Embed Size (px)
Citation preview

of April 10, 2019.This information is current as
or Mouse LeukocytesCXCR7 Protein Is Not Expressed on Human
Juan C. Jaen and Thomas J. SchallPereda, Jessica Dias, Igor I. Slukvin, Kathleen E. McGrath,Susanna Lewén, Yu Wang, Zhenhua Miao, Lin Gan, Jaime Robert D. Berahovich, Brian A. Zabel, Mark E. T. Penfold,
http://www.jimmunol.org/content/185/9/5130doi: 10.4049/jimmunol.1001660October 2010;
2010; 185:5130-5139; Prepublished online 1J Immunol
Referenceshttp://www.jimmunol.org/content/185/9/5130.full#ref-list-1
, 14 of which you can access for free at: cites 31 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2010 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
CXCR7 Protein Is Not Expressed on Human or MouseLeukocytes
Robert D. Berahovich,* Brian A. Zabel,* Mark E. T. Penfold,* Susanna Lewen,*
Yu Wang,* Zhenhua Miao,* Lin Gan,† Jaime Pereda,‡ Jessica Dias,x Igor I. Slukvin,x
Kathleen E. McGrath,{ Juan C. Jaen,* and Thomas J. Schall*
Since the discovery that CXCR7 binds to CXCL12/SDF-1a, the role of CXCR7 in CXCL12-mediated biological processes has been
under intensive scrutiny. However, there is no consensus in the literature on the expression of CXCR7 protein by peripheral
blood cells. In this study we analyzed human and mouse leukocytes and erythrocytes for CXCR7 protein expression, using a
competitive CXCL12 binding assay as well as by flow cytometry and immunohistochemistry using multiple CXCR7 Abs.
CXCR72/2 mice were used as negative controls. Together, these methods indicate that CXCR7 protein is not expressed by
human peripheral blood T cells, B cells, NK cells, or monocytes, or by mouse peripheral blood leukocytes. CXCR7 protein is,
however, expressed on mouse primitive erythroid cells, which supply oxygen to the embryo during early stages of development.
These studies therefore suggest that, whereas CXCR7 protein is expressed by primitive RBCs during murine embryonic de-
velopment, in adult mammals CXCR7 protein is not expressed by normal peripheral blood cells. The Journal of Immunology,
2010, 185: 5130–5139.
The interaction between the chemokine CXCL12/SDF-1and its receptor CXCR4 has been the subject of experi-mental investigation for almost 15 years. This receptor–
ligand pair has been implicated in a wide range of biologicalprocesses, including development, stem cell biology, angiogene-sis, leukocyte trafficking, cancer, neurodegenerative disorders, in-flammatory disorders, atherosclerosis, and HIV pathology (1–9).Until recently, CXCL12 and CXCR4 were thought to be exclusivepartners—a relative rarity in the chemokine network. However, asecond chemokine receptor, CXCR7, was identified, which bindsto CXCL12 with even higher affinity than CXCR4 (10, 11).CXCR7 also binds to a second chemokine, CXCL11/I-TAC, oneof the ligands for CXCR3 (10). Unlike CXCR4, which triggerscalcium mobilization and chemotaxis in response to CXCL12,CXCR7 does not effectively initiate these processes. In fact, theCXCR7-mediated cellular responses to CXC12 and CXCL11 arelargely unknown, although ligand binding does trigger b-arrestin2association with CXCR7 and ligand/receptor internalization in vitro(12–14). CXCR7 regulates CXCR4/CXCL12-mediated processes,such as transendothelial migration of tumor cells and human renalprogenitors (14, 15) and primordial germ cell migration duringzebrafish development (16, 17), underlining a complex interplayamong CXCL12, CXCR4, and CXCR7.Most leukocyte subsets express CXCR4 and respond function-
ally to CXCL12 in vitro and in vivo. To determine whether these
leukocytes also express CXCR7, we and others have analyzedthe cells by flow cytometry or immunocytochemistry with CXCR7-specific mAbs. Four CXCR7 mouse mAbs have been described:11G8, generated byDNA immunization (10, 18); 9C4, generated byN-terminal peptide immunization (11, 19); and 8F11 and 358426,generated by immunization with CXCR7 cell transfectants (14, 20).Analysis of CXCR7 expression on leukocyte subsets with theseAbs has yielded inconsistent results (11, 19–21). Balabanian et al.(11) reported that mAb 9C4 stained a majority of cultured humanT cells by immunocytochemistry. Infantino et al. (19) reported that9C4 stained all human peripheral B cells and monocytes, but onlya small fraction of peripheral T cells, by flow cytometry. Hartmannet al. (20) reported that 11G8, 9C4, and 358426 did not stain pu-rified human T cells by flow cytometry. Instead, that group foundthat the mAbs stained the cells after permeabilization, indicatingthat the purified T cells expressed CXCR7 only intracellularly (20).Because CXCL12 is reported to bind only to CXCR4 and
CXCR7 with high affinity, CXCR7 protein can be detected usinga competitive [125I]–CXCL12 binding assay. In this assay, CXCR7is distinguished from CXCR4 by the competition of CXCL11,CXCL12, or CXCR7-specific compounds (such as CCX733 andCCX771) for the same binding site, and the lack of competition bythe CXCR4 antagonist AMD3100 (10, 18). Indeed, the originalhypothesis that there existed a second receptor for CXCL12emerged from observations at ChemoCentryx that [125I]–CXCL12could bind to mouse embryonic day (E)13 fetal liver cells fromCXCR42/2 mice (10). Although the CXCL12 binding assay hasbeen used to identify CXCR7 protein expression by these fetalliver cells, renal cell progenitors, vascular smooth muscle cells,and certain tumor lines (10, 13, 15), it has not yet been applied toleukocytes.In this study, we used the competitive [125I]–CXCL12 binding
assay and flow cytometry with all four CXCR7 mAbs to evaluateCXCR7 protein expression by normal human and mouse leuko-cytes. T cells from CXCR72/2 mice were used to assess the spec-ificity of the flow cytometric method employed by other in-vestigators to detect CXCR7 protein after prolonged incubation(20). We also used the competitive CXCL12 binding assay, flow
*ChemoCentryx, Inc., Mountain View, CA 94043; †Flaum Eye Institute and {Centerfor Pediatric Biomedical Research, University of Rochester, Rochester, NY 14642;‡Human Embryology Unit, University of Santiago de Chile, Santiago, Chile; andxPathology and Laboratory Medicine, University of Wisconsin, Madison, WI 53706
Received for publication May 19, 2010. Accepted for publication August 20, 2010.
Address correspondence and reprint requests to Dr. Robert D. Berahovich, Chemo-Centryx, Inc., 850 Maude Avenue, Mountain View, CA 94043. E-mail address:[email protected]
Abbreviations used in this paper: E, embryonic day; ESC, embryonic stem cell; IHC,immunohistochemistry.
Copyright� 2010 by TheAmericanAssociation of Immunologists, Inc. 0022-1767/10/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1001660
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

cytometry, and immunohistochemistry (IHC) to determine whichcells in mouse fetal liver express CXCR7.
Materials and MethodsPrimary Abs and reagents
CXCR7 mAb 11G8 was generated at ChemoCentryx (Mountain View, CA)and has been described previously (10, 18). mAb 9C4 was purchased fromMBL International (Woburn, MA). mAb 8F11 and isotype control mAbIgG2b were purchased from BioLegend (San Diego, CA). mAb 358426,isotype control mAbs IgG1 and IgG2a, and recombinant CXCL11 andCXCL12 were purchased fromR&DSystems (Minneapolis, MN). A humanBcl-2–specific mAb was purchased from BD Biosciences (San Jose, CA).[125I]–CXCL12 (labeling performed by the lactoperoxidase procedure) waspurchased from PerkinElmer (Wellesley, MA). AMD3100 was purchasedfromSigma-Aldrich (St. Louis,MO).CXCR7-specific compoundsCCX451,CCX733, CCX754, and CCX771 were generated by the ChemoCentryxMedicinal Chemistry Department and described previously (10, 14).
Cells and tissues
The human breast tumor cell line MDA MB 435s and its CXCR7 trans-fectant (“435-CXCR7”) cells have been described previously (10, 18). 293-mCXCR7 cells were generated by electroporation of HEK293 cells withthe pcDNA3.1 vector (Invitrogen, Carlsbad, CA) expressing murineCXCR7 and cultured in DMEM containing 10% FBS (Mediatech, Wash-ington, DC). B lymphoblast Raji and prostate cancer PC3 cell lines wereobtained from the American Type Culture Collection (Manassas, VA) andcultured in RPMI 1640 medium (Mediatech) containing 10% FBS. Theosteosarcoma K7M2 cell line was obtained from the American TypeCulture Collection and cultured in DMEM containing 10% FBS. Theglioma U343 cell line was donated by Christine Hsieh (University ofCalifornia, San Francisco, CA) and cultured in DMEM containing 10%FBS. Human normal bone marrow cells were purchased from AllCells(Emeryville, CA). Human thymocytes were obtained by mechanical dis-ruption of fetal thymus tissue purchased from Advanced BioscienceResources (Alameda, CA). Human PBMCs were collected from buffycoats (Stanford Blood Center, Stanford, CA) by centrifugation on Ficoll-Paque (GE Healthcare, Freiburg, Germany) and rinsed with PBS. T cells,B cells, and monocytes were isolated from the PBMCs with CD3, CD19,and CD14 (respectively) magnetic microbeads according to the manu-facturer’s instructions (Miltenyi Biotec, Auburn, CA). PBLs were isolatedfrom the PBMCs by depletion with CD14 microbeads. Mouse peripheralblood leukocytes were prepared from freshly drawn blood of BALB/c miceby erythrocyte lysis with Pharmlyse (BD Biosciences). Splenocytes wereprepared from BALB/c spleens by mechanical disruption on 70-mm nylonstrainers (BD Biosciences) and erythrocyte lysis with RBC Lysing Buffer(Sigma-Aldrich). Mouse E13 blood cells were collected from pregnantmice, as follows. Uteri were rinsed in PBS and opened along the length ofmaternal uterine muscle to expose the intact conceptuses, with decidualtissues and placenta attached. The yolk sac was punctured and peeled backto reveal its intact connection with the placenta. The umbilical vesselswere squeezed close near the abdomen with tweezers, and the embryo wasimmersed briefly in PBS to remove maternal blood cells. The embryo wasthen transferred to PBS containing 12.5 mg/ml heparin, and the tweezerswere removed, allowing the embryo blood to flow out of the umbilicalvessels. Afterward, the blood cells were collected by centrifugation andrinsed with PBS. Freshly collected c57BL/6 mouse and human blood weremixed with Pharmlyse (BD Biosciences) to lyse erythrocytes, after whichthe leukocytes were collected by centrifugation and rinsed with PBS.Collection of cells from mice was performed in accordance with theChemoCentryx Institutional Animal Care and Use Committee. An intacthuman embryo with normal appearance was obtained from an emergencysalpingectomy performed for tubal ectopic pregnancy at the Department ofObstetrics and Gynecology, Barros-Luco Hospital, Santiago, Chile. Theage of the embryo was determined to be 32 d postconception by extrap-olation from the patient’s last menstrual period, and then, with use ofa stereomicroscope, confirmed by the anatomical characteristics of thespecimen. The study was approved by the Ethics Committee of the Facultyof Medical Sciences, Santiago de Chile University. Human primitiveerythroid cells were generated in culture as follows. The human embryonicstem cell (ESC) line H1 (passage 31–47; WiCell Research Institute,Madison, WI) was maintained in an undifferentiated state by coculturewith mouse embryonic fibroblasts (WiCell Research Institute). To inducehematopoietic differentiation, H1 cells were cocultured with OP9 cells(provided by Toru Nakano at Osaka University, Osaka, Japan), as pre-viously described (22). After 8 d of coculture, cells were harvested by
treatment with 1 mg/ml collagenase (Invitrogen) for 25 min, followed by0.05% trypsin + 0.5 EDTA (Invitrogen) for 20 min at 37˚C. Cells werewashed with a-modified Eagle medium (Invitrogen) containing 10% FBS,filtered through a 70-mm cell strainer and cultured in a poly 2-hydrox-yethyl methacrylate (Sigma-Aldrich)-coated flask with serum-free medium(Stem Cell Technologies, Vancouver, BC, Canada) containing 0.3%Ex-Cyte (Millipore, Bedford, MA), 1 mg/ml iron-saturated transferrin(Sigma-Aldrich), 1026 M dexamethasone (Sigma-Aldrich), 20 ng/ml in-sulin (Sigma-Aldrich), 50 ng/ml stem cell factor, 2 U/ml erythropoietin, 50ng/ml thrombopoietin, 5 ng/ml IL-3, 10 ng/ml IL-6, and 200 ng/ml Flt3L(all from PeproTech, Rocky Hill, NJ). After 2 d, the culture was harvestedand mechanically disaggregated. Subsequently, the cells were spun over
FIGURE 1. CXCR7 mAb 11G8 stains mouse primitive erythroid cells
by IHC. Whole-mount sections of E13 mouse embryos were analyzed by
IHC using mAb 11G8. Staining in liver sinuses was detected by fluores-
cence IHC, using the Alexa Fluor 488 fluorochrome (green, top). Staining
in heart lumen was detected by enzymatic IHC, using the fuchsin stain
(red, bottom). Mouse IgG1 was used as an isotype control Ab. Original
magnification 3400.
FIGURE 2. Mouse primitive erythroid cells express CXCR7. A, Freshly
isolated, nonpermeabilized blood cells from mouse E13 embryos were
analyzed by flow cytometry. The cells were stained by mAb 11G8 (purple
histogram), but not by the mouse IgG1 isotype control Ab (green overlay).
B, Freshly isolated blood cells from mouse E13 embryos were analyzed by
the competitive [125I]–CXCL12 binding assay. [125I]–CXCL12 binding
was inhibited by CXCL12, CXCL11, and the CXCR7 antagonist CCX733,
but not by the CXCR4 antagonist AMD3100. The means 6 SE of four
wells for each concentration of inhibitor are shown.
The Journal of Immunology 5131
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
20% Percoll (Sigma-Aldrich) to remove dead cells and cell aggregates, asdescribed previously (23). The viable cells were then cultured on a layer ofirradiated MS-5 stromal cells (DSMZ, Braunschweig, Germany) using themedium described above but lacking thrombopoietin, IL-3, and IL-6. Half-medium changes were performed every 2 d thereafter, maintaining the celldensity near 1–2 3 106 cells/ml. Cells were collected after 15 d of ex-pansion for analysis.
Construction and analysis of CXCR7 knockout mice
To generate a CXCR7lacZ targeting construct, the 2.9-kb 59 flanking se-quence immediately upstream of the CXCR7 translation initiation codonand the 4.3-kb 39 flanking sequence starting 1.1 kb downstream of theCXCR7 translation initiation codon were inserted at the NheI site and theSalI–KpnI sites of the pKII-lacZ vector containing lacZ (L. Gan, un-published observations), respectively. The targeting construct thus containsthe lacZ reporter gene under the control of 59 and 39 regulatory sequences
of CXCR7. To generate CXCR7lacZ knockin mice, the targeting constructwas linearized at the KpnI site and inserted into W4 ESCs (Taconic Farms,Germantown, NY) by electroporation. Five targeted mouse ESC cloneswere obtained from a total of 192 G418-resistant ESC clones. The targetedclones were confirmed by Southern blotting and PCR genotyping, andinjected into C57BL/6J blastocysts to generate mouse chimeras. CMV-Cremice (The Jackson Laboratory, Bar Harbor, ME) were used to remove theneomycin resistance gene to generate the CXCR7lacZ allele. CXCR7lacZheterozygous and homozygous mice were generated and maintained ina mixed 129S6 and C57BL/6J background.
RT-PCR
Total RNA was isolated from human PBLs, purified human leukocytesubsets, mouse peripheral blood leukocytes, mouse splenocytes, humanESC-derived erythroid cells, and endogenous CXCR7-expressing humanand mouse cell lines, using the RNAeasy Kit (Qiagen, Valencia, CA). First-strand cDNAwas generated using the GeneAmp RNA PCR Core Kit withpoly dTTT primers (Applied Biosystems, Foster City, CA). PCR of cDNAsamples was performed using Taq PCR Master Mix Kit (Qiagen). Primersets were as follows: human CXCR7, 59-AGCACAGCCAGGAAGG-CGAG-39 and 59-TCATAGCCTGTGGTCTTGGC-39; mouse CXCR7, 59-GGAGCCTGCAGCGCTCACCG-39 and 59-CTTAGCCTGGATATTCA-CCC-39; human ε-globin, 59-GCCTGTGGAGCAAGATGAAT-39 and 59-GCGGGCTTGAGGTTGT-39; universal GAPDH, 59-CATTGACCTCAA-CTACATGG-39 and 59-GGGCCATCCACAGTCTTCTG-39.
QuantiGene analysis
435-CXCR7 cells, untransfected 435 cells (20,000 cells per reaction), andhuman PBLs (150,000 cells per reaction) were analyzed for CXCR7,
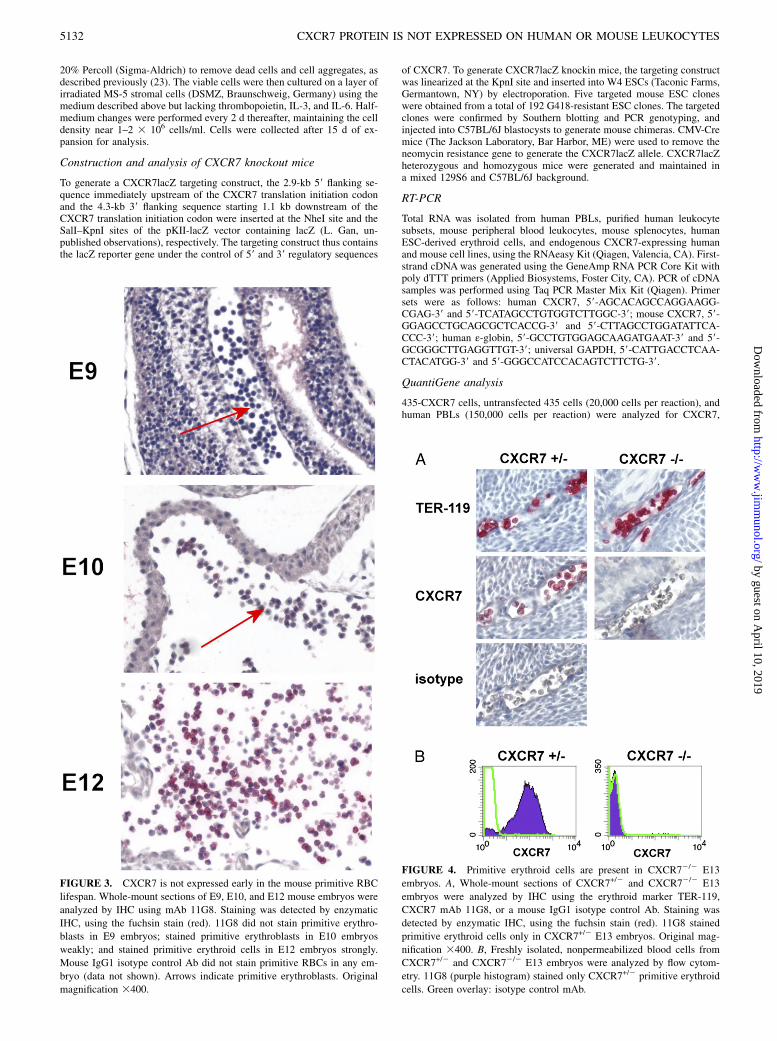
FIGURE 3. CXCR7 is not expressed early in the mouse primitive RBC
lifespan. Whole-mount sections of E9, E10, and E12 mouse embryos were
analyzed by IHC using mAb 11G8. Staining was detected by enzymatic
IHC, using the fuchsin stain (red). 11G8 did not stain primitive erythro-
blasts in E9 embryos; stained primitive erythroblasts in E10 embryos
weakly; and stained primitive erythroid cells in E12 embryos strongly.
Mouse IgG1 isotype control Ab did not stain primitive RBCs in any em-
bryo (data not shown). Arrows indicate primitive erythroblasts. Original
magnification 3400.
FIGURE 4. Primitive erythroid cells are present in CXCR72/2 E13
embryos. A, Whole-mount sections of CXCR7+/2 and CXCR72/2 E13
embryos were analyzed by IHC using the erythroid marker TER-119,
CXCR7 mAb 11G8, or a mouse IgG1 isotype control Ab. Staining was
detected by enzymatic IHC, using the fuchsin stain (red). 11G8 stained
primitive erythroid cells only in CXCR7+/2 E13 embryos. Original mag-
nification 3400. B, Freshly isolated, nonpermeabilized blood cells from
CXCR7+/2 and CXCR72/2 E13 embryos were analyzed by flow cytom-
etry. 11G8 (purple histogram) stained only CXCR7+/2 primitive erythroid
cells. Green overlay: isotype control mAb.
5132 CXCR7 PROTEIN IS NOT EXPRESSED ON HUMAN OR MOUSE LEUKOCYTES
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
CXCR4, and b-actin mRNA with the branched DNA-based QuantiGeneAssay according to the manufacturer’s instructions (Genospectra, Fremont,CA). Briefly, cell lysates were hybridized with specific probe sets over-night at 53˚C, after which amplifier reagents and labeled probe sets wereadded and incubated for 1 h at 53˚C after washing. Substrate reagents wereadded and incubated for 30 min, after which the plates were analyzed forchemiluminescence.
Flow cytometry
Mouse E13 blood cells, human thymocytes, and cells derived from humanESCs were stained with 11G8 or mouse IgG1 isotype control mAbs at 10mg/ml on ice for 20 min and then rinsed with cold buffer (PBS containing2% FBS). Cells were then incubated on ice for 20 min with PE-conjugated F(ab9)2 fragments of donkey anti-mouse IgG (Jackson Immu-noResearch Laboratories, West Grove, PA) and rinsed with cold buffer.Human peripheral blood leukocytes were stained with CXCR7 mAbs,isotype control mAbs, or Bcl-2 mAbs, as described above, then incubatedon ice for 10 min with mouse serum (final concentration 5%) (JacksonImmunoResearch Laboratories) and mixed with fluorochrome-conjugatedAbs (FITC anti-human CD3, PE-Cy5 anti-human CD19, Alexa Fluor 647anti-human CD16, and allophycocyanin-Cy7 anti-human CD14, all fromBD Biosciences) on ice for 30 min. Human bone marrow leukocytes werestained with 11G8 in a similar fashion, except the lineage Abs were FITCanti-human CD3, allophycocyanin anti-human CD19, allophycocyanin-Cy7 anti-human CD14, and PE-Cy5 anti-human CD34 (all from BDBiosciences). Mouse leukocytes were stained with CXCR7 mAbs or iso-type control mAbs at 10 mg/ml on ice for 20 min and then rinsed with coldbuffer. Cells were then incubated on ice for 30 min with FITC-conjugatedanti-mouse Ly6G, PE-conjugated anti-mouse CD3, PE-Cy5–conjugatedanti-mouse B220 (all from BD Biosciences), and allophycocyanin-conjugated F(ab9)2 fragments of donkey anti-mouse IgG (Jackson Immu-noResearch Laboratories). Cells were rinsed with cold buffer and analyzedwith a FACScan (BD Biosciences). For permeabilization experiments,human leukocytes were first incubated in PBS containing 3% para-formaldehyde (Sigma-Aldrich) for 5 min, collected by centrifugation, andthen incubated in PBS containing 0.2% saponin (Sigma-Aldrich) for 10min. Cells were then rinsed with buffer and stained with Abs, as indicatedabove. For prolonged incubation experiments, allophycocyanin-conjugated
11G8 (R&D Systems) was added to total leukocytes from CXCR7+/+ andCXCR72/2 mixed background mice (see above) at 10 mg/ml in the ab-sence or presence of CXCR7-specific compounds at 2.5 mM, and the cellswere incubated at 37˚C for 2 h. Cells were collected by centrifugation,suspended in cold buffer, and incubated on ice for 30 min with FITC-conjugated anti-mouse CD3 (BD Biosciences). Cells were rinsed coldwith buffer and analyzed with the FACScan.
Immunohistochemistry
Mouse and human embryos were fixed in 10% neutral-buffered formalin(Sigma-Aldrich), dehydrated, and embedded in paraffin, and then 5-mmsections were cut and transferred to glass slides. Sections were deparaffi-nized with xylenes (Sigma-Aldrich) and hydrated through graded alcohols(Sigma-Aldrich) by conventional methods. For enzymatic IHC, sectionswere rinsed in water and immersed in TBS containing 1% BSA (Sigma-Aldrich) for 10 min. CXCR7 mAbs or isotype control Abs were added tothe sections at 10 mg/ml and incubated at room temperature for 1 h.Sections were rinsed thoroughly in TBST and incubated with biotinylatedanti-mouse IgG (Jackson ImmunoResearch Laboratories) for 30 min.Sections were rinsed thoroughly in TBST and incubated with streptavidin-conjugated alkaline phosphatase (Dako, Glostrup, Denmark) for 20 min.Sections were rinsed thoroughly in TBST and incubated with Fuchsin+
reagent (Dako) for 5–10 min. Sections were rinsed with water and coun-terstained with Mayer’s hematoxylin (Sigma-Aldrich) for 3 min. Sectionswere rinsed with water and coverslipped with Faramount (Dako). Forfluorescence IHC, sections were rinsed in water and immersed in TBScontaining 1% BSA, 0.1% Triton X-100, and 0.05% Tween-20 (all fromSigma-Aldrich) for 30 min. CXCR7 mAbs or isotype control Abs wereadded to the sections at 10 mg/ml and incubated at room temperature for1 h. Sections were rinsed thoroughly in TBST and incubated with AlexaFluor 488-conjugated anti-mouse IgG (Invitrogen) for 30 min. Sectionswere rinsed thoroughly in PBS and incubated with DAPI (Invitrogen) for15 min. Sections were rinsed thoroughly in PBS, transferred to water,and coverslipped with GelMount (VWR, West Chester, PA). All imageswere acquired on a Nikon Eclipse E800 microscope and captured witha SPOT RT-3 camera and imaging software (Diagnostic Instruments,Twinsburg, OH).
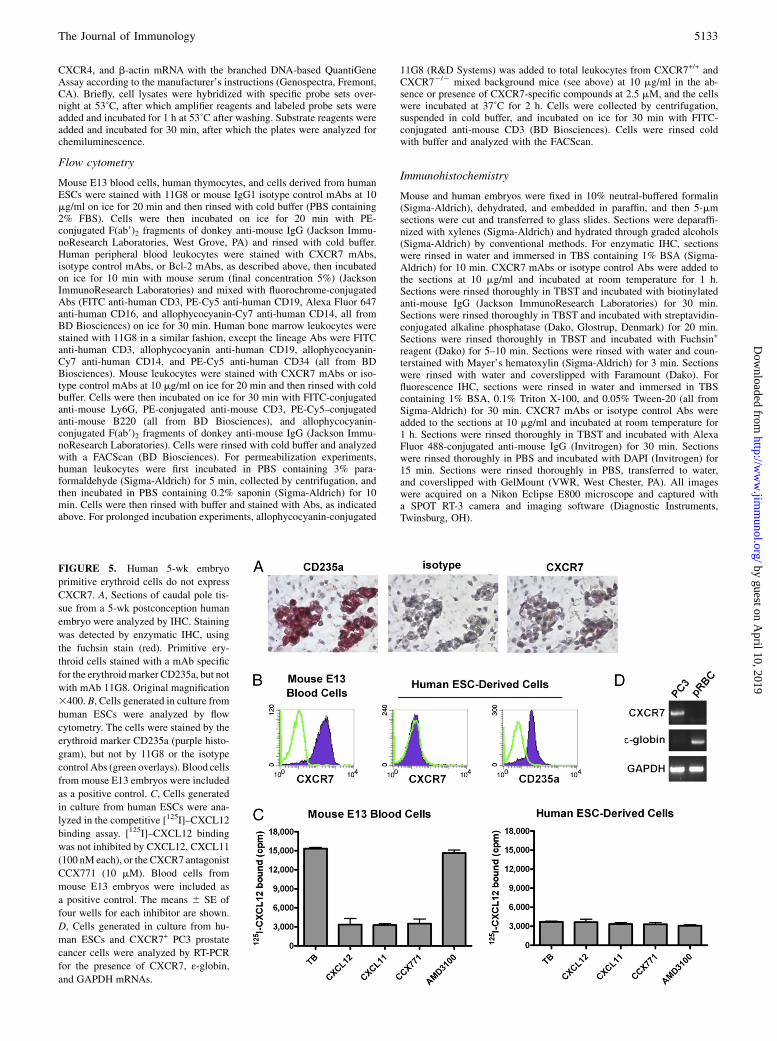
FIGURE 5. Human 5-wk embryo
primitive erythroid cells do not express
CXCR7. A, Sections of caudal pole tis-
sue from a 5-wk postconception human
embryo were analyzed by IHC. Staining
was detected by enzymatic IHC, using
the fuchsin stain (red). Primitive ery-
throid cells stained with a mAb specific
for the erythroidmarker CD235a, but not
with mAb 11G8. Original magnification
3400. B, Cells generated in culture from
human ESCs were analyzed by flow
cytometry. The cells were stained by the
erythroid marker CD235a (purple histo-
gram), but not by 11G8 or the isotype
control Abs (green overlays). Blood cells
from mouse E13 embryos were included
as a positive control. C, Cells generated
in culture from human ESCs were ana-
lyzed in the competitive [125I]–CXCL12
binding assay. [125I]–CXCL12 binding
was not inhibited by CXCL12, CXCL11
(100 nM each), or the CXCR7 antagonist
CCX771 (10 mM). Blood cells from
mouse E13 embryos were included as
a positive control. The means 6 SE of
four wells for each inhibitor are shown.
D, Cells generated in culture from hu-
man ESCs and CXCR7+ PC3 prostate
cancer cells were analyzed by RT-PCR
for the presence of CXCR7, ε-globin,and GAPDH mRNAs.
The Journal of Immunology 5133
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

CXCL12 binding assay
Cells were suspended at 1–5 million/ml in HBSS containing 0.1% BSA andtransferred to 96-well plates (100 ml/well) containing 5 ml/well recombi-nant CXCL11 or CXCL12 (final concentration 100 nM in single-doseassays) or compounds (final concentration 10 mM in single-dose assays).Then 100 ml/well cold buffer containing 0.025 mCi (∼1 nM) [125I]–CXCL12 was added, and the plates were shaken at 4˚C for 3 h. Cells weretransferred onto polyethyleneimine-treated GF/B glass fiber filters (Per-kinElmer) with a cell harvester (Tomtec, Hamden, CT) and washed twicewith washing buffer (25 mM HEPES, 500 mM NaCl, 1 mM CaCl2, 5 mMMgCl2, pH 7.1). Next 50 ml/well MicroScint-20 (PerkinElmer) was added
to the filters, and counts per minute were measured on a Packard TopCountScintillation Counter (PerkinElmer).
ResultsCXCR7 protein is expressed on mouse primitive erythroid cells
We previously reported that total liver cells from mouse E13embryos express CXCR7 protein, as determined by a competitive[125I]–CXCL12 binding assay and by flow cytometry with mAb11G8 (10). We next sought to identify the cell type in the fetalliver that expresses CXCR7 protein. By IHC on E13 mouse
FIGURE 6. Adult human and mouse peripheral blood cells do not bind to CXCR7-specific mAbs. A, Freshly isolated mouse blood cells were analyzed by
flow cytometry using CXCR7 mAbs 11G8, 9C4, and 8F11. Abs specific for T cells (CD3) and neutrophils (Ly6G) were included in the stains. Open
histograms indicate CXCR7 mAb staining; color-matched filled histograms indicate isotype control Ab staining. HEK293 cells transfected with mouse
CXCR7 (293-mCXCR7) were included as a positive control. CXCR7 mAb 358426 did not stain 293-mCXCR7 cells, indicating that this mAb does not
recognize mouse CXCR7. B and C, Freshly isolated human blood was depleted of erythrocytes and analyzed by flow cytometry using CXCR7 mAbs 11G8,
9C4, 8F11, and 358426 as well as a mAb specific for the intracellular protein Bcl-2. Abs specific for T cells (CD3), B cells (CD19), monocytes (CD14), and
NK cells (CD16) were included in the stains. Neutrophils were identified based on light scatter properties and CD16 expression. Open histograms indicate
CXCR7 mAb staining; color-matched filled histograms indicate isotype control Ab staining. MDA MB 435 cells transfected with human CXCR7 (435-
CXCR7) were included as a positive control. Surface staining is shown in B, and staining following permeabilization is shown in C. D, Freshly isolated
blood from CXCR72/2 mice or CXCR7+/+ littermates was depleted of erythrocytes, incubated with CXCR7 mAb 11G8 or isotype control mAb at 37˚C for
2 h in the presence or absence of CXCR7-specific compounds CCX733 or CCX771, and analyzed by flow cytometry for mAb binding to T cells. The
means 6 SE of four wells for each inhibitor are shown.
5134 CXCR7 PROTEIN IS NOT EXPRESSED ON HUMAN OR MOUSE LEUKOCYTES
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

embryos, 11G8 did not stain fetal hepatocytes, but rather stainednearly all of the primitive erythroid cells present in the large sinusesof the liver (Fig. 1). 11G8 also stained primitive erythroid cellselsewhere in the embryo, both inside (Fig. 1) and outside the vas-culature (data not shown). 11G8 stained primitive RBCs by flowcytometry as well (Fig. 2A). [125I]–CXCL12 binding to primitiveerythroid cells was inhibited by CXCL12 (IC50 value 0.2 nM),CXCL11 (IC50 value 0.4 nM), and the CXCR7-selective compoundCCX733 (IC50 value 2.4 nM) but not by the CXCR4 antagonistAMD3100, thus confirming CXCR7 protein expression (Fig. 2B).In contrast, definitive (fetal) erythrocytes did not stain with 11G8by IHC, either during development in erythroblast islands in thefetal liver or after release into the vasculature (data not shown).To determine if CXCR7 is expressed early in the mouse primitive
erythroid cell lifespan,weanalyzedembryos fromE9 toE12by IHC.11G8did not stain primitive erythroblasts inmouseE9 embryos, andonly weakly stained these cells in E10 embryos (Fig. 3). The in-tensity of staining increased thereafter, with strong staining inprimitive erythroblasts in E12 embryos (Fig. 3). These data indicatethat CXCR7 is not expressed by immature primitive erythroblasts,but is instead induced as these cells mature over time during em-bryonic development.To investigate the role of CXCR7 on primitive erythroid cells,
we generated CXCR7-deficient mice. Primitive erythroblasts, de-fined as nucleus-containing TER-119+ cells, were present in bothCXCR7+/2 and CXCR72/2 E13 embryos (Fig. 4). 11G8 did notstain primitive erythroid cells of E13 CXCR72/2 embryos byeither IHC or flow cytometry (Fig. 4). E13 CXCR72/2 embryosdid not exhibit any gross histological or morphological changescompared with CXCR7+/2 or CXCR7+/+ embryos, and containednormal numbers of primitive erythroid cells with unaltered mor-phology (data not shown). These results confirm that CXCR7protein is expressed by mouse primitive erythroid cells, and in-dicate that CXCR7 expression is not required for the developmentor survival of these cells.
To determine whether CXCR7 is expressed on human primitiveerythroid cells, we used two approaches. First, we performed IHCon sections of the caudal pole from a human embryo 5 wk post-conception, an age at which all blood cells are primitive erythro-blasts. Human primitive erythroblasts were identified by stainingwith CD235a, a surface sialoglycoprotein expressed during allstages of erythropoeisis, and by the presence of a nucleus. 11G8 didnot stain human primitive erythroblasts present in this tissue (Fig.5A). Next, we analyzed erythroid cells generated in vitro fromhuman ESCs. CD235a+ cells generated from the H1 ESC line didnot stain with 11G8 (Fig. 5B), did not exhibit a CXCR7 profile inthe competitive CXCL12 binding assay (Fig. 5C), and did notcontain detectable CXCR7 mRNA (Fig. 5D). These cells wereconfirmed to be primitive erythroid cells by the presence of he-moglobin ε mRNA (Fig. 5D).
CXCR7 protein is not expressed on or in primary leukocytes
We next evaluated CXCR7 protein expression in definitive (adult)mouse erythrocytes and in mouse and human leukocyte subsetsby flow cytometry, using all available CXCR7 mAbs. First, wedetermined the specificity of the mAbs, using CXCR72 cellstransfected with CXCR7 or empty expression vectors. For mouseCXCR7, mAbs 11G8, 9C4, and 8F11 stained 293-mCXCR7 cells(Fig. 6A), but not 293-empty vector cells (data not shown). mAb358426 did not stain 293-mCXCR7 cells (Fig. 6A), indicating thatit does not bind to mouse CXCR7. For human CXCR7, mAbs11G8, 9C4, 8F11, and 358426 stained 435-CXCR7 cells (Fig. 6B),but not 435-empty vector cells (data not shown). On primary cells,mAbs 11G8, 9C4, and 8F11 did not stain mouse peripheral blooderythrocytes, T cells, or neutrophils (Fig. 6A). Similarly, mAbs11G8, 9C4, and 8F11 did not stain T cells, B cells, NK cells,monocytes, or neutrophils from freshly drawn human blood (Fig.6B). mAb 358426 stained human monocytes but did not stain theother leukocyte subsets (Fig. 6B). In addition, all four CXCR7mAbs failed to stain each human leukocyte subset after per-
FIGURE 7. Human and mouse peripheral blood leukocytes do not express CXCR7 protein. Magnetic bead-purified human T cells, B cells, and
monocytes and erythrocyte-depleted total mouse blood leukocytes were assessed for CXCR7 protein expression by the competitive [125I]–CXCL12 binding
assay. [125I]–CXCL12 binding was not inhibited by CXCL12, CXCL11 (100 nM each), or the CXCR7 antagonists CCX451 or CCX733 (10 mM) in any
sample. 435-CXCR7 cells were included as a positive control. The means 6 SE of four wells for each inhibitor are shown.
The Journal of Immunology 5135
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

meabilization; effective permeabilization was confirmed by posi-tive staining for the intracellular protein Bcl-2 (Fig. 6C). mAb358426 failed to stain 435-CXCR7 cells and monocytes afterpermeabilization, likely indicating that the target epitope is alteredby fixation (Fig. 6C).Other investigators have reported that, although 11G8 failed to
stain intact purified human T cells under normal conditions, the Abdid stain the cells if they were incubated together for 2 h at 37˚C(20). We attempted to reproduce this finding but also employedleukocytes from both wild-type and CXCR72/2 mice as a speci-ficity control. After 2 h at 37˚C, 11G8 stained wild-type mouseT cells to a small degree (Fig. 6D). However, not only wasstaining not inhibited by CXCR7-specific compounds, but stainingalso occurred on T cells from CXCR72/2 mice (Fig. 6D). Thesedata suggest that the 11G8 staining seen after prolonged incu-bation at 37˚C is artifactual or nonspecific.In addition, we evaluated plasma membrane expression of
CXCR7 on purified human T cells, B cells, and monocytes, as wellas total mouse peripheral blood leukocytes, with the competitive[125I]–CXCL12 binding assay. Although CXCL12, CXCL11, andthe CXCR7-specific small molecule CCX733 blocked [125I]–CXCL12 binding to 435-CXCR7 cells, none of the primary cell
types displayed a CXCR7 binding profile, confirming that these-cells do not express cell-surface CXCR7 protein (Fig. 7).We also analyzed leukocytes in human bone marrow and thy-
mus for CXCR7 protein expression (Fig. 8). By flow cytometry,CXCR7 mAbs 11G8, 9C4, and 8F11 did not stain any cell type
FIGURE 8. Human bone marrow leukocytes and thymocytes do not express CXCR7 protein. Freshly isolated human bone marrow cells (A) and freshly
isolated human fetal thymocytes (B) were analyzed in the competitive [125I]–CXCL12 binding assay (left) and by flow cytometry using CXCR7 mAb 11G8
(right). [125I]–CXCL12 binding was not inhibited by CXCL12, CXCL11 (100 nM each), or the CXCR7 antagonists CCX754 or CCX771 (10 mM) in either
sample. 435-CXCR7 cells (C) were used as a positive control. The means6 SE of four wells for each inhibitor are shown. 11G8 (purple histograms) did not
stain bone marrow hematopoietic progenitor cells (CD34+), T cells (CD3+), B cells (CD19+), monocytes (CD14+), or granulocytes (high side scatter), nor
did 11G8 stain CD4+CD8+ immature thymocytes or CD4+CD82 or CD42CD8+ mature thymocytes. Green overlays indicate isotype control Ab staining.
CXCR7 mAbs 9C4 and 8F11 did not stain any cell subset either (not shown). 435-CXCR7 cells (C) were used as a positive control.
FIGURE 9. CXCR7+/+ and CXCR72/2 mouse leukocytes respond
equally to CXCL12. Freshly isolated mouse bone marrow leukocytes from
CXCR7+/+ and CXCR72/2 mice were tested for chemotaxis to varying
concentrations of CXCL12 in vitro. Equal numbers of CXCR7+/+ and
CXCR72/2 cells migrated to CXCL12, and the potency of CXCL12 was
equal for CXCR7+/+ and CXCR72/2 cells.
5136 CXCR7 PROTEIN IS NOT EXPRESSED ON HUMAN OR MOUSE LEUKOCYTES
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

(T cells, B cells, monocytes, granulocytes, and CD34+ progenitorcells) in bone marrow. In addition, 11G8, 9C4, and 8F11 did notstain immature CD4+CD8+, mature CD4+CD82, or mature CD42
CD8+ thymocytes. Bone marrow leukocytes and thymocytes didnot display the CXCR7 profile in the competitive [125I]–CXCL12binding assay, indicating that leukocytes in these two organs donot express cell-surface CXCR7 protein.Because CXCR7 has been shown to affect CXCR4-mediated
responses to CXCL12 (14, 15, 20, 24), we reasoned that if mouseleukocytes do not express CXCR7, then genetic ablation of CXCR7should not affect leukocyte responses to CXCL12. Indeed, bonemarrow leukocytes from CXCR7+/+ and CXCR72/2mice migratedto CXCL12 equally well in chemotaxis assays in vitro (Fig. 9).To determinewhether human andmouse leukocytes lack CXCR7
protein because they lack CXCR7 mRNA, we analyzed these cellsfor CXCR7 mRNA expression (Fig. 10). In total human PBLs,CXCR7mRNAwas undetectable byRT-PCR andQuantiGene anal-ysis. In addition, little to no CXCR7mRNAwas detected in purifiedhuman T cells, B cells, and monocytes by RT-PCR. CXCR7mRNAwas also not detected in total peripheral blood cells or splenocytesfrom BALB/c mice. These data indicate that the lack of CXCR7protein in human andmouse leukocytes is primarily due to transcrip-tional, not translational, regulation.
DiscussionIn these studies, we used a competitive [125I]–CXCL12 bindingassay, flow cytometry, and IHC to assess human and mouse normalblood cells for expression of CXCR7 protein. We first followed upon our initial observation that cells in mouse E13 embryo liver ex-press CXCR7 protein (10). In this paper, we show that these cells are
primitive erythroid cells. Primitive erythroid cells collected fromwild-type, but not CXCR72/2, mouse E13 embryos exhibited thesignature CXCR7 binding profile in the [125I]–CXCL12 bindingassay: [125I]–CXCL12 binding was inhibited by CXCL11- andCXCR7-specific compounds, but not by the CXCR4 antagonistAMD3100. In addition, CXCR7 mAb 11G8 stained wild-type,but not CXCR72/2, primitive erythroid cells by IHC and flowcytometry.The role of CXCR7 on murine primitive erythroid cells is un-
certain. CXCR72/2 embryos were unaltered in the numbers ofprimitive erythroid cells, and no gross changes in morphology wereobserved. Indeed, most of our CXCR72/2 mice do not die untilsoon after birth, similarly to other CXCR72/2 mice (25, 26). HenceCXCR7 is not required for the development, proliferation, or sur-vival of primitive erythroid cells. CXCR7 is also not likely nec-essary for primitive erythrocyte function, as mutations that disruptprimitive erythrocyte function cause mouse embryo lethality byE10.5 (27). Interestingly, CXCR7 protein was not expressed bywild-type mouse primitive erythroid cells as they develop in theyolk sac and enter circulation (which occurs at E9). Instead, lowlevels of CXCR7 protein appeared on the cell surface at E10 (Fig.3), and the staining intensity increased over time, reaching a max-imum at E13. The ability to detect CXCR7 in liver or blood cellsis lost by E17 (10), whether through downregulation on primitiveerythrocytes or owing to the presence of vast numbers of (CXCR72)definitive erythrocytes produced by this stage of development.Although we were able to detect enucleated CXCR72/2 primitiveerythrocytes, it is possible that CXCR7 serves to enhance primi-tive RBC enucleation, which occurs between E12.5 and E17.5 (27).It is also possible that CXCR7 plays an even later role in the
FIGURE 10. Human and mouse leukocytes express virtually no CXCR7 mRNA. Human and mouse leukocyte subsets were assessed for CXCR7 mRNA
expression by RT-PCR and QuantiGene analysis. A, RNA preparations from human PBLs (left gel) and purified T cells, B cells, and monocytes (right gel)
were analyzed by RT-PCR and agarose gel electrophoresis. Two donors are shown for each cell population. CXCR7 mRNA was either not detected or
detected at low levels in each sample. Positive controls included CXCR7+ Raji cells and U343 cells. GAPDH control reactions demonstrated that the
substrate RNAs were intact. B, RNA preparations from mouse peripheral blood leukocytes and splenocytes were analyzed by RT-PCR and agarose gel
electrophoresis. Mouse CXCR7 mRNA was either not detected or detected at low levels in each sample. The positive control was K7M2 cells. GAPDH
control reactions demonstrated that the substrate RNAs were intact. C, QuantiGene analysis was performed on human PBLs, with 435-CXCR7 cells and
untransfected 435 cells serving as positive and negative control cells, respectively. CXCR7 mRNA was not detected in any PBL sample. CXCR4 and
b-actin control reactions demonstrated that the substrate RNAs were intact. The means 6 SE of three wells for each reaction are shown.
The Journal of Immunology 5137
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

primitive erythrocyte lifespan, although it is not known whetherprimitive erythrocytes are actively removed from the embryo orare simply swamped out by the definitive erythrocytes (28).In contrast to mouse primitive erythroid cells, human primitive
erythroid cells do not appear to express CXCR7 at the same stage ofdevelopment. Primitive erythroblasts present in a 5-wk-old humanfetus (Theiler stage 15, which corresponds to mouse E12) did notstain with CXCR7 mAb 11G8. In addition, primitive erythroblastsgenerated in culture from human ESCs did not stain with 11G8 orexhibit the CXCR7 profile in the competitive CXCL12 bindingassay. However, it is possible that CXCR7 protein is induced onhuman primitive erythroid cells at earlier or later stages oferythropoeisis than in the mouse.Wewere unable to procure humanembryos containing primitive erythroid cells at these other stages.In this study, we also determined that CXCR7 protein is not
expressed on adult human or mouse leukocytes. T cells, B cells,NK cells, monocytes, and neutrophils in freshly isolated humanblood and bone marrow each failed to stain with CXCR7 mAbs11G8, 9C4, and 8F11 by flow cytometry. These three mAbs alsofailed to stain human thymocytes and mouse peripheral blooderythrocytes, T cells, and neutrophils. CXCR7 mAb 358426, soldby R&D Systems, stains human monocytes but also binds to celllines lacking cell-surface CXCR7 and fails to bind to tumor celllines endogenously expressing CXCR7 (21). CXCR7 polyclonalAbs ab12870 and ab38039, sold by Abcam (Cambridge, MA),stains tissues from CXCR72/2 mice by IHC (21), indicatingthat not all CXCR7 Abs are reactive or specific for CXCR7.Moreover, by IHC, none of the CXCR7 mAbs (including 9C4)stained leukocytes in human tissue sections (R.D. Berahovich, un-published observations), in contrast to a previous report wherein9C4 was observed to stain follicular B cells in human tonsil (19).Finally, purified human T cells, B cells, and monocytes, as well astotal leukocytes in human bone marrow, thymus, and mouse pe-ripheral blood, each did not display the CXCR7 profile in thecompetitive [125I]–CXCL12 binding assay. In fact, most humanand mouse leukocyte samples did not contain detectable CXCR7mRNA either, as assessed by RT-PCR and QuantiGene analysis.CXCR7 mRNA expression levels in human leukocytes have pre-viously been reported, but with inconsistent results (19, 29–31).These results differ from results in several published reports (11,
19, 20), which in turn differ among themselves. Those reportsstate the following: CXCR7 is expressed on the surface of mostT cells (where it was reported to mediate chemotaxis) (11); on allB cells and monocytes, but only a small subset of T cells (19); andinside (but not on the surface of) all T cells (20). In the last ofthese reports, the authors performed flow cytometry on T cellsseveral hours after blood draw, after multiple steps of enrichment(20). Because we did not observe CXCR7 mAb staining insideT cells in freshly drawn blood, it is possible that the staining seenby these investigators (20) was due to extensive manipulation ofthe cells. The authors also showed that 11G8 could stain theirpurified T cells without permeabilization if incubated for 2 h at37˚C. We showed that under these unusual conditions, 11G8equally stains T cells from wild-type and CXCR72/2 mice.Finally, we used a functional assay to explore whether mouse
leukocytes express CXCR7 protein. Because CXCR7 modulatesCXCR4 functional responses to CXCL12, we reasoned that ifCXCR7 protein is present on mouse leukocytes, leukocytes fromCXCR7+/+ and CXCR72/2 mice should respond differently toCXCL12. However, leukocytes from CXCR7+/+ and CXCR72/2
mice migrated with equal efficacy and potency to CXCL12 inchemotaxis assays in vitro. Thus, using the most sensitive detectionmethods available, we have demonstrated that CXCR7 protein is notexpressed by normal mouse or human peripheral blood leukocytes.
DisclosuresThe authors have no financial conflicts of interest.
References1. Alkhatib, G. 2009. The biology of CCR5 and CXCR4. Curr Opin HIV AIDS 4:
96–103.2. Braunersreuther, V., F. Mach, and S. Steffens. 2007. The specific role of che-
mokines in atherosclerosis. Thromb. Haemost. 97: 714–721.3. Gelmini, S., M. Mangoni, M. Serio, P. Romagnani, and E. Lazzeri. 2008. The
critical role of SDF-1/CXCR4 axis in cancer and cancer stem cells metastasis. J.Endocrinol. Invest. 31: 809–819.
4. Iwamoto, T., H. Okamoto, Y. Toyama, and S.Momohara. 2008. Molecular aspectsof rheumatoid arthritis: chemokines in the joints of patients. FEBS J. 275: 4448–4455.
5. Levesque, J. P., and I. G. Winkler. 2008. Mobilization of hematopoietic stemcells: state of the art. Curr. Opin. Organ Transplant. 13: 53–58.
6. Miller, R. J., G. Banisadr, and B. J. Bhattacharyya. 2008. CXCR4 signaling inthe regulation of stem cell migration and development. J. Neuroimmunol. 198:31–38.
7. Mines, M., Y. Ding, and G. H. Fan. 2007. The many roles of chemokinereceptors in neurodegenerative disorders: emerging new therapeutical strategies.Curr. Med. Chem. 14: 2456–2470.
8. Schober, A. 2008. Chemokines in vascular dysfunction and remodeling. Arte-rioscler. Thromb. Vasc. Biol. 28: 1950–1959.
9. Vandercappellen, J., J. Van Damme, and S. Struyf. 2008. The role of CXCchemokines and their receptors in cancer. Cancer Lett. 267: 226–244.
10. Burns, J. M., B. C. Summers, Y. Wang, A. Melikian, R. Berahovich, Z. Miao,M. E. Penfold, M. J. Sunshine, D. R. Littman, C. J. Kuo, et al. 2006. A novelchemokine receptor for SDF-1 and I-TAC involved in cell survival, cell adhe-sion, and tumor development. J. Exp. Med. 203: 2201–2213.
11. Balabanian, K., B. Lagane, S. Infantino, K. Y. Chow, J. Harriague, B. Moepps,F. Arenzana-Seisdedos, M. Thelen, and F. Bachelerie. 2005. The chemokineSDF-1/CXCL12 binds to and signals through the orphan receptor RDC1 inT lymphocytes. J. Biol. Chem. 280: 35760–35766.
12. Luker, K. E., M. Gupta, J. M. Steele, B. R. Foerster, and G. D. Luker. 2009.Imaging ligand-dependent activation of CXCR7. Neoplasia 11: 1022–1035.
13. Rajagopal, S., J. Kim, S. Ahn, S. Craig, C. M. Lam, N. P. Gerard, C. Gerard, andR. J. Lefkowitz. 2010. Beta-arrestin- but not G protein-mediated signaling by the“decoy” receptor CXCR7. Proc. Natl. Acad. Sci. USA 107: 628–632.
14. Zabel, B. A., Y. Wang, S. Lewen, R. D. Berahovich, M. E. Penfold, P. Zhang,J. Powers, B. C. Summers, Z. Miao, B. Zhao, et al. 2009. Elucidation of CXCR7-mediated signaling events and inhibition of CXCR4-mediated tumor celltransendothelial migration by CXCR7 ligands. J. Immunol. 183: 3204–3211.
15. Mazzinghi, B., E. Ronconi, E. Lazzeri, C. Sagrinati, L. Ballerini, M. L. Angelotti,E. Parente, R. Mancina, G. S. Netti, F. Becherucci, et al. 2008. Essential butdifferential role for CXCR4 and CXCR7 in the therapeutic homing of human renalprogenitor cells. J. Exp. Med. 205: 479–490.
16. Boldajipour, B., H. Mahabaleshwar, E. Kardash, M. Reichman-Fried, H. Blaser,S. Minina, D. Wilson, Q. Xu, and E. Raz. 2008. Control of chemokine-guided cell migration by ligand sequestration. Cell 132: 463–473.
17. Dambly-Chaudiere, C., N. Cubedo, and A. Ghysen. 2007. Control of cell mi-gration in the development of the posterior lateral line: antagonistic interactionsbetween the chemokine receptors CXCR4 and CXCR7/RDC1. BMC Dev. Biol.7: 23.
18. Miao, Z., K. E. Luker, B. C. Summers, R. Berahovich, M. S. Bhojani,A. Rehemtulla, C. G. Kleer, J. J. Essner, A. Nasevicius, G. D. Luker, et al. 2007.CXCR7 (RDC1) promotes breast and lung tumor growth in vivo and is expressedon tumor-associated vasculature. Proc. Natl. Acad. Sci. USA 104: 15735–15740.
19. Infantino, S., B. Moepps, and M. Thelen. 2006. Expression and regulation of theorphan receptor RDC1 and its putative ligand in human dendritic and B cells. J.Immunol. 176: 2197–2207.
20. Hartmann, T. N., V. Grabovsky, R. Pasvolsky, Z. Shulman, E. C. Buss, A. Spiegel,A. Nagler, T. Lapidot, M. Thelen, and R. Alon. 2008. A crosstalk between in-tracellular CXCR7 and CXCR4 involved in rapid CXCL12-triggered integrinactivation but not in chemokine-triggered motility of human T lymphocytes andCD34+ cells. J. Leukoc. Biol. 84: 1130–1140.
21. Berahovich, R. D., M. E. Penfold, and T. J. Schall. 2010. Nonspecific CXCR7antibodies. Immunol. Lett.133: 112–114.
22. Vodyanik, M. A., J. A. Bork, J. A. Thomson, and I. I. Slukvin. 2005. Humanembryonic stem cell-derived CD34+ cells: efficient production in the coculturewith OP9 stromal cells and analysis of lymphohematopoietic potential. Blood105: 617–626.
23. Vodyanik, M. A., and I. I. Slukvin. 2007. Directed differentiation of humanembryonic stem cells to dendritic cells. Methods Mol. Biol. 407: 275–293.
24. Levoye, A., K. Balabanian, F. Baleux, F. Bachelerie, and B. Lagane. 2009.CXCR7 heterodimerizes with CXCR4 and regulates CXCL12-mediated G pro-tein signaling. Blood 113: 6085–6093.
25. Gerrits, H., D. S. van Ingen Schenau, N. E. Bakker, A. J. van Disseldorp,A. Strik, L. S. Hermens, T. B. Koenen, M. A. Krajnc-Franken, and J. A. Gossen.2008. Early postnatal lethality and cardiovascular defects in CXCR7-deficientmice. Genesis 46: 235–245.
26. Sierro, F., C. Biben, L. Martınez-Munoz, M. Mellado, R. M. Ransohoff, M. Li,B. Woehl, H. Leung, J. Groom, M. Batten, et al. 2007. Disrupted cardiac de-velopment but normal hematopoiesis in mice deficient in the second CXCL12/SDF-1 receptor, CXCR7. Proc. Natl. Acad. Sci. USA 104: 14759–14764.
5138 CXCR7 PROTEIN IS NOT EXPRESSED ON HUMAN OR MOUSE LEUKOCYTES
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

27. McGrath, K., and J. Palis. 2008. Ontogeny of erythropoiesis in the mammalianembryo. Curr. Top. Dev. Biol. 82: 1–22.
28. Kingsley,P.D., J.Malik,K.A.Fantauzzo, and J.Palis. 2004.Yolk sac-derivedprimitiveerythroblasts enucleate during mammalian embryogenesis. Blood 104: 19–25.
29. Jin, Z., D. Nagakubo, A. K. Shirakawa, T. Nakayama, A. Shigeta, K. Hieshima,Y. Yamada, and O. Yoshie. 2009. CXCR7 is inducible byHTLV-1 Tax and promotesgrowth and survival of HTLV-1-infected T cells. Int. J. Cancer 125: 2229–2235.
30. Loos, T., A. Mortier, M. Gouwy, I. Ronsse, W. Put, J. P. Lenaerts, J. Van Damme,and P. Proost. 2008. Citrullination of CXCL10 and CXCL11 by peptidylargininedeiminase: a naturally occurring posttranslational modification of chemokinesand new dimension of immunoregulation. Blood 112: 2648–2656.
31. Shimizu, N., Y. Soda, K. Kanbe, H. Y. Liu, R. Mukai, T. Kitamura, and H. Hoshino.2000. A putative G protein-coupled receptor, RDC1, is a novel coreceptor forhuman and simian immunodeficiency viruses. J. Virol. 74: 619–626.
The Journal of Immunology 5139
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from