Embed Size (px)
Citation preview

Mark Botirius
Describe the steps in CTAB DNA extraction and explain what is happening in each step
1. Establish the big picture.
The first step when performing a CTAB DNA extraction is to consider those factors that may affect how you proceed with the extraction. These factors include such things as what type of tissue is represented in the sample, the size of the organism’s genome, how fresh the sample is, how much tissue there is to work with, and even such things as how many centrifuge tubes will be needed and labeling. For example, is the sample from a plant, animal, or fungus? Each of these organisms have different cell wall characteristics that may alter how those steps designed to release the DNA into the extraction buffer are executed. Plants and fungi have cell walls composed of cellulose and chitin respectively, in addition to membranes that must be broken down to allow access to the DNA, whereas most animals only have membranes. Consequently, if the sample is from a plant, it would need to be ground up, while a muscle sample may only need to have its cell membranes disrupted by a detergent, such as CTAB. Another example is whether or not the sample is fresh or frozen, and how old the sample is. Fresh samples, of course, contain the highest quality of DNA, whereas older DNA is subject to degradation that varies with age and the conditions in which the DNA was preserved. Applying this step to the extraction we did in class reveals that the organism was a plant (Vicia faba) and the sample was fresh. Also, we had plenty of material to work with. With these factors in mind, it was determined that we had to freeze the tissue solid in dry ice and then grind it in a coffee grinder. Why? Because of the tissue type factor. Plant tissue means the cells have cell walls made of cellulose, freezing makes them brittle (and helps preserve the DNA), and grinding breaks them apart. If we had been working with animal cells, for example, we may have been able to skip these steps, and went straight to the detergent step since it is possible that the only thing needed is to disrupt the membranes. Having established that Vicia faba is my model organism for this discussion and that all of my prerequisite preparations have been completed (such as labeling all tubes and such), it is time for the next step. (Rogers, 2016)
2. Gain access to the DNA
Great. I have plenty of Vicia faba tissue, but it is all sequestered behind protective cell walls and membranes. I need to gain access to that DNA (I can’t extract something that I have no access to) while at the same time, protecting it from damage and degradation. How this is done, as I have already pointed out, can vary depending on several circumstances that I have already described. In this case, I have plant tissue, so for this step the tissue is frozen in liquid nitrogen (or dry ice, as was done in lab), and everything that comes into contact with the tissue is frozen as well. As I have already mentioned, this makes the cell walls brittle and helps preserve the DNA. The frozen tissue is then ground up (either manually in a mortar and pestle, or by using a small coffee grinder) to break the already brittle walls and membranes apart and expose the DNA. Of course, the dry ice is allowed to sublime before going further. (Rogers, 2016)
3. Break apart the membranes, protect the DNA and keep it in solution.
Okay. I have succeeded in getting past the cell walls. However, much of the DNA may still be protected behind nuclear and cellular membranes. Therefore, I must take steps to break apart those membranes and then protect the DNA without delay. To accomplish this, immediately after
Page 1 of 8

Mark Botirius
grinding the DNA in dry ice and allowing the ice to sublime, an equal volume of hot (65 ) ℃ 2X CTAB buffer is added to the tissue. Cell membranes consist of a phospholipid bilayer. Because CTAB is a detergent, it solubilizes the phospholipids and causes the membranes to disintegrate, exposing the DNA. Unfortunately, the DNA is now exposed to anything that can damage or degrade it, such as nucleases. Fortunately, CTAB buffer also contains EDTA, which chelates the cofactors needed by these nucleases (Mg2+) and thereby stopping them from degrading the DNA. Tris is also present to maintain a stable pH (around 8.0 to further protect the DNA from degradation). Lastly, it is necessary to keep the DNA in solution. If it were to precipitate out at this time, it would simply be a part of all of the other junk solids present in the sample and it would be impossible to isolate. Therefore, CTAB buffer also contains 1.4M NaCl. This creates a sodium salt of DNA which keeps it in solution. In order to ensure that the CTAB buffer has sufficient time to solubilize the membranes the solution is placed in a water bath (65℃) for one to five minutes. (Rogers, 2016)
4. Extract proteins
At this step in the procedure, I have an aqueous solution that also has a lot of junk solids that were never in solution (such as plant fibers) and some molecules that have precipitated out by interacting with CTAB (CTAB, being a cationic molecule, will complex with some proteins and polysaccharides that have a negative charge somehow associated with the molecule, taking them out of solution). Regrettably, however, cells naturally contain a whole host of water soluble proteins because, let’s face it, much of the cell is an aqueous environment. Which means that, although I have a lot of unwanted stuff that is not in solution and therefore can be removed, I still have a lot of unwanted proteins in solution along with my DNA, and I need to separate those proteins from my DNA. In other words, I need to extract the proteins, and leave the DNA salt in solution. But how can this be done? I do this by adding another phase to my solution.
So far, my solution has consisted of only one phase, an aqueous phase, represented by the CTAB buffer solution. Most salts are happy in aqueous phases, and right now, my DNA exists as a sodium salt of DNA. To extract the aqueous proteins, I need to add another phase, an organic phase. Aqueous phases are polar (water is a polar molecule), and most organic phases are non-polar (or less polar than water). This difference in polarity is the basis behind separating two different substances in a solution, or extracting one substance from another. Since the two solvents have different polarities, they also have different solubility, and will separate in solution. For example, suppose I have a solution of benzoic acid, m-nitroaniline, and naphthalene in a non-polar organic phase such as ether. Ether, is non-polar. To separate out the benzoic acid, I add a NaOH aqueous phase. The polar, aqueous phase and the non-polar organic phase form two layers. When they are forced to mix, however, the benzoic acid in the organic phase (ether) reacts with the NaOH in the aqueous phase, forming a salt (benzoate- + Na+) which is highly soluble in the aqueous phase (not unlike our DNA salt) and insoluble in the organic phase. This causes the benzoic acid to transfer to the aqueous phase as benzoate (the conjugate base of benzoic acid) from the organic (ether) phase. To get the benzoic acid back, simply siphon off the aqueous phase (remember, the two phases don’t mix, but form two layers) and add HCl to convert the benzoate back to benzoic acid. Benzoic acid has just been extracted from the ether. The ether solution now primarily contains only m-nitroaniline and naphthalene. (Chung, 2015)
Page 2 of 8

Mark Botirius
The principles behind extracting the proteins from our CTAB solution are very similar. Instead of extracting out of an organic phase into an aqueous phase, however, we are extracting out of an aqueous phase (CTAB) into an organic one (chloroform). In addition, in order to get benzoic acid to change its solubility from ether to water we used NaOH to change it to benzoate (its conjugate base). Similarly, the proteins in our CTAB solution also change. However, this time acid-base chemistry isn’t used, the property that proteins can change conformation is what is used. When the chloroform (organic phase) is mixed with the CTAB (aqueous phase) to form an emulsion, the chloroform causes the proteins to denature thus adopting a radically different conformation. They
don’t just adopt any old conformation, however, they adopt a non-polar one, thereby changing their solubility from polar (aqueous/CTAB) to a non-polar (organic/chloroform) causing them to move from the CTAB to the chloroform solution. (Oswald, 2016) Now that the proteins are in the chloroform, the aqueous/CTAB phase contains mostly just the DNA salt. The CTAB/chloroform solution is centrifuged to separate the two phases (chloroform is a little heavier) and the phase containing the DNA (aqueous, or top) is siphoned off. Of course, no chemical reaction is 100%, and so to maximize the removal of proteins, the chloroform extraction is performed twice. (Rogers, 2016)
5. Precipitate the DNA
Awesome, now I have removed all of the plant debris, and also removed most of the soluble proteins. My aqueous solution, however, most likely still contains impurities. For example, although chloroform is much less polar than water, it nonetheless has a definite dipole moment that is apparent when looking at the molecular structure. Chlorine is more electronegative than carbon, and therefore the electrons spend more time at the chlorine atoms than they do at the hydrogen atom. This allows the molecule to participate in some hydrogen bonding, although not nearly as much as water. Which means that chloroform is
slightly soluble in water, and no matter how much the solution is centrifuged to remove it, there is probably some
dissolved in solution regardless. A good way to get rid of the impurities still left in solution, is to precipitate the DNA out of solution, and discard the solution along with its impurities, leaving only the DNA behind. To do this, CTAB precipitation buffer is added to the aqueous phase. This buffer is the same as the CTAB buffer, only the CTAB precipitation buffer has no NaCl. As a result, the DNA forms a salt with the CTAB molecule itself instead of the Na+. Recall that CTAB is cationic, and therefore in the absence of the sodium cation, it will form a salt with
Page 3 of 8
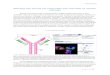
Figure 1. Polar proteins that reside in the aqueous portion of the cytoplasm denature to become non-polar. This causes them to transfer from an aqueous phase to an organic phase, where they can be removed from solution. This figure is from BitesizeBio.com
Figure 2. Chloroform, the picture is from Wikipedia

Mark Botirius
the DNA molecule as well. However, the solubility of ionic substances (such as salts) exhibit great variability. For example, 36 grams of NaCl can be dissolved in 100 mL of water, while only 0.002 grams of calcium phosphate can be dissolved. To see why the CTAB salt of DNA is much less soluble than the sodium salt, one need only look at the sodium and CTAB ions.
Obviously, the CTAB cation has a relatively huge nonpolar “tail” 16 carbons long. There is no doubt, this would affect the solubility of the salt it forms with DNA. After the precipitation buffer is added and the CTAB salt of DNA precipitates out, DNA is pelleted in a centrifuge and the supernatant is discarded (along with any remaining impurities, such as proteins, or even chloroform). (Rogers, 2016)
6 Convert back to a sodium salt of DNA
It looks like I should be done. I have a relatively pure pellet of DNA. There is only one problem, however. CTAB is, after all, a cationic detergent, and as such, isn’t acceptable in a lot of biological procedures. Therefore, in this form, the DNA is pretty much useless. To be useful, it needs to be converted back to a sodium salt of DNA. This is not a problem; the DNA pellet is simply dissolved in a high sodium salt buffer called “high salt TE”. This buffer not only contains 1M of salt, it also has Tris (to maintain pH), and EDTA (to protect the DNA from nucleases). In the high sodium solution, the CTAB ion is replaced with sodium, and the DNA goes back in solution. (Rogers, 2016)
7. Re-pellet the sodium salt of DNA
Once again, we have our sodium salt of DNA, however it is in solution. We cannot add CTAB to precipitate our DNA, so we change our solvent instead. Last time we used CTAB to make our molecule more non-polar in a highly polar solvent. This time, we change our solvent to be less polar when our molecule is highly polar. This, along with lower temperature, gives us the same result. Our DNA precipitates out of solution and can be pelleted. The less polar solvent used is 100% cold ethanol. The DNA pellet is centrifuged, and again the supernatant is discarded. Finally, our DNA pellet is “washed” in cold 80% ethanol and centrifuged. Finally, it is rehydrated in 0.1X TE when it is to be used in molecular biology methods. (Rogers, 2016)
Define and explain protein trafficking, as it exists in eukaryotic cells. Be sure to address the movement of all of the cell compartments that are the destinations of various proteins.
Proteins serve a wide variety of functions. They serve enzymatic, structural, movement, defense, storage, and regulatory functions just to name a few. Even though they serve many different functions throughout the organism, they are all synthesized by the ribosome (non-ribosomal peptides notwithstanding). Because their functions are so varied, the locations where
Page 4 of 8
Figure 3. The difference between a sodium ion and CTAB ion clearly illustrate why the CTAB salt is less soluble.VS Na+

Mark Botirius
they serve these functions are equally varied, and so there must be some way to direct a particular protein from its site of synthesis (the ribosome) to its site of function. For example, a membrane protein must be directed from the ribosome to the membrane, while an enzymatic protein synthesized at the ribosome must be directed to, for the sake of example, the cytoplasm (I am not saying here, that all enzymes wind up in the cytoplasm). Therefore, protein trafficking refers to the transport of proteins from where they are synthesized to a particular destination where they are needed. This can be an extremely complex question because, for example, in humans there are well over 100,000 proteins each with its own function and destination. Luckily, however, almost all of those proteins can be placed in one of two categories; those proteins that move from the ribosome to the lumen of the endoplasmic reticulum, and those that move from the ribosome to the cytosol. Proteins that travel through the endoplasmic reticulum either stay in the endomembrane system, become part of the membranes themselves, or are secreted from the cell. On the other hand, those proteins that travel into the cytosol usually remain in the cell and either stay in the cytosol or they travel to various cellular destinations such as the organelles, or the cytoskeleton to name a few. (Rogers, 2016)
Proteins that travel to the lumen of the endoplasmic reticulum
Although it is true that the rough endoplasmic reticulum is a place where the ribosomes are attached to the cytosolic surface of the endoplasmic reticulum, it should be noted that these ribosomes are not permanently attached. In theory, all ribosomes are actually free. The attachment of the ribosome is actually part of protein trafficking. It is where protein trafficking begins for those proteins that must travel into the lumen of the endoplasmic reticulum. In protein synthesis, amino acids are added to the carboxyl end of the growing peptide chain, which means that the amino end of the protein first emerges from the ribosome. Those amino acids that are destined for the ER lumen contain a signal sequence close to the amino end that is recognized by a signal recognition protein in the ER membrane that binds the sequence thereby stopping translation. When the SRP comes into contact with an SRP receptor, it binds to the receptor and in so doing attaches the ribosome to the ER cytosolic membrane. SRP receptors are located next to transmembrane proteins called translocons, and the bound ribosome, now attached to the ER, continues translation as the growing peptide is fed into the translocon and through the ER membrane into the lumen. For this reason, this process is called co-translational translocation. If, on the other hand, the final destination of the protein happens to be inside the ER membrane itself, it will contain a hydrophobic sequence that causes the translocon to eject
Page 5 of 8
Figure 5. When the translocon (the blue transmembrane protein) encounters a hydrophobic domain of the growing peptide, it expels the nascent protein to the side and into the ER membrane. This figure is from www.zoology.ubc.ca
Figure 4. Those proteins that are trafficked into the ER lumen travel through the Golgi complex and have one of three fates. They either become an integral member protein, stay in the ER lumen, or are packaged into a secretory vesicle and leave the cell. This figure is from the class lecture.

Mark Botirius
the growing peptide laterally and into the ER membrane where it becomes an integral membrane protein. (Iwasa, 2016)
The trafficking of nascent proteins that wind up within the ER lumen is done by vesicular transport. The destination of these proteins is determined by various sequences. For example, those proteins destined to stay within the ER lumen contain a short retrieval sequence of lys-asp-glu-leu (KDEL in canonical nomenclature) that is recognized by a receptor in a COPI coated vesicle that ensures the protein will stay in the ER where it belongs. Those proteins without KDEL sequences continue on to the Golgi complex in COPII coated vesicles. In the Golgi complex, proteins are identified by certain sorting signals and processed for trafficking accordingly. For example, those proteins destined for the lysozyme contain phosphorylated mannose residues that are recognized by receptors in membranes that form clathryn coated vesicles that bud off from the Golgi complex and travel to the lysosome. Most proteins without these signal residues end up in a secretory vesicle or they have signal sequences in their cytoplasmic membrane domains that destine them to become integral cellular membrane proteins. (Iwasa, 2016)
That pretty much sums up the trafficking associated with those proteins that travel into the lumen of the ER. However, there is one last thing that I think I should address that does not involve proteins directly, but nonetheless plays a role in their transport. As I have previously stated, proteins that end up in the ER lumen are trafficked in transport vesicles that move from the ER to the Golgi complex and beyond. These vesicles also move the opposite direction, as in the case of COPI vesicles. Although these vesicles aren’t proteins themselves, they have protein “cargo”. So the question is, how do these protein transporting vesicles move? (Iwasa, 2016)
Most likely, they move along microtubule “tracks” that exist within the cell. Microtubules are cytoskeletal elements constructed of α and β tubulin subunits that give the microtubule polarity. Microtubules are often synthesized so that their “plus” end points away from the center of the cell which means their “minus” ends point towards the center. This is important because there are two motor proteins associated with microtubules that use ATP to move cell elements such as vesicles. Kinesin is the motor protein responsible for moving vesicles towards the plus end of the microtubule (towards the outside of the cell) and dynein is responsible for moving vesicles towards the minus end (towards the inside of the cell). This means, that COPII and clathryn coated vesicles are most likely powered by kinesin, while COPI coated vesicles are powered by dynein. The association of a particular vesicle with a particular direction is mediated by vesicular proteins (such as “Rabs” and “SNARE’s”) that set the vesicular specificity. (Iwasa, 2016)
Proteins that move from the ribosome to the cytosol
Those proteins that do not have a signal sequence recognized by an ER membrane receptor are released by the ribosome into the cytosol and they have different sequences at their amino ends that are recognized by receptors located on the organelles they are destined for. Generally speaking, those target organelles are the peroxisome, the mitochondrion, the chloroplast, and the nucleus. (Rogers, 2016) (Iwasa, 2016)
Those proteins intended for the peroxisome have a peroxisomal targeting signal sequence that is recognized by a
Page 6 of 8
Figure 6. Those proteins that go from the ribosome directly into the cytosol are destined for cellular organelles. This figure is from the class lecture.

Mark Botirius
membrane receptor on the peroxisome. When the protein encounters the peroxisome receptor (also called an importomer) it is shuttled directly into the peroxisome in its completed, folded tertiary configuration. How this is done is not completely understood. With regards to the mitochondrion, the nascent proteins contain a removable sequence (called a presequence) and they associate with molecular chaperones (Hsp70 and Hsp90) that help them maintain a relatively unfolded configuration so that they can make it through the mitochondrial membrane. When they encounter a mitochondrial receptor (called a TOM complex) they are translocated across the membrane.
Figure 7. This figure is from class illustrating mitochondrial protein trafficking
Once inside the intermembrane space, those proteins destined to become inner membrane proteins enter the TIM22 complex, whereas those destined for the mitochondrial matrix pass through the TIM23 complex. Once inside the matrix, their presequence is spliced off. Chloroplast protein trafficking is very similar. It also relies on translocation processes and peptide signal sequences, with the main difference having to do with the fact that the chloroplast has six sub compartments to the four possessed by mitochondria. Finally, proteins destined for the nucleus are similar to those destined for the other cellular organelles in that they also contain a sequence that serves as an “address” directing them to their destination. (Iwasa, 2016)
Page 7 of 8

Mark Botirius
ReferencesChung. (2015, 9 25). Organic Chemistry Class Lecture on Extraction Lab. Bowling Green, Ohio, United
States of America.
Iwasa, J. a. (2016). Karp's Cell and Molecular Biology (8th ed.). Hoboken: Wiley.
Oswald, N. (2016, July 16). Retrieved from BitesizeBio: http://bitesizebio.com/384/the-basics-how-phenol-extraction-works
Rogers, S. (2016, October). Class lecture. Bowling Green, Ohio.
Page 8 of 8