Embed Size (px)
Citation preview

Decolourization of synthetic dyes by a newly isolated strain of Serratia marcescens
Pradeep Verma*,� and Datta MadamwarPost-Graduate Department of Biosciences, Sardar Patel University, Vallabh Vidyanagar 388 120, Gujarat, India*Author for correspondence: E-mail: [email protected]�Present address: Institute of Microbiology, Academy of Sciences of the Czech Republic, Videnska 1083, 14220 Prague4, Czech Republic
Received 18 November 2002; accepted 15 March 2003
Keywords: Decolourization, laccase, manganese peroxidase, Serratia marcescens
Summary
A novel dye-decolourizing strain of the bacterium Serratia marcescens efficiently decolourized two chemicallydifferent dyes Ranocid Fast Blue (RFB) and Procion Brilliant Blue-H-GR (PBB-HGR) belonging respectively tothe azo and anthraquinone groups. Extracellular laccase and manganese peroxidase (MnP) activity were detectedduring dye decolourization. The involvement of MnP activity was found in the decolourization of both dyes.More than 90% decolourization of PBB-HGR and RFB was obtained on days 8 and 5, respectively at 26 �C understatic conditions at pH 7.0. MnP activity was increased by the addition of Mn2+. At 50 lM Mn2+, high MnP(55.3 U/ml) but low laccase activity (8.3 U/ml) was observed. Influence of oxalic acid on MnP activity was alsoobserved.
Introduction
The growth of the chemical industry has generated avariety of anthropogenic compounds. The production ofcertain chemicals is often associated with the formationof large amounts of undesired byproducts and wastes.Synthetic dyes are extensively used in textile dyeing,paper, printing, colour photography, pharmaceutics,cosmetics and other industries (Marmion 1991). Dur-ing production, 10–15% of synthetic dyes producedare discharged as such into the environment (Meyer1981). Since some of the dyes are harmful, dye-containingwastes pose an important environmental problem.Microorganisms are capable of utilizing a variety of
complex chemicals including dyes as sole source ofcarbon, but only meagre data are available on thebacterial breakdown of azo and anthraquinone dyes(Sarnaik & Kanekar 1995; Chang & Lin 2000). Manysynthetic dyes are recalcitrant to microbial degradation(Chung 1993). Microbial decolourization is an environ-ment-friendly and cost-competitive alternative to chem-ical decomposition processes. Attempts are being madeto develop bacterial strains for aerobic dye degradation.However, in many cases, adsorption of dyes to microbialsurfaces is the primary mechanism of decolourization(Wang & Yu 1998).The abilities of white rot fungi to degrade a wide
range of synthetic chemicals including dyes have been
widely reported, and the involvement of their non-specific lignin-degrading system for degradation ofsynthetic dyes has been demonstrated (Rodriguez et al.1999). Fungi are a promising group of organisms for dyedegradation, but their filamentous growth and lesshomogeneous substrate colonization pattern in additionto slow growth and oxygen limitation due to viscosity insubmerged conditions pose problems in their applica-tion. Hence, replacement of fungi with bacteria with agood degradation activity would offer special advanta-ges.Neem (Azadirachta indica) hull wastes are generated
by the neem oil industries and are essentially com-posed of lignocellulose and, hence, not easily degrad-able (Verma & Madamwar 2002b). It has beenpreviously reported that S. marcescens shows lignin-degrading activity (Rhoades et al. 1995). During thedegradation of neem hull waste we isolated thebacterial strain Serratia marcescens showing ligninoly-tic activity. Ranocid Fast Blue (RFB) and ProcionBrilliant Blue-HGR (PBB-HGR) were selected fordecolourization studies because of their wide use inthe textile industry as well as their known structure.This paper deals with biological decolourization ofselected dyes by the new isolate. An attempt has beenmade to optimize conditions for the efficient decolou-rization and the exploitation of the microorganism fordye removal.
World Journal of Microbiology & Biotechnology 19: 615–618, 2003. 615� 2003 Kluwer Academic Publishers. Printed in the Netherlands.
Materials and methods
Chemicals
ABTS (2,2¢-azino-bis-ethylbenzothiazoline sulphonicacid), veratryl alcohol (3,4-dimethoxybenzyl alcohol),o-dianisidine were from Sigma (USA). All other chem-icals were of analytical grade. The dyes RFB (kmax 538)and PBB-HGR (CI 61210) (kmax 604) were of industrialgrade procured from Aravind Mills, Ahemdabad India.The trivial names of dyes were used for convenience.
Organism and culture conditions
A bacterium was isolated from decomposed neem hullwaste. The characterization and identification of theisolate Serratia marcescens was done by The MicrobialType Culture Collection (MTCC), Institute of MicrobialTechnology, Chandigarh, India. The organism wasmaintained on nutrient medium containing 0.1% yeastextract, 0.1% glucose and 50 mg dye/l. It was sub-cultured once every 2 months.Two different nutrient media compositions were used
to select a medium for decolourization of synthetic dyes.Medium I: Nutrient-limited medium (NLM) used hadthe following composition (g/l): 1.0 KH2PO4, 0.5 KCl,0.5 MgSO4, 1.0 yeast extract, 1.0 glucose and final pH7.0. Medium II: Nutrient high medium (NHM) had thefollowing composition (g/l): 0.2 MgSO4, 1.0 K2HPO4,1.0 KH2PO4, 0.02 CaCl2, 0.05 FeCl3, 1.0 NH4NO3, 1.0yeast extract, 1.0 glucose and final pH 7.0.
Decolourization and enzyme activity assays
The flasks containing medium were sterilized byautoclaving. Inoculation of freshly grown bacterialsuspension (0.5 ml) was done in 50 ml medium con-taining the individual dye (100 mg/l final concentra-tion) in 250 ml Erlenmeyer flasks, the flasks wereincubated under static conditions at 26 �C. At regularintervals, 3 ml samples were withdrawn from the flasksand centrifuged at 8000 rev/min for 20 min; the super-natant was analysed for remaining dye content andenzyme activities. A control set without dye wasinoculated and the supernatant was analysed for theenzyme activities. Adsorption of dyes to bacterialbiomass was analysed using washed culture sedimentin distilled water. The absorbance was determined atRFB (kmax 538 nm) and PBB-HGR (kmax 604 nm)using spectrophotometer. All experiments were per-formed in quadruplicate.The decolourization was measured as the decrease in
absorbance at the absorbance maxima (kmax) of eachdye using u.v.–visible spectrophotometer (Hewlett–Packard 8452). Laccase activity was determined byoxidation of o-dianisidine according to Palmieri et al.(1993). MnP activity was determined according toKatagiri et al. (1995). Lignin peroxidase (LiP) activitywas assayed according to Tien & Kirk (1988).
Effect of pH, temperature, manganese and oxalic acid
The effect of pH on the decolourization and enzymeactivities was studied. The pH was adjusted with NaOHor HCl in the range of 3.0–8.0. The effect of temperaturewas studied during the incubation at 26, 37 and 45 �C.The effect of manganese (10–100 lM) and oxalic acid(10–100 lM) on decolourization and enzyme activitieswas also studied.
Detection of metabolites
The transformation products were analysed usingHPTLC on precoated Silica Gel 60 F (20 · 20) (EMerck, Germany). About 10 ll samples were appliedbandwise by Linomat IV Camag sampler, band length8 mm, track distance10 mm, distance from the side20 mm, distance from lower edge 10 mm, delivery speed15 s/ll, temperature 25 �C. The plate was developed in apresaturated (15 min) twin chamber containing solventsystems of ammonia:propanol (2:1 v/v) and the runningdistance was 80 mm for the separation of transforma-tion products. Spots were analysed by a Camag Scanner.
Results and discussion
In NHM complete decolourization of RFB and PBB-HGR was observed on days 5 and 8, respectively understatic conditions. In NLM highest decolourization ofRFB and PBB-HGR was observed on days 6 and 10under static conditions. The decolourization was drasti-cally reduced using agitated conditions (data not shown).Further studies were carried out using NHM because
of rapid decolourization in that medium. As shown in
0
6
12
18
24
30
0 2 4 6 8 10 12 14
Time (Days)
Enz
yme
Act
ivity
(U
/ml)
0
25
50
75
100(%
) D
ecol
oriz
atio
n
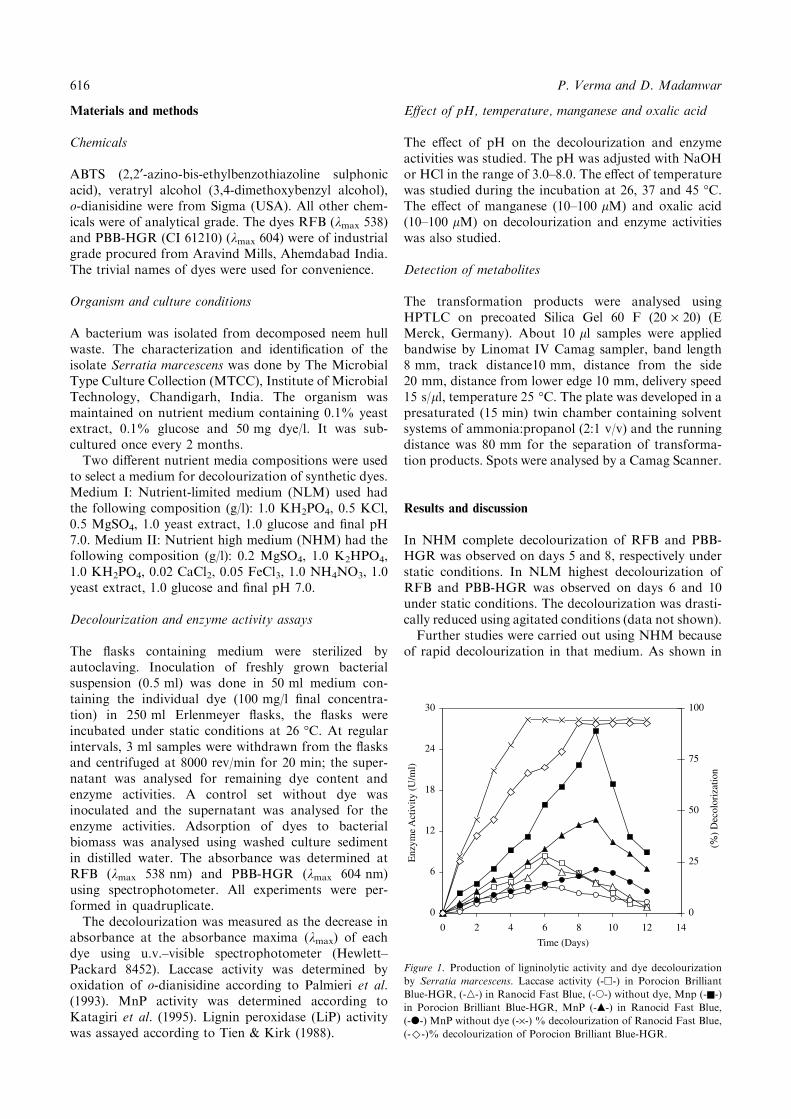
Figure 1. Production of ligninolytic activity and dye decolourization
by Serratia marcescens. Laccase activity (-h-) in Porocion Brilliant
Blue-HGR, (-n-) in Ranocid Fast Blue, (-s-) without dye, Mnp (-n-)
in Porocion Brilliant Blue-HGR, MnP (-m-) in Ranocid Fast Blue,
(-d-) MnP without dye (-·-) % decolourization of Ranocid Fast Blue,
(-)-)% decolourization of Porocion Brilliant Blue-HGR.
616 P. Verma and D. Madamwar

Figure 1, laccase and MnP activities were detectedduring dye decolourization both in presence and absenceof both dyes. LiP activity was not detected in the culturesupernatant. Laccase and MnP were detected from thefirst day. The laccase activity reached its peak on day 6(8.4 U/ml) while MnP reached the maximum (26.8 U/ml) on day 9 in the presence of PBB-HGR. Butsignificantly lower activities of both laccase and MnPwere detected in the absence of the dyes. S. marcescensshowed MnP activity four times higher in the presenceof PBB-HGR and two-fold higher in the presence ofRFB. Laccase had two-fold higher activity in thepresence of PBB-HGR. Laccase and MnP activitieswere lower in the presence of RFB compared to PBB-HGR. More than 70% decolourization of RFB wasobtained within 3 days while only 30–40% decolouriza-tion of PBB-HGR was obtained. Adsorption of dyes tobacterial biomass was not observed in the present study.It has been reported that anthraquinone dyes are directsubstrates of laccase (Schiliephake & Langergan 1998).Presence of higher laccase activity in PBB-HGR sug-gested that it is a substrate of laccase. On the other handRFB is an azo dye and is not a substrate of laccase(Pasti-Grigsby et al. 1992). Perestelo et al. (1994)reported laccase activity during dye decolourization byS. marcescens. To the best of our knowledge, our resultis the first report that MnP activity was detected in S.marcescens during dye decolourization.As shown in Figure 2, more than 90% decolouriza-
tion of RFB and PBB-HGR (100 mg/l final concentra-tion) was obtained by S. marcescens at pH 7.0 within 5and 8 days, respectively. In the range of temperaturetested the optimum temperature for decolourization andenzyme production was 26 �C.Recently, several reports on white rot fungi have
demonstrated the involvement of MnP during the
decolourization of synthetic dyes (Heinfling et al.1998). Culture liquid from S. marcescens with highestMnP activity was used in order to check the catalyticactivity of the extracellular enzymes produced by theorganism. Each dye (50 mg/l final concentration) wasprepared in acetate buffer (50 mM, pH 5.0). Manganese(1 mM final concentration), culture supernatant, andH2O2 (1 mM final concentration) were mixed andcomplete decolourization was obtained with both dyes.However, the supernatant did not decolourize the dyesin the absence of H2O2 or Mn2þ. This suggests theinvolvement of MnP in decolourization. The addition ofMn2+ to media increased decolourization rates andenzyme activities. The optimum concentration was50 lM for decolourization as well as enzyme production(Table 1).
Table 1. Effect of Mn2+ on dye decolourization and enzyme activities.
Mn2+ (lM) Laccase (lmol/ml) MnP (lmol/ml) RFB (kmax = 538)
(% decolourization)
Procion Brilliant Blue
(kmax = 604)
(% decolourization)
0 6.7 ± 0.9 26.8 ± 1.9 79.6 ± 2.4 82.7 ± 2.4
10 7.5 ± 1.1 45.5 ± 2.4 85.9 ± 1.8 87.3 ± 3.1
25 8.4 ± 1.3 48.9 ± 1.7 89.7 ± 3.1 90.4 ± 2.8
50 8.3 ± 0.8 55.3 ± 2.7 94.9 ± 1.9 93.5 ± 2.7
100 8.9 ± 1.5 54.4 ± 2.1 94.9 ± 2.7 92.4 ± 2.6
Table 2. Influences of oxalic acid on dye decolourization and enzyme activities.
Oxalic acid (lM) Laccase (lmol/ml) MnP (lmol/ml) RFB (kmax = 538)
(% decolourization)
Procion Brilliant Blue
(kmax = 604)
(% decolourization)
0 6.5 ± 1.1 25.9 ± 1.9 89.6 ± 1.7 82.7 ± 3.3
10 6.2 ± 0.9 28.9 ± 2.1 95.9 ± 2.5 86.5 ± 2.4
25 5.7 ± 1.4 34.7 ± 1.2 78.9 ± 1.6 73.7 ± 3.2
50 5.4 ± 1.7 21.9 ± 3.4 74.6 ± 1.8 65.8 ± 2.1
100 ND 8.9 ± 2.9 34.4 ± 3.2 29.8 ± 2.7
ND – Not detected.
0
20
40
60
80
100
3 3.5 4 4.5 5 5.5 6 6.5 7 7.5 8pH
% D
ecol
oriz
atio
n
Figure 2. Effect of initial pH media on dye decolourization by Serratia
marcescens on day 5 and 8 respectively. (-n-) PBB-HGR, (-h-) RFB.
Dye decolourization by Serratia marcescens 617

It has been reported that oxalic acid plays animportant role in degradation of recalcitrant com-pounds (e.g. lignin, PAHs) in wood-rotting fungi (Barr& Aust 1994; Green et al. 1995). The optimum concen-tration of oxalic acid for enzyme production was 25 lM(Table 2). High concentrations of oxalic acid may causea partial inhibition of bacterial growth (Roy & Archi-bald 1993).After the transformation of RFB (Rf ¼ 0.71), three
metabolites (Rf ¼ 0.23, 0.31 and 0.68) were detectedwhereas, in PBB-HGR (Rf ¼ 0.41), four different meta-bolites (Rf ¼ 0.25, 0.32, 0.63 and 0.79) were detected. Onemetabolite of RFB transformation might be metanalicacid withRf ¼ 0.68 and this was further confirmed by thepresence of a peak in the u.v. region. Detection ofmetanalic acid was same as in the case of Phanerochaetechrysosporium (Verma & Madamwar 2002a).The present study confirms that the use of the newly
isolated Serratia marcescens strain for decolourizationof structurally different synthetic dyes is possible. Thedye decolourization was very fast compared to white rotfungi. Nevertheless, the fungal-based treatment systemis less efficient and time-consuming due to the slowgrowth rate of fungi as well as high adsorption to thebiomass. The exploitation of S. marcescens wouldbe help to overcome the difficulties associated with theuse of mycelial organisms for dye decolourization andto develop an efficient technology for the removal ofdyes from dyes-containing effluents using a bacterialspecies.
Acknowledgements
This work was supported by Department of Biotech-nology, Government of India, New Delhi, India. Theauthor is thankful to Dr Petr Baldrian for his valuablesuggestions
References
Barr, D.P. & Aust, S.D. 1994 Mechanisms white rot fungi use to
degrade pollutants. Environmental Science and Technology 28,
79A–87A.
Chang, J. & Lin Y. 2000 Fed-batch bioreactor strategies for microbial
decolourization of azo dye using a Pseudomonas luteola strain.
Biotechnology Progress 16, 979–985.
Chung, K.T. & Stevens, S.E. Jr. 1993 Decolourization of azo dyes by
environmental microorganisms and helminths. Environmental
Toxicology and Chemistry 12, 2121–2132.
Green, F., Clausen, C.A., Larsen, M.J. & Highely, T.L. 1995 Induction
of polygalacturonase and the formation of oxalic acid by pectin in
brown rot fungi. World Journal of Microbiology and Biotechnology
11, 519–524.
Heinfling, A., Martinez, M.J., Martinez, A.T., Bergbauer, M. &
Szewzyk, U. 1998 Transformation of industrial dyes by manganese
peroxidases from Bjerkandera adusta and Pleurotus eryngii in a
manganese-independent reaction. Applied and Environmental
Microbiology 64, 2788–2793.
Katagiri, N., Tsutsumi, Y. & Nishida, T. 1995 Correlation of
brightening with cumulative enzyme activity related to lignin
biodegradation during biobleaching of kraft pulp by white rot
fungi in the solid-state fermentation system. Applied and Environ-
mental Microbiology 61, 617–627.
Marmion, D.M. 1991 Handbook of US Colorants, 3rd edn, pp. 573.
New York: John Wiley and Sons, Inc. ISBN 0-471-50074-7.
Meyer, U. 1981 Biodegradation of synthetic organic colorants. In
Microbial Degradation of Xenobiotics and Recalcitrant Com-
pounds, eds. Leisinger, T., Cook, A.M., Hutter, R. & Nuesch, J.
pp. 387–399. London: Academic Press. FEMS Symposium 12,
ISBN 0-12-442920-3.
Palmieri, G., Giardina, P., Marzullo, L., Desiderio, B., Nitti, B.,
Cannio, R. & Sannia, G. 1993 Stability and activity of phenol
oxidase from lignolytic fungus Pleurotus osteratus. Applied Micro-
biology and Biotechnology 39, 632–636.
Pasti-Grigsby, M.B., Paszczynski, A., Goszczynski, S., Crawford, D.L.
& Crawford, R.L. 1992 Influence of aromatic substitution patterns
on azo dye degradability by Streptomyces spp. and Phanerochaete
chrysosporium. Applied and Environmental Microbiology 58, 3605–
3613.
Perestelo, F., Falcon, M.A., Carnicero, A., Rodriguez, A. & Da La
Fuente, G. 1994 Limited degradation of industrial synthetic and
natural lignin by Serratia marcescens. Biotechnology Letters 16(3),
299–302.
Rhoades, T.L., Mikells, A.T., Jr. & Eley, M.H. 1995 Investigation of
the lignin-degrading activity of Serratia marcescens: biochemical
screening and ultrastructural evidence. Candian Journal of Micro-
biology 41(7), 592–600.
Rodriguez, E., Pickard, M.A. & Duhalt, R.V. 1999 Industrial dye
decolourization by laccases from lignolytic fungi. Current Micro-
biology 38, 27–32.
Roy, B.P. & Archibald, F. 1993 Effect of kraft pulp and lignin on
Trametes versicolor carbon catabolism. Applied and Environmental
Microbiology 59, 1855–1863.
Sarnaik, S. & Kanekar, P. 1995 Bioremediation of color of methyl
violet and phenol from a dye-industry waste effluent using
Pseudomonas sp. isolated from factory soil. Journal of Applied
Bacteriology 79, 459–469.
Schiliephake, K. & Langergan, G.T. 1998 Laccase variation during dye
decolourization in packed bed bioreactor. Applied and Environ-
mental Microbiology 18, 881–886.
Tien, M. & Krik, T.K. 1988 Lignin peroxidase of Phanerochaete
chrysosporium. Methods in Enzymology 161, 238–243.
Verma, P. & Madamwar, D. 2002a Decolourization of synthetic dyes
by lignin peroxidase of Phanerochaete chrysosporium. Folia Mi-
crobiologica 47, 283–286.
Verma, P. & Madamwar, D. 2002b Production of ligninolytic enzyme
by co-cultivation of P. ostreatus and P. chrysosporium and its
application towards dyes decolourization. Applied Biochemistry
and Biotechnology 102–103, 109–118.
Wang, Y. & Yu, J. 1998 Adsorption and degradation of synthetic dyes
on the mycelium of Trametes versicolor. Water Science and
Technology 38, 233–238.
618 P. Verma and D. Madamwar




























