) which
overwhelmed the even distribution of individuals at the intermediate
velocity. The absence of significance for any test in the gravel
versus cobble contrast (Table 3.1B) indicated no substratum selection
25

at any velocity. Current velocity influenced the direction of
substratum selection in the pebble and cobble contrast (Table 3.1 C).
No selectivity could be detected at fast or intermediate velocities;
however, in the slow velocity, small cobble was preferred over small
pebble.
Discussion
Laboratory experiments concerning lotic organisms can
clarify and complement research conducted in field settings
(Cummins, 1962). Sites and Willig (1991) presented quantitative
analyses of microhabitat associations of each of these three naucorid
species; however, they did not uncouple the effects of the substratum
and velocity on microhabitat associations. Because our study
regulates both parameters independently, it refines our
understanding of specific substratum associations as potentially
modified by current speed. These experiments addressed the manner
in which current velocity could alter substratum selection. Velocity
preferences were not evaluated per se.
In each experiment involving A. circumcinctus, selection was
in the direction of the larger available rock class. This resulted in a
hierarchical arrangement of substratum selectivity (cobble > pebble
> gravel), which was unaltered by current velocity. In comparison,
Sites and Willig (1991) found that mean rock size as well as current
speed were significant variables influencing the microdistribution of
A. circumcinctus (mean rock size and mean current velocity in the
quadrats containing this species were found to be 184.7 mm and 37.8
em/sec, respectively). Rock size and current velocity in their field
26

situations exceeded those of our largest rock category (small cobble,
upper range of 128 mm) and the fastest current (13.7 em/sec).
Apparently current speeds up to 13.7 em/sec do not alter substratum
selectivity.
Cryphocricos hungerfordi often inhabits swift, deep waters in
which frequent surface visits to replenish a compressible air store
would cause downstream displacement. It has been shown that adult
C. hungerfordi and C. barozzii Signoret have cuticular modifications
necessary for plastral respiration (Parsons and Hewson, 197 4 ),
thereby eliminating the need for frequent surfacing and enabling
them to exploit habitats such as large rocks in swift currents. These
habitats are generally unavailable to species that utilize compressible
air bubbles. As a consequence, C. hungerfordi may be associated
with large rock sizes and swifter currents because it experiences
ecological release from interspecific competition with other
naucorids. A significant pooled effect existed in all three
experiments involving C. hungerfordi; however, a current
substratum interaction was observed between pebble and gravel in
which the direction of selection was affected by current. Strong
association with the larger of the available substrata occurs at fast
(81.3%) and intermediate (100%) velocities; however, this
relationship is vitiated by slow velocities at which no substratum
preference was shown. Similarly, Sites and Willig (1991) found that
both rock size and current velocity were important variables in
determining the microdistributions of .C.. hungerfordi in field
situations. Mean rock size in the quadrats containing this species was
189.0 mm, with an average current velocity of 37.2 em/sec. This
27

rock size and current velocity exceeded those tested herein. In our
experiments, preference only for the small cobble could be
delineated over the other substrata. At faster current velocities,
substratum-current interactions may become more prevalent and
might confound field interpretations.
Although L. lutzi has been viewed as a substratum generalist
with sensitivity to current velocity (Sites and Willig, 1991), it did
exhibit some preferential substratum selection in our study. In fact,
current velocity influenced substratum selection in the gravel versus
pebble contrast and in the pebble versus cobble contrast. Clearly,
current velocity plays an important role in modifying substratum
selection for this species, a view which did not emerge from a field
study in which their separate effects could not be evaluated.
Field observations suggest that niche partitioning is based
primarily on flow characteristics which separate the three naucorid
species tested here from the other five that occur in the South Llano
River (see Sites and Willig, 1991). The former three naucorid
species occur in association with rapidly flowing water, whereas the
other five species generally occur in shoreline and backwater
situations, frequently among algae and aquatic macrophytes.
Shoreline algae often grow among moderate to large-sized alluvium
as well as in silt, consequently substratum size exploited by those five
species is variable.
The fmdings in this study strongly paralleled those of Sites and
Willig (1991); however, important differences were observed. For
example, substratum size was relatively unimportant for L. hUti in
field situations; however, laboratory analyses suggested that at least
28

some consistent substratum bias existed. Similarly, field analyses
suggested an important role of current velocity on the
microdistribution of C. hun~erfordi. This was expected to occur
considering the respiratory adaptations of this species for swift
waters. Contrary to expectation, current velocity only minimally
influenced substratum selectivity in the laboratory. Ironically, this
occurred at rock sizes (gravel and pebble), which do not occur in
common habitat for C. hun~erfordi. In reality, a tradeoff may exist
between substratum size and current velocity. At small rocks sizes
the importance of substratum size is dependent upon current.
Conversely, at larger rock sizes, current does not alter the
preference for larger substrata.
Historically, stream ecologists have suggested that substratum
characteristics are primarily responsible for determining
microspatial arrangements of stream invertebrates [see Minshall
(1984) and citations therein]. However, more recent work (e.g.,
Statzner et al., 1988) has redirected attention to flow attributes as
primary factors affecting lotic community structure. Strong
evidence herein suggests that substratum selection by macrobenthic
predators varies interspecifically with respect to rock size and can be
altered by current velocity. The inability to uncouple the effects of
current velocity on substratum selection in natural environments,
where the two may be correlated, could give rise to erroneous
conclusions concerning the relative importance of hydraulic or
substratum attributes. Even in situations when current velocity may
not appear to directly affect the presence or absence of species, it
may have an indirect effect by modifying substratum selection.
29

Thus, the uncoupling of abiotic factors when determining
microhabitat selection of benthic organisms is imperative.
30

Literature Cited
Allan, D. J. 1975. The distributional ecology and diversity of benthic insects in Cement Creek, Colorado. Ecology 56:1040-1053.
Constantz, G. D. 1974. The mating behavior of a creeping water bug, Ambrysus occidentalis (Hemiptera: Naucoridae ). Am. Midi. Nat. 92:234-239.
Cummins, K. W. 1962. An evaluation of some techniques for the collection and analysis of benthic samples with special emphasis on lotic waters. Am. Midi. Nat. 67:477-504.
Cummins, K. W. and G. H. Lauff. 1969. The influence of substrate particle size on the microdistribution of stream macrobenthos. Hydrobiologia 34:145-181.
Davis, J. R. 1986. New distributional records of N aucoridae (Hemiptera) from the United States and Mexico, with morphological and taxonomic notes. Southwest. Nat. 31:535-539.
de March, B.G.E. 1976. Spatial and temporal patterns in macrobenthic stream diversity. J. Fish. Res. Board Can. 33:1261-1270.
Erman, D.C. and N.A. Erman. 1984. The response of stream macroinvertebrates to substrate size and heterogeneity. Hydrobiologia 108:75-82.
Fuller, R. L. and P. S. Rand. 1990. Influence of substrate type on vulnerability of prey to predacious aquatic insects. J. N. Am. Benthol. Soc. 9:1-8.
Gonsoulin, G. J. 1973. Seven families of aquatic and semiaquatic Hemiptera in Louisiana. Entomol. News 84:83-88.
Harman, W. N. 1972. Benthic substrates: Their effect on freshwater Mollusca. Ecology 53:271-277.
31

Herrmann, D.P., R. W. Sites and M. R. Willig. 1992. A laboratory flow tank with variable current and depths for replicating riffles and shallow streams. Texas J. Sci. 44:89-94.
Lamberti, G.A. and V. H. Resh. 1979. Substrate relationships, spatial distribution patterns, and sampling variability in a stream caddisfly population. Environ. Entomol. 8:561-567.
La Rivers, I. 1951. A revision of the Genus Ambrysus in the United States (Hemiptera: Naucoridae ). Univ. Calif. Pub I. Entomol. 8:277-338.
La Rivers, I. 1971. Studies of Naucoridae (Hemiptera). Memoir 2, Bioi. Soc. Nevada, Verdi.
La Rivers, I. 1974. Catalogue of taxa described in the family Naucoridae (Hemiptera) supplement no. 1: Corrections, emendations and additions, with descriptions of new species. Occas. Pap. 38, Bioi. Soc. Nevada, Verdi.
La Rivers, I. 1976. Supplement no. 2 to the catalogue described in the family Naucoridae (Hemiptera), with descriptions of new species. Occas. Pap. 41, Bioi. Soc. Nevada, Verdi.
Lauff, G. H. and K. W. Cummins. 1964. A model stream for studies in lotic ecology. Ecology 45:188-191.
Mackay, R. J. and J. Kalff. 1969. Seasonal variation in standing crop and species diversity of insect communities in a small Quebec stream. Ecology 50:101-109.
Minshall, G. W. 1984. Aquatic insect-substratum relationships, pp. 358-400. In V. H. Resh and D. M. Rosenberg (eds.). The ecology of aquatic insects. Praeger, New York.
Minshall, G. W. and J. N. Minshall. 1977. Microdistribution of benthic invertebrates in a Rocky Mountain (U.S.A.) stream. Hydrobiologia 55:231-249.
Moon, H. P. 1939. Aspects of the ecology of aquatic insects. Trans. Brit. Entomol. Soc. 6:39-49.
32

Parsons, M. C. and R. J. Hewson. 1974. Plastral respiratory devices in adult Cryphocricos (Naucoridae: Heteroptera). Psyche 81:510-527.
Polhemus, D. A. and J. T. Polhemus. 1988. Family Naucoridae Leach 1815: the creeping water bugs, pp. 521-527. InT. J. Henry and R. C. Froeschner (eds.). Catalogue of the Heteroptera, or true bugs, of Canada and the continental United States. Brill, Leiden.
Reice, S. R. 1980. The role of substratum in benthic macroinvertebrate microdistribution and litter decomposition in a woodland stream. Ecology 61:580-590.
Sites, R. W. 1990. Morphological variations in the hemelytra of Cryphocricos hungerfordi Usinger (Heteroptera: Naucoridae). Proc. Entomol. Soc. Wash. 92:111-114.
Sites, R. W. and M. R. Willig. 1991. Microhabitat associations of three sympatric species of Naucoridae (Insecta: Hemiptera). Environ. Entomol. 20:127-134.
Sokal, R. R. and F. J. Rohlf. 1981. Biometry: the principles and practice of statistics in biological research, 2nd ed. Freeman, San Francisco.
Statzner, B., J. A. Gore and V. H. Resh. 1988. Hydraulic stream ecology: observed patterns and potential applications. J. N. Am. Benthol. Soc. 7:307-360.
Usinger, R. L. 1947. Classification of the Cryphocricinae (Hemiptera: Naucoridae ). Ann. Entomol. Soc. Am. 40:329-343.
Venkatesan, P. and A. Cloarec. 1988. Density dependent prey selection in Dyocoris (N aucoridae ). Aquatic Insects 10:105-116.
33

Wentworth, C. K. 1922. A scale of grade and class terms for cladistic sediments. J. Geol. 30:377-392.
Wiley, M. J. 1981. Interacting influences of density and preference of the emigration rates of some lotic chironomid larvae (Diptera: Chironomidae ). Ecology 62:426-438.
Williams, D. D. 1980. Some relationships between stream benthos and substrate heterogeneity. Limnol. Oceanogr. 25: 166-172.
34

CHAPTER IV
DENSITY-DEPENDENT HABITAT SELECTION
AND COMPETITION AMONG THREE
SYMPATRIC NAUCORIDS
Abstract
The role of competition for space in structuring stream communities
remains unclear. Much of the evidence for competition in streams has been
derived from work concerning sessile or slow moving benthos. Relatively
few studies have focused on mobile predators which may compete in more
complex manners than their sessile counterparts. As such, three naucorid
species (Ambrysus circumcinctus Montandon, Cryphocricos hungerfordi
U singer, and Limnocoris lutzi La Rivers) were subjected to laboratory
experiments to determine the influence of intra- and interspecific
competitive interactions on habitat selection. Intraspecific experiments
evaluated the effects of density per se; whereas, interspecific experiments
evaluated the effects of competition between naucorids beyond those
associated with an increase in density. The results suggest that naucorids
respond to both density and interspecific competitors in terms of habitat
selection; however, these relationships often varied among substratum
comparisons, with current velocity and interspecifically. This variation
illustrates the complex nature of interacting variables in stream communities
and offers evidence that competition is one mediator of invertebrate habitat
associations.
Introduction
Much of contemporary ecology focuses on the factors which facilitate
the coexistence of ecologically similar species. A popular view holds that
35

disturbance and predation can prevent the over-exploitation of space by
suppressing the density of competitive dominants (Connell, 1978; Sousa,
1984 ), which in tum, frees resources for less dominant species (McAuliffe,
1984 ). Alternatively, competition may mediate species distributions and
alter an organism's perception of habitat suitability (Schoener, 1983;
Hemphill, 1991 ). However, the relevance of competition in nature has been
questioned (Connell, 1975; 1978; 1980; Pulliam, 1983) and viewed as an
mtermittent phenomenon. Nevertheless, the mechanistic roles of
competition in affecting the abundance and distribution of species warrant
continued exploration.
Within stream ecosystems, the potential contributions of the
substratum and current to community structure have been intensively
investigated (Minshall, 1984; Statzner et al., 1988; Ward, 1992).
Consequently, both are considered to be primary determinants of the
abundance and distribution of stream invertebrates. Macrobenthic
populations frequently exist at high densities and extensive overlap in
resource utilization occurs between potential competitors (Dudley et al.,
1990). Thus, intra- and interspecific competition for these resources as well
as space in general may contribute to the overall structure and function of
stream communities (Harding and Colbo, 1981; McAuliffe, 1984; Dudley et
al., 1986; Hemphill, 1988; 1991). In theory, the effects of these
contributions on community organization may differ fundamentally. An
increase in a population's density may be conducive to generalistic resource
utilization, whereas, the presence of interspecific competitors may force a
generalistic species to specialize on a limited range of resources.
Comparisons of intra- and interspecific competition may offer insight into
the coexistence and abundance of competitors. For example, if a
36

hypothetical species A is a superior competitor of species B, one might
conclude that A will eventually eliminate B, unless the process is interrupted
by other factors (Connell, 1983). If, however, species A is subjected to
intraspecific pressures greater than the interspecific pressures imposed by
species B, species A may be self-limited to a density which can not eliminate
species B (Connell, 1983).
Intraspecific competition for space may be considered to be density
dependent. Contemporary habitat selection theory based on Fretwell and
Lucas (1970) predicts that individuals select habitats based on fitness. As
the density of a species increases within a habitat, individual fitness
generally decreases and individuals may select what was initially suboptimal
habitat as a consequence of cost-benefit analyses. Evidence for intraspecific
competition in stream communities is variable. Caddisflies (Glass and
Bovbjerg, 1969) and black flies (Zahar, 1951) are thought to alter their
microdistributions through territorial spacing, whereas, chironomids respond
to elevated density via alteration of emigration rates (McLachlan, 1969).
Wiley (1981) established an interaction between sediment type and
chironomid density, even though emigration rates were unpredictable based
on either factor alone.
Several groups of sessile and slow moving benthos contribute the
majority of information on interspecific competition in stream systems.
Hemphill (1988) established a competitive dominance between Hydropsyche
oslari and Simulium vir~atum on the basis of former's ability to initiate
aggressive encounters and preempt boulder habitat. Competition between
the two species was less prevalent during winter months when floods
lowered the densities of Hydropsyche (Hemphill, 1991). McAuliffe (1983)
established a negative correlation between the colonization rates of Baetis
37

and Simulium and the density of the caddisfly Leucotrichia. Again,
disturbance mediated density reductions of Leucotrichia, thereby preventing
the monopolization of space (McAuliffe, 1984 ).
Many stream benthos that have been shown to compete, do so through
interference mechanisms including overgrowth, crushing, and undercutting
(Hemphill, 1991). Highly mobile species, which do not interact in those
manners have received little attention in terms of competition. Furthermore,
many studies have focused on species for which competition was expected
to occur. For this reason, the prevalence of competition in nature may be
overestimated and represent a strong bias (Connell, 1983). A wider range of
species must be examined to alleviate this concern.
Creeping water bugs (Naucoridae: Hemiptera) are prominent keystone
members of stream ecosystems in the southwestern U.S.; however, little is
known about their basic biology (Gonsoulin, 1973). Most studies of
naucorid ecology have focused on abiotic interactions. Stout (1981)
examined flooding effects associated with two tropical species, Limnocoris
insularis Champion and Czyphocricos latus Usinger. Sites and Willig (1991)
examined abiotic factors that contribute to the microdistributions of riffle
inhabiting naucorids (see also Chapter III). Relatively few studies have
focused on biotic interactions associated with the group. Of those, two
(V enkatesan and Cloarec, 1988; Camilo and Willig, unpublished data)
examined naucorid prey selection. The former examined density-dependent
prey selection in a European species, llyocoris cimicoides, and the latter
evaluated prey selection by three Texas species. Other ecological efforts
have focused on life histories (Constantz, 1974; McPherson et al., 1987;
Sites and Nichols, 1990); however, none has focused on competition among
naucorids. The South llano River (SLR) of central Texas harbors eight
38

naucorid species (Sites and Willig, 1991), which is the most species rich
assemblage known to occur in the U.S. The SLR lies on Cretaceous
limestone and is characterized by well-defined riffle and pool habitats.
Naucorids can be found within the substratum in these habitats and among
aquatic macrophytes along stream margins. These naucorids feed on a
variety of invertebrates including immature mayflies and caddisflies (Camilo
and Willig, unpublished data). Three species, Ambrysus circumcinctus
Montandon, Limnocoris lutzi La Rivers, and Cryphocricos hungerfordi
Usinger, are common riffle inhabitants which coexist at high densities and
exhibit partial overlap in substratum (Chapter ill), current (Sites and Willig,
1991) and prey (Camilo and Willig, unpublished data) utilization. The
degree to which these insects compete for spatial resources and the
mechanisms which allow them to coexist remains poorly understood. The
following study attempted to evaluate the effects of both density and
interspecific competition on habitat selection by the three naucorid species
and offer potential mechanisms by which they compete for spatial resources.
Materials and Methods
Laboratory Experiments
A method of stream simulation described in detail by Herrmann et al.
(1992) was used in laboratory experiments (see Chapter IT). The design
involved a simple water fall system that could produce current velocities
from 0-45 em/sec. Each stream contained four paired quadrats (30.5 X 30.5
em) in which substratum could be positioned. Each of three naucorid
species (A. circumcinctus, .C. hun~:erfordi and L.lmW was subjected to
every paired contrast of three substratum categories [from the Wentworth
39

(1922) classification scheme after Minshall (1984)]: (1) coarse gravel (CG;
8-16 mm rocks), (2) small pebble (small pebble; 16-32 mm rocks), and (3)
small cobble (SC; 64-128 mm rocks) within the streams. The amount of
substratum in each category was standardized according to volumetric
displacement (CG, 900 ml; SP, 1200 ml, SC, 1500 ml). For each contrast,
three trials were conducted, each at a different current velocity (slow, 6.6
em/sec; intermediate, 9.8 em/sec; and fast, 13.7 em/sec). To reduce the
potential for biased selection related to complex hydraulic features, the two
substratum categories in each replicate were alternated in a checkerboard
pattern. Water used in the experiments was collected from a riffle in the
SLR and current depth was standardized with a 2 em lower weir (see
Chapter II). Each trial was conducted for 32 hrs in one of 6 randomly
assigned streams. At the conclusion of each experiment, the location of each
individual naucorid was determined by visual inspection of the substrata.
Individuals found on the upper and lower retaining screens were discarded
and not included in subsequent analyses.
All naucorids were collected from various riffles in the SLR. Because
of the large numbers of individuals used in experimentation, most were
released after use; however, none was used more than once. To determine
the importance of density-dependent intraspecific interactions on substratum
selection, the aforementioned experiment was conducted at three densities
representing a geometric progression (N=18, N=36, N=54). To determine
the influence of interspecific interactions, experiments were conducted with
all possible pair-wise combinations of species [e.g., A. circumcinctus
(N=18) + k lutzi (N=18) or~- hun&erfordi (N=18) + L. lutzi (N=18)], and
one experiment involving all three species, each at N=18.
40

Statistical Analyses
A series of hierarchical log linear models (SPSS Inc., 1988) was used
to test hypotheses that three- and two-way interactions involving current
velocity, density treatment, and substratum selection were negligible for
each species. If significant three-way interactions occurred, then substratum
selection was density dependent in a manner that varied with current
velocity. Thereafter, the model was decomposed via a series of .a priori
contrasts to evaluate the manner in which competition occurred. If no three
way interaction was detected, that interaction term was not included in
subsequent models. A series of models then systematically evaluated two
way interactions. To test for a current velocity-substratum selection
interaction, that two-way interaction term was eliminated from the model.
Similarly, the two-way interaction between density and substratum selection
was tested by evaluating a model with the corresponding two-way
interaction term absent. If both possible two-way interactions were
significant, then substratum selection consistently varied with density
regardless velocity, and the effects of velocity on substratum selection were
consistent at all densities. As a consequence, only one series of .a priori
contrasts would be needed to determine the relationship between density and
substratum selection. If only a current velocity-substratum selection
interaction was significant, current velocity affected substratum selection in
the same way, regardless of density treatment. Thus, a priori contrasts were
not appropriate. If only a density-substratum selection interaction was
significant, it was concluded that density affected the way in which
substratum selection occurred, independent of current velocity.
Subsequently, i priori contrasts were implemented to compare substratum
selection at different densities. If neither three- nor two-way interactions
41

occurred, then substratum selection was considered independent of current
velocity and density regime.
If a density-substratum selection interaction was detected, the two
way interaction was resolved via non-orthogonal i! priori contrasts involving
Z scores (SPSS Inc., 1988). Two intraspecific comparisons of substratum
selection were conducted between: (1) experiments involving 18 and 36
individuals, and (2) experiments involving 18 and 54 individuals.
Comparisons of substratum selection when interspecific competitors were
present were of two types: those involving two species and those involving
three species. In all cases, density was constant within a particular contrast.
For example, substratum selection by A. circumcinctus when N=36 was
compared to substratum selection when N=18 for A. circumcinctus and
N=18 for ,C. hungerfordi. Similarly, substratum selection by A.
circumcinctus when N=36 was compared to substratum selection by A.
circumcinctus when its density and that of L.lutzi were each 18. In the
three species experiment, substratum selection for a focal species (N=54 ),
was compared to substratum selection by that same species when each of the
three species occurred at equal densities (N=18). For each comparison,
significance implied that doubling or tripling density, or that the presence of
interspecific competitors altered microhabitat associations. For the five D.
priori contrasts, comparison-wise error rate was held at 0.05; and not all
possible contrasts were made in order to reduce experiment-wise error rate.
Intraspecific comparisons evaluated density w g, whereas interspecific
comparisons held density constant but varied species composition.
42

Results
Ambrysus circumcinctus
For all three paired contrasts of substratum sizes, density affected
substratum selection in a consistent fashion, regardless of current velocity
(absence of three-way interaction with a significant density by substratum
interaction; Table 4.1 and Table 4.2). Nonetheless, A. circumcinctus
responded in a complex fashion to the various density treatments. In the
pebble vs. cobble contrast, velocity affected substratum selection in a
consistent fashion regardless of density. More powerful a priori contrasts
revealed the manner in which density affected substratum selection (Table
4.3). Increased density of conspecifics modified substratum selection only
in one of the pebble vs. cobble contrasts (18 vs. 36). The presence of L. lutzi
affected substratum selection by A. circumcinctus in all but the gravel vs.
pebble contrast; conversely, the joint presence of C. hungerfordi and L. lutzi
only affected substratum selection in the gravel vs. pebble contrast.
Cryphocricos hungerfordi
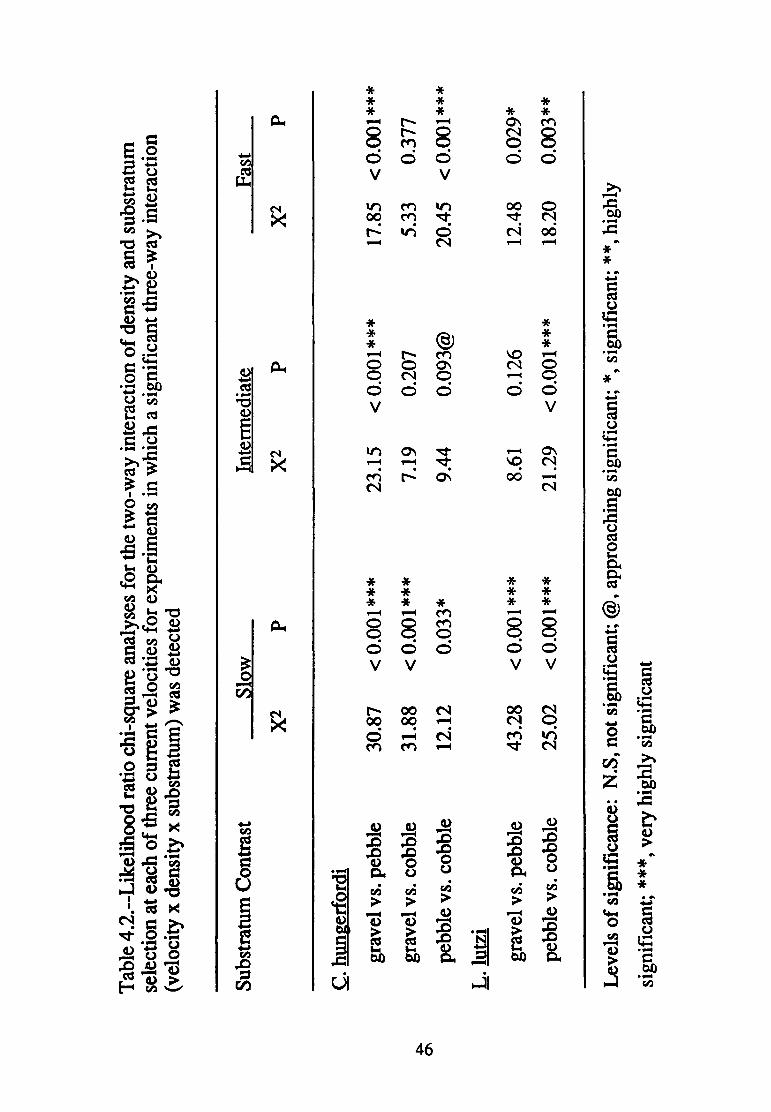
Velocity affected the manner in which density treatment modified
substratum selection in all three paired contrasts of substratum sizes
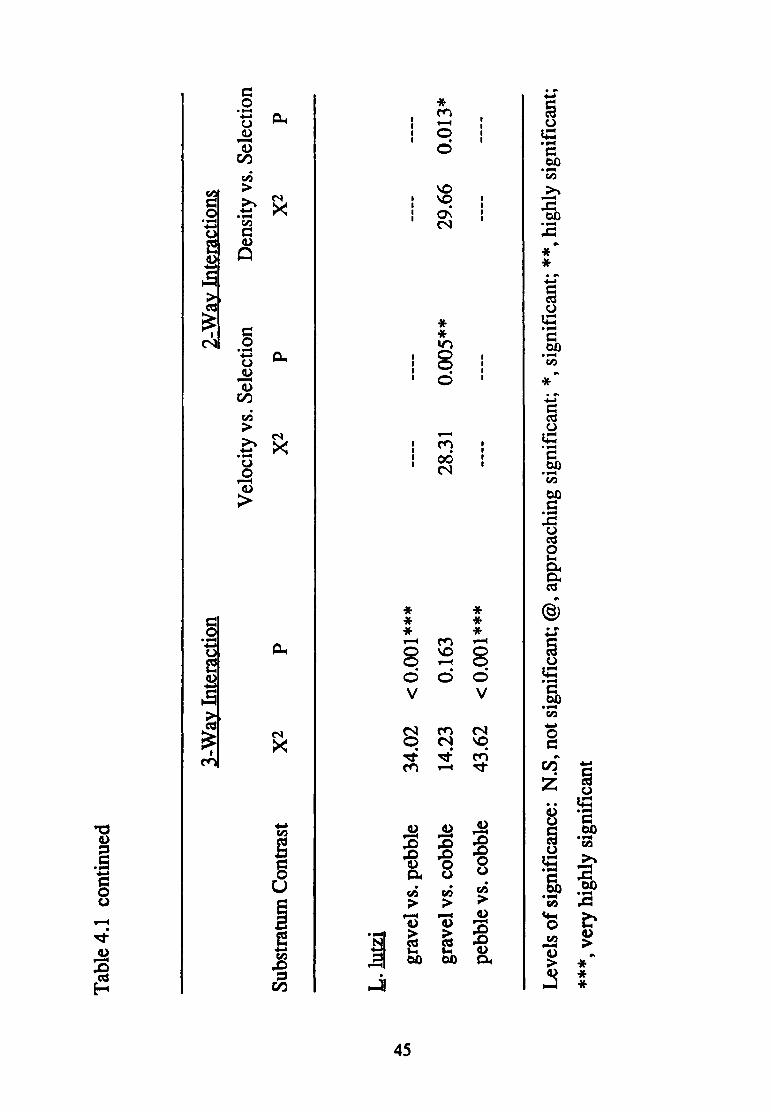
(presence of three-way interactions in each case; Table 4.1). Nonetheless,
differences in density treatment at least approached significance in
modifying substratum selection in all but the intermediate and fast velocity
experiments involving the gravel vs. cobble contrast (Table 4.2).
Again, patterns of significance related to density treatment are complex
(Table 4.3). Substratum selection was modified by increased numbers of
conspecifics at slow velocities in all but the pebble vs. cobble contrast,
43
t
Tab
le 4
.1.-
-Lik
elih
ood
rati
o ch
i-sq
uare
ana
lysi
s fo
r ea
ch c
ontr
ast o
f sub
stra
tum
siz
e fo
r A
. cir
cum
cinc
tus,
.C..
hung
erfo
rdi,
and
L. l
utzi
. H
a t
hree
-way
inte
ract
ion
(vel
ocit
y x
dens
ity
x se
lect
ion)
was
pre
sent
, sep
arat
e an
alys
is o
f the
den
sity
by
subs
trat
um i
nter
acti
on a
t eac
h cu
rren
t vel
ocit
y w
as r
equi
red.
D
ensi
ty a
nd c
urre
nt
velo
city
are
ind
epen
dent
var
iabl
es, a
nd c
anno
t app
ear
in tw
o-w
ay i
nter
acti
on te
rms
3-W
ay I
nter
acti
on
2-W
ay I
nter
acti
ons
Vel
ocit
y vs
. S
elec
tion
D
ensi
ty v
s. S
elec
tion
Sub
stra
tum
Con
tras
t X
2 p
X2
p X
2 p
A. c
ircu
mci
nctu
s
grav
el v
s. p
ebbl
e 9.
07
0.52
6 9.
10
0.69
4 30
.41
0.02
4*
grav
el v
s. c
obbl
e 11
.83
0.29
7 12
.44
0.41
1 28
.08
0.04
4*
pebb
le v
s. c
obbl
e 15
.78
0.10
6 25
.69
0.01
2**
28.5
6 0.
018*
C. h
une:
erfo
rdi
grav
el v
s. p
ebbl
e 56
.19
< 0
.001
***
grav
el v
s. c
obbl
e 36
.09
< 0
.001
***
pebb
le v
s. c
obbl
e 30
.03
< 0
.001
***
~
VI
Tab
le 4
.1
cont
inue
d
3-W
ay I
nter
acti
on
2-W
ay I
nter
acti
ons
Vel
ocit
y vs
. S
elec
tion
D
ensi
ty v
s. S
elec
tion
Sub
stra
tum
Con
tras
t X
2 p
X2
p X
2 p
L.m
w gr
avel
vs.
peb
ble
34.0
2 <
0.0
01 *
**
grav
el v
s. c
obbl
e 14
.23
0.16
3 28
.31
0.00
5**
29.6
6 0.
013*
pebb
le v
s. c
obbl
e 43
.62
< 0
.001
***
Lev
els
of s
igni
fica
nce:
N
.S, n
ot s
igni
fica
nt;@
, app
roac
hing
sig
nifi
cant
; *,
sign
ific
ant;
**,
high
ly s
igni
fica
nt;
***,
ver
y hi
ghly
sig
nifi
cant
~
0\
Tab
le 4
.2.-
-Lik
elih
ood
rati
o ch
i-sq
uare
ana
lyse
s fo
r th
e tw
o-w
ay i
nter
acti
on o
f den
sity
and
sub
stra
tum
se
lect
ion
at e
ach
of t
hree
cur
rent
vel
ocit
ies
for
expe
rim
ents
in
whi
ch a
sig
nifi
cant
thre
e-w
ay i
nter
acti
on
(vel
ocit
y x
dens
ity
x su
bstr
atum
) w
as d
etec
ted
Sub
stra
tum
Con
tras
t S
low
In
term
edia
te
Fas
t
X2
p X
2 p
X2
.C. h
unge
rfor
di
grav
el v
s. p
ebbl
e 30
.87
< 0
.001
***
23
.15
< 0
.001
***
17
.85
grav
el v
s. c
obbl
e 31
.88
< 0
.001
***
7.
19
0.20
7 5.
33
pebb
le v
s. c
obbl
e 12
.12
0.03
3*
9.44
0.
093@
20
.45
L.h
!W
grav
el v
s. p
ebbl
e 43
.28
< 0
.001
***
8.
61
0.12
6 12
.48
pebb
le v
s. c
obbl
e 25
.02
< 0
.001
***
21
.29
< 0
.001
***
18
.20
Lev
els
of s
igni
fica
nce:
N
.S, n
ot s
igni
fica
nt;@
, app
roac
hing
sig
nifi
cant
; *,
sign
ific
ant;
**, h
ighl
y
sign
ific
ant;
***
,ver
y hi
ghly
sig
nifi
cant
p
< 0
.001
***
0.37
7
< 0
.001
***
0.02
9*
0.00
3**
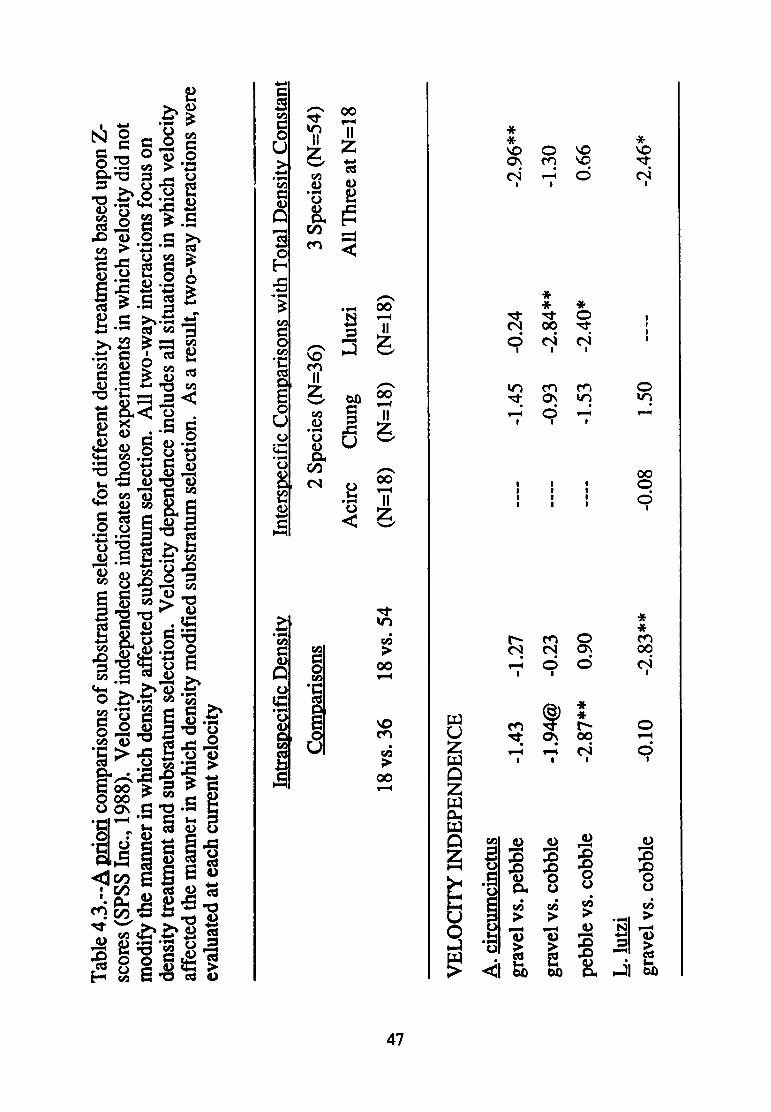
:!j
Tab
le 4
.3.-
-A p
rior
i com
pari
sons
of
subs
trat
um s
elec
tion
for
dif
fere
nt d
ensi
ty t
reat
men
ts b
ased
upo
n Z
sc
ores
(S
PS
S I
nc.,
1988
). V
eloc
ity
inde
pend
ence
ind
icat
es th
ose
expe
rim
ents
in
whi
ch v
eloc
ity
did
not
mod
ify
the
man
ner i
n w
hich
den
sity
aff
ecte
d su
bstr
atum
sel
ecti
on.
All
two-
way
int
erac
tion
s fo
cus
on
dens
ity
trea
tmen
t and
sub
stra
tum
sel
ecti
on.
Vel
ocit
y de
pend
ence
inc
lude
s al
l si
tuat
ions
in w
hich
vel
ocit
y af
fect
ed th
e m
anne
r in
whi
ch d
ensi
ty m
odif
ied
subs
trat
um s
elec
tion
. A
s a
resu
lt,
two-
way
inte
ract
ions
wer
e ev
alua
ted
at e
ach
curr
ent v
eloc
ity
Intr
aspe
cifi
c D
ensi
ty
Com
pari
sons
18 v
s. 3
6 18
vs.
54
VE
LO
CIT
Y I
ND
EP
EN
DE
NC
E
A.ci
rcur
nnci
n~tu
s
grav
el v
s. p
ebbl
e -1
.43
-1.2
7
grav
el v
s. c
obbl
e -1
.94@
-0
.23
pebb
le v
s. c
obbl
e -2
.87*
* 0.
90
L.l
utzi
gr
avel
vs.
cob
ble
-0.1
0 -2
.83*
*
Inte
rspe
cifi
c C
ompa
riso
ns w
ith
Tot
al D
ensi
ty C
onst
ant
2 S
peci
es (
N=
36)
Aci
rc
Chu
ng
Llu
tzi
(N=
18)
(N=
18)
(N=
18)
----
-1.4
5 -0
.24
----
-0.9
3 -2
.84*
*
----
-1.5
3 -2
.40*
-0.0
8 1.
50
----
3 S
peci
es (
N =
54)
All
Thr
ee a
t N=
18
-2.9
6**
-1.3
0
0.66
-2.4
6*
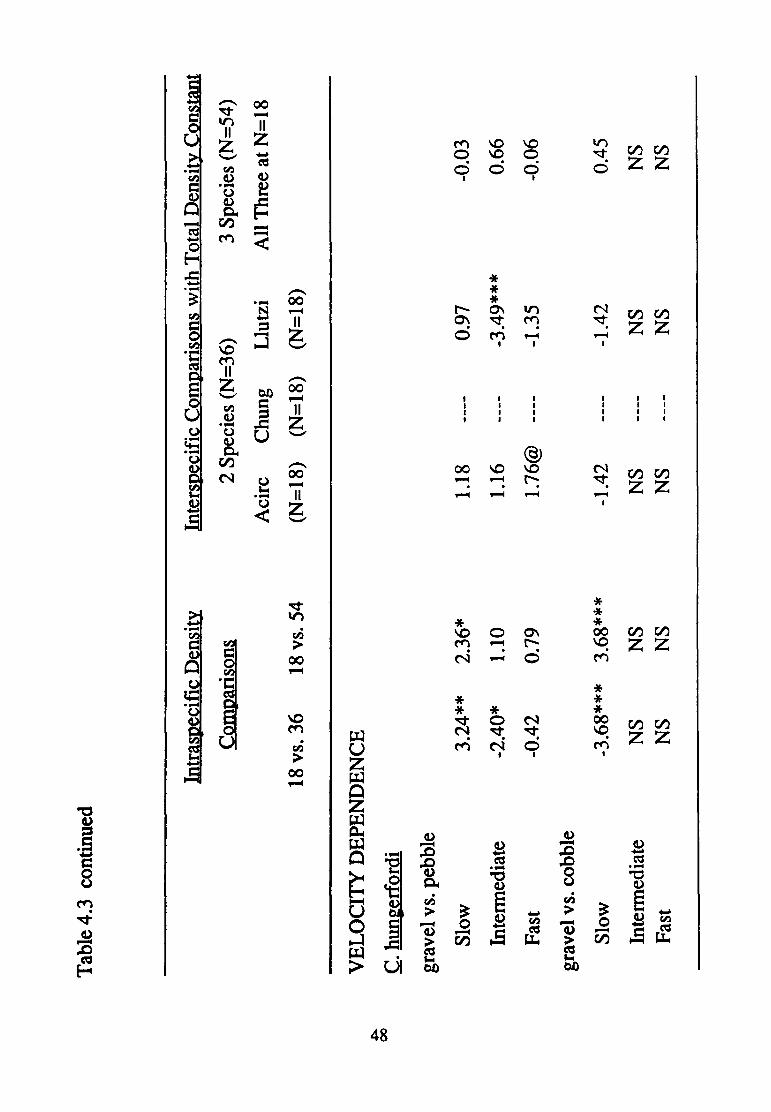
Tab
le 4
.3 c
onti
nued
Intr
as.p
ecif
ic D
ensi
ty
Com
pari
sons
18 v
s. 3
6 18
vs.
54
VE
LO
CIT
Y D
EP
EN
DE
NC
E
~
00
C.
hun
gerf
ordi
grav
el v
s. p
ebbl
e
Slo
w
3.24
**
2.36
*
Inte
rmed
iate
-2
.40*
1.
10
Fas
t -0
.42
0.79
grav
el v
s. c
obbl
e
Slo
w
-3.6
8***
3.
68**
*
Inte
rmed
iate
N
S N
S F
ast
NS
NS
Inte
rspe
cifi
c C
ompa
riso
ns w
ith T
otal
Den
sity
Con
stan
t
2 S
peci
es (
N=
36)
Aci
rc
Chu
ng
Llu
tzi
(N=
18)
(N=
18)
(N=
18)
1.18
---
-0.
97
1.16
---
--3
.49*
**
1.76
@
----
-1.3
5
-1.4
2 ---
--1
.42
NS
----
NS
NS
----
NS
3 S
peci
es (
N =
54)
All
Thr
ee a
t N
=18
-0.0
3
0.66
-0.0
6
0.45
NS
NS
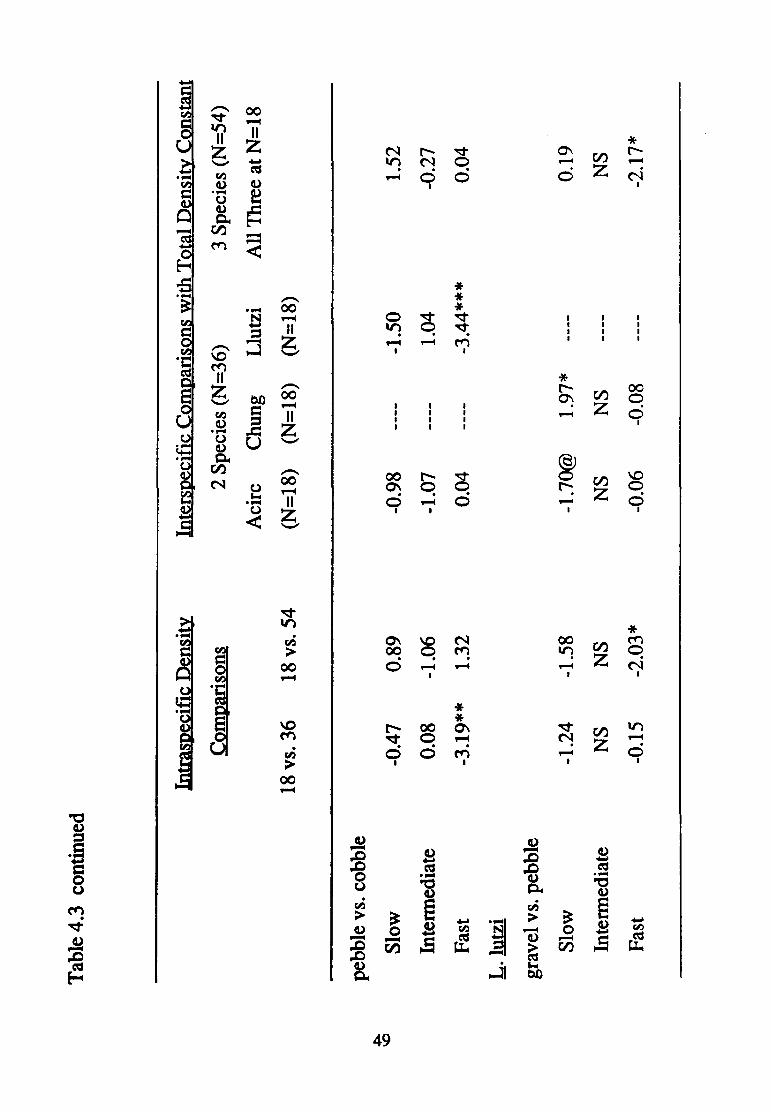
Tab
le 4
.3 c
onti
nued
Intr
aspe
cifi
c D
ensi
ty
Com
pari
sons
18 v
s. 3
6 18
vs.
54
pebb
le v
s. c
obbl
e ~
\0
Slo
w
-0.4
7 0.
89
Inte
rmed
iate
0.
08
-1.0
6
Fas
t -3
.19*
* 1.
32
L.m
m
grav
el v
s. p
ebbl
e
Slo
w
-1.2
4 -1
.58
Inte
rmed
iate
N
S N
S
Fas
t -0
.15
-2.0
3*
Inte
rspe
cifi
c C
ompa
riso
ns w
ith T
otal
Den
sity
Con
stan
t
2 S
peci
es (
N=
36)
Aci
rc
Chu
ng
Llu
tzi
(N=
18)
(N=
18)
(N=
18)
-0.9
8 ---
--1
.50
-1.0
7 ---
-1.
04
0.04
---
--3
.44*
**
-1.7
0@
1.97
* ---
-N
S N
S ---
-
-0.0
6 -0
.08
----
3 S
peci
es (
N=
54)
All
Thr
ee a
t N=
18
1.52
-0.2
7
0.04
0.19
NS
-2.1
7*
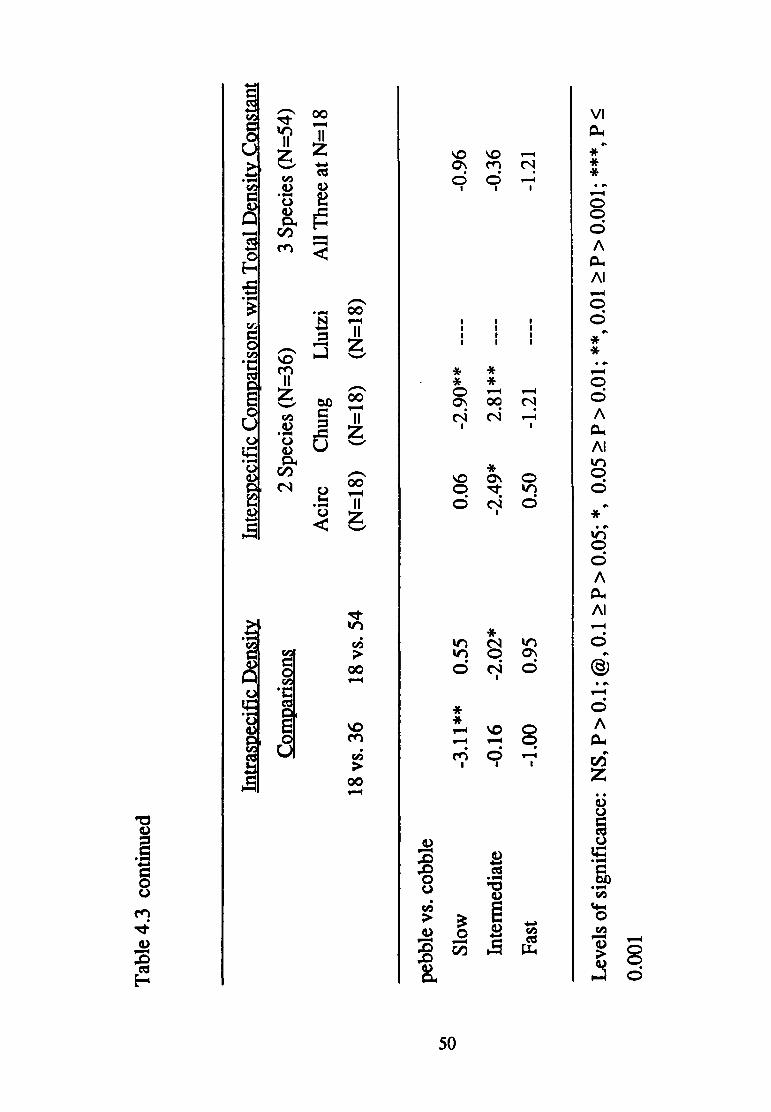
VI
0
Tab
le 4
.3 c
onti
nued
pebb
le v
s. c
obbl
e
Slo
w
Inte
rmed
iate
Fas
t
Intr
as.p
ecif
ic D
ensi
ty
Com
pari
sons
18 v
s. 3
6 18
vs.
54
-3.1
1 **
0.
55
-0.1
6
-1.0
0
-2.0
2*
0.95
Inte
rspe
cifi
c Comparison~ w
ith
Tot
al D
ensi
ty C
onst
ant
2 S
peci
es (
N=
36)
Aci
rc
Chu
ng
Llu
tzi
(N=
18)
(N=
18)
(N=
18)
0.06
-2
.90*
*
-2.4
9*
2.81
**
0.50
-1
.21
3 S
peci
es (
N -5
4)
All
Thr
ee a
t N
= 18
-0.9
6
-0.3
6
-1.2
1
Lev
elso
fsig
nifi
canc
e: N
S,P
>0
.1;@
,0.1
>P
>0
.05
; *,
0.0
5>
P>
0.0
1;
**,0
.01
>P
>0
.00
1;
**
*,P
<
0.00
1

whereas doubling (but not tripling) the density of .C. hungerfordi affected
substratum selection only in the fast current. Interspecific competition with
A. circumcinctus never significantly modified substratum selection by .C.
hungerfordi. However, the presence of L . .1Y1zi did modify substratum
selection in some substratum size combinations in intermediate and fast
current velocities. The simultaneous presence of all three naucorid species
did not alter substratum selection compared to situations with similarly
elevated densities of C. hungerfordi.
Limnocoris lutzi
Current velocity affected the way in which density treatments
modified substratum selection in all contrasts except those involving gravel
and cobble, where the effects of both density treatment and velocity were
consistent (Table 4.1). In cases where a three-way interaction existed,
substratum selection usually varied among the density treatments, except at
the intermediate velocity and the gravel vs. pebble contrast. Tripling the
number of L. lutzi consistently modified substratum selection in the gravel
vs. cobble contrast, but at only one of the three ve~ocities in each of the
other two substratum combinations. The addition of a single competing
species modified substratum selection, although not at fast current speeds or
in the gravel vs. cobble contrast. In the simultaneous presence of both other
species of naucorid, substratum selection by L. lutzi changed consistently
in the gravel vs. cobble contrast, but modified substratum selection in the
gravel vs. pebble contrast only at fast current velocities.
51

Discussion
Early work in stream ecology emphasized substratum characteristics
as factors largely responsible for the microdistribution of stream benthos.
Recently, researchers have stressed the importance of flow characteristics
(Statzner et al., 1988) and questioned the relevance of the substratum.
Clearly substratum and current attributes are confounded in natural streams
(Moon, 1939; Reice, 1980), yet have important independent implications
(Chapter lll). The premise for these experiments was based on competition
for space. Because naucorids respond to both current velocity and
substratum character (Chapter ill) in terms of habitat selection, variation in
both was included in these experiments.
Velocity effects
The role of current velocity in altering substratum selectivity for each
of the three naucorid species has been previously established (Chapter ill).
The way in which velocity affected substratum selectivity differed among
species. In the extreme case, substratum selection by A. circumcinctus did
not change in response current velocity. Similarly, substratum selection by
C. hun~erfordi, was fairly unresponsive to changes in current speeds. Those
responses that did occur were associated with small rock sizes which
constitute uncommon habitat for this species in the field. In comparison, the
manner in which L.lutzi selected substrata often depended on current
velocity. Given that velocity can alter habitat selection (at least for some
species), does this relationship remain constant over a realm of biotic
interactions? More specifically, does velocity affect habitat associations of a
species in the same manner, regardless of the type and magnitude of
competition that occurs within a community?
52

These results revealed a complex relationship, in which velocity often
affected the manner in which competition altered substratum selectivity. In
biological terms, a species response to an increase in its own density or the
presence of another competitor may vary, depending on current velocity.
Perhaps at fast currents, one species is able to cling to the substratum and
displace (exploitative or interference) other competitors, whereas, at slower
currents, adaptations for clinging or displacement may not suffice to preempt
a microhabitat. Instead, a second species (which is perhaps more mobile at
slow currents) may exhibit a higher degree of exploitative efficiency, and
may dominate the competitive interaction. The idea of environment
dependent competitive dominance is not new. Park (1954) showed how two
species of Tribolium could vary in their competitive dominance depending
on temperature. Similarly, Connell (1961) established a competitive
dominance between barnacle species which varied with altitude.
Of the three naucorid species tested, two (C. hungerfordi and L.lutzi)
exhibited complex responses to current and competition. For the former, a
three-way interaction between current velocity, density, and substratum
selection was consistently detected for every substratum contrast. For the
latter, a three-way response was detected in two of the three substratum
contrasts, reflecting the intimate relationship between this species and
current.
Density effects
The proponents of density-dependent habitat selection hold that at
high densities, a species will occupy a wider range of habitats (i.e., initially
suboptimal habitat will be selected as density increases). As the number of
individuals increases, the proportion of individuals in preferred to initially
53

unpreferred habitat will gradually decrease. As a consequence, each
naucorid species should respond to an increase in density by exhibiting
diminished substratum associations (see Chapter ill).
In nature, one would expect intraspecific competition to affect habitat
associations of a species more dramatically at high densities. These data
support the presence of intraspecific competition, but they do not advocate
the idea that highest densities affect habitat associations the most. For
example, ~. hun~erfordi was quite responsive to an increase density, but the
majority of these responses were detected when densities were doubled.
Similarly, A. circumcinctus changed its habitat associations in response to
doubled densities, but never responded to tripled densities. In comparison,
responses by L. lutzi to increases in density were less frequent than by C.
hun~erfordi, but tripled densities modified habitat associations more
frequently than at doubled densities. The frequency of responses by A.
circumcinctus and C. hun~erfordi to doubled densities over that of tripled
densities reflects some non-linearity of density effects on habitat selection.
The density of a species in a habitat that was initially suboptimal may reach
a threshold that can not sustain increasing densities without severe
consequences to an individual's fitness. At this point, the initially preferred
habitat offers higher fitness and individuals opt for this habitat secondarily.
This may explain those cases in which doubled densities altered habitat
associations, while tripling a species density failed to alter habitat
associations.
The degree and manner by which species altered their behavior in
response to density varied among species. Cryphocricos hun~erfordi was
quite responsive to an increase in density (in 42.9% of the i priori
intraspecific comparisons, competition was detected); whereas, A.
54

circumcinctus responded to an increase in density in a low 16.7% of the
time. Limnocoris ~was intermediate in response to increases in density;
this species altered its microdistribution in one-third of those l! priori
comparisons. Habitat selection is dynamic; a species perception of habitat
quality may depend on the density of conspecifics in that habitat. The
consequence of increasing population density on habitat selection may not
be a function of linearity as shown by these naucorids. Furthermore, the
relationships between density and habitat selection can vary among species.
Interspecific effects
Interspecific competition in nature appears to be mostly asymmetrical
(Lawton and Hassell, 1981; McAuliffe, 1984; Hemphill, 1988); however,
many studies focus on effects of competition on one target species, in tum,
neglecting reciprocal responses by other species (Schoener, 1983; Connell,
1983 ). Dominant hierarchies of competition and symmetry of responses are
impossible to establish in this manner. Approaches that measure the effects
of interspecific competition on all species concerned may offer insight into
mechanisms by which species coexist.
Responses by A. circumcinctus to the presence of other naucorids was
highly variable. Evidence from the two-species interaction experiments
indicated that A. circumcinctus was virtually unresponsive to~.
hun~erfordi; however, the presence of L . .him frequently altered habitat
associations. Similarly, the joint occurrence of all three species affected
habitat associations of A. circumcinctus. In all likelihood this response was
induced by L. lutzi. rather than the simultaneous presence of all three
species, considering that A. circumcinctus was unresponsive to ~.
hun~erfordi in previous experiments. Limnocoris ~ responded to
55

interspecific pressures in a third of the two-species a priori contrasts. The
majority of those responses were detected in the presence of .C. hungerfordi.
Czyphocricos hungerfordi was by far the least sensitive species to
interspecific competition. Habitat associations were never modified in the
presence of L. ~ or in the sirilultaneous presence of all three-species.
Ecologists frequently measure competitive dominance (in tenns of
competition for space) by the ability of a species to impose habitat
restrictions on other species while retaining its own habitat associations.
Given that the outcomes of competitive interactions are ultimately dependent
upon physical factors (Connell, 1983), such an approach may oversimplify
competitive relationships between species. The relationship between C.
hungerfordi and L. lutzi illustrates this point nicely. Czyphocricos.
hungerfordi was unresponsive to the presence L.lutzi. Conversely, L. lutzi
altered its patterns of habitat associations in the presence of ,C. hungerfordi
at larger rock sizes (i.e., pebble vs. cobble) but not in other contrasts. The
ability of~- hungerfordi to affect the distribution of L. lutzi at large rock
sizes, but not at small sizes illustrates how physical factors can detennine
competitive outcomes. If an attempt was made to determine competition for
smaller rocks between the two species, competition may not have been
detected, when in fact these species compete intensely for larger rock sizes.
Potential Mechanisms and Observations
Few studies isolate competitive mechanisms (interference vs.
exploitation vs. preemption) in stream systems, in part because of the
difficulty to quantify shared resources (Dudley et al., 1990). The difficulty
to detect these resources increases as the mobility of an organism increases.
For example, a sessile (or semi-sessile) benthic inhabitant is limited to a set
56

of resources defined by those it encounters in the three dimensional column
in which it inhabits. As mobility increases, the choices offered to an animal
is not limited to those encountered at the site of attachment. The ability to
forage enables a species to avoid deleterious effects of overcrowding thereby
optimizing a habitats ability to sustain diversity and high population
densities. Naucorids are among the most mobile lotic benthos in
southwestern stream systems. Furthennore the three species tested exhibit
extensive overlap in substratum, current, and prey utilization. This overlap
may explain the complex competitive relationships among the naucorids but
fails to offer mechanisms by which naucorids compete. Through
observation of naucorid behavior in closed containers and at slow velocities
in the artificial streams, this author offers two potential mechanisms by
which naucorids compete for space. The first entails a direct interference
mechanism. Intra- and interspecific encounters by A. circumcinctus and C.
hun~erfordi are not passive. Upon an encounter, two individuals grasp the
pro legs of one another. The individuals remain interlocked (often seconds at
a time) until one individual relinquishes its hold and continues its movement
to an unoccupied substratum. Another mechanism involves avoidance
behavior. When an individual L. ~ encounter conspecifics or other
naucorid species, the individual moves up into the water column and swims
in a random direction to a new site of settlement. If the new site of
settlement was occupied this behavior was frequently repeated until an
unoccupied microhabitat was identified. This avoidance behavior may serve
to segregate the species and explain why L.bmi altered its substratum
associations in the presence of other species.
Naucorids may minimize interspecific interactions by exploiting
different regions of substratum particles. For instance, ~. hun2erfordi was
57

rarely viewed clinging to the top or sides of individual rocks. Instead they
were found associated with under surfaces of larger rocks. In contrast, L.
lutzi frequently existed at the water-substratum interface where it minimized
contact with C. hungerfordi. This "particle partitioning" may minimize
conflicting interactions in tum facilitating the coexistence of the two species.
Competition among naucorids is complex. Naucorids can exhibit
variation in habitat associations depending upon current velocity, substratum
size distributions, density, and interspecific competition. The ability of
substrata to supply food may also influence habitat associations of naucorids
especially if the abundance and distribution of prey species are correlated
with flow and substratum characteristics. Furthermore, density reductions
caused by flooding events suppress intraspecific (Feminella and Resh, 1990)
and interspecific (Hemphill, 1991) competition in stream communities. The
SLR as well as other streams on the Edwards plateau is frequently subjected
to spates and high intensity floods which affect naucorid densities (York et
al., unpublished data). These events may reduce competition among
naucorid species and ensure species richness in the SLR.
58

Literature Cited
Connell, J. H. 1961. The influence of interspecific competition and other factors on the distribution of the barnacle Chthamalus stellatus. Ecology 42:710-723
Connell, J. H. 1975. Some mechanisms producing structure in natural communities: a model and evidence from field experiments. pp. 460-490. in M. L. Ecology and evolution of communities. Belknap Press, Cambridge, Massachusetts, U.S.A.
Connell, J. H. 1978. Diversity in tropical rainforests and coral reefs. Science 199:1302-1310.
Connell, J. H. 1980. Diversity and the coevolution of competitors, or the ghost of competition past. Oikos 35:131-138.
Connell, J. H. 1983. On the prevalence and relative importance of interspecific competition: evidence from field experiments. Am. Nat. 122:661-669.
Constantz, G. D. 1974. The mating behavior of a creeping water bug, Ambzysus occidentalis (Hemiptera: Naucoridae ). Am. Midi. Nat. 92:234-239.
Dudley, T. L., S.D. Cooper, and N. Hemphill. 1986. Effects of macroalgae on a stream invertebrate community. J. N. Am. Benthol. Soc. 5:93-106.
Dudley, T. L., C. M. D'Antonio, & S.D. Cooper. 1990. Mechanisms and consequences of interspecific competition between two stream insects. I. Anim. Ecol. 59:849-866.
Feminella, I. W., & V. H. Resh. 1990. Hydrologic influences, disturbance and intraspecific competition in a stream caddisfly population. Ecology 71:2083-2094.
Fretwell, S.D., and H. L. Lucas, Jr. 1970. On territorial behavior and other factors influencing habitat distribution in birds. I. Theoretical development. Acta Biotheoretica 19:16-36.
59

•
Glass, L.W. and R.V. Bovbjerg. 1969. Density and dispersion in laboratory populations of caddisfly larvae (Cheumatopsyche, Hydropsychidae). Ecology 50:1082-1084.
Gonsoulin, G. J. 1973. Seven families of aquatic and semiaquatic Hemiptera in Louisiana. Entomol. News 84:83-88.
Harding, J. and M. H. Colbo. 1981. Competition for attachment sites between larvae of Simuliidae (Diptera). Can. Ent. 113:761-763.
Hemphill, N. 1988. Competition between two stream dwelling filterfeeders, Hydropsyche oslari and Simulium virgatum. Oecologia 77:73-80.
Hemphill, N. 1991. Disturbance and variation in competition between two stream insects. Ecology 72:864-872.
Herrmann, D.P., R. W. Sites and M. R. Willig. 1992. A laboratory flow tank with variable currents and depths for replicating riffles and shallow streams. Texas J. Sci. 44:89-94.
Lawton, J. H., and M.P. Hassell. 1981. Asymmetrical competition in insects. Nature 289:793-795.
McAuliffe, J. R. 1983. Competition, colonization patterns and disturbance in stream benthic communities. pp. 137-156. in J. R. Barnes and G. W. Minshall ( eds. ). Stream ecology: application and testing of general ecological theory. Plenum, New York, New York.
McAuliffe, J. R. 1984. Competition for space, disturbance, and the structure of a benthic stream community. Ecology 65:894-908.
McLachlan, A. J. 1969. Substrate preferences and invasion behaviour exhibited by larvae of Nilodorum breyibucca Freeman (Chironomidae) under experimental conditions. Hydrobiologia 33:237-249.
60

McPherson J. E., R. J. Packauskas, and P. P. Korch, ill. 1987. Life history and laboratory rearing of Pelocoris femoratus (Hemiptera: Naucoridae), with descriptions of immature stages. Proc. Entomol. Soc. Wash. 89:288-295.
Minshall, G. W. 1984. Aquatic insect-substratum relationships, pp. 358-400. in V. H. Resh and D. M. Rosenberg (eds.). The ecology of aquatic insects. Praeger, New York.
Moon, H.P. 1939. Aspects of the ecology of aquatic insects. Trans. Brit. Entomol. Soc. 6:39-49.
Park, T. 1954. Experimental studies of interspecific competition. II. Temperature, humidity and competition in two species of Tribolium. Physiol. Zool. 27:177-238.
Pulliam, H. R. 1983. Ecological community theory and the coexistence of sparrows. Ecology 64:45-52
Reice, S. R. 1980. The role of substratum in benthic macroinvertebrate microdistribution and litter decomposition in a woodland stream. Ecology 61 :580-590.
Schoener, T. W. 1983. Field experiments on interspecific competition. Amer. Nat. 122:240-285.
Sites, R. W. and B. J. Nichols. 1990. Life history and descriptions of immature stages of Ambrysus lunatus lunatus (Hemiptera: Naucoridae). Ann. Entomol. Soc. Am. 83:800-808.
Sites, R. W. and M. R. Willig. 1991. Microhabitat associations of three sympatric species of Naucoridae (Insecta: Hemiptera). Environ. Entomol. 20:127-134.
Sousa, W. P. 1984. The role of disturbance in natural communities. Ann. Rev. Ecol. Sys. 15:353-391.
SPSS Inc. Advanced statistics guide. 2nd Edition. pp. 527. Chicago n.
61

Stout, R. J. 1981. How abiotic factors affect the distribution of two species of tropical predaceous aquatic bugs (Family: Naucoridae ). Ecology 62:1170-1178
Venkatesan, P. and A. Cloarec. 1988. Density dependent prey selection in llyocoris (Naucoridae). Aquatic Insects 10:105-116.
Ward, J. V. 1992. Aquatic insect ecology. 1. Biology and habitat. Wiley and Sons, Inc New York.
Wentworth, C. K. 1922. A scale of grade and class terms for cladistic sediments. J. Geol. 30:377-392.
Wiley, M. J. 1981. Interacting influences of density and preference of the emigration rates of some lotic chironomid latvae (Diptera: Chironomidae). Ecology 62:426-438.
Zahar, A. R. 1951. The ecology and distribution of black-flies (Simuliidae) in southeast Scotland. J. Anim. Ecol. 20:33-62.
62

CHAPTER V
EFFECTS OF SUBSTRATUM SIZE AND HETEROGENEITY
ON MACROBENTHIC DNERSITY AND
COMPOSffiON
Abstract
Studies concerning diversity and taxonomic composition associated
with substratum characteristics in temperate streams have received little
attention despite the ascendancy of biodiversity as a central theme in
contemporary ecology. This study evaluated the effects of substratum
characteristics on species diversity and taxonomic composition. In
particular, colonization of four rock size classes (gravel, pebble, cobble and
a heterogeneous combination of all three) by macrobenthos was quantified in
the South and North Llano Rivers of central Texas during the summer of
1991. In each of six experimental treatments, the effect of substratum type
on species diversity and composition was evaluated by placing 25 pairs of
trays into riffle habitat within each river. Each treatment was defmed by one
paired contrast between two of the four rock classes. Trays were left
undisturbed for three days after which all macroinvertebrates were extracted
and preserved. The Shannon index of diversity was used to estimate the
diversity of all macroinvertebrates within each tray, and a paired-t test was
used to evaluate if statistically significant differences in species diversity
existed between substrata. Similarly, compositional differences between
substrata within each river were evaluated by log-linear models (SPSS Inc,
1988). Highly significant differences in taxonomic composition between
substrata occurred for all rock contrasts in both rivers. However, substratum
size and heterogeneity had little or no effect on species diversity.
63

Introduction
Biodiversity is a central theme in contemporary ecology. As a
consequence, ecologists have developed a variety of methods to measure
diversity. These measures have potential application to conservation, land
management, and environmental monitoring (Magurran, 1988). In fact,
considerable debate surrounds the appropriateness of diversity measures. In
conseiVation, measurements are based largely on species richness, with little
regard to relative abundance of constituent taxa. In environmental
monitoring, measures are used that include both relative abundances and
species richness. Nevertheless, studies that examine multiple aspects of
community complexity and structure are few. The isolation of factors that
contribute to species diversity, and taxonomic composition is desirable
because these factors reflect the mechanisms contributing to community
level processes.
A thorough approach for the description of diversity data involves
species abundances models [e.g., log-nonnal model (Sugihara, 1980),
logarithmic series model (Magurran, 1988), and random niche boundary
hypothesis (MacArthur, 1957)]; however, these approaches can involve
tedious model fitting, and problems arise when two communities that are
being compared do not fit the same model. A simpler and more popular
approach involves indices that are based on proportional abundances. These
measures are desirable because they not only account for eveness (a measure
of the distribution of abundances) but also account for species richness.
Nevertheless, examination of only this type of measure may not be
conducive to a fundamental understanding of diversity because of their
generality. The Shannon (H') infonnation index (Krebs, 1985) has become
64

one of the most widely used indices. If the Shannon index is applied to a
number of samples, the measures themselves are nonnally distributed
(Magurran, 1988). This allows for the use of parametric statistics. Although
information indices such as H' are popular they remain taxonomically
insensitive. For example, the Shannon index will render the same value
given that the number of species and their proportional abundances remain
constant (Magurran, 1988) even though constituent taxa are completely
different. Approaches that are sensitive to taxonomic composition may offer
additional insight into species diversity.
Many stream species exploit benthic modes of existence (Ward,
1992). As a consequence, it is not surprising that substratum characteristics
are major determinants of the distribution and abundance of stream
organisms (Minshall, 1984 ). Particle size and heterogeneity are important
components of the substratum (Cummins and Lauff, 1969, Minshall and
Minshall, 1977, Lamberti and Resh, 1979, Fuller and Rand, 1990, and many
others). Nonetheless, the effects of particle size and heterogeneity on stream
benthic species composition are often confounded by factors such as current
association, food habits, respiratory requirements, and other substratum
characteristics (Ward, 1992). As such, careful observations and analyses are
required to interpret substratum effects on community organization.
The idea that complex habitats offer resources that are unavailable to
species in simple habitats is almost universally accepted (Minshall, 1984 ).
In stream environments, a positive correlation may exist between particle
heterogeneity and species richness (Allan, 1975; de March, 1976; Williams,
1980). Nevertheless, a direct correlation between substratum heterogeneity
and density is speculative (Williams, 1980; Ennan and Ennan, 1984).
65

Mean particle size also affects the abundance and distribution of
stream benthos, but size and heterogeneity are often correlated. As the
median particle size of sediments increases, sediment complexity can
increase as well (Reice, 1974). For example, cobble substrata have
propensities for tiny gravel to become lodged in interstitial spaces. The
resulting mixture of particles minimizes gravel-current interactions, thereby
reducing the ability of current to displace gravel. Consequently, cobble is
often considered a conglomeration of particle sizes (Ward, 1992). Although
particle size and heterogeneity are positively correlated, many investigations
have failed to control for their confounding effects on species diversity and
composition. As a result, attempts (e.g., Erman and Erman, 1984) have
focused on elucidating the separate effects of particle size and complexity on
macro benthic assemblages. The effects of heterogeneity on species diversity
may indeed be confounded with particle size (Erman and Ennan, 1984 ). The
following study was aimed at determining the effects of substratum size and
heterogeneity on species diversity and taxonomic composition in two similar
river ecosystems.
Materials and Methods
Study Sites
The South (SLR) and North (NLR) Llano Rivers are located on the
Edwards Plateau of central Texas. The NLR originates in Sutton Co. and
flows eastward ca. 80 km to its confluence with the SLR in Kimble Co. The
SLR originates in Edwards Co., and meanders ca. 17 5 km NE at which point
it joins the NLR. At the confluence, the Llano River emerges and drains into
the Colorado River basin. The SLR and NLR lie on Cretaceous limestone
and are typical of streams on the Edwards Plateau in having gravel and
66

cobble substrata with well-defined riffle and pool habitats. The watersheds
of each river receive substantial quantities of rainfall in the spring and early
fall as a result of moisture-laden air derived from the tropical Gulf storm
tract and convective thunderstorm activity (Baker, 1977). Although streams
in close proximity often exhibit similar flow regimes, the SLR and NLR
vary in both point of origin and flow direction. Consequently, flow regimes
and floods vary with respect to frequency and magnitude in accordance with
the location and prevalence of thunderstorm activity.
Strikingly different habitat structure characterizes channel vs.
marginal habitats in each river, and contributes to contrasting biota in bank
and mid-channel areas. Emergent angiosperms, algae, and silt are common
components in marginal areas. In contrast, exposed cobble and gravel
substrata dominate riffle habitat. Riffles are often vegetatively
impoverished; however, a plethora of invertebrates inhabit these areas.
Colonization
Substratum trays (30.5 x 30.5 x 2.5 em) were constructed from
aluminum flashing, and were used in field experiments as colonization
platforms. Pairs of trays filled with substratum were placed into both the
SLR and NLR during the Summer of 1991. Six treatments were defmed by
all paired contrasts between four substratum classes: (1) coarse gravel (CG;
8-16 mm rocks), (2) small pebble (SP; 16-32 mm rocks), (3) small cobble
(SC; 64-128 mm rocks) and ( 4) a heterogeneous mix (HM) [the first three
size categories from the Wentworth (1922) classification scheme after
Minshall (1984)]. Volumetric displacement standardized the amount of
substrata used in each category (CG, 900 ml; SP, 1200 ml; SC, 1500 m1 and
HM, 1/3 of each homogenous category). Trays were left undisturbed for
67

three days, after which all macroinvertebrates were extracted and preserved
in 70% ethanol. The Shannon index of diversity (Krebs, 1985) estimated the
diversity of all macroinvertebrates within each tray, and a paired-t test
(Sokal and Rohlf, 1981) evaluated differences in species diversity between
substratum types (25 paired samples). Similarly, differences in taxonomic
composition between paired substrata were evaluated for each rock contrast
with log-linear models (SPSS Inc., 1988).
Results and Discussion
Site comparison
Mean temperature, depth, and current velocity were similar at sites in
the SLR and NLR (Table 5.1); however, statistically significant differences
in habitat characteristics were detected by both multivariate (P<0.001) and
univariate tests (temperature, P<0.001; depth, P=0.007; current velocity,
P=0.005). These differences may contribute to different faunal compositions
between rivers.
Riffle faunas
The riffle faunas within the SLR and NLR sites are characteristically
litho- and psephophilic (Table 5.2). Nonetheless, riffle faunas differ
dramatically with respect to the abundances of several species. For instance,
a caddisfly, Marilia sp., was virtually absent (only one specimen collected)
from the SLR; however, it constituted 4.8% of all individuals collected in the
NLR. Conversely, a mayfly Traverella presidiana, was scarce in the NLR
(only two specimens collected); however, it comprised 1.7% of all
individuals collected within the SLR.
68
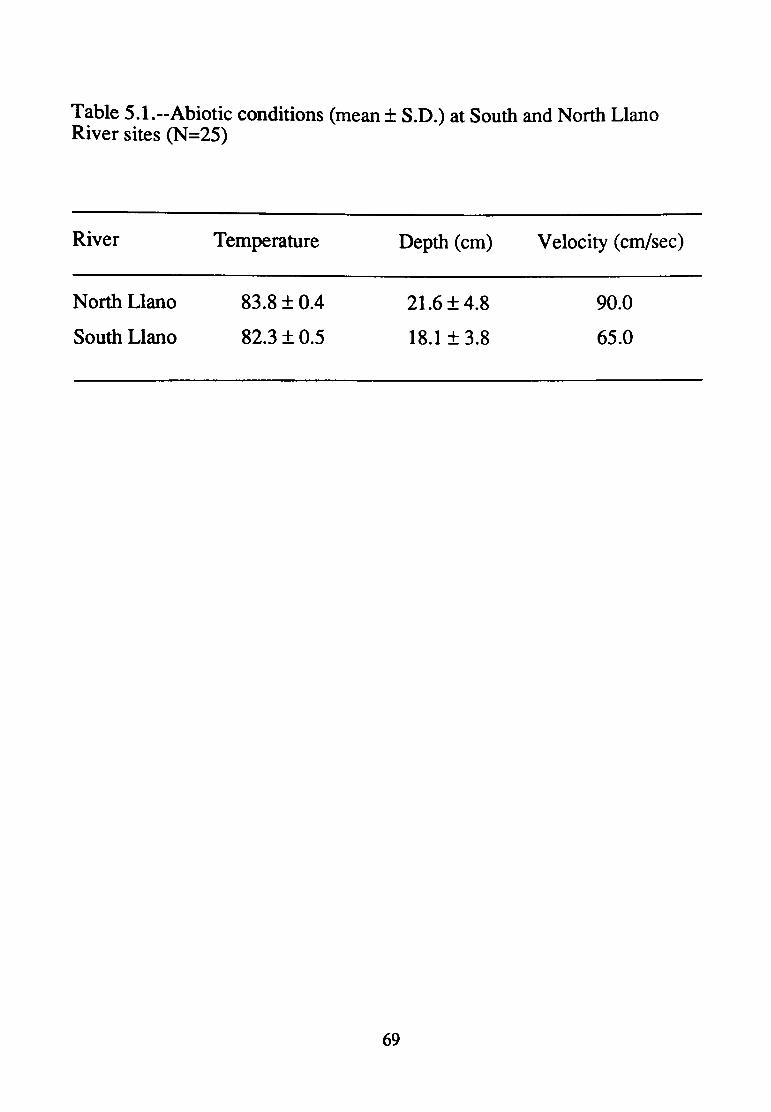
Table 5.1.--Abiotic conditions (mean+ S.D.) at South and North Llano River sites (N=25)
River Temperature
North Llano 83.8 + 0.4
South Llano 82.3 + 0.5
Depth (em)
69
21.6 + 4.8
18.1 + 3.8
Velocity (em/sec)
90.0
65.0
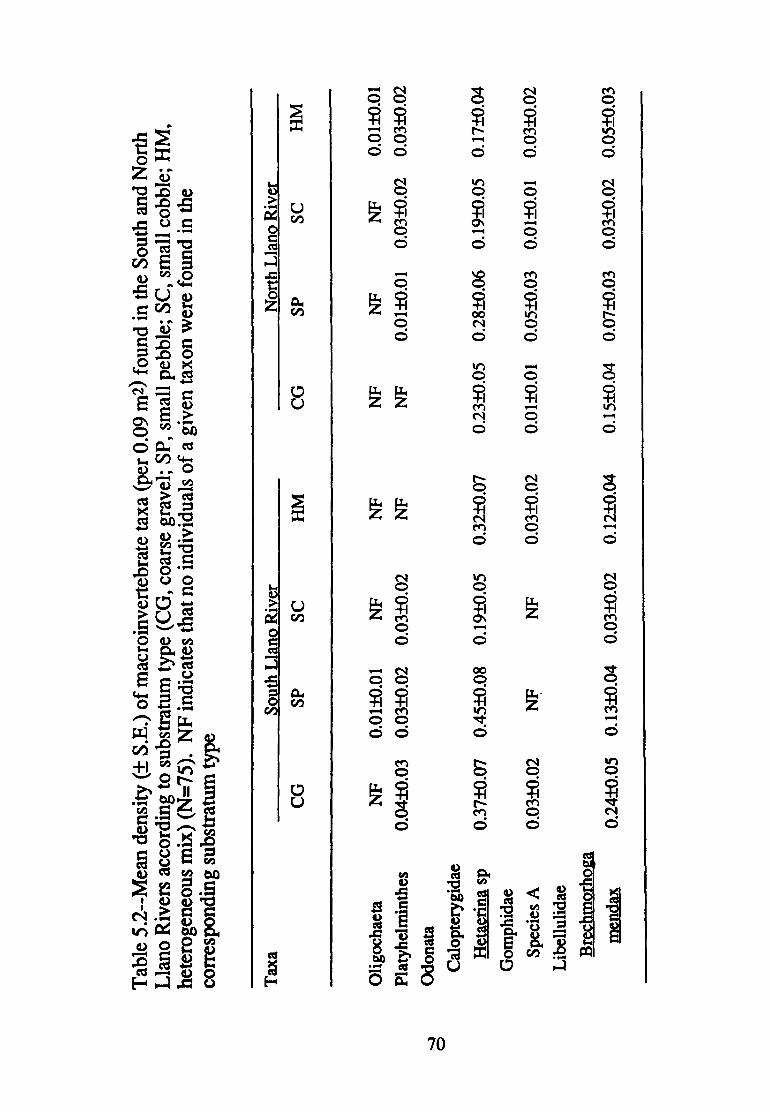
...J
0
Tab
le 5
.2--
Mea
n de
nsit
y(+
S.E
.) o
fmac
roin
vert
ebra
te ta
xa (
per 0
.09
m2)
fou
nd i
n th
e S
outh
and
Nor
th
Lla
no R
iver
s ac
cord
ing
to s
ubst
ratu
m ty
pe (
CG
, coa
rse
grav
el;
SP
, sm
all p
ebbl
e; S
C, s
mal
l cob
ble;
HM
, he
tero
gene
ous
mix
) (N
=75
). N
F in
dica
tes
that
no
indi
vidu
als
of a
giv
en ta
xon
wer
e fo
und
in th
e co
rres
pond
ing
subs
trat
um ty
pe
Tax
a S
outh
Lla
no R
iver
N
orth
_Lla
no R
ive_
r__
CG
S
P
SC
H
M
CG
S
P
SC
H
M
Oli
goch
aeta
N
F
0.01
±0.
01
NF
N
F
NF
N
F
NF
0.
01±
0.01
Pla
tyhe
lmin
thes
0.
04±
0.03
0.
03±
0.02
0.
03±
0.02
N
F
NF
0.
01±
0.01
0.
03±
0.02
0.
03±
0.02
Odo
nata
Cal
opte
rygi
dae
Het
aeri
na s
p
0.37
±0.
07
0.45
±0.
08
0.19
±0.
05
0.32
±0.
07
0.23
±0.
05
0.28
±0.
06
0.19
±0.
05
0.17
±0.
04
Gom
phid
ae
Spe
cies
A
0.03
±0.
02
NF
N
F
0.03
±0.
02
0.01
±0.
01
0.05
±0.
03
0.01
±0.
01
0.03
±0.
02
Lib
ellu
lida
e
Br~hmQrhQ&il
men
dax
0.24
±0.
05
0.13
±0.
04
0.03
±0.
02
0.12
±0.
04
0.15
±0.
04
0.07
±0.
03
0.03
±0.
02
0.05
±0.
03

Tab
le 5
.2 c
onti
nued
Tax
a So
uth
Lla
no R
iver
N
orth
Lla
no R
iver
CG
SP
SC
HM
CG
SP
SC
H
M
Coe
nagr
ioni
dae
Spec
ies
A
0.13
±0.
05
0.29
±0.
06
0.19
±0.
05
0.12
±0.
04
0.19
±0.
05
0.13
±0.
04
0.09
±0.
03
0.15
±0.
05
Spec
ies
B
0.04
±0.
02
NF
0.
01±
0.01
0.
09±
0.03
0.
08±
0.04
0.
07±
0.03
N
F
0.11
±0.
04
Eph
emer
opte
ra
Bae
tidae
-..
J B
aetis
sp
0.07
±0.
03
0.05
±0.
03
0.16
±0.
05
0.08
±0.
04
0.03
±0.
02
NF
0.
03±
0.02
0.
03±
0.02
-
Da~~
lob~
&eti
s
mex
ican
us
0.13
±0.
06
0.08
±0.
03
0.28
±0.
08
0.16
±0.
05
0.23
±0.
06
0.12
±0.
05
0.73
±0.
27
0.16
±0.
05
Fa.Il
eceo
n
quill
eri
0.23
±0.
07
0.21
±0.
05
0.68
±0.
17
0.53
±0.
10
0.61
±0.
13
1.07
±0.
22
1.27
±0.
21
1.00
±0.
15
Lep
toph
lebi
idae
Ihra
ulod
es
2onz
ales
i 1.
48±
0.19
1.
89±
0.22
0.
61±
0.10
1.
43±
0.20
2.
99±
0.33
2.
64±
0.28
0.
93±
0.14
2.
23±
0.24

Tab
le 5
.2 c
onti
nued
Tax
a S
outh
Lla
no R
iver
N
orth
Lla
no R
iver
CG
S
P
SC
H
M
co
SP
S
C
HM
Tra
yere
lla
pres
idia
na
0.49
±0.
12
0.40
±0.
08
0.31
±0.
08
0.71
±0.
11
0.01
±0.
01
NF
N
F
0.01
±0.
01
Spe
cies
C
0.01
±0.
01
0.03
±0.
02
NF
0.
04±
0.02
0.
32±
0.10
0.
37±
0.08
0.
11±
0.04
0.
44±
0.09
Oli
gone
urid
ae
Isonychia~
7.48
±0.
86
15.0
4±1.
18
7.41
±0.
88
13.4
0±1.
27
4.40
i0.6
0 8.
23±
0.88
1.
64±
0.28
5.
53±
0.74
-..
.J T
rico
ryth
idae
N
wtohmh~s
spA
0.
69±
0.17
0.
33±
0.07
0.
43±
0.10
0.
63±
0.10
0.
19±
0.05
0.
08±
0.03
0.
08±
0.04
0.
21±
0.06
Le~1
2hmh
~s
spB
0.
68±
0.17
0.
39±
0.10
0.
85±
0.20
0.
75±
0.16
0.
27±
0.08
0.
45±
0.11
0.
48±
0.12
0.
32±
0.09
Tri~
QQth
od~~
sp
0.09
±0.
04
0.16
±0.
11
0.11
±0.
04
0.19
±0.
05
0.61
±0.
17
0.36
±0.
11
0.29
±0.
06
0.28
±0.
13
Tri
chop
tera
Hel
icop
sych
idae
Hel
icoo
svch
e sp
0.
40±
0.12
0.
24±
0.06
0.
15±
0.05
0.
29±
0.12
0.
81±
0.19
0.
41±
0.10
0.
28±
0.09
0.
56±
0.12
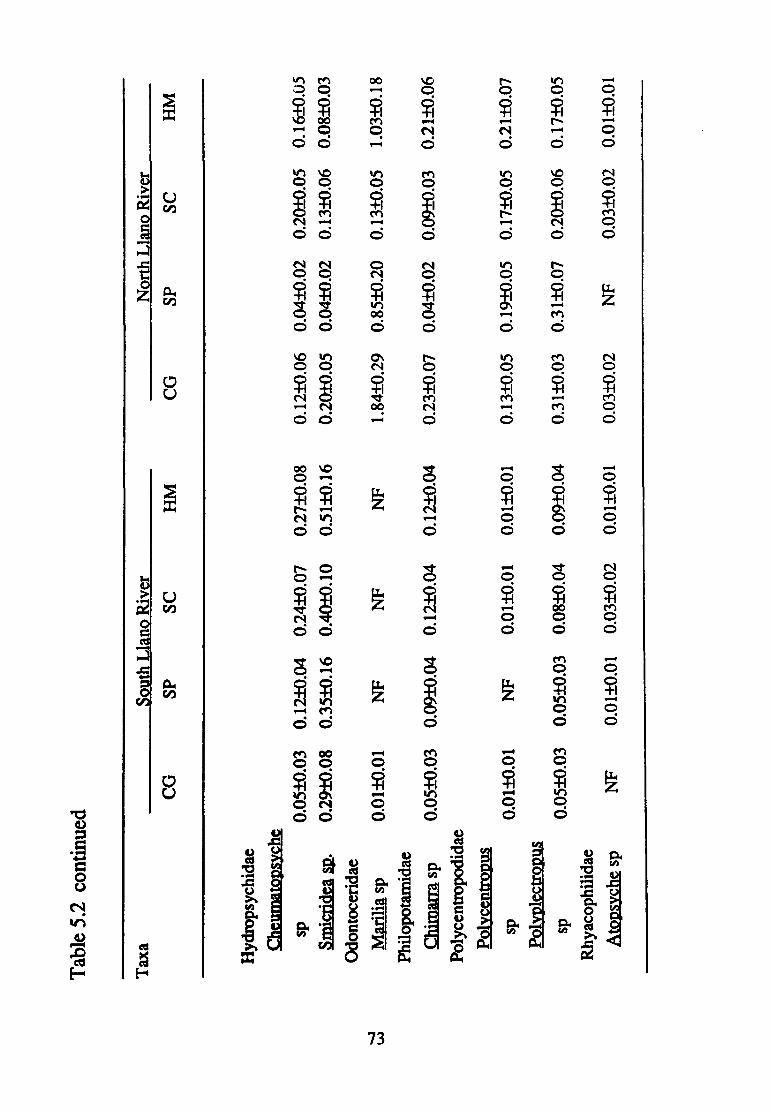
Tab
le 5
.2 c
onti
nued
Tax
a S
outh
Lla
no R
iver
N
orth
Lla
no R
iver
co
SP
S
C
HM
co
S
P
SC
H
M
Hyd
rops
ychi
dae
Cheumatop~~b~
sp
0.05
±0.
03
0.12
±0.
04
0.24
±0.
07
0.27
±0.
08
0.12
±0.
06
0.04
±0.
02
0.20
±0.
05
0.16
±0.
05
Sm
icri
dea
21·
0.29
±0.
08
0.35
±0.
16
0.40
±0.
10
0.51
±0.
16
0.20
±0.
05
0.04
±0.
02
0.13
±0.
06
0.08
±0.
03
Odo
ntoc
erid
ae
....,J
M
aril
ia s
p 0.
01±
0.01
N
F
NF
N
F
1.84
±0.
29
0.85
±0.
20
0.13
±0.
05
1.03
±0.
18
Vol
Phi
lopo
tam
idae
Cbi
mar
ra s
p 0.
05±
0.03
0.
09±
0.04
0.
12±
0.04
0.
12±
0.04
0.
23±
0.07
0.
04±
0.02
0.
09±
0.03
0.
21±
0.06
Pol
ycen
trop
odid
ae
PQI!~~ntropys
sp
0.01
±0.
01
NF
0.
01±
0.01
0.
01±
0.01
0.
13±
0.05
0.
19±
0.05
0.
17±
0.05
0.
21±
0.07
Polm
l~~t
tQPY
S
sp
0.05
±0.
03
0.05
±0.
03
0.08
±0.
04
0.09
±0.
04
0.31
±0.
03
0.31
±0.
07
0.20
±0.
06
0.17
±0.
05
Rhy
acop
hili
dae
Ato
osvc
he s
p N
F
0.01
±0.
01
0.03
±0.
02
0.01
±0.
01
0.03
±0.
02
NF
0.
03±
0.02
0.
01±
0.01
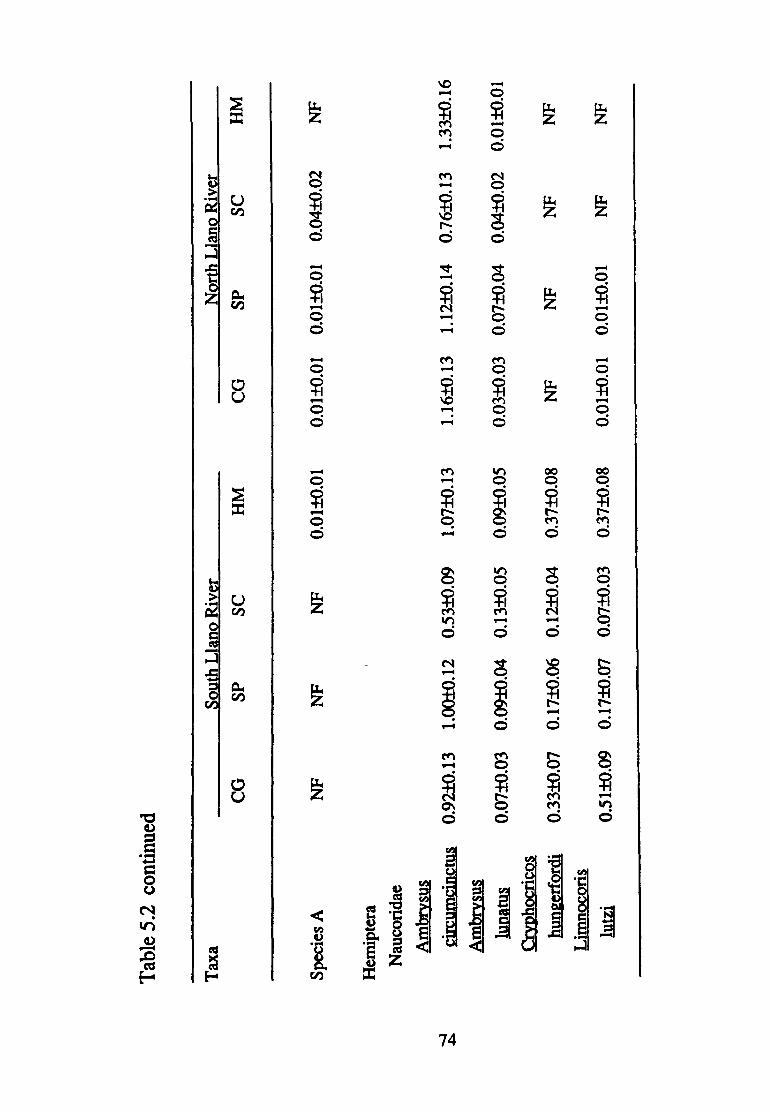
Tab
le 5
.2 c
onti
nued
Tax
a So
11th
Lla
no R
iver
N
orth
Lla
no_R
iver
CG
SP
SC
HM
CG
SP
SC
H
M
Spec
ies
A
NF
N
F
NF
0.
01±
0.01
0.
01±
o.01
0.
01±
o.01
0.
04±
0.02
N
F
Hem
ipte
ra
Nau
cori
dae
Am
bt:y
sus
-..J
ci
rc:u
mcin
~tus
0.
92±
0.13
1.
00±
0.12
0.
53±
0.09
1.
07±
0.13
1.
16±
o.13
1.
12±
0.14
0.
76±
o.13
1.
33±
0.16
~
Am
bt:y
sus
luna
tus
0.07
±0.
03
0.09
±0.
04
0.13
±0.
05
0.09
±0.
05
0.03
±0.
03
0.07
±0.
04
0.04
±0.
02
0.01
±0.
01
Ccyp
hocr
i~QS
bYn&
erfQ
rdi
0.33
±0.
07
0.17
±0.
06
0.12
±0.
04
0.37
±0.
08
NF
N
F
NF
N
F
Lim
noco
ris
hlW
0.
51±
0.09
0.
17±
0.07
0.
07±
0.03
0.
37±
0.08
0.
01±
o.01
0.
01±
0.01
N
F
NF
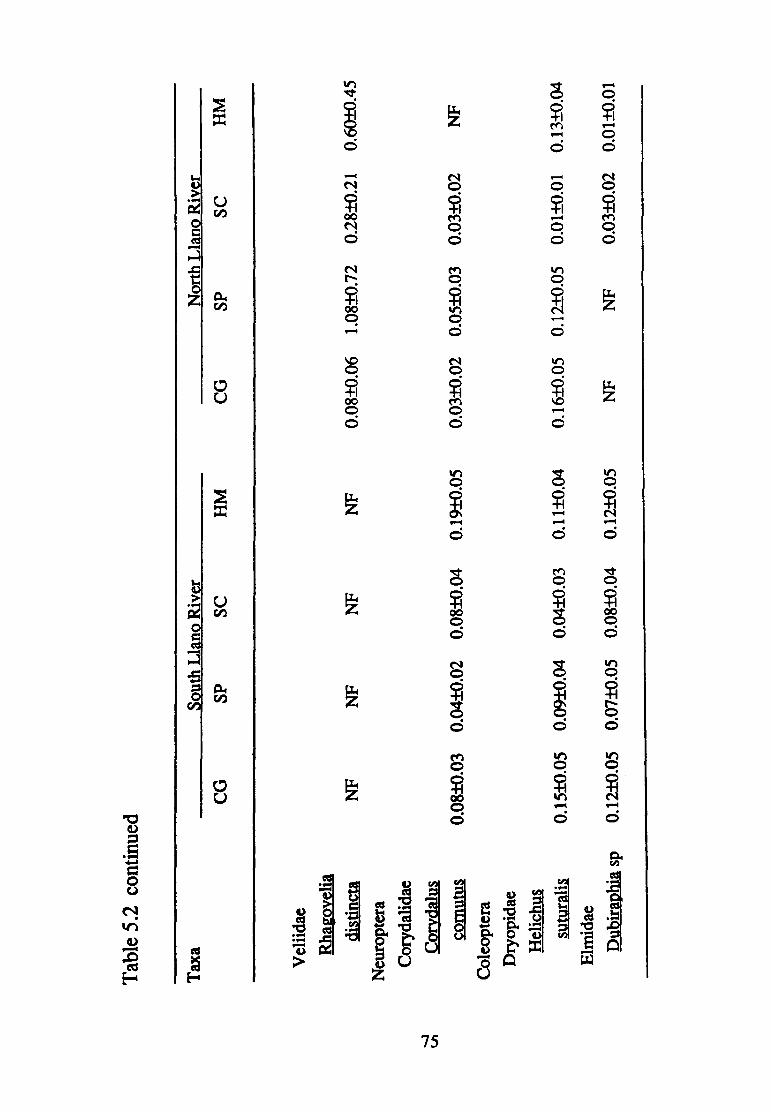
Tab
le 5
.2 c
ontin
ued
Tax
a
Vel
iidae
Rha
gove
lia
dist
inct
a
Neu
ropt
era
Cor
ydal
idae
CG
NF
Sout
h L
lano
Riv
er
SP
SC
HM
NF
N
F
NF
......,J
v.
C
ot:y
dalu
s
corn
utus
Col
eopt
era
Dry
opid
ae
Hel
ie bu
s
sutu
ral i
s
Elm
idae
0.08
±0.0
3 0.
04±0
.02
0.08
±0.0
4 0.
19±0
.05
0.15
±0.0
5 0.
09±0
.04
0.04
±0.0
3 0.
11±0
.04
Dub
irap
hia
sp
0.12
:t.i).
05
0.07
±0.0
5 0.
08±0
.04
0.12
±0.0
5
Nor
th L
lanQ
RiY
er
CG
SP
SC
HM
0.08
±0.0
6 1.
08±0
.72
0.28
±0.2
1 0.
60±0
.45
0.03
±0.0
2 0.
05±0
.03
0.03
±0.0
2 N
F
0.16
±0.0
5 0.
12±0
.05
0.01
±0.0
1 0.
13±0
.04
NF
N
F
0.03
±0.0
2 0.
01±0
.01
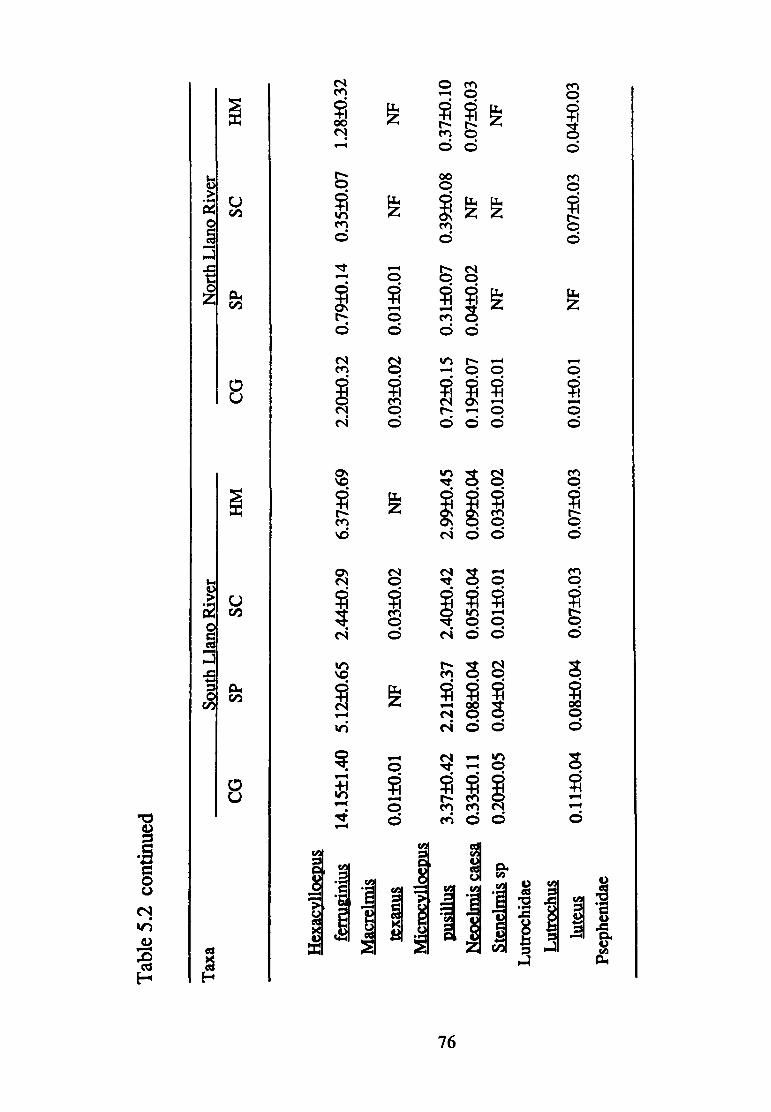
Tab
le 5
.2 c
onti
nued
Tax
a S
outh
Lla
no R
iver
N
orth
Lla
no R
iver
CG
SP
S
C
HM
CG
S
P
SC
H
M
H~x~
~!ll
oe.a
:us
ferr
u&in
ius
14.1
5±1.
40
5.12
±0.
65
2.44
±0.
29
6.37
±0.
69
2.20
±0.
32
0.79
±0.
14
0.35
±0.
07
1.28
±0.
32
Mac
relm
is
texa
nys
0.01
±0.
01
NF
0.
03±
0.02
N
F
0.03
±0.
02
0.01
±0.
01
NF
N
F
Mi~mc!lloe,ay:i
-...J
pusi
llus
3.
37±
0.42
2.
21±
0.37
2.
40±
().4
2 2.
99±
0.45
0.
72±
0.15
0.
31±
0.07
0.
39±
0.08
0.
37±
0.10
0
'\
Neo
elm
is ~
0.33
±0.
11
0.08
±0.
04
0.05
±0.
04
0.09
±0.
04
0.19
±0.
07
0.04
±0.
02
NF
0.
07±
0.03
Ste
nelm
is s
p 0.
20±
0.05
0.
04±
0.02
0.
01±
0.01
0.
03±
0.02
0.
01±
0.01
N
F
NF
N
F
Lut
roch
idae
Lut
roch
:us
lyte
ys
0.11
±0.
04
0.08
±0.
04
0.07
±0.
03
0.07
±0.
03
0.01
±0.
01
NF
0.
07±
0.03
0.
04±
0.03
Pse
phen
idae
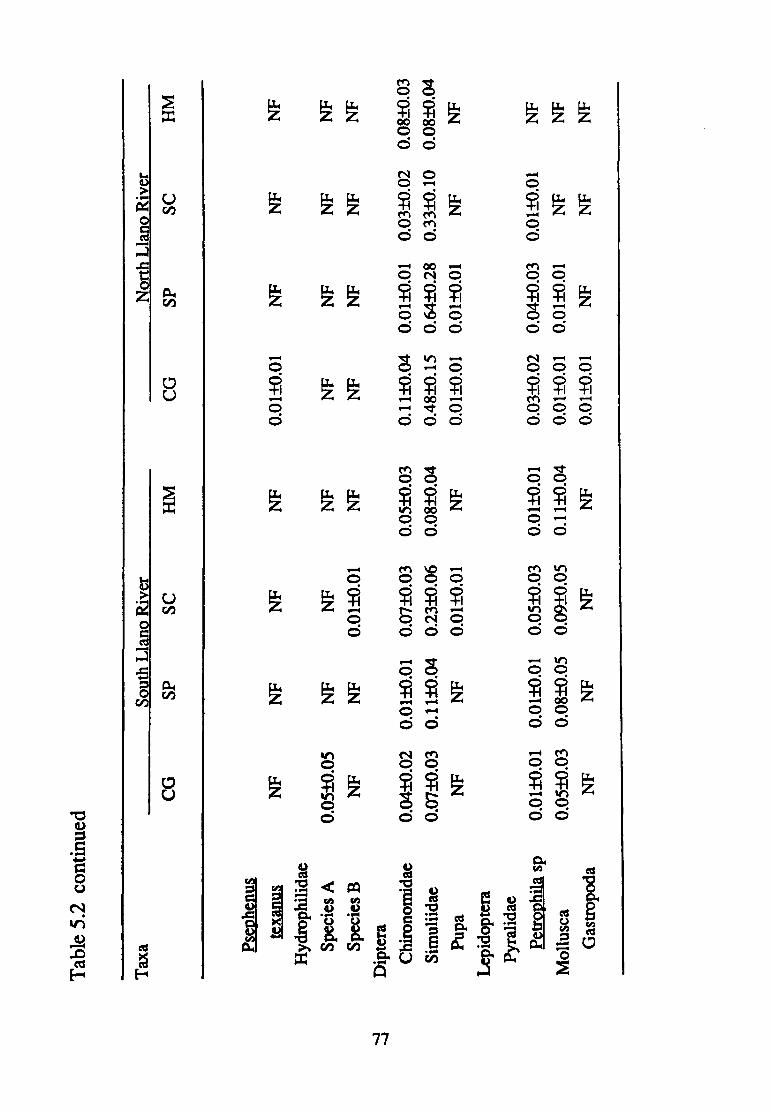
Tab
le 5
.2 c
onti
nued
Tax
a So
uthL
lano
Riv
er
Nor
th L
lano
.RiY
er
CG
S
P
SC
H
M
CG
S
P
SC
H
M
Psm
>hen
us
texa
nus
NF
N
F
NF
N
F
0.01
±0.
01
NF
N
F
NF
Hyd
roph
ilid
ae
Spe
cies
A
0.05
±0.
05
NF
N
F
NF
N
F
NF
N
F
NF
Spe
cies
B
NF
N
F
0.01
±0.
01
NF
N
F
NF
N
F
NF
...
.,J
Dip
tera
...
.,J
Chi
rono
mid
ae
0.04
±0.
02
0.01
±0.
01
0.07
±0.
03
0.05
±0.
03
0.11
±0.
04
0.01
±0.
01
0.03
±0.
02
0.08
±0.
03
Sim
ulii
dae
0.07
±0.
03
0.11
±0.
04
0.23
±0.
06
0.08
±0.
04
0.48
±0.
15
0.64
±0.
28
0.33
±0.
10
0.08
±0.
04
Pup
a N
F
NF
0.
01±
0.01
N
F
0.01
±0.
01
0.01
±0.
01
NF
N
F
Lep
idop
tera
Pyr
alid
ae
Pet
ro.p
hila
sp
0.01
±0.
01
0.01
±0.
01
0.05
±0.
03
0.01
±0.
01
0.03
±0.
02
0.04
±0.
03
0.01
±0.
01
NF
Mol
lusc
a 0.
05±
0.03
0.
08±
0.05
0.
09±
0.05
0.
11±
0.04
0.
01±
0.01
0.
01±
0.01
N
F
NF
Gas
trop
oda
NF
N
F
NF
N
F
0.01
±0.
01
NF
N
F
NF

Dryopoid beetles were well-represented within the S:...R (8 species)
and NLR (9 species) sites; however, the waterpenny CPsephenus texanus),
was represented by only one immature specimen in the NLR. Caddisflies
were represented by six families and eight genera, most prominently by
hydropsychids. Mayflies comprised the most individuals from each site;
taxonomic composition included common species (8 genera and 4 families)
of the Edwards Plateau. The SLR harbors eight naucorid (Hemiptera)
species (Sites and Willig, 1991) which is most species-rich temperate
naucorid fauna. Four naucorids were found in the SLR riffle site and three
in the NLR site.
The most obvious difference in riffle biota between the NLR and SLR
involved the total density of invertebrates. Macroinvertebrate densities in
the SLR site were twice that of the NLR site. This difference was
accentuated by the presence of several common species (including two
beetles and one mayfly), whose densities were consistently higher in the
SLR. In addition, conspicuous differences in certain predator densities were
detected between sites. For instance, five invertebrate predators appear to
dominate the SLR. The hellgrammite, Cozydalus comutus (Corydalidae), is
the top invertebrate consumer, feeding on all consumer trophic levels
(Camilo and Willig, unpublished data). Similarly, four naucorids (A.
circumcinctus, A. lunatus, C. hun~erfordi and L.lutzil are prevalent
consumers that feed at more than one trophic level (Camilo and Willig,
unpublished data). A contrasting scenario existed at the NLR, wherein only
one predator (A. circumcinctus) occurred in substantial numbers. This
species existed at lower densities in the SLR than in the NLR. Consistently
lower densities in the SLR may be a result of interspecific interactions
(Chapter IV), physical differences between the riffles (Table 5.1 ),
78

differences in time since perturbation (York et al., unpublished data) or a
combination of each. Nevertheless, the faunas in the SLR and NLR are
diverse and constituent species are abundant. Overall the faunas are
strikingly similar; however, the biotic structure of each site differs in various
respects.
South Llano River
Forty-six taxa were collected from samples in the SLR (Table 5.2).
The mayfly, Isonychia sicca clearly dominated the riffle by contributing
37.9% of the total individuals. Four of the 46 species a. sicca,
Hexacylloepus ferruginius (24.6%), Microcylloepus pusillus (9.6%) and
Thraulodes gonzalesi ( 4. 7%) comprised 76.8% of all individuals.
North Llano River
Forty-seven taxa were collected from the NLR (Table 5.2), 44 of
which co-occurred in the SLR. Again, I. sicca dominated the riffle
comprising 29.9% of all individuals. Four species[! . .s,im, T. gonzalesi
(13.3%), H. ferruginius (7.0%), M. pusillus (2.7%)] accounted for 52.9% of
all individuals collected.
Taxonomic composition
Substratum size
For every paired contrast of substratum size, taxonomic composition
varied very highly significantly (P<O.OOl) between the SLR and NLR (Table
5.3). Within the gravel vs. pebble, strong association by I. sicca with the
pebble substratum and by H. ferru~inius for the gravel substratum
contributed to much of the compositional variation between substrata at both
79
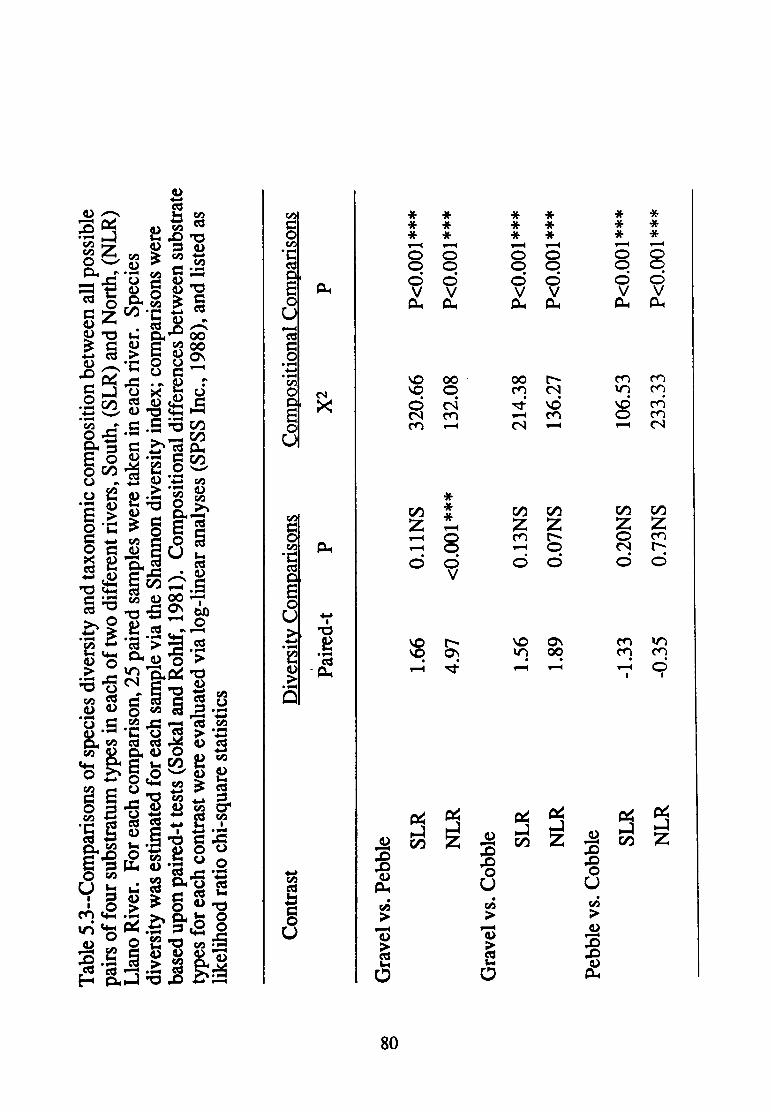
00
0
Tab
le 5
.3--
Com
pari
sons
of s
peci
es d
iver
sity
and
taxo
nom
ic c
ompo
siti
on b
etw
een
all p
ossi
ble
pair
s o
f fou
r su
bstr
atum
type
s in
eac
h o
f tw
o di
ffer
ent r
iver
s, S
outh
, (S
LR
) an
d N
orth
, (N
LR
) L
lano
Riv
er.
For
eac
h co
mpa
riso
n, 2
5 pa
ired
sam
ples
wer
e ta
ken
in e
ach
rive
r. S
peci
es
dive
rsit
y w
as e
stim
ated
for
eac
h sa
mpl
e vi
a th
e S
hann
on d
iver
sity
inde
x; c
ompa
riso
ns w
ere
base
d up
on p
aire
d-t t
ests
(S
okal
and
Roh
lf,
1981
). C
ompo
siti
onal
dif
fere
nces
bet
wee
n su
bstr
ate
type
s fo
r ea
ch c
ontr
ast w
ere
eval
uate
d vi
a lo
g-li
near
ana
lyse
s (S
PSS
Inc.
, 19
88),
and
list
ed a
s li
keli
hood
rat
io c
hi-s
quar
e st
atis
tics
Con
tras
t D
iver
sity
Com
pari
sons
C
ompo
siti
onal
Com
pari
sons
' Pai
red-
t p
X2
p
Gra
vel v
s. P
ebbl
e SL
R
1.66
0.
11N
S
320.
66
P<0.
001 **
* N
LR
4.
97
<0.0
01 **
* 13
2.08
P<
0.00
1 **
* G
rave
l vs.
Cob
ble SL
R
1.56
0.
13N
S
214.
38
P<0.
001 **
* N
LR
1.
89
0.07
NS
13
6.27
P<
0.00
1 **
* P
ebbl
e vs
. Cob
ble SL
R
-1.3
3 0.
20N
S
106.
53
P<O
.OO
l ***
N
LR
-0
.35
0.73
NS
23
3.33
P<
O.O
Ol *
**
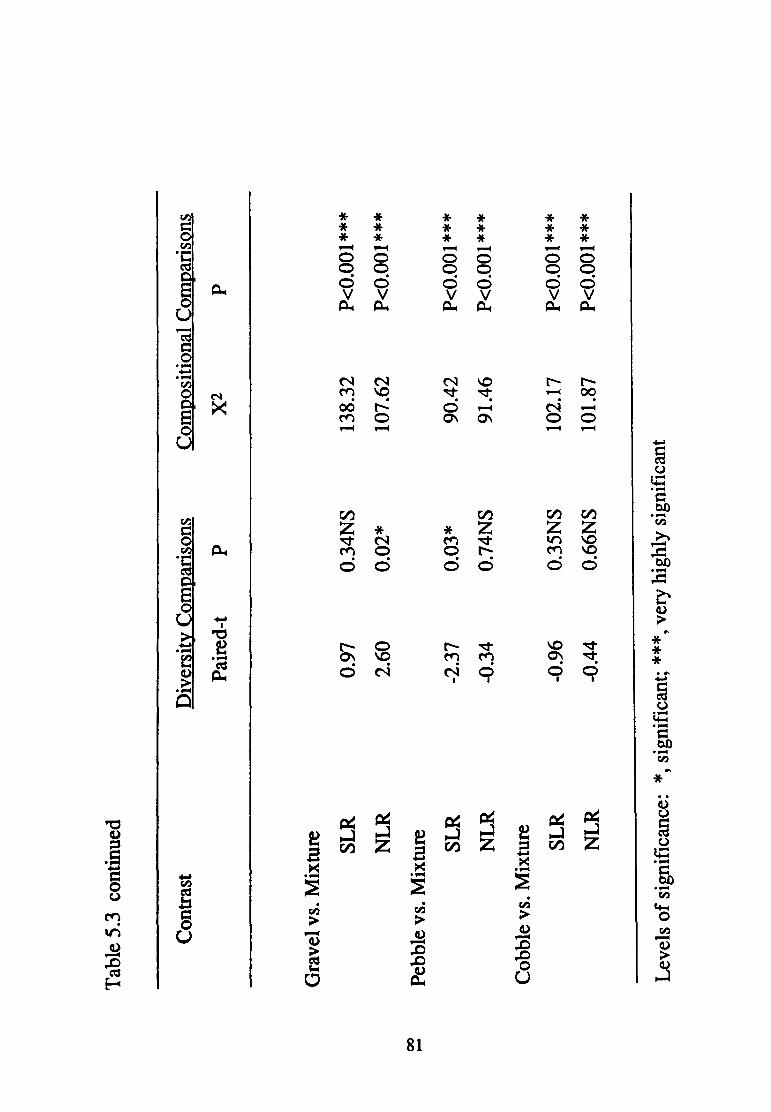
Tab
le 5
.3 c
onti
nued
Con
tras
t D
iver
sity
Com
pari
sons
C
ompo
siti
onal
Com
pari
sons
Pai
red-
t p
X2
p
Gra
vel v
s. M
ixtu
re
SL
R
0.97
0.
34N
S
138.
32
P<
0.00
1 **
*
NL
R
2.60
0.
02*
107.
62
P<
0.00
1 **
* 0
0
Peb
ble
vs.
Mix
ture
-
SL
R
-2.3
7 0.
03*
90.4
2 P<
O.O
Ol *
**
NL
R
-0.3
4 0.
74N
S
91.4
6 P<
O.O
Ol *
**
Cob
ble
vs.
Mix
ture
SL
R
-0.9
6 0.
35N
S
102.
17
P<0.
001
***
NL
R
-0.4
4 0.
66N
S
101.
87
P<
0.00
1 **
*
Lev
els
of
sign
ific
ance
: *,
sig
nifi
cant
; **
*, v
ery
high
ly s
igni
fica
nt

sites. In the gravel vs. cobble contrast in the NLR, Leptohyphes sp.B. and
Dactylobaetis mexicanus occurred more frequently in the cobble over the
gravel; this significantly contributed to the overall difference in taxonomic
composition between the substrata. Within the SLR, a strong affmity by H.
ferru~inius for gravel compared with cobble was again evident and
significantly contributed to compositional variation between substrata. In
the pebble vs. cobble contrast, strong substratum associations by four species
of mayflies significantly contributed to the overall variation in compositional
differences between substrata. Influences by I. sicca (pebble) and D.
mexicanus (cobble) in the SLR and Falleceon quilleri (cobble) and
Leptohyphes sp. B (cobble) in the NLR were most evident.
Hetero~eneity
In the gravel vs. heterogeneous mixture contrast, much of the
variation involved with compositional differences at each site was affected
by strong associations of I. sicca with the heterogeneous substratum. The
strong associations of Marilia sp. and H. ferru~inius for the gravel within the
NLR and SLR, respectively, also contributed to the variation. A caddis fly,
Helicopsyche sp. had little effect on compositional variation between
substrata in previous contrasts; however, it significantly contributed to
differences in the pebble vs. heterogeneous mixture contrast in the NLR as
evidenced by its strong association with cobble. In the SLR, the mayfly, T.
gonzalesi (pebble), and a trichopteran, Smicridea sp. (heterogeneous
mixture), each exhibited distinct habitat associations and contributed
substantially to overall differences between substrata. In the cobble vs.
heterogeneous mixture, a strong association between H. ferruginius and the
82

heterogeneous substratum in both rivers accentuated compositional
differences between substrata.
Diversity comparisons
Although highly significant compositional differences associated with
substratum size existed in all cases, species diversity differed between
substratum types in only one instance within the SLR (Table 5.3). This
difference occurred between the pebble and the heterogeneous mixture; the
latter supporting significantly (P=0.03) higher diversity than the former.
Results obtained in the NLR differed from those in the SLR. Two instances
were detected in which a particular substratum supported higher species
diversities than its paired counterpart. In the gravel vs. pebble contrast,
species diversity was significantly higher (P<O.OOl) in the gravel
substratum. Similarly, in the coarse gravel vs. heterogeneous mixture
contrast, the gravel substratum yielded a higher species diversity than did the
heterogeneous mixture.
Although three instances were detected in which substratum type
affected species diversity, no pattern existed in both rivers. Moreover, those
instances in which species diversities differed between substrata provided
little support for hypotheses that favor heterogeneity as a contributor to
species diversity in benthic communities. In fact, in one instance,
significantly higher species diversity was detected for a homogeneous
substratum (gravel) over the heterogeneous mixture in the NLR. Substratum
size and heterogeneity influence stream biota in many fashions. Most
notable are those involved with taxonomic composition. This study
illustrates how differences in habitat structure may support equally diverse
assemblages yet may lead to highly significant differences in taxonomic
83

composition. Approaches that examine species diversity from multiple
perspectives including taxonomic considerations may facilitate a
fundamental understanding of biodiversity and help elucidate community
level processes.
84

Literature Cited
Allan, _D. J. 1 ?75. The distributional ecology and diversity of benthic msects tn Cement Creek, Colorado. Ecology 56:1040-1053.
Baker, V. R. 1977. Stream-channel response to floods, with examples from central Texas. Geol. Soc. Am. Bull. 88:1057-1071.
Cummins, K. W. and G. H. Lauff. 1969. The influence of substrate particle size on the microdistribution of stream macrobenthos. Hydrobiologia 34:145-181.
de March, B.G.E. 1976. Spatial and temporal patterns in macrobenthic stream diversity. J. Fish. Res. Board Can. 33:1261-1270.
Erman, D.C. and N.A. Ennan. 1984. The response of stream macroinvertebrates to substrate size and heterogeneity. Hydrobiologia 108:75-82.
Fuller, R. L. and P. S. Rand. 1990. Influence of substrate type on vulnerability of prey to predacious aquatic insects. J. N. Am. Benthol. Soc. 9:1-8.
Krebs, C. J. 1985. Ecology: the experimental analysis of distribution and abundance. Harper and Row, New York.
Lamberti, G.A. and V. H. Resh. 1979. Substrate relationships, spatial distribution patterns, and sampling variability in a stream caddisfly population. Environ. Entomol. 8:561-567.
MacArthur, R. H. 1957. On the relative abundance of bird species. Proc. Nat. Acad. Sci. 43:293-295.
Magurran, A. E. 1988. Ecological diversity and its measurement. Princeton University Press, Princeton, New Jersey.
Minshall, G. W. and J. N. Minshall, 1977. Microdistribution of benthic invertebrates in a Rocky Mountain (U.S.A.) stream. Hydrobiologia 55:324-355.
85

Minshall, G. W. 1984. Aquatic insect-substratum relationships, pp. 358-400 . .m.v. H. Resh and D. M. Rosenberg (eds.). The ecology of aquat1c msects. Praeger, New York.
Reice, S. R. 197 4. Environmental patchiness and the breakdown of leaf litter in a woodland stream. Ecology 55:1271-1282.
Sites, R. W. and M. R. Willig. 1991. Microhabitat associations of three sympatric species of Naucoridae (Insecta: Hemiptera). Environ. Entomol. 20: 127-134.
Sokal, R. R. and F. J. Rohlf. 1981. Biometry: the principles and practice of statistics in biological research, 2nd ed. Freeman, San Francisco.
SPSS Inc. Advanced statistics guide. 2nd Edition. pp. 527. Chicago n.
Sugihara, G. 1980. Minimal community structure: an explanation of species abundance patterns. Amer. Nat. 116:770-787.
Ward, J. V. 1992. Aquatic insect ecology. 1. Biology and habitat. Wiley and Sons, Inc. New York.
Wentworth, C. K. 1922. A scale of grade and class terms for cladistic sediments. J. Geol. 30:377-392.
Williams, D. D. 1980. Some relationships between stream benthos and substrate heterogeneity. Limnol. Oceanogr. 25: 166-172.
86