Embed Size (px)
Citation preview

Cellular basis of sex disparities in human cardiac
electrophysiology
A. O. Verkerk,1,2* R. Wilders,2* W. de Geringel2 and H. L. Tan1
1 Department of Clinical and Experimental Cardiology, Academic Medical Center, University of Amsterdam, Amsterdam, The
Netherlands
2 Department of Physiology, Academic Medical Center, University of Amsterdam, Amsterdam, The Netherlands
Received 15 September 2005,
revision requested 14 December
2005,
revision received 28 February
2006,
accepted 21 March 2006
Correspondence: A. Verkerk,
Department of Physiology,
Academic Medical Center, Room
M01-217, Meibergdreef 15, 1105
AZ Amsterdam, The Netherlands.
E-mail: [email protected]
*Both authors contributed equally
to the study.
Abstract
Aim: Sex disparities in electrocardiogram variables and dysrhythmia sus-
ceptibility exist, notably in long QT syndrome (LQTS) and Brugada syn-
drome, but the underlying mechanisms in man are unknown. We studied the
cellular basis of sex distinctions in human cardiac electrophysiology and
dysrhythmia susceptibility using mathematical models of human ventricular
myocytes.
Methods: We implemented sex differences in the Priebe–Beuckelmann and
ten Tusscher–Noble–Noble–Panfilov human ventricular cell models by
modifying densities of the L-type Ca2+ current (ICa,L), transient outward K+
current (Ito), and rapid delayed rectifier K+ current (IKr), according to
experimental data from male and female hearts of various species. Sex dis-
parities in transmural repolarization were studied in transmural strands of
cells with ion current densities based on canine experimental data.
Results: Female cells have longer action potential duration (APD), steeper
APD-heart rate relationship, larger transmural APD heterogeneity, and a
greater susceptibility to pro-dysrhythmogenic early afterdepolarizations
(EADs) than male cells. Conversely, male cells have more prominent phase-1
repolarization and are more susceptible to all-or-none repolarization.
Conclusion: Sex differences in ICa,L, Ito and IKr densities may explain sex
disparities in human cardiac electrophysiology. Female cells exhibit a limited
‘repolarization reserve’ as demonstrated by their larger susceptibility to
EADs, which, combined with their larger transmural electrical heterogeneity,
renders them more vulnerable to tachydysrhythmias in LQTS. Conversely,
male cells have a limited ‘depolarization reserve’, as shown by their larger
susceptibility to all-or-none repolarization, which facilitates tachydysrhyth-
mias in Brugada syndrome. These general principles may also apply to
dysrhythmia susceptibility in common disease.
Keywords action potentials, dysrhythmias, Brugada syndrome, computer
simulation, electrophysiology, ionic currents, long QT syndrome.
Sex differences exist in physiology and pathophysiology
of the cardiovascular system (Mendelsohn & Karas
2005), including human electrocardiogram (ECG) vari-
ables (Pham & Rosen 2002, James et al. 2006). Women
have faster resting heart rates and longer rate-corrected
QT intervals (QTc) than men (Bazett 1920, Merri et al.
1989, Rautaharju et al. 1992) and also manifest a
greater lengthening of the QT interval at longer cycle
lengths, rendering sex-related differences in QT inter-
vals most prominent at slow heart rates (Kligfield et al.
1996, Stramba-Badiale et al. 1997). Moreover, there
are also striking sex disparities in the incidence of
Acta Physiol 2006, 187, 459–477
� 2006 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2006.01586.x 459

cardiac dysrhythmias (James et al. 2006). The most
notable examples are inherited long QT syndrome
(LQTS) and Brugada syndrome, where prominent sex
disparities in clinical expressivity exist, despite equal
genetic transmission (autosomal dominant inheritance).
Thus, women are more likely to sustain ‘Torsade de
Pointes’ (TdP) ventricular tachycardia in LQTS (Zareba
et al. 1995, Locati et al. 1998). Excessive action
potential (AP) prolongation culminating in early after-
depolarizations (EADs) is involved in TdP (Tan et al.
1995). Conversely, women are less susceptible to
dysrhythmias in Brugada syndrome (Matsuo et al.
2001). This syndrome encompasses sudden cardiac
death due to ventricular tachydysrhythmias, and a
distinctive ECG signature (ST-segment elevation in
right precordial leads). These features have been
attributed in part to severe repolarization at the end
of phase-1 of the subepicardial AP, leading to excessive
AP shortening (all-or-none repolarization) (Yan &
Antzelevitch 1999, Tan et al. 2003).
Clinical observations suggest that male sex hormones
are involved in these sex disparities. While QTc is
indistinguishable between boys and girls before the age
of 15, it shortens in men from puberty through
adulthood, but remains unchanged in women
(Rautaharju et al. 1992). Furthermore, the sex differ-
ence in QTc in adults was abolished by castration of
men, and restored by subsequent application of testos-
terone (Bidoggia et al. 2000). Similar developmental
components exist in the dysrhythmias of LQTS and
Brugada syndrome. While boys and girls before age 15
have similar incidences of TdP in LQTS, men have a
reduced incidence of TdP from puberty through adult-
hood (Zareba et al. 1995). Conversely, the incidence of
dysrhythmias in Brugada syndrome is higher in men
than in women aged 20–60 (Matsuo et al. 2001).
Interestingly, castration of men abolished the Brugada
syndrome ECG pattern (Matsuo et al. 2003).
The cellular basis of these sex-related distinctions in
human cardiac electrophysiology is unresolved. Animal
studies demonstrate the importance of sex-related
differences in membrane current densities in sex dis-
tinctions in cardiac electrophysiology (Pham & Rosen
2002). We propose that sex disparities in sarcolemmal
ion current densities may also account for the sex
differences in ECG variables and dysrhythmia suscept-
ibility in humans. However, to date, human data on sex
differences in membrane current densities of undiseased
hearts are not available (Verkerk et al. 2005), in large
part because of limited availability of human cardio-
myocytes for experimental research (ten Tusscher et al.
2006). We studied the cellular basis of sex distinctions
in human cardiac electrophysiology and dysrhythmia
susceptibility using mathematical models of human
ventricular myocytes in which we modified ion current
densities based on experimental data obtained from
male and female hearts of various species. Thus, we
studied AP characteristics, transmural electrophysiolog-
ical heterogeneity, and susceptibility to EADs (LQTS)
and all-or-none repolarization (Brugada syndrome).
Materials and methods
Human ventricular cell models
We studied the cellular basis of sex distinctions in
human cardiac electrophysiology and dysrhythmia sus-
ceptibility using mathematical models of human ven-
tricular myocytes. To date, three distinct human
ventricular cell models exist: the Priebe–Beuckelmann
(PB) model (Priebe & Beuckelmann 1998), the
ten Tusscher–Noble–Noble–Panfilov (TNNP) model
(ten Tusscher et al. 2004), and the Iyer–Mazhari–
Winslow model (Iyer et al. 2004). The large number
of variables (67) and the small time step required for
integration make the Iyer–Mazhari–Winslow model
computationally extremely demanding, with simula-
tions running about 900 times slower than either the PB
or TNNP model (ten Tusscher et al. 2006). Therefore,
we refrained from using this model in our study.
The PB model and the TNNP model provide the basis
for the simulations in this study. The PB model was the
first model for human ventricular myocytes. It is largely
based on the Luo–Rudy phase II model for guinea pig
ventricular cells (Luo & Rudy 1994) in which formula-
tions for the major ionic currents were adjusted to the
scarce data available for human ventricular cells at that
time (Bernus et al. 2002; ten Tusscher et al. 2006). The
TNNP model is a recently developed human ventricular
cell model, which uses new formulations for all major
ionic currents based on a now much wider basis of
experimental data, largely from human ventricular cell
experiments but also from ion channel expression
experiments (for overview about human ventricular cell
models, see ten Tusscher et al. 2006). Consequent to the
differences in membrane current formulations, AP
shape and duration, AP restitution, and relative import-
ance of ionic currents in the AP generation differs
between the PB and TNNP model (ten Tusscher et al.
2006). To study sex distinctions in human cardiac
electrophysiology rather than model-based effects, we
used both the PB and TNNP model.
The PB model and the TNNP model are both
importantly based on experimental data from single
ventricular myocytes isolated from explanted human
hearts. As these myocytes were generally isolated from
midmyocardial areas of the left ventricle (LV) of male
patients (Priebe & Beuckelmann 1998), the PB model
and the midmyocardial variant of the TNNP model
(ten Tusscher et al. 2004) are, in effect, models of male
460 � 2006 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2006.01586.x
Sex and human cardiac electrophysiology Æ A O Verkerk et al. Acta Physiol 2006, 187, 459–477

midmyocardial ventricular myocytes. To study sex
disparities in human cardiac electrophysiology and the
susceptibility to dysrhythmias, we incorporated experi-
mentally observed disparities in ion current densities
between sexes and myocardial layers of various species
into these models. These disparities are discussed below
and summarized in Table 1 as conductances relative to
male midmyocardial myocytes. In the midmyocardial
TNNP model, we replaced the relatively simple intra-
cellular calcium handling by the more sophisticated
calcium dynamics of the updated Luo–Rudy model
(Faber & Rudy 2000) to increase its stability, especially
in the right ventricular (RV) models.
Sex disparities in ion current densities
Sex distinctions in human cardiac electrophysiology and
dysrhythmia susceptibility were studied using the PB and
TNNP human ventricular myocyte models in which we
modified ion current densities based on experimental
data obtained from male and female hearts of various
species. Therefore, we reviewed all studies into sex
disparities in ion current densities, conducted in single
ventricular myocytes (Table 1). These data were pub-
lished for dog, rabbit, guinea pig and mouse hearts, but
not for human hearts. L-type Ca2+ current (ICa,L) in
female subepicardium and midmyocardium is 1.32 times
that of males (Pham et al. 2002). Transient outward K+
current (Ito) in females is 0.75 times that of males in all cell
layers (Di Diego et al. 2002, Wu & Anderson 2002),
although this is not a consistent finding (Leblanc et al.
1998, Trepanier-Boulay et al. 2001). The rapid compo-
nent of the delayed rectifier K+ current (IKr) in females is
0.83 times that of males (Liu et al. 1998).
Although the ultrarapid component of the delayed
rectifier K+ current (IKur) and the ATP-regulated K+
current (IK,ATP) also exhibit sex disparities (Ranki et al.
2001, Trepanier-Boulay et al. 2001), these disparities
were not incorporated, because IKur and IK,ATP are not
present or functional under normal conditions in human
ventricular myocytes (Li et al. 1996). Similarly, we did
not modify Na+ current (INa), slow component of the
delayed rectifier K+ current (IKs), inward rectifier K+
current (IK1) and Na+-Ca2+ exchange current (INaCa),
because they are equal in male and female hearts
(Leblanc et al. 1998, Liu et al. 1998, Trepanier-Boulay
et al. 2001, Li et al. 2002) [equal INaCa was inferred
from equal baseline intracellular Na+ and Ca2+ concen-
trations (Ranki et al. 2001, Sugishita et al. 2001, Chen
et al. 2003)].
Transmural heterogeneity in ion current densities
Ventricular dysrhythmias in LQTS (Akar et al. 2002)
and Brugada syndrome (Yan & Antzelevitch 1999,
Tan et al. 2003) are in large part due to electrophys-
iological heterogeneities across the ventricular wall. To
study sex disparities in the susceptibility to these
dysrhythmias, we incorporated the differences in ion
current densities between subepicardium, midmyocar-
dium and subendocardium, as summarized in Table 1,
according to data from isolated canine ventricular
myocytes. These data were used, because data from
human are limited. ICa,L in subepicardium and mid-
myocardium is 1.32 times that in subendocardium in
females, but not in males (Pham et al. 2002). In both
sexes, Ito in subepicardium and midmyocardium is
twice that in subendocardium (Liu et al. 1993, Li
et al. 2002), while IKs in subepicardium and subendo-
cardium is 1.42 times that in midmyocardium
(Li et al. 2002). INaCa in subendocardium is 0.69
times that in the other layers (Zygmunt et al. 2000).
Although a late component of INa is 27% smaller in
subepicardium and subendocardium than in midmyo-
cardium (Zygmunt et al. 2001), this transmural het-
erogeneity was not incorporated, because such a
component is not present in the PB and TNNP
models. Similarly, no transmural heterogeneities in
INa, IKr, and IK1 were reported (Liu et al. 1993,
Zygmunt et al. 2001, Li et al. 2002).
Table 1 Membrane current magnitude of male and female myocytes
Male Female
Epicardial Midmyocardial Endocardial Epicardial Midmyocardial Endocardial
INa 1 1 1 1 1 1
ICa,L 1 1 1 1.32 1.32 1
Ito 1 1 0.5 0.75 0.75 0.375
IKr 1 1 1 0.83 0.83 0.83
IKs 1.42 1 1.42 1.42 1 1.42
IK1 1 1 1 1 1 1
INaCa 1 1 0.69 1 1 0.69
Values are current density relative to male midmyocardial myocytes.
� 2006 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2006.01586.x 461
Acta Physiol 2006, 187, 459–477 A O Verkerk et al. Æ Sex and human cardiac electrophysiology

Simulation protocols
In our single cell simulations, APs were elicited with a
2-ms, 3-nA (c. 20% suprathreshold) stimulus pulse.
Transmural conduction was studied using a heteroge-
neous linear strand of 600 transversally coupled ven-
tricular cells as diagrammed in Figure 1. This strand
represents a LV wall thickness of 1.2 cm [normal range
0.9–1.4 cm (Feigenbaum 1994)], with the transverse
orientation of cells based on data from human and
canine LV (Streeter & Hanna 1973, Greenbaum et al.
1981). The strand was stimulated from the endocardial
side by injecting a c. 20% suprathreshold stimulus of
2 ms duration into the first of 300 cells of the
endocardial type. The next 240 cells of the strand were
M type cells and the final 60 cells were epicardial type
cells. This distribution of 50% endocardial type cells,
40% M type cells and 10% epicardial type cells was
based on data from human LV (Drouin et al. 1995).
The intercellular coupling conductance (Gc) was set to
5 or 10 lS, which is well within the estimated range of
3–12 lS for the gap junctional conductance between
human ventricular myocytes (Jongsma & Wilders
2000). The cytoplasmic resistivity was set to 150 Xcm.
Models were coded using Compaq Visual Fortran
6.6C (Hewlett-Packard, Palo Alto, CA, USA) and run
on a 3-GHz Intel Pentium-4 processor workstation as a
32-bit Windows application, applying a simple and
efficient Euler-type integration scheme with a 5-ls time
step. The spatial discretization step in the strand
simulations was 20 lm, i.e. the width of a single cell,
which was treated as isopotential. Stimulus and end
effects, which were restricted to <10 cells, were
minimized by discarding data obtained from cells
1–10 and 591–600 when assessing dispersion in
repolarization time (RT). All figures show steady-state
AP characteristics, obtained at 1–7 min after onset of
stimulation.
Results
We used our ‘male’ and ‘female’ models of subepicardial,
midmyocardial, and subendocardial myocytes to assess
sex disparities in the action potential duration (APD)-
heart rate relationship, transmural APD dispersion and
susceptibility to EADs and all-or-none repolarization.
Sex differences in AP duration
In both PB and TNNP based models, simulations at
different stimulation frequencies revealed significant sex
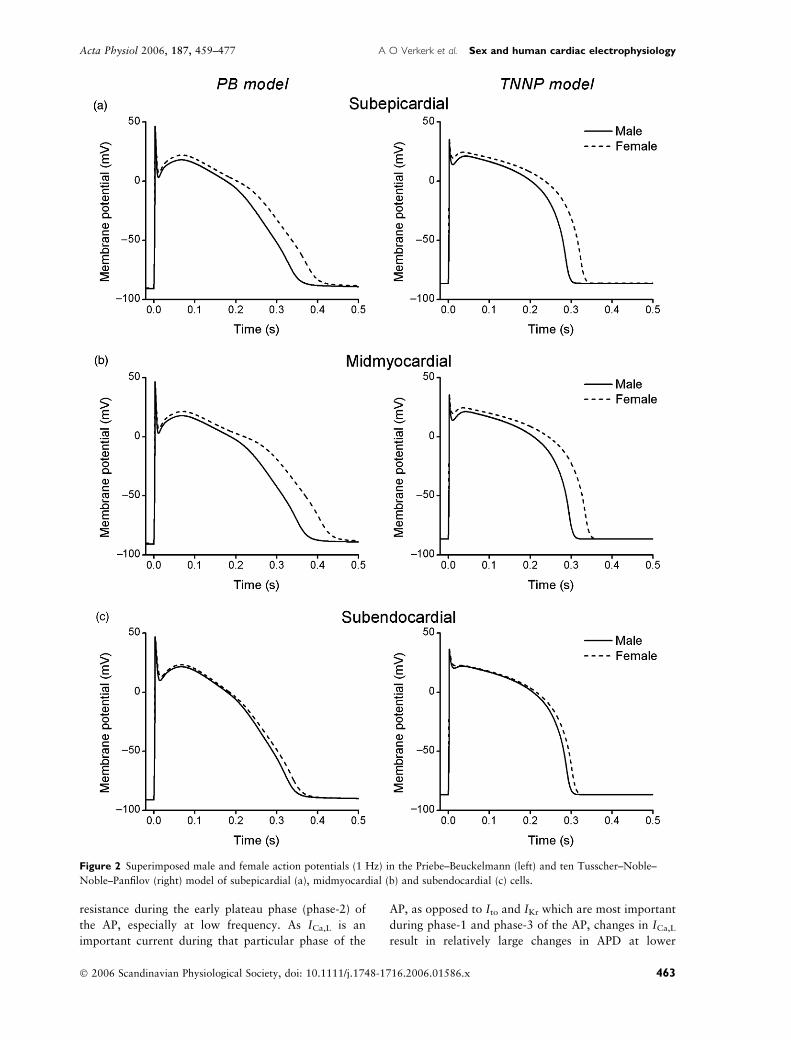
differences in APD. Figure 2 shows superimposed APs
at 1 Hz in the female and male PB (left) and TNNP
(right) cells of subepicardial, midmyocardial, and sub-
endocardial origin. In all cell types, the APs were
significantly longer in females than males. Figure 3
summarizes the frequency dependence of APD at 90%
repolarization (APD90) of the male and female PB (left)
and TNNP (right) cells. At all frequencies, APD90 of
subepicardial, midmyocardial and subendocardial cells
was longer in females than in males. Moreover, APD90
decreased with increasing stimulus frequency in all cell
types. Linear regression analysis indicated that APD-
heart rate relationship was steeper in females, except in
subendocardial cells.
Ionic basis of sex differences in response to rate
The combined effect of the differences in ionic current
densities between male and female cells (Table 1) is a
longer AP and a steeper APD-heart rate relationship in
female subepicardial and midmyocardial but not sub-
endocardial cells (Fig. 3). To investigate the contribu-
tion of the individual ion currents involved, we carried
out simulations in which we selectively increased ICa,L
density, decreased IKr density, or decreased Ito density.
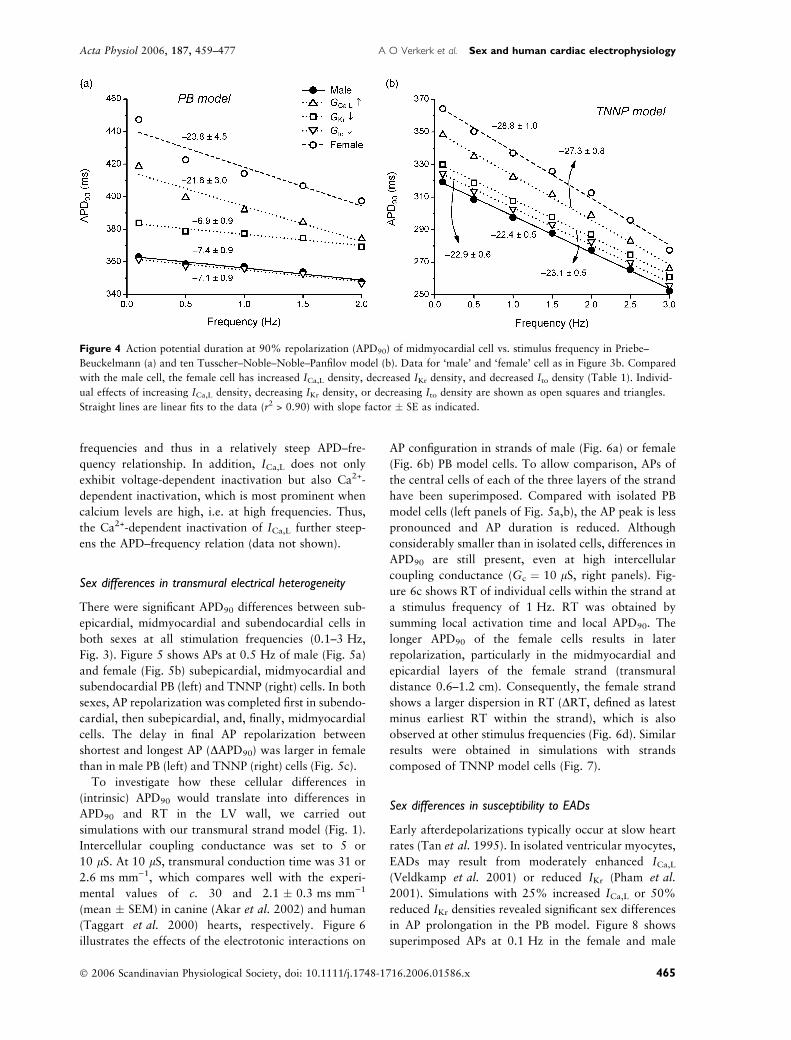
As illustrated in Figure 4 for midmyocardial cells, the
APD was importantly increased by both an increased
ICa,L density and a decreased IKr density, but not by a
decrease in Ito density. Also illustrated in Figure 4, the
steepness of the APD–frequency relationship is largely
determined by ICa,L. Changes in either IKr or Ito density
had only little effect on the steepness of this relation-
ship. This readily explains the similar slopes in male and
female cells of endocardial origin, as these cells have
identical ICa,L density (Fig. 3). The larger sensitivity to
changes in ICa,L can be explained by the high membrane
Figure 1 Diagram of linear strand model. The transmural
strand is composed of 600 human left ventricular myocytes,
which are arranged transversally and coupled by an intercel-
lular coupling conductance Gc. Cytoplasmic resistivity is set to
150 Xcm. Action potential propagation is initiated by applying
an external stimulus at the endocardial side of the strand. See
Materials and methods for details.
462 � 2006 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2006.01586.x
Sex and human cardiac electrophysiology Æ A O Verkerk et al. Acta Physiol 2006, 187, 459–477
resistance during the early plateau phase (phase-2) of
the AP, especially at low frequency. As ICa,L is an
important current during that particular phase of the
AP, as opposed to Ito and IKr which are most important
during phase-1 and phase-3 of the AP, changes in ICa,L
result in relatively large changes in APD at lower
Figure 2 Superimposed male and female action potentials (1 Hz) in the Priebe–Beuckelmann (left) and ten Tusscher–Noble–
Noble–Panfilov (right) model of subepicardial (a), midmyocardial (b) and subendocardial (c) cells.
� 2006 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2006.01586.x 463
Acta Physiol 2006, 187, 459–477 A O Verkerk et al. Æ Sex and human cardiac electrophysiology
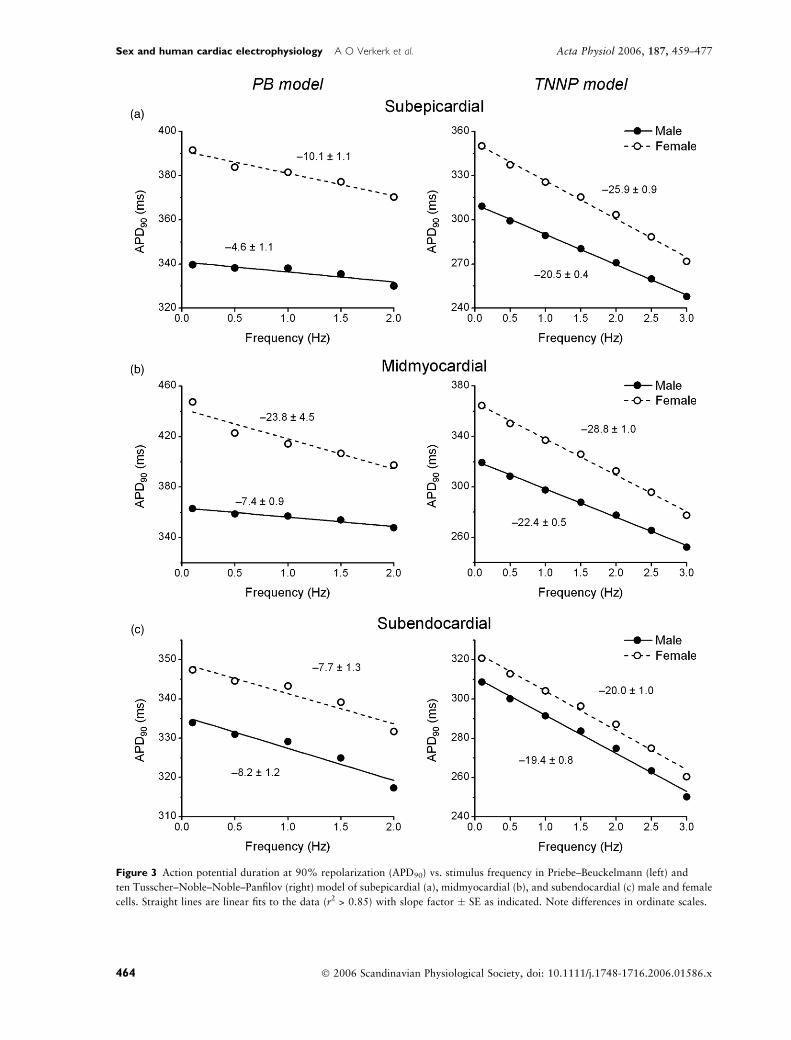
Figure 3 Action potential duration at 90% repolarization (APD90) vs. stimulus frequency in Priebe–Beuckelmann (left) and
ten Tusscher–Noble–Noble–Panfilov (right) model of subepicardial (a), midmyocardial (b), and subendocardial (c) male and female
cells. Straight lines are linear fits to the data (r2 > 0.85) with slope factor � SE as indicated. Note differences in ordinate scales.
464 � 2006 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2006.01586.x
Sex and human cardiac electrophysiology Æ A O Verkerk et al. Acta Physiol 2006, 187, 459–477
frequencies and thus in a relatively steep APD–fre-
quency relationship. In addition, ICa,L does not only
exhibit voltage-dependent inactivation but also Ca2+-
dependent inactivation, which is most prominent when
calcium levels are high, i.e. at high frequencies. Thus,
the Ca2+-dependent inactivation of ICa,L further steep-
ens the APD–frequency relation (data not shown).
Sex differences in transmural electrical heterogeneity
There were significant APD90 differences between sub-
epicardial, midmyocardial and subendocardial cells in
both sexes at all stimulation frequencies (0.1–3 Hz,
Fig. 3). Figure 5 shows APs at 0.5 Hz of male (Fig. 5a)
and female (Fig. 5b) subepicardial, midmyocardial and
subendocardial PB (left) and TNNP (right) cells. In both
sexes, AP repolarization was completed first in subendo-
cardial, then subepicardial, and, finally, midmyocardial
cells. The delay in final AP repolarization between
shortest and longest AP (DAPD90) was larger in female
than in male PB (left) and TNNP (right) cells (Fig. 5c).
To investigate how these cellular differences in
(intrinsic) APD90 would translate into differences in
APD90 and RT in the LV wall, we carried out
simulations with our transmural strand model (Fig. 1).
Intercellular coupling conductance was set to 5 or
10 lS. At 10 lS, transmural conduction time was 31 or
2.6 ms mm)1, which compares well with the experi-
mental values of c. 30 and 2.1 � 0.3 ms mm)1
(mean � SEM) in canine (Akar et al. 2002) and human
(Taggart et al. 2000) hearts, respectively. Figure 6
illustrates the effects of the electrotonic interactions on
AP configuration in strands of male (Fig. 6a) or female
(Fig. 6b) PB model cells. To allow comparison, APs of
the central cells of each of the three layers of the strand
have been superimposed. Compared with isolated PB
model cells (left panels of Fig. 5a,b), the AP peak is less
pronounced and AP duration is reduced. Although
considerably smaller than in isolated cells, differences in
APD90 are still present, even at high intercellular
coupling conductance (Gc ¼ 10 lS, right panels). Fig-
ure 6c shows RT of individual cells within the strand at
a stimulus frequency of 1 Hz. RT was obtained by
summing local activation time and local APD90. The
longer APD90 of the female cells results in later
repolarization, particularly in the midmyocardial and
epicardial layers of the female strand (transmural
distance 0.6–1.2 cm). Consequently, the female strand
shows a larger dispersion in RT (DRT, defined as latest
minus earliest RT within the strand), which is also
observed at other stimulus frequencies (Fig. 6d). Similar
results were obtained in simulations with strands
composed of TNNP model cells (Fig. 7).
Sex differences in susceptibility to EADs
Early afterdepolarizations typically occur at slow heart
rates (Tan et al. 1995). In isolated ventricular myocytes,
EADs may result from moderately enhanced ICa,L
(Veldkamp et al. 2001) or reduced IKr (Pham et al.
2001). Simulations with 25% increased ICa,L or 50%
reduced IKr densities revealed significant sex differences
in AP prolongation in the PB model. Figure 8 shows
superimposed APs at 0.1 Hz in the female and male
Figure 4 Action potential duration at 90% repolarization (APD90) of midmyocardial cell vs. stimulus frequency in Priebe–
Beuckelmann (a) and ten Tusscher–Noble–Noble–Panfilov model (b). Data for ‘male’ and ‘female’ cell as in Figure 3b. Compared
with the male cell, the female cell has increased ICa,L density, decreased IKr density, and decreased Ito density (Table 1). Individ-
ual effects of increasing ICa,L density, decreasing IKr density, or decreasing Ito density are shown as open squares and triangles.
Straight lines are linear fits to the data (r2 > 0.90) with slope factor � SE as indicated.
� 2006 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2006.01586.x 465
Acta Physiol 2006, 187, 459–477 A O Verkerk et al. Æ Sex and human cardiac electrophysiology
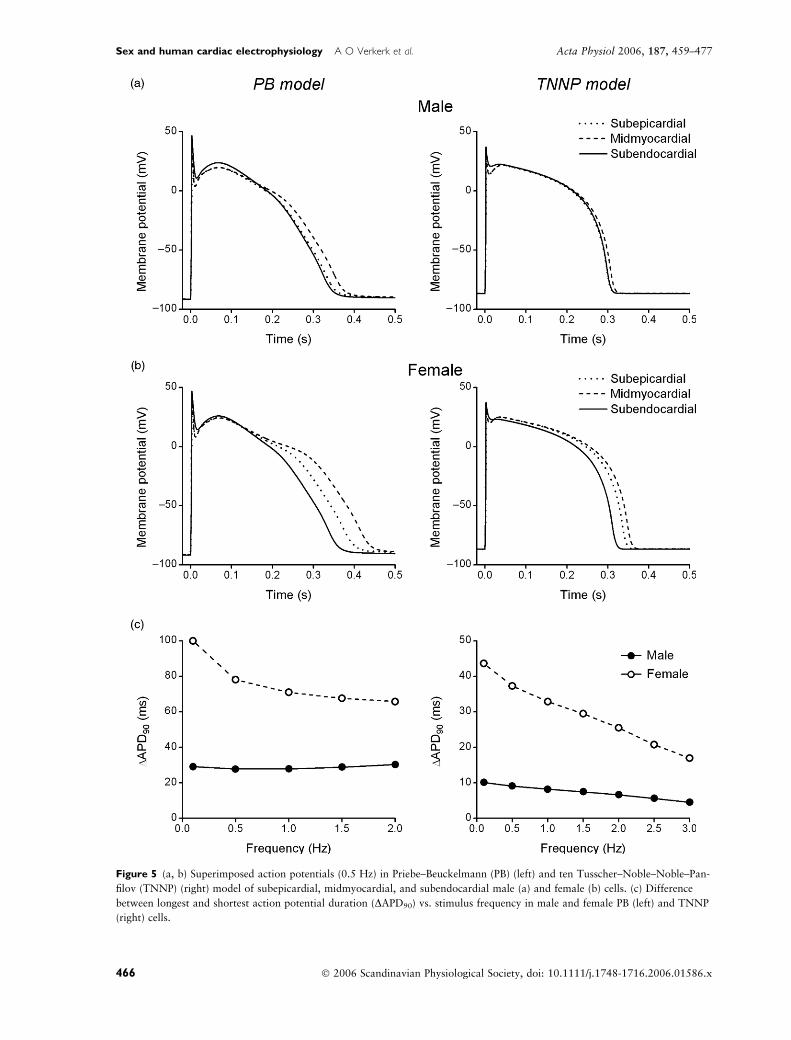
Figure 5 (a, b) Superimposed action potentials (0.5 Hz) in Priebe–Beuckelmann (PB) (left) and ten Tusscher–Noble–Noble–Pan-
filov (TNNP) (right) model of subepicardial, midmyocardial, and subendocardial male (a) and female (b) cells. (c) Difference
between longest and shortest action potential duration (DAPD90) vs. stimulus frequency in male and female PB (left) and TNNP
(right) cells.
466 � 2006 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2006.01586.x
Sex and human cardiac electrophysiology Æ A O Verkerk et al. Acta Physiol 2006, 187, 459–477
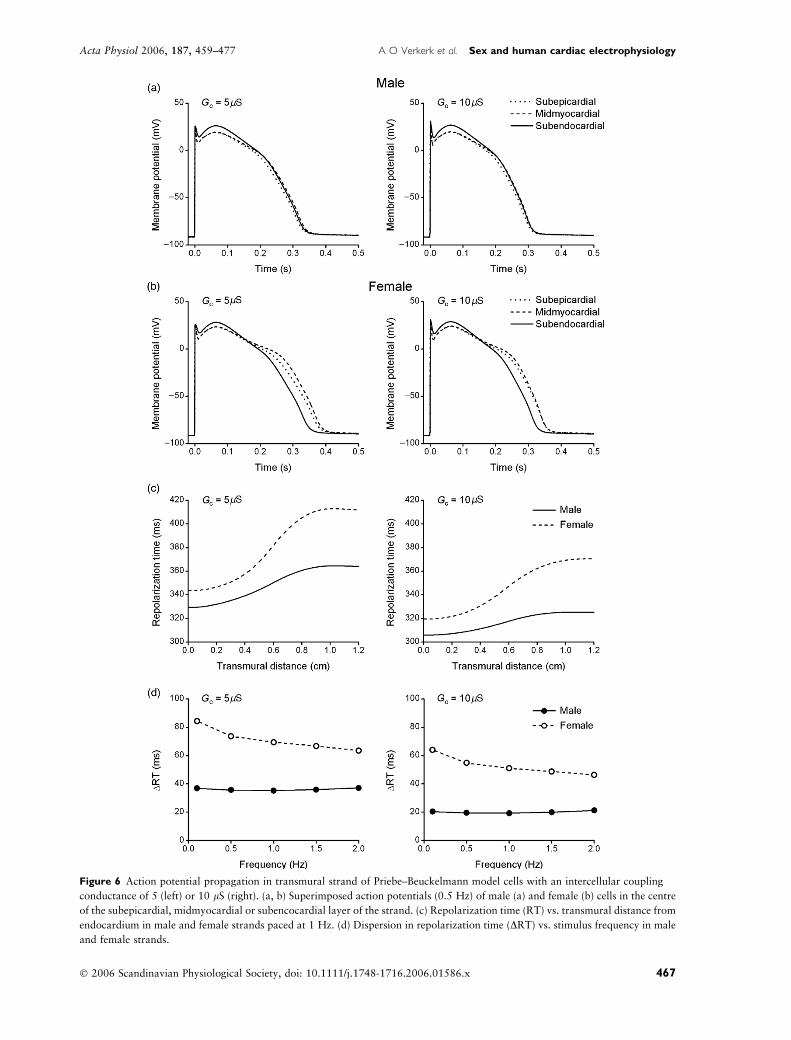
Figure 6 Action potential propagation in transmural strand of Priebe–Beuckelmann model cells with an intercellular coupling
conductance of 5 (left) or 10 lS (right). (a, b) Superimposed action potentials (0.5 Hz) of male (a) and female (b) cells in the centre
of the subepicardial, midmyocardial or subencocardial layer of the strand. (c) Repolarization time (RT) vs. transmural distance from
endocardium in male and female strands paced at 1 Hz. (d) Dispersion in repolarization time (DRT) vs. stimulus frequency in male
and female strands.
� 2006 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2006.01586.x 467
Acta Physiol 2006, 187, 459–477 A O Verkerk et al. Æ Sex and human cardiac electrophysiology

midmyocardial PB (left) and TNNP (right) cell resulting
from increased ICa,L (Fig. 8a) or reduced IKr (Fig. 8b).
In the PB model, the excessive prolongation in females
culminated in subthreshold ‘humps’ during phase-3
repolarization (arrows), which are markers of suscep-
tibility to the formation of EADs (Cranefield 1977,
Binah & Rosen 1992). The TNNP model did not
exhibit such humps, most likely due to its particular
ICa,L inactivation kinetics that effectively prevent
reactivation of ICa,L (see Discussion). Multicellular
simulations at physiological pacing rate (1 Hz) and
intercellular coupling conductance (10 lS) demonstra-
ted an increased sex disparity in dispersion of RT with
enhanced ICa,L (Fig. 8c), both with PB (left) and TNNP
(right) model cells. In the midmyocardial and epicardial
layers of transmural strands of female, but not male, PB
model cells EADs occurred with enhanced ICa,L (Fig. 8c,
left inset, arrows).
Figure 9 illustrates the mechanism underlying the
subthreshold humps during phase-3 repolarization
observed in isolated female PB model cells (Fig. 8a) as
well as the EADs observed in strands of female PB cells
(Fig. 8c, inset). Although Ca2+-induced inactivation of
ICa,L is more prominent in the female cell because of the
larger intracellular free calcium concentration ([Ca2+ ]i)
in the female cell (Fig. 9b), the current reactivates
(Fig. 9c) and thus produces the subthreshold humps in
the single cell (Fig. 9a) and the EADs in the strand
(Fig. 9a, inset). In the male cell (Fig. 9, left), there is also
AP prolongation, but this prolongation is not large
enough to result in subthreshold humps in the single cell
(Fig. 9a) or EADs in the strand (Fig. 9a, inset). The
increased calcium release of the female cell not only
contributes to the AP shape by modulating ICa,L, but
also by increasing the sodium–calcium exchange current
(INaCa), which results in an additional inward current
during the AP plateau phase, in particular at higher
frequencies as in the strand simulations (Fig. 9d, insets).
Additional simulations in which we selectively increased
ICa,L density, decreased IKr density, or decreased Itodensity demonstrate that the smaller female IKr facili-
tates AP prolongation, but is not crucial to the prefer-
ential occurrence of subthreshold humps and EADs in
the female cell, and that Ito only plays a minor role (data
not shown).
Sex differences in susceptibility to all-or-none
repolarization
The basis of Brugada syndrome resides in the RV
(Tukkie et al. 2004). One proposed cellular mechanism
revolves around accentuation of the AP notch (end of
phase-1) and loss of the AP dome (all-or-none repolar-
ization) in RV subepicardial myocytes with concomitant
preservation of the AP dome in subendocardium. The
resulting transmural voltage gradient causes ECG ST
elevations and reentrant tachydysrhythmias (Yan &
Antzelevitch 1999). These changes may become man-
ifest when net depolarizing current is reduced [Na+
channel mutations (Tan et al. 2003)] or net repolarizing
current is enhanced during the early phases of the AP.
Repolarizing forces in RV are larger than in LV,
because IKs density is two times larger, and Ito density
is 2.5 to four times larger in RV (Volders et al. 1999,
Di Diego et al. 2002). Accordingly, we implemented
RV models of both sexes by amplifying IKs and Itodensities by factors 2 and 4, respectively. Our simula-
tions revealed significant differences in AP configura-
tions between RV and LV. Figure 10a shows
superimposed APs at 0.5 Hz in male (left) and female
(right) TNNP models of subepicardial and subendo-
cardial RV and LV myocytes. AP duration was shorter
and phase-1 repolarization was more pronounced in
RV than LV subepicardium. The AP notch clearly was
more prominent in males than females in subepicardial
RV.
An experimental study demonstrated that more
prominent phase-1 repolarization may result in all-or-
none repolarization, when an IK,ATP agonist is
administered (Di Diego et al. 2002). Accordingly, we
incorporated IK,ATP into our models to study suscepti-
bility to all-or-none repolarization. Figure 10b shows
superimposed APs from male and female TNNP cells
before and after introduction of IK,ATP. Introducing
IK,ATP (ohmic conductance: 0.8 nS) hardly affected the
AP dome in LV of both sexes (not shown). However, in
the male RV subepicardial cell (left), IK,ATP caused all-
or-none repolarization, while it evoked only a slight
depression of the AP plateau in females (right). Because
experimental studies have reported that IK,ATP density is
2.5 times larger in females than males (Ranki et al.
2001), we repeated these simulations with increased
IK,ATP (2.0 nS) in females. These simulations still
resulted in a less prominent reduction of the dome
and abbreviation of the AP than simulations with the
small IK,ATP in males. Introduction of IK,ATP into RV
subendocardial cells (Fig. 10c) hardly affected the AP
dome in males (left) and females (right). Similar results
were obtained with the PB model (not shown). To
investigate the contribution of the individual ion
currents involved in the sex disparity in susceptibility
to all-or-none repolarization, we carried out simulations
in which we selectively increased ICa,L density,
decreased IKr density, or decreased Ito density. We
found that the occurrence of all-or-none repolarization
was critically dependent on the high Ito density in the
male subepicardial cell, and facilitated by the lower
ICa,L density and higher IKr density in the male, which
both enhance the repolarizing forces of the AP (data not
shown).
468 � 2006 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2006.01586.x
Sex and human cardiac electrophysiology Æ A O Verkerk et al. Acta Physiol 2006, 187, 459–477
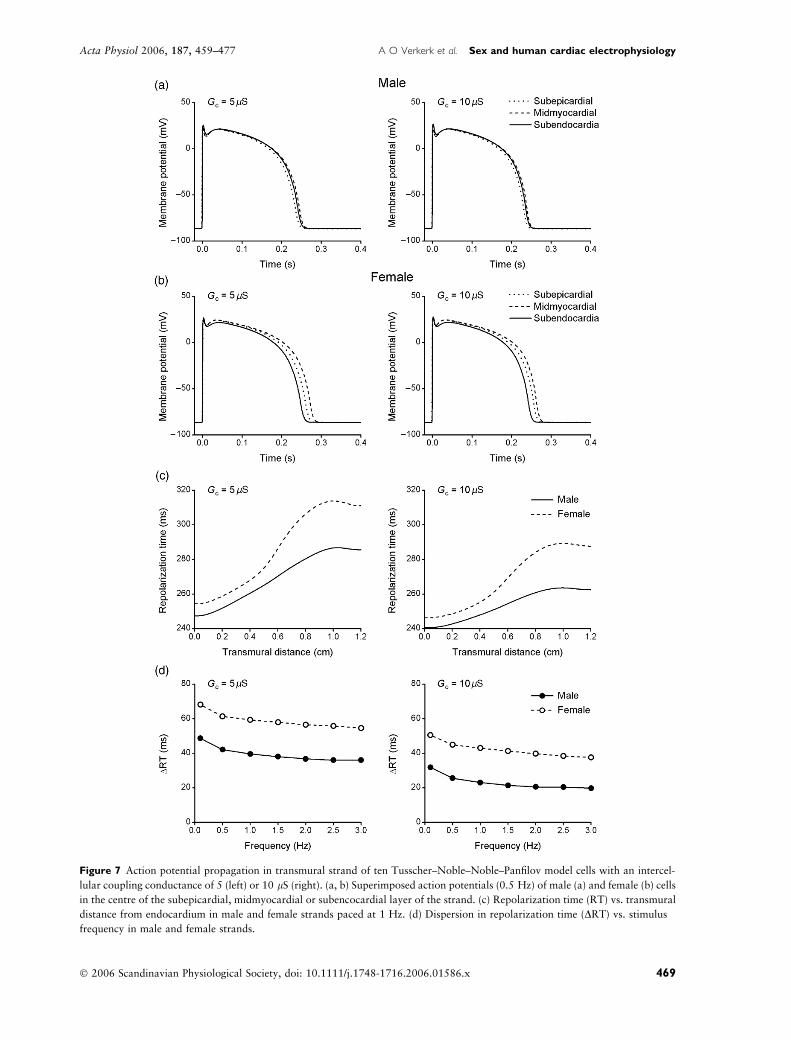
Figure 7 Action potential propagation in transmural strand of ten Tusscher–Noble–Noble–Panfilov model cells with an intercel-
lular coupling conductance of 5 (left) or 10 lS (right). (a, b) Superimposed action potentials (0.5 Hz) of male (a) and female (b) cells
in the centre of the subepicardial, midmyocardial or subencocardial layer of the strand. (c) Repolarization time (RT) vs. transmural
distance from endocardium in male and female strands paced at 1 Hz. (d) Dispersion in repolarization time (DRT) vs. stimulus
frequency in male and female strands.
� 2006 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2006.01586.x 469
Acta Physiol 2006, 187, 459–477 A O Verkerk et al. Æ Sex and human cardiac electrophysiology
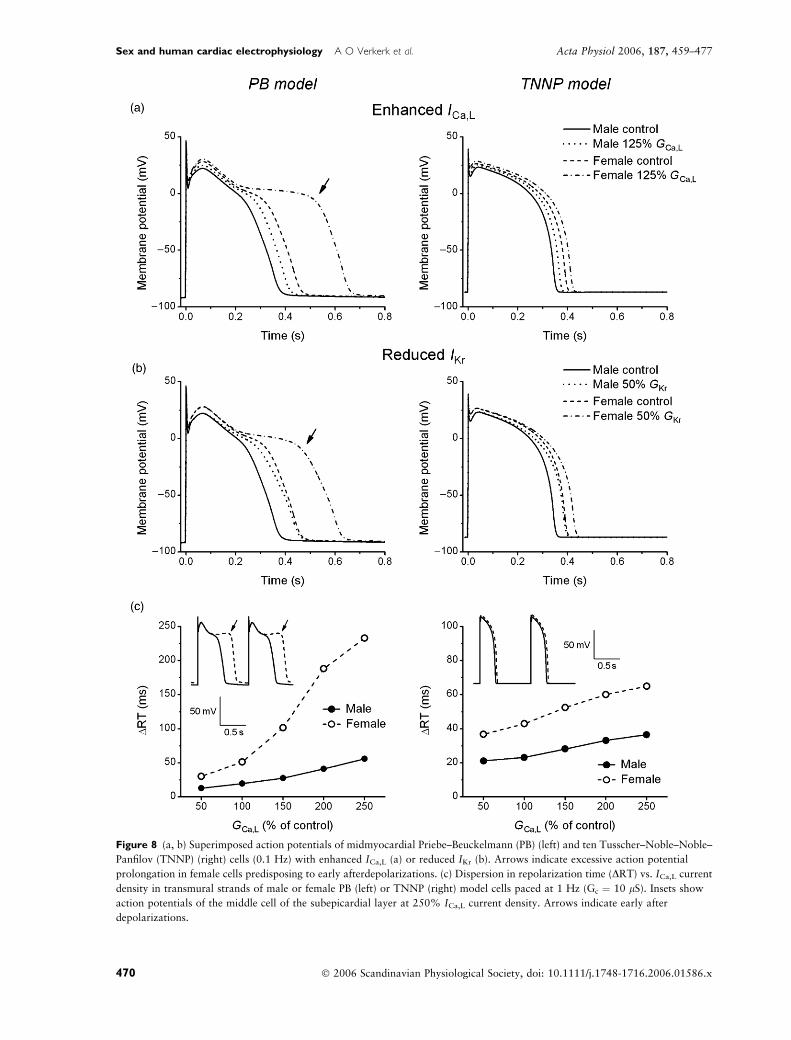
Figure 8 (a, b) Superimposed action potentials of midmyocardial Priebe–Beuckelmann (PB) (left) and ten Tusscher–Noble–Noble–
Panfilov (TNNP) (right) cells (0.1 Hz) with enhanced ICa,L (a) or reduced IKr (b). Arrows indicate excessive action potential
prolongation in female cells predisposing to early afterdepolarizations. (c) Dispersion in repolarization time (DRT) vs. ICa,L current
density in transmural strands of male or female PB (left) or TNNP (right) model cells paced at 1 Hz (Gc ¼ 10 lS). Insets show
action potentials of the middle cell of the subepicardial layer at 250% ICa,L current density. Arrows indicate early after
depolarizations.
470 � 2006 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2006.01586.x
Sex and human cardiac electrophysiology Æ A O Verkerk et al. Acta Physiol 2006, 187, 459–477

To test whether preferential all-or-none repolariza-
tion of subepicardial cells in response to enhanced net
repolarizing current was still possible in case of
electrotonic interactions occurring in the well-coupled
myocardium, we also carried out simulations with
strands composed of RV male or female cells. For these
simulations, we reduced the number of cells in the
transmural strand from 600, as used for LV, to 210 (70
in each layer), based on the RV wall thickness of
4 � 1 mm observed in the human heart (Gottdiener
et al. 1985). With the IK,ATP conductance increased to
4 nS, we found preferential loss of the AP dome in the
subepicardium of the male strand (Fig. 10d).
Discussion
Our studies provide insights into the cellular basis of sex
disparities in cardiac electrophysiology and dysrhyth-
mia susceptibility. Of note, there was general consis-
tency between the PB and TNNP human ventricular cell
models.
Sex differences in AP duration
We found that female LV cells had longer intrinsic AP
duration than their male counterparts (Figs 2 and 3),
resulting in later RT (Figs 6 and 7). Although experi-
mental findings of sex disparity in AP duration of
undiseased human hearts are not available, our results
are in agreement with experimental findings in animals
and diseased human myocardium. In mouse subepicar-
dial myocytes (Trepanier-Boulay et al. 2001, Wu &
Anderson 2002), rabbit LV subendocardial myocytes
(Valverde et al. 2003), and human failing LV myocytes
(Verkerk et al. 2005), APs were significantly longer in
females than in males. Our results are also in agreement
with the clinical observations that women have longer
QTc intervals (Bazett 1920, Merri et al. 1989,
Rautaharju et al. 1992). The longer APD of the female
cell is importantly due to its larger ICa,L density and
smaller IKr density (Fig. 4). Decreased Ito density hardly
affected the APD, consistent with the findings of several
studies using mathematical modelling (Priebe & Beuck-
elmann 1998, Winslow et al. 1999; Gima & Rudy
2002; ten Tusscher et al. 2004), and a ‘dynamic clamp’
study in which a computed Ito was introduced in real
myocytes (Sun & Wang 2005).
The female LV cells had steeper APD–frequency
relationships than males (Fig. 3), largely due to their
larger ICa,L (Fig. 4). Although not studied in detail,
experimental findings in animals provide supportive
results. In rabbits, APD at 20%, 50% and 90%
repolarization is similar in males and females at a cycle
length of 300 ms, but different at longer cycle lengths
(Valverde et al. 2003). Again, these results are in
keeping with clinical observations: a steeper QT–heart
rate relationship in women than men (Kligfield et al.
1996, Stramba-Badiale et al. 1997).
Sex differences in TdP
We found that female cells exhibit greater differences in
intrinsic APD90 between subendocardium, midmyocar-
dium and subepicardium (Fig. 5), resulting in greater
transmural dispersion of repolarization (Figs 6d and 7d).
Moreover, we found that increasing ICa,L or reducing IKr
prolonged APs significantly more in females than males
(Fig. 8a,b). Increasing ICa,L resulted in EAD formation in
female, but not male, PB cells (Fig. 8c). These findings are
in agreement with experimental findings in rabbits (Pham
et al. 2001) and support the concept of limited ‘repolar-
ization reserve’ in females (Roden 2004). In our simula-
tions, the limited ‘repolarization reserve’ of female cells
was due to their larger ICa,L density and smaller IKr
density. Taken together, these observations may explain
why female sex is an independent risk factor for TdP
(Zareba et al. 1995, Locati et al. 1998). Clinical
(Schwartz et al. 1975) and experimental (Antzelevitch
& Sicouri 1994, Akar et al. 2002) studies have produced
two theories regarding the electrophysiological basis of
TdP. One theory holds that TdP arises from triggered
activity in competing ventricular foci. Evidence stems
from experimental observations (Antzelevitch & Sicouri
1994) and computer models (Zeng & Rudy 1995)
demonstrating an enhanced potential of cardiac myo-
cytes for EADs in response to factors that prolong APD.
Under circumstances of AP prolongation, the AP may
remain long enough at plateau level to permit recovery
from ICa,L inactivation during early repolarization, with
subsequent reactivation of ICa,L, resulting in interruption
of final repolarization and transient depolarization
(January & Riddle 1989, Zeng & Rudy 1995). The other
theory emphasizes dispersion of repolarization, suggest-
ing involvement of reentrant excitation (Akar et al.
2002). Our study links the observed female predomin-
ance in TdP to both theories by demonstrating both an
enhanced susceptibility to EAD formation and a larger
transmural dispersion of repolarization in female cells.
Sex influence on the clinical phenotype of Brugada
syndrome
In RV cells, we documented more negative notch
potentials in subepicardial than subendocardial APs.
Furthermore, the most negative notch potential was
found in males. Incorporation of IK,ATP caused all-or-
none repolarization in the male, but not the female,
subepicardial cell (Fig. 9). This finding is consistent
with experimental findings in dogs, where the IK,ATP
agonist pinacidil resulted in loss of the AP dome in male
� 2006 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2006.01586.x 471
Acta Physiol 2006, 187, 459–477 A O Verkerk et al. Æ Sex and human cardiac electrophysiology
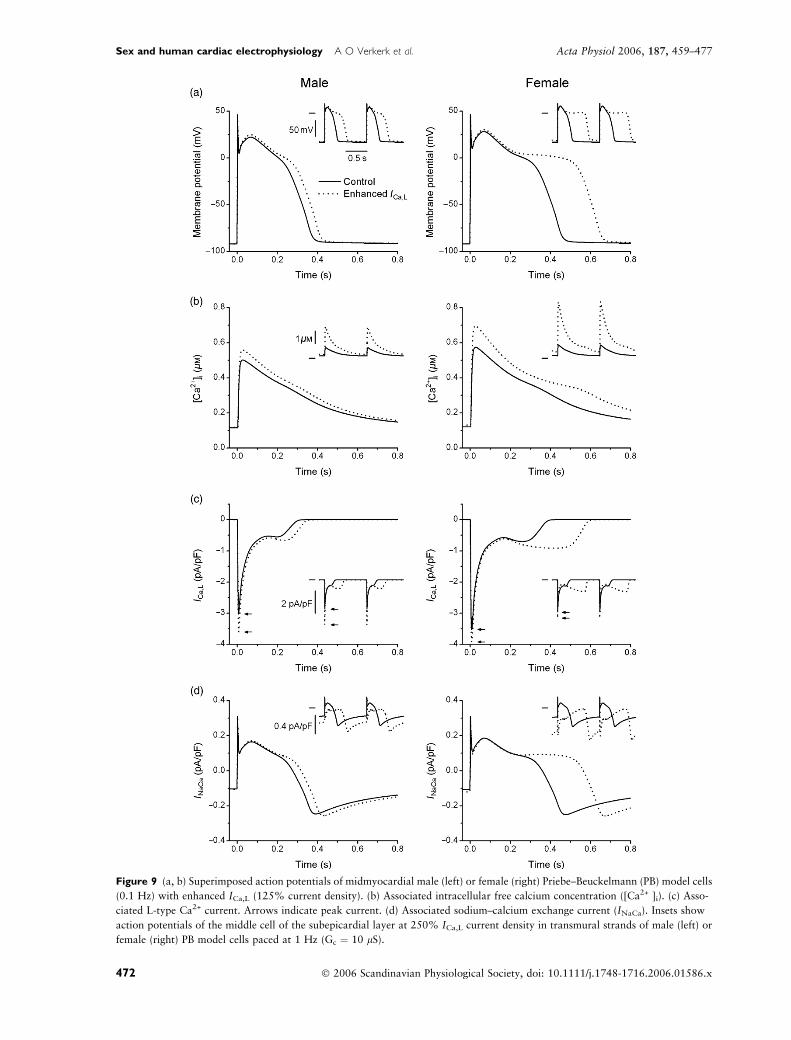
Figure 9 (a, b) Superimposed action potentials of midmyocardial male (left) or female (right) Priebe–Beuckelmann (PB) model cells
(0.1 Hz) with enhanced ICa,L (125% current density). (b) Associated intracellular free calcium concentration ([Ca2+ ]i). (c) Asso-
ciated L-type Ca2+ current. Arrows indicate peak current. (d) Associated sodium–calcium exchange current (INaCa). Insets show
action potentials of the middle cell of the subepicardial layer at 250% ICa,L current density in transmural strands of male (left) or
female (right) PB model cells paced at 1 Hz (Gc ¼ 10 lS).
472 � 2006 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2006.01586.x
Sex and human cardiac electrophysiology Æ A O Verkerk et al. Acta Physiol 2006, 187, 459–477

but not in female ventricular myocytes (Di Diego et al.
2002). IK,ATP itself is not believed to be involved in the
dysrhythmogenesis of the Brugada syndrome, but its
introduction into our simulations mimics the experi-
mental approach of blocking inward currents or
enhancing outward currents to unmask the Brugada
syndrome (Fish & Antzelevitch 2004).
Our findings demonstrate that males are exquisitely
sensitive to disruptions of the balance between depolar-
izing and repolarizing forces during the early phases of
the AP. The loss of the AP dome following small
reductions in net depolarizing force reveals that males
have a limited ‘depolarization reserve’. In our simula-
tions, the limited ‘depolarization reserve’ of male cells
was in large part because of their high Ito density in
combination with their smaller ICa,L density and larger
IKr density. This theory of limited ‘depolarization reserve’
in males is further substantiated in Brugada syndrome,
where reductions in depolarizing force result from Na+
channel mutations which reduce INa (Tan et al. 2003)
and/or the use of Na+ channel blocking drugs (class I
antidysrhythmic drugs). Thus, the concept of limited
‘depolarization reserve’ in males may explain why the
clinical phenotype of Brugada syndrome is more prom-
inent in males than females (Matsuo et al. 2001).
Order of repolarization
In our single cell simulations, AP repolarization was
completed first in subendocardial, then subepicardial,
and, finally, midmyocardial cells (Fig. 5). Such obser-
vations, with longer APD for epicardial than endocar-
dial cells, have also been made in isolated ventricular
myocytes [e.g. by Li et al. (2002)], although the
opposite (shorter APD for epicardial than endocardial
cells) has also been reported [e.g. by Liu et al. (1993)]. If
these cells are coupled together into a transmural
strand, there is only a moderate difference in APD
between the cells of the transmural layers (Figs 6a,b and
7a,b), and the repolarization wave proceeds from
endocardium to epicardium, as illustrated by the
increase in RT from endocardium to epicardium
(Figs 6c and 7c). This seems at odds with the textbook
notion of T-wave genesis, in which repolarization is
understood to begin and be completed first in the
subepicardium. However, as recently reviewed by
Conrath & Opthof (2006) with emphasis on the human
heart, there is a growing body of experimental data
suggesting that the transmural gradient in repolariza-
tion runs from endocardium to epicardium rather than
in the opposite direction. Among these data are
transmural recordings made in canine (Anyukhovsky
et al. 1996, Janse et al. 2005) and human (Conrath
et al. 2004) hearts that show little or no variation in
APD90 or activation–recovery intervals across the
ventricular wall, resulting in a repolarization wave
travelling from endocardium to epicardium. Specific-
ally, the recent experimental study by Janse et al. (2005)
demonstrates that the T wave is determined by apico-
basal gradients in RT and not by transmural gradients.
Limitations
Computer simulation studies have improved our under-
standing of the ways in which various aspects of ion
channel function (gating and expression) may alter
cardiac excitability (Kleber & Rudy 2004, Rudy 2004),
but they also have some shortcomings (Priebe &
Beuckelmann 1998, ten Tusscher et al. 2004). Our
observation that the TNNP model exhibited no EADs,
whereas the PB model did, but only with significantly
enhanced ICa,L, points to a shortcoming of all current
human ventricular cell models. As set out in detail by
Iyer et al. (2004), these models require a dominant
voltage-dependent inactivation of ICa,L to assure AP
stability. Yet, recent experimental data show that
voltage-dependent inactivation of ICa,L is slow and
incomplete, whereas Ca2+-mediated inactivation is
strong and dominates the inactivation process (Carme-
liet 2004, Iyer et al. 2004). The artificial dominant
voltage-dependent inactivation of ICa,L tends to prevent
the formation of EADs by inhibiting reactivation of
ICa,L during the plateau phase of the AP. Moreover, the
relative absence of Ca2+-mediated ICa,L inactivation
indicates that the modulatory role of Ca2+ handling
system on the AP configuration is underestimated.
In this study, we have simulated transmural conduc-
tion of the AP using linear strands with a simple and
uniform arrangement of cells. Such simulations repre-
sent the transmural conduction of broad planar activa-
tion and repolarization wavefronts during normal
ventricular excitation. However, the complex structure
of the ventricular myocardium is much simplified and
some important determinants of transmural conduc-
tion, including current-to-load mismatch and wavefront
curvature, have been excluded from our analysis.
Our recapitulation of the male predominance in the
clinical expressivity of Brugada syndrome is based on the
hypothesis that all-or-none repolarization underlies this
disorder. However, this concept is controversial
(Meregalli et al. 2005) and it has recently been demon-
strated that selective activation slowing in the RV may
also contribute to its pathophysiology (Martini 2004,
Tukkie et al. 2004). While we have not validated this
concept in this study, it is compatible with limited
‘depolarization reserve’ in males, given that conduction
velocity is primarily determined by depolarizing forces
(Tan et al. 2001). Accordingly, selective conduction
slowing in male RV may be explained by the reported
electrophysiological properties of the RV which may
� 2006 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2006.01586.x 473
Acta Physiol 2006, 187, 459–477 A O Verkerk et al. Æ Sex and human cardiac electrophysiology
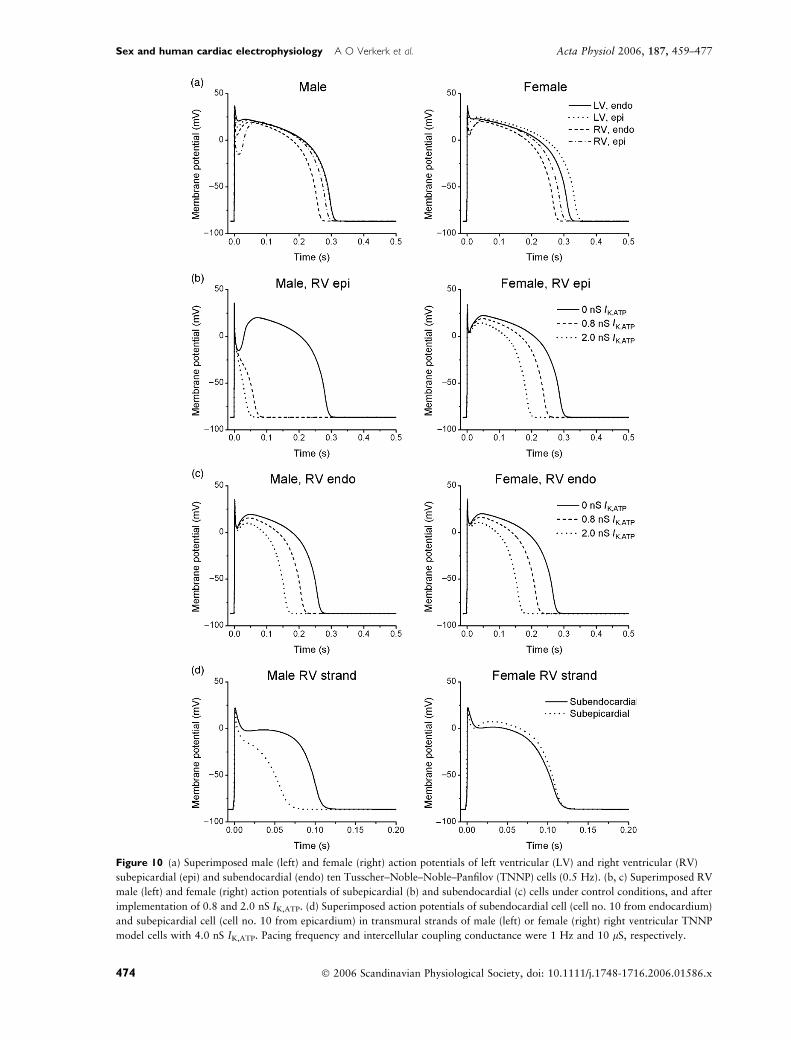
Figure 10 (a) Superimposed male (left) and female (right) action potentials of left ventricular (LV) and right ventricular (RV)
subepicardial (epi) and subendocardial (endo) ten Tusscher–Noble–Noble–Panfilov (TNNP) cells (0.5 Hz). (b, c) Superimposed RV
male (left) and female (right) action potentials of subepicardial (b) and subendocardial (c) cells under control conditions, and after
implementation of 0.8 and 2.0 nS IK,ATP. (d) Superimposed action potentials of subendocardial cell (cell no. 10 from endocardium)
and subepicardial cell (cell no. 10 from epicardium) in transmural strands of male (left) or female (right) right ventricular TNNP
model cells with 4.0 nS IK,ATP. Pacing frequency and intercellular coupling conductance were 1 Hz and 10 lS, respectively.
474 � 2006 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2006.01586.x
Sex and human cardiac electrophysiology Æ A O Verkerk et al. Acta Physiol 2006, 187, 459–477

constitute the background against which sex disparities
in net depolarizing forces produce the male predomin-
ance of disease in Brugada syndrome. Similarly, we have
recently provided evidence that RV fibrosis may be
another element in the pathophysiology of Brugada
syndrome that may be conducive to the initiation and
maintenance of reentrant tachydysrhythmias (Coronel
et al. 2005). Subsequently, in a study including 18
Brugada syndrome patients, Frustaci et al. (2005) showed
that, despite an apparently normal heart at non-invasive
evaluation, endomyocardial biopsy detected structural
alterations in all 18 patients. Thus, structural derange-
ments may also be a backdrop for the critical reduction in
depolarizing forces and the consequent increase in dys-
rhythmia susceptibility of males in Brugada syndrome.
Conclusion
The sex-related distinctions in ECG variables and
dysrhythmia susceptibility in humans may be explained
by sex disparities in ICa,L, Ito and IKr conductances,
which result in limited ‘repolarization reserve’ in
females and limited ‘depolarization reserve’ in males.
While these concepts have now been demonstrated in
model diseases (LQTS, Brugada syndrome), they may
also apply to common diseases. For instance, clinical
observations reveal that females have an increased
dysrhythmia susceptibility in common disorders where
repolarization is impaired by electrolyte disturbances or
drug use (acquired LQTS) (Lehmann et al. 1996).
Conversely, a male predominance in dysrhythmia sus-
ceptibility and sudden death may exist in heart failure
(Kannel & Schatzkin 1985), a condition which may
reduce depolarizing forces. Whether increased dysrhyth-
mia susceptibility in heart failure consistently afflicts
males, is, however, difficult to predict because of the
constellation of opposing electrophysiological changes
in heart failure. Thus, while males may be more
vulnerable to reentrant tachydysrhythmias because of
reductions in depolarizing forces, this effect may be
offset by concurrent reductions in repolarizing forces
(Armoundas et al. 2001), which may render females
more susceptible to EADs and TdP. Still, it is clear that
insights into sex disparities in dysrhythmia susceptibility
in common disease may contribute to further refine-
ments in dysrhythmia management in inherited and
common acquired diseases.
Conflict of interest
There are no conflicts of interest.
Dr Tan was supported by a fellowship of the Royal Netherlands
Academy of Arts and Sciences (KNAW), the Netherlands Heart
Foundation (NHS-2002B191), and the Bekales Foundation.
References
Akar, F.G., Yan, G.-X., Antzelevitch, C. & Rosenbaum, D.S.
2002. Unique topographical distribution of M cells underlies
reentrant mechanism of Torsade de Pointes in the long-QT
syndrome. Circulation 105, 1247–1253.
Antzelevitch, C. & Sicouri, S. 1994. Clinical relevance of car-
diac arrhythmias generated by afterdepolarizations: role of
M cells in the generation of U waves, triggered activity and
Torsade de Pointes. J Am Coll Cardiol 23, 259–277.
Anyukhovsky, E.P., Sosunov, E.A. & Rosen, M.R. 1996.
Regional differences in electrophysiological properties of
epicardium, midmyocardium, and endocardium. In vitro and
in vivo correlations. Circulation 94, 1981–1988.
Armoundas, A.A., Wu, R., Juang, G., Marban, E. & Tomaselli,
G.F. 2001. Electrical and structural remodeling of the failing
ventricle. Pharmacol Therapeut 92, 213–230.
Bazett, H.C. 1920. An analysis of the time relations of elec-
trocardiograms. Heart. 7, 353–367.
Bernus, O., Wilders, R., Zemlin, C.W., Verschelde, H. &
Panfilov, A.V. 2002. A computationally efficient electro-
physiological model of human ventricular cells. Am J Physiol
Heart Circ Physiol 282, H2296–H2308.
Bidoggia, H., Maciel, J.P., Capalozza, N. et al. 2000. Sex dif-
ferences on the electrocardiographic pattern of cardiac
repolarization: possible role of testosterone. AmHeart J 140,
678–683.
Binah, O. & Rosen, M.R. 1992. Mechanisms of ventricular
arrhythmias. Circulation 85(Suppl. 1), I25–I31.
Carmeliet, E. 2004. Intracellular Ca2+ concentration and rate
adaptation of the cardiac action potential. Cell Calcium 35,
557–573.
Chen, J., Petranka, J., Yamamura, K., London, R.E., Steen-
bergen, C. & Murphy, E. 2003. Gender differences in sar-
coplasmic reticulum calcium loading after isoproterenol.
Am J Physiol Heart Circ Physiol 285, H2657–H2662.
Conrath, C.E. & Opthof, T. 2006. Ventricular repolarization:
an overview of (patho)physiology, sympathetic effects and
genetic aspects. Prog Biophys Mol Biol in press.
Conrath, C.E., Wilders, R., Coronel, R. et al. 2004. Intercel-
lular coupling through gap junctions masks M cells in the
human heart. Cardiovasc Res 62, 407–414.
Coronel, R., Casini, S., Koopmann, T.T. et al. 2005. Right
ventricular fibrosis and conduction delay in a patient with
clinical signs of Brugada syndrome: a combined electro-
physiological, genetic, histopathologic, and computational
study. Circulation 112, 2769–2777.
Cranefield, P.F. 1977. Action potentials, afterpotentials, and
arrhythmias. Circ Res 41, 415–423.
Di Diego, J.M., Cordeiro, J.M., Goodrow, R.J. et al. 2002.
Ionic and cellular basis for the predominance of the Brugada
syndrome phenotype in males. Circulation 106, 2004–2011.
Drouin, E., Charpentier, F., Gauthier, C., Laurent, K. & Le
Marec, H. 1995. Electrophysiological characteristics of cells
spanning the left ventricular wall of human heart: evidence
for the presence of M cells. J Am Coll Cardiol 26, 185–192.
Faber, G.M. & Rudy, Y. 2000. Action potential and con-
tractility changes in [Na+ ]i overloaded cardiac myocytes:
a simulation study. Biophys J 78, 2392–2404.
� 2006 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2006.01586.x 475
Acta Physiol 2006, 187, 459–477 A O Verkerk et al. Æ Sex and human cardiac electrophysiology

Feigenbaum, H. 1994. Echocardiography, 5th edn. Lee and
Febiger, Philadelphia.
Fish, J.M. & Antzelevitch, C. 2004. Role of sodium and cal-
cium channel block in unmasking the Brugada syndrome.
Heart Rhythm 1, 210–217.
Frustaci, A., Priori, S.G., Pieroni, M. et al. 2005. Cardiac his-
tological substrate in patients with clinical phenotype of
Brugada syndrome. Circulation 112, 3680–3687.
Gima, K. & Rudy, Y. 2002. Ionic current basis of electro-
cardiographic waveforms: a model study. Circ Res 90, 889–
896.
Gottdiener, J.S., Gay, J.A., Maron, B.J. & Fletcher, R.D. 1985.
Increased right ventricular wall thickness in left ventricular
pressure overload: echocardiographic determination of
hypertrophic response of the ‘nonstressed’ ventricle. J Am
Coll Cardiol 6, 550–555.
Greenbaum, R.A., Ho, S.Y., Gibson, D.G., Becker, A.E. &
Anderson, R.H. 1981. Left ventricular fibre architecture in
man. Br Heart J 45, 248–263.
Iyer, V., Mazhari, R. & Winslow, R.L. 2004. A computational
model of the human left-ventricular epicardial myocyte.
Biophys J 87, 1507–1525.
James, A.F. Choisy, S.C. & Hancox, J.C. 2006. Recent
advances in understanding sex differences in cardiac re-
polarization. Prog Biophys Mol Biol in press.
Janse, M.J., Sosunov, E.A., Coronel, R. et al. 2005. Repolari-
zation gradients in the canine left ventricle before and after
induction of short-term cardiac memory. Circulation 112,
1711–1718.
January, C.T. & Riddle, J.M. 1989. Early afterdepolarizations:
mechanism of induction and block. A role for L-type Ca2+
current. Circ Res 64, 977–990.
Jongsma, H.J. & Wilders, R. 2000. Gap junctions in cardio-
vascular disease. Circ Res 86, 1193–1197.
Kannel, W.B. & Schatzkin, A. 1985. Sudden death: lessons
from subsets in population studies. J Am Coll Cardiol
5(Suppl.), 141B–149B.
Kleber, A.G. & Rudy, Y. 2004. Basic mechanisms of cardiac
impulse propagation and associated arrhythmias. Physiol
Rev 84, 431–488.
Kligfield, P., Lax, K.G. & Okin, P.M. 1996. QT interval-heart
rate relation during exercise in normal men and women:
definition by linear regression analysis. J Am Coll Cardiol
28, 1547–1555.
Leblanc, N., Chartier, D., Gosselin, H. & Rouleau, J.L. 1998.
Age and gender differences in excitation-contraction cou-
pling of the rat ventricle. J Physiol 511, 533–548.
Lehmann, M.H., Hardy, S., Archibald, D., Quart, B. &
MacNeil, D.J. 1996. Sex difference in risk of torsade de
pointes with d,l-sotalol. Circulation 94, 2535–2541.
Li, G.R., Feng, J., Yue, L. & Nattel, S. 1996. Evidence for two
components of delayed rectifier K+ current in human ven-
tricular myocytes. Circ Res 78, 689–696.
Li, G.R., Lau, C.P., Ducharme, A., Tardif, J.C. & Nattel, S.
2002. Transmural action potential and ionic current
remodeling in ventricles of failing canine hearts. Am J
Physiol Heart Circ Physiol 283, H1031–H1041.
Liu, D.W., Gintant, G.A. & Antzelevitch, C. 1993. Ionic
bases for electrophysiological distinctions among epicardial,
midmyocardial, and endocardial myocytes from the free wall
of the canine left ventricle. Circ Res 72, 671–687.
Liu, X.K., Katchman, A., Drici, M.D. et al. 1998. Gen-
der difference in the cycle length-dependent QT and
potassium currents in rabbits. J Pharmacol Exp Ther 285,
672–679.
Locati, E.H., Zareba, W., Moss, A.J. et al. 1998. Age- and sex-
related differences in clinical manifestations in patients with
congenital long-QT syndrome: findings from the Interna-
tional LQTS Registry. Circulation 97, 2237–2244.
Luo, C.H. & Rudy, Y. 1994. A dynamic model of the cardiac
ventricular action potential. I. Simulations of ionic currents
and concentration changes. Circ Res 74, 1071–1096.
Martini, B. 2004. Further confirmation that a conduction
disturbance underlies the electrocardiographic pattern of the
so-called Brugada syndrome. Circulation 110, e53.
Matsuo, K., Akahoshi, M., Nakashima, E. et al. 2001. The
prevalence, incidence and prognostic value of the Brugada-
type electrocardiogram: a population-based study of four
decades. J Am Coll Cardiol 38, 765–770.
Matsuo, K., Akahoshi, M., Seto, S. & Yano, K. 2003.
Disappearance of the Brugada-type electrocardiogram after
surgical castration: a role for testosterone and an explana-
tion for the male preponderance. Pacing Clin Electrophysiol
26, 1551–1553.
Mendelsohn, M.E. & Karas, R.H. 2005. Molecular and cel-
lular basis of cardiovascular gender differences. Science 308,
1583–1587.
Meregalli, P.G., Wilde, A.A.M. & Tan, H.L. 2005. Patho-
physiological mechanisms of Brugada syndrome: depolar-
ization disorder, repolarization disorder, or more?
Cardiovasc Res 67, 367–378.
Merri, M., Benhorin, J., Alberti, M., Locati, E. & Moss, A.J.
1989. Electrocardiographic quantitation of ventricular
repolarization. Circulation 80, 1301–1308.
Pham, T.V. & Rosen, M.R. 2002. Sex, hormones, and
repolarization. Cardiovasc Res 53, 740–751.
Pham, T.V., Sosunov, E.A., Gainullin, R.Z., Danilo, P. Jr &
Rosen, M.R. 2001. Impact of sex and gonadal steroids on
prolongation of ventricular repolarization and arrhythmias
induced by IK-blocking drugs. Circulation 103, 2207–
2212.
Pham, T.V., Robinson, R.B., Danilo, P. Jr & Rosen, M.R.
2002. Effects of gonadal steroids on gender-related differ-
ences in transmural dispersion of L-type calcium current.
Cardiovasc Res 53, 752–762.
Priebe, L. & Beuckelmann, D.J. 1998. Simulation study of
cellular electric properties in heart failure. Circ Res 82,
1206–1223.
Ranki, H.J., Budas, G.R., Crawford, R.M. & Jovanovic, A.
2001. Gender-specific difference in cardiac ATP-sensitive K+
channels. J Am Coll Cardiol 38, 906–915.
Rautaharju, P.M., Zhou, S.H., Wong, S. et al. 1992. Sex
differences in the evolution of the electrocardiographic QT
interval with age. Can J Cardiol 8, 690–695.
Roden, D.M. 2004. Drug-induced prolongation of the QT
interval. N Engl J Med 350, 1013–1022.
Rudy, Y. 2004. From genetics to cellular function using com-
putational biology. Ann N Y Acad Sci 1015, 261–270.
476 � 2006 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2006.01586.x
Sex and human cardiac electrophysiology Æ A O Verkerk et al. Acta Physiol 2006, 187, 459–477

Schwartz, P.J., Periti, M. & Malliani, A. 1975. Fundamentals
of clinical cardiology: the long Q-T syndrome. Am Heart J
89, 378–390.
Stramba-Badiale, M., Locati, E.H., Martinelli, A., Courville, J.
& Schwartz, P.J. 1997. Gender and the relationship between
ventricular repolarization and cardiac cycle length during
24-h Holter recordings. Eur Heart J 18, 1000–1006.
Streeter, D.D. & Hanna, W.T. 1973. Engineering mechanics
for successive states in canine left ventricular myocardium II.
Fiber angle and sarcomere length. Circ Res 33, 656–664.
Sugishita, K., Su, Z., Li, F., Philipson, K.D. & Barry, W.H.
2001. Gender influences [Ca2+ ]i during metabolic inhibition
in myocytes overexpressing the Na+-Ca2+ exchanger.
Circulation 104, 2101–2106.
Sun, X. & Wang, H.-S. 2005. Role of the transient outward
current (Ito) in shaping canine ventricular action potential – a
dynamic clamp study. J Physiol 564, 411–419.
Taggart, P., Sutton, P.M., Opthof, T. et al. 2000. Inhomoge-
neous transmural conduction during early ischaemia in pa-
tients with coronary artery disease. J Mol Cell Cardiol 32,
621–630.
Tan, H.L., Hou, C.J.Y., Lauer, M.R. & Sung, R.J. 1995.
Electrophysiologic mechanisms of the long QT interval
syndromes and torsade de pointes. Ann Intern Med 122,
701–714.
Tan, H.L., Bink-Boelkens, M.T., Bezzina, C.R. et al. 2001. A
sodium-channel mutation causes isolated cardiac conduction
disease. Nature 409, 1043–1047.
Tan, H.L., Bezzina, C.R., Smits, J.P.P., Verkerk, A.O. &
Wilde, A.A.M. 2003. Genetic control of sodium channel
function. Cardiovasc Res 57, 961–973.
Trepanier-Boulay, V., St-Michel, C., Tremblay, A. & Fiset, C.
2001. Gender-based differences in cardiac repolarization in
mouse ventricle. Circ Res 89, 437–444.
Tukkie, R., Sogaard, P., Vleugels, J., de Groot, I.K., Wilde,
A.A.M. & Tan, H.L. 2004. Delay in right ventricular
activation contributes to Brugada syndrome. Circulation
109, 1272–1277.
ten Tusscher, K.H.W.J., Noble, D., Noble, P.J. & Panfilov,
A.V. 2004. A model for human ventricular tissue. Am J
Physiol Heart Circ Physiol 286, H1573–H1589.
ten Tusscher, K.H.W.J., Bernus, O., Hren, R. & Panfilov, A.V.
2006. Comparison of electrophysiological models for human
ventricular cells and tissues. Prog Biophys Mol Biol 90,
326–345.
Valverde, E.R., Biagetti, M.O., Bertran, G.R., Arini, P.D., Bi-
doggia, H. & Quinteiro, R.A. 2003. Developmental changes
of cardiac repolarization in rabbits: implications for the role
of sex hormones. Cardiovasc Res 57, 625–631.
Veldkamp, M.W., Verkerk, A.O., van Ginneken, A.C.G. et al.
2001. Norepinephrine induces action potential prolongation
and early afterdepolarizations in ventricular myocytes iso-
lated from human end-stage failing hearts. Eur Heart J 22,
955–963.
Verkerk, A.O., Wilders, R., Veldkamp, M.W., de Geringel, W.,
Kirkels, J.H. & Tan, H.L. 2005. Gender disparities in car-
diac cellular electrophysiology and arrhythmia susceptibility
in human failing ventricular myocytes. Int Heart J 46, 1105–
1118.
Volders, P.G., Sipido, K.R., Carmeliet, E., Spatjens, R.L.,
Wellens, H.J. & Vos, M.A. 1999. Repolarizing K+ currents
ITO1 and IKs are larger in right than left canine ventricular
midmyocardium. Circulation 99, 206–210.
Winslow, R.L., Rice, J., Jafri, S., Marban, E. & O’Rourke, B.
1999. Mechanisms of altered excitation-contraction coup-
ling in canine tachycardia-induced heart failure, II: model
studies. Circ Res 84, 571–586.
Wu, Y. & Anderson, M.E. 2002. Reduced repolarization
reserve in ventricular myocytes from female mice. Cardio-
vasc Res 53, 763–769.
Yan, G.-X. & Antzelevitch, C. 1999. Cellular basis for
the Brugada syndrome and other mechanisms of
arrhythmogenesis associated with ST segment elevation.
Circulation 100, 1660–1666.
Zareba, W., Moss, A.J., le Cessie, S. et al. 1995. Risk of car-
diac events in family members of patients with long QT
syndrome. J Am Coll Cardiol 26, 1685–1691.
Zeng, J. & Rudy, Y. 1995. Early afterdepolarizations in car-
diac myocytes: mechanism and rate dependence. Biophys J
68, 949–964.
Zygmunt, A.C., Goodrow, R.J. & Antzelevitch, C. 2000. INaCa
contributes to electrical heterogeneity within the canine
ventricle. Am J Physiol Heart Circ Physiol 278, H1671–
H1678.
Zygmunt, A.C., Eddlestone, G.T., Thomas, G.P., Nesterenko,
V.V. & Antzelevitch, C. 2001. Larger late sodium con-
ductance in M cells contributes to electrical heterogeneity in
canine ventricle. Am J Physiol Heart Circ Physiol 281,
H689–H697.
� 2006 Scandinavian Physiological Society, doi: 10.1111/j.1748-1716.2006.01586.x 477
Acta Physiol 2006, 187, 459–477 A O Verkerk et al. Æ Sex and human cardiac electrophysiology