Embed Size (px)
Citation preview

Mark Botirius
Describe how the immune system recognizes foreign organisms, reacts to the foreign organisms, and then immobilizes and kills them.
The immune system is a highly specialized and complex entity that utilizes a variety of strategies for the recognition, immobilization, and destruction of foreign organisms. Although it is complex, the strategies of the immune system can be divided into two general categories: those strategies employed by the innate system, and those employed by the adaptive system. Dividing the immune system into these two general categories has the added benefit of bestowing a chronological component to the immune system, which provides another way to organize and make sense of its extremely complicated structure because, generally speaking, the innate system reacts first, followed by the adaptive system. Those elements that work to eliminate pathogens that do not involve any cellular participation will be excluded, simply because I do not perceive them to be relevant to the answer the question is looking for. Specifically, those parts of the immune system related to anatomical barriers to infection (e.g. the skin), non-specific physical and chemical defenses (e.g. mucous, saliva, antimicrobial peptides found in sweat and/or tears etc.) and the complement system (a totally passive system that does not actively recognize or react to microbes, but does so passively) will not be discussed. 1
In order to describe how the innate system reacts, it is necessary to first describe what is doing the reacting (in other words, who are the “players” of the system?) followed by how the system reacts. Therefore, what is to follow will be a description of the following “players” of the innate system: granulocytes (neutrophils, basophils, mast cells, and eosinophils), myeloid antigen presenting cells (macrophages and dendritic cells), and NK (natural killer) cells. The discussion of some of these cells (for example, dendritic cells) will overlap into the adaptive system because, of course, activation of the innate system leads to the activation of the adaptive system.
Granulocytes
The common defining characteristic of granulocytes is, as the name indicates, the presence of granules that are released to kill any pathogens that they encounter. In addition, granulocytes exhibit odd shaped, multi-lobed nuclei in contrast to the round nuclei found in lymphocytes. While all granulocytes possess granules, only neutrophils and eosinophils are phagocytic. There are many different types of granules secreted by granulocytes. Some examples are: proteases (elastase, collagenase) antimicrobial proteins (defensins, lysozyme), histamine, cytokines, and chemokines. Histamines, cytokines, and chemokines are particularly important. Cytokines up regulate the immune response while chemokines are a chemical signal that attracts more white blood cells. Histamines are important because they promote vasodilation and inflammation that brings more blood to the site of infection. The effects of these three proteins add together, to result in recruiting more cells to the site of infection in order to combat the invading pathogens. Lastly, another
1 I began to include the complement system in my original draft and it became increasingly clear to me that, although the complement system plays a major role in the immune response, it nonetheless plays a passive, cooperative role with the innate and adaptive systems, not an active one. As a result, it didn’t add much to the answer and I deemed it to be ancillary to what the question was asking.
P a g e 1 | 9

Mark Botirius
important chemokine secreted by granulocytes are cell adhesion molecules (CAM’s) which cause newly arrived white blood cells to slowly “roll” along the blood vessels at the site of infection and extravasate into the infected tissues. (Owen, 2013, p. 168)
Myeloid Antigen Presenting Cells
The principal importance of myeloid antigen presenting cells is to take in the antigens released by pathogens and then migrate to the secondary lymph organs to present these antigens to activate the T cells and B-cells of the adaptive system. There are several ways that APC’s can accomplish this. Macrophages are phagocytic and are not only able to provide a first line of defense against invading microbes, but are also able to internalize and process invaders into antigens for presentation to T cells. Dendritic cells are the most highly specialized and effective APC’s that continually scan for antigens to take in by phagocytoses, pinocytosis, or receptor mediated endocytosis. Once they have their antigen, dendritic cells migrate into the secondary lymphoid organs (such as the lymph nodes, to name one) to present their antigen to T cells and B-cells. (Owen,2013, p. 33)
Natural Killer (NK) cells and NKT cells
Although natural killer cells are lymphoid in lineage they are considered to be part of the innate system nonetheless because they do not exhibit antigen specific receptors, which is one of the defining characteristics of the adaptive system. Explaining why these cells are part of the innate system provides me an excellent segue into an important distinction between the innate and adaptive systems. That is the difference in how the two systems recognize pathogens. The innate system is the first responder when it comes to infection. The question is, how does the system
recognize something that it has never seen before? The answer is that a great deal of invading microbes is mostly comprised of four general types: viruses, fungi, parasites,
and bacteria. Fortunately, these four types of pathogens have a great deal in common, which has provided evolutionary selective
pressure for immune systems to develop ways to combat these microbes by recognizing what they have in common. In the language of immunology, these common characteristics shared by a particular group of microbes are called “Pathogen Associated Molecular Patterns” (PAMPs). Cells of the innate system have evolved receptors that recognize these common patterns. These receptors are called “Pattern Recognition Receptors” (PRRs). (Iwasa, 2016, p. 663) How effective are PRRs? Consider that across all bacteria that possess a cell wall (which is almost all that we know of) there are over 100 different types of peptidoglycan. Even with all of this diversity, they are all nonetheless composed of N-acetylglucosamine and N-acetylmuramic acid and are connected with a β-1,4 linkage. (Madigan, 2012, p. 58) With a single PRR (i.e. TLR-2), the immune system can identify any bacteria with a peptidoglycan cell wall (which is almost all of them), and this is just one example. Because PRRs have evolved to recognize a large portion of invading microbes that share a common characteristic, it would be counterproductive for them to be adaptive and changing. Therefore, PRRs are germ line encoded and invariable which is not the case with B-cell and T-cell receptors which are variable and
constructed via somatic DNA recombination. The reason natural killer cells are part of the innate system, is because they also identify their targets using an invariable strategy
P a g e 2 | 9
Table 1. This is just a small sample of only one category of PRR’s known as “Toll Like Receptors”. (Owen, 2013, p. 155)
TLR's TLR1TLR2

Mark Botirius
that is not adaptive in nature. In the case of NK cells, they primarily identify their targets via the absence of MHC class I proteins that is expressed on almost all cells. In the case of NKT cells they do have T-cell receptors, however they are not very diverse, and they are limited in what they recognize (they only recognize particular lipids and glycolipids). In other words, their receptors can be classified as TCR’s, although they are not really the same as the receptors found on actual T-cells. (Owen, 2013, p. 40) NKTs are therefore considered to be a hybrid, spanning both the innate and adaptive categories. Lastly, like granulocytes, NK and NKT cells kill their targets by releasing granules containing antimicrobial proteins. Having described the players involved in the innate response and how they recognize foreign organisms, I can now describe how the innate system reacts and kills foreign organisms.
Leukocytes (mostly neutrophils which comprise 50% to 70%) (Owen, 2013, p. 33)routinely travel around the circulatory system and body tissues searching for pathogens. Suppose someone cuts themselves. PAMP’s of invading pathogens such as bacteria are recognized by the PRRs of leukocytes residing in the area and are destroyed using methods that depend on the type of cell. Neutrophils and eosinophils will begin phagocytizing and using their granules, while basophils and mast cells will attack using their granules as well. In addition to phagocytizing and releasing granules, these cells will also release chemokines (such as IL-8) that bring more leukocytes into the battle and cytokines (such as IL-4, IL-10, and IL-13) that alert other leukocytes of the invasion, thereby up regulating their activity (Owen, 2013, p. 35). Histidine and CAM proteins are also released to aid in the fight by increasing blood flow, causing inflammation, and signaling where arriving leukocytes will begin to roll along the blood vessel epithelium eventually coming to a stop and transmigrating into the infected tissue to join the fight. Meanwhile, APC’s in the area such as dendritic cells and macrophages are taking up antigens released in the battle and are migrating to secondary lymph tissues such as the lymph nodes (or the spleen in the case of blood borne pathogens) to present these antigens to T-cells. Notice that B-cells and T-cells are entirely absent in the fight. In fact, at this point in the explanation, T-cells are literally unable to join the fight, because they do not have PRR’s. T-cells must be activated, and naïve T-cell receptors can only recognize one thing, that is an MHC/antigen combination presented by an APC. Although it is true that B-cells do have some PRRs, they need, for the most part, help from T-cells to become fully activated. This brings me to the question of how the adaptive immune system recognizes pathogens when they do not, overall, use PRRs.
Cells of the adaptive immune system: B-cell and T-cell Lymphocytes
Unlike the PRR’s used by the leukocytes of the innate system, the receptors used by the lymphocytes of the adaptive system are very specific. In other words, they have a very specific target (instead of targeting a general characteristic of all bacteria, it would target a specific protein of a particular bacterium for example). The question is, “How can the target be so specific, in light of the tremendous diversity in the pathogens that attack the host?” The answer lies in how these receptors are generated. Lymphocytes generate their receptors through genetic recombination (Iwasa, 2016, p. 682). I won’t go into the specifics here (it is well beyond the scope of this question) however it is important to note that the result is that an enormous variety of cells expressing specific receptors are generated that can target practically any enemy. To avoid the possibility that a self- targeting receptor is created, lymphocytes are tested and any self-reactive cells are destroyed. Those that remain have the ability to bind a single antigen that isn’t self. It is important to note, that each B-cell and T-cell can bind only one antigen (known as allelic exclusion) (Owen, 2013, p. 303).
P a g e 3 | 9

Mark Botirius
After establishing its particular receptor (that can bind only one antigen) and reaching maturity (another process well beyond the limits of this essay) the mature lymphocytes travel the bloodstream and visit various secondary lymph tissues such as the lymph nodes where the B-cells occupy the primary cortex and T-cells the paracortex, which is where they enter the fight against infection.
Recall that I left off at the point where the granulocytes and macrophages were destroying the invading pathogens, and the APC’s where engulfing antigens and taking them to the secondary lymph tissues (lymph nodes). In the lymph node (or some other secondary lymph tissue) the T-cells and B-cells are browsing through all of the countless MHC/antigen complexes (both class one and
class two) being presented by all of the APC’s until they find one that matches their uniquely generated receptor. When they do find a match, they become activated. This is the mechanism by which the cells of the adaptive system gain the ability to recognize pathogens. It is important to note here, that activation of a T-cell by an APC is the primary event that begins the adaptive response. When a naïve CD4 T-cell encounters an APC that is presenting an antigen that its receptor recognizes, it becomes activated, and it is the CD4 T-cells that fully activate B-cells and CD8 T-cells (T cytotoxic cells). 2 Without these CD4 cells to activate the others, the adaptive response is shut down. This is the reason AIDS is so deadly.
Hence the dendritic cells that were at the site of infection have engulfed antigen, traveled to the
lymph nodes, and were discovered by a T-cell with a receptor that matched the antigen it was presenting. This T-cell differentiates into a CD4 cell that fully activates B-cells and CD8 cells. The B-cells multiply and some of them become plasma cells that continually secret antibodies against the invading pathogens. Other B-cells become memory cells, and still some others set up germinal centers that are the sites for somatic hypermutation. The antigens secreted by the plasma cells circulate and destroy the invading pathogens in several ways. 1. Neutralization. The antigen binds directly to the pathogen blocking the receptors it needs to gain entry into the host. 2. Opsonization. 3. Complement fixation/ activation. 4. Antibody Dependent Cell Mediated Cytotoxicity (ADCC). (Owen, 2013, p. 417)
So far, all of the “players” in my immunological scenario have been addressed with one exception. I have yet to address the role of cytotoxic T-cells (CD8). The reason is because, in the scenario I have created, CD8 cells don’t really play a role. Why? Recall that T-cells are MHC 2 I should note that I am generalizing here, which is why I use the words “fully activate” The immune system is so complex, that it is impossible for me to cover all of the details in a short essay. For example, it is not an absolute requirement for CD8 cells to be activated by CD4 cells (although it is the most efficient). The only requirement is that the CD8 cell is activated by a licensed APC and that it receives three signals which I will not go into. However, without CD4 activation, CD8 cells are unable to develop memory cells and therefore cannot mount a secondary response to infection, hence the reason I consider them to not be “fully activated”. For the purpose of brevity, I am also generalizing regarding certain aspects of B-cell activation. In both cases, I am nonetheless painting a true picture of what is taking place, exceptions notwithstanding.
P a g e 4 | 9
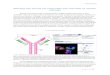
Figure 1. T-cell activation by an Antigen Presenting Cell. This is from the Dr. Rogers class lecture.

Mark Botirius
restricted. That is, CD4 cells can only recognize antigen presented via an MHC class II molecule, and CD8 cells can only recognize antigen presented via MHC class I molecules. MHC class II molecules are presented by APC’s, whereas MHC class I molecules are found in almost all nucleated host cells. What this means, is that MHC class II molecules mediate the cellular response to extracellular pathogens, and MHC class I molecules mediate the response to intracellular pathogens. (Owen, 2013, p. 278) In my scenario, there are no intracellular viral or bacterial pathogens, and so little was said regarding CD8 responses. In fact, the role of CD8 cells surpasses mere response to pathogens. Since all nucleated cells exhibit MHC class I on their cellular membranes, they are not only presenting antigen relevant to pathogens, they are presenting antigen relevant to all of their cellular processes. Therefore, this can be considered to be some kind of “readout” that indicates everything going on inside the cell. If the cell is in some kind of distress, whether infected or not, it will indicate this to the “outside world” via the antigens it is presenting via its MHC class I molecules. Many cells in the beginning stages of becoming cancerous, in fact, will present antigens with their MHC class I molecules indicating that they are on the road to becoming cancerous. These signals are detected by CD8 cells and are destroyed. Of course, not all cells are detected, which leads to full blown cancers.
CD8 cells recognize their targets by recognizing the MHC class I/antigen presented in the cell membrane, and they kill their target in one of two ways, both of which cause the cell to enter apoptosis (which is the reason it has been called the “kiss of death”). (Owen, 2013, p. 432) In both cases, the CD8 cell binds the target cell and releases granules directed towards its target containing ligands that direct the target cell to enter apoptosis. In many cancer cells, the pathway to apoptosis is mutated, which provides one way that cancerous cells can survive efforts by the immune system to eradicate it. (Owen, 2013, p. 644)
In summary, upon infection leukocytes and APC’s recognize pathogens’ PAMPs with their PRRs. They react to those pathogens by phagocytizing and releasing granules that lyse the invaders. They also release cytokines and chemokines that bring more leukocytes to the infection site, cause an inflammatory response, and upregulate the activity of other leukocytes. APC’s such as dendritic cells take in pathogen proteins, process them into “epitopes”, and present them on their cell membranes via MHC class I and MHC class II molecules. APCs then travel to secondary lymph tissues such as lymph nodes, where naïve T-cells and B-cells browse the APCs until they encounter one carrying the antigen their receptor recognizes. When this happens, the APCs activate the B-cells and T-cells via co-stimulatory signals. Activated B-cells and T-cells then begin to proliferate, producing more clones of themselves capable of engaging the particular antigen that binds their receptor. Some of the B-cells become plasma cells and continually excrete antibodies that fight the infection, some form germinal centers and engage in somatic hypermutation, and the remainder become memory B-cells. Regarding T-cells, the population of T-cells differentiates into two main sub types known as “helper” T-cells (CD4) and “cytotoxic” T-cells (CD8). These two types differentiate further into several other types that are beyond the scope of this question. CD4 cells mediate the cellular response by activating other B-cells and T-cells. CD8 cells, once fully activated, travel to the site of infection where they identify infected cells by the MHC class I/antigen the cells are presenting in their membranes. The CD8 cells then bind to the infected cells and release granules that cause the cell to undergo apoptosis. Finally, it should be noted that lymphocytes such as B-cells and T-cells require “survival” signals provided by the presence of antigen. When antigen is no longer present (because the pathogen has been eliminated) the lymphocytes begin to die off, with the exception of
P a g e 5 | 9

Mark Botirius
the memory lymphocytes that are left behind to provide a secondary response should one be needed. This ends the immune response to this particular infection.
Describe the processes by which DNA can be repaired following a mutation.
The six best understood and well known processes by which DNA can be repaired following mutation are direct reversal, mismatch repair, base excision repair, nucleotide excision repair, non-homologous end joining, and homologous recombination. (Rogers, 2017, p. 181) I will therefore describe each in the order that I have listed them.
Direct Reversal
Many DNA repair processes involve a change to the DNA as a result of the repair. Direct reversal, however, is an exception. As its name implies, the process of direct reversal simply undoes the mutation, restoring the DNA to its original state. Two examples of mutations that can be
repaired using the process of direct reversal are photoreactivation and the demethylation of O6 – methylguanine. (Watson, 2014, p. 325). In photoreactivation, the two neighboring pyrimidine rings present in DNA absorb UV light (primarily UV-B) causing the double bonds within the rings to break and forming new covalent bonds between the two neighboring rings. These new bonds form a cyclobutane dimer that creates a lesion in the DNA that will adversely affect replication and translation. The direct repair process for this mutation begins with the enzyme photolyase. Photolyase has two chromophores that also absorb UV light. When photolyase encounters a pyrimidine dimer, it flips the lesion away from the DNA interior and into the catalytic
pocket of the enzyme. It then takes the light energy absorbed by its two chromophores and transfers it to an FADH (an electron carrier) coenzyme. FADH then uses the energy to break the cyclobutane bonds between the neighboring pyrimidines thus restoring the DNA to its original state. (Rajeshwar P Sinha, 2002)
Another example of the direct reversal process is the repair of a methylated guanine. The O6 - methylguanine mutation is often formed by alkylating chemicals such as nitrosamines. These chemicals can cause the keto group at carbon 6 of guanine to become methylated forming O6-methylguanine that causes the guanine to pair with thymine instead of cytosine. (Watson, 2014, p. 322) This mutation is repaired by a methyltransferase. When it encounters O6-methylguanine, it simply transfers the methyl group to itself, reversing the damage and completely repairing the DNA.
Mismatch Repair
P a g e 6 | 9
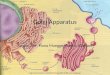
Figure 2. Two neighboring pyrimidine rings absorb a UV-B photon, causing the double bonds between C5 and C6 to break. As a result, a new covalent bonds are formed between the two rings, forming a cyclobutane dimer. (Watson, 2014, p. 322)

Mark Botirius
With a diploid genome size of over 6 billion base pairs, it is understandable that replication errors in humans will occur. In fact, DNA polymerase makes a mistake once every 105 to 106 bases. Fortunately, DNA polymerase has a proofreading ability that can correct many of these mistakes before they are incorporated into the newly synthesized strand. This ability, along with the help of some accessory proteins can lower the rate of error to one in 1010 bases. (Rogers, 2017, p. 180) Those mismatches that escape proofreading are repaired using the mismatch repair process, which begins with a protein called MutS. MutS routinely scans newly replicated DNA looking for mispaired bases. When it detects one (mismatched bases will distort the DNA somewhat, making them detectable by MutS) MutS binds to the lesion and recruits another protein, MutL, which in turn recruits MutH. MutH is an endonuclease that makes a cut in the new strand slightly upstream from the mismatch. MutH differentiates the new strand from the old strand because newly replicated DNA exists in a hemimethylated state. Prior to replication, both strands of the DNA duplex have a characteristic methylation pattern where the adenine residues contained in 5’-GATC-3’ are methylated by Dam methylase. However, immediately after replication, the new strand does not exhibit this methylation pattern, while the old strand does (hence, the strand is “hemimethylated”). It is this difference between the methylation patterns of the two strands that causes MutH to differentiate between them and selectively cut the new strand. This hemimethylated state does not last forever, however, because Dam methylase routinely scans DNA, and when it comes across the unmethylated new strand, it will methylate it, and MutH will be unable to distinguish which strand is the new strand, and which strand is the old one. Therefore, for mismatch repair to work properly, it must detect the error quickly. Once the new strand has been nicked by MutH, it is unwound by the helicase UvrD. Once unwound, the single strand containing the error is degraded by exonucleases, leaving a gap that spans the mismatch. The gap is then reconstructed by DNA polymerase III and reconnected to the rest of the strand with ligase. Ligation finishes the repair. (Watson, 2014, p. 316)
Base Excision Repair and Nucleotide Excision Repair
While mismatch repair deals with replication errors between bases, excision repair pathways such as base excision repair and nucleotide excision repair deal with mutation that damage or alter the bases themselves. An example of this kind of mutation is the deamination of cytosine. When cytosine loses its amino group, it becomes uracil, which occasionally happens spontaneously by hydrolysis. In the case of base excision repair, the damaged base is removed by a glycosylase specific for this particular type of base damage (specifically, uracil glycosylase). There are numerous
kinds of damaged bases, and each is recognized by a particular glycosylase. So far, we have discovered eleven different types. (Watson, 2014, p. 326) When uracil glycosylase discovers the damaged base, it cleaves the glycosidic bond between the damaged base and the ribose of the DNA backbone, thus removing the base but leaving the backbone intact. This creates an apyrimidinic site that is then cut out by an AP endonuclease at the 5’ end
P a g e 7 | 9

Mark Botirius
and an exonuclease at the 3’ end, leaving a gap. The gap is then repaired by DNA polymerase I, thereby completing the repair.
In the case of nucleotide excision repair, the section of DNA containing the damaged base is recognized by how the DNA strand is distorted by the damaged base. The damaged base itself, is not recognized. The distortion is recognized by a protein complex consisting of four protein subunits. Two of the protein subunits are UvrA, and the other two are UvrB. When the protein complex discovers the distortion, the two UvrB subunits bind to the distortion, and the UvrA subunits are realeased. UvrB denatures the DNA and recruits UvrC, which cuts the DNA on both sides of the distortion and the entire section containing the distortion is carried away by UvrD. Dna polymerase I then synthesizes the missing section, and the damage is repaired.
Non Homologous End Joining and Homologous Recombination
The last two repair processes to be discussed, non-homologous end joining (NHEJ) and homologous
recombination deal with the repair of double stranded breaks in DNA. There are several things that can cause DSBs and they include radiation, reactive oxygen species, and chemicals, such as bleomycin. (Iwasa, 2016, p. 534)
In non – homologous end joining, the proteins Ku70 and Ku80 bind to the broken ends of the DNA and recruits the kinase DNA – PKcs which subsequently recruits a protein called Artemis. Artemis and DNA – PKcs form a complex that processes the broken ends by removing a little bit of DNA from each end. This processing then enables a ligase protein complex consisting of the proteins Ligase IV, Cernunnos - XLF, and XRCC4. (Watson, 2014, p. 332) It should be noted that, since this process removes a little bit of DNA from each end, it is mutagenic in that some information is deleted from the genome.
Unlike non – homologous end joining, which uses no information to repair a double stranded break, homologous recombination uses the information from a sister chromosome to fix the break. It does this by first aligning the region containing the break with a nearly identical region from its sister chromosome. Then, a DNA nuclease processes the end of the break to create a region of single stranded DNA. This single strand is then stabilized
by strand exchange proteins which also promote strand invasion. In strand invasion, the single strand invades the homologous strand of the other chromosome, causing the helix to separate in the process forming a “D loop” wherein each strand from one chromosome pairs with the strands of its homologue.
P a g e 8 | 9
Figure 3. Uracil glycosylase cleaves the glycosidic bond between the ribose of the DNA backbone and the uracil base. (Watson, 2014, p. 327)

Mark Botirius
The area where the DNA strands cross is called a “Holliday Junction”. In addition, the resulting hybrid strand formed from the two strands is called a
“heteroduplex” and it can resolve in one of two ways depending on where the DNA is cut. Figure 5 depicts the two different ways to cut the DNA. In the method on the left (the horizontal dashed cut line) the DNA is cut in such a way that the homologues stay with their respective chromosomes. This creates a patch of recombination (depicted in the center of the two DNA strands at the bottom). This is therefore called the “Patch Product”. In the method on the right (the vertical cut line) the homologues exchange chromosomes, creating an actual crossover of homologues. Therefore, this is called a
“Crossover Product”. Upon resolution of the crossover, the repair is complete.
ReferencesIwasa, J. a. (2016). Karp's Cell and Molecular Biology (8th ed.). Hoboken: Wiley.
Madigan, M. S. (2012). Brock Biology of Microorganisms. San Francisco: Pearson Education Inc.
Owen, P. S. (2013). Kuby Immunology. New York: W.H. Freeman and Company.
Rajeshwar P Sinha, D. H. (2002). UV - Induced DNA damage and repair: a review. Photochemical and Photobiological Sciences.
Rogers, S. O. (2017). Integrated Molecular Evolution (2nd ed.). Boca Raton: CRC Press.
Watson, e. a. (2014). Molecular Biology of the Gene (7th ed.). Boston: Pearson.
P a g e 9 | 9
Figure 4. This figure illustrates the mechanism of strand invasion. Sections of single stranded DNA pair with the homologous strand of their sister chromosome. (Rogers, 2017, p. 182)
Figure 5. Depicted in this figure are the two ways that DNA can be cut to separate the duplexes. This figure is from Dr. Rogers’ class lecture