Embed Size (px)
Citation preview

Copyright 0 1994 by the Genetics Society of America
Complete DNA Sequence of the Mitochondrial Genome of the Black Chiton, Katharina tunicata
Jeffrey L. Boore' and Wesley M. Brown
Department of Biology and Museum of Zoology, University of Michigan, Ann Arbor, Michigan 48109-1048 Manuscript received March 10, 1994 Accepted for publication July 8, 1994
ABSTRACT The DNA sequence of the 15,532-base pair (bp) mitochondrial DNA (mtDNA) of the chiton Katharina
tunicata has been determined. The 37 genes typical of metazoan mtDNA are present: 13 for protein subunits involved in oxidative phosphorylation, 2 for rRNAs and 22 for t R N A s . The gene arrangement resembles those of arthropods much more than that of another mollusc, the bivalve Mytilus edulis. Most genes abut directly or overlap, and abbreviated stop codons are inferred for four genes. Four junctions between adjacent pairs of protein genes lack intervening tRNA genes; however, at each of these junctions there is a sequence immediately adjacent to the start codon of the downstream gene that is capable of forming a stem-and-loop structure. Analysis of the tRNA gene sequences suggests that the D arm is un- paired in tRNASer(AGN), which is typical of metazoan mtDNAs, and also in tRNAxr(UCN), a condition found previously only in nematode mtDNAs. There are two additional sequences in Katharina mtDNA that can be folded into structures resembling tRNAs; whether these are functional genes is unknown. All possible codons except the stop codons TAA and TAG are used in the protein-encoding genes, and Katharina mtDNA appears to use the same variation of the mitochondrial genetic code that is used in Drosophila and Mytilus. Translation initiates at the codons ATG, ATA and GTG. A + T richness appears to have affected codon usage patterns and, perhaps, the amino acid composition of the encoded proteins. A 142-bp non-coding region between tRNA@'" and C03 contains a 72-bp tract of alternating A and T.
M ETAZOAN mitochondrial DNA (mtDNA) is a closed-circular molecule, except in some hydro-
zoan cnidarians where it is one or two linear molecules (WARRIOR and GALL 1985; BRIDGE et al. 1992). Metazoan mtDNAs vary in size from ca. 14-42 kilobases (kb) (MORITZ et al. 1987; WOLSTENHOLME 1992; SNYDER et al. 1987). Usually this variation is due to differences in non- coding regions (BROWN 1985; HARRISON 1989), but oc- casionally it is due to duplications or multiplications of coding regions (AZEVEDO and HW 1993; FULLER and ZOUROS 1993; MORITZ and BROWN 1986, 1987; ZEVERING et al. 1991).
The gene content of metazoan mtDNA is highly con- served, and typically consists of genes for 2 ribosomal subunit RNAs [small- and large-rRNA (+rRNA and 1-rRNA) 1, for 22 tRNAs, and for 13 protein subunits [cy- tochrome c oxidase subunits 1-111 (CO1-3), cytochrome b apoenzyme (Cytb), ATP synthase subunits 6 and 8 (ATPase6 and ATPase8), and NADH dehydrogenase subunits 1-6 and 4L (NDl-6, 4L)I. In addition, there is at least one sequence ofvariable length which does not encode any structural genes and which, in vertebrates (BOGENHAGEN et al. 1985; CLAWON 1991, 1992; FORAN et al. 1988; KING and LOW 1987; MONTOYA et al. 1982) and insects (CLARY and WOLSTENHOLME 1985a), has been
Present address: Department of Cell Biology and Neuroanatomy, Univer- sity of Minnesota, 4135 Church Street SE, Minneapolis, Minnesota 55455.
Genetics 138: 423-443 (October, 1994)
shown to include elements for the initiation and control of replication and transcription.
Mitochondrial gene arrangements are usually quite similar among animals within the same phylum. All 37 genes are arranged in the same relative order among most vertebrates (ANDERSON et al. 1981, 1982; ARNASON
et al. 1991; &ASON and J ~ H N S S ~ N 1992; BIBB et al. 1981; CHANG et al. 1994; GADALETA et al. 1989; ROE et al. 1985;
JOHANSEN et al. 1990; TZENG et al. 1992), although minor rearrangements are found in marsupials (Pmo et al. 1991) and birds (DESJARDINS and MORAIS 1990; DESJARDINS et al. 1990). Likewise, gene arrangements are very similar among echinoderms, with only a single in- version differentiating sea urchins from sea stars UACOBS et al. 1988; CANTATORE et al. 1987b, 1989; DE GIORGI et al. 1991; HIMENO et al. 1987; SMITH et al. 1989,1990,1993). Within arthropods, the mitochondrial gene arrange- ments of Drosophila (CLARY and WOLSTENHOLME 1985a; DE BRUIJN 1983; GARESSE 1988) and Apis (CROZIER and CROZIER 1993), and partial gene arrangements of others (BATUCAS et al. 1988; DUBIN et al. 1986; HSUCHEN et al. 1984; MCCRACKEN et al. 1987; PASHLEY and KE 1992; UHLENBUSCH et al. 1987; L. DAEHLER, D. STANTON and W. BROWN, unpublished data) differ only by one to a few tRNA transpositions. The mtDNAs of two nematodes, Caenorhabditis elegans and Ascaris mum, have nearly identical gene arrangements (WOLSTENHOLME et al. 1987; OKIMOTO et al. 1992), although that of a third,

424 J. L. Boore and W. M. Brown
Meloidogyne javanica, is radically different (OKIMOTO et al. 1991). Nematodes are also unusual in lacking a mitochondrially encoded ATPuse8. In contrast to the general similarity of gene arrangements within a phy- lum, there are substantial differences among phyla.
Comparison of the relative arrangements of mito- chondrial genes may be useful for evaluating phyloge- netic relationships among major metazoan groups (see BOORE and BROWN 1994). The gene content of metazoan mtDNAs is nearly unvarying, so a comparable data set is available for all taxa. Mitochondrial genomes appear to undergo rearrangement on a time scale appropriate for resolving ancient phylogenetic relationships. The great number of theoretically possible gene arrangements makes it unlikely that two or more taxa will indepen- dently converge on an identical order, thus, identical gene arrangements will generally be shared only by com- mon ancestry. Comparisons of mitochondrial gene ar- rangements have been useful so far in addressing the evolutionary relationships among echinoderm classes (SMITH et al. 1993). As this data set expands, we antici- pate that mitochondrial gene arrangements, as well as many aspects of the molecular biology of mtDNA ( i. e . , alternative start codons, unusual tRNA and rRNA struc- tures, genetic code variations, and features of mtDNA replication and transcription) will reveal patterns that signal the evolutionary relationships of many major metazoan groups.
Mytilus edulis is the first mollusc whose mitochon- drial gene arrangement was determined (HOFFMANN et al. 1992). That arrangement is radically different from any previously reported. Mytilus mtDNA, like that of nematodes, lacks A TPase8, and it has several other atypi- cal features, including a second tRNAmet gene, several lengthy unassigned intergenic sequences, and signifi- cant departure in the size of several of its protein- encoding genes from those of other metazoans. It was unclear whether these features of Mytilus mtDNA, which belongs to the class Bivalvia, are typical of molluscs, or of a more phylogenetically restricted group. To evaluate this, to add to the rapidly growing database of mitochon- drial gene arrangements, and to detail aspects of the molecular biology of mtDNA, many of which promise to be useful for evolutionary comparisons, we determined the complete mtDNA sequence for the chiton Katha- rina tunicata, a representative of the molluscan class Polyplacophora.
MATERIALS AND METHODS
Mitochondrial DNA from the chiton K. tunicata was iso- lated and purified as described and referenced in WRIGHT et al. (1983), but with a twofold increased concentration of ethidium bromide substituted for propidium iodide. A de- tailed map of the cleavage sites for 12 restriction endonucle- ases was constructed, using a combination of single and double digests (see Figure 1). DNA fragments resulting from the di- gests were labeled with [32P]dNTP~ (Amersham Corp.) using the Klenow fragment of DNA polymerase (Boehringer Mann-
heim) according to the protocol in SAMBROOK et al. (1989); after labeling, digests were split and subjected to electrophore- sis in 1% agarose and 3.5% polyacrylamide gels, using condi- tions (BROWN 1980) that allowed detection of fragments as small as 40 base pairs (bp) in the polyacrylamide gels.
Katharina mtDNA was cloned into the A bacteriophage vec- tor EMBL3 (Stratagene) as a single insert, using the unique BamHI site at position 669-674 (see APPENDIX). Approximately 50 ng of digested mtDNA and 1 pg of EMBLS arms were mixed with DNA ligase and incubated overnight at 14" (SAMBROOK et al. 1989). The ligation mix was added to a packaging system (Stratagene) and used to transfect a culture of Escherichia coli strain P2-392. After initially screening for insert size, plaque- purified recombinant phage were tested for the presence of the entire mtDNA insert by restriction enzyme fragment com- parisons with Katharina mtDNA.
Five insertcontaining DNA fragments, of approximately 7.0,3.9,3.1,1.1 and 0.7 kilobases (kb), were produced by com- bined digestion of the recombinant DNA with BamHI and EcoRI. The fragments were separated by electrophoresis in 0.8% agarose, excised from the gel, and extracted and purified using Geneclean (Bio 101, Inc.) according to supplier's in- structions. A 50- to 500-ng sample of each fragment was ligated to an equimolar amount of the plasmid vector pBluescript I1 KS- (Stratagene) which had been digested with BamHl and EcoR1, or with EcoRI alone. The recombinant plasmids, des- ignated K7.0, K3.9, K3.1, K1.l and K0.7, respectively, were transformed into the XL1 Blue strain of E . coli, which had been made transformationcompetent by treatment with poly- ethylene glycol (CHUNG and MILLER 1988). The cleavage sites of 20 restriction enzymes were mapped in the inserts, and fur- ther subcloning yielded a total of 41 additional clones (see Figure 1). White, ampicillin-resistant colonies were selected and the identities of the inserts in the recombinant plasmids were verified by comparing restriction enzyme maps of puri- fied plasmid DNA with those of native mtDNA.
DNA sequences were determined according to procedures adapted from SANCER et al. (1977), using modified T7 DNA polymerase (Sequenase 2.0, U. S. Biochemical Corp.) and [35S]dATP (Amersham) . Double-stranded sequencing tem- plates were prepared according to DEL SAL et al. (1988), LISZEWSKI et al. (1989), or the CsCl banding technique of SAMBROOK et al. (1989). Some clones were sequenced from single-stranded templates generated according to SAMBROOK et al. (1989). Products of sequencing reactions were separated by electrophoresis in 4% or 6% polyacrylamide gels; dITP was used, when necessary, to resolve compressions. The sequence of ca. 400 nucleotides (nt) from each end of an insert could usually be determined using the T7 or T3 priming sites in pBluescript 11; additional sequence was obtained using oligo- nucleotide primers of 17 nt, designed on the basis of the se- quence obtained (see Figure 1).
Open reading frames (ORFs) were identified with the pro- gram IBI MacVector (version 3.5), using the genetic code in- ferred for Drosophila mtDNA (CLARY and WOLSTENHOLME 1985a). The identity of the ORFs was determined by the simi- larity of their inferred amino acid sequences to those of the corresponding mitochondrial genes of Drosophila yakuba (CLARY and WOLSTENHOLME 1985a). In the case of ND4L, ND6 andATPase8, which show only weak amino acid sequence simi- larity between Katharina and Drosophila, identities were con- firmed by comparisons of hydrophilicity profiles (KYTE and DOOLITTLE 1982; see Figure 3). Sequences were identified ge- nerically as tRNA genes by their potential to be folded into secondary structures characteristic of mt-tRNAs, and specifi- cally by the triplet sequence in their putative anticodon loop. The rRNA genes were identified by their similarity in sequence

Chiton Mitochondrial Genome
Katharina tunicata
425
d -/
F W ) ? S 7 E H T P u y ME€ KRI S(AGN)
D m LfQM ! a G A N
(K7.0) 1 K0.7 I K3.1 I K1.l I K3.9 K7.0 < > < - < - ___ < - - < - < -
- > >
>
~" " - - - - - ___ _ _ " - - - - " ___- "
" - "
"7 7 ""2 -7 7 " " r 7 7 - 77
7 - 3 - "" 7 " " 7 - "" 7
c" " 2 L
L
" L 2 -7" " "- 2 "- 7"
7
I I IB TCKE X T I W WREI R W H PRlA KSEX H H H i H S I S S R R H t W X I C I S X J I l l Ill I I I I II 1 1 1 1 I l l I I I 1 1 1 1 I I II I I I I I I I1 II u 1 1 1 1 I II I I
5000 1 0 0 0 0 15000
FIGURE 1.-Mitochondrial DNA of the chiton K. tunicata, showing subcloning and sequencing strategies. The circular mtDNA map has been linearized between CO1 and ND2. Genes are designated by their products: CO1-3, subunits 1-111 of cytochrome c oxidase; ND1-6 and ND4L, subunits 1-6 and 4L of NADH dehydrogenase; CYTB, cytochrome b apoenzyme; A6 and A8, subunits 6 and 8 of the mitochondrial ATP synthase; s-rRNA and 1-rRNA, small and large ribosomal R N A s , respectively; tRNA genes are identified by the single letter amino acid code; F(UUU)? and S(UCU)?, tRNA genelike sequences (see text and Figure 5); UNK (unknown), the longest region of unassigned DNA (424 bp); A/T, a 72-nt tract of alternating As and Ts. All genes are transcribed from left to right, except those shown under the arrows (they are underlined for emphasis). K0.7, K3.1, K1.l, K3.9 and K7.0 are the five DNA fragments subcloned from the original recombinant EMBLS bacteriophage; vertical bars show their boundaries. The horizontal bars immediately below the K clones depict further subclones. The symbols < and > at the extreme right and left ends of the lines of subclones indicate that the adjacent subclone spans the arbitrary break made to linearize the circular map. Sequences obtained using the pBluescript T3 or T7 primers are depicted by arrows with downward facing barbs; arrows with upward facing barbs depict those obtained using primers designed from the Katharina sequence. Sites of cleavage by restriction enzymes employed in cloning are indicated by short vertical bars above the scale bar at the bottom. Restriction enzyme abbreviations: A, SacII; B, BamHI;C, CZaI; E, EcoRI; H, HindIII; I, HindII; K, KpnI; N, NheI; P, PstI; R, EcoRV; S, SpeI; T, Sad; X, XbaI. The BamHI site at position 669-674 (see APPENDIX) was used to insert the entire genome into the bacteriophage vector EMBL3. The scale, shown at the bottom, is in nucleotides.
and in potential secondary structure to their D. yakuba coun- terparts (CLARY and WOLSTENHOLME 1985b).
RESULTS AND DISCUSSION
Genome composition: The size of the cloned Katha- rina mtDNA is 15,532 bp. As is typical of metazoan mtDNAs, most intergenic regions are small (<lo bp) or absent, and there are no introns. Thirty-one of the genes abut directly, and seven have small (1-8 nt) overlaps.
The nucleotidesA and T make up 69% of the genome, less than for Drosophila mtDNA (78.6%; CLARY and WOLSTENHOLME 1985a), much less than for Apis mtDNA (84.9%; CROZIER and CROZIER 1993), but somewhat more than for the sequenced portions of Mytilus mtDNA (62%; HOFFMANN et ul. 1992). The rRNA genes are slightly more (A + T)-rich (72.2% for s-rRNA and 74.1 % for 1-rRNA ) and the proteinencoding genes slightly less (A + T)-rich (67.6%) than the genome average.
An analysis of dinucleotide frequencies shows that of CG to be 1.4%, only 0.64 of the value expected, given the individual nucleotide frequencies (C = 11.9% and G =
18.6% for the strand shown in the APPENDIX) and assum- ing random distribution. The ratio of 0.64 for the o b served to expected frequency of CG is the lowest such ratio among all dinucleotide pairs in this mtDNA, as is typical among metazoan mtDNAs [e.g., D. yakuba = 0.69, human = 0.64, sea urchin (Strongylocentrotuspur- purutus) 0.57 and nematode ( C. eleguns) = 0.601. CG frequency is low in the proteinencoding genes (0.56 of expectation) and in the large unassigned region (0.40 of expectation), but not in the rRNA genes (1.13 and 0.98 of expectation for s- and 1-rRNA genes, respec- tively). CG dinucleotides are underrepresented in ver- tebrate nuclear genomes (RUSSELL et ul. 1976), where their potential for hypermutability due to C methylation appears to be selected against (BIRD 1986; DROUIN 1991). However, there is evidence that vertebrate mtDNA is unmethylated (reviewed in BROWN 1985), and this may be true of metazoan mtDNAs in general. Therefore, the low CG frequency in metazoan mtDNAs demands an alternative explanation.
Gene content and arrangement: Katharina mtDNA contains genes for the 13 protein subunits [l-6 and 4L

426
Mytilus
Katharina
D
L(UUR) K G ANE H S(W) UCLLCU Y I M Wy -
Drosophila d ! i 1111 I 1 4 13 ICYTB 1 U l j - r R N d
I- - ND2 4 h
S(AGN) Q G
FIGURE 2.-Comparison of mitochondrial gene arrangements among K. tunicata, M . edulis (HOFFMANN et al. 1992) and D. yakuba (CLARYand WoLsTENHoLME 1985a). Each mtDNAis shownwith the 5"end of C O l to the left. All Mytilus genes are transcribed from left to right, as are all Katharina and Drosophila genes except those underlined. Lines connecting gene pairs (or blocks of contiguous genes, when marked by a bar) show rearrangements needed to interconvert the (protein + rRNA) gene arrangements; tRNA gene rearrangements, which are numerous, have been ignored. Lines with encircling arrows are inversions; lines without them are transpositions. Gene designations as in FIGURE 1. except that M(AUA) in M y t h s denotes a supernumerary tRNAmet-like gene with the anticodon TAT.
of NADH dehydrogenase (ND1-6, 4L), 1-111 of cyto- chrome c oxidase (CO1-3), 6 and 8 of ATP synthase (ATPase6 and 8), and cytochrome b apoenzyme (Cytb)] , 2 rRNAs and 22 tRNh that are found in most metazoan mtDNAs. It is notable that in this it differs from another mollusc, the bivalve M . edulis, whose mtDNA does not contain ATPase8 (HOFFMANN et al. 1992). Unlike other metazoan mtDNAs examined, Katharina mtDNA con- tains two additional sequences which can be folded into tRNA-like structures, although it is questionable whether either is a functional tRNA gene. Mytilus also has a sequence that may be a supernumerary tRNA gene (tRNAmet(*'*); HOFFMANN et al. 1992), but it is not ho- mologous to either of those found in Katharina
The Katharina gene arrangement is notably similar to those of the arthropods Drosophila (CLARY and WOLSTENHOLME 1985a) and Apis (CROZIER and CROZIER 1993). If one considers only the 15 genes encoding rR- NAs and proteins, only two rearrangements are required
(tRNAphe(uuu) and mAser(ucu) ).
to interconvert the Katharina and arthropod gene or- ders: a transposition of the block C03-ND3, and an in- version of the block ND6-Cytb (see Figure 2). As noted previously (CLARY and WOLSTENHOLME 1985a; MORITZ et al. 1987), tRNA genes appear to rearrange at a higher frequency: 10 differ in their relative locations between Katharina and Drosophila mtDNAs [those for leu (UUR) , lys, asp, gly, ser(AGN) , glu, ile, gln, met and trp] .
The similarity of the Katharina gene arrangement with those of the arthropods Apis and Drosophila is in marked contrast to its dissimilaritywith the arrangement of the bivalve mollusc Mytilus. The only Katharina gene boundaries that are shared by Mytilus are those between tRNAth' and ND4L and among tRNA"uiCuN), tRNA"(uUR' and NDI. The results of these gene arrangement com- parisons, which run counter to all expectations based on presently accepted phylogenetic affinities among these taxa, are discussed further, in conjunction with other comparisons, in the section on phylogenetic implications.

Chiton Mitochondrial Genome 427
ND4L ND6 ATPase8
Katharina
10 20 30 40 50 60 70 80 93 100 20 40 60 80 100 120 140 160 10 20 30 40 50
Drosophila
10 20 30 40 50 60 70 80 90 M 40 60 80 100 120 140 160 10 20 30 40 50
FIGURE 3.-Hydrophilicity profiles of the mitochondrially encoded ND4L, ND6 and ATPase8 proteins of K. tunicata (above) compared with those of D. yakuba (below). Hydrophilicity was computed by the method of KWE and DOOLITTLE (1982), with a search window setting of seven. The y-axis represents the hydrophilicity scale ranging from -5.0 to +5.0; x-axis numbers designate amino acid positions in each protein.
Thirty-one Katharina mtDNA genes either abut di- rectly or have some noncoding sequence separating them. In seven cases, however, there are overlaps be- tween adjacent genes. In two, the genes are encoded on opposite strands ( tRNASe'(UCN) overlaps tRNA"' by 1 nt, and tRNAP'" overlaps NDI by 8 nt) , which presents no problems for the resolution of their respective tran- scripts. In five, however, the overlapping genes are en- coded by the same strand. The gene pairs tRNA1e"(CuN)-
and tRNAtYr-tRNAcYs each overlap by 1 nt. Their transcripts could be resolved if the processing point alternated by 1 nt, if the transcripts were length- ened by enzymatic editing, or if the unpaired nucleotide at the 3'end of the upstream tRNA were unnecessary to the downstream tRNA. However, it is more difficult to explain how the transcripts for the two gene pairs with 2-nt overlaps ( tRNAgin-tRNAi* and tRNAaSn-tRNA'le) might be resolved. An alternating point of transcript processing, transcript editing, or relaxation of tRNA structure could again be invoked. An argument against the latter is that all Katharina tRNA genes, including these four, are invariant in having the potential for standard, seven-member amino-acyl stems. Finally, tRNAIYY' and tRNA"'" overlap on the same DNA strand by 4 nt; for these, alternation of transcript processing or transcript editing are the only explanations we can propose.
In mammalian mitochondria, the tRNA portions of the polycistronic transcript have been suggested to serve as processing signals for its cleavage into gene-specific RNAs (BATTEY and CLAWON 1980; OJALA et al . 1980, 1981). In cases where proteinencoding genes abut di- rectly, sequences adjacent to the genejunctions have the potential to form stem-loop structures which, when tran- scribed, may serve as alternative signals for RNA pro- cessing enzymes. Either tRNA genes or sequences with the potential to form stem-loop structures have been found at nearly all mitochondrial gene boundaries in mammals ( M u s domesticus, Homo sapiens and Bos tau- rus; BIBB et al. 1981), in the insects D. yakuba (CLARY and WOISTENHOLME 1985a) and D. melanogaster (DE BRUIJN 1983), in the sea urchin Paracentrotus lividus (CANTATORE et al. 198713, 1989), and in the nematodes
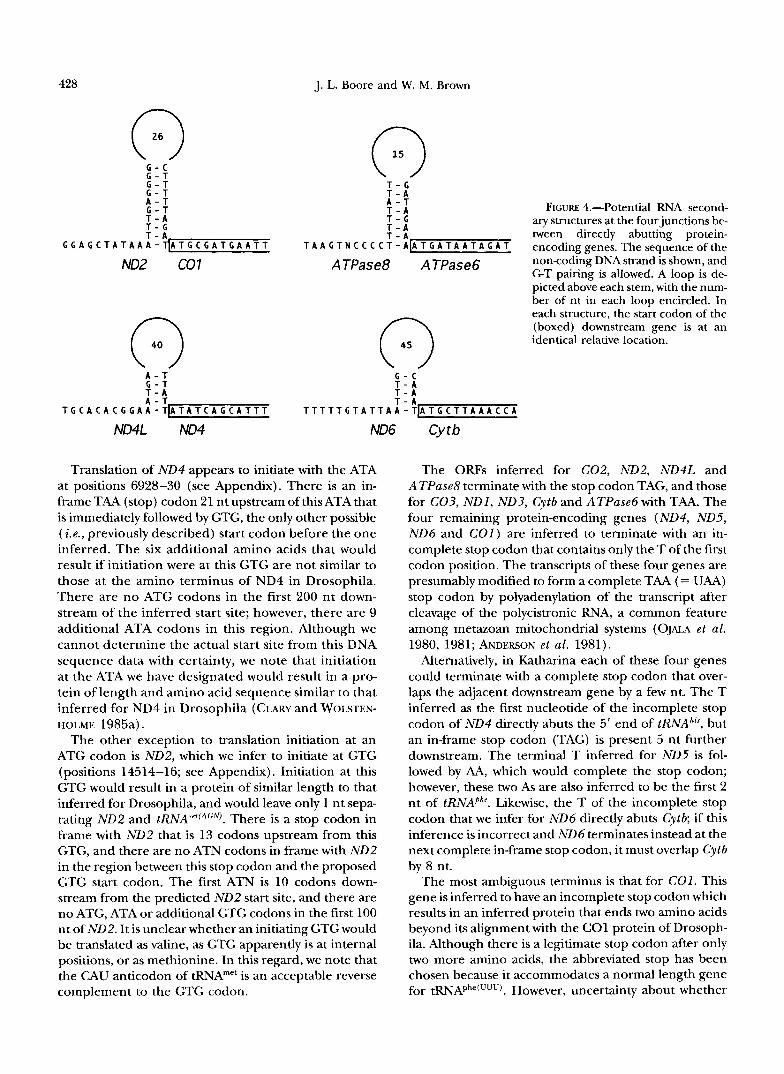
C. elegans and Ascaris suum (OKIMOTO et al . 1992). In Katharina mtDNA there are four pairs of protein- encoding genes that abut without an intervening tRNA: NDZ-COl, ATPase8-ATPase6, ND4L-ND4 and ND6- Cytb. At each of these four gene junctions there is a sequence with the potential for forming a stem-loop structure, and in each case the start codon of the down- stream gene is positioned identically relative to that structure (see Figure 4). Although sequences with the potential to form other stem-loop structures also occur near the gene boundaries, there is no commonality to their positions relative to the gene termini, and none of them is predicted to be more stable then the structures in Figure 4. Although additional structuring (not shown) is possible within each of the loops, there is no apparent similarity in this respect among the several loops. The variability in loop size and, in the cases of the ND4-ND4L and ND6-Cytbjunctions, the short stems ar- gue against the hypothesis that these structures are sig- nificant in vivo. If one assumes a random sequence model, it is likely that a reverse complement to the 5 nt preceding ND4 (TTATT) would occur by chance alone within the 50 nt upstream of ND4 ( P = 0.78), and some- what likely for the 5 nt upstream of Cytb (CAAAT) ( P = 0.1). However, the likelihood of chance matches as ex- planations for the potential secondary structures be- tween NDZ-COl and ATPase8-ATPase6 is much lower ( P = 0.006). Thus, the occurrence of these structures is suggestive of function. However, whether the structures actually form in vivo and fimction in transcript processing is purely speculative, since no experimental data exist.
Translation initiation and termination: An ATG codon is at the inferred initiation site in 11 of the 13 protein-encoding genes (see Table 2). Translation of the other two appears to initiate at ATA (ND4) and GTG (ND2), both of which have been invoked as alternative start codons in other animal mitochondrial systems (MONTOYA et al. 1981; ANDERSON et al. 1981, 1982; BIBB et al. 1981; CLARY and WOLSTENHOLME 1985a; GADALETA et al. 1988, 1989; JACOBS et al. 1988; GAREY and WOLSTENHOLME 1989; DESJARDINS and MORAIS 1990; JOHANSEN et al. 1990; HOFFMANN et al. 1992) and in some prokaryotes (e.g., STORMO et al. 1982).
428
G - C G - T
G - T 6 - T
J. L. Boore and W. M. Brown
T - A 1 - 6
A - T G - T T - A
T - A T - G
G G A G C T A T A A A - T ~ A T G C G A T G A A T T
ND2 CO7
0 A - T
T - A G - T
A - T T G C A C A C G G A A - T ~ A T A T C A G C A T T T
ND4L ND4
A - 7 1-5 ary structures at the four iunctions be- T - A FIGURE 4.-Potential RNA second- l - A T - A tween directly abutting protein-
- T A A G T N C C C C T - A t A T G A T A A T A G A T encodinggenes.Thesequenceofthe
noncoding DNA strand is shown, and G T pairing is allowed. A loop is de- A TPase8 A TPase6 picted above each stem, with the num- ber of nt in each loop encircled. In each structure, the start codon of the (boxed) downstream gene is at an identical relative location.
G - C
T - A T - A
T - A T T T T T G T A T T A A - T ~ A T G C T T A A A C C A
ND6
Translation of ND4 appears to initiate with the ATA at positions 6928-30 (see Appendix). There is an in- frame TAA (stop) codon 21 nt upstream of this ATA that is immediately followed by GTG, the only other possible ( i.e., previously described) start codon before the one inferred. The six additional amino acids that would result if initiation were at this GTG are not similar to those at the amino terminus of ND4 in Drosophila. There are no ATG codons in the first 200 nt down- stream of the inferred start site; however, there are 9 additional ATA codons in this region. Although we cannot determine the actual start site from this DNA sequence data with certainty, we note that initiation at the ATA we have designated would result in a pro- tein of length and amino acid sequence similar to that inferred for ND4 in Drosophila (CLARY and WOLSTEN- HOLME 1985a).
The other exception to translation initiation at an ATG codon is ND2, which we infer to initiate at GTG (positions 14514-16; see Appendix). Initiation at this GTG would result in a protein of similar length to that inferred for Drosophila, and would leave only 1 nt sepa- rating ND2 and tRNAserlAGN). There is a stop codon in frame with ND2 that is 13 codons upstream from this GTG, and there are no ATN codons in frame with ND2 in the region between this stop codon and the proposed GTG start codon. The first ATN is 10 codons down- stream from the predicted ND2 start site, and there are no ATG, ATA or additional GTG codons in the first 100 nt of ND2. It is unclear whether an initiating GTG would be translated as valine, as GTG apparently is at internal positions, or as methionine. In this regard, we note that the CAU anticodon of tRNA"" is an acceptable reverse complement to the GTG codon.
Cytb
The ORFs inferred for C02, ND2, ND4L and A TPase8 terminate with the stop codon TAG, and those for C03, ND1, ND3, Cytb and ATPaseGwith TAA. The four remaining protein-encoding genes (ND4, NDS, ND6 and COl) are inferred to terminate with an in- complete stop codon that contains only the T of the first codon position. The transcripts of these four genes are presumably modified to form a complete TAA (= UAA) stop codon by polyadenylation of the transcript after cleavage of the polycistronic RNA, a common feature among metazoan mitochondrial systems (OJALA et al. 1980, 1981; ANDERSON et al. 1981).
Alternatively, in Katharina each of these four genes could terminate with a complete stop codon that over- laps the adjacent downstream gene by a few nt. The T inferred as the first nucleotide of the incomplete stop codon of ND4 directly abuts the 5' end of tRNAhiS, but an in-frame stop codon (TAG) is present 5 nt further downstream. The terminal T inferred for NDS is fol- lowed by AA, which would complete the stop codon; however, these two As are also inferred to be the first 2 nt of tRNAPhe. Likewise, the T of the incomplete stop codon that we infer for ND6 directly abuts Cytb; if this inference is incorrect and ND6 terminates instead at the next complete in-frame stop codon, it must overlap Cytb by 8 nt.
The most ambiguous terminus is that for CO1. This gene is inferred to have an incomplete stop codon which results in an inferred protein that ends two amino acids beyond its alignment with the CO1 protein of Drosoph- ila. Although there is a legitimate stop codon after only two more amino acids, the abbreviated stop has been chosen because it accommodates a normal length gene for tRNAphe(UUU). However, uncertainty about whether
Chiton Mitochondrial Genome 429
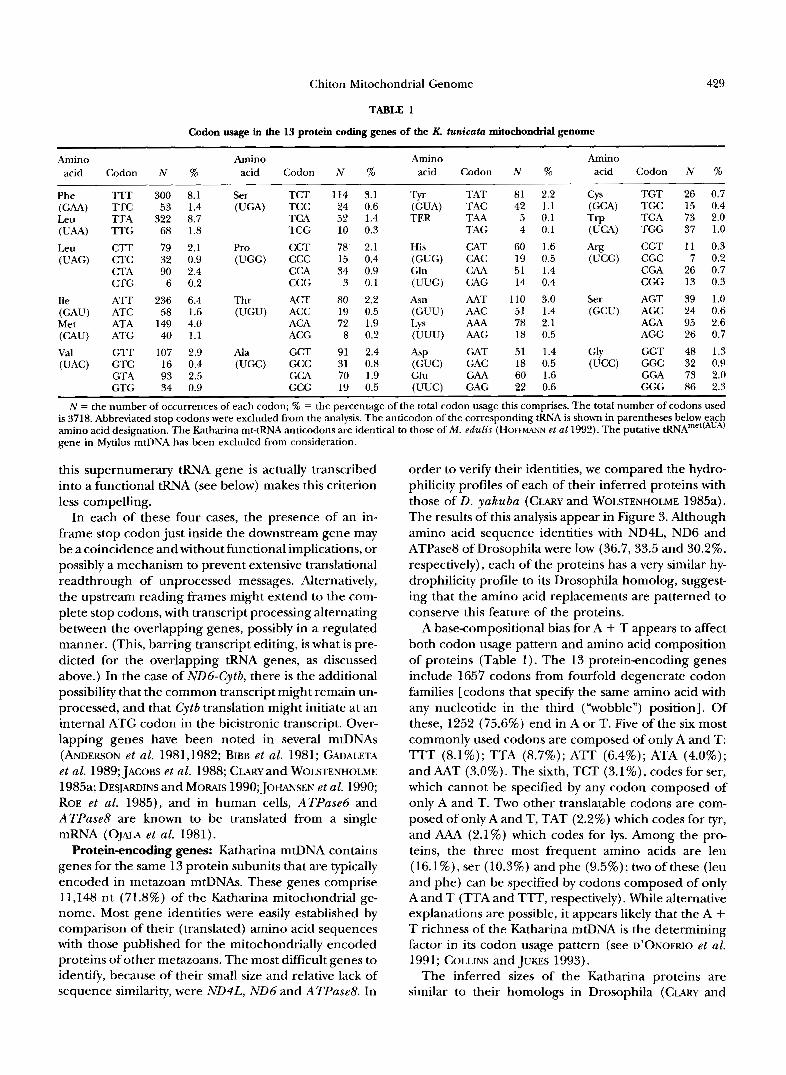
TABLE 1
Codon usage in the 13 protein coding genes of the K. tunicata mitochondrial genome
Codon
TIT TTC TTA TTG CTT CTC CTA CTG
ATT ATC ATA ATG
Gl" GTC GTA GTG
N
300 53
322 68
79 32 90 6
236 58
149 40
107 16 93 34
-
Amino % acid Codon N 5%
8.1 Ser TCT 114 3.1 1.4 (UGA) TCC 24 0.6 8.7 TCA 52 1.4 1.8 TCG 10 0.3
2.1 Pro CCT 78' 2.1 0.9 (UGG) CCC 15 0.4 2.4 CCA 34 0.9 0.2 CCG 3 0.1 6.4 Thr ACT 80 2.2 1.6 (UGU) ACC 19 0.5 4.0 ACA 72 1.9 1.1 ACG 8 0.2 2.9 Ala GCT 91 2.4 0.4 (UGC) GCC 31 0.8 2.5 GCA 70 1.9 0.9 GCG 19 0.5
Codon N
TAT 81 TAC 42 TAA 5 TAG 4 CAT 60 CAC 19 CAA 51 GAG 14 AAT 110 AAC 51 AAA 78 AAG 18
GAT 51 GAC 18 GAA 60 GAG 22
%
2.2 1.1 0.1 0.1 1.6 0.5 1.4 0.4 3.0 1.4 2.1 0.5 1.4 0.5 1.6 0.6
Amino acid Codon
TGT TGC TGA TGG
CGT CGC CGA CGG AGT AGC AGA AGG
GGT GGC GGA GGG
N
26 15 73 37 11 7
26 13
39 24 95 26 48 32 73 86
~
%
0.7 0.4 2.0 1.0
0.3 0.2 0.7 0.3
1 .o 0.6 2.6 0.7 1.3 0.9 2.0 2.3
N = the number of occurrences of each codon; % = the percentage of the total codon usage this comprises. The total number of codons used is 3718. Abbreviated stop codons were excluded from the analysis. The anticodon of the corresponding tRNA is shown in parentheses below each amino acid designation. The Katharina mt-tRNA anticodons are identical to those of M . edulis (HOFFMANN et a1 1992). The putative tRNAme'(AUA) gene in MytilusmtDNA has been excluded from consideration.
this supernumerary tRNA gene is actually transcribed into a functional tRNA (see below) makes this criterion less compelling.
In each of these four cases, the presence of an in- frame stop codon just inside the downstream gene may be a coincidence and without functional implications, or possibly a mechanism to prevent extensive translational readthrough of unprocessed messages. Alternatively, the upstream reading frames might extend to the com- plete stop codons, with transcript processing alternating between the overlapping genes, possibly in a regulated manner. (This, barring transcript editing, is what is pre- dicted for the overlapping tRNA genes, as discussed above.) In the case of ND6-Cytb, there is the additional possibility that the common transcript might remain un- processed, and that Cytb translation might initiate at an internal ATG codon in the bicistronic transcript. Over- lapping genes have been noted in several mtDNAs (ANDERSON et al. 1981,1982; BIBB et al. 1981; GADALETA et al. 1989; JACOBS et al. 1988; CLARY and WOLSTENHOLME 1985a; DESJARDINS and MORAIS 1990; JOHANSEN et al. 1990; ROE et al. 1985), and in human cells, ATPase6 and ATPase8 are known to be translated from a single mRNA (OJALA et al. 1981).
Protein-encoding genes: Katharina mtDNA contains genes for the same 13 protein subunits that are typically encoded in metazoan mtDNAs. These genes comprise 11,148 nt (71.8%) of the Katharina mitochondrial ge- nome. Most gene identities were easily established by comparison of their (translated) amino acid sequences with those published for the mitochondrially encoded proteins of other metazoans. The most difficult genes to identify, because of their small size and relative lack of sequence similarity, were ND4L, ND6 and ATPase8. In
order to verify their identities, we compared the hydro- philicity profiles of each of their inferred proteins with those of D. yakuba (CLARY and WOLSTENHOLME 1985a). The results of this analysis appear in Figure 3. Although amino acid sequence identities with ND4L, ND6 and ATPase8 of Drosophila were low (36.7, 33.5 and 30.2%, respectively), each of the proteins has a very similar hy- drophilicity profile to its Drosophila homolog, suggest- ing that the amino acid replacements are patterned to conserve this feature of the proteins.
A base-compositional bias for A + T appears to affect both codon usage pattern and amino acid composition of proteins (Table 1). The 13 proteinencoding genes include 1657 codons from fourfold degenerate codon families [codons that specify the same amino acid with any nucleotide in the third ("wobble") position]. Of these, 1252 (75.6%) end in A or T. Five of the six most commonly used codons are composed of only A and T: TTT (8.1%); TTA (8.7%); ATT (6.4%); ATA (4.0%); and AAT (3.0%). The sixth, TCT (3.1%), codes for ser, which cannot be specified by any codon composed of only A and T. Two other translatable codons are com- posed of only A and T, TAT (2.2%) which codes for tyr, and AAA (2.1%) which codes for lys. Among the pro- teins, the three most frequent amino acids are leu (16.1%), ser (10.3%) and phe (9.5%); two ofthese (leu and phe) can be specified by codons composed of only A and T (TTA and TTT, respectively). While alternative explanations are possible, it appears likely that the A + T richness of the Katharina mtDNA is the determining factor in its codon usage pattern (see D'ONOFRIO et al. 1991; COLLINS and JUKES 1993).
The inferred sizes of the Katharina proteins are similar to their homologs in Drosophila (CLARY and

430 J. L. Boore and W. M. Brown
TABLE 2
Comparisons among the mitochondrial protein coding genes of K. tunicata, D . yakuba and M. edulis
Protein
ATPase6 ATPase8 co1 c02 C03 Cytb ND1 ND2 ND3 ND4 ND4L ND5 ND6
Number of amino acids
Katharina Drosophila Mytilus
230 224 238 53 53
513 512 Not present
229 Partial (129)
228 259
Partial (193) 262 264
379 378 316
Partial (272) 324
338 341 Partial (221)
120 Partial (217)
117 116 442 446 100
Partial (287) 96 93
571 573 166
Partial (522) 174 158
Percent amino acid identity
Katharina/ Katharina/ Drosophila Mytilus
37.8 32.7 30.2 76.7
Not present 39.5
62.6 37.3 64.1 46.6 68.7 47.4 54.4 46.2 38.9 37.3 48.1 44.8 47.3 35.5 36.7 39.8 44.1 41.2 33.5 22.8
Predicted initiation and termination codons
Katharina Drosophila Mytilus
ATG TAA ATG TAa ATG TAG ATG TAG A'M TAA Not present ATG T a ATAA TAA - TAA ATG TAG ATG Ta ATG TAG ATG TAA ATG TAA ATG TAA ATG TAA ATG TAA ATG TAA
GTG TAG A l T T a ATG TAG ATG TAA A'M TAA ATG TAA ATA Ta ATG T" ATG TAA ATG TAG ATG TA" ATG TAA ATG T a A'M T" ATA TA" ATG Ta ATT TAA ATG TAA
ATG TAA ATA TAA - TA'
Inferred sizes, degree of amino acid identity and predicted initiation and termination codons were compared for the mitochondrially encoded protein genes of K . tunicata, D. yakuba (CLARY and WOLSTENHOLME 1985a) and M . edulis (HOFFMANN et al. 1992). Amino acid sequences were aligned pairwise using the PAM250 matrix (IBI MacVector; version 3.5). Percentage identity was calculated by dividing the number of identical amino acid positions by the average length of the sequences compared. Numbers in parentheses are the number of inferred amino acids compared from partially sequenced genes of Mytilus. No ATPase8 is present in Mytilus mtDNA.
a Incomplete termination codons presumably completed by polyadenylation (see text). *The CO1 and ND1 initiation codons of Mytilus could not be identified with confidence.
WOLSTENHOLME 1985a) and to the five proteins whose genes have been completely sequenced in Mytilus (HOFFMANN et al. 1992; see Table 2). ND1 differs sig- nificantly in size between Drosophila and Mytilus, and this may also be true of CO1 and C03, as inferred from differences in their initiation and termination points (HOFFMANN et al. 1992); in these respects, the three Katha- rina genes are more similar to Drosophila than to Mytilus.
The percent amino acid sequence identity of Katha- rina, Drosophila and Mytilus proteins are compared in Table 2. Complete sequences are available for five genes in all three species; for four of them, Katharina is more similar to Drosophila than to Mytilus. A similar result is obtained in comparisons of all of the remaining genes, which are only partially sequenced for Mytilus. It may be argued that these comparisons are biased because the Mytilus sequences are from gene termini, which are of- ten especially variable (e .g . , see the protein alignments in JACOBS et al. 1988). In order to test this, Mytilus and Katharina C03 amino acid sequences were aligned, using the PAM 250 matrix of the IBI MacVector pro- gram, and analyzed for patterns of amino acid identity. [C03 was chosen because it has the least ambiguous alignment of the five genes completely sequenced in Mytilus.] Fifty-seven (43%) of the 132 aligned positions in the central region are identical, whereas 65 (49%) of the 132 aligned positions nearest the ends ( L e . , the 66 aligned positions nearest each end) are identical, sug- gesting that comparisons of the terminal amino acid se- quences of Mytilus proteins may be representative of the overall degree of variation.
Genetic code: Similarities in codon usage patterns, tRNA anticodons and amino acid sequence alignments
suggest that the mitochondrial genetic code of Katha- rina is identical to those of Drosophila and Mytilus. All codons occur within the protein genes of Katharina mtDNA except the stop codons TAA and TAG. In par- ticular, three codons (TGA, AGG and AGA) that appear to function as stop codons in the mitochondrial systems of some metazoans occur frequently in Katharina pro- tein genes.
AGA and AGG, like AGY codons, specifr serine: AGA and AGG code for arg in the "universal" genetic code and in the mitochondrial codes of protozoa (PRITCHARD et al. 1990; ZIAIE and SUYAMA 1987), fungi (BONITZ et al. 1980) and plants (see JUKES and OSAWA 1990; OHYM et al. 1991), for gly in ascidian mtDNA (YOKOBORI et al. 1993), and are designated as stop codons in vertebrate mtDNA (ANDERSON et al. 1981, 1982; BIBB et al. 1981; GADALETA et al. 1989; ROE et al. 1985; JOHANSEN et al. 1990). In mitochondria, tRNAser(AGYJ appears also to rec- ognize AGA and AGG in flatworms (Fasciola; GAREY and WOLSTENHOLME 1989), echinoderms UACOBS et al. 1988; CANTATORE et al. 1989; DE GIORCI et al. 1991; HIMENO et al. 1987; SMITH et al. 1989, 1990), Mytilus (HOFFMANN et al. 1992) and nematodes (OKIMOTO et al. 1992). AGA also codes for serin Drosophila mtDNA, but AGG codons are not used (CLARY and WOLSTENHOLME 1985a). Of the 95 in-frame AGA codons in Katharina mtDNA, 34 corre- spond in position to ser in Drosophila proteins (1 1 to AGA, 8 to AGT, 15 to TCN) , but none correspond to arg. AGA codons in Katharina mtDNA correspond to asp positions in Drosophila proteins 11 times and 6 times each to gly, met and thr. Of the 26 in-frame AGG codons in Katharina mtDNA, 6 correspond in position to ser in Drosophila proteins (2 to AGA, 3 to AGT, 1 to T W ) ,

Chiton Mitochondrial Genome 431
but none correspond to arg. Katharina AGG codons correspond in position to thr in Drosophila proteins five times, and to no other amino acid more than three times.
Although AGG codons in Katharina correspond al- most equally to thr and ser positions in Drosophila pro- teins, we do not regard this as significant. Katharina mtDNA encodes a tRNA with the anticodon UGU, typi- cal for the tRNAth‘ which decodes the ACN codon family. This tRNA would not be expected to recognize the codon AGG, nor does it seem likely that any other Ka- tharina tRNA would be able to discriminate AGG codons from others in the AGN family. It seems most likely, therefore, that AGG specifies ser in Katharina mtDNA, and that the nearly identical number of matches to ser and thr positions in Drosophila proteins is due to (1) the similar chemical nature of ser and thr, (2) the lack of AGG codons in Drosophila, and (3) the potential for one-step interconversion of thr and ser codons (thr = ACN; ser = AGN).
The anticodon of tRNAse‘(AGN) is GCU. Since this is the only tRNA with an NCU anticodon and since AGR codons occur frequently in Katharina mitochondrial protein genes, this tRNA presumably recognizes all AGN codons as ser. It is not obvious how the G in this anticodon would pair with A or G to recognize both AGA and AGG codons, although this also appears to be the case in several other metazoan mitochondrial systems. This may be at- tributable to posttranscriptional modification of nucleoti- des, which is a common feature of nuclearencoded tRNAs and has been demonstrated in a mitochondrial tRNA from a marsupial (JANKE and PAABo 1993).
TGA, like TGG codons, specify tryptophan: TGA is a stop codon in the “universal” genetic code and in the mitochondrial code of plants, but specifies trp in the mitochondria of fungi (CUMMINGS and DOMENICO 1988), protozoa (PRITCHARD et al. 1990; ZIAIE and SUYAMA 1987) and all metazoans so far reported (see JUKES and OSAWA 1990). TGA codons occur 73 times in the mitochondrial genes of Katharina; 51 of these correspond in position to trp in Drosophila proteins. Two tRNAs are identified with NCA anticodons, suggesting that the TGN codon family is twofold degenerate: tRNAV, with anticodon GCA, would be expected to recognize TGT and TGC; tRNAT, with anticodon UCA, would be expected to rec- ognize TGA and TGG.
A T A codons specify methionine rather than isoleucine: ATA codes for ile in the “universa1”genetic code, but for met in the mitochondria of many organisms. Of 149 ATA codon occurrences in Katharina mtDNA, 53 cor- respond in position to met in Drosophila proteins and only 14 correspond to ile, suggesting that ATA codes for met in Katharina mtDNA. ATA codons in Katharina mtDNA also correspond to leu positions in Drosophila proteins in 36 of the 149 occurrences. It seems unlikely, however, that this is significant. Leu is the most abun-
dant amino acid in Katharina mitochondrial proteins (16.1 % of all amino acids). Only 1 nt change is required to convert the leu codons CTA and TTA to ATA, and both CTA and TTA are present in abundance (2.4% and 8.7% of all codons, respectively). Further, no tRNA with an anticodon that could discriminate an ATA from ATG, ATC and ATT has been identified. The anticodon of tRNA1le is GAU, so the G in the wobble position is ex- pected to discriminate the ATT and ATC codons. The anticodon of tRNAm“is CAU, so C in the wobble position presumably pairs with either A or G of ATA and ATG codons. While it is not obvious how the GA pairing is achieved, this appears to be the general case for tRNAmet in mitochondrial systems.
Code evidence from tRNA anticodons: Identity of the mitochondrial genetic code in Drosophila and Katha- rina is also indicated by comparisons of their cognate tRNA anticodons, which are identical in all but one case. The exceptional tRNA in Katharina has the anticodon UUU, and presumably recognizes the lys codons AAA and AAG; the U in the wobble position could do this efficiently. The cognate tRNA anticodon in Drosophila is CUU (CLARY and WOLSTENHOLME 1985a), but in Myti- lus it is also UUU (HOFFMANN et al. 1992).
Other than in (see above), G only occurs in the wobble position of tRNAs that specifically recog- nize NNYcodons (those for asp, asn, cys, his, ile, phe and tyr) . C in the wobble position of tRNAmet must pair with both the A in ATA and the G in ATG. U is in the wobble position of all other WAS; these either do not discrimi- nate among nucleotides in the third codon position (fourfold degenerate codon families) or specifically rec- ognize NNR codons (glu, gln, leu(UUR), lys and trp).
Novel tRNA genes do not suggest a code change: Based on their folding potential, two sequences in Katharina mtDNA may encode two additional tRNAs, with AAA and AGA anticodons (see below and Figure 5) . It is not known whether either of these sequences is transcribed or produces a functional tRNA, but sequence consider- ations (discussed below) argue against the latter. If they encoded functional WAS, their products would be ex- pected to recognize the codons TTT and TCT and to specify phe and ser, respectively. However, it is theo- retically possible that these codons would be uniquely recognized by the two tRNAs and might specify different amino acids. To test this unlikely hypothesis, we com- pared the correspondence of TIT and TCT codons in Katharina mtDNA to Drosophila amino acid positions. Out of 300 Katharina TTT codons, 153 corresponded to phe, 38 to leu, 25 to ile, 21 to tyr, and no more than 12 to any other amino acid. Out of 114 Katharina TCT codons, 60 corresponded to ser (30 to TCT codons and 30 to one of the other 7 ser-specifying codons), and no more than 7 to any other amino acid. Therefore, it appears likely that TTT and TCT specify phe and ser, respectively.

432 J. L. Boore and W. M. Brown
Alanine c - 2 A - T c -c A-T C -C T T A -T A
T T C C C C
G I l l 1 A A T T T C A I I I I I A
A C C C C
T A A A A C C A T C
T - T - A T - A A - T
T T A - T
T A T C C
Proline c -2 A-T C - C A- T T - A
C- C
A A C - C
T A T C C
Arginine c-E
C - C T - A
T - A T - A A - T
A T T C T C I A - T T T
T A C T C A bb!!hI I A
A T C A T ' A A T~ T'C'
I I I I
T T T - A C - C
T C C - C
T O T C G
A Glutamine :I;
C C A
A
A - T
A - T A - T
A - T C -C T
T T T T T T C
A A A A A C A a. I I I I I I
T C T C
C C A C A A C '
( ( 1 1 T
T - A A A A - T C - C
A A
C - C A- T
T A T T C
Asparagine T-:
A-T 0 - C
A-T T - A A -T T - A
A T A T 0 T C T C G A I I I
c I I I I A T A
T T C A C C ~ A
T - A c A c G
T - A T T
G - C
T A T A
G - C
C T T
serine A-: S e r i n e ( ~ ~ ~ ) (AGN) :I: Folding A - T
G - C Alternative A-T A- T
T T A - 1 G-T - A- T C - T CCw T T C C C T T A A T- 6 C C T T A '
I I I I I G C G I l l l l c A C C C G
T T c O T T T I I I I
C C C G T I
'T T T - A A ~ ~
T - A T - A T - A
G - T T - A C - T
C A T A
C - C G - C C A
C C T T A
C C T
A
A-T T - A
C-C A-T
A - T C-C
Tyrosine A - T
T T T C C T A C C C G C I I I I
C A G G T A I l l
A & A G C T C G - TT TA
A - T A - T
T - A
C A T A
A - T
C T A
T
I T I A
A
A - T A - T A - T A - T
C - C T - A
Valine T - A
A A A I I I I A C A C A ' A
T - A T - A T - A
T A - T C T A
T A C
Aspartate C-: A-T C -C A -T
A -T A -T
T - A A T T C A A
A C T T A A A I I I I A
A A T T C
A T A A A C A - ~ A G T
A - T C - C C - T
T A T A
T - A
C T C
A Histidine ;I;
T - A A -T C -C A -T T - A
T A A I I I I T T T C A
A A C T A T T T A
A A A a T - A A A C
T - A A - T A - T
T G - c T T C
C T G
Methionine T-i C-C
A-T T - A
A - T A - T A - T
T C T C T
T !).hi T C T C C A G A G A
A A A I I I I
T A A A T C T T- A G A G - C
T C C- C
T A C A T
A
A C
T A
T
T T
Cysteine A-: A -1 C -c T - A c -0 T -A A - T
T T C T T A C A A
A I I I I T T T C
A - T T - A
A - T T A - T A T A
C C A
T Isoleucine ;I;
T - A c -c T - A
i-A A - T c - c
T T A - T
T C C A T
C - C C - C
A - T T C T C
6 A A
Threonine c-; C-C T - A A-T A-T
T- A
C A
C- c
T A
T - A
T C A
C - C
C A
C - C T - A
T A T G T
T - G
T T T - A
A C A A A
T T A A
A- T
C C A C A
FIGURE 5.-Twenty-two Katharina mtDNA sequence elements folded into secondary structures resembling mitochondrial WAS. DNA sequences of the non-coding strand are shown. Bars connecting nucleotide pairs indicate base pairing, with GT pairs allowed. Dashed bars indicate potential base pairing within loops. Heavy lines indicate short complementary sequences within the large loops of serine tRNA. WAxr(AGN) is shown in both the form that has been hypothesized in other animal mtDNA studies and in an alternative form with a folded D arm. The folded version is shifted 2-3 nucleotides relative to the version lacking a D arm. Two additional tRNA-like structures are shown in the box at the lower right (see text).

Chiton Mitochondrial Genome 433
Transfer RNAs Twenty-four sequences that could be folded into tRNA-like structures with acceptable antic- odons were identified. Of these, 22 corresponded to the standard set of tRNA genes found in other metazoan mDNAs; these are discussed in the first part of this sec- tion. The two supernumerary sequences, which have no counterparts in other reported metazoan mtDNAs, are discussed in the latter part of this section.
The standard tRNA gene set: The corresponding mi- tochondrial tRNA genes of Katharina and Mytilus have identical anticodons, and in Drosophila only the tRNA'Ys anticodon differs from these. The Katharina tRNA'F an- ticodon is UUU, as in vertebrate (see references above), Apis (CROZIER and CROZIER 1993) and nematode mtDNAs (OKIMOTO et al. 1992), whereas it is CUU in Drosophila, mosquito (HSUCHEN et al. 1983) and Fas- ciola mtDNAs (GAREY and WOLSTENHOLME 1989). The codons M A and AAG specify lys, so a U, as occurs in Katharina, would be expected in the wobble position of the anticodon. The C found at this position in some organisms is unexpected and must, under suitable cir- cumstances, be able to pair with both A and G (this is also true for the CAU anticodon of tRNAme', which recog- nizes both ATA and ATG) and to discriminate against the codons AAT and AAC, which specify asp and whose cognate tRNA has the anticodon GUU.
As shown in Figure 5, all Katharina mt-tRNAs have a seven-member amino-acyl stem, a five-member antic- odon stem, and a seven-member anticodon loop. The variable arm sizes range from 3 to 6 nt, with 4 nt being most common. The D arm (shown to the left in each tRNA in Figure 5) has a paired stem that varies from 3 to 5 nt, without mismatch, and an unpaired loop offrom 3 to 8 nt. The opposite (TWC) arm has a paired stem that varies, except in tRNAglu, from 3 to 6 nt, without mis- match, and an unpaired loop of from 3 to 8 nt. The TWC stem of tRNAg'" is only 2 nt; in all other respects the structure is conventional, but this short stem, coupled with the failure of this gene to directly abut a neigh- boring gene at either end, led us to search for an alter- native sequence. All 'TTC trinucleotides in intergenic re- gions of either strand were investigated as possible tRNAgl' anticodons, but none with flanking sequences that could be folded into a suitable tRNA structure were found.
Five tRNA genes have one mismatch each in the amino-acyl stem; in four, the mismatch is between the bases of the 6th nt pair. Six genes have mismatches in the anticodon stem: tRNAwg has two mismatches, be- tween the first 2 nt pairs at the top of the stem; tRNAmet also has two, but between nt pairs 1 and 3; of the re- maining four, three have a mismatch between the 1st nt pair, and two between the 2nd nt pair.
All 22 tRNA genes have a T immediately preceding the anticodon; immediately following it, 17 have an A and 5 have a G. A single purine nucleotide separates the anticodon stem from the D arm in 19 of the tRNAgenes.
The TWC arm derives its name from the nearly universal presence, in nuclearencoded tRNAgenes, of TTC at the first three positions in its loop (SPRINZL et al. 1991) ; in the transcript, the first U is methylated to ribothymidine (T) and the second is rearranged to pseudouridine (Y) . A TTC motif occurs at this position in only two Katharina mt-tRNA, tRNAEr(AGN) and tRNAser(UCN), wherein it is fol- lowed by the dinucleotide RA (R = A or G), as is usual for nuclear tRNA. The dinucleotide separating the amino-acyl stem from the D arm is most commonly TA (conventionally referred to as T, and b), but is TG in three of the WAS. In nuclearencoded WAS, T, usu- ally pairs with A,, to generate part of an Lshaped tertiary structure. There is potential base pairing between po- sitions 8 and 14 in 20 of the Katharina mt-tRNA: T8-Al4 in 17, &-T,, in two, and C8-G,, in one. T,-Al4 base pairing is also possible in both of the supernumerary tRNA-like structures (Figure 5).
Except for Mytilus, all metazoan mtDNAs that have been sequenced encode only one tRNA"'", which pre- sumably functions in both initiation and elongation. DUBIN and HSUCHEN (1984) analyzed the mt-tRNAmet of Aedes (mosquito) in detail, and found that it has both the U preceding the anticodon, as is typical of conven- tional elongator tRNAs, and the three G C pairs in the anticodon stem, as is typical of initiator tRNA, consis- tent with a dual role. Except for a GA mismatch in the anticodon stem, mt-tRNAmet of Katharina shares these features, along with the two T (= U) nucleotides in the anticodon stem that have been shown to be postran- scriptionally modified to pseudouridine in the Aedes mt-tRNAmet and a nearly perfect match to all nt in the D arms of Aedes and Drosophila mt-tFtNAmet (DUBIN and HSUCHEN 1984; CLARY and WOLSTENHOLME 1985a).
In all metazoan mtDNAs examined, the D arm of the
1989). In Katharina mtDNA, the can be folded into a structure resembling a conventional tRNA, complete with a paired D arm. Alternatively, by slightly altering the inferred size and position of this gene (add- ing 3 nt at the 3' end and omitting 2 nt at the 5' end) a tRNA-like structure without a paired D arm, similar to the tRNAser(AGN) of Drosophila, can be formed. Both structures are presented in Figure 5. Favoring the struc- ture lacking a paired D arm is that it most closely matches those proposed for mt-tRNAser(AGN) in other in- vertebrates; this similarity extends to the number of un- paired nucleotides. In this, but not in the structure with the paired D arm, there is the potential for interaction of T, and A,, and, perhaps more significantly, the TWC loop exactly matches the sequence of this loop in the
(T. COLLINS and W. BROWN, unpublished data), in which an alternative pairing of the D arm is not possible. As an additional ambiguity, there is potential for up to 4 ad- ditional base pairs in the anticodon stem of tRNAser(AGN).
mAser(AGN) is unpaired (see GAREY and WOLSTENHOLME
mAser(AGN) of another mollusc, the snail Plicopurpura

434 J. L. Boore and W. M. Brown
A variant secondary structure with a six-member antic- odon stem has been suggested for both insect and mam- malian tRNAser(AGN) (DUBIN et al. 1984 and references therein) and demonstrated for bovine tRNAser(UCN) (YOKOGAWA et al. 1991).
In nematodes, but not in other metazoans, mt- tRNAsr(UCN) also lacks a paired D arm (WOLSTENHOLME et al. 1987). Surprisingly, the D arm of the mt- tRNAser(UCN) of Katharina is also unpaired. Finally, the seven-member TlIrC loop of Katharina mt-tRNASer(UCN) matches the T W loop of Katharina tRNAser(AGN) at six of seven positions.
Supernumerary tRNA genelike sequences: Two addi- tional sequences can be folded into secondary structures resembling tRNAs (Figure 5). If these function as tRNA genes, they presumably recognize the codons TTT and TCT, which correspond to phe and ser, respectively. They are, therefore, provisionally identified as
suggest that they are non-functional, at least as tRNA genes: Each has an unprecedented anticodon sequence, AAA and AGA, with A in the wobble position; no tRNA encoded in Katharina, Mytilus, Apis or Drosophila mtDNAs has an A in this position, although there is an A in the wobble position of mt-tRNA”‘g in Caenorhab- ditis and Ascaris. Each has several mismatches within stems, and tRNAPhe(UUU) has only 2 nt in the variable arm. Finally, neither has a T preceding the anticodon, as is found in all other Katharina WAS.
If these two sequences do not encode functional tRNA, how might one explain this similarity? The tRNAscr(UCUi sequence occupies the region between C02 and ATPase8, except for 3 nt flanking each end; the tRNAf’he((UUU) sequence is positioned between CO1 and tRNAQSf’, ending 3 nt from CO1 and 10 nt from tRNA””. One hypothesis is that these sequences originated from tRNA gene duplications, that either they and/or their functional counterparts were transposed from the du- plication sites, and that their tRNA-like structures have been conserved because they mediate other functions (e.g. , replication, termination of D-loop DNA synthesis, transcript processing). Sequence comparisons with the other Katharina mt-tRNA genes provide no support for a duplication hypothesis; however, if the duplication events were ancient, the initial sequence resemblances would be eroded by substitutions. Experimental data on replication and transcription initiation and on tran- script processing in Katharina, as well as additional com- parisons of mtDNA sequences from other species of chi- tons and representatives of other molluscan classes are needed to clanfy this.
Ribosomal RNAs Katharina mtDNA contains genes for the s-rRNA and 1-rRNA of mitochondrial ribosomes. The precise boundaries of these genes cannot be de- termined with certainty on the basis of sequence com- parisons among species. If one assumes that the mt-
&NAphe(UUU) and tRNASer(UCU) . H owever, the following
rRNA genes extend to the exact boundaries of their adjacent genes, then genes of similar length and good match in primary sequence to those of Drosophila are obtained. Also, corresponding portions of the se- quences can be folded into secondary structures that are similar to those proposed for other metazoan s- and 1-rRNAs (Drosophila, e.g.; CLARY and WOLSTENHOLME 1985b). Interpreted in this way, the mt-+rRNA gene comprises 826 nt and the mt-1-rRNA gene comprises 1275 nt in Katharina [compared, respectively, with 789 and 1326 nt in Drosophila (CLARY and WOLSTENHOLME 1985b), ca. 819 and 1640 nt in Xenopus (ROE et al. 1985), ca. 880 and 1525 nt in echinoderms (JACOBS et al. 1988; CANTATORE et al. 1989; DE GIORGI et al. 1991), ca. 955 and 1570 nt in mammals (BIBB et al. 1981), and ca. 945 and 1245 nt in Mytilus (although there is significant ambiguity in determining the 3’ end of the 1-rRNA gene; HOFFMANN et al. 1992)].
Functionally unassigned sequences: The largest un- assigned sequence, 424 nt, is between tRNA* and C02 (AP PENDIx, positions 1683-2106). This sequence is 75.3% A -t T, with most of the C and G present in homopolymer tracts. No open reading frames of more than 100 nt (33 amino acids) are present. Four sequences that can be folded into hairpin structures with stems 210 nt and loops 550 nt are present; their stems are formed by base-pairing between positions 1740-1756 and 1758-1774 (15/16 match), 1769-1780 and 1803-1814, 1833-1847 and 1849-1863, and 2034-2044 and 2073-2083 (see APPENDIX).
The second largest unassigned sequence, of 141 nt, is positioned between tRNA@u and C03. This location co- incides with one of the two boundaries between oppo- sitely transcribed portions of the genome (Figure 1). 128 of the 141 nt are As or Ts, 72 of which occur in a tract of 36 consecutive AT pairs. Although alternating AT tracts are found in the mtDNAs of C. elegans, Ascaris suum (Om- MOTO et al. 1992), SpodOptera fmgiperda (PASHEY and KE 1992), and D. yduba (CLARY and WOLSTESHOLME 1985a), that in Katharina is the longest reported. A tract of 17 As in this region is immediately upstream of CO3; part of this tract and its flanking sequence (positions 12922-12944; see APPENDIX) are an 18/23 match to the reverse comple- ment of positions 1940-1918 in the largest unassigned se- quence. At the other end of this 141-nt region, positions 12810-12815 are an exact match to positions 1744-1749 in the largest unassigned region. Whether either match has significance is unknown.
Two other lengthy regions (those in which the two tRNA-like structures designated tRNAs’(Ucu) and tRNAPhe(U”U) occur) may also belong in the unassigned category. The remaining 120 unassigned nt occur in- tergenically, in short segments of 1-28 nt. No commonly held sequence elements or other features were found among them.
Phylogenetic implications: The arrangement of genes in Katharina mtDNA is quite similar to those of

Chiton Mitochondrial Genome 435
several other metazoan phyla. This was unexpected, given the marked dissimilarity in arrangement between another mollusc, the bivalve M. edulis, and these same phyla, and given the much closer phylogenetic affinity that is thought to exist between Mytilus and Katharina. Ignoring tRNA genes, two rearrangement events are necessary to interconvert the gene arrangements of Ka- tharina and Drosophila (Figure 2), whereas many (210, including loss of A TPase8) are required to interconvert those of Katharina and Mytilus. Moreover, neither Ka- tharina nor Mytilus share any unique feature (apomor- phy) that is in either of their respective mtDNAs (e.g. , protein gene sizes; possible supernumerary tRNA genes). Finally, as discussed above, the proteinencoding gene sequences of Katharina and Drosophila are more similar than those of Katharina and Mytilus or of Dro- sophila and Mytilus (Table 2).
One explanation for these observations is that the as- sumed relationship of bivalves and chitons as taxa (classes) within the phylum Mollusca is incorrect. This appears to be unlikely, however; the placement of bi- valves within the Mollusca is strongly supported by com- parative morphology (BRUSCA and BRUSCA 1990; see p. 762). Another explanation is that rates of both substi- tution and rearrangement in mtDNA are or have been grossly different in the lineages leading to Katharina and Mytilus. If true, then both rates must have accelerated in the lineage leading to Mytilus, since its sequence and gene arrangement are more derived than Katharina's. We have analyzed all published mtDNA gene arrange- ments using cladistic methods (HENNIG 1966); none of the gene arrangements shared between Katharina and Drosophila mtDNAs supports their having a common ancestor to the exclusion of Mytilus (J. L. BOORE and W. M. BROWN, manuscript in preparation; BOORE and BROWN 1994). Expansion of the mtDNA data set and comparisons of conserved nuclear genes from these and additional taxa may clarify this.
In all vertebrate mtDNAs examined tRNA"u(UURI is between I-rRNA and N D I , whereas in insect mtDNAs (CLARY and WOLSTENHOLME 1985a; CROZIER and CROZIER 1993; PAsHLmand KE 1992; HSUCHEN et all984) tRNA'"'cw is at this location (see Figure 6). Although codon switch- ing was invoked to explain this (CANTATORE et al. 1987a), the Katharina and Mytilus gene arrangements suggest an- other, more plausible explanation. In Katharina the ar- rangement is l-rRNA-tRNA'"~Cw-tRNA'"(~)-~l; in Myti- lus it is tRNA'"(Cw-tRNA'"(m-hDl (I-rRNA is located elsewhere). The arrangement in which both leucine tRNA genes are between ML" and h D I may represent the primitive coelomate condition. Iftrue, then in the lineage leading to vertebrates tRNA'"(cciN) must have translocated to a position between tRNAMAGy) and hD5 and in the lineage leading to insects tRNA"(m' must have translocated to a position between COl and C02. We believe this hypothesis to be more plausible than codon switching, because it in-
W N UUR
I-rRNA N)1 L L
Vertebrates Arthropods Molluscs UUR W N CUN UUR
I-rRNA L W1 L I-rRNA M1 L I-rRNA W1 L
AGY CUN
N)4 H S N)S H N)4 NDS N)4 H NDS L
UCN UUR uuu CO1 CO2 UNK D F? CO1 m2 L c o l CO2 D S
FIGURE 6.-Evolutionary hypothesis for the positioning of the leucine tRNA genes in mollusc, vertebrate and insect mtDNAs. The positions of tRNAku(C"N) and tRNA'cUfUuR) relative to flanking genes are depicted for each of these mtDNAs. The gene arrangement in the hypothetical ancestor of vertebrates, arthropods and molluscs is depicted at the top. The molluscan arrangements, shown on the right, are unchanged (at least in this respect) from the ancestral state. In the lineage leading to vertebrates, tRNA'cu(CUN) was translocated to a new position be- tween and ND5, as shown on the left. In the lineage leading to arthropods, however, the other tRNA"" gene [ tRNA'eu(""R)] was translocated to a new position, between GO1 and CO2, resulting in the arrangements shown in the middle. Genes are designated as in Figure 1.
vokes a frequently observed phenomenon (tRNA transpe sitions; see MORITZ et al 1987), rather than one that (while theoretically possible) has not been observed elsewhere among metazoan mtDNAs.
We anticipate that as more metazoan mtDNA gene arrangements become known, it will be possible to dis- cern other patterns of evolutionary rearrangement. At least two factors suggest that gene arrangement com- parisons will be useful for inferring the phylogeny of major metazan groups. (1) The very large number of theoretically possible arrangements makes it unlikely that various organisms would convergently adopt iden- tical gene orders; thus, shared derived gene orders are very likely to indicate a common evolutionary history. (2) In general, rearrangements appear to occur infre- quently, on a time scale that is appropriate for address- ing these ancient divergences.
This work was supported by grants from the National Institutes of Health (GM30144; GM07544), the National Science Foundation (BSR-9107306; DEB-9220640) and the H. H. Rackham Graduate School of the University of Michigan. We thank R. Cox for collecting and purifymg mtDNA from K. tunicata, L. LYNNE DAEHLER for expert technical assistance, and T. COLLINS, S. FUERSTENBERG, R. HOFFMANN, E. F. KRAus and G. P. NAYLOR for many helpful comments. The Katharina mtDNA sequence has been placed in GenBank under accession num- ber U09810.
LITERATURE CITED
ANDERSON, S., A. T. BANNER, B. G. BARRELL, M. H. L. DE BRUIJN, A. R. COUL~ON et al., 1981 Sequence and organization of the human mitochondrial genome. Nature 490: 457-465.
ANDERSON, S., M. H. L. DE BRUIJN, A. R. COULSON, I. C. EPERON, F. SANCER et al., 1982 Complete sequence of bovine mitochondrial DNA
J. Mol. Biol. 156 683-717. conserved features of the mammalian mitochondrial genome.

436 J. L. Boore and W. M. Brown
ARNASON, U., and E. JOHNSON, 1992 The complete mitochondrial DNAsequence of the harbor seal, Phoca vitulina. J. Mol. Evol. 34:
ARNASON, U., A. GULLBERC and B. WIDEGREN, 1991 The complete nucleotide sequence of the mitochondrial DNA of the fin whale, Balaenoptera physalus. J. Mol. Evol. 33: 556-568.
AZEVEDO, J. L., and B. C. H m , 1993 Molecular characterization of lengthy mitochondrial DNA duplications from the parasitic nematode Romanomermis culicivorax. Genetics 133: 933-942.
BATTEY, J., and D. A. CLAYTON, 1980 The transcripion map of human mitochondrial DNA implicates transfer RNA excision as a major processing event. J. Biol. Chem. 255: 11599-11606.
BATUCAS, B., R. GARESSE, M. CALLEJA, J. R. VALVERDE and R. MARCO, 1988 Genome organization of Artemia mitochondrial DNA. Nucleic Acids Res. 16: 6515-6529.
BIBB, M. J., R. A. VAN ETTEN, C. T. WRIGHT, M. W. WALBERG and D. A. CLAYTON, 1981 Sequence and gene organization of mouse mi- tochondrial DNA. Cell 26: 167-180.
BIRD, A. P., 1986 CpGrich islands and the function of DNA methy- lation. Nature 321: 209-213.
BOGENHAGEN, D. F., S. S. CAIRNS and B. K. You, 1985 Nucleotide se- quences involved in the control of transcription and displacement loop DNA synthesis for Xenopus leavis mtDNA, pp. 175-182 in Achievements and Perspectives of Mitochondrial Research, Vol. II: Biogenesis, edited by E. QUAGLIARIELLO et al. Elsevier Science Pub- lishers, New York.
BONITZ, S. G., R. BERLANI, G. CORUZZI, M. LI, G. MACINO et al.,
Acad. Sci. USA 77: 3167-3170. 1980 Codon recognition rules in yeast mitochondria. Proc. Natl.
BOORE, J. L., and W. M. BROWN, 1994 Mitochondrial genomes and the phylogeny of mollusks. Nautilus (in press).
Buss, 1992 Class-level relationships in the phylum Cnidaria: evi- dence from mitochondrial genome structure. Proc. Natl. Acad. Sci. USA 8 9 8750-8753.
BROWN, W. M., 1980 Polymorphism in mitochondrial DNA of hu- mans as revealed by restriction endonuclease analysis. Proc. Natl. Acad. Sci. USA 77: 3605-3609.
BROWN, W. M., 1985 The mitochondrial genome of animals, pp. 95- 130 in MolecularEvolutionary Genetics, edited by R. J. MACINTYRE. Plenum Press, New York.
BRUSCA, R. C., and G. J. BRUSCA, 1990 Invertebrates. Sinauer, Sun- derland MA.
CANTATORE, P., M. N. GADALETA, M. ROBERTI, C. SACCONE and A. C. WIL- SON, 1987a Duplication and remoulding of tRNA genes during the evolutionary rearrangement of mitochondrial genomes. Na- ture 329: 853-855.
CANTATORE, P., M. ROBERTI, P. MORISCO, G. RAINALDI, M. N. GA~ALETA et al., 19871, A novel gene order in the Paracentrotus lividus mitochondrial genome. Gene 5 3 41-54.
CANTATORE, P., M. ROBERTI, G. RAINALDI, M. N. GADALETA and C. SAC- CONE, 1989 The complete nucleotide sequence, gene order and genetic code of the mitochondrial genome of Paracentrotus livi- dus. J. Biol. Chem. 264: 10965-10975.
CHANG, Y.-S, F.-L. HumcandT.-B. Lo, 1994 The complete nucleotide sequence and gene organization of carp (Cyprinus carpio) mi- tochondrial genome. J. Mol. Evol. 38: 138-155.
CHUNG, C. T., and R. H. MILLER, 1988 Arapid and convenient method for the preparation and storage of competent bacterial cells. Nucleic Acids Res. 16: 3580.
CLARY, D. O., and D. R. WOLSTENHOLME, 1985a The mitochondrial DNA molecule of Drosophila yakuba: nucleotide sequence, gene
CLARY, D. O., and D. R. WOLSTENHOLME, 1985b The ribosomal RNA organization, and genetic code. J. Mol. Evol. 2 2 252-271.
genes of Drosophila mitochondrial DNA. Nucleic Acids Res. 13: 4029-4044.
CLAYTON, D. A,, 1991 Replication and transcription of vertebrate mi- tochondrial DNA. Annu. Rev. Cell Biol. 7: 453-478.
CLAKON, D. A,, 1992 Transcription and replication of animal mito- chondrial DNA. Int. Rev. Cytol. 141: 217-232.
COLLINS, D. W., and T. H. JUKES, 1993 Relationship between G + C in silent sites of codons and amino acid composition of human proteins. J. Mol. Evol. 36: 201-213.
CROZIER, R. H, and Y. C. CROZIER, 1993 The mitochondrial genome of the honeybee Apis mellifera: complete sequence and genome organization. Genetics 133: 97-117.
493-505.
BRIDGE, D., c . W. CUNNINGHAM, B. SCHIERWATER, R. DESALLE and L. W.
CUMMINGS D. H., and J. M. DOMENICO, 1988 Sequence analysis of mito- chondrial DNA from Pcdospora ansoina. J. Mol. Biol. 204: 815-839.
DE BRUIJN, M. H. L., 1983 Drosophila melampfmmitochondrid DNA, a novel organization and genetic code. Nature 304: 234-241.
DE GIORGI, c . , C. LANAVE, M. D. MUXI and C. SACCONE, 1991 Mito- chondrial DNA in the sea urchin Arbacia lixula: evolutionary inferences from nucleotide sequence analysis. Mol. Biol. Evol. 8
DEL SAL, G., G. MANFIOLETTI and C. SCHNEIDER, 1988 A one-tube plas- mid DNA mini-preparation suitable for sequencing. Nucleic Acids Res. 1 6 9878.
DESJARDINS, P., and R. MOMS, 1990 Sequence and gene organization of the chicken mitochondrial genome: a novel gene order in higher vertebrates. J. Mol. Biol. 212: 599-634.
DESJARDINS, P., V. RAMIWZ and R. MouIs, 1990 Gene organization of the Peking duck mitochondrial genome. Curr. Genet. 17: 515-518.
D'ONOFRIO, G., D. MOUCHIROUD, B. AISSANI, C. GAUTIER and G. BERNARDI, 1991 Correlations between the compositional properties of hu- man genes, codon usage, and amino acid composition of pro- teins. J. Mol. Evol. 32: 504-510.
DROUIN, G., 1991 Nonrandom CpG mutations affect the synonymous codon usage of moderately GGrich single copy actin genes.
DUBIN, D. T., and C.-C. HSUCHEN, 1984 Sequence and structure of a methionine transfer RNA from mosquito mitochondria. Nucleic Acids Res. 12: 4185-4189.
DUBIN, D. T., C . 4 . HSUCHEN, G. R. CLEAVES and K. D. TIMKO, 1984 Se- quence and structure of a serine transfer RNA with GCU antic- odon from mosquito mitochondria. J. Mol. BIOI. 176: 251-260.
DUBIN, D. T., C.-C. HSUCHEN and L. E. TILLOTSON, 1986 Mosquito mitochondrial tRNks for valine, glycine and glutamate: RNA and gene sequences and vicinal genome organization. Curr. Genet.
FORAN, D. R., J. E. HIXSON and W. M. BROWN, 1988 Comparisons of ape and human sequences that regulate mitochondrial DNA transcription and D-Loop DNA synthesis. Nucleic Acids Res. 1 6
FULLER, K. M., and E. ZOUROS, 1993 Dispersed length polymorphism of mitochondrial DNA in the scallop Placopecten magellanicus (Gmelin). Curr. Genet. 2 3 365-369.
GADALETA, G., G. PEPE, G. DE CANDIA, C. QUAGLIARIELLO, E. SBISA et al., 1988 Nucleotide sequence of rat mitochondrial NADH dehy- drogenase subunit 1. GTG, a new initiator codon in vertebrate mitochondrial genome. Nucleic Acids Res. 16: 6233.
GADALETA, G., G. PEPE, G. DE CANDIA, C. QUAGLIARIELLO, E. SBISA et al., 1989 The complete nucleotide sequence of the Rattus norue- gicus mitochondrial genome: cryptic signals revealed by compara- tive analysis between vertebrates. J. Mol. Evol. 28: 497-516.
GARESSE, R., 1988 Drosophila melanogaster mitochondrial DNA gene organization and evolutionary considerations. Genetics 118 649-663.
GAREY, J. R., and D. R. WOLSTENHOLME, 1989 Platyhelminth mitochon- drial DNA evidence for early evolutionary origin of a tRNA"'AGN that contains a dihydrouridine arm replacement loop and of serine-specifymg AGAand AGG codons. J. Mol. Evol. 28: 374-387.
HARRISON, R. G., 1989 Animal mitochondrial DNA as a genetic marker in population and evolutionary biology. Trends Ecol.
HENNIG, W., 1966 Phylogenetic Systematics. University of Illinois
HIMENO, H., H. MASIKI, T. UWAI, T. OHTA, I. KUMAGAI et al., 1987 Un- Press, Urbana, Ill.
usual genetic codes and a novel gene structure for tRNASer(*"' in starfish mitochondrial DNA. Gene 56: 219-230.
HOFFMANN, R. J., J. L. BOORE and W. M. BROWN, 1992 A novel mito- chondrial genome organization for the blue mussel, Mytilus edu- lis. Genetics 131: 397-412.
HSUCHEN, C.-C., G. R. CLEAVES and D. T. DUBIN, 1983 A major lysine tRNA with a CUU anticodon in insect mitochondria. Nucleic AC- ids Res. 11: 8659-8662.
HSUCHEN, C.C. , R. M. KOTIN and D. T. DUBIN, 1984 Sequencesofthe cod- ing and flanking regions of the large ribosomal subunit RNA gene of mosquito mitochondria. Nucleic Acids Res. 1 2 7771-7785.
JACOBS, H. T., D. J. ELLIOTT, V. B. MATH and A. FARQUARSON, 1988 Nucleotide sequence and gene organization of sea urchin mitochondrial DNA. J. Mol. Biol. 202: 185-217.
515-529.
J. Mol. EvoI. 33: 237-240.
10: 701-707.
5841-5861.
Evol. 4 6-11.

Chiton Mitochondrial Genome 437
JANKE, A,, and S. P&o, 1993 Editing of a tRNA anticodon in mar- supial mitochondria changes its codon recognition. Nucleic Acids Res. 21: 1523-1525.
JOHANSEN, H., P. H. GunDAL and T. JOHANSEN, 1990 Organization of the mitochondrial genome of Atlantic cod, Gadus morhua. Nucleic Acids Res. 18: 411-419.
JUKES, T. H., and S. OSAWA, 1990 The genetic code in mitochondria and chloroplasts. Experientia 46: 1117-1 126.
KING, T. C., and R. L. Low, 1987 Mapping of control elements in the displacement loop region of bovine mitochondrial DNA. J. Biol. Chem. 13: 6204-6213.
KYTE, J., and R. F. DooLITTLE, 1982 A simple method for displaying the hydropathic character ofa protein. J. Mol. Biol. 157: 105-132.
LISZEWSKI, M. K., V. KUMAR and J. P. ATKINSON, 1989 “Midiprep” iso- lation of plasmid DNA in less than two hours for sequencing, subcloning and hybridizations. Biotechniques 7: 1079-1081.
MCCRACKEN, A,, I. UHLENBUSCH and G. GELLISSEN, 1987 Structure of the cloned Locusta migratoria mitochondrial genome: restriction mapping and sequence ofitsND-1 (URF-1) gene. Curr. Genet. 11: 625-630.
MONTOYA, J., D. OJU and G. ATTARDI, 1981 Distinctive features of the 5”terminal sequences of the human mitochondrial mRNAs. Na- ture 290: 465-470.
MONTOYA, J., T. CHRISTIANSON, D. LEVENS, M. RAFXNOWITZ and G. A n m I , 1982 Identification of initiation sites for heavy-strand and light- strand transcription in human mitochondrial DNA. Proc. Natl. Acad. Sci. USA 79: 7195-7199.
MOIUTZ, C., and W. M. BROW, 1986 Tandem duplications of D-loop and ribosomal RNA sequences in lizard mitochondrial DNA. Science 233 1425-1427.
MORITZ, C., and W. M. BROW, 1987 Tandem duplications in animal mitochondrial DNAs: variation in incidence and gene content among lizards. Proc. Natl. Acad. Sci. USA 84: 7183-7187.
MORITZ, C., T. E. DOWLING and W. M. BROWN, 1987 Evolution of ani- mal mitochondrial DNA relevance for population biology and systematics. Annu. Rev. Ecol. Syst. 18: 269-292.
OHYAMA, K., Y. OGURA, K ODA, K. YAMATO, E. OHTA et al., 1991 Evo- lution of organellar genomes, pp. 187-198 in Evolution ofLife: Fossils, Molecules, and Culture, edited by S. OSAWA and T. HONJO. Springer-Verlag, Tokyo.
OJAIA, D., C. MERKEL, R. GELFAND and G. A n m I , 1980 The tRNA genes punctuate the reading of genetic information in human mitochondrial DNA. Cell 22 393-403.
OJAIA, D., J. MONTOVA and G. AT~ARDI, 1981 tRNA punctuation model of RNA processing in human mitochondria. Nature 290: 470-474.
OKIMOTO, R., H. M. CHAMBERLIN,J. L. MACFARLANE and D. R. WOLSTEN- HOLME, 1991 Repeated sequence sets in mitochondrial DNA molecules of root knot nematodes (Meloidogyne): nucleotide se- quences, genome location and potential for host race identifica- tion. Nucleic Acids Res. 19: 1619-1626.
OKIMOTO, R., J. L. MACFARLANE, D. 0. Cumand D. R. WOLSTENHOLME, 1992 The mitochondrial genomes of two nematodes, Caenorh- abditis elegans and Ascaris m u m . Genetics 130: 471-498.
P f i ~ o , S., W. K. THOMAS, K. M. WHITFIELD, Y. KUMAZAWA and A. C . WIL- SON, 1991 Rearrangements of mitochondrial transfer RNA genes in marsupials. J. Mol. Evol. 33: 426-430.
PASHLEY, D. P., and L. D. K E , 1992 Sequence evolution in mitochon- drial ribosomal and ND-1 genes in lepidoptera: implications for phylogenetic analyses. Mol. Biol. Evol. 9: 1061-1075
PRITCHARD, A. E., J. J. SEILHAMER, R. M A H A L I N G A M , C. L. SABLE, S. E. VENUTI et al., 1990 Nucleotide sequence of the mitochondrial genome of Paramecium. Nucleic Acids Res. 18(1): 173-180.
ROE, B. A,, D.-P. MA, R. K. WIWN and J. F.-H. WONG, 1985 The com- plete nucleotide sequence of the Xenopus laeuis mitochondrial genome. J. Biol. Chem. 260: 9759-9774.
RUSSELL, G. J., P. M. B. WALKER, R. A. ELTON and J. H. SUBAK-SHARPE, 1976 Doublet frequency analysis of fractionated vertebrate nuclear DNA. J. Mol. Biol. 108 1-23.
SAMBROOK, J., E. F. FRITSCH and T. MANIATIS, 1989 Molecular Cloning: A Laboratory Manual, Ed. 2. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
SANGER, F., S. NICKLEN and A. R. COULSON, 1977 DNA sequencingwith chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74: 5463- 5467.
SMITH, M. J., D. K. BANFIELD, K. DOTEVAL, S. FORSKI and D. J. KOWBEL, 1989 Gene arrangement in sea star mitochondrial DNA dem- onstrates a major inversion event during echinoderm evolution. Gene 76: 181-185.
SMITH, M. J., D. K. BANFIELD, K. DOTEVAL, S. FOR~KJ and D. J. KOWBEL, 1990 Nucleotide sequence of nine proteincoding genes and 22 tRNAs in the mitochondrial DNA of the sea star Pisaster ochra- ceus. J. Mol. Evol. 31: 195-204.
SMITH, M. J.. A. ARNDT, S. GORSKI and E. FAJBER, 1993 The phylogeny of echinoderm classes based on mitochondrial gene arrange- ments. J. Mol. Evol. 36 545-554.
SNYDER, M., A. R. FRASER, J. LAROCHE, K. E. GARTNER-KEPKAY and E. ZOUROS, 1987 Atypical mitochondrial DNA from the deepsea scallop Placopecten magellanicus. Proc. Natl. Acad. Sci. USA 84: 7595-7599.
SPRINZL, M., N. DANK, S. NOCK and A. SCHON, 1991 Compilation of tRNA sequences and sequences of tRNAgenes. Nucleic Acids Res. 17: rl-rl71.
STORMO, G. D., T. D. SCHNEIDER and L. M . GOLD, 1982 Characteriza- tion of translation initiation sites in E. coli. Nucleic Acids Res. 10:
TZENG, C.S., C.-F. HUI, S.-C. SHEN and P. C. HUANG, 1992 The com- plete nucleotide sequence of the Crossostome lacustre mitochon-
Nucleic Acids Res. 20: 4853-4858. drial genome: conservation and variations among vertebrates.
UHLENBUSCH, I., A. MCCRACKEN and G. GELLISSEN, 1987 The gene for the large (16s) ribosomal RNA from the Locusta migratoria mi- tochondrial genome. Curr. Genet. ll: 621-638.
WARRIOR, R., and J. GALL, 1985 The mitochondrial DNA of Hydra attenuata and Hydra littoralis consists of two linear molecules. Arch. Sci. 38: 439-445.
WOUTENHOLME, D. R., 1992 Animal mitochondrial DNA structure and evolution. Int. Rev. Cytol. 141: 173-216.
WOLSTENHOLME, D. R., J. L. MACFARLANE, R. OKIMOTO, D. 0. CLARY and J. A. WAHLEITHNER, 1987 Bizarre tRNAs inferred from DNA se- quences of mitochondrial genomes of nematode worms. Proc. Natl. Acad. Sci. USA 8 4 : 1324-1328.
WRIGHT, J. W., C. SPOLSKV and W. M. BROWN, 1983 The origin of the parthenogenetic lizard Cnemidophorus laredoensis inferred from mitochondrial DNA analysis. Herpetologica 39: 410-416.
YOKOBORI, S., T. UEDA and K. WATANABE, 1993 Codon AGA and AGG are read as glycine in ascidian mitochondria. J. Mol. Evol. 36: 1-8.
YOKOGAWA, T., Y.4. WATANABE, Y. KUMAWWA, T. UEDA, I. Hlwo et al.,
chondrial tRNRer(UCN). Nucleic Acids Res. 19: 6101-6105. 1991 A novel cloverleaf structure found in mammalian mito-
ZEMRING, C. E., C. MORITZ, A. HEIDEMAN and R. A. STURM, 1991 Par- allel origins of duplications and the formation of pseudogenes in mitochondrial DNA from parthenogenetic lizards (Heteronotia binoei: Gekkonidae). J. Mol. Evol. 33: 431-441.
ZIAIE, Z., and Y. SUYAMA, 1987 The cytochrome oxidase subunit I gene of Tetrahymena: a 57 amino acid NHZ-terminal extension and a 108 amino acid insert. Curr. Genet. 1 2 357-368.
2971-2996.
Communicating editor: A. G. CLARK
APPENDIX
Sequence of K. tunicata mtDNA is given in Figure 7 (pp. 438-443). Orientation is as in Figure 1. Gene products are shown above the sequence; arrows indicate direction of transcription. Genes are abbreviated as in the text, except that three letter amino acid designations are used to denote the corresponding tRNA genes. tRNA gene positions are denoted by overlining; anticodons are underlined. Precise termini of rRNA genes are unknown; dashes indicate approximate termini. Asterisks denote stop codons and abbreviated stop codons. Protein genes are translated, using single letter amino acid designations; these are centered below the 2nd n t of the corresponding codon. N denotes an unidentified nt; X denotes an unidentified amino acid.

438 J. L. Boore and W. M. Brown
COl-") A T G C G A T G A A T T T T T T C I A ~ T C A T A A A G A T A T T G G M R W I F S T N H K D I G T L Y I L F G I W A G L V G T A L S L L
T T C G T G C A G A G C T A G G T C A A C C A ~ T T T A T T G G G G G T I R A E L G Q P G A L L G D D Q L Y N V I V T A H A F V M I F F L V
T A n ; C C T A T A A T A A T T G G G G ~ C ~ T A G T G C C ~ T A A T ~ T ~ G n ; C C G ~ A n ; G C T T T C C ~ ~ T T A A A T A A T A T R A G T T T ~ G A M P M M I G G F G N W L V P L M L G V P D M A F P R L N N M S F W
C T ~ T G C C T C C G G C A T T A T G T C T T T T G T T A G C T T C A G G G L L P P A L C L L L A S G A V E S G A G T G W T V Y P P L A G N V
GGCATGCTGG~;GATCTGTTGA~T~TATTTTTT~TTACATTTAG~GGA~ATCGTCTAT~AGG~T~TAATTTTATTACTACAAT~;TAAA G H A G G S V D L A I F S L H L A G V S S I L G A V N F I T T I V N
T A T A C G A A G A G A G A G C G A T T A C C T P T C T M R S E G M Q L E R L P L F V W S V K I T A I L L L L S L P V L A
G G A G G A A T T A C A A T A T T G T T A A C T G A T C G T A A T T T T A A T A G T G G I T M L L T D R N F N T S F F D P A G G G D P I L Y Q H L F W
T T T T T G G P C A T C C T G R A G T ~ A T A T C T T A A T T T T A C C T C F F G H P E V Y I L I L P G F G M I S H I V M H Y S S K K E T F G T
TTTAGGAATAATTTATGCGATATTAGCTATTGGTCTTTT~GATTTATn;TTTGAGCACATCATAn;T~GTAGTAGGGATGGAT~TGAT~TCG~T L G M I Y A M L A I G L L G F I V W A H H M F V V G M D V D T R A
TATTTTACTGCGGCTACTATGATTATTGCTGTTCCTACTGGAATTAAGATT~TA~G~T~TAC~TTTA~G~CGGATTGGPAGGGAGACTC Y F T A A T M I I A V P T G I K I F S W L A T I Y G A R I G S E T
CTATAATGn;GGCTITAGGATTTATTTT~TATTTA~G~G~~~A~GGTATT~G~ATC~TTCTTCTTTAGATATTAT~TACAn;ATAG P M M W A L G F I F L F T V G G L T G I V L S N S S L D I M L H D S
G T A T T A C G T A ~ T G C T C A T T T T C A T T A T ~ T ~ A T C G A T A Y Y V V A H F H Y V L S M G A V F A L F G A F N Y W Y P L M T G L
A G T C T T C A T G A A C G A T G A T T ~ ~ A ~ G A T A ~ T ~ A ~ ~ T A A T T T A A C ~ T T T T ~ C T C ~ C A C T ~ T T G G G G T T A A G A G G G A T G C S L H E R W T K S H F V V M F L G V N L T F F P Q H F L G L S G M
C T C G A C G T T A T P C T G A ~ A T C T G A ~ G ~ A T A T T ~ T ~ T G ~ G ~ T ~ T C T A T T G G T T C T T T A A T T T C T ~ ~ T G G C ~ ~ T ~ T T T T T A T A T T P R R Y S D Y P D C Y I K W N V V S S I G S L I S F V A V L F F M F
T A T C G T T T G G G A G ~ T T A ~ T T C T C A G C G G G G G G T G A T T C G I V W E S L V S Q R G V I W S S H L S S A L E W D N S I P L G F T
tRNA phe (VnO) ? +- G A T C T G A T G A A G G G G A A G T T A T n ; T T T G T T A A T ~ ~ T T ~ A ~ G T A G T T T A T ~ C ~ A n ; T C C A ~ T ~ T A G C G G G G T T ~ ~ G T A T T G ~ G G D L M K G K L L F V N L I '
tRNA asp -> G A T T A T A A A T T A A ~ T A G G A G A A A T T A G ~ ~ T A A T G A A G ~ T n ; ~ ~ T ~ T A G T T A C ~ ~ G T A T ~ C T T T G T A T T ~ G ~ A A T A G A T
T T A T T T I A T T T A T T A T T G T A G T W U L T T A n ; T G T T A T T T G A T A G ~ A T ~ ~ T ~ T T G G ~ G T T R A G T T ~ T T A T ~ T T T T ~ T A T C
T T ~ T ~ A G C T I ~ ~ G T A T T T T G T T A T A A G A A R A T T T
A A A C T A A A T T T T G T T G G T T ~ T ~ A ~ ~ T A G C A C ~ ; T A G C ~ ~ T T T G T T ~ ; T G A G T G T A T A ~ T ~ C T C C ~ G A T G G ~ T ~ C T C C ~ ~ T A G
G G G G T ' P T A A T T A G A A A T T A ~ A ~ ~ T T T A ~ T G T A T ~ T ~ G A T T T A T ~ T T T T G G T G T G T A T T T ~ T A T ~ A T G T A T C A C C ~ A ~ G ~ ~ A A
A A G A A A A T G G C I T ~ T G A G T G A G ~ T ~ C A G G A c02 . M A F W S Q W G F Q D G A S P I M E Q L I F F H D H A M L I L
T T A T A A T T A T T A G T P T A T T G C T T A T ~ ~ G G T T T ~ T T ~ T A A A T A A T T ~ T T T ~ T T C G ~ T T C T A C A T T A ~ ~ ~ ~ T ~ A ~ T ~ T A T G I M I I S L 1 , S Y G A V S L M N N S F L S R S T L E S Q E I E I V W
GAffATCTn;CCTGCGGTA~~~AATTTTTTTGGCATTTCC~CTTTACAATTAT~TATTTGTTAGATG~Tn;~~~T~T~AACTATTAAG T I L P A V V L I F L A F P S L Q L L Y L L D E L E E P A L T I K
G T G G T G G G G C A T C A A T G A T A C T G G A G G T A T G A G T A T T C T T T T T T A C A T A T T C G A A G A C T T A G ~ G A G ~ A G V V G H Q W Y W S Y E Y S D F L N L E F D S Y M I S L E D L E E G
A T T A C C G G C T A ~ A G A A G T G A T C A C C G ~ G G G T G G ~ ~ A T G ~ A ~ A A A G ~ C ~ ~ T ~ A ~ T A C T G C T ~ ~ T ~ A C T T C A ~ C ~ G ~ C T G T D Y R L L E V D H R S V V P M K T K V R V L V T A A D V L H S W T V
100
200
300
400
500
600
700
800
900
1000
1100
1200
1300
1400
1500
1600
1700
1800
1900
2000
2100
2200
2300
2400
2500
2600
FIGURE 7.-Sequence of K. tunicata mtDNA.

Chiton Mitochondrial Genome 439
T C ~ T C I T ~ ~ ~ ~ T ~ ~ ~ ~ A ~ A A G A T ~ A A T T ~ C C I ~ G T I ~ A T ~ ~ ~ ~ T G ~ P S L G V K A D A V P G R L N Q L S F F A N Y P G V F Y G Q C S E
A ~ G I Y ; G G G C A A A C C A T T C T ~ A T A C C A A T A C ~ T I G T G T T A G A ~ A ~ G A T ~ ~ ~ A T I A A A ~ T T A T A T T I A A ~ ~ ~ - I C G A N H S F M P I V L E V V D S S S F I K W I M F N G E A * * *
tR?JA 8ar (UCU) ?e ATP8808--") . T A A A T T r A T A A G A A A T T A G P T A G A T R T A T ~ ~ T I T A T I T ~ T C T T ~ ~ T I A ~ ~ A ~ C T r T A T ~ A ~ T I A ~ ~ T A T ~ T I G A A T
M P Q L A P M N W I
~ A ~ T T ~ T ~ ~ ~ ~ G ~ ~ ~ ~ C I A ~ ~ ~ T C I ~ ~ ~ ~ T C T ~ ~ T F L L L F F W S N V F C L G V C V W W V K S S S Y S F L S K S V S
A!fP88e6 d NCCCCm T r A T I T I ~ A T G A G T I T G A T f f i ~ ~ T A A T A G A T A T T T I T T C T ~ T I T G A ~ T A A T ~ T I T T A ~ ~ m A T A C C A C ~ m ~ A T T X P F Y F K K W V W * * * M M M D I F S S F D D N N F T L F S F S Y
T G A T T ~ T T I ~ ~ A T G T ~ T A A T T A G T C ~ ~ ~ A A T A ~ T T I T ~ T I A ~ T C I C G T T I T T A T ~ ~ ~ A A T G A T ~ A T I T ~ T ~ T I T L I W V L G L A P V L I S Q N M F W F S K S R F Y S M I L F P K N F
T A ~ G G G C A G G T T A T G T ~ A ~ ~ T A T T A G ~ T T A ~ ~ T A T U ; T G A T T A ~ T I T ~ A C T r T T A A m T A G T I A A T T r A T C A M G G Q V M R A T G K N I S G F I N M V I S I F L L L I L V N L S
G G G C T T C T T C C T I A T G T C T T A ~ T T A A G A A G R C A T T T T ~ ~ ~ ~ ~ T T ~ T ~ T I ~ T ~ ~ ~ C I C T I T ~ T C I T T I C ~ ~ T T T A T G L L P Y V F S L S S H L S F A F C F G L P F W L S L I F S G D L
A T ~ T T C T G A G G I ~ T G T A T I ~ C T A G G G G G G C Y V S E V V S H L L P S G A P G I L N P F L V L V E T V S I S V R P
T A I T A C I T T A T C I ~ C G A C T T G C T G C T A A T A T A A G G C I T L S V R L A A N M S A G H I I L G L I G S Y L S V G L F C Y P
A T T C T T G m T ~ T T G T T A ~ A T T r ~ C A A G ~ T T r T A T T r T ~ A ~ ~ T T G G T ~ T ~ T A A ~ ~ C A T A T A T T r T ~ C ~ A T T A G T r A I L V L G L L L F V Q V F Y F L F E I G V S L I Q A Y I F S L L V
f" tRNA phe GGTIATACTCAGATGAT~ATIAATTAAAmATACCATAATTATTmATTCIATAGTAACIACG~T~~G~TCCATT~A~~~T~A~G S L Y S D D H P Y * * *
C T C I G A T A T A A G C T A C T T A G T T A A G T A A A T C A T A T A G T A T * T F W M T V S I F L M S L L T L S I F L N F L N S Q
G A T I T T TTTGATrAATTGTTGTAAGTGTTAT~TTITATTrAGIA~~TCCC~CCGATAGTITCCATT~CC~TCT~C~T~~TGIA N K Q N I T T L T M I K N L L G E G G I T E M W G L D L S K F T Y
G A G P C C I G T A T A A A G A G A T C I ~ C I G ~ T A T A T T A T T T T L G T Y L S S K S T M N N S L N I M F W M N F F F N N I I S H S I
T T A A G T T T G T A T ~ ~ T I G C I ~ ~ A A C ~ ~ ~ T ~ ~ T A T T ~ T I T A ~ T A T T A T ~ A T T ~ G I ~ G T I G K L E L K F L K Y T I A A G T L T A F T A M L K N M M S I L P L T S
A G G G G G T T A A A A T A A G T C A T G ~ C T C T ~ A A T I ~ C C ~ G C I ~ A ~ A T G G T ~ T ~ ~ A A T A A T ~ ~ A T C T ~ ~ C A G ~ T P T L I L W S F V A G S T I A G A A L M T L P A I I S N D E D S T
T C T G A C T A C P C P T G A T T G T G A T T T I T G A T ~ G ~ T ~ T ~ A T A ~ G ~ f f i A ~ A T C ~ ~ G ~ ~ A ~ G T I ~ ~ T A T A T I ~ S V V S S Q T M K S W F I F L S L R A S Y S A T L I T A L F M N L
G A A A T T A G A A A A T T A T T I T G G T T A A A T A T I A T I A ~ ~ T ~ T T ~ A ~ T ~ A G A G T ~ T ~ T G C T A T ~ ~ ~ ~ C ~ ~ G ~ ~ T I T G S I L F N N Q N F M M M E I I V D K S Y F G A M F P F G C L A L N A
C A A T G T I G A A G C A T G T T C T T A T T I G A A T ~ A T ~ ~ ~ C ~ A ~ C ~ A T A T ~ T r G T r A T I A ~ ~ ~ ~ T A A T A T T ~ C ~ C I N F C T S S F P M Q I W L H S M K R I D Q N N N H S H I I N G A
G C A A A U ; A A T A G G A G G G C T T I T A A A T A T T ~ A ~ A G I G A A T ~ T G ~ T A ~ T A ~ ~ T A ~ C ~ ~ T C T I A C I C T I A A T A T I A T ~ T C C T ~ T C L F L L A K F M A H T F L H F L A L S F L G L S V S L M M V G L
T G G C T T A A A G T G T T ~ ~ A T C I ~ T T C ~ T I T ~ T ~ T G ~ ~ T A T T A C T A T ~ ~ ~ ~ A T T C T ~ ~ T ~ ~ A T I ~ T C Q S L T S L A I I K K L D T E F N A S I G A M V M T M S S I I M L S
T T G I ~ A A A T A A A T G T T C T ~ G ~ T ~ T I ~ A T r ~ T ~ C I C ~ G ~ ~ T ~ T ~ G I A ~ G A G ~ ~ T ~ ~ ~ C T S F L H S N S L F P Y F Q I L L F V G A T V L T S S H V L A S V
A G G G G T A G G U ~ C G C A T C ~ ~ T G G T ~ T ~ A ~ T C I T T ~ ~ ~ ~ ~ A T ~ C ~ C T ~ T A ~ T T A T ~ ~ A ~ A G ~ A A T G T I P T P A A M A A P L W S C F P L Q A S K T M G A L L I M S I T L T
A T T G I T G G G G T T C T T C A T A T A A A A A G M ; C A T T G A A G T G C C M T P T S W M F L A N F H G Q S F T V S I A L L I A V D G I R N M F
FIGURE 7.-Continued.
2700
2800
2 900
3000
3100
3200
3300
3400
3500
3600
3700
3800
3900
4000
4100
4200
4300
4400
4500
4 600
4700
4800
4900
5000
5100

440 J. L. Boore and W. M. Brown
A T ~ G T T A T T A G C C C G T A A A G A ~ A T A G ~ ~ G A T A A T A G A T T A C T A A G C ~ A T A C T A G C C ~ A G C C ~ T ~ C ~ ~ ~ T A A ~ A T M L G A G L S K Y N Q Y Y I V L C F S V L G L G D W G L L L T
T G C T A A ~ T T A ~ A G A ~ A ~ G A G P L ; T A C A A A G P L ; T A ~ ~ G A T T C A f f i T A A A T C G ~ G A T A A A G A T A T ~ ~ C T T A ~ A A ~ A L S P I F I L L N M S L V F L M V I W T F R K I F I D Q S M Y N
T T A G T A A A A A T T A T T A C T C T T G C G G A G A T A T A A C A T A C A A K T F I M V S A S I Y C V I A S F L A T T W N I L I P L T I T V S N
T A A A G T T G T A G A A A C A A A A T T C T A G T A A A A T A A T C T T G T T F N Y F C F E L L I I K N T T L T Y M A T S F S L L S H S L L I V
. f"-ND5 -tRNA hie TGTTGCTIGTGAAGTACTIAAAAATATTATIT~ATT~TAGATAA~TTGTACTTACTTTI~~~AA~TAATATT~TTAT~TTAATCTAA T A Q S T S L F M M K M - CAAATTGTT~TCTTAATAAAAGTTAAAAG~GTGCTG~~CAAT~~TIA~TTGTAATATAAATGGGTTIGAAATIA~~~AACAA * I T T S L L I L L Q A P I W H L F I V N Y Y L P K F N N F P N V F
A T G A A G A A A T T T ~ C C A T G T T G T ~ A ~ ~ ~ T ~ ~ T A G ~ T A T A A A C ~ C C G A A A ~ A A T ~ A T A A ~ ~ A ~ ~ T I A T C A G A A A T T T T ~ S S I K G H Q T N T F L F L T Y V A S V F A M L A L L I M L F K S
G A T A G A R A C T G T A C T A A T I A ~ ~ C T G C T T ~ C T I ~ ~ T T G A T T G A ~ A G C ~ T A T ~ ~ T ~ ~ A T A T T A A ~ G A T A ~ C T A I S V T S I M L G A E S L L N I S P P A A M N I S C M L F F S M S
A A A G T T G G A A C T A A T G A G A T A A A C C C T T T T G T A A T A A A A A F T P V L S I F G K T I F L S R S G S K E Y M M N S M S F L G S S S
AAAGACCGTGA~TTATTATTAATAAAGCTATTTTT~CCAAATTCAAATCTGCTGAGGA~CCT~A~ATGAGTCCTATATGACC~TGAG~ L G H A I M M L L A M K L G F E F S S L L G A L M L G M H G V S S
ATAAGCGATTAATGCITATGTCTCTIT~C~AGACAAATTAGACTTCTTAATACTCCTCCTACT~GCCAAGTGTGATAAT~TPL;TTI~TA~ Y A I L A K M D S Q R L C I L S S L V G G V L G L T I I F Y N L V
AAATIGTAT~GAAAATATTATTATTAATTAAT~TATTAATAC~T~CAC~A~TTTAffiAG~~T~TAGAATTATAGAGCC~TACAGATGGGC~ F N Y K F I N N N I T N I G Y G G L K L L L A A L I M S G A V P A E
C T A C A T G A G C T T T A G G G A G T C A T A G A T G A A A A G A G T A A A T V H A K P L W L H F S Y I P L K V L F V L I I F L W W F S I M A Q
A T A A G G T A T A A T T G G T A G G C T T A T T A T A A ~ A ~ ~ T T ~ T G A ~ A ~ A T G T ~ T ~ T ~ G A T ~ T A A T T A A ~ ~ ~ G C T C ~ ~ Y P M I P L S M M I L L N L H A N H S Y I F F I N I L F P L A G L
A T A ~ G T A G A T T A T T A T A T A T A T C C C ~ T I G A A G G C ~ T ~ ~ T ~ T A C C ~ C A T A T T A A A A T T ~ G A ~ A A T G T A ~ G A T T A A G G A ~ T I C ~ I T Y I M M Y M G A Q L R E P Q Y G W M L I L I L T P I L S A E F F
A A A T A T A T A A T G A A A A T P L ; T C T T T I T ~ A ~ ~ T A C T A T A A T A A T A A T ~ T T T ~ ~ A T I A ~ T T I A ~ ~ T C T T ~ T P L ; A T T A T I A T T I Y L S F L S K Q T F V M I I I F N L I M V N L L F F S V L N N N
T I T T A G A A T T I T A T A T C T G C T T A T ~ T I A T I ~ C C A ~ ~ T T ~ C A T C T T ~ T A T T A T T ~ T A C f f i ~ T T ~ C A C T ~ T C G T A A T T A K L I K Y S S M I M L G T I W C S L M M L P V S F N D S F X E Y N
ATTTTAGGGATGTTTATTGGTGA~TTTAATGTAATTAT~TAT~TGA~TT~GCTTATTTTGTAT~TITAT~TGCT~TTTATTA I K P I N M P S F F N L T I M F F M F H S I W S M K Y W N M K S N K
-ND4 . T I G T T A T A A C G G T C G A A A A T G C T G A T A T A A T A A A A A A A G C T M V T S F A S M * * * C K H I S I S S V Y D N G H A R I L I
ACTAATATTGCGAGCCCGATGCTAGCTTCn;AAGC~AGAGTAA~~A~TIATTAACCCT~ATIT~TATAAGGTITGATAGGGATACTA V L M A L G I S A E S A A L T L L V F I M L G E N S M L N S L S V L
ATAATAAAAAAATICTTACTATTAAGATTTCTAA~TAATAAGGT~TAAGAA~TGTTTIC~T~A~AAATIGTTATIA~GCTCT~TCCTA~ L L F I S V M L I E L A L L T N L L H K R Q L C I T M L A S I S S
;f"ND4L tRNA thr T A A R G C T C C A A G C G T G A A T A A T A T G G G T T ~ T T T A T C A T G T C T ~ T ~ G C ~ T A T A G C ~ T G G T C T T ~ ~ C ~ A T A ~ T ~ T T ~ L A G L T F L P N F F N M M
-tRNA ser (UCN)
T T I C T T T T A G T A T G C C G T T A A ~ G A A T ~ ~ C A C T T C T T I A T I G T I T ~ ~ A ~ ~ ~ ~ C C A ~ T A A G C G G ~ ~ ~ G T I G A T T ~ T C A G T * * * F N I T W D
C T C A T A T T T T m T G T I A G A G A T I A ~ T A A C A G ~ T A T A C A A A T ~ T ~ T T I ~ C A A T T A G A A T ~ ~ C T A ~ G G C T ~ G C W M K K T L P I M L F L L F Y V F T L I Q G I L I F P D E V P Q A
A C C A A T T C A A G T T A A A A G T ~ G A C ~ A C A C T A ~ C T T C A G T A T ~ ~ T G A G T I A C A G G G T ~ ~ T A ~ T ~ G A A T I T T C C T ~ ~ T ~ T T G I W T L L L F V S V L S W Y L I Q T V P Y F S I T R F K G L N I
5200
5300
5400
5500
5600
5700
5800
5900
6000
6100
6200
6300
64 00
6500
6600
6700
6800
6900
7000
7100
7 200
7 300
7400
7500
7600

Chiton Mitochondrial Genome 441
A A T G G G A C A A A A A A T A A T A ~ A A T ~ ~ T ~ ~ T A A T T T A T T ~ A A ~ A T C ~ ~ C A T f f i ~ A A T A ~ T 7700 L P V F F L I V I S M L L G V V G G L K N P I S R L I A Y A F L F Y
A C C A T T C T G G C T G G A T A T G T ~ T A A ( 3 T A G A G G G T 7800 W E P Q I H T P T V L P N A P I F N D P H A L L N P D L F V L L L
T A A A A A A A T T A ( 3 T A T T A C T ~ ~ C C C ~ f f i A ~ T T A G T G T G T A ~ T G A G T ~ T A G T A A C P T ~ ~ C T A T C P ( 3 T A ~ G A T T C C T A A ~ ~ G 7 900 L F I V M V V F G V L D K L T Y F S H I T V K E S D S N I G L P N
T T A G A C C C T G T T I C A T G T G T A ~ A ~ T G C T ~ A ~ A A T A A T ~ C A A G A T A A T ~ T G ~ T C G T G ~ A A A G ~ 8000 N S G T E H L F L I H L V S A A A I I F P A L Y H F S F F R T L T A
C G T I A T C T A C T G C A A A T C C C T C A ~ ~ A ~ G A A ( 3 T A ~ G ~ G ~ ~ T A T A A G ~ T C G C T ~ T A T T A A G T T T G T A A T A A C ~ ~ ~ C T C ~ 8100 N D V A F G G W I W Q V M T T G I Y P I A S M L N T I V T A G W F
A G A T A T C T G T C C P C A ~ A G T A C ~ A A C C G A T A A A A G C 8200 S M Q G W P L V Y G I F A T G M T L F L L I V G V X W T H I N V F
G A G C C A T A A T A T A T C C C G C G C ~ T A T G G A A A T A A A G G C A 8300 S G Y Y M G R G I H F Y L C I F F L S G G N A H A A R I L W G Y N V
C G T C T f f i A C A G A T A T G T G C A A C f f i A A G ~ A A G T T C G A T A ~ ~ T ~ T G T A T T ~ T f f i A A A G ~ A ~ T ~ T ~ ~ T A A T A A ~ A 8400 D R C I H A V S S F A L E I N T A Y H M A L F L G T L I Q I I L C
T A G G C ~ ~ G T G A C C C A T C A T ~ T T G A T A A 8500 L G L L S G F N W W I S L N S P S P L D I L S G N I I K F V P H S
f - " C s b G A T C G A A T T G G ~ T A A G C A T A T T T G T A G G G G C G G A G A G G T 8600 S R I P K L M * K Y P R L P G N S F Y C I K V V C I L V F F L I L A
A A T C T T A C A A T A T ~ ~ ~ T A A A ~ T T C C T A A G ~ T I ~ A A T T T C A T A ~ T A T T A ~ G T I T G T T C C T C P A 8700 L S V I I L L N F P S F L S I G L H N F F S N W L F M M K T Q E E L
ATAAAGTTCCAATGAAGTCTTT~TAAAGTATTAATAACC~AATACPGTT~ACA~~~CTTGAATT~~TTT~TAAATATT~TT 8800 L T G I F D S L Y L M L L W F V T S V P F F V Q I L N K K F M L N
T G G I A T I A A G G C T G T A T A A G C G A A T A T T A C T A A A A 8900 P M L A T V Y A F M V L L G G I Y I L F L I F S F W T S S G F F I
A A G A T A C T A A T A A A T A G C C T A T ~ T ~ A G T ~ T ~ ~ C P A A T T ~ T ~ A G T T ~ G A G G T A A T G ~ C A C ~ G ~ T I A ~ G A T A 9000 L I S I F L S M F L L L L G L S I P Q T L L P L S F A C S F I L S M
" N D C tRNA pro T T G T T A A T A A G A A T G T C A T A T T T C A ~ T ~ T f f i T T I A A G ~ T A T T ~ A T T ~ ~ C P A A A ~ T T ~ ~ T T ~ m G T T A T C P G A ~ A A T ~ G G T T 9100 T L L F T M * * * S L L L T
G T T A C T ATAAATTTT~GATA~TTTCATATT~TTATT~T~TACT~Gffi~TCTTAGTC~TTAATITTATTAATAGAT~TATC~~ 9200 T V M F K A S L N W M L I M L S V P L F S L W T L K M L L D Y R L R
G A G G A A A ~ A C C T C G A A C T C A A A T ~ C ~ ~ A T ~ G T A A ( 3 T T T T A ( 3 T C ~ T ~ G T T A G T T ~ T T ~ A A T C ~ C ~ T C C T A A 9300 P F T G R V W I F V F A I F T V K V W F L F N T L I W S D G A G L
A A A G A G A A T P G T C G P T A T T A T P e r C A T G M T ~ T T C T T G C ~ A T T C ~ A A ~ T A A T A G A ~ G ~ ~ C T T C ~ T A C T C G A T G T T ~ C C T 9400 F L I T T M M S M F L I S A Y E A L F L L A F S G S G Y E I N F G
G A T A C T A G T T C A G A T T C P C ( 3 T T C P ~ ~ ~ T ~ ~ T G T C T C T ~ T A A G C A G A C T A C A A T T C A T A T T A G T ~ A A T ~ G ~ A T I A ~ 9500 S V L E S E G E A F D F P A R N T E A L C V V I W M L L L P T M L I
T T C P G T I TCAGTATAGTAATTGATGGGATG~~AAAT~~TTIAAATTIAGGATGAAC~AGAGCA~~A~AG~T~TAACGTTACTIC 9600 S N W Y L L Q H S S I Q N L N L S S S L A A S S L L V L F L T V E
G T A A G A A A T G G ? T T G A G C A T G C A C G G ~ G C A ~ T A G A A 9700 Y S I T Q A V A R L A G L L A Y K S N S A W G A I M T G Y V S L A
G A T G T G C A T A G A C ~ T T A A A A T A T C 9800 S T C L F F L V G F N F Y W L T F D S W Y L S W L C L S L I L S L V
C T G G T G C T G T G T A A T A A G G A A G ~ G T T T G A A T T A T A A A W L T 9900 P A T Y Y P L K N S N Y L N N E E K L F L K L A D A L P Q A L G F
A A T I C T T A ~ T T G ~ G G T C T ~ T C G A A A C ~ A A T A T A ~ C ~ ~ C T I T C C G ~ C T A A T A A A G ~ T G C T A ( ~ T G C T A A T A A G A T A ~ T A T A G 10000 I S V K N P G K R F Q I Y G L G K R E L L T F F A V A L L I S I Y
. "Dl tRNA leu (UUR) G C A A T T A C T A T T C A A G ~ G A T A G A A A A A G A T A A G A A C A T A A C T A A A A 10100 A I V M W S F S L F M
FIGURE 7.Pontinued.

442 J. L. Boore and W. M. Brown
+ tRNA leu (CUN) [-----l-rmA-
T A T T A ~ G A G A T T ~ ~ T C G T A T A A T A A T T ~ A A A T T A T T T ~ C T T ~ T T T G C I ~ ~ T A A T A T A ~ A A A T T A T T T G T T A ~ T T T ~ ~ T T 10~00
C G T A C T A A A A C A T A A A T A A T T A G C I C ~ G A T ~ C C ~ ~ ~ T G ~ T R C ~ ~ C ~ C T C A G A T C A T G T A A A G G T T T A A ~ C U \ A C A G A C C ~ 10300
C A T T ~ A A G C T T ~ G T R C T T T A A T C C A A T C G ~ ~ ~ T T T T C T T C G A T T A ~ C T C T C T G ~ A T G C T G T T A T 10400
C C C T A C G G T A G C P T T T T C T T A A T C A T A A T T A T G G C T C 10500
A A A A A T G G A T A T A A A m T G T T T T T ~ ~ G A T m A T T T A T 10600
C A C T T ~ T T A A T T T T T A T T T A A T G T A A T A G A G A C A G C 10700
T T A G C A C A G T C A a ; G T A c T ~ ~ G T T T A A A A T T T C R C T ~ ~ A C G A C T ~ A T T A A T A A T T T T C A A A G ~ G A T G T T ~ T ~ A ~ ~ 10800
a ; C T A G A A T T I ~ G A G T T C C ~ T T A C A T T T A T A A T A T T T T T T T ~ T ~ ~ ~ A T T C T T ~ T T A T A R A ~ A T A m G A ~ T A A T R C T C A 10900
TTTTAACATARACTAATTTAAGPTTTTAAATTTTAAATAA~GATATT~~TTAGGTAT~TTTTTGAT~ATT~TTAAA~CTATTA 11000
ATATGATAGCIACTATTAA~RCATTAATTAAAGGTT~AATTTTCTTTTAAATTTTATT~TTAGCCTAATAAAAT~TAATTTAAATTA~TA 11100
TATTAAAAAATAATTTTTAGTAATTT~TTAGTAcTTATATACTTC~AACTAGCTATC~~CffiCAT~TTTAACTTCTATAAATA 11200
AATCATTAATTTAATTTn;CTACATTAATAAA~ATATATAGCTCCTTTT~TCT~TGATATTATAATAT~TAATCT~TTAAAC~TAATGC 11300
A A A R G G T A C T A G T G T P A A T C G T A C P T ~ T T T A T T C ~ T A C A G T R C T G T ~ T T T ~ T T ~ T T T T T T ~ A T A C T A A T T T A T A T A A 11400 . f" "1-rRNA-] f" tRNA val
G G G T T T T G A A T T A A A A A A T T T G T A A ~ T T A T T G G A A G G T 11500 [ """ #-rRNA-- -
TTTTAAGGAAGCTTC~TACTCCTA~ATGTTRC~CTTATCT~AAAAAG~AGTGAC~ATATGTA~CGTCTT~GCTTTTTCAAA 11 600
ATAATGTTCATAA~ATTTTTA~T~~TCC~TTGATT~TATAAT~C~T~ATTTCC~ATTTTAAATTTAAATTGTAACCCTAATATTAT 11700
~TTACTATAACTGGCGTGACCTGTCATAATA~~GTTGTCT~AGTAGTTAACTPATGRCG~~ATACAAACTAGT~TTTAAGTAAAG~ 11800
A G G T A A A G C G C G G G T T G T C G A G T T A A A A T A C A G G T T C C C C C 11900
T C T G G T ~AATGATTTAATTTTTTAATAGTG~TATCTAATCCCAGTTTTA~T~TTTT~T~CGTC~TTTAGTAAGAA~ffiT~T~ 12000
AATGTAAAmTACPTT~CATTTTTTATAATCTGACATC~AT~TATACCTTTTACCGTTTAGATTAT~AAC~GTATAACC~~ 12100
T P G C I ~ A C A A A T T T A A C T A G T T C T G A A A T C T A m C T A A 12200
AATTGATATCTTAAAAATAAATATTTTTTATAATTTATACAAAT~TTTATTTTTA~TRCAGCTTC~~~A~ATGTGTTTCT~G~ 12300 . f" --.-rRNA-] e tRNA mat
T T A A T T A A T T A T A A A C T A G A C A A A T T T A T T T G T A A A A G 12400 +tRNA tyr-
f" tRNA cy# GIAAACTCTAACAATCAGATGTAACTTTAAATTT~TTTAATATTTTTTAAACTAT~~TAATAG~GATATCCTCATAAATARATP~A~ 12500
f- tRNA trp
TTACCACCTTTTATCGGCCACT~ATTCAAAACAC~CATTTTAT~ATAGAAAGCTTT~~~~CTATTAGTT~TTAACTT~~T~T~GAA 12600
AAAGAGTTAC~TTTTTC~AATTCAAATTCCTAC~~~TAC~RCTTTTTATTATTT~TTTAAATGATTAAACTTTTTA~TT~~A 12700
AGTGTACTAATTATAC.:N\AGAAACATAGTTGAATAC~TTATCAGATAGATATTTATATTPATTCTTT~TAAAT~TTAGATGTG~TATTTAACRC 12800
f-" tRNA gln -tRNA glY-
. . " *tRNA glu
TACTTATCTGATAGCATTTATATATATATATATATATATATATATATATATATATATATATATATATATATATATATATATATATATATAT~TAU\ATA 12900 .CO3 -
T G A C C A T A T A T T T A T A T A T C T T A A A G ~ T A G ~ T G A T T C ~ T C C G ~ C A T T T A G T A G A A T T T ~ C C T ~ T T T A ~ 13000 M I R N P F H L V E F S P W P L V
G G G G T C T A TAGGCGCGTTTAA~~GCTTAGCAGCT~A~CAT~GTT~CTTTATTTTTAGTATG~T~GTTATTTTAATTTTA~A 13100 G S M G A F C L T V G L A A W F H G F S L F L V W V G V I L I L L
R C A A T G G T T C A A T G G T G G C G A G A T G T A A T T C G P G A A G G G A 13200 T M V Q W W R D V I R E G T F Q G Y H T L R V S R G L R W G M I L
TTATTACTTCTU\AGTACTATTTTTT~GCATTTTTTPG~CTTA~TCCATTCTAGATTGGCCCCTTGTT~AAATT~TCT~TGA~TCCACA 13300 F I T S E V L F F F A F F W A Y F H S S L A P C L E I G S C W P P Q
AGGAGTTGCTCCITTAAATCcTTTTCAAG~CTTTATTGAATRCAGC~GTTATTAGCGTC~TTAGTGTTACPTGAGCGCRCCATAGTTTAATP 13400 G V A P L N P F O V P L L N T A V L L A S G V S V T W A H H S L I

Chiton Mitochondrial Genome 443
G A T G G G G A T C ~ ~ G T ~ ~ A ~ ~ A C A G I T A T T T T A ~ ~ G T A ~ T T A C G I T ~ ~ A ~ ~ ~ T A T ~ A ~ ~ A ~ 13500 D G D Q G G A N I S L L T T V I L G A Y F T F L Q A G E Y L E T S
T T A C T A T T ~ C G A ~ ~ G T T A ~ A T C A A C A ~ ~ T T ~ A ~ A ~ G G G ~ C A T ~ T T ~ T G I G T T A G T A ~ ~ A ~ ~ A ~ ~ T T A ~ 13600 F T I A D S C Y G S T F F V A T G F H G F H V L V G S L F L L V T L
G T G A C G A A A T T T T A G T T G T C ~ ~ C ~ C C A ~ ~ ~ ~ ~ C ~ C T G C T T G G T ~ T ~ A T T T T ~ G A T G T A ~ ~ T T G T T T ~ A 13700 W R N F S C H F S S S H H F G F E A A A W Y W H F V D V V W L F L
tRNA lys TATAT~cTATTTACTGGTGGGGT~TAA~TATATC(A 13800
Y . 1 S I Y W W G S*** tRNA ala-”.)
- tRNA arg *- G G A A C T A T A C T T P A A G T A A A A G G T T T A A T T A A ~ ~ A ~ T ~ T A ~ ~ ~ T ~ ~ C C A G I ~ ~ G I G A T T A T T ~ T T T T T ~ T ~ ~ C 13900
tRNA am----) C A ~ A C G T A G C A G A G A A G P T C T G T m A A G A ( 3 T T C T G A A T A 14000
tRNA 110 + ND3 *
A R T I ” r A T A T A ~ ~ T G T T A C ~ C ~ T ~ ~ ~ T A C A T T G A ~ G T T G I A ~ T ~ T A A G ~ T ~ ~ C T T T ~ C A ~ ~ ~ T T T ~ T G I A ~ ~ T ~ G 14100 M F F V L S L
G I ~ G I T T A C T T T T T T A T T A A G A T T A T T 14200 V L F T F L L S L V L L S V S L S L T K K K M M N R E K S S P F E
G T G G G T T T G A T C C T A A A A G T C ~ C ~ ~ T ~ C ~ T T T ~ A ~ G G T ~ ~ T T A A ~ A ~ G T A G I T T T C T T A ~ ~ T ~ T G I n ; A ~ T ~ T T T T ~ T 14 300 C G F D P K S S A R L P F S M R F F L I T V V F L V F D V E I V L L
T T T G C C C T A T T T A T T T T C T A G T G G C n ; A A G A A T C G A T G T 14400 L P Y L F S S G W S I D V F S L V G S M M I L V I L I I G V L H E
tRNA ser(AGN) __)__ T G G I C n ; A A G G G A G G T T G T ~ T ~ T ~ A ~ ~ ~ T T A A G T T ~ T ~ T T T G G G A ~ T T T T G G C T G C ~ A A ~ ~ ~ T A A ~ G ~ T ~ ~ C 14 500 W S E G S L E W F S S S N * * *
ND2 j . C G T T T P f C T T T T T G P G T T T T A A A T T T T C C ~ T G T T ~ ~ T A T T T A T T T T T A T ~ T A T T ~ T ~ ~ A ~ ~ A T ~ T ~ T T A T ~ T ~ A T T C A T ~ A ~ C ~ G 14600
V L N F P F V G L F I F I L F F G T L F S L S S I H W F G
G P I T G G C T n ; G ~ T A G A A T T A A A T T T A A T A T C C 14700 V W L G L E L N L M G F I P V M V Q K S T S E E T E S G V K Y F L
T T C A A G C T G T T G G C T C A G C T T T P r r G T T T T T G P T T G G A T T G A 14800 V Q A V G S A L F L F G L M L M N W N F C C W E L N F F S G F S X S
A G G T T T A A T T T T T T T T G G T T ~ ~ T ~ ~ T ~ ~ ~ C A ~ A ~ ~ ~ ~ ~ A G G G T T G T A ~ ~ T A T ~ ~ A A T A ~ A A T ~ T 14 900 G L I . F F G . L L M F L G A . A P F . H F W Y P S V . V A G . L S W M S N F .
T T ~ T A T T A A C A G ~ C A A A A A A T T G C T C C A T T G m A T G G T 15000 L L L T V Q K I A P L F M V C W Y L N L S S F L L L I L V F L S S
T A T P T G G A G G A G I C G G A G G G G T T A A ~ A ~ C ~ C ~ T A C G C 15100 L F G G V G G V N Q T S V R A L I A Y S S I L H M G W M L X G A S A
G G G A T G G A G C A G T A T T ~ T T T T T A C ~ T T T T T T T A C T 15200 G W S S I F F Y F F F Y C F I L G F I A Y L M G L D E S F N M S C
T T T A G A A G A G T G I A T G n ; T G A A A T T ~ T A T T C G C G P A A A 15300 F S S V Y V W N S Y S R N F L V F M L L S L G G L P P L L G F F G
A G ~ A G T G T P A G T T A G A T T ~ A T C T T T ~ G ~ T T T A G T T T T A A ~ T T A T T T T A G T T ~ T ~ ~ T A T ~ T T ~ A T T A T A T T A T T A T T T ~ T ~ T 15400 K W L V L V S L L S L G N L V L S I I L V C G S M I S L Y Y Y L V L
T A G T T T T T C A ~ A ~ T T T A A G C A A A G G G A G G T G A A G A 15500 S F S L L L S K G S W S S G A L V S K N L M W F G G A M N L S G L
GGTGTITTATTGTGGTIAAGTACll’TlTAGAT 15532 G V L L W L S T F * * *
FIGURE 7.-Continued.