Embed Size (px)
Citation preview

lable at ScienceDirect
Environmental Pollution 272 (2021) 116005
Contents lists avai
Environmental Pollution
journal homepage: www.elsevier .com/locate/envpol
DEHP-elicited small extracellular vesicles miR-26a-5p promotedmetastasis in nearby normal A549 cells*
Yifei Qin a, b, Jing Zhang a, c, Ricardo David Avell�an-Llaguno a, b, Xu Zhang a, b,Qiansheng Huang a, d, *
a Key Lab of Urban Environment and Health, Institute of Urban Environment, Chinese Academy of Sciences, Xiamen, 361021, PR Chinab College of Resources and Environment, University of Chinese Academy of Sciences, Beijing, 100049, PR Chinac Laboratory of Molecular Biology, Department of Biochemistry, Anhui Medical University, Hefei, 230032, PR Chinad State Key Laboratory of Molecular Vaccinology and Molecular Diagnostics, Xiamen University, Xiamen, 361102, PR China
a r t i c l e i n f o
Article history:Received 30 August 2020Received in revised form21 October 2020Accepted 4 November 2020Available online 6 November 2020
Keywords:DEHPEpithelial to mesenchymal transformationSmall extracellular vesiclesmicroRNA
* This paper has been recommended for acceptanc* Corresponding author. Key Lab of Urban Environm
Urban Environment, Chinese Academy of Sciences, XiE-mail address: [email protected] (Q. Huang).
https://doi.org/10.1016/j.envpol.2020.1160050269-7491/© 2020 Elsevier Ltd. All rights reserved.
a b s t r a c t
Small extracellular vesicles (sEV) are small lipid bilayer particles released by cells. sEV have been shownto play critical roles in intercellular communication. Di (2-ethylhexyl) phthalate (DEHP), widely used asplasticizers, has been detected in the environment and human beings. DEHP was found to exist in the airparticles and showed pulmonary toxicity. However, there’s little knowledge about the role of sEV inmediating the toxicity of DEHP-induced lung toxicity. We hypothesized that sEV mediated the toxicity ofDEHP through their cargo. To validate this, lung epithelial cells (A549) were exposed to various con-centrations (0, 0.2, 2 and 20 mM) of DEHP for 48 h. sEV extracted from DEHP-exposed A549 cells werecultured with unexposed A549 cells. Results showed that DEHP induced the epithelial-mesenchymaltransition (EMT) and promoted the migration and invasion ability of A549 cells. The number ofreleased sEV significantly increased in the culture media in DEHP-exposed groups compared to unex-posed groups. The sEV can enter the unexposed A549 cells and enhance its EMT and the ability ofmigration and invasion. Treatment with GW4869 in DEHP-exposed A549 cells almost blocked the effectsof DEHP-elicited sEV in normal A549 cells. Sequencing and functional analysis showed that theenrichment of significantly differentially expressed sEV miRNAs were related to tumor etiology. MiR-26a-5p was significantly enriched in DEHP-elicited sEV. Inhibition of miR-26a-5p in DEHP-exposed cells ledto the downregulation of miR-26a-5p in sEV, and thus abolished the effects of DEHP-elicited sEV innormal A549 cells, whereas overexpression of miR-26a-5p restored the effects. The transcription factorstwist is one of the downstream targets in the effects of sEV-miR-26a-5p on EMT process. In all, ourresults showed that DEHP exposure promoted the secretion of miR-26a-5p in sEV, which subsequentlyenhanced the EMT, migration and invasion ability in neighboring normal cells via the twist.
© 2020 Elsevier Ltd. All rights reserved.
Author contribution
Yifei Qin, Conceptualization, Methodology, Writing, Jing Zhang,Formal analysis, Methodology, drafting, Ricardo David Avell�an-Llaguno, drafting. Xu Zhang, Revision, Methodology, QianshengHuang, Study conception, Formal analysis, writing.
e by Dr. Sarah Harmon.ent and Health, Institute of
amen, 361021, PR China.
1. Introduction
DEHP is one type of the most common manufactured chemicalswhich is used as a plasticizer in various products like beveragebottle, food containers, medical instruments, cosmetics, andclothes. DEHP is easily eluted, leached, and evaporated from plas-tics and prevalent in the atmosphere (H. Y. Kim, 2016). Moreover,DEHP was also widely detected in the airborne fine particulatematter (PM2.5). Among PM2.5-bound phthalates in Beijing, di-n-butyl phthalate (DBP) and DEHP which accounting for 76.3%e97.7% were the most abundant (Chen et al., 2018). People wereestimated to expose to 18 ng per 8 h of DEHP through inhalingPM2.5 from Mexico in May 2013 which contained 229.7 mg DEHP

Y. Qin, J. Zhang, R.D. Avell�an-Llaguno et al. Environmental Pollution 272 (2021) 116005
per gram of particles (Quintana-Belmares et al., 2018).As inhalation is a significant exposure route for DEHP, their ef-
fect on respiratory toxicity, especially in promoting lung cancermetastasis, has received widespread attention. The estimation ofincremental lifetime cancer risks (ILCR) from DEHP inhalationexposure for children and adolescents (ages of 1e21 years) wereseveral times higher than the U.S. EPA’s acceptable limit (Pei et al.,2013).In an enroll with 559 elderly participants, high urine levels ofphthalate metabolite were closely related to the lower lung func-tion and an increased rate of decline in lung function (K. N. Kimet al., 2018). Moreover, there’s an association between phthalateexposure and asthma risk. For example, metabolites of DEHP,butylbenzyl phthalate (BBzP), and di-n-butyl phthalate (DnBP) inprenatal maternal urinary have been associated with childhoodwheeze and asthma (Gascon et al., 2015; Ku et al., 2015). In a panelresearch on 418 individuals over 60 years old in Seoul, Korea, uri-nary phthalate including MBzP, MEHHP, MECPP, MCNP, and DEHPmetabolite are associated with decreasing pulmonary function(Park et al., 2013). Furthermore, DEHP and its metabolite mono-ethylhexyl phthalate (MEHP) have the capacity to change thestructure and elevate migration ability of A549 cells (Rafael-Vazquez et al., 2018). DEHP exposure promoted the phosphoryla-tion, nuclear localization, and transcriptional activities of p65which is one of the major parts of NF-kB complex, and then stim-ulated the migration and invasion of non-small cell lung cancer(NSCLC) via NF-kB-IL-6 pathway (Y. Wang et al., 2017). During thedevelopment of these pathological process in respiratory systeminduced by DEHP, lung epithelial cells not only act as the first lineresponding to environmental stresses but also participate in therelease of diverse cytokines, chemokines, and mediators, whichpass the first signaling to inner cell layer and recruit immune cells.However, it is still unclear how the toxicity of DEHP is transferredacross the exposed lung epithelial cells to unexposed lung epithe-lial cells.
Extracellular vesicles (EVs) are small nanometre-sized mem-brane vesicles which can be secreted by a wide range of cells(Mathieu et al., 2019). According to the sizes, the InternationalSociety for Extracellular Vesicles classified the EVs into small(sEV < 200 nm) and medium (mEV> 200 nm) EVs (Thery et al.,2018). Secreted sEV with donor cell-derived bioactive molecules,such as nucleic acids, lipids, and proteins, will be endocytosed and/or phagocytosed by target cells, thus play essential roles in variouscellular processes in recipient cells (Qin et al., 2020). Among thenumerous molecular cargos, microRNAs (miRNAs) are regarded asone of the main compositions of sEV and play a vital part in thefunction of sEV.
A variety of epidemiological and toxicological evidence hasproved that sEV were involved in the toxicology of environmentalexposure. Environmental toxicants directly affect the secretion ofsEV, as well as the components of sEV cargo (Benedikter et al.,2019). The altered sEV are capable of activating or silencingdownstream signaling pathways in recipient cells and affect thefunction of these cells. Previous studies have shown that severalenvironmental toxins or carcinogens like cigarette smoking (Xuet al., 2018) and heavy metal (Gao et al., 2020) transduce theircytotoxicity and carcinogenicity in cell-to-cell way through sEV.
In all, there are still no reports on the modified sEV mediatingthe physio-pathological activity from exposed cells to unexposedcells under DEHP exposure. We presented the use of DEHP-treatedA549 derived sEV and nearby normal A549 as a co-culture systemto study the role of sEV in transferring the toxicity signal uponDEHP exposure.
2
2. Materials and methods
2.1. Cell culture and treatment
A549 cells were obtained from the Cell Bank of Type CultureCollection, Chinese Academy of Sciences (Shanghai, China) andmaintained in DME/F12 medium (HyClone, Logan, UT) supple-mented with 10% Fetal Bovine Serum (FBS, HyClone, Victoria,Australia). Cells were cultured at 37 �C in a humidified incubatorcontaining 5% CO2. After reaching 70e80% confluence, cellsmonolayers were washed twice with phosphate-buffered saline(PBS, HyClone, Victoria, Australia) and were treated with serialconcentrations of DEHP (0.2, 2 and 20 mM) which were first dis-solved in DMSO as storage solution and then added into the culturesupplemented with 10% FBS (depleted of sEV by ultracentrifugationat 120,000 g overnight). 0.1% of DMSO was set up as a vehiclecontrol group. After 48 h incubation, cells and culture media wereharvested for next step assay as indicated.
2.2. Calculation of DEHP exposure concentration
The concentrations of phthalates acid esters (PAE) in Chinaranged from 4.08 to 4790 ng/m3 (Chen et al., 2018; Li et al., 2019; B.Ma et al., 2020; J. Ma et al., 2014; Zhang et al., 2019b). The volume ofan adult breathes for 24 h is about 20 m3, and the volume of airwayand lung lining fluids is estimated to be around 12.4 ml (Sun et al.,2001). DEHP was the abundant exposure species, which contrib-uted 15.6%e19.5% of the total PAEs (Chen et al., 2018).
The concentration of DEHP exposure in the lung (CDEHP) can becalculated by equation((f91)(1):
CDEHP ¼�Cair,V1,1000
V2,M
�,20%#ð1Þ
Cair is the concentration of phthalate in the air (ng/m3), V1 is thevolume of an adult breathes for 24 h (m3), V2 is the volume of lunglining fluids (ml), M is the molecular mass of DEHP. For facilitatecalculation, the content of DEHP in PAE was regarded as 20%. Ac-cording to the equation (1), the DEHP exposure concentration inlung tissue in 24 h ranged from 0.011 mM to 4 mM. Thu, the exposureconcentration of DEHP we choose is 0.2, 2, 20 mM.
2.3. Cell proliferation analysis
A549 cell proliferation was determined using a CellTiter 96®AQueous One Solution Cell Proliferation Assay (MTS) (Promega,USA). Cells were plated in a 96-well plate and exposed to DEHP for48 h, with five biological replicates for each group. 20 mL MTS re-agent was added to each well and cultured at 37 �C for 1 h. Theabsorbance was quantified at 490 nm by a microplate spectro-photometer (Tecan, Austria). The proliferation of cells wasexpressed as the percentage of control.
2.4. Isolation of sEV
sEV were isolated and purified from the culture medium ofA549 cells using a differential centrifugation method based onprevious protocols (Lobb et al., 2015). In short, after removal oflarge dead cells (500 g, 10 min), supernatants were centrifuged(3000 g, 10 min, followed by 10,000 g, 45 min) to eliminate big celldebris. The supernatant was then filtered with an ultrafine 0.22 mmPVDF filter (Millipore), and then ultracentrifuged at 120,000 g for
Y. Qin, J. Zhang, R.D. Avell�an-Llaguno et al. Environmental Pollution 272 (2021) 116005
1.5 h at 4 �C to pellet the small vesicles. The pellet containing sEVwas washed in a large volume of PBS and centrifuged at 120,000 gfor 1.5 h to eliminate contaminating proteins. After discard super-natant, the final pellet was finally re-suspended in PBS.
2.5. Nanoparticle tracking analysis
The number and size distribution of sEV were analyzed withNanoparticle Tracking Analysis (NTA) technology with ZetaViewPMX 110 (Particle Metrix, Meerbusch, Germany) and correspondingsoftware ZetaView 8.04.02. NTA measurement was recorded andanalyzed at 11 positions. The ZetaView systemwas calibrated using110 nm polystyrene particles.
2.6. Transmission electron microscopy (TEM)
sEV pellets were fixed with 4% paraformaldehyde (Sigma) in0.1 M phosphate buffer (pH 7.4) and solution was placed on thecopper mesh. Then, they were visualized using the FEI Tecnai G2Spirit transmission electron microscope (Hitachi, JPN).
2.7. Western blot
Total proteins in A549 cells and sEV were extracted by Pierce®RIPA buffer (Thermo Scientific, IL, USA) containing protease inhib-itor (Thermo Scientific, IL, USA) and quantified by Pierce® Micro-plate BCA Protein Assay Kit (Thermo Scientific, IL, USA). Equalamounts of protein (20e50 mg) were loaded for 10% SDS-PAGEelectrophoresis for western blot analysis according to the stan-dard protocols.
The manufactures and catalogue numbers of antibodies used in
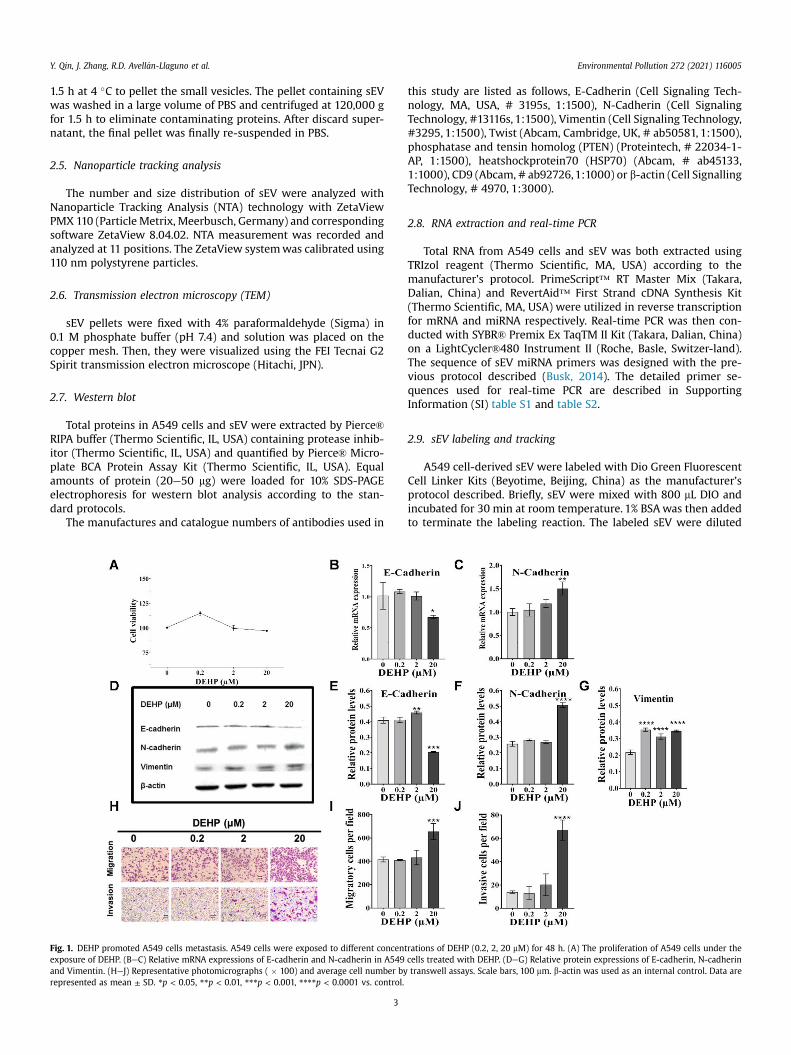
Fig. 1. DEHP promoted A549 cells metastasis. A549 cells were exposed to different concentexposure of DEHP. (BeC) Relative mRNA expressions of E-cadherin and N-cadherin in A549and Vimentin. (HeJ) Representative photomicrographs ( � 100) and average cell number byrepresented as mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 vs. control.
3
this study are listed as follows, E-Cadherin (Cell Signaling Tech-nology, MA, USA, # 3195s, 1:1500), N-Cadherin (Cell SignalingTechnology, #13116s, 1:1500), Vimentin (Cell Signaling Technology,#3295, 1:1500), Twist (Abcam, Cambridge, UK, # ab50581, 1:1500),phosphatase and tensin homolog (PTEN) (Proteintech, # 22034-1-AP, 1:1500), heatshockprotein70 (HSP70) (Abcam, # ab45133,1:1000), CD9 (Abcam, # ab92726,1:1000) or b-actin (Cell SignallingTechnology, # 4970, 1:3000).
2.8. RNA extraction and real-time PCR
Total RNA from A549 cells and sEV was both extracted usingTRIzol reagent (Thermo Scientific, MA, USA) according to themanufacturer’s protocol. PrimeScript™ RT Master Mix (Takara,Dalian, China) and RevertAid™ First Strand cDNA Synthesis Kit(Thermo Scientific, MA, USA) were utilized in reverse transcriptionfor mRNA and miRNA respectively. Real-time PCR was then con-ducted with SYBR® Premix Ex TaqTM II Kit (Takara, Dalian, China)on a LightCycler®480 Instrument II (Roche, Basle, Switzer-land).The sequence of sEV miRNA primers was designed with the pre-vious protocol described (Busk, 2014). The detailed primer se-quences used for real-time PCR are described in SupportingInformation (SI) table S1 and table S2.
2.9. sEV labeling and tracking
A549 cell-derived sEV were labeled with Dio Green FluorescentCell Linker Kits (Beyotime, Beijing, China) as the manufacturer’sprotocol described. Briefly, sEV were mixed with 800 mL DIO andincubated for 30 min at room temperature. 1% BSAwas then addedto terminate the labeling reaction. The labeled sEV were diluted
rations of DEHP (0.2, 2, 20 mM) for 48 h. (A) The proliferation of A549 cells under thecells treated with DEHP. (DeG) Relative protein expressions of E-cadherin, N-cadherintranswell assays. Scale bars, 100 mm. b-actin was used as an internal control. Data are
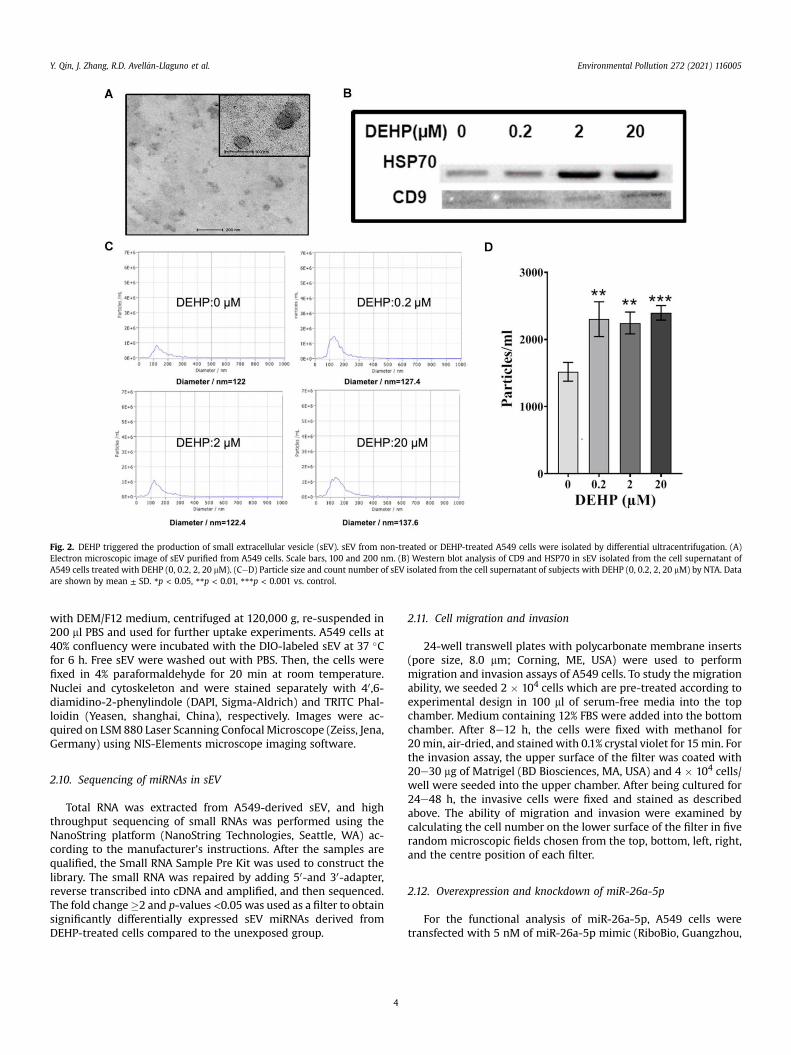
Fig. 2. DEHP triggered the production of small extracellular vesicle (sEV). sEV from non-treated or DEHP-treated A549 cells were isolated by differential ultracentrifugation. (A)Electron microscopic image of sEV purified from A549 cells. Scale bars, 100 and 200 nm. (B) Western blot analysis of CD9 and HSP70 in sEV isolated from the cell supernatant ofA549 cells treated with DEHP (0, 0.2, 2, 20 mM). (CeD) Particle size and count number of sEV isolated from the cell supernatant of subjects with DEHP (0, 0.2, 2, 20 mM) by NTA. Dataare shown by mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001 vs. control.
Y. Qin, J. Zhang, R.D. Avell�an-Llaguno et al. Environmental Pollution 272 (2021) 116005
with DEM/F12 medium, centrifuged at 120,000 g, re-suspended in200 ml PBS and used for further uptake experiments. A549 cells at40% confluency were incubated with the DIO-labeled sEV at 37 �Cfor 6 h. Free sEV were washed out with PBS. Then, the cells werefixed in 4% paraformaldehyde for 20 min at room temperature.Nuclei and cytoskeleton and were stained separately with 40,6-diamidino-2-phenylindole (DAPI, Sigma-Aldrich) and TRITC Phal-loidin (Yeasen, shanghai, China), respectively. Images were ac-quired on LSM 880 Laser Scanning Confocal Microscope (Zeiss, Jena,Germany) using NIS-Elements microscope imaging software.
2.10. Sequencing of miRNAs in sEV
Total RNA was extracted from A549-derived sEV, and highthroughput sequencing of small RNAs was performed using theNanoString platform (NanoString Technologies, Seattle, WA) ac-cording to the manufacturer’s instructions. After the samples arequalified, the Small RNA Sample Pre Kit was used to construct thelibrary. The small RNA was repaired by adding 50-and 30-adapter,reverse transcribed into cDNA and amplified, and then sequenced.The fold change�2 and p-values <0.05 was used as a filter to obtainsignificantly differentially expressed sEV miRNAs derived fromDEHP-treated cells compared to the unexposed group.
4
2.11. Cell migration and invasion
24-well transwell plates with polycarbonate membrane inserts(pore size, 8.0 mm; Corning, ME, USA) were used to performmigration and invasion assays of A549 cells. To study the migrationability, we seeded 2 � 104 cells which are pre-treated according toexperimental design in 100 ml of serum-free media into the topchamber. Medium containing 12% FBS were added into the bottomchamber. After 8e12 h, the cells were fixed with methanol for20min, air-dried, and stainedwith 0.1% crystal violet for 15min. Forthe invasion assay, the upper surface of the filter was coated with20e30 mg of Matrigel (BD Biosciences, MA, USA) and 4 � 104 cells/well were seeded into the upper chamber. After being cultured for24e48 h, the invasive cells were fixed and stained as describedabove. The ability of migration and invasion were examined bycalculating the cell number on the lower surface of the filter in fiverandom microscopic fields chosen from the top, bottom, left, right,and the centre position of each filter.
2.12. Overexpression and knockdown of miR-26a-5p
For the functional analysis of miR-26a-5p, A549 cells weretransfected with 5 nM of miR-26a-5p mimic (RiboBio, Guangzhou,
Y. Qin, J. Zhang, R.D. Avell�an-Llaguno et al. Environmental Pollution 272 (2021) 116005
China) or miR-26a-5p inhibitor with LipofectamineTM 3000Transfection Reagent (Thermo Scientific, MA, USA) following themanufacturer’s instructions for 48 h.
2.13. Inhibition of the secretion of sEV
A549 cells were exposed to DEHP for 48 h, and 10 mM GW4869(Sigma-Aldrich) or 0.1% dimethyl sulfoxide (vehicle) was added 3 hbefore the end of the exposure period. The culture mediawere thencollected, and sEV were isolated using a differential centrifugationmethod.
2.14. Statistical analyses
All statistical analyses were performed by GraphPad 7.0 soft-ware. The results are presented as mean ± standard deviation (SD).After verifying the normality and equality, the differences amongtwo groups and multiple groups were conducted with unpairedStudent’s t-test and one-way analysis of variance (ANOVA) followedby Bonferroni’s post hoc test, respectively. Individual data that does
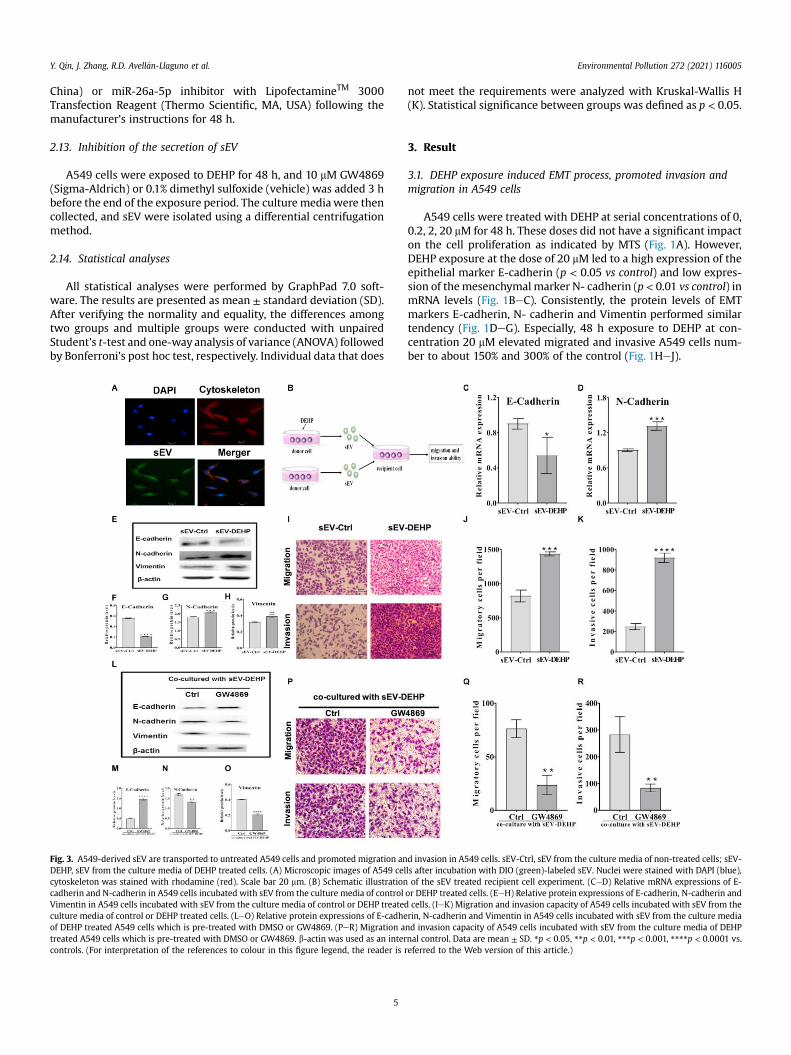
Fig. 3. A549-derived sEV are transported to untreated A549 cells and promoted migration anDEHP, sEV from the culture media of DEHP treated cells. (A) Microscopic images of A549 celcytoskeleton was stained with rhodamine (red). Scale bar 20 mm. (B) Schematic illustrationcadherin and N-cadherin in A549 cells incubated with sEV from the culture media of controlVimentin in A549 cells incubated with sEV from the culture media of control or DEHP treatedculture media of control or DEHP treated cells. (LeO) Relative protein expressions of E-cadheof DEHP treated A549 cells which is pre-treated with DMSO or GW4869. (PeR) Migration atreated A549 cells which is pre-treated with DMSO or GW4869. b-actin was used as an intercontrols. (For interpretation of the references to colour in this figure legend, the reader is r
5
not meet the requirements were analyzed with Kruskal-Wallis H(K). Statistical significance between groups was defined as p < 0.05.
3. Result
3.1. DEHP exposure induced EMT process, promoted invasion andmigration in A549 cells
A549 cells were treated with DEHP at serial concentrations of 0,0.2, 2, 20 mM for 48 h. These doses did not have a significant impacton the cell proliferation as indicated by MTS (Fig. 1A). However,DEHP exposure at the dose of 20 mM led to a high expression of theepithelial marker E-cadherin (p < 0.05 vs control) and low expres-sion of themesenchymal marker N- cadherin (p < 0.01 vs control) inmRNA levels (Fig. 1BeC). Consistently, the protein levels of EMTmarkers E-cadherin, N- cadherin and Vimentin performed similartendency (Fig. 1DeG). Especially, 48 h exposure to DEHP at con-centration 20 mM elevated migrated and invasive A549 cells num-ber to about 150% and 300% of the control (Fig. 1HeJ).
d invasion in A549 cells. sEV-Ctrl, sEV from the culture media of non-treated cells; sEV-ls after incubation with DIO (green)-labeled sEV. Nuclei were stained with DAPI (blue),of the sEV treated recipient cell experiment. (CeD) Relative mRNA expressions of E-
or DEHP treated cells. (EeH) Relative protein expressions of E-cadherin, N-cadherin andcells. (IeK) Migration and invasion capacity of A549 cells incubated with sEV from therin, N-cadherin and Vimentin in A549 cells incubated with sEV from the culture mediand invasion capacity of A549 cells incubated with sEV from the culture media of DEHPnal control. Data are mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 vs.eferred to the Web version of this article.)
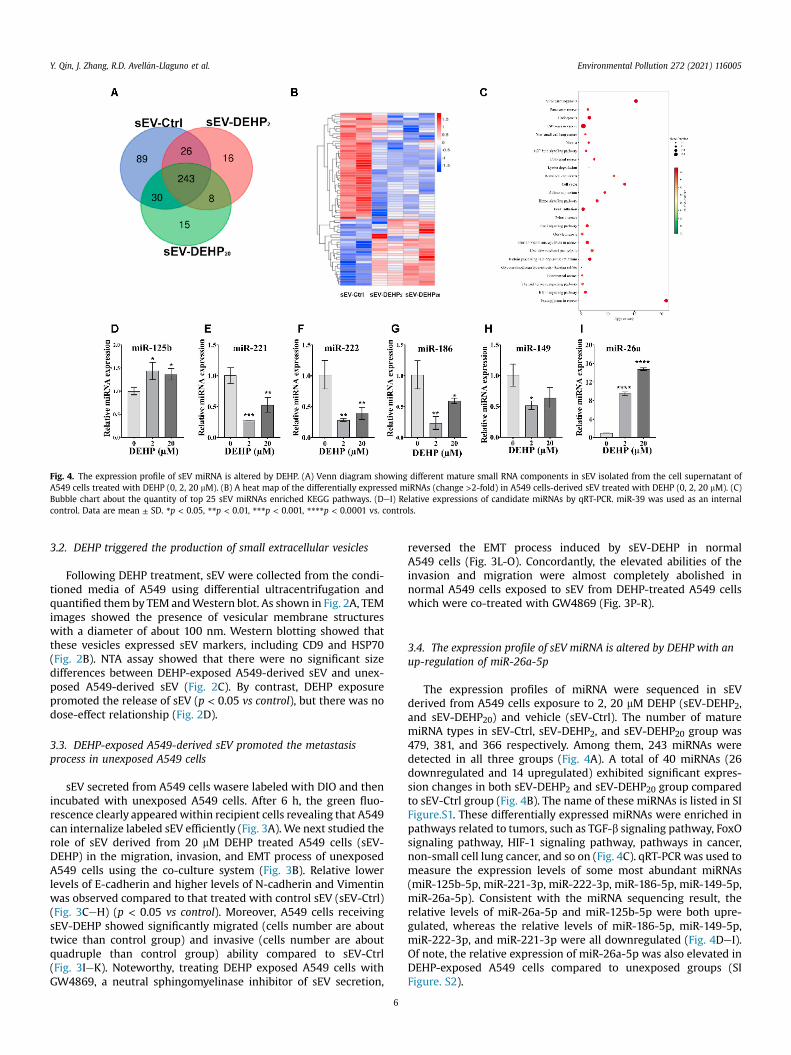
Fig. 4. The expression profile of sEV miRNA is altered by DEHP. (A) Venn diagram showing different mature small RNA components in sEV isolated from the cell supernatant ofA549 cells treated with DEHP (0, 2, 20 mM). (B) A heat map of the differentially expressed miRNAs (change >2-fold) in A549 cells-derived sEV treated with DEHP (0, 2, 20 mM). (C)Bubble chart about the quantity of top 25 sEV miRNAs enriched KEGG pathways. (DeI) Relative expressions of candidate miRNAs by qRT-PCR. miR-39 was used as an internalcontrol. Data are mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 vs. controls.
Y. Qin, J. Zhang, R.D. Avell�an-Llaguno et al. Environmental Pollution 272 (2021) 116005
3.2. DEHP triggered the production of small extracellular vesicles
Following DEHP treatment, sEV were collected from the condi-tioned media of A549 using differential ultracentrifugation andquantified them by TEM andWestern blot. As shown in Fig. 2A, TEMimages showed the presence of vesicular membrane structureswith a diameter of about 100 nm. Western blotting showed thatthese vesicles expressed sEV markers, including CD9 and HSP70(Fig. 2B). NTA assay showed that there were no significant sizedifferences between DEHP-exposed A549-derived sEV and unex-posed A549-derived sEV (Fig. 2C). By contrast, DEHP exposurepromoted the release of sEV (p < 0.05 vs control), but there was nodose-effect relationship (Fig. 2D).
3.3. DEHP-exposed A549-derived sEV promoted the metastasisprocess in unexposed A549 cells
sEV secreted from A549 cells wasere labeled with DIO and thenincubated with unexposed A549 cells. After 6 h, the green fluo-rescence clearly appearedwithin recipient cells revealing that A549can internalize labeled sEV efficiently (Fig. 3A). We next studied therole of sEV derived from 20 mM DEHP treated A549 cells (sEV-DEHP) in the migration, invasion, and EMT process of unexposedA549 cells using the co-culture system (Fig. 3B). Relative lowerlevels of E-cadherin and higher levels of N-cadherin and Vimentinwas observed compared to that treated with control sEV (sEV-Ctrl)(Fig. 3CeH) (p < 0.05 vs control). Moreover, A549 cells receivingsEV-DEHP showed significantly migrated (cells number are abouttwice than control group) and invasive (cells number are aboutquadruple than control group) ability compared to sEV-Ctrl(Fig. 3IeK). Noteworthy, treating DEHP exposed A549 cells withGW4869, a neutral sphingomyelinase inhibitor of sEV secretion,
6
reversed the EMT process induced by sEV-DEHP in normalA549 cells (Fig. 3L-O). Concordantly, the elevated abilities of theinvasion and migration were almost completely abolished innormal A549 cells exposed to sEV from DEHP-treated A549 cellswhich were co-treated with GW4869 (Fig. 3P-R).
3.4. The expression profile of sEV miRNA is altered by DEHP with anup-regulation of miR-26a-5p
The expression profiles of miRNA were sequenced in sEVderived from A549 cells exposure to 2, 20 mM DEHP (sEV-DEHP2,and sEV-DEHP20) and vehicle (sEV-Ctrl). The number of maturemiRNA types in sEV-Ctrl, sEV-DEHP2, and sEV-DEHP20 group was479, 381, and 366 respectively. Among them, 243 miRNAs weredetected in all three groups (Fig. 4A). A total of 40 miRNAs (26downregulated and 14 upregulated) exhibited significant expres-sion changes in both sEV-DEHP2 and sEV-DEHP20 group comparedto sEV-Ctrl group (Fig. 4B). The name of these miRNAs is listed in SIFigure.S1. These differentially expressed miRNAs were enriched inpathways related to tumors, such as TGF-b signaling pathway, FoxOsignaling pathway, HIF-1 signaling pathway, pathways in cancer,non-small cell lung cancer, and so on (Fig. 4C). qRT-PCR was used tomeasure the expression levels of some most abundant miRNAs(miR-125b-5p, miR-221-3p, miR-222-3p, miR-186-5p, miR-149-5p,miR-26a-5p). Consistent with the miRNA sequencing result, therelative levels of miR-26a-5p and miR-125b-5p were both upre-gulated, whereas the relative levels of miR-186-5p, miR-149-5p,miR-222-3p, and miR-221-3p were all downregulated (Fig. 4DeI).Of note, the relative expression of miR-26a-5p was also elevated inDEHP-exposed A549 cells compared to unexposed groups (SIFigure. S2).
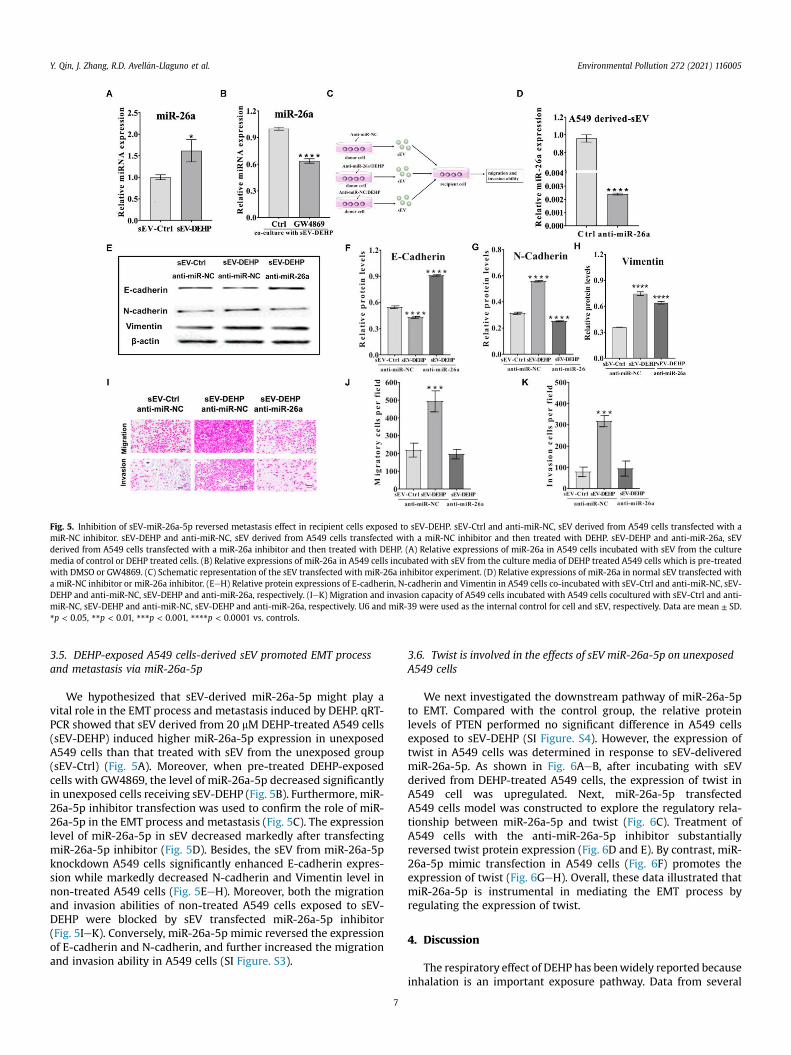
Fig. 5. Inhibition of sEV-miR-26a-5p reversed metastasis effect in recipient cells exposed to sEV-DEHP. sEV-Ctrl and anti-miR-NC, sEV derived from A549 cells transfected with amiR-NC inhibitor. sEV-DEHP and anti-miR-NC, sEV derived from A549 cells transfected with a miR-NC inhibitor and then treated with DEHP. sEV-DEHP and anti-miR-26a, sEVderived from A549 cells transfected with a miR-26a inhibitor and then treated with DEHP. (A) Relative expressions of miR-26a in A549 cells incubated with sEV from the culturemedia of control or DEHP treated cells. (B) Relative expressions of miR-26a in A549 cells incubated with sEV from the culture media of DEHP treated A549 cells which is pre-treatedwith DMSO or GW4869. (C) Schematic representation of the sEV transfected with miR-26a inhibitor experiment. (D) Relative expressions of miR-26a in normal sEV transfected witha miR-NC inhibitor or miR-26a inhibitor. (EeH) Relative protein expressions of E-cadherin, N-cadherin and Vimentin in A549 cells co-incubated with sEV-Ctrl and anti-miR-NC, sEV-DEHP and anti-miR-NC, sEV-DEHP and anti-miR-26a, respectively. (IeK) Migration and invasion capacity of A549 cells incubated with A549 cells cocultured with sEV-Ctrl and anti-miR-NC, sEV-DEHP and anti-miR-NC, sEV-DEHP and anti-miR-26a, respectively. U6 and miR-39 were used as the internal control for cell and sEV, respectively. Data are mean ± SD.*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 vs. controls.
Y. Qin, J. Zhang, R.D. Avell�an-Llaguno et al. Environmental Pollution 272 (2021) 116005
3.5. DEHP-exposed A549 cells-derived sEV promoted EMT processand metastasis via miR-26a-5p
We hypothesized that sEV-derived miR-26a-5p might play avital role in the EMT process and metastasis induced by DEHP. qRT-PCR showed that sEV derived from 20 mM DEHP-treated A549 cells(sEV-DEHP) induced higher miR-26a-5p expression in unexposedA549 cells than that treated with sEV from the unexposed group(sEV-Ctrl) (Fig. 5A). Moreover, when pre-treated DEHP-exposedcells with GW4869, the level of miR-26a-5p decreased significantlyin unexposed cells receiving sEV-DEHP (Fig. 5B). Furthermore, miR-26a-5p inhibitor transfection was used to confirm the role of miR-26a-5p in the EMT process and metastasis (Fig. 5C). The expressionlevel of miR-26a-5p in sEV decreased markedly after transfectingmiR-26a-5p inhibitor (Fig. 5D). Besides, the sEV from miR-26a-5pknockdown A549 cells significantly enhanced E-cadherin expres-sion while markedly decreased N-cadherin and Vimentin level innon-treated A549 cells (Fig. 5EeH). Moreover, both the migrationand invasion abilities of non-treated A549 cells exposed to sEV-DEHP were blocked by sEV transfected miR-26a-5p inhibitor(Fig. 5IeK). Conversely, miR-26a-5p mimic reversed the expressionof E-cadherin and N-cadherin, and further increased the migrationand invasion ability in A549 cells (SI Figure. S3).
7
3.6. Twist is involved in the effects of sEV miR-26a-5p on unexposedA549 cells
We next investigated the downstream pathway of miR-26a-5pto EMT. Compared with the control group, the relative proteinlevels of PTEN performed no significant difference in A549 cellsexposed to sEV-DEHP (SI Figure. S4). However, the expression oftwist in A549 cells was determined in response to sEV-deliveredmiR-26a-5p. As shown in Fig. 6AeB, after incubating with sEVderived from DEHP-treated A549 cells, the expression of twist inA549 cell was upregulated. Next, miR-26a-5p transfectedA549 cells model was constructed to explore the regulatory rela-tionship between miR-26a-5p and twist (Fig. 6C). Treatment ofA549 cells with the anti-miR-26a-5p inhibitor substantiallyreversed twist protein expression (Fig. 6D and E). By contrast, miR-26a-5p mimic transfection in A549 cells (Fig. 6F) promotes theexpression of twist (Fig. 6GeH). Overall, these data illustrated thatmiR-26a-5p is instrumental in mediating the EMT process byregulating the expression of twist.
4. Discussion
The respiratory effect of DEHP has beenwidely reported becauseinhalation is an important exposure pathway. Data from several
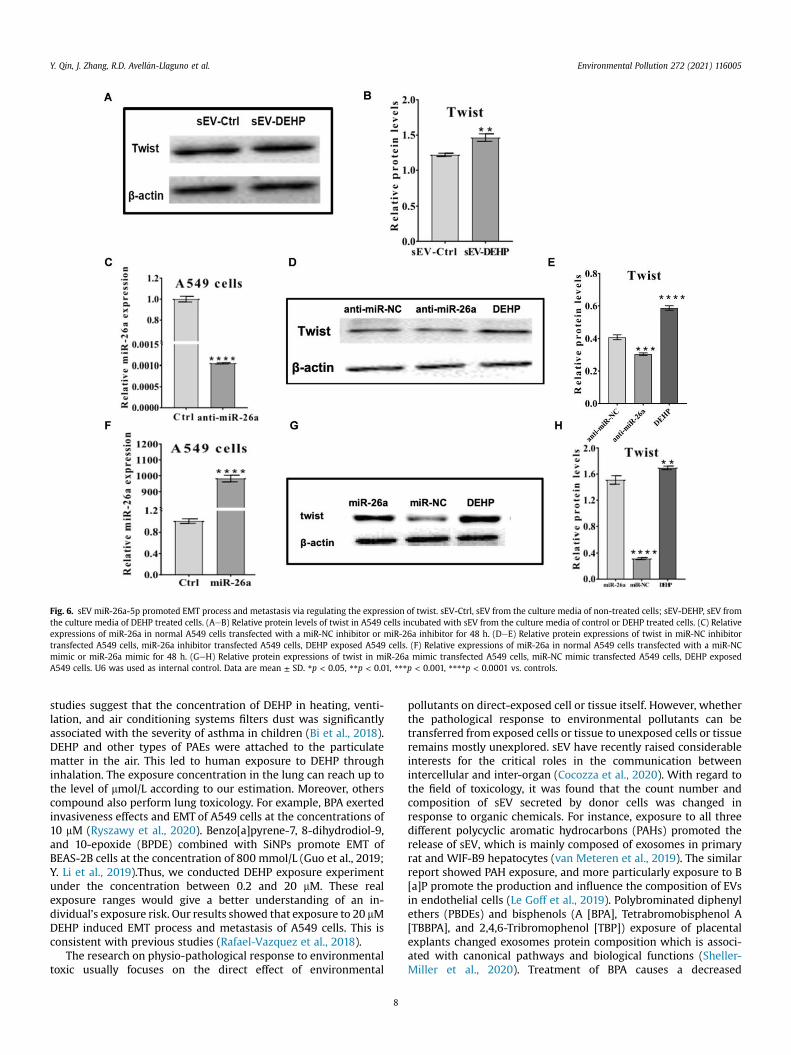
Fig. 6. sEV miR-26a-5p promoted EMT process and metastasis via regulating the expression of twist. sEV-Ctrl, sEV from the culture media of non-treated cells; sEV-DEHP, sEV fromthe culture media of DEHP treated cells. (AeB) Relative protein levels of twist in A549 cells incubated with sEV from the culture media of control or DEHP treated cells. (C) Relativeexpressions of miR-26a in normal A549 cells transfected with a miR-NC inhibitor or miR-26a inhibitor for 48 h. (DeE) Relative protein expressions of twist in miR-NC inhibitortransfected A549 cells, miR-26a inhibitor transfected A549 cells, DEHP exposed A549 cells. (F) Relative expressions of miR-26a in normal A549 cells transfected with a miR-NCmimic or miR-26a mimic for 48 h. (GeH) Relative protein expressions of twist in miR-26a mimic transfected A549 cells, miR-NC mimic transfected A549 cells, DEHP exposedA549 cells. U6 was used as internal control. Data are mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 vs. controls.
Y. Qin, J. Zhang, R.D. Avell�an-Llaguno et al. Environmental Pollution 272 (2021) 116005
studies suggest that the concentration of DEHP in heating, venti-lation, and air conditioning systems filters dust was significantlyassociated with the severity of asthma in children (Bi et al., 2018).DEHP and other types of PAEs were attached to the particulatematter in the air. This led to human exposure to DEHP throughinhalation. The exposure concentration in the lung can reach up tothe level of mmol/L according to our estimation. Moreover, otherscompound also perform lung toxicology. For example, BPA exertedinvasiveness effects and EMT of A549 cells at the concentrations of10 mM (Ryszawy et al., 2020). Benzo[a]pyrene-7, 8-dihydrodiol-9,and 10-epoxide (BPDE) combined with SiNPs promote EMT ofBEAS-2B cells at the concentration of 800 mmol/L (Guo et al., 2019;Y. Li et al., 2019).Thus, we conducted DEHP exposure experimentunder the concentration between 0.2 and 20 mM. These realexposure ranges would give a better understanding of an in-dividual’s exposure risk. Our results showed that exposure to 20 mMDEHP induced EMT process and metastasis of A549 cells. This isconsistent with previous studies (Rafael-Vazquez et al., 2018).
The research on physio-pathological response to environmentaltoxic usually focuses on the direct effect of environmental
8
pollutants on direct-exposed cell or tissue itself. However, whetherthe pathological response to environmental pollutants can betransferred from exposed cells or tissue to unexposed cells or tissueremains mostly unexplored. sEV have recently raised considerableinterests for the critical roles in the communication betweenintercellular and inter-organ (Cocozza et al., 2020). With regard tothe field of toxicology, it was found that the count number andcomposition of sEV secreted by donor cells was changed inresponse to organic chemicals. For instance, exposure to all threedifferent polycyclic aromatic hydrocarbons (PAHs) promoted therelease of sEV, which is mainly composed of exosomes in primaryrat and WIF-B9 hepatocytes (van Meteren et al., 2019). The similarreport showed PAH exposure, and more particularly exposure to B[a]P promote the production and influence the composition of EVsin endothelial cells (Le Goff et al., 2019). Polybrominated diphenylethers (PBDEs) and bisphenols (A [BPA], Tetrabromobisphenol A[TBBPA], and 2,4,6-Tribromophenol [TBP]) exposure of placentalexplants changed exosomes protein composition which is associ-ated with canonical pathways and biological functions (Sheller-Miller et al., 2020). Treatment of BPA causes a decreased

Y. Qin, J. Zhang, R.D. Avell�an-Llaguno et al. Environmental Pollution 272 (2021) 116005
expression of granulosa cells derived EV-miRNA-27b-3p, andincreased levels of relevant genes (Rodosthenous et al., 2019).Toluene exposure resulted in dysregulation of exosomal miRNAs ofHL-60 cell, which are related to various disease (Lim et al., 2017). Inline with previous research, our results showed DEHP treatmenttriggered the generation of A549 cells-derived sEV compared totheir respective control groups. Moreover, from miRNA profilinganalysis, we found that both the abundance of mature miRNA andthe expression levels of differentially expressed miRNA havechanged significantly in sEV-Ctrl, sEV-DEHP2, and sEV-DEHP20groups, suggesting a pathological role for sEV in lung epithelialcells. However, whether the aberrant expression of EVs cargomediated the toxicity of these organic chemicals is still not yetconfirmed.
To investigate the role of lung epithelial cell-derived sEVinvolved in the paracrine actions of nearby normal lung epithelialcells, we firstly examined whether A549-derived sEV can beengulfed by neighboring cells. Treating A549 cells with DIO-labeledsEV caused significantly green fluorescence accumulation withincells, demonstrating A549 cells-derived sEV can be absorbed byrecipient cells efficiently. Based on the sEV uptake experiments, weassumed that sEV modified by DEHP could transfer the damagebetween the exposed cell and the nearby normal cell. Second, in thecontext of sEV-DEHP exposure, accelerating of EMT process, as wellas promoting the invasion and migration ability were performed inA549 cells. Finally, once sEV release was blocked by GW4869 in theco-culture system, these effects were profoundly diminished. Theseresults illustrated that the DEHP-treated A549 cells are primed todeliver signals molecular into healthy A549 cells, and furthertransfer effect via sEV release.
Among various active components within sEV, miRNA is one ofthe main compositions of sEV and has excellent potential as lungcancer therapeutics and biomarkers (Zhang et al., 2019a). Our datashowed that a set of miRNAs was obviously changed in DEHP-treated A549 cells derived sEV, in comparison with control cells.Specifically, significant overexpression of miR-26a-5p was identi-fied in the DEHP-A549-derived sEV. Interestingly, the expression ofmiR-26a-5p also increased in the DEHP-exposed A549 cells, sug-gesting miR-26a-5p was mainly attributed to the metastasisinduced by DEHP directly or indirectly. Moreover, inhibition of sEVmiR-26a-5p blocked the changes of migration and invasion ability,and EMT process in normal A549 cells caused by sEV derived fromDEHP-treated A549 cells, but upregulation of sEV miR-26a-5prestored these effects. Also, some studies have shown that over-expression of miR-26a-5p exerted oncogenic functions via regu-lating as a series of target genes in different types of cancers. Forexample, PTEN is the first recognized human tumor suppressorgene with phosphatase activity (Zeitels et al., 2014). In our study,sEV derived from DEHP exposed A549 cells did not significantlyaffect the protein levels of PTEN. Twist has been known as a tran-scription factor which plays an essential role in the occurrence ofEMT, and thereby influenced the invasion and metastasis of tumorcells (Yang et al., 2004). In this study, treatment with miR-26a-5pinhibitor downregulated twist expression, and further reducedthe migration and invasion of A549 cells.
sEV not only serves as a carrier of signaling molecules tomediate cell-to-cell communication but also can be used as a nat-ural nanocarrier for substance delivery. After exposure to 20 nmgold nanoparticles (AuNPs) in human primary macrophages, re-searchers found AuNPs are uptaken by macrophages and then canbe released into the extracellular environment via sEV (Logozziet al., 2019). Only recently, scientists designed sEV derived frommacrophage RAW264.7 cells as a curcumin carrier which featuredhighly effective blood-brain barrier-crossing and preventedneuronal death in vitro. Studies have demonstrated that
9
encapsulation into the sEV increased curcumin solubility from 1.8to 18.5 mg mL�1 and evaluated curcumin stability (H. Wang et al.,2019). These studies raised the assumption that DEHP might beincorporated, or even enriched into sEV, thus transferred acrosscells, and finally led to toxicity. The possibility increased as DEHP isa lipophilic chemical. In our companion study, rats were gavage-administrated with 50 and 500 mg/kg of DEHP. After 4 h, theconcentrations of DEHPwere detected in serum and serum-derivedsEV. Results showed that DEHP was not enriched in sEV (data notshown). Additionally, knockdown of miR-26a-5p diminished themetastasis effects of sEV derived from DEHP-exposed A549 cells.We believed that sEV mediated the toxicity by transferring itsbiological contents rather than external chemicals in this study.
There are several limitation in the study. First, in addition tomiR-26a-5p, there could exist other miRNAs or even other mole-cules that can also contribute to the mediating roles of sEV in thetoxicity of DEHP. Second, Twist is not the direct target gene of miR-26a-5p. The downstream pathway of miR-26a-5p still needs to beinvestigated.
5. Conclusion
This study demonstrated that sEV played a vital role in trans-ferring the promoting effects on EMT, migration and invasion fromDEHP-exposed A549 cells to unexposed A549 cells. MiR-26a-5pfrom DEHP-exposed cells-derived sEV regulated the expression oftwist, and then activate the EMT pathway in recipient cells. DEHPcan induce pulmonary toxicity by affecting the intercellularcommunication within cells.
Declaration of competing interest
The authors declare that they have no known competingfinancial interests or personal relationships that could haveappeared to influence the work reported in this paper.
Acknowledges
This work was supported by the National Key R&D Program ofChina (2018YFE0103300), the Youth Innovation Promotion Associ-ation CAS (2018343), Xiamen Municipal Bureau of Science andTechnology Program (3502Z20203079), the International Partner-ship Program of Chinese Academy of Sciences(132C35KYSB20180006), the Open Research Fund of State KeyLaboratory of Molecular Vaccinology and Molecular Diagnostics(SKLVD2020KF04).
Appendix A. Supplementary data
Supplementary data to this article can be found online athttps://doi.org/10.1016/j.envpol.2020.116005.
References
Benedikter, B.J., Bouwman, F.G., Heinzmann, A.C.A., et al., 2019. Proteomic analysisreveals procoagulant properties of cigarette smoke-induced extracellular vesi-cles. J. Extracell. Vesicles 8 (1), 1585163. https://doi.org/10.1080/20013078.2019.1585163.
Bi, C., Maestre, J.P., Li, H., et al., 2018. Phthalates and organophosphates in settleddust and HVAC filter dust of U.S. low-income homes: association with season,building characteristics, and childhood asthma. Environ. Int. 121 (Pt 1),916e930. https://doi.org/10.1016/j.envint.2018.09.013.
Busk, P.K., 2014. A tool for design of primers for microRNA-specific quantitative RT-qPCR. BMC Bioinf. 15 (1), 29. https://doi.org/10.1186/1471-2105-15-29.
Chen, Y., Lv, D., Li, X., et al., 2018. PM2.5-bound phthalates in indoor and outdoor airin Beijing: seasonal distributions and human exposure via inhalation. Environ.Pollut. 241, 369e377. https://doi.org/10.1016/j.envpol.2018.05.081.
Cocozza, F., Grisard, E., Martin-Jaular, L., et al., 2020. SnapShot: extracellular

Y. Qin, J. Zhang, R.D. Avell�an-Llaguno et al. Environmental Pollution 272 (2021) 116005
vesicles, 262 e261 Cell 182 (1), 262. https://doi.org/10.1016/j.cell.2020.04.054.Gao, M., Dong, Z., Sun, J., et al., 2020. Liver-derived exosome-laden lncRNA MT1DP
aggravates cadmium-induced nephrotoxicity. Environ. Pollut. 258, 113717.https://doi.org/10.1016/j.envpol.2019.113717.
Gascon, M., Casas, M., Morales, E., et al., 2015. Prenatal exposure to bisphenol A andphthalates and childhood respiratory tract infections and allergy. J. Allergy Clin.Immunol. 135 (2), 370e378. https://doi.org/10.1016/j.jaci.2014.09.030.
Kim, H.Y., 2016. Risk assessment of di(2-ethylhexyl) phthalate in the workplace.Environ Health Toxicol 31, e2016011. https://doi.org/10.5620/eht.e2016011.
Kim, K.N., Lee, M.R., Choi, Y.H., et al., 2018. Association between phthalate exposureand lower lung function in an urban elderly population: a repeated-measureslongitudinal study. Environ. Int. 113, 177e183. https://doi.org/10.1016/j.envint.2018.02.004.
Ku, H.Y., Su, P.H., Wen, H.J., et al., 2015. Prenatal and postnatal exposure to phthalateesters and asthma: a 9-year follow-up study of a taiwanese birth cohort. PloSOne 10 (4), e0123309. https://doi.org/10.1371/journal.pone.0123309.
Le Goff, M., Lagadic-Gossmann, D., Latour, R., et al., 2019. PAHs increase the pro-duction of extracellular vesicles both in vitro in endothelial cells and in vivo inurines from rats. Environ. Pollut. 255 (Pt 1), 113171. https://doi.org/10.1016/j.envpol.2019.113171.
Li, P.H., Jia, H.Y., Wang, Y., et al., 2019. Characterization of PM2.5-bound phthalic acidesters (PAEs) at regional background site in northern China: long-rangetransport and risk assessment. Sci. Total Environ. 659, 140e149. https://doi.org/10.1016/j.scitotenv.2018.12.246.
Lim, J.H., Song, M.K., Cho, Y., et al., 2017. Comparative analysis of microRNA andmRNA expression profiles in cells and exosomes under toluene exposure.Toxicol. Vitro 41, 92e101. https://doi.org/10.1016/j.tiv.2017.02.020.
Lobb, R.J., Becker, M., Wen, S.W., et al., 2015. Optimized exosome isolation protocolfor cell culture supernatant and human plasma. J. Extracell. Vesicles 4, 27031.https://doi.org/10.3402/jev.v4.27031.
Logozzi, M., Mizzoni, D., Bocca, B., et al., 2019. Human primary macrophagesscavenge AuNPs and eliminate it through exosomes. A natural shuttling fornanomaterials. Eur. J. Pharm. Biopharm. 137, 23e36. https://doi.org/10.1016/j.ejpb.2019.02.014.
Ma, B., Wang, L., Tao, W., et al., 2020. Phthalate esters in atmospheric PM2.5 andPM10 in the semi-arid city of Xi’an, Northwest China: pollution characteristics,sources, health risks, and relationships with meteorological factors. Chemo-sphere 242, 125226. https://doi.org/10.1016/j.chemosphere.2019.125226.
Ma, J., Chen, L.L., Guo, Y., et al., 2014. Phthalate diesters in airborne PM(2.5) andPM(10) in a suburban area of Shanghai: seasonal distribution and risk assess-ment. Sci. Total Environ. 497e498, 467e474. https://doi.org/10.1016/j.scitotenv.2014.08.012.
Mathieu, M., Martin-Jaular, L., Lavieu, G., et al., 2019. Specificities of secretion anduptake of exosomes and other extracellular vesicles for cell-to-cell communi-cation. Nat. Cell Biol. 21 (1), 9e17. https://doi.org/10.1038/s41556-018-0250-9.
Park, H.Y., Kim, J.H., Lim, Y.H., et al., 2013. Influence of genetic polymorphisms onthe association between phthalate exposure and pulmonary function in theelderly. Environ. Res. 122, 18e24. https://doi.org/10.1016/j.envres.2012.11.004.
Pei, X.Q., Song, M., Guo, M., et al., 2013. Concentration and risk assessment ofphthalates present in indoor air from newly decorated apartments. Atmos.Environ. 68, 17e23. https://doi.org/10.1016/j.atmosenv.2012.11.039.
Qin, Y.F., Long, L., Huang, Q.S., 2020. Extracellular vesicles in toxicological studies:key roles in communication between environmental stress and adverse out-comes. J. Appl. Toxicol. 40, 9. https://doi.org/10.1002/jat.3963.
Quintana-Belmares, R.O., Krais, A.M., Esfahani, B.K., et al., 2018. Phthalate esters onurban airborne particles: levels in PM10 and PM2.5 from Mexico City and
10
theoretical assessment of lung exposure. Environ. Res. 161, 439e445. https://doi.org/10.1016/j.envres.2017.11.039.
Rafael-Vazquez, L., Garcia-Trejo, S., Aztatzi-Aguilar, O.G., et al., 2018. Exposure todiethylhexyl phthalate (DEHP) and monoethylhexyl phthalate (MEHP) pro-motes the loss of alveolar epithelial phenotype of A549 cells. Toxicol. Lett. 294,135e144. https://doi.org/10.1016/j.toxlet.2018.05.012.
Rodosthenous, R.S., Baccarelli, A.A., Mansour, A., et al., 2019. Supraphysiologicalconcentrations of bisphenol A alter the expression of extracellular vesicle-enriched miRNAs from human primary granulosa cells. Toxicol. Sci. 169 (1),5e13. https://doi.org/10.1093/toxsci/kfz020.
Ryszawy, D., Pudelek, M., Kochanowski, P., et al., 2020. High bisphenol A concen-trations augment the invasiveness of tumor cells through Snail-1/Cx43/ERRgamma-dependent epithelial-mesenchymal transition. Toxicol. Vitro 62,104676. https://doi.org/10.1016/j.tiv.2019.104676.
Sheller-Miller, S., Radnaa, E., Arita, Y., et al., 2020. Environmental pollutant inducedcellular injury is reflected in exosomes from placental explants. Placenta 89,42e49. https://doi.org/10.1016/j.placenta.2019.10.008.
Sun, G., Crissman, K., Norwood, J., et al., 2001. Oxidative interactions of syntheticlung epithelial lining fluid with metal-containing particulate matter. Am. J.Physiol. Lung Cell Mol. Physiol. 281 (4), L807eL815. https://doi.org/10.1152/ajplung.2001.281.4.L807.
Thery, C., Witwer, K.W., Aikawa, E., et al., 2018. Minimal information for studies ofextracellular vesicles 2018 (MISEV2018): a position statement of the Interna-tional Society for Extracellular Vesicles and update of the MISEV2014 guide-lines. J. Extracell. Vesicles 7 (1), 1535750. https://doi.org/10.1080/20013078.2018.1535750.
van Meteren, N., Lagadic-Gossmann, D., Chevanne, M., et al., 2019. Polycyclic aro-matic hydrocarbons can trigger hepatocyte release of extracellular vesicles byvarious mechanisms of action depending on their affinity for the aryl hydro-carbon receptor. Toxicol. Sci. (2), 2. https://doi.org/10.1093/toxsci/kfz157.
Wang, H., Sui, H., Zheng, Y., et al., 2019. Curcumin-primed exosomes potentlyameliorate cognitive function in AD mice by inhibiting hyperphosphorylation ofthe Tau protein through the AKT/GSK-3beta pathway. Nanoscale 11 (15),7481e7496. https://doi.org/10.1039/c9nr01255a.
Wang, Y., Zhao, M., Liu, J., et al., 2017. Up regulation of IL-6 is involved in di (2-ethylhexyl) phthalate (DEHP) induced migration and invasion of non smallcell lung cancer (NSCLC) cells. Biomed. Pharmacother. 89, 1037e1044. https://doi.org/10.1016/j.biopha.2017.02.107.
Xu, H., Ling, M., Xue, J.C., et al., 2018. Exosomal microRNA-21 derived from bronchialepithelial cells is involved in aberrant epithelium-fibroblast cross-talk in COPDinduced by cigarette smoking. Theranostics 8 (19), 5419e5433. https://doi.org/10.7150/thno.27876.
Yang, J., Mani, S.A., Donaher, J.L., et al., 2004. Twist, a master regulator ofmorphogenesis, plays an essential role in tumor metastasis. Cell 117 (7),927e939. https://doi.org/10.1016/j.cell.2004.06.006.
Zeitels, L.R., Acharya, A., Shi, G., et al., 2014. Tumor suppression by miR-26 overridespotential oncogenic activity in intestinal tumorigenesis. Genes Dev. 28 (23),2585e2590. https://doi.org/10.1101/gad.250951.114.
Zhang, X., Sai, B., Wang, F., et al., 2019a. Hypoxic BMSC-derived exosomal miRNAspromote metastasis of lung cancer cells via STAT3-induced EMT. Mol. Canc. 18(1), 40. https://doi.org/10.1186/s12943-019-0959-5.
Zhang, X., Wang, Q., Qiu, T., et al., 2019b. PM2.5 bound phthalates in four metro-politan cities of China: concentration, seasonal pattern and health risk viainhalation. Sci. Total Environ. 696, 133982. https://doi.org/10.1016/j.scitotenv.2019.133982.
























![26a Augustan Program[1]](https://img.pdfslide.net/doc/110x75/577ce3c61a28abf1038cfa67/26a-augustan-program1.jpg)




