Embed Size (px)
Citation preview

doi:10.1016/j.jmb.2009.08.048 J. Mol. Biol. (2009) 393, 898–908
Available online at www.sciencedirect.com
Crystal Structure of the Antitoxin–Toxin ProteinComplex RelB–RelE from Methanococcus jannaschii
Djordje Francuski and Wolfram Saenger⁎
Kristallographie, Institut fürChemie und Biochemie, FreieUniversität Berlin, Takustrasse6, 14195 Berlin, Germany
Received 22 March 2009;received in revised form4 August 2009;accepted 18 August 2009Available online25 August 2009
*Corresponding author. E-mail [email protected] used: TA, toxin–an
RelE from Methanococcus jannaschii;Pyrococcus horikoshii; DLS, dynamicnoncrystallographic symmetry; MPDpentanediol.
0022-2836/$ - see front matter © 2009 E
Here we present the crystal structure of the Methanococcus jannaschiiRelE–RelB (RelBE) toxin–antitoxin (TA) protein complex determined bythe MIRAS (multiple isomorphous replacement with anomalous signal)method. The genes encoding this TA system are located in the chromosomeof this archaeon and involved in stress response. RelE acts as an endo-ribonuclease that cleaves mRNA on the ribosome, and we compare theRelBE complex to the known structures of other TA systems belonging tothis group and to endoribonucleases. M. jannaschii RelBE forms a hetero-tetramer with the antitoxin in the centre of the complex, a configuration thatdiffers vastly from the heterotetramer structure of the previously publishedRelBE from another archaeon, Pyrococcus horikoshii. The long N-terminal α-helix of the tightly bound M. jannaschii antitoxin RelB covers the presumedactive site of the toxin RelE that is formed by a central β-sheet, a loop on oneside and a C-terminal α-helix on the other side. The active site of the M.jannaschii toxin RelE harbours positive charges that are thought to neutralizethe negative charges of the substrate mRNA, including Arg62 that waschanged to Ser62 by the Escherichia coli expression system, thereby leading toinactive toxin RelE. Comparative studies suggest that Asp43 and His79 arealso involved in the activity of the toxin.
© 2009 Elsevier Ltd. All rights reserved.
Edited by J. Doudna
Keywords: RelBE; RelE; antitoxin–toxin; addiction module; endoribonucleaseIntroduction
For a series of bacterial toxin–antitoxin (TA)systems, the genes encoding the TA proteins wereinitially identified on plasmids due to the ability oftheir gene products to prevent the formation ofplasmid-free progeny.1,2 If a cell loses the plasmidcarrying genes encoding a TA system, the antitoxin issusceptible to rapid degradation by endogenousproteases, but the free toxin persists and kills the cell,so that only those cells that harbour the plasmidsurvive. When genes of homologous TA systemswere later discovered in chromosomes of many bac-teria and archaeons3,4 that have a so called “delayedrelaxed” phenotype, it was evident that these TAs
ress:
titoxin; MjRelE, toxinPhRelE, RelE fromlight scattering; NCS,, 2-methyl-2,4-
lsevier Ltd. All rights reserve
may act as cellular regulators that help the cells toadapt their gene expression to environmental stress.5
The currently known toxins can be subdivided intofive superfamilies called CcdB, RelE, Doc, HipA andζ. A proposed sixth superfamily represented by toxinVapC that has not yet been studied thoroughly6
consists of anN-terminal domain (PilT-N or PIN) thatis also found in proteins of the eukaryotic nonsense-mediated RNA decay system.6,7 An overview of theknown TA systems is provided in Table 1.
The RelE toxin superfamily
The toxin RelE from Methanococcus jannaschii (de-noted as MjRelE) described here belongs to a largersuperfamily (Table 1) having a polypeptide foldsimilar to the fold of RelE from Pyrococcus horikoshii11
(denoted as PhRelE) and YoeB from Escherichia coli12
that act as endoribonucleases and cleave mRNA.Structurally related proteins that are not part of TAsystems but share the endoribonuclease fold arecolicins D and E5 (both tRNases21,22), RNase Sa fromStreptomyces aureofaciens (and members of its family)and RNase RegB that is involved in the control of themultiplication cycle23 of bacteriophage T4.
d.

Table 1. Overview of the known TA systems
Toxin/antitoxin
Toxin superfamily Antitoxin superfamily
Toxin targetToxin (T)–antitoxin (A)
oligomer state Spread and gene locationMazF RelE Doc PIN HipA ζ AbrB MetJ/Arc YefM/Phd HTH motif
CcdB/CcdA + + Replication, gyrase T2A2T28 Plasmid of Gram-negative bacteriaKid/Kis + + Translation, mRNA TA2T concentration
dependent9Plasmid of Gram-negative and
Gram-positive bacteriaMazF/MazE + + Translation, mRNA T2A2T210 Chromosome of Gram-negative and
Gram-positive bacteriaChpBK/ChpBI + + Translation, mRNA Chromosome of Gram-negative bacteria
RelE/RelB + + Translation, mRNA TAAT(this work and11)
Chromosome of Gram-negative andGram-positive bacteria, Archaea
YoeB/YefM + + Translation, mRNA TA2T12 Chromosome of Gram-negative bacteriaYafQ/DinJ + + Translation, mRNA13 Chromosome of Gram-negative bacteriaHigB/HigA + + Plasmid of Gram-negative and
Gram-positive bacteriaParE/ParD + + Replication, gyrase Plasmid of Gram-negative and
Gram-positive bacteriaYhaV/PrlF + + (TAT)2 14 Chromosome of Gram-negative bacteriaTxe/Axe + + Plasmid of Gram-negative and
Gram-positive bacteria
Doc/Phd + + Translation, ribosome15 TA216 Chromosome and Plasmids of Gram-negativeand Gram-positive bacteria, Archaea
VapC/VapB + + TA17 Chromosome of Gram-negative andGram-positive bacteria, Archaea
HipA/HipB + + TAAT18 Chromosome of Gram-negative bacteria
ζ/ɛ (ω)a + + TA2T19 Chromosome and Plasmids ofGram-positive bacteria
In the column “Toxin(T)–antitoxin (A) oligomer state” the experimentally verified compositions are given (structural data are indicated in bold face). The “Spread and gene location” column shows thephylogenetic distribution of the gene families. (Table based on data from Refs. 5, 6, and 20). Boldface indicates the family name of the TA systems.
a The promoter regulation is achieved via a third protein; see the text for details.
899CrystalS
trutureof
Methanococcus
jannaschiiRelB
E

900 Crystal Struture of Methanococcus jannaschii RelBE
The toxins of the RelE superfamily are charac-terized by a five-stranded β-sheet with four of thestrands antiparallel to each other, while the first andlast strands are parallel. The β-sheet is flanked onone side by two α-helices α1 and α2 and on the otherside by α-helix α3 that, together with the loop con-necting strands β2 and β3, forms a deep cleft thatencompasses the RNase active site located on thefive-stranded β-sheet as described below in moredetail. A comparable architecture is also found ingenuine RNases that have very low sequence homo-logy to RelE.The relBE promoter is strongly activated during
nutritional stress, and protease Lon is required24 fordegradation of antitoxin RelB. There is competitionbetween the translation release factor RF1 and RelEfor ribosome binding, and it was suggested thatRelE enters the ribosomal A-site to cleave mRNA25
at the second position of the A-site codon, preferen-tially at stop codons (UAG, UAA, UGA) but at sensecodons (UCG, CAG) as well,25,26 leading to globalinhibition of protein synthesis and eventually to cellgrowth arrest. This toxic effect may be reversed byexpression of tmRNA (which is also cleaved byRelE)26 that recycles ribosomes by entering the A-site of ribosomes stalled on truncated mRNAs.
Fig. 1. Arrangement of toxin and antitoxin chains in the heCartoon representation of the heterotetramer formed by M. jantwo views, on (top) and along (bottom) the NCS axis relating thform the U-shaped base on which the β-sheet is placed. The lonantiparallel β-sheet, the loop β2–β3, and C-terminal helix α3antitoxin being wrapped around the toxin. Strand β2 of MjMjRelE, and strand β1 ofMjRelB forms an intermolecular β-shheterodimer interaction is primarily stabilized through MjRelB(cyan) wraps around the toxin PhRelE (red). The PhRelBEheterotetramer. [All figures were prepared with PyMOL (DeL
It was suggested11 that the toxic action of RelEis counteracted by antitoxin RelB, which wrapsaround RelE, blocks its active site and preventssterically the binding to the ribosomal A-site. Thetranscription of the relBE operon is autoregulatedby RelB that acts as transcriptional repressor, andRelE enhances the binding of RelB to thepromoter.27,28
A number of practical applications have been pro-posed4,29 for TA systems in general or are already inuse30 (such as CcdB in the Gateway™ cloning fromInvitrogen). Some other applications have been pro-posed specifically for the RelBE TA system.31The work presented here describes the crystal
structure of the nontoxic TA complex MjRelBER62S
from M. jannaschii and compares it to the knownstructures of this toxin class.
Results
Circular dichroism and dynamic light-scatteringmeasurements
To determine the α/β contents and to assess thethermal stability of the purified RelBE complex, cir-
terotetramer formed by MjRelBE (a) and PhRelBE (b). (a)naschii toxin MjRelE (red) and antitoxin MjRelB (blue), ine twoMjRelBE heterodimers. Helices α1 and α2 ofMjRelEg helix α1 of the antitoxinMjRelB is tightly gripped by theof MjRelE (both indicated by arrows), with the rest of theRelB forms an intermolecular β-sheet with strand β1 ofeet with β1′ of the NCS-relatedMjRelB. The heterodimer––MjRelB interaction. (b) The P. horikoshii antitoxin PhRelBheterotetramer form is very different from the MjRelBEano Scientific, 2006)].

Fig. 2. Topographical representation showing the fold-ing ofMjRelE (red) andMjRelB (blue). Residue numbers ofN- and C-termini of α-helices and β-strands are labelled.Strand β2 of antitoxinMjRelB forms an antiparallel β-sheetwith strand β1 of toxinMjRelE, and the helix α1 ofMjRelBis positioned above the five-stranded central β-sheet ofMjRelE and gripped from both sides by helix α3 and theβ2–β3 loop ofMjRelE. Two of these heterodimers form theheterotetramer (MjRelBE)2 by the interaction of theantitoxins MjRelB and MjRelB′ that form an antiparallelβ-sheet with their β1 strands, and by contacts between α3ofMjRelEI and α1 ofMjRelBI. Some functionally importantresidues are marked (graphic prepared with TopDraw32).
901Crystal Struture of Methanococcus jannaschii RelBE
cular dichroism (CD) spectra were recorded at diffe-rent temperatures. As expected for proteins from anarchaeon, MjRelBE proved to be very heat stable upto the limits of the instrumental capabilities (88 °C).Since MjRelBE tends to form higher oligomeri-
zation states at ionic strengths below 400 mM NaCl,as indicated by dynamic light scattering (DLS; notshown), the crystallization trials were performed inbuffer C containing 500 mM NaCl.
Description of the structure
MjRelBE forms a heterotetramer consisting of twoheterodimers that are related by a noncrystallo-graphic symmetry (NCS) axis (Fig. 1a). HeterodimerI (MjRelBEI) consists of MjRelBI, MjRelEI, andheterodimer II (MjRelBEII) of MjRelBII, MjRelEII. Inthe heterotetramer, the N-terminal β-strands β1 andα-helices α1 of the antitoxins MjRelBI and MjRelBIIare connected by hydrogen bonds and van derWaals interactions (Fig. 1a.), and helix α1 of RelBis tightly bound to a cleft (see below) of RelE. Theremaining two loops, strand β2 and helix α2 of theantitoxinsMjRelBI,II are tightly wrapped around thetoxins MjRelEI and MjRelEII in each of the MjRelBEheterodimers.N-terminal sequencing and MALDI-TOF (matrix-
assisted laser desorption/ionization time-of-flight)spectra showed that full-length antitoxin MjRelBwas present before and after crystallization, whilethe first two residues (Met–Lys) of the toxin MjRelEwere missing probably due to protease clipping. Theelectron density was not interpretable for residues1–7 and 49–52 of MjRelBI and for residues 1–8 and48–52 of MjRelBII, while the electron density fortoxin MjRelEI was not interpretable for Asp39.The two MjRelBE heterodimers forming the
heterotetramer are related by a noncrystallographic2-fold symmetry axis (Fig. 1a) and, consequently,not entirely identical (Supplemental Fig. 1). The Cα
traces are very similar in the core parts of theproteins, while in the loops and terminal regions thetemperature factors and conformations are gov-erned by crystal contacts. In both toxin moleculesMjRelEI and MjRelEII, the flexible loops (residues35–43) and the C-termini (residues 80–88), showlarger discrepancy in position and B-factors thatdiffer up to 10 Å2, with molecule I being better de-fined in the loop regions and molecule II at theC-terminus.Since the topographies of the heterodimers
MjRelBEI andMjRelBEII are similar, only the formeris described here. The core of the toxin MjRelE isformed by an antiparallel five-stranded β-sheet ex-cept for strand β1 that is parallel to the adjoiningstrand β5. The β-sheet is flanked by three helices(Figs. 1a and 2), helices α1 and α2 being on one sideforming a U-shaped base, while the C-terminal helixα3 is on the other side of the β-sheet. Helix α3 andthe loop connecting strands β2 and β3 form a deepcleft accommodating the active site (see below) thatis blocked by the long helix α1 of antitoxin MjRelB(Figs. 3 and 4).
The antitoxin MjRelB is wrapped around toxinMjRelE, with strand β2 forming an antiparallel sheetwith β1 of the toxin MjRelE, while strands β1 andβ1′ (prime indicating the NCS-related partner) ofthe two antitoxins form another short antiparallelβ-sheet that contributes to the stabilization of theheterotetramer (Fig. 1).
MjRelE–MjRelB complex formation
Besides the large contact surface between the cleftformed by MjRelE and the long helix α1 of MjRelB,further interactions involve the long loop (residues34–39) of MjRelB, its short strand β2 and the shortC-terminal helix α2. In the heterodimer, the inter-actions between the toxin and antitoxin bury 2740Å2 of the solvent-accessible area. The contributionof the buried surface to the dissociation free energyis around 17 kcal/mol (the exact value variesslightly between the two heterodimers due to smallstructural differences), as calculated by the PISAservice.33
The cleft formed by MjRelE (Figs. 3 and 4) ispositively charged with Lys7, Lys45, Lys48, Arg55,Arg57, Arg62 (this position is replaced by Ser62 inthe inactive variant MjRelBER62S described here),Arg75, Arg76 and Arg80. Additionally, there arepolar residues Tyr50, Tyr53 and Tyr86 and nonpolar

Fig. 3. Superposition of MjRelBE (red, blue) andPhRelBE (grey, cyan) heterodimers. β0 of PhRelB and β2of MjRelB as well as α4 of PhRelB and α2 of MjRelBoverlap fairly well, while the rest of the antitoxin chainsdiffer radically in wrapping around the cognate toxins.The loop β2–β3 and the C-terminal helix α3 ofMjRelE thatembrace helix α1 of MjRelB are both indicated by thickarrows.
902 Crystal Struture of Methanococcus jannaschii RelBE
residues Phe87 and Pro88. The interactions withMjRelB are mainly electrostatic and involve, onMjRelBI, Glu21, Asp25 and Lys30, and on MjRelBII,Glu21, Glu22, Asp25 and Glu34 (Supplementary
Fig. 4. Surface charge representation with negative (red) an(white). (a)MjRelE with α1 ofMjRelB (yellow stick representatThe arginines on the far ends of the cleft, Arg55, Arg57, Arg62Arg76 at the bottom render the cleft positively charged. Theform both sides of the cleft are labelled. (b) Surface electrostatiyellow stick representation in (a). Secondary-structure elemesentation of interacting parts of MjRelE (grey, with functional
Table 1). The loop between helix α1 and strand β2 ofMjRelB interacts with MjRelE through hydrogenbonds between MjRelB Asp38OD and Glu39O andMjRelE Lys7N, Thr8N and Thr8Oγ. Additionalhydrogen bonds are found in the β-sheet formedby strands β2 ofMjRelB (Glu39O, Leu41N, Leu41O)and β1 of MjRelE (Phe5O, Phe5N, Lys7N).
Heterotetramer formation
Analytical ultracentrifugation (see SupplementaryFig. 2) confirmed an equilibrium between hetero-tetramers and heterodimers in a wide variety (0.5 to2.75 g/L) of protein concentrations and showed thiscomplex to be not very stable, with a dissociationconstant of 75.6±13 μM. The mass of the hetero-dimer and heterotetramer would only be obtainableafter extrapolation to very low and very high con-centrations, respectively, of MjRelBE.In the heterotetramer, the antitoxins MjRelBI and
MjRelBII form an antiparallel β-sheet with strandsβ1 and β1′. Further interactions involve hydropho-bic contacts between helices α1 and α1′ throughresidues Ile20, Ile23 and Ile20′, Ile23′ and interac-tions between residues Phe10, Tyr17 and Phe10′,Tyr17′.Interactions between the heterodimers MjRelBEI
and MjRelBEII include salt bridges and hydrogenbonds between Arg80, Lys81 and Tyr84 of theC-terminal helix α3 of MjRelEI and Glu16, Lys19of helix α1′ of MjRelBII (Supplementary Table 1).This interaction is not symmetric in the hetero-tetramer because the interaction between MjRelEI
d positive (blue) charges indicated, and neutral segmentsion) located in the deep positively charged cleft ofMjRelE.(replaced here by Ser62), Arg80 at the top and Lys7, Arg75,tyrosines (Tyr50, Tyr53 and Tyr86), Pro88 and Phe87 thatcs of the antitoxin MjRelB rotated 180° with respect to thents of MjRelB are marked, and cartoon and stick repre-ly important residues marked).

903Crystal Struture of Methanococcus jannaschii RelBE
Arg80, Tyr84 and MjRelBII Glu16 is direct, whilethis interaction between MjRelEII and MjRelBIis bridged by a water molecule positioned in themiddle between MjRelEII Arg80, Tyr84 and MjRelBIGlu16.
Comparison of the MjRelBE and PhRelBEstructures
We expected to find a heterotetramer of the kindas reported for the PhRelBE homolog, which, how-ever, proved to be wrong as shown by the models(Fig. 1) that differ grossly. The antitoxin PhRelB ofP. horikoshii is longer (67 residues) than MjRelB (52residues), and its helix α2 is positioned in the cleft ofthe toxin PhRelE (Figs. 1b and 3). This is in constrastto MjRelB that is bound with helix α1 to MjRelE.Starting from the C-termini of the antitoxins, theyshare similar features (Fig. 1a and b). They both havea short C-terminal helix followed by a short β-strand(β2 inMjRelB and β0 in PhRelB) that is participatingin β-sheet formation with the toxin strand β1(Fig. 3). From there toward the N-termini, the Cα
traces of the antitoxins take different paths. PhRelBforms a short helix α3 (not present in MjRelB) andcontinues to a longer helix α2 positioned within thecleft formed by toxin PhRelE that is wider than thecleft in MjRelE (Fig. 3).Toward the N-termini, the polypeptide chains of
the antitoxins take opposite directions; the MjRelBhelix α1 is much longer than the cleft of MjRelE andends with N-terminal strand β1, whereas helix α2 ofPhRelB ends within the cleft and is connected by aloop with the long N-terminal helix α1.The folds of the RelE toxins are fairly well con-
served with root mean square deviation (r.m.s.d) of1.3 Å (see Discussion), the main differences occur-ring in the position of loop β2–β3 and helix α3 thatform the cleft of MjRelE, while in PhRelE the cleft isshallow and formed only by the bent β-sheet (Fig. 3).In the MjRelBE heterotetramer, the contact betweenthe two heterodimers is formed between the anti-toxins and there is no MjRelEI–MjRelEII interaction,an arrangement that contrasts PhRelBE where thetwo PhRelE toxins share a common interaction sur-face (Fig. 1a and b, bottom panels).
Discussion
MjRelE from M. jannaschii is the second structur-ally characterized member of the RelE toxin familyand the third member of the RelE superfamily(PhRelE and YoeB, respectively). The structure ofMjRelBE differs from that of P. horikoshii PhRelBE inthe arrangement of the heterodimer, the depth of thecleft binding the antitoxin and the presence orposition of some crucial residues (His79 and Tyr84,respectively) that are similar in MjRelE and YoeBbut not in PhRelE. Structure and amino acidsequence superpositions of the RelE toxins fromM. jannaschii, P. horikoshii, E. coli, and toxin YoeBshow that there are several highly conserved resi-
dues, Arg57, Arg62 (essential for toxic function),Tyr84 and Asp43 (Glu46 in YoeB) (RelE numbering,Fig. 5a and c). Other residues are type conserved.These toxins share low sequence identity but a
common fold. The r.m.s.d. of the superimposedmain-chain atom positions obtained by the TOPprogram36 is 1.3 Å for MjRelE–PhRelE (26.7% se-quence identity for 60 overlapping residues), 1.8 Åfor MjRelE–YoeB (13.4% sequence identity for 67overlapping residues) and 1.5 Å for PhRelE–YoeB(15.2% sequence identity for 66 overlapping resi-dues). The r.m.s.d. for the superposition of the toxinswith fold-related ribonucleases is 1.8 Å for MjRelE–RegB (34 residues overlapping), 1.9 Å for MjRelE–RNase Sa (33 residues overlapping) and 1.5 Å forMjRelE–colicin E5 (31 residues overlapping). Muta-tional studies11,12 confirmed that the positivelycharged cleft (Fig. 4) of the toxin is essential for thetoxic effects of RelE and its homologs.One crucial residue is Arg62 in MjRelE of
M. jannaschii and Arg56 of E. coli EcRelE (Table 2),which corresponds to Arg65 of YoeB and Arg65 ofPhRelE that probably serves as phosphate-bindingresidue and would stabilize the negative charge ofthe 2′,3′-cyclic phosphate intermediate as in colicinE5.22 The same arginine is also conserved amongtoxins Txe, YhaV and HigB. In both M. jannaschiiMjRelE and YoeB, mutation of this Arg renders thetoxin inactive (this work and that of Kamada andHanaoka12).During our work we noticed the spontaneous
mutation in MjRelE of Arg62 to Ser62 or Lys62 thatinactivated the toxin, and hence it was possible toclone MjRelER62S alone into another vector and toexpress it without its cognate antitoxin MjRelB. Alltrials to revert this mutation and to clone wild-typeMjRelE even together with the antitoxin MjRelBfailed, and the transformation of cells was neversuccessful. We have learned that Gerdes' group inOdense, Denmark (S. Christensen, personal com-munication) encountered the same problem. This isa puzzling effect because we had no problems incloning wild-type EcRelBE of E. coli in expressionstrains. The toxin MjRelE is evidently more toxic toE. coli cells because even basal expression of theprotein caused cell stasis or death.The mechanism of RNA cleavage among endo-
ribonucleases includes an acid–base catalytic stepwhere the general acid–base pair consists of Gluand His, but it is possible that an Arg–Asp pairmay take this catalytic role.37 In the tRNase colicinE5, the proposed mechanism for RNA cleavageinvolves Asp46 and Arg4822 and does not requireany His.Comparison of the structures of YoeB and MjRelE
(Fig. 5a) shows that the residues denoted to be cru-cial or important for the function as an endoribonu-clease are conserved between YoeB and MjRelE(Table 2). His83 and Arg65 of YoeB are likely to beinvolved in phosphate binding12 and may serve asgeneral acids in the catalytic reaction. The mostprominent differences among the conserved resi-dues are the orientations of MjRelE His79 and YoeB
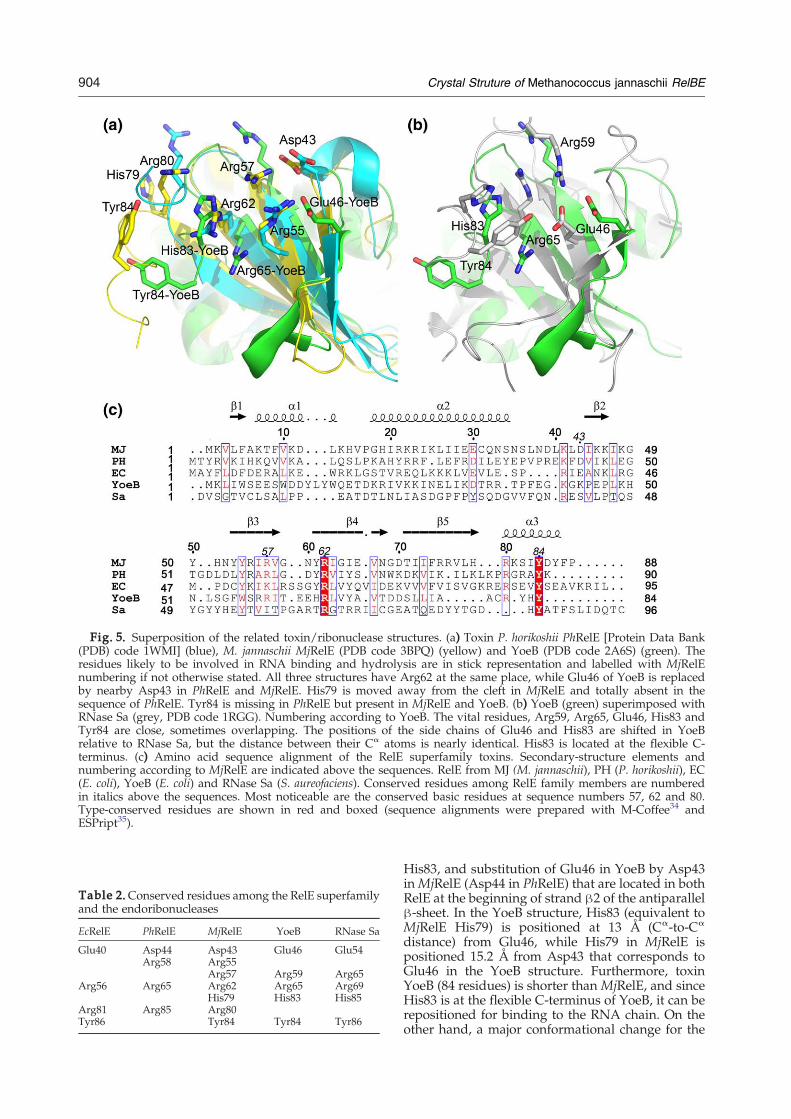
Fig. 5. Superposition of the related toxin/ribonuclease structures. (a) Toxin P. horikoshii PhRelE [Protein Data Bank(PDB) code 1WMI] (blue), M. jannaschii MjRelE (PDB code 3BPQ) (yellow) and YoeB (PDB code 2A6S) (green). Theresidues likely to be involved in RNA binding and hydrolysis are in stick representation and labelled with MjRelEnumbering if not otherwise stated. All three structures have Arg62 at the same place, while Glu46 of YoeB is replacedby nearby Asp43 in PhRelE and MjRelE. His79 is moved away from the cleft in MjRelE and totally absent in thesequence of PhRelE. Tyr84 is missing in PhRelE but present in MjRelE and YoeB. (b) YoeB (green) superimposed withRNase Sa (grey, PDB code 1RGG). Numbering according to YoeB. The vital residues, Arg59, Arg65, Glu46, His83 andTyr84 are close, sometimes overlapping. The positions of the side chains of Glu46 and His83 are shifted in YoeBrelative to RNase Sa, but the distance between their Cα atoms is nearly identical. His83 is located at the flexible C-terminus. (c) Amino acid sequence alignment of the RelE superfamily toxins. Secondary-structure elements andnumbering according to MjRelE are indicated above the sequences. RelE from MJ (M. jannaschii), PH (P. horikoshii), EC(E. coli), YoeB (E. coli) and RNase Sa (S. aureofaciens). Conserved residues among RelE family members are numberedin italics above the sequences. Most noticeable are the conserved basic residues at sequence numbers 57, 62 and 80.Type-conserved residues are shown in red and boxed (sequence alignments were prepared with M-Coffee34 andESPript35).
Table 2.Conserved residues among the RelE superfamilyand the endoribonucleases
EcRelE PhRelE MjRelE YoeB RNase Sa
Glu40 Asp44 Asp43 Glu46 Glu54Arg58 Arg55
Arg57 Arg59 Arg65Arg56 Arg65 Arg62 Arg65 Arg69
His79 His83 His85Arg81 Arg85 Arg80Tyr86 Tyr84 Tyr84 Tyr86
904 Crystal Struture of Methanococcus jannaschii RelBE
His83, and substitution of Glu46 in YoeB by Asp43inMjRelE (Asp44 in PhRelE) that are located in bothRelE at the beginning of strand β2 of the antiparallelβ-sheet. In the YoeB structure, His83 (equivalent toMjRelE His79) is positioned at 13 Å (Cα-to-Cα
distance) from Glu46, while His79 in MjRelE ispositioned 15.2 Å from Asp43 that corresponds toGlu46 in the YoeB structure. Furthermore, toxinYoeB (84 residues) is shorter than MjRelE, and sinceHis83 is at the flexible C-terminus of YoeB, it can berepositioned for binding to the RNA chain. On theother hand, a major conformational change for the

905Crystal Struture of Methanococcus jannaschii RelBE
equivalent histidine (His79) in MjRelE would berequired to occupy the same position as that ofHis83 in YoeB. The same applies for Tyr84 ofMjRelEthat corresponds to Tyr84 of YoeB.It is noticeable that His83 present in YoeB and the
equivalent His79 in MjRelE is missing (Fig. 5 andTable 2) in the sequences of E. coli and P. horikoshiiRelE. Since His83 is essential for the toxicity ofYoeB,12 this suggests that the E. coli and P. horikoshiiRelE are missing a residue (that is possibly sup-plemented by the ribosome) to form an activeribonuclease.As noted in Table 2, Arg62 inMjRelE and Arg65 in
PhRelE are at the same location and essential for theactivity of these toxins. In YoeB, Glu46 and Arg65are required for toxicity.12 Glu46 is near the positionwhere Asp43 is located in the structure of MjRelE,and Arg65 in YoeB is placed in exactly the sameposition as the essential Arg62 in MjRelE and Arg65in PhRelE (Fig. 5a and Table 2), suggesting thatresidues Asp43/Arg62 inMjRelE and Asp44/Arg65in PhRelE could be the most probable catalyticresidues for RNA cleavage. YoeB, a purine-specificribonuclease,12 has a “classical” catalytical site, asshown in the comparison with RNase Sa (Fig. 5b).YoeB residues Arg59 and Arg65 overlap with Arg65and Arg69 from RNase Sa, while His83 and Glu46 ofYoeB are at similar distances compared to thepositions of the equivalent His85 and Glu54 inRNase Sa. The distances between these residues are10.4 Å in YoeB and 9.9 Å (measured from Cγ to Cγ)in RNase Sa. His83 and Tyr84 of YoeB are placed atthe C-terminus of the polypeptide chain, and theirpositions vary in the antitoxin bound and freeforms,12 indicating that their distance to Glu46most probably changes upon RNA binding.12
In colicin E5 and RegB, the active site is located atdifferent places on the β-sheets of the toxins andsurrounding helices (Supplementary Fig. 3). Both ofthese proteins have endoribonuclease activity sim-ilar to that of YoeB. Therefore, the positions of thecatalytically active residues are not restricted to oneplace on the surface of the cleft but can be placed atanother part of the β-sheet, depending on thespecificity of the endoribonuclease. The substratesfor colicin E5 are tRNAs that are preferentiallycleaved at anticodons coding for tyrosine, histidine,glutamic acid and aspartic acid, while RegB cleavesin the Shine–Dalgarno sequence GGAG. Bothenzymes have, besides the preferential target se-quence, the requirement for a particular secondarystructure of tRNA in the vicinity of the cleavagesite,22,38 which has not yet been established for YoeBor RelE.The shape of the cleft harbouring the active site,
the arrangement of secondary-structure elementsand the positioning of positively charged residuesare the essential ingredients for RNA binding. Thecommon fold between the TA systems and otherendoribonucleases is associated with the substrate(RNA) and, therefore, widely spread. The position-ing of the residues responsible for RNA cleavagediffers regardless of the common fold.
Materials and Methods
Cloning, overexpression and purification of RelBE
The group of Kenn Gerdes, Odense, Denmark kindlysupplied the plasmid pSC2526MJ#2 containing the M.jannaschii (strain DSM2661) chromosomally encoded relBEgenes. E. coli strains DH5a and TOP10 were used for thesubsequent subcloning.We constructed plasmid pET21relBE_MJ by insertion of
the relBE genes, taken from pSC2526MJ#2, in pET21b, andexpressed the encoded proteins in E. coli Rosetta strainfrom Novagen (that carries the pRARE helper plasmid).For mutagenesis, we used the TOPO Cloning Kit fromInvitrogen and a variation of the megaprimer protocol.An overnight culture of the E. coli Rosetta strain trans-
formed with pET21relBE_MJ was grown in LB mediumwith ampicillin (100 μg/ml) and chloramphenicol (20 μg/ml) at 37 °C. The growth culture was obtained by 1:100dilution of the overnight culture. Cells were grown in LBmedium with ampicillin (100 μg/ml) and chloram-phenicol (20 μg/ml) at 37 °C till they reached an OD600of 0.6, and 1 mM IPTG was then added to the culture forinduction. The growth was continued for 3.5 h foroverexpression, after which the culture was centrifugedat 6000g; the obtained pellet was resuspended in buffer A[50 mM NaCl, 50 mM Tris (pH 8.0)] and frozen at −20 °Cor used right away. The same procedure was followedwith the inactive Arg62Ser mutant of theM. jannaschii relEgene without gene relB.Following cell lysis and centrifugation, theM. jannaschii
RelBE protein complex was purified on a heparin columnwith elution buffer B [2 M NaCl, 50 mM Tris (pH 8.0)],yielding highly purified MjRelBE. After gel-filtrationchromatography in buffer C [500 mM NaCl, 20 mM 4-(2-hydroxyethyl)-1-piperazine-ethanesulfonic acid (Hepes)(pH 7.5)], N95% pure RelBE was obtained, concentratedto 10 mg/ml and used for crystallization.
Isoelectric focusing, CD, DLS and absorbancemeasurements
Isoelectric focusing experiments were performed withprecast gels (GE Healthcare) in a PhastGel system fromPharmacia (GE Healthcare). CD spectra of M. jannaschiiMjRelBE were measured in 500 mM NaCl, 20 mM phos-phate buffer (pH 7.5) at a protein concentration of 1.4 mg/ml with a JASCO-J600 CD spectrometer. The secondary-structure content was calculated with the K2D program.39DLS measurements were performed with a Laser Spec-
troscatter 201 from RiNA, Berlin, in 20 mM Tris (pH 8.0)and protein concentrations from 2 to 10mg/ml at differentNaCl concentrations in the range of 50 mM to 1 M NaCl.Protein concentrations were measured by UV absor-
bance of the pure protein solution at 280 nm in the abovebuffers. The extinction coefficient used, derived fromthe amino acid composition, is 10,240 (cm M)−1 for M.jannaschii MjRelBE.
Crystallization and heavy-atom derivatisation ofM. jannaschii MjRelBE
Initial crystalline clusters were found in a sitting-dropexperiment with Hampton Crystal Screen HT in 20%ethanol and 0.1 M Tris (pH 8.0). Starting crystals forseeding were grown by the hanging-drop method by the

Table 3. Data collection and refinement statistics
Native Ethylmercury phosphate Dichlorodiammine-platinum
Data collectionSpace group P21 P21 P21Cell parameters
a, b, c (Å) 52.79, 57.99, 58.75 52.04, 58.37, 59.75 52.69, 57.92, 57.66α, β, γ (°) 90.00, 92.30, 90.00 90.00, 91.72, 90.00 90.00, 92.74, 90.00
Wavelength (Å) 0.9490 1.0074 1.0722Resolution limits (Å) 50.0–2.13 (2.13–2.18) 50.0−2.6 (2.60−2.70) 50.0−2.7 (2.70−2.80)Rsym
a 0.050 (0.42) 0.050 (0.34) 0.064 (0.35)I/σ(I) 27.03 (3.13) 28 (6.50) 18.88 (4.14)Completeness (%) 99.6 (98.9) 99.9 (100.0) 99.7 (99.6)Redundancy 4.2 7.6 5.7
RefinementResolution (Å) 20.0−2.2Unique reflections 17,159Rwork (%)b/Rfree (%)c 23.8/27.4No. of atoms
Protein 2191Water 56
B-factors (average) (Å2) 50.04r.m.s.d.'s
Bond lengths (Å) 0.008Bond angles (°) 1.058
Geometry (%)Most favored 93.8Additionally allowed 5.7Generously allowed 0.4
a Rsym=∑hkl∑j|Ij− ⟨I⟩|/∑hkl∑jIj, where ⟨I⟩ is the mean intensity of reflection hkl.b R‐factor=∑hkl||Fobs|−|Fcalc||/∑hkl|Fobs|; where Fobs and Fcalc are the observed and calculated structure factor amplitudes,
respectively, for reflections hkl included in the refinement.c Rfree is the same as the R‐factor but calculated over a randomly selected fraction (5%) of reflection data not included in the refinement.
906 Crystal Struture of Methanococcus jannaschii RelBE
addition of 2.5 μl of protein complex in buffer C and 2.5 μlof reservoir solution containing 10% 2-methyl-2,4-penta-nediol (MPD) and 0.1 M Hepes (pH 7.5). The drop wasequilibrated over 500 μl of reservoir solution; after 2 days,single, irregularly shaped crystals grew to a size of 0.4–0.9 mm. For seeding experiments, a single crystal wasground with a spatula in the crystallization drop. Thedebris were diluted with 200 μl of stabilizing solutionconsisting of 22% MPD, 250 mM NaCl and 0.1 M Hepes(pH 7.5), vortexed and further used as seed stock. Finalcrystals were prepared by streak seeding in a hangingdrop over a 400-μl reservoir consisting of 20% MPD and0.1 M Bis–Tris (pH 5.5). The crystallization drop wasprepared with 2.5 μl of protein solution in buffer C and2.5 μl of reservoir solution. Streak-seeding was performedby dipping a horse tail hair in the seed stock and streakingwith it through three prepared drops in a row in order toobtain a dilution series of the seeding. The best crystalswere obtained after 2 days in the last seeded drop. Theyshowed a regular rhombohedral habitus and were directlyflash-frozen without further cryoprotectant. The crystalsdiffracted to 2.1 Å resolution at synchrotron BESSY2(Table 3).For heavy-atom derivatisation, ethylmercury phos-
phate (C2H7HgO4P) and cis-dichlorodiammineplatinum(III) [cis-Pt(NH3)2Cl2] were cocrystallized with MjRelBE,and MAD (multiwavelength anomalous diffraction) datawere collected with a resolution up to 2.6 and 2.7 Å reso-lution, respectively.
Structure determination and refinement
X-ray diffraction data were integrated with DENZO andscaled with SCALE.40 Since molecular replacement using
the structure of the homologous RelBE complex from P.horikoshii11 (PhRelBE) as search model (24% sequenceidentity for all residues of the toxin) was not successful,crystals were derivatised with heavy atoms. Since MADphasing was unsuccessful due to poor anomalous signal,phasing was performed with MIRAS (multiple isomor-phous replacement with anomalous signal), using themercury and platinum derivative data sets. Heavy-atomsearch, substructure refinement and electron densitymodification were done in SHARP.41 Initial modelbuilding was done with RESOLVE,42 and the completemodel was built manually using COOT.43 The model wasrefined with REFMAC544 (including TLS refinement) fromthe CCP4 package45 and using Anneal, Refine andBindividual from the CNS46 package. Details of datacollection and refinement statistics are given in Table 3.
PDB accession numbers
X-ray coordinates and structure factors have beendeposited at the PDB and are available as entry 3BPQ.
Acknowledgements
Thisworkwas performed in the frame of EUprojectQLK3-CT-2001-00277. We thank Caterina Farnleitnerfor the pET21relBE_MJ construct, Dr. WernerSchröder for N-terminal sequencing, Dr. Peter Frankefor MALDI-TOF analysis, and Prof. J. Behlke for ana-lytical ultracentrifugation analyses of M. jannaschii

907Crystal Struture of Methanococcus jannaschii RelBE
MjRelBE. Allocation of PSF beamline BL1 of FreieUniversität Berlin at BESSY/Berlin is gratefullyacknowledged. F.D. would like to thank the Ministryfor Science and Technological Development of theRepublic of Serbia for support thru Project 142010.
Supplementary Data
Supplementary data associated with this articlecan be found, in the online version, at doi:10.1016/j.jmb.2009.08.048
References
1. Gerdes, K., Rasmussen, P. B. & Molin, S. (1986). Uni-que type of plasmid maintenance function: post-segregational killing of plasmid-free cells. Proc. NatlAcad. Sci. USA, 83, 3116–3120.
2. Ogura, T. & Hiraga, S. (1983). Mini-F plasmid genesthat couple host cell division to plasmid proliferation.Proc. Natl Acad. Sci. USA, 80, 4784–4788.
3. Masuda, Y., Miyakawa, K., Nishimura, Y. & Ohtsubo,E. (1993). chpA and chpB, Escherichia coli chromo-somal homologs of the pem locus responsible forstable maintenance of plasmid R100. J. Bacteriol. 175,6850–6856.
4. Alonso, J. C., Balsa, D., Cherny, I., Christensen, S. K.,Francuski, D., Gazit, E. et al. (2007). Bacterial toxin–antitoxin systems as targets for the development ofnovel antibiotics. In Enzyme-Mediated Resistance toAntibiotics: Mechanism, Dissemination, and Prospects forInhibition (Bonomo, R. A. & Tolmasky, M. E., eds),pp. 313–329, ASM Press, Washington, DC.
5. Gerdes, K., Christensen, S. K. & Løbner-Olesen, A.(2005). Prokaryotic toxin–antitoxin stress response loci.Nat. Rev. Microbiol. 3, 371–382.
6. Anantharaman, V. & Aravind, L. (2003). New con-nections in the prokaryotic toxin–antitoxin network:relationship with the eukaryotic nonsense-mediatedRNA decay system. Genome Biol. 4, R81.
7. Shyu, A., Wilkinson, M. F. & van Hoof, A. (2008).Messenger RNA regulation: to translate or to degrade.EMBO J. 27, 471–481.
8. Buts, L., De Jonge, N., Loris, R., Wyns, L. & Dao-Thi,M. (2005). Crystallization of the C-terminal domainof the addiction antidote CcdA in complex with itstoxin CcdB. Acta Crystallogr., Sect. F: Struct. Biol.Cryst. Commun. 61, 949–952.
9. Kamphuis, M. B., Monti, M. C., van den Heuvel,R. H. H., López-Villarejo, J., Díaz-Orejas, R. & Boelens,R. (2007). Structure and function of bacterial kid-kisand related toxin–antitoxin systems. Protein Pept. Lett.14, 113–124.
10. Kamada, K., Hanaoka, F. & Burley, S. K. (2003). Crystalstructure of the MazE/MazF complex: molecular basesof antidote–toxin recognition. Mol. Cell, 11, 875–884.
11. Takagi, H., Kakuta, Y., Okada, T., Yao, M., Tanaka, I.& Kimura, M. (2005). Crystal structure of archaealtoxin–antitoxin RelE–RelB complex with implicationsfor toxin activity and antitoxin effects.Nat. Struct. Mol.Biol. 12, 327–331.
12. Kamada, K. & Hanaoka, F. (2005). Conformationalchange in the catalytic site of the ribonuclease YoeBtoxin by YefM antitoxin. Mol. Cell, 19, 497–509.
13. Prysak, M. H., Mozdzierz, C. J., Cook, A. M., Zhu, L.,Zhang, Y., Inouye, M. & Woychik, N. A. (2009). Bacte-
rial toxin YafQ is an endoribonuclease that associateswith the ribosome and blocks translation elongationthrough sequence-specific and frame-dependentmRNA cleavage. Mol. Microbiol. 71, 1071–1087.
14. Schmidt, O., Schuenemann, V. J., Hand, N. J., Silhavy,T. J., Martin, J., Lupas, A. N & Djuranovic, S. (2007).prlF and yhaV encode a new toxin–antitoxin system inEscherichia coli. J. Mol. Biol. 372, 894–905.
15. Liu, M., Zhang, Y., Inouye, M. & Woychik, N. A.(2008). Bacterial addiction module toxin Doc inhibitstranslation elongation through its association with the30S ribosomal subunit. Proc. Natl Acad. Sci. USA, 105,5885–5890.
16. Gazit, E. & Sauer, R. T. (1999). The Doc toxin and Phdantidote proteins of the bacteriophage P1 plasmidaddiction system form a heterotrimeric complex. J. Biol.Chem. 274, 16813–16818.
17. Miallau, L., Faller, M., Chiang, J., Arbing, M., Guo, F.,Cascio, D & Eisenberg, D. (2009). Structure andproposed activity of a member of the VapBC familyof toxin–antitoxin systems. VapBC-5 from Mycobacte-rium tuberculosis. J. Biol. Chem. 284, 276–283.
18. Schumacher, M. A., Piro, K. M., Xu, W., Hansen, S.,Lewis, K. & Brennan, R. G. (2009). Molecular mecha-nisms of HipA-mediated multidrug tolerance and itsneutralization by HipB. Science, 323, 396–401.
19. Meinhart, A., Alonso, J. C., Sträter, N. & Saenger, W.(2003). Crystal structure of the plasmid maintenancesystem epsilon/zeta: functional mechanism of toxinzeta and inactivation by epsilon 2 zeta 2 complexformation. Proc. Natl Acad. Sci. USA, 100, 1661–1666.
20. Sevin, E. W. & Barloy-Hubler, F. (2007). RASTA-Bacteria: a web-based tool for identifying toxin–antitoxin loci in prokaryotes. Genome Biol. 8, R155.
21. Graille, M., Mora, L., Buckingham, R. H., vanTilbeurgh, H. & de Zamaroczy, M. (2004). Structuralinhibition of the colicin D tRNase by the tRNA-mimicking immunity protein. EMBO J. 23, 1474–1482.
22. Lin, Y., Elias, Y. & Huang, R. H. (2005). Structural andmutational studies of the catalytic domain of colicinE5: a tRNA-specific ribonuclease. Biochemistry, 44,10494–10500.
23. Uzan, M., Favre, R. & Brody, E. (1988). A nuclease thatcuts specifically in the ribosome binding site of someT4 mRNAs. Proc. Natl Acad. Sci. USA, 85, 8895–8899.
24. Christensen, S. K., Mikkelsen, M., Pedersen, K. &Gerdes, K. (2001). RelE, a global inhibitor of transla-tion, is activated during nutritional stress. Proc. NatlAcad. Sci. USA, 98, 14328–14333.
25. Pedersen, K., Zavialov, A. V., Pavlov, M. Y., Elf, J.,Gerdes, K. & Ehrenberg, M. (2003). The bacterial toxinRelE displays codon-specific cleavage of mRNAs inthe ribosomal A site. Cell, 112, 131–140.
26. Christensen, S. K. & Gerdes, K. (2003). RelE toxinsfrom bacteria and Archaea cleave mRNAs on trans-lating ribosomes, which are rescued by tmRNA. Mol.Microbiol. 48, 1389–1400.
27. Overgaard, M., Borch, J., Jørgensen, M. G. & Gerdes,K. (2008). Messenger RNA interferase RelE controlsrelBE transcription by conditional cooperativity. Mol.Microbiol. 69, 841–857.
28. Li, G., Zhang, Y., Inouye, M. & Ikura, M. (2008).Structural mechanism of transcriptional autorepres-sion of the Escherichia coli RelB/RelE antitoxin/toxinmodule. J. Mol. Biol. 380, 107–119.
29. Kedzierska, B., Lian, L. & Hayes, F. (2007). Toxin–antitoxin regulation: bimodal interaction of YefM–YoeB with paired DNA palindromes exerts transcrip-tional autorepression. Nucleic Acids Res. 35, 325–339.

908 Crystal Struture of Methanococcus jannaschii RelBE
30. Bernard, P., Gabant, P., Bahassi, E. M. & Couturier, M.(1994). Positive-selection vectors using the F plasmidccdB killer gene. Gene, 148, 71–74.
31. Andreev, D., Hauryliuk, V., Terenin, I., Dmitriev, S.,Ehrenberg, M. & Shatsky, I. (2008). The bacterial toxinRelE induces specific mRNA cleavage in the A site ofthe eukaryote ribosome. RNA, 14, 233–239.
32. Bond, C. S. (2003). TopDraw: a sketchpad for proteinstructure topology cartoons. Bioinformatics, 19, 311–312.
33. Krissinel, E. & Henrick, K. (2007). Inference of macro-molecular assemblies from crystalline state. J. Mol.Biol. 372, 774–797.
34. Moretti, S., Armougom, F., Wallace, I. M., Higgins,D. G., Jongeneel, C. V. & Notredame, C. (2007). The M-Coffee Web server: a meta-method for computingmultiple sequence alignments by combining alternativealignment methods. Nucleic Acids Res. 35, W645–W648.
35. Gouet, P., Courcelle, E., Stuart, D. I. & Métoz, F.(1999). ESPript: analysis of multiple sequence align-ments in PostScript. Bioinformatics, 15, 305–308.
36. Lu, G. (2000). TOP: a new method for protein struc-ture comparisons and similarity searches. J. Appl.Crystallogr. 33, 176–183.
37. Saïda, F. & Odaert, B. (2007). RNA recognition andcleavage by sequence-specific endoribonucleases.Protein Pept. Lett. 14, 103–111.
38. Lebars, I., Hu, R. M., Lallemand, J. Y., Uzan, M. &Bontems, F. (2001). Role of the substrate confor-mation and of the S1 protein in the cleavage effi-ciency of the T4 endoribonuclease RegB. J. Biol. Chem.276, 13264–13272.
39. Andrade, M., Chacon, P., Merelo, J. & Moran, F.(1993). Evaluation of secondary structure of proteinsfrom UV circular dichroism spectra using an unsu-pervised learning neural network. Protein Eng. 6,383–390.
40. Otwinowski, Z. & Minor, W. (1997). Processing ofX-ray diffraction data collected in oscillation mode.Methods Enzymol. 276, 307–326.
41. Vonrhein, C., Blanc, E., Roversi, P. & Bricogne, G.(2007). Automated structure solution with auto-SHARP. Methods Mol. Biol. 364, 215–230.
42. Terwilliger, T. C. (2003). Automated main-chainmodel building by template matching and iterativefragment extension. Acta Crystallogr., Sect. D: Biol.Crystallogr. 59, 38–44.
43. Emsley, P. & Cowtan, K. (2004). Coot: model-buildingtools for molecular graphics. Acta Crystallogr., Sect. D:Biol. Crystallogr. 60, 2126–2132.
44. Murshudov, G. N., Vagin, A. A. & Dodson, E. J.(1997). Refinement of macromolecular structures bythe maximum-likelihood method. Acta Crystallogr.,Sect. D: Biol. Crystallogr. 53, 240–255.
45. Collaborative Computational Project, No. 4. (1994).The CCP4 suite: programs for protein crystallo-graphy. Acta Crystallogr., Sect. D: Bio. Crystallogr. 50,760–763.
46. Brünger, A. T., Adams, P. D., Clore, G. M., DeLano,W. L., Gros, P., Grosse-Kunstleve, R. W. et al. (1998).Crystallography & NMR system: a new software suitefor macromolecular structure determination. ActaCrystallogr., Sect. D: Biol. Crystallogr. 54, 905–921.





























