Embed Size (px)
Citation preview

doi:10.1093/brain/awl184 Brain (2006), 129, 2471–2483
Comparison of the fastest regenerating motorand sensory myelinated axons in the sameperipheral nerve
Mihai Moldovan,1 Jesper Sørensen1,2 and Christian Krarup1,3
1Division of Neurophysiology, Institute of Medical Physiology, Panum Institute, University of Copenhagen, Departments of,2Plastic Surgery and 3Clinical Neurophysiology, The Neuroscience Center, Rigshospitalet, Copenhagen, Denmark
Correspondence to: Christian Krarup, MD, DMSc, FRCP, Department of Clinical Neurophysiology, The NeuroscienceCenter, NF3063, Rigshospitalet, 9 Blegdamsvej, 2100 Copenhagen, DenmarkE-mail: [email protected]
Functional outcome after peripheral nerve regeneration is often poor, particularly involving nerve injuries farfrom their targets. Comparison of sensory and motor axon regeneration before target reinnervation is notpossible in the clinical setting, and previous experimental studies addressing the question of differences ingrowth rates of different nerve fibre populations led to conflicting results. We developed an animal model tocompare growth and maturation of the fastest growing sensory and motor fibres within the same mixed nerveafter Wallerian degeneration. Regeneration of cat tibial nerve after crush (n = 13) and section (n = 7) wasmonitored for up to 140 days, using implanted cuff electrodes placed around the sciatic and tibial nerves andwire electrodes at plantar muscles. To distinguish between sensory and motor fibres, recordings were carriedout from L6–S2 spinal roots using cuff electrodes. The timing of laminectomy was based on the presence ofregenerating fibres along the nerve within the tibial cuff. Stimulation of unlesioned tibial nerves (n = 6) evokedthe largest motor response in S1 ventral root and the largest sensory response in L7 dorsal root. Growth rateswere compared by mapping the regenerating nerve fibres within the tibial nerve cuff to all ventral or dorsalroots and, regardless of the lesion type, the fastest growth was similar in sensory and motor fibres. Maturationwas assessed as recovery of themaximummotor and sensory conduction velocities (CVs) within the tibial nervecuff. Throughout the observation period the CV was �14% faster in regenerated sensory fibres than in motorfibres in accordancewith the difference observed in control nerves. Recovery of amplitude was only partial aftersection, whereas the root distribution pattern was restored. Our data suggest that the fastest growth andmaturation rates that can be achieved during regeneration are similar formotor and sensorymyelinated fibres.
Keywords: nerve; regeneration; laminectomy; cuff electrodes; cat
Abbreviations: CMAP = compound muscle action potential; CNAP = compound nerve action potentials; CRP = compoundroot action potential; CV = conduction velocity
Received March 28, 2006. Revised May 30, 2006. Accepted June 15, 2006. Advance Access publication August 10, 2006.
IntroductionAfter a nerve injury, both motor and sensory axons have the
ability to regenerate and, given a proper pathway, reconnect
with targets. Despite this capacity, the functional outcome of
peripheral nerve regeneration is often poor, particularly after
nerve injuries that sever peripheral nerves far from their
targets (Lundborg, 2003). In a long-term study of nerve
regeneration in non-human primates the lesion and repair
type, through the time to muscle reinnervation, influenced
recovery assessed by physiological methods (Krarup et al.,
2002). Thus, both the distance and rate of regeneration
probably play important roles in determining the outcome
of nerve regeneration.
Axonal transport responsible for nerve regeneration is slow
(Wujek and Lasek, 1983) for both motor and sensory axons
(Braendgaard and Sidenius, 1986). The fastest growth rates
approach 4 mm/day depending on the regeneration environ-
ment (Fugleholm et al., 1994, 1998, 2000; Sorensen et al.,
2001). In the mouse distal stump, Schwann cells previously
associated with motor axons retain specialized features
(Martini et al., 1992), and this was suggested to contribute
to preferential reinnervation of motor pathways by motor
axons (Brushart, 1988), which may improve functional out-
come of motor regeneration after section (Brushart, 1993). It
is unknown whether such environmental specializations
# The Author (2006). Published by Oxford University Press on behalf of the Guarantors of Brain. All rights reserved. For Permissions, please email: [email protected]
Dow
nloaded from https://academ
ic.oup.com/brain/article-abstract/129/9/2471/332714 by guest on 23 N
ovember 2018

accelerate motor axon growth rates compared with sensory
axon growth rates.
Comparison of sensory and motor axon regeneration
before target reinnervation is not possible in the clinical
setting, and previous experimental studies addressing the
question of differences in growth rates of different nerve
fibre populations led to conflicting results. In some studies,
axons in muscle or mixed nerves were found to regenerate
faster than fibres in pure sensory nerves (Jenq and Coggeshall,
1985; Chen and Bisby, 1993). A better motor than sensory
regeneration was also suggested in the same mixed nerve (da
Silva et al., 1985; Madison et al., 1988). Other studies, how-
ever, indicated that sensory fibres dominate in the early stages
of regeneration (Sanger et al., 1991; Madorsky et al., 1998;
Suzuki et al., 1998; Kawasaki et al., 2000). Nevertheless, the
fastest growing motor fibres were found to regenerate as fast
as the sensory fibres (Forman and Berenberg, 1978; Forman
et al., 1979). The reason for these controversies may be attrib-
uted to the technical difficulties of identifying the fastest
growing fibres at the front of regeneration.
The aim of this study was to compare, in a large animal
model, the fastest rates of regeneration that are achieved by
sensory and motor fibres within the same mixed nerve both
after crush and section. We previously developed an electro-
physiological method in cat to monitor growth of axons in
the tibial nerve through a series of electrodes implanted
around the nerves in a silicone cuff and confirmed by his-
tological studies (Fugleholm et al., 1994, 1998; Sorensen et al.,
2001). In this study, to distinguish between motor and sen-
sory fibres, we recorded evoked action potentials from all the
ventral and dorsal roots containing fibres projecting to
the tibial nerve. To compare rates of growth, we mapped
the distances reached within the tibial nerve cuff by fibres
belonging to different spinal roots. Furthermore, to compare
maturation, recovery of motor and sensory conduction velo-
cities (CVs) within the tibial nerve cuff was investigated for
up to 5 months of regeneration.
Material and methodsAnimals, experimental design and anaesthesiaElectrodes were implanted in both legs in the plantar muscles and
around the sciatic and tibial nerves of 15 adult female cats (2.5–
3.2 kg, Iffa-Credo, France). After implantation, all animals regained
normal movements and gait and stayed healthy during the experi-
ment. Data from four legs were excluded from the analysis because
the cat destroyed the cabling connecting the electrodes.
The study was performed in five consecutive steps: (i) surgery for
electrode implantation and nerve lesions; (ii) electrophysiological
validation of the nerve lesion; (iii) serial recordings to identify the
outgrowth of the tibial nerve fibres; (iv) terminal laminectomies to
compare growth of sensory and motor fibres in the tibial cuff after
crush (n = 5) and section (n = 4); (v) terminal laminectomies
to compare maturation of sensory and motor axons after target
reinnervation after crush (n = 8) and section (n = 3). Recordings
from roots were performed in six unlesioned tibial nerves for control
1 week after the electrode implantation.
Chronic serial investigations were carried out under anaesthesia
by a mixture of 0.7 ml Ketamine (10 mg/kg) and 0.4 ml Xylazine
(2 mg/kg) delivered by intramuscular injection and maintained by
subcutaneous injection as needed. Electrode implantations and
laminectomies were carried out during deep anaesthesia induced
with pentobarbital 40 mg/kg, 2.5 ml intraperitoneal, and maintained
by repeated intravenous injection of 1 ml from the 10% dilution.
During anaesthesia, the core temperature was controlled by placing
the animal on a thermostat-heated rubber pad (37�C) and the per-
ipheral temperature was maintained using a thermostat-controlled
heating lamp (37�C). At the completion of the investigations, the
cats were killed by an overdose of 250 mg pentobarbital i.v. The
experiments were performed with approval of the Danish National
Animal Experiment Committee.
Electrode implantations and nerve lesionsProcedures of nerve lesions and electrode implantation (Krarup
and Loeb, 1988) were described in detail previously (Fugleholm
et al., 1994, 1998, 2000; Sorensen et al., 2001). Briefly, after exposure
of the tibial nerves over �70 mm, crush lesions were carried out by
clamping a silicone coated forceps for 2 min, and complete transec-
tion lesions were immediately co-adapted using microsurgical tech-
niques by four epineural sutures (10-0 Taper Point, S&T Neuhausen,
Switzerland). Eight-lead semicircular cuff electrodes (Larsen et al.,
1998) with leads spaced 7.5 mm apart were implanted around the
tibial nerves (Fig. 1A) and positioned so that the first electrode (T1)
was 10 mm distal to the lesion site (marked with a 6-0 suture). Six-
lead cuff electrodes were implanted around the sciatic nerves (two
pairs of three leads, with 18 mm between the centre recording
leads). The electrode placed around the tibial nerve had an internal
diameter of 3 mm and the cuff around the sciatic nerve had an
internal diameter of 4 mm, which was at least 30% larger than
the diameter of the nerves to avoid compression. Wire electrodes
for recording of plantar muscle activity were sutured to the fascia
of the flexor digitorum brevis muscle and subcutaneously on the
dorsum of the paw. A ground wire electrode was implanted sub-
cutaneously. The connecting teflon-coated stainless steel wires (AS
631, Cooner WIRE Co., Chatsworth, CA, USA) connecting the elec-
trodes were passed subcutaneously to the back, resurfaced through a
skin excision and mounted on an integrated circuit board protected
by an aluminium shield.
The integrity of the implanted electrodes and electrical connec-
tions was tested in situ by measuring the impedance between the
leads and ground. Complete axonal loss after the nerve lesion was
ascertained electrophysiologically 1 week after the lesion.
Serial investigations to identify the growthfront of the tibial nerve fibresDuring the first month after the lesion, serial electrophysiological
studies were carried out every 4–6 days. An illustration of the
recording set-up is presented in Fig. 1A. Supramaximal stimulation
(10 mA at 0.5–3 Hz) was delivered consecutively to the T1–T7 leads
in the tibial cuff with the anode placed one lead distal to the cathode.
Biphasic rectangular negative-positive pulses (duration = 0.1 ms)
were generated from a photo-isolated battery-powered, constant
current stimulator. The ascending evoked compound nerve action
potentials (CNAPs) were recorded (10C02 Dantec, 0.2–6 kHz)
in two tripolar electrode configurations (R1 and R2) in the
sciatic cuff. The stimulus was first applied to T1, and if a clearly
2472 Brain (2006), 129, 2471–2483 M. Moldovan et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article-abstract/129/9/2471/332714 by guest on 23 N
ovember 2018
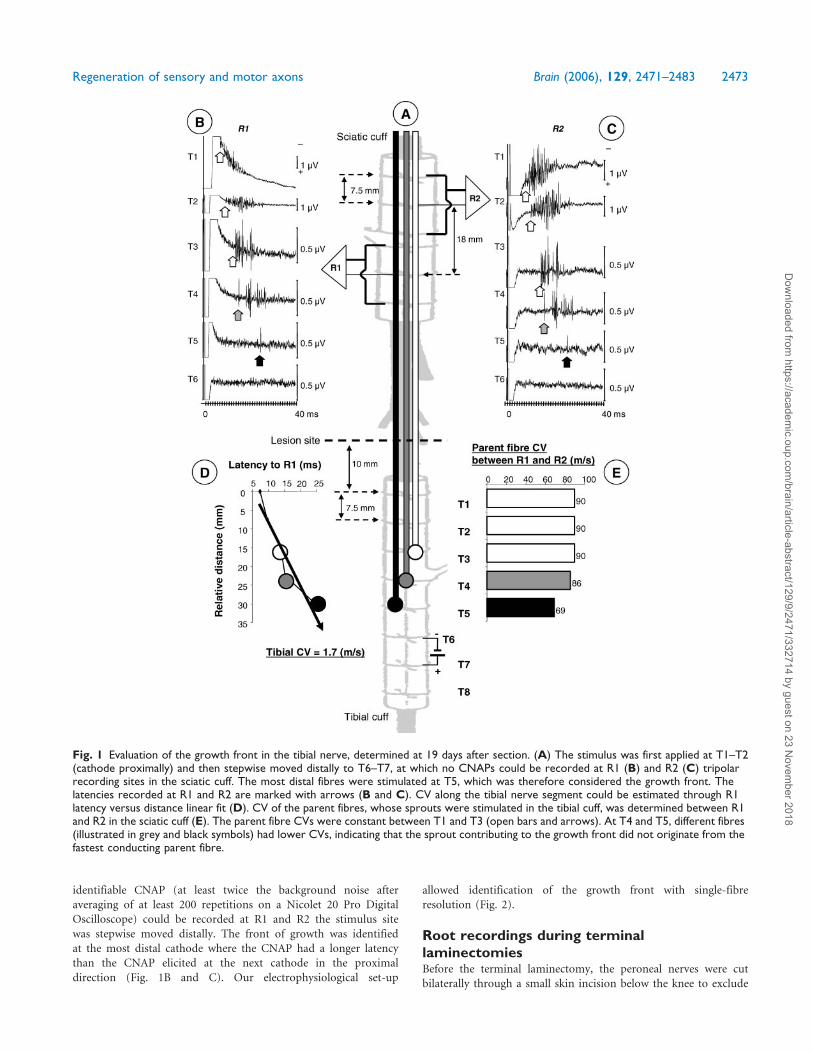
identifiable CNAP (at least twice the background noise after
averaging of at least 200 repetitions on a Nicolet 20 Pro Digital
Oscilloscope) could be recorded at R1 and R2 the stimulus site
was stepwise moved distally. The front of growth was identified
at the most distal cathode where the CNAP had a longer latency
than the CNAP elicited at the next cathode in the proximal
direction (Fig. 1B and C). Our electrophysiological set-up
allowed identification of the growth front with single-fibre
resolution (Fig. 2).
Root recordings during terminallaminectomiesBefore the terminal laminectomy, the peroneal nerves were cut
bilaterally through a small skin incision below the knee to exclude
Fig. 1 Evaluation of the growth front in the tibial nerve, determined at 19 days after section. (A) The stimulus was first applied at T1–T2(cathode proximally) and then stepwise moved distally to T6–T7, at which no CNAPs could be recorded at R1 (B) and R2 (C) tripolarrecording sites in the sciatic cuff. The most distal fibres were stimulated at T5, which was therefore considered the growth front. Thelatencies recorded at R1 and R2 are marked with arrows (B and C). CV along the tibial nerve segment could be estimated through R1latency versus distance linear fit (D). CV of the parent fibres, whose sprouts were stimulated in the tibial cuff, was determined between R1and R2 in the sciatic cuff (E). The parent fibre CVs were constant between T1 and T3 (open bars and arrows). At T4 and T5, different fibres(illustrated in grey and black symbols) had lower CVs, indicating that the sprout contributing to the growth front did not originate from thefastest conducting parent fibre.
Regeneration of sensory and motor axons Brain (2006), 129, 2471–2483 2473
Dow
nloaded from https://academ
ic.oup.com/brain/article-abstract/129/9/2471/332714 by guest on 23 N
ovember 2018
erroneous conduction to the roots. The cat was placed in a stereo-
taxic frame and the vertebrae were exposed by stepwise removal
of fascia, muscles and ligaments adjacent to os sacrum and the
vertebrae. The L6–S2 (Highland et al., 1990; Bailey et al., 1992)
ventral and dorsal roots were separated (polished forceps, type
FRS-15 & RM-8; S&T Neuhausen, Switzerland) under optic mag-
nification and cut as close to the spinal cord as possible (�3 cm free
length). In order to ensure fixed geometry of the recording electro-
des, cuffs rather than hooks were used. Each ventral or dorsal root
was pulled gently through the tubular cuff (3 circular leads 7.5 mm
apart) by a thin thread (5–0) attached to its free tip. The internal cuff
diameter was 1.0 mm (around S1 and S2 roots) and 2.0 mm (around
the L7 and L6 roots), owing to the differences in root thickness. The
contact between the root and the leads in the cuff was ensured by
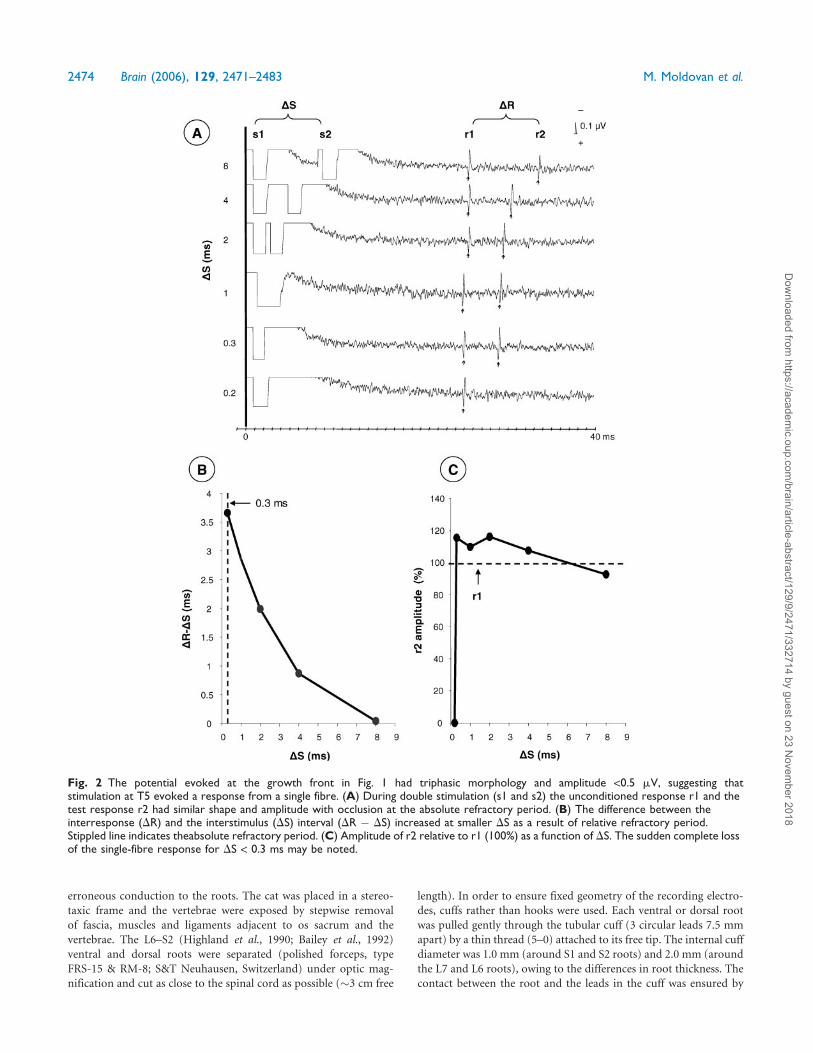
Fig. 2 The potential evoked at the growth front in Fig. 1 had triphasic morphology and amplitude <0.5 mV, suggesting thatstimulation at T5 evoked a response from a single fibre. (A) During double stimulation (s1 and s2) the unconditioned response r1 and thetest response r2 had similar shape and amplitude with occlusion at the absolute refractory period. (B) The difference between theinterresponse (DR) and the interstimulus (DS) interval (DR � DS) increased at smaller DS as a result of relative refractory period.Stippled line indicates theabsolute refractory period. (C) Amplitude of r2 relative to r1 (100%) as a function of DS. The sudden complete lossof the single-fibre response for DS < 0.3 ms may be noted.
2474 Brain (2006), 129, 2471–2483 M. Moldovan et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article-abstract/129/9/2471/332714 by guest on 23 N
ovember 2018

physiological saline solution maintained by the capillarity of the cuff.
In order to prevent tissue dry-out the remaining exposed spinal cord
was covered with gauze soaked in saline.
The ascending ventral and dorsal compound root action poten-
tials (CRPs) were recorded (10C02 Dantec, 0.2–6 kHz) in tripolar
electrode configurations identical to the R1 and R2 sciatic nerve
recording sites. The integrity of the root recordings was ascertained
by stimulation in the sciatic cuff with cathode placed at R2 and R1
electrode positions (Fig. 3). Sciatic stimulation evoked CRPs from all
ventral and dorsal L6, L7, S1 and S2, whereas no CRPs could be
Fig. 3 Comparison of distances attained by motor and sensory fibres for the tibial nerve in Fig. 1 was carried out during a terminallaminectomy (A). Tripolar recordings were carried out from cuffs placed around ventral (B) and dorsal (C) roots L6–S2. The integrityof root recordings was ascertained by stimulating in the sciatic cuff at R2 and R1 positions. Tibial nerve stimuli were firstapplied at T1–T2 (cathode proximally) and were then stepwise moved distally until no clearly identifiable CRPs—at least twice thebackground noise after averaging 500 repetitions–could be recorded at each of the investigated roots. The motor growth front wasidentified at T4 by S1V recording. The sensory growth front was identified at T5 by L6D recording. It may be noted that on thebasis of the refractory period measurements in Fig. 2 sensory growth front at T5 was represented by a single fibre that could be identifiedby CRP recordings.
Regeneration of sensory and motor axons Brain (2006), 129, 2471–2483 2475
Dow
nloaded from https://academ
ic.oup.com/brain/article-abstract/129/9/2471/332714 by guest on 23 N
ovember 2018

recorded from S2 dorsal root at tibial stimulation in any of the
investigated nerves. The largest CRP amplitudes were at least as
large as the corresponding R1 and R2 recordings and allowed
identification at the root level of the single fibre(s) at the front of
regeneration (Fig. 3).
Motor and sensory growth frontsLaminectomies were carried so that the growth front approached but
did not exceed the distal end of the tibial cuff. Stimuli were first
applied at T1–T2 (cathode proximally) and then stepwise moved
distally until no clearly identifiable CRP (at least twice the back-
ground noise after averaging 500 repetitions) could be recorded at
each of the investigated roots (Fig. 3). The motor growth front
was identified at the most distal cathode position where stimulation
evoked ventral root CRPs (Fig. 3B). Similarly, the sensory growth
front was identified at the most distal cathode position where
stimulation evoked dorsal root CRPs (Fig. 3C). The stability of
the growth front was ascertained after the 6–10 h of recordings
during laminectomy. The actual growth distance, and subsequently
the growth rate, could only be determined at the completion of the
study when the nerves were exposed and T1 position in respect to the
lesion mark was reassessed to allow corrections due to the sliding of
the cuff. At the completion of the experiment, the distances varied
from 8 to 14 mm, and the measured distance was used to calculate
growth rates. The slight displacement probably occurred during the
first week of the experiment when collagen formed around the
electrode (Fugleholm et al., 1994).
Motor and sensory CVsMaturation of the fastest axons was compared by investigating
recovery in motor and sensory CVs after target reinnervation.
Laminectomies were performed between 52 and 139 days after
the lesion, when compound muscle action potentials (CMAPs)
could be recorded (10C02 Dantec, 0.01–10 kHz) after tibial stimula-
tion. The latencies of the CNAPs and CRPs were measured to the
first positive peak and latencies of the CMAPs were measured to the
take-off of the negative phase to calculate the CVs of the fastest
fibres. Because of known distances between electrodes relative to
T1, the CV along the nerve segment in the tibial cuff could be
estimated from the slope of the latency versus distance linear fit
without measuring the distances to the recording sites. The
motor CV was estimated as the fastest CV among all ventral
roots. Similarly, the sensory CV was estimated as the fastest CV
to all dorsal roots. This method of CV calculation was validated
by comparing the motor CV with the plantar CV in the same nerve
both in regenerated (Fig. 4) and control nerves (data not shown).
Motor and sensory amplitudesThe amplitudes of CRPs and CNAPs were measured peak to
peak. Amplitude measurements depend on the number, distribution
and temporal dispersion of the different single-fibre potentials con-
tributing to the compound response. Since we could not compare
the ‘control’ and ‘regenerated’ amplitudes in the same nerve, we
limited our investigations to the recovery and distribution of the
maximum sensory and motor amplitude that could be attained
(among all stimulation sites and among all roots).
StatisticsResults are given as mean 6 standard error of measurement (SEM).
Owing to the small sample size, comparisons were performed using
non-parametric statistics (SPSS Inc. SPSS for Windows, release 13,
2005, Chicago). Individual tests are mentioned where used.
ResultsFastest rate of growth in the tibial nerveBy serial observations during the first month after the
lesion, the progression of the growth front without distinc-
tion between sensory and motor fibres could be followed
along the tibial nerve (Fig. 1). At the completion of the
experiments, the growth rate in the tibial cuff was calculated
from the slope of the linear fit of the growth distance versus
time after lesion plot (Fig. 5). The crushed fibres had a growth
rate through the cuff of �4 mm/day (Fig. 5A) and the sec-
tioned fibres slightly lower at 3.7 mm/day (Fig. 5B). The
intercept was�8 days both after crush and section, indicating
a similar delay between lesion and sprouting (Fig. 5A and B).
These values were in agreement with previous findings on the
same model (Krarup et al., 1989; Fugleholm et al., 1994).
Motor and sensory growth frontsTo compare the distances reached by sensory and motor
fibres within the tibial cuff a laminectomy was performed
between 18 and 21 days after the lesion (Fig. 3). The distances
reached by the motor and sensory fibres at the growth front
were similar both after crush (Fig. 5C) and section (Fig. 5D).
The differences between motor and sensory growth fronts
never exceeded one stimulation level (7.5 mm).
At the termination of the experiment after measurement
of the tibial cuff placement in respect to the lesion mark,
differences between the motor and sensory fibre growth
were compared with the variability of the overall fastest
growth rate in the tibial cuff (Fig. 5A and B). After crush,
the difference between the motor and the sensory growth
fronts remained within the 95% confidence limits
(67.3 mm, alpha = 0.05) of the growth front estimated
from serial observations. After section, the variability was
greater but still no significant differences between motor
and sensory fibres could be detected.
Parent fibre CVParent fibre CV of the axons stimulated in the tibial cuff could
be measured between the R1 and R2 sciatic recording sites
(Fig. 1E). Since only one recording site was feasible at the
root cuff, it was not possible to distinguish the parent fibre
CV of sensory and motor fibres in our study. In controls,
sciatic nerve CV was 85 6 1 m/s. During early outgrowth
(Fig. 1E) and during post-reinnervation maturation (data
not shown), parent fibre CVs at the sciatic nerve were in
the normal range. Nevertheless, when single fibres were acti-
vated at the tip of the regeneration front, their parent fibre CVs
were lower than the parent velocities of fibres activated at more
proximal stimulation sites with lower growth rates (Figs 1E
and 2). These observations indicated that it was not the fastest
fibres (parent fibre CV) that constituted the growth front.
2476 Brain (2006), 129, 2471–2483 M. Moldovan et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article-abstract/129/9/2471/332714 by guest on 23 N
ovember 2018
Recovery of CV along the tibial nervesIn control nerves, average motor CV (77 6 3 m/s; n = 6) was
lower than the sensory CV (956 4 m/s, P < 0.05, Wilcoxon).
During outgrowth through the tibial cuff the motor and
sensory CVs along the regenerating nerve segments were
<3 m/s both after crush and section (Fig. 1D). At this
early time point, latency changes were uneven as different
axons with different CVs reached different cathodes (Fig. 1A)
and no consistent differences were detected between the
sensory and motor mean conduction velocities estimated
from the linear slopes. After 7 weeks of regeneration, the
latency–distance relationships in the tibial cuff were linear
both after crush and section, and CVs could be accurately
estimated (Fig. 4). The recovery of CV from pooled observa-
tions collected at different laminectomies could best be
described by a logarithmic function (Fig. 6A) both for
motor CV (R2 = 0.94) and sensory CV (R2 = 0.98). The
slope seemed steeper for sensory (42 m/s/day) than for
motor (35 m/s/day), indicating that the absolute difference
between the motor and the sensory CVs increased with time.
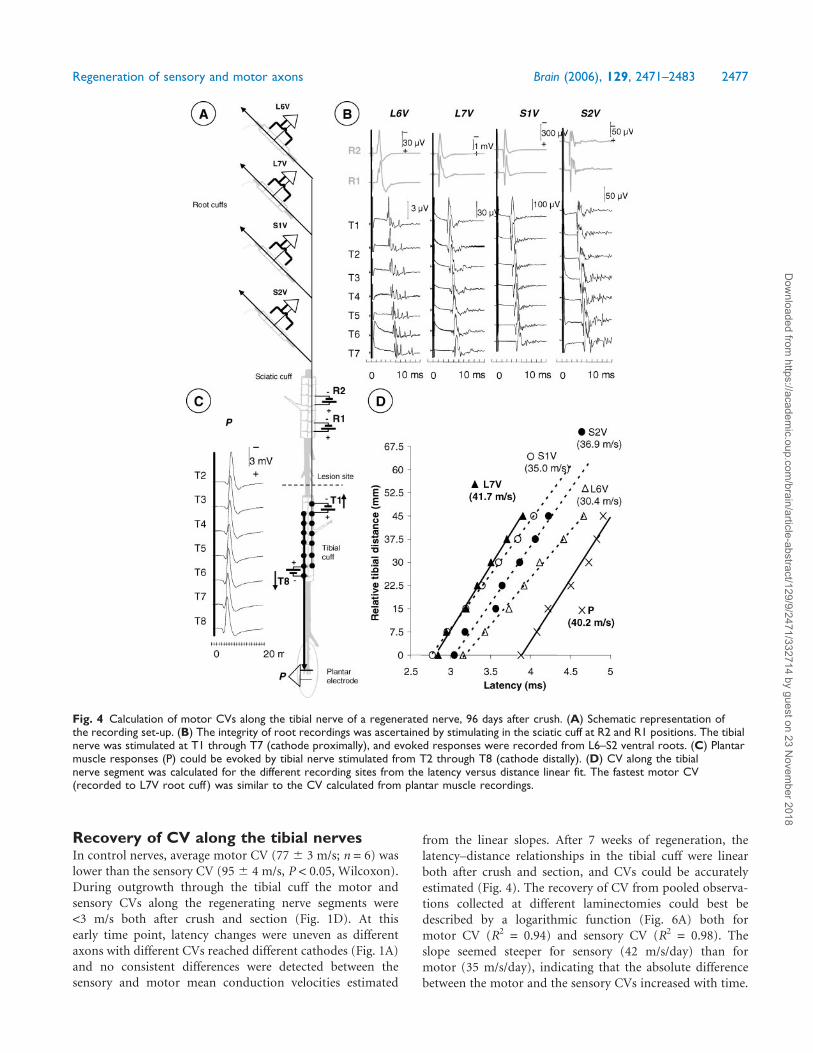
Fig. 4 Calculation of motor CVs along the tibial nerve of a regenerated nerve, 96 days after crush. (A) Schematic representation ofthe recording set-up. (B) The integrity of root recordings was ascertained by stimulating in the sciatic cuff at R2 and R1 positions. The tibialnerve was stimulated at T1 through T7 (cathode proximally), and evoked responses were recorded from L6–S2 ventral roots. (C) Plantarmuscle responses (P) could be evoked by tibial nerve stimulated from T2 through T8 (cathode distally). (D) CV along the tibialnerve segment was calculated for the different recording sites from the latency versus distance linear fit. The fastest motor CV(recorded to L7V root cuff ) was similar to the CV calculated from plantar muscle recordings.
Regeneration of sensory and motor axons Brain (2006), 129, 2471–2483 2477
Dow
nloaded from https://academ
ic.oup.com/brain/article-abstract/129/9/2471/332714 by guest on 23 N
ovember 2018
For example, at 52 days after crush, motor CV was 13 m/s and
sensory CV was 15 m/s, whereas at 139 days after crush motor
CV was 49 m/s and sensory CV was 56 m/s.
During post-reinnervation maturation we found that the
sensory CV was faster at any time point (Fig. 6A). Never-
theless, the relative recovery in conduction was similar in
motor and sensory axons. For example, at 52 days conduction
recovered �16% and at 139 days conduction recovered
�60% relative to the respective motor and sensory controls.
In fact, in all nerves investigated after target reinnervation we
found a high correlation (R2 = 0.98) between motor CVs and
the corresponding sensory CVs (Fig. 7A), indicating that
sensory CVs were �14% faster than the corresponding
motor CVs. This finding was also consistent in the sectioned
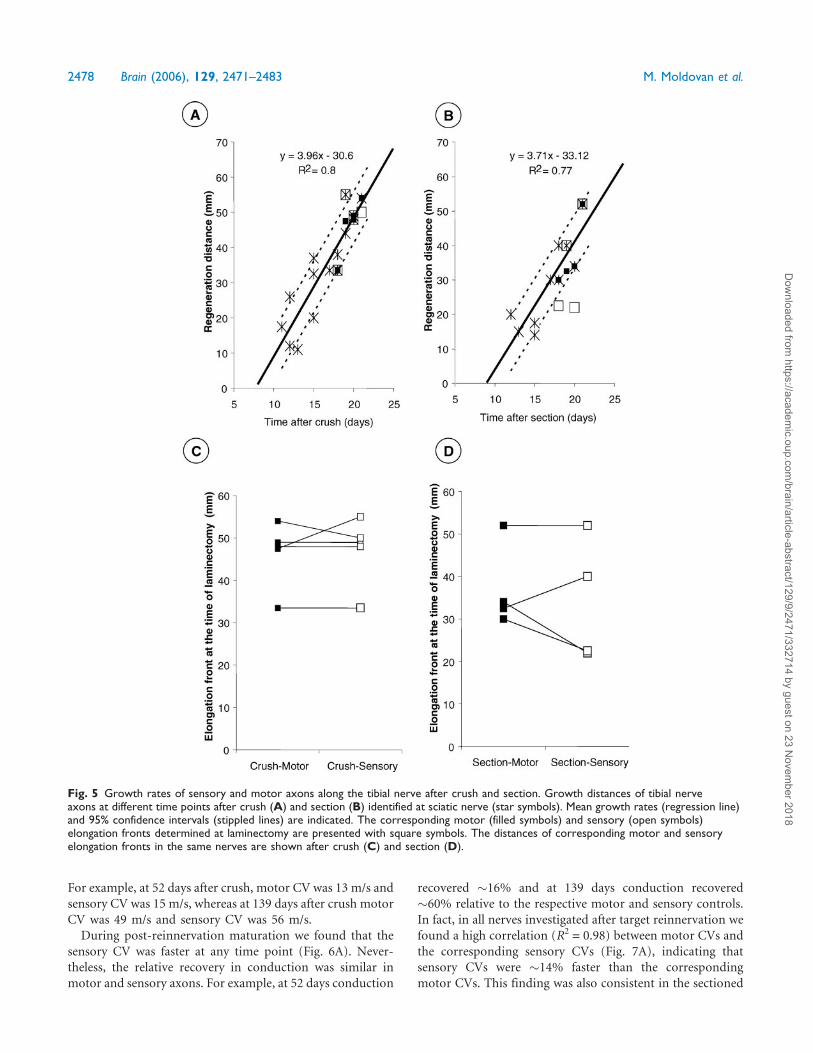
Fig. 5 Growth rates of sensory and motor axons along the tibial nerve after crush and section. Growth distances of tibial nerveaxons at different time points after crush (A) and section (B) identified at sciatic nerve (star symbols). Mean growth rates (regression line)and 95% confidence intervals (stippled lines) are indicated. The corresponding motor (filled symbols) and sensory (open symbols)elongation fronts determined at laminectomy are presented with square symbols. The distances of corresponding motor and sensoryelongation fronts in the same nerves are shown after crush (C) and section (D).
2478 Brain (2006), 129, 2471–2483 M. Moldovan et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article-abstract/129/9/2471/332714 by guest on 23 N
ovember 2018
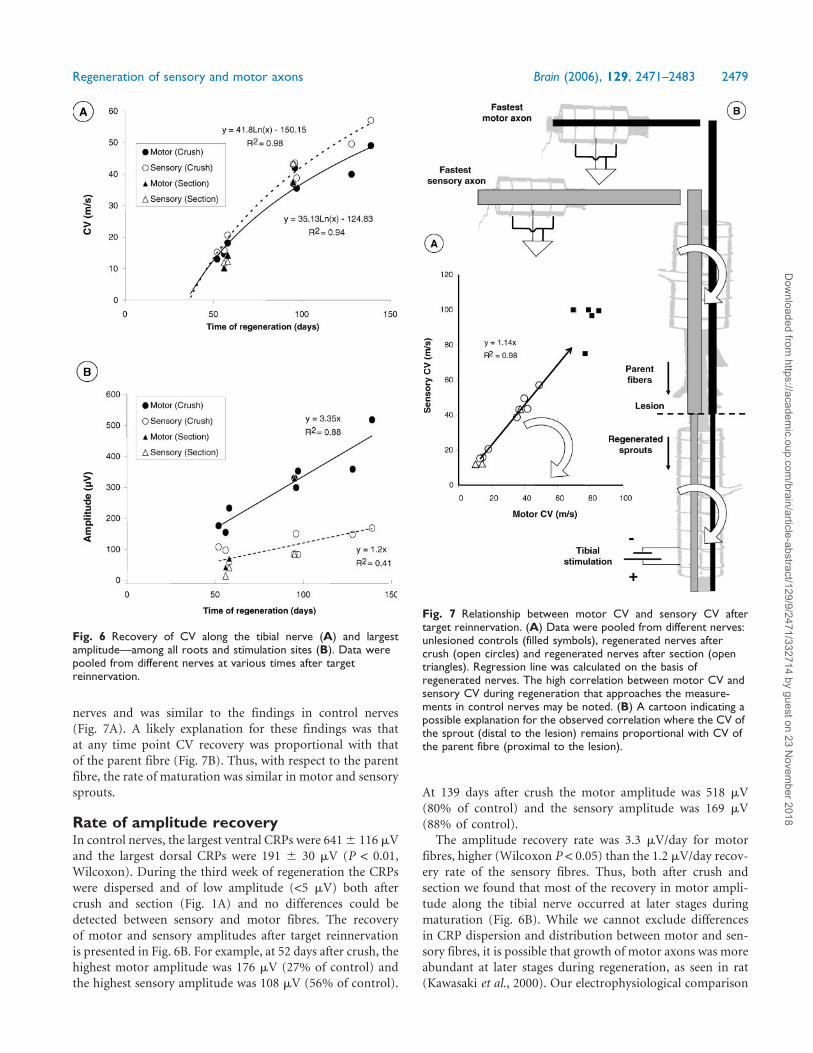
nerves and was similar to the findings in control nerves
(Fig. 7A). A likely explanation for these findings was that
at any time point CV recovery was proportional with that
of the parent fibre (Fig. 7B). Thus, with respect to the parent
fibre, the rate of maturation was similar in motor and sensory
sprouts.
Rate of amplitude recoveryIn control nerves, the largest ventral CRPs were 6416 116 mV
and the largest dorsal CRPs were 191 6 30 mV (P < 0.01,
Wilcoxon). During the third week of regeneration the CRPs
were dispersed and of low amplitude (<5 mV) both after
crush and section (Fig. 1A) and no differences could be
detected between sensory and motor fibres. The recovery
of motor and sensory amplitudes after target reinnervation
is presented in Fig. 6B. For example, at 52 days after crush, the
highest motor amplitude was 176 mV (27% of control) and
the highest sensory amplitude was 108 mV (56% of control).
At 139 days after crush the motor amplitude was 518 mV
(80% of control) and the sensory amplitude was 169 mV
(88% of control).
The amplitude recovery rate was 3.3 mV/day for motor
fibres, higher (Wilcoxon P < 0.05) than the 1.2 mV/day recov-
ery rate of the sensory fibres. Thus, both after crush and
section we found that most of the recovery in motor ampli-
tude along the tibial nerve occurred at later stages during
maturation (Fig. 6B). While we cannot exclude differences
in CRP dispersion and distribution between motor and sen-
sory fibres, it is possible that growth of motor axons was more
abundant at later stages during regeneration, as seen in rat
(Kawasaki et al., 2000). Our electrophysiological comparison
Fig. 6 Recovery of CV along the tibial nerve (A) and largestamplitude—among all roots and stimulation sites (B). Data werepooled from different nerves at various times after targetreinnervation.
Fig. 7 Relationship between motor CV and sensory CV aftertarget reinnervation. (A) Data were pooled from different nerves:unlesioned controls (filled symbols), regenerated nerves aftercrush (open circles) and regenerated nerves after section (opentriangles). Regression line was calculated on the basis ofregenerated nerves. The high correlation between motor CV andsensory CV during regeneration that approaches the measure-ments in control nerves may be noted. (B) A cartoon indicating apossible explanation for the observed correlation where the CV ofthe sprout (distal to the lesion) remains proportional with CV ofthe parent fibre (proximal to the lesion).
Regeneration of sensory and motor axons Brain (2006), 129, 2471–2483 2479
Dow
nloaded from https://academ
ic.oup.com/brain/article-abstract/129/9/2471/332714 by guest on 23 N
ovember 2018
was, however, aimed at the fastest regenerating motor and
sensory axons that were not influenced by such regenerative
differences within the motor and sensory fibre populations.
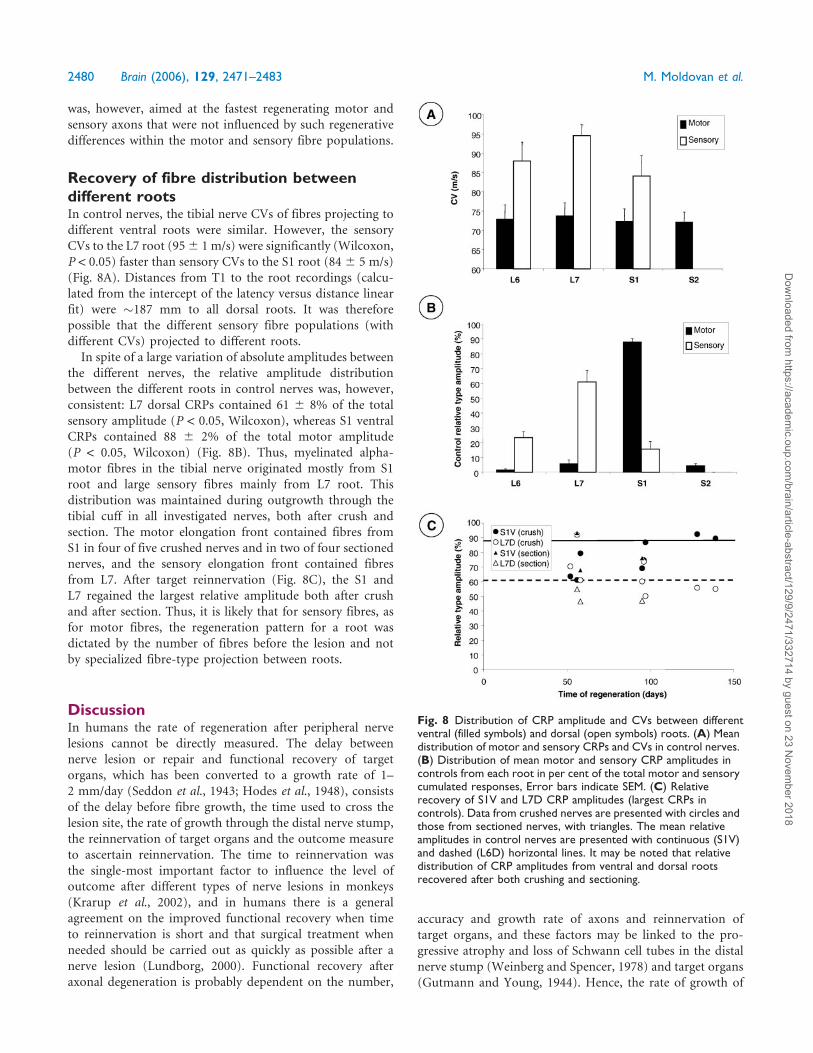
Recovery of fibre distribution betweendifferent rootsIn control nerves, the tibial nerve CVs of fibres projecting to
different ventral roots were similar. However, the sensory
CVs to the L7 root (956 1 m/s) were significantly (Wilcoxon,
P < 0.05) faster than sensory CVs to the S1 root (846 5 m/s)
(Fig. 8A). Distances from T1 to the root recordings (calcu-
lated from the intercept of the latency versus distance linear
fit) were �187 mm to all dorsal roots. It was therefore
possible that the different sensory fibre populations (with
different CVs) projected to different roots.
In spite of a large variation of absolute amplitudes between
the different nerves, the relative amplitude distribution
between the different roots in control nerves was, however,
consistent: L7 dorsal CRPs contained 61 6 8% of the total
sensory amplitude (P < 0.05, Wilcoxon), whereas S1 ventral
CRPs contained 88 6 2% of the total motor amplitude
(P < 0.05, Wilcoxon) (Fig. 8B). Thus, myelinated alpha-
motor fibres in the tibial nerve originated mostly from S1
root and large sensory fibres mainly from L7 root. This
distribution was maintained during outgrowth through the
tibial cuff in all investigated nerves, both after crush and
section. The motor elongation front contained fibres from
S1 in four of five crushed nerves and in two of four sectioned
nerves, and the sensory elongation front contained fibres
from L7. After target reinnervation (Fig. 8C), the S1 and
L7 regained the largest relative amplitude both after crush
and after section. Thus, it is likely that for sensory fibres, as
for motor fibres, the regeneration pattern for a root was
dictated by the number of fibres before the lesion and not
by specialized fibre-type projection between roots.
DiscussionIn humans the rate of regeneration after peripheral nerve
lesions cannot be directly measured. The delay between
nerve lesion or repair and functional recovery of target
organs, which has been converted to a growth rate of 1–
2 mm/day (Seddon et al., 1943; Hodes et al., 1948), consists
of the delay before fibre growth, the time used to cross the
lesion site, the rate of growth through the distal nerve stump,
the reinnervation of target organs and the outcome measure
to ascertain reinnervation. The time to reinnervation was
the single-most important factor to influence the level of
outcome after different types of nerve lesions in monkeys
(Krarup et al., 2002), and in humans there is a general
agreement on the improved functional recovery when time
to reinnervation is short and that surgical treatment when
needed should be carried out as quickly as possible after a
nerve lesion (Lundborg, 2000). Functional recovery after
axonal degeneration is probably dependent on the number,
accuracy and growth rate of axons and reinnervation of
target organs, and these factors may be linked to the pro-
gressive atrophy and loss of Schwann cell tubes in the distal
nerve stump (Weinberg and Spencer, 1978) and target organs
(Gutmann and Young, 1944). Hence, the rate of growth of
Fig. 8 Distribution of CRP amplitude and CVs between differentventral (filled symbols) and dorsal (open symbols) roots. (A) Meandistribution of motor and sensory CRPs and CVs in control nerves.(B) Distribution of mean motor and sensory CRP amplitudes incontrols from each root in per cent of the total motor and sensorycumulated responses, Error bars indicate SEM. (C) Relativerecovery of S1V and L7D CRP amplitudes (largest CRPs incontrols). Data from crushed nerves are presented with circles andthose from sectioned nerves, with triangles. The mean relativeamplitudes in control nerves are presented with continuous (S1V)and dashed (L6D) horizontal lines. It may be noted that relativedistribution of CRP amplitudes from ventral and dorsal rootsrecovered after both crushing and sectioning.
2480 Brain (2006), 129, 2471–2483 M. Moldovan et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article-abstract/129/9/2471/332714 by guest on 23 N
ovember 2018

motor and sensory axons may have direct implications
on functional recovery. Since the reinnervation of sensory
receptors is considerably more difficult to ascertain than
muscle fibre reinnervation in humans and non-human
primates owing to the small sensory nerve and receptor
responses (Buchthal and Kuhl, 1979; Krarup et al., 1990,
2002), direct comparison of motor and sensory regeneration
rates is inaccurate. We therefore extended a method to
monitor growth of axons during regeneration after Wallerian
degeneration to directly compare elongation and maturation
of motor and sensory fibres within the same mixed tibial
nerve. Both after crush and section we found that similar
growth and maturation rates were attained by myelinated
motor and sensory fibres.
Growth of sensory and motor axonsWe could not directly measure the rate of growth of sensory
and motor axons in serial studies. We compared instead
the distances reached by the motor and sensory fibres at
specified times during regeneration in different nerves by
stimulating in the tibial cuff and recording from the L6–S2
dorsal and ventral roots. The spatial and temporal resolutions
of our method were determined by the length of the tibial
cuff, the inter-electrode spacing and the intervals between
studies. During serial studies we found that fastest outgrowth
started after �8 days and progressed at a rate of �4 mm/day,
which was largely in agreement with our previous observa-
tions (Fugleholm et al., 1994). Under these circumstances, to
discriminate a distance difference of one stimulation level
(7.5 mm) after 20 days of regeneration, the difference in
growth rates should have been >0.5 mm/day (4 versus
3.5 mm/day).
We found that the fastest growth rates achieved by
myelinated fibres were similar in motor and sensory fibres
populations and fell within the confidence intervals of the
mixed nerve growth rates (Fig. 5). This indicated that
growth rates of motor and sensory fibres were similar within
0.5 mm/day accuracy. While it was not possible to increase
the length of the cuff owing to anatomical constraints, one
would expect that an even greater accuracy could have been
obtained by reducing the inter-electrode distance. We
previously verified by electron microscopy that the 10 mA
stimulation current required to excite the immature axons
could accurately predict the growth front within 7.5 mm
resolution (Fugleholm et al., 1994). By reducing inter-
electrode distance, however, stimulus spread could have
been a source of error.
Axons contributing to the growth frontWe stimulated the immature axons at the growth front in
the tibial and recorded the evoked responses in cuff electro-
des. Our electrophysiological approach was constrained by
the methodological limitation that action potentials could be
recorded from single myelinated fibres with diameters larger
than 5–7 mm (Rindos et al., 1984; Krarup and Loeb, 1988).
Thus, our method allowed us to investigate alpha-
motor fibres but was limited to only the larger sensory fibres.
A functional study in rat showed that responses from
nociceptive and sudomotor fibres occur slightly earlier and
to a greater extent than recordings of compound muscle and
nerve action potentials from large myelinated fibres (Navarro
et al., 1994). It is, however, unlikely that small sensory fibres
regenerate strikingly faster than large sensory fibres since a
recent retrograde tracer study showed that regeneration of
unmyelinated sensory fibres and large sensory fibres in rat
had similar rates (Lozeron et al., 2004).
However contraintuitive, our study did not support
the assumption that the largest fibres have the fastest growth
rate. While our method favoured detection of regeneration of
the largest fibres with the largest action potential amplitudes,
we found evidence that within the respective sensory and
motor fibre populations it was not the largest and fastest
conducting fibres that grew first (Figs 1E, 2 and 8A), and
that motor and sensory fibres made similar contributions to
the growth front (Fig. 5). It may therefore be speculated that
growth rates were maximized for an ‘optimal’ relationship
between radial growth and longitudinal growth, which is
similar in sensory and motor axons.
Maturation of sensory and motor axonsMaturation (the increase in axonal calibre and myelination)
occurs slower (Wujek et al., 1986; Zhu et al., 1997) and
independently (Fried et al., 1991; Macias et al., 1998) of
growth. In regenerating cat axons (Fugleholm et al., 2000)
CV remains proportional with the fibre diameter and we
could compare maturation of sensory and motor axons
by investigating recovery in CV. After target reinnervation,
the change in latency along the tibial cuff was linear and we
could calculate the average CV along the tibial nerve as the
latency versus distance linear fit and then determine the
fastest motor and sensory CV among the different roots.
This method of calculation was validated by comparing
the motor CV estimated to the roots with the CV to the
plantar muscles. Furthermore, this method of calculation
was able to detect the slightly faster sensory than motor
CV corresponding to the differences in largest diameters of
sensory and motor fibres (Hursh, 1939).
We found that at various times during maturation, the
sensory CV was slightly faster than the motor CV (Fig. 6A).
This did not indicate that sensory maturation occurred at a
faster rate, because a similar difference in CV was also seen in
control nerves, and the ratio between sensory and motor CV
in control nerves was preserved during regeneration. It was
more likely that maturation occurred at a rate proportional to
the diameter of the parent fibre (Wujek et al., 1986). Further-
more, early studies in rat sensory fibres showed that CV of the
sprout was proportional with CV of its parent fibre (Devor
and Govrin-Lippmann, 1979). Thus, in respect to the parent
fibre diameters, maturation rate was similar between sensory
and motor fibres.
Regeneration of sensory and motor axons Brain (2006), 129, 2471–2483 2481
Dow
nloaded from https://academ
ic.oup.com/brain/article-abstract/129/9/2471/332714 by guest on 23 N
ovember 2018

Crush versus sectionWhile the theoretical rate-limiting factor of axonal regenera-
tion may be the speed of up to 6.5 mm/day at which slow
axonal transport moves cytoskeletal elements involved in the
motility of the growth cone and elongation of the axon
(Wujek and Lasek, 1983), the linear growth rate is much
slower, dependent on the growth cone environment
(Fugleholm et al., 1994, 1998, 2000; Sorensen et al., 2001).
The fastest rates of regeneration could be attained after crush,
where growth cones move along their original Schwann
cell tubes. We found that in these ideal conditions the
growth rates were similar between motor and sensory fibres.
In the clinical situation of a co-adapted nerve section, regen-
erating fibres are more likely to grow through unfamiliar
pathways that may impair the growth rates. In the mouse
motor axons preferentially reinnervated motor pathways
(Brushart, 1988) after section. It was therefore possible
that such environmental specialization may favour the
growth of motor fibres. Our data suggests that after section,
as after crush, the fastest growth rates that could be attained
by motor and sensory fibres were similar. Thus, even in the
clinically relevant regenerative environment some motor
and sensory axons retain similar growth capacity and will
simultaneously compete for target selection.
AcknowledgementsThe project was supported by Lundbeck Foundation, the
Novo Nordisk Foundation, the Danish Medical Research
Council, the Ludvig and Sarah Elsass Foundation and the
Foundation for Research in Neurology.
ReferencesBailey CS, Kitchell RL, Guinan MJ, Sharp JW. Dorsal nerve root origins of
the cutaneous nerves of the feline pelvic limb. Anat Histol Embryol 1992;
21: 23–31.
Braendgaard H, Sidenius P. Anterograde components of axonal transport in
motor and sensory nerves in experimental 2,5–hexanedione neuropathy.
J Neurochem 1986; 47: 31–7.
Brushart TM. Preferential reinnervation of motor nerves by regenerating
motor axons. J Neurosci 1988; 8: 1026–31.
Brushart TM. Motor axons preferentially reinnervate motor pathways.
J Neurosci 1993; 13: 2730–8.
Buchthal F, Kuhl V. Nerve conduction, tactile sensibility, and the
electromyogram after suture or compression of peripheral nerve: a
longitudinal study in man. J Neurol Neurosurg Psychiatry 1979; 42:
436–51.
Chen S, Bisby MA. Impaired motor axon regeneration in the C57BL/Ola
mouse. J Comp Neurol 1993; 333: 449–54.
da Silva CF, Madison R, Dikkes P, Chiu TH, Sidman RL. An in vivo model
to quantify motor and sensory peripheral nerve regeneration using
bioresorbable nerve guide tubes. Brain Res 1985; 342: 307–15.
Devor M, Govrin-Lippmann R. Maturation of axonal sprouts after nerve
crush. Exp Neurol 1979; 64: 260–70.
Forman DS, Berenberg RA. Regeneration of motor axons in the rat sciatic
nerve studied by labeling with axonally transported radioactive proteins.
Brain Res 1978; 156: 213–25.
Forman DS, Wood DK, DeSilva S. Rate of regeneration of sensory
axons in transected rat sciatic nerve repaired with epineurial sutures.
J Neurol Sci 1979; 44: 55–9.
Fried K, Govrin-Lippmann R, Rosenthal F, Ellisman MH, Devor M.
Ultrastructure of afferent axon endings in a neuroma. J Neurocytol 1991;
20: 682–701.
Fugleholm K, Schmalbruch H, Krarup C. Early peripheral nerve regenera-
tion after crushing, sectioning, and freeze studied by implanted electrodes
in the cat. J Neurosci 1994; 14: 2659–73.
Fugleholm K, Sorensen J, Schmalbruch H, Krarup C. Axonal elongation
through acellular nerve segments of the cat tibial nerve: importance of
the near-nerve environment. Brain Res 1998; 792: 309–18.
Fugleholm K, Schmalbruch H, Krarup C. Post reinnervation maturation of
myelinated nerve fibers in the cat tibial nerve: chronic electrophysiological
and morphometric studies. J Peripher Nerv Syst 2000; 5: 82–95.
Gutmann E, Young JZ. The re-innervation of muscle after various periods
of atrophy. J Anat 1944; 78: 15–44.
Highland TR, Ottoni L, Lamont RL, Goshgarian HG. The contribution of
the dorsal and ventral roots of the tibial nerve in the propagation of the
spinal somatosensory evoked potential in cats. Spine 1990; 15: 560–6.
Hodes R, Larrabee MG, German W. The human electromyogram in
response to nerve stimulation and the conduction velocity of motor axons.
Arch Neurol Psychiat 1948; 60: 340–65.
Hursh JB. Conduction velocity and diameter of nerve fibers. Am J Physiol
1939; 127: 131–9.
Jenq CB, Coggeshall RE. Numbers of regenerated axons in tributary
nerves following neonatal sciatic nerve crush in rat. Neurosci Lett 1985;
61: 43–8.
Kawasaki Y, Yoshimura K, Harii K, Park S. Identification of myelinatedmotor
and sensory axons in a regenerating mixed nerve. J Hand Surg [Am] 2000;
25: 104–11.
Krarup C, Loeb GE. Conduction studies in peripheral cat nerve using
implanted electrodes: I. Methods and findings in controls. Muscle Nerve
1988; 11: 922–32.
Krarup C, Loeb GE, Pezeshkpour GH. Conduction studies in peripheral cat
nerve using implanted electrodes: III. The effects of prolonged constriction
on the distal nerve segment. Muscle Nerve 1989; 12: 915–28.
Krarup C, Upton J, Creager MA. Nerve regeneration and reinnervation after
limb amputation and replantation: clinical and physiological findings.
Muscle Nerve 1990; 13: 291–304.
Krarup C, Archibald SJ, Madison RD. Factors that influence peripheral nerve
regeneration: an electrophysiological study of the monkey median nerve.
Ann Neurol 2002; 51: 69–81.
Larsen JO, Thomsen M, Haugland M, Sinkjaer T. Degeneration and
regeneration in rabbit peripheral nerve with long-term nerve cuff electrode
implant: a stereological study of myelinated and unmyelinated axons. Acta
Neuropathol (Berl) 1998; 96: 365–78.
Lozeron P, Krarup C, Schmalbruch H. Regeneration of unmyelinated and
myelinated sensory nerve fibres studied by a retrograde tracer method.
J Neurosci Methods 2004; 138: 225–32.
Lundborg G. A 25-year perspective of peripheral nerve surgery: evolving
neuroscientific concepts and clinical significance. J Hand Surg [Am] 2000;
25: 391–414.
Lundborg G, Richard P. Bunge memorial lecture. Nerve injury and repair—a
challenge to the plastic brain. J Peripher Nerv Syst 2003; 8: 209–226.
Macias MY, Lehman CT, Sanger JR, Riley DA. Myelinated sensory and alpha
motor axon regeneration in peripheral nerve neuromas. Muscle Nerve
1998; 21: 1748–58.
Madison RD, da Silva CF, Dikkes P. Entubulation repair with protein
additives increases the maximum nerve gap distance successfully bridged
with tubular prostheses. Brain Res 1988; 447: 325–34.
Madorsky SJ, Swett JE, Crumley RL. Motor versus sensory neuron
regeneration through collagen tubules. Plast Reconstr Surg 1998; 102:
430–36.
Martini R, Xin Y, Schmitz B, Schachner M. The L2/HNK-1 carbohydrate
epitope is involved in the preferential outgrowth of motor neurons on
ventral roots and motor nerves. Eur J Neurosci 1992; 4: 628–39.
Navarro X, Verdu E, Buti M. Comparison of regenerative and reinnervating
capabilities of different functional types of nerve fibers. Exp Neurol 1994;
129: 217–24.
2482 Brain (2006), 129, 2471–2483 M. Moldovan et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article-abstract/129/9/2471/332714 by guest on 23 N
ovember 2018

Rindos AJ, Loeb GE, Levitan H. Conduction velocity changes along lumbar
primary afferent fibers in cats. Exp Neurol 1984; 86: 208–26.
Sanger JR, Riley DA, Matloub HS, Yousif NJ, Bain JL, Moore GH. Effects
of axotomy on the cholinesterase and carbonic anhydrase activities of
axons in the proximal and distal stumps of rabbit sciatic nerves: a temporal
study. Plast Reconstr Surg 1991; 87: 726–38.
Seddon HJ, Medawar PB, Smith H. Rate of regeneration of peripheral
nerves in man. J Physiol 1943; 102: 191–215.
Sorensen J, Fugleholm K, Moldovan M, Schmalbruch H, Krarup C.
Axona elongation through long acellular nerve segments depends on
recruitment of phagocytic cells from the near-nerve environment.
Electrophysiological and morphological studies in the cat. Brain Res
2001; 903: 185–97.
Suzuki G, Ochi M, Shu N, Uchio Y, Matsuura Y. Sensory neurons regenerate
more dominantly than motoneurons during the initial stage of the reg-
enerating process after peripheral axotomy. Neuroreport 1998; 9: 3487–92.
Weinberg HJ, Spencer PS. The fate of Schwann cells isolated from axonal
contact. J Neurocytol 1978; 7: 555–69.
Wujek JR, Lasek RJ. Correlation of axonal regeneration and slow
component B in two branches of a single axon. J Neurosci 1983; 3: 243–51.
Wujek JR, Lasek RJ, Gambetti P. The amount of slow axonal transport is
proportional to the radial dimensions of the axon. J Neurocytol 1986;
15: 75–83.
Zhu Q, Couillard-Despres S, Julien JP. Delayed maturation of regenerating
myelinated axons in mice lacking neurofilaments. Exp Neurol 1997;
148: 299–316.
Regeneration of sensory and motor axons Brain (2006), 129, 2471–2483 2483
Dow
nloaded from https://academ
ic.oup.com/brain/article-abstract/129/9/2471/332714 by guest on 23 N
ovember 2018





























