Embed Size (px)
Citation preview

Divergent Regulation of Terpenoid Metabolism in theTrichomes of Wild and Cultivated Tomato Species1[W][OA]
Katrin Besser, Andrea Harper2, Nicholas Welsby3, Ines Schauvinhold, Stephen Slocombe, Yi Li,Richard A. Dixon, and Pierre Broun4*
Center for Novel Agricultural Products, Department of Biology, University of York, York YO10 5YW, UnitedKingdom (K.B., A.H., N.W., I.S., S.S., Y.L., R.A.D., P.B.); and Plant Biology Division, Samuel Roberts NobleFoundation, Ardmore, Oklahoma 73401 (R.A.D.)
The diversification of chemical production in glandular trichomes is important in the development of resistance againstpathogens and pests in two species of tomato. We have used genetic and genomic approaches to uncover some of thebiochemical and molecular mechanisms that underlie the divergence in trichome metabolism between the wild speciesSolanum habrochaites LA1777 and its cultivated relative, Solanum lycopersicum. LA1777 produces high amounts of insecticidalsesquiterpene carboxylic acids (SCAs), whereas cultivated tomatoes lack SCAs and are more susceptible to pests. We show thattrichomes of the two species have nearly opposite terpenoid profiles, consisting mainly of monoterpenes and low levels ofsesquiterpenes in S. lycopersicum and mainly of SCAs and very low monoterpene levels in LA1777. The accumulation patternsof these terpenoids are different during development, in contrast to the developmental expression profiles of terpenoidpathway genes, which are similar in the two species, but they do not correlate in either case with terpenoid accumulation.However, our data suggest that the accumulation of monoterpenes in S. lycopersicum and major sesquiterpenes in LA1777 arelinked both genetically and biochemically. Metabolite analyses after targeted gene silencing, inhibitor treatments, andprecursor feeding all show that sesquiterpene biosynthesis relies mainly on products from the plastidic 2-C-methyl-D-erythritol-4-phosphate pathway in LA1777 but less so in the cultivated species. Furthermore, two classes of sesquiterpenesproduced by the wild species may be synthesized from distinct pools of precursors via cytosolic and plastidial cyclases.However, highly trichome-expressed sesquiterpene cyclase-like enzymes were ruled out as being involved in the production ofmajor LA1777 sesquiterpenes.
Plants collectively produce a large diversity of sec-ondary metabolites as part of a defense strategyagainst pests, diseases, and different forms of abioticstress. Many of these compounds are produced at thesurface of the plant by epidermal secretory structurescalled glandular trichomes. These hair- or gland-likestructures typically consist of a unicellular or multi-cellular stalk capped by one or more secretory cellsthat accumulate or exude chemicals (McCaskill andCroteau, 1999; Werker, 2000). In two species of tomato,
the glandular type VI trichomes are particularly abun-dant on the leaves and stems of the cultivated speciesSolanum lycopersicum (formerly Lycopersicon esculentum)and its wild relative Solanum habrochaites (formerlyLycopersicon hirsutum) f. typicum LA1777 (Luckwill,1943; Gianfagna et al., 1992; Fig. 1, A and B). Theyhave been shown to accumulate monoterpenes in S.lycopersicum and very high levels of sesquiterpenes,mostly in the form of insecticidal carboxylic acid de-rivatives, in LA1777 (Coates et al., 1988, Frelichowskiand Juvik, 2001; Li et al., 2004; Fig. 1D).
On the basis of genetic evidence and heterologousexpression, it has been proposed that enzymes relatedto germacrene C synthase, corresponding to a locus onchromosome 6, mediate the accumulation of a groupof structurally similar compounds termed class I ses-quiterpenes (cI-Ss) in LA1777 and S. lycopersicum(Colby et al., 1998; van der Hoeven et al., 2000; Fig.1C). This group comprises germacrenes as well asa-humulene and b-caryophyllene. A distinct but so farunidentified enzyme or group of enzymes, associatedwith a locus on chromosome 8, has been hypothesizedto catalyze the formation of class II sesquiterpenes(cII-Ss) in LA1777 but not in S. lycopersicum (van derHoeven et al., 2000). These compounds are also struc-turally different from cI-Ss and include a-santalene,a-bergamotene, and b-bergamotene, which are thoughtto be further modified to form the sesquiterpene
1 This work was supported by a grant from the Samuel RobertsNoble Foundation.
2 Present address: School of Biosciences, University of Birming-ham, Birmingham B15 2TT, United Kingdom.
3 Present address: Physiology Laboratory, Downing Street, Cam-bridge CB2 3EG, United Kingdom.
4 Present address: Nestle R&D Center Tours, Plant Science andTechnology, 101 Avenue Gustave Eiffel, 37390 Notre-Dame D’Oe,France.
* Corresponding author; e-mail [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policydescribed in the Instructions for Authors (www.plantphysiol.org) is:Pierre Broun ([email protected]).
[W] The online version of this article contains Web-only data.[OA] Open Access articles can be viewed online without a sub-
scription.www.plantphysiol.org/cgi/doi/10.1104/pp.108.126276
Plant Physiology, January 2009, Vol. 149, pp. 499–514, www.plantphysiol.org � 2008 American Society of Plant Biologists 499
Dow
nloaded from https://academ
ic.oup.com/plphys/article/149/1/499/6107851 by guest on 27 N
ovember 2021

carboxylic acids (SCAs) a-santalenoic and a- andb-bergamotenoic acid, which have strong insecticidalproperties (Coates et al., 1988; Frelichowski and Juvik,2001; Fig. 1D).
Terpenoid production in plants is largely dependenton flux through two pathways that provide isopen-tenyl diphosphate (IPP) and dimethylallyl diphos-phate (DMAPP). Initially, it was suggested that themevalonate (MVA) pathway, which operates in thecytosol, supplies acetyl CoA-derived precursors forthe production of sesquiterpenes and triterpenes,whereas the plastid-localized 2-C-methyl-D-erythritol-4-phosphate (MEP) pathway provides pyruvate-derived IPP and DMAPP for the biosynthesis ofhemiterpenes, monoterpenes, diterpenes, and tetrater-penes (Lichtenthaler, 1999; Newman and Chappell,1999; Fig. 1E). However, this might be an oversimpli-
fication, and the contribution of the pathways tospecific compounds may be much more flexible andvariable. In lima bean (Phaseolus lunatus), for example,different stresses can differentially allocate precursorsto the synthesis of induced terpenoids, and limitingconditions in one pathway can be overcome usingprecursors supplied by the other pathway (Piel et al.,1998; Jux et al., 2001; Bartram et al., 2006). The sesqui-terpenes from mint (Mentha spp.) essential oils havebeen found to derive from the MEP pathway, like theemitted floral sesquiterpenes from snapdragon (Antir-rhinum majus) flowers, due to inactive MVA pathways(McCaskill and Croteau, 1995; Lange et al., 2000;Dudareva et al., 2005). Similarly, it has been shownthat some monoterpenes in strawberry (Fragaria spp.)have an exclusive MVA origin (Hampel et al., 2006).Sesquiterpene precursors in carrot (Daucus carota),
Figure 1. Sesquiterpene production in the glan-dular trichomes of S. lycopersicum and S. habro-chaites f. typicum LA1777. A and B, Scanningelectron microscopy images of stem surfaces of S.lycopersicum (cv M82; A) and LA1777 (B). Themost abundant trichomes (types IV, V ,and VI) areindicated by arrows. Bars = 1mm. C and D,Majorclass I (C) and class II (D) sesquiterpene deriva-tives found in the trichomes of S. lycopersicumand LA1777, respectively. b-car, b-Caryophyl-lene; a-hum, a-humulene; a-cS, a-cis santalene;a-cSA, a-cis santalenoic acid; a-cB, a-cis berga-motene; a-cBA, a-cis bergamotenoic acid. E,Current model for terpenoid biosynthesis in plants.DXP, Deoxyxylulose phosphate; fos, fosmidomycin,inhibitor of DXR; G3P, glyceraldehyde-3-phosphate;GPP, geranyl diphosphate; MTS, monoterpenesynthases; SST, sesquiterpene synthases.
Besser et al.
500 Plant Physiol. Vol. 149, 2009
Dow
nloaded from https://academ
ic.oup.com/plphys/article/149/1/499/6107851 by guest on 27 N
ovember 2021

chamomile (Matricaria chamomilla), and Solidago havebeen shown to be derived in part from both the MEPand MVA pathways (Adam et al., 1999; Steliopouloset al., 2002; Hampel et al., 2005). This metabolic crosstalk between cytosolic and chloroplastic compart-ments mediated by precursors such as IPP seems tohave a preferred direction of flux from plastids to thecytoplasm (Estevez et al., 2000; Bick and Lange, 2003;Hemmerlin et al., 2003; Laule et al., 2003; Flugge andGao, 2005).The study of metabolic pathway control in glandu-
lar trichomes has been hindered by the lack of tools forthe rapid evaluation of gene function. Recently, virus-induced gene silencing (VIGS) has emerged as a usefultool for the study of gene function in the Solanaceaespecies and other species (Burch-Smith et al., 2004;Robertson, 2004). However, there are still few exam-ples of its use in metabolic studies, and there is noevidence that the method can be used for studyingtrichome metabolism in cultivated and wild tomatospecies (Chen et al., 2004; Page et al., 2004; Park et al.,2005; van Schie et al., 2007).To investigate further the divergence in terpenoid
production between S. lycopersicum and S. habrochaites,we have undertaken a comparative study based on acombination of genetic, genomic, and physiologicalapproaches. We show that there is a close relationshipbetween monoterpene and cII-S metabolism at boththe genetic and biochemical levels. We also show thatcII-Ss in LA1777 are mostly derived from MEP pre-cursors, whereas the MVA pathway contributes moresignificantly to sesquiterpene production in S. lycoper-sicum. Our results further suggest that S. habrochaitesproduces its two classes of sesquiterpenes from dis-tinct precursor pools derived from the MVA and MEPpathways, respectively, which implies the existence ofa mechanism for partitioning sesquiterpene produc-tion, most likely by a different subcellular localizationof the corresponding cyclases.
RESULTS
Divergence in Terpenoid Quantity and Quality inTrichomes of S. lycopersicum and S. habrochaites LA1777
To date, the profiles of monoterpenes, sesquiter-penes, and their carboxylic acids (SCAs) in S. lycoper-sicum and LA1777 have been obtained in separateexperiments (Coates et al., 1988; van der Hoeven et al.,2000; Fridman et al., 2005). To obtain a completepicture of terpenoid accumulation in the two species,we profiled surface extracts of S. lycopersicum (cv M82)and LA1777 from comparable stem internodes. Mono-terpenes were prevalent in S. lycopersicum, with ses-quiterpenes as only minor components. The majormonoterpenes represented 90.5% of all terpenoidsmeasured, whereas known cI-Ss represented 0.5% ofthe total and cII-Ss were completely absent (Table I). InLA1777, sesquiterpenes and especially cII-S deriva-tives were the major components, with monoterpenes
either present in small amounts or absent (Table I). ThecII-SCAs represented 93.5%, and their presumed pre-cursors, the cII-Ss a-santalene, a-cis-bergamotene, anda-trans-bergamotene, together formed 0.7% of thetotal terpenoids, whereas the cI-Ss constituted 0.6%of the terpenoids measured (Table I).
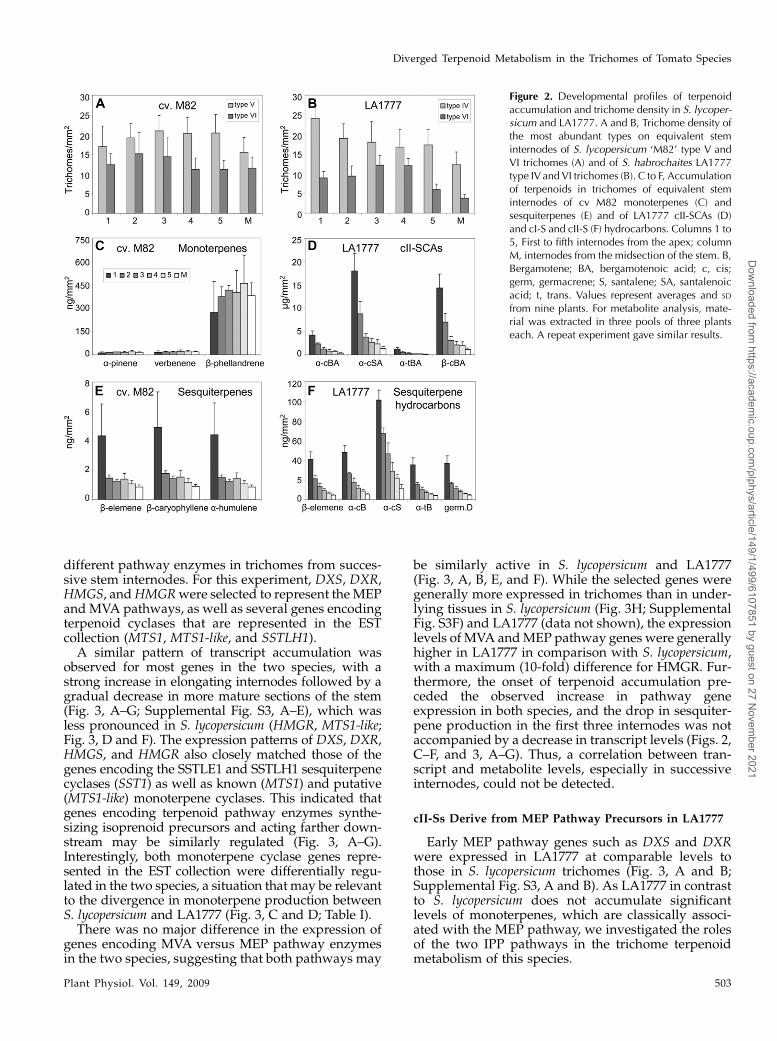
As shown in Figure 1B, there are two abundant typesof glandular trichomes present on stems of LA1777. Thetype VI trichomes are the second most abundant tri-chome type after glandular type IV in LA1777 and afternonglandular type V in S. lycopersicum (Figs. 1, A and B,and 2, A and B). Careful sampling of metabolites fromindividual glands with a capillary revealed that thecomposition of terpenoids in type VI trichome headswas nearly identical to that of extracts derived fromwhole tissue after immersion in chloroform, whichsuggested that all surface terpenoids measured areaccumulated in type VI trichomes in LA1777 (Supple-mental Fig. S1). The same result was obtained bysampling type VI glands from S. lycopersicum (datanot shown). Although type VI trichome densities onequivalent internodes were slightly lower in LA1777(Figs. 1, A and B, and 2, A and B), the trichomes ofLA1777 accumulated 8-fold higher levels of free terpe-noids per surface area of stems of greenhouse-grownplants than those of S. lycopersicum (Table I).
The Developmental Profiles of Terpenoid AccumulationAre Different in S. lycopersicum and LA1777
To determine whether fundamental differences in thedevelopmental pattern of terpenoid accumulation couldcontribute to the highly divergent metabolite profilesbetween the wild and cultivated species, terpenoidaccumulation was measured in successive internodesof 3-month-old S. lycopersicum and LA1777 plants.
Monoterpene accumulation remained fairly con-stant on an area basis in successive internodes of S.lycopersicum (Fig. 2C). In contrast, sesquiterpene levelswere much higher in younger tissue and decreasedfrom one internode to the next in LA1777 (Fig. 2, D andF). As trichome density in the uppermost internodesdid not change significantly despite the increase ininternode length (Fig. 2, A and B), terpenoid accumu-lation in trichomes was lower in older stem segmentsin the wild but not in the cultivated species. Interest-ingly, sesquiterpene levels likewise decreased fromyounger to older internodes in S. lycopersicum, espe-cially in the most apical internodes (Fig. 2E). Thereforesesquiterpene and monoterpene accumulation appearto be differentially regulated in tomato, although theaccumulation of each compound class may be simi-larly controlled in LA1777 and S. lycopersicum.
Genomic Analysis of S. habrochaitesGlandular Trichomes
To shed more light on the molecular basis of terpe-noid production in the wild species, we examined acollection of 2,435 public ESTs produced from LA1777trichomes (R.S. van der Hoeven, unpublished data).
Diverged Terpenoid Metabolism in the Trichomes of Tomato Species
Plant Physiol. Vol. 149, 2009 501
Dow
nloaded from https://academ
ic.oup.com/plphys/article/149/1/499/6107851 by guest on 27 N
ovember 2021

Cluster analysis produced a total of 1,641 unigenes,suggesting a relatively low level of redundancy amongthe expressed sequences. Some of the ESTs, however,were disproportionately represented, and interest-ingly, many of these corresponded to genes that couldbe associated with terpenoid production. In particular,ESTs for SSTLH1, which has been predicted to encodea cI-S cyclase (van der Hoeven et al., 2000), representedover 5% of all ESTs in the collection. Additionalpathway enzymes were also represented by multi-ple ESTs of the corresponding genes, for example3-hydroxy-3-methylglutaryl-CoA synthase (HMGS;eight ESTs), 3-hydroxy-3-methylglutaryl-CoA reductase(HMGR; seven ESTs), 1-deoxy-D-xylulose-5-phosphatesynthase (DXS; six ESTs), and 1-deoxy-D-xylulose-5-phosphate reductoisomerase (DXR; five ESTs; Sup-plemental Table S1). In total, 9% of the ESTs of the 100most highly represented genes could be associatedwith terpenoid metabolism, representing the highest
proportion for a single functional class (SupplementalFig. S2). Based on the predicted functions of theencoded proteins, many more of the 100 most highlyrepresented genes could be associated with otheraspects of metabolism, in particular lipid and cellwall metabolism (6% and 5% of the ESTs, respectively).A significant proportion of the EST collection alsoappeared to represent genes involved in stress re-sponses (8%) or defense (4%; Supplemental Fig. S2).These predicted functions were generally consistentwith the role of trichomes as epidermal defensivestructures actively secreting secondary metabolites.
Regulation of Terpenoid Pathway Gene Expression inTomato Trichomes
To determine whether transcriptional regulation islikely to play a role in controlling terpenoid produc-tion, we examined the expression of genes encoding
Table I. Composition of trichome-produced terpenoids in S. lycopersicum cv M82 and S. habrochaitesf. typicum LA1777
Terpenoids were from surface extracts of the fifth youngest stem internode of 3-month-old greenhouse-grown plants. Values are averages and SE (in parentheses) for nine plants. Only a selection of terpenoids isshown.
Compound LA1777 cv M82
ng mm22 % of total a ng mm22 % of total a
MonoterpenesLimonene 2.2 (0.1)b NDa-Pinene ND 19.8 (3.0)Verbenene ND 20.5 (3.0)d-2-Carene ND 150.1 (24.4)b-Phellandrene ND 458.4 (63.7)Total 2.2 (0.1)b 0.04 (0.007) 648.7 (94.1) 90.48 (1.49)
cI-Ssb-Elemene 6.4 (0.6) 1.1 (0.1)g-Elemene 4.8 (1.4) NDGermacrene D 6.0 (0.5) NDGermacrene B 2.4 (0.5) NDb-Caryophyllene 4.1 (0.3) 1.2 (0.1)a-Humulene 3.7 (0.2) 1.1 (0.1)Total 27.4 (3.5) 0.55 (0.09) 3.3 (0.24) 0.52 (0.06)
cII-S hydrocarbonsa-cis-Bergamotene 8.6 (0.8) NDa-Santalene 21.8 (2.2) NDa-trans-Bergamotene 5.3 (0.4) NDTotal 35.7 (3.4) 0.71 (0.08) 0.0 0.00
Carboxylic acidsa-cis-Bergamotenoic acid 713.8 (103.1) NDa-Santalenoic acid 2,340.6 (359.3) NDa-trans-Bergamotenoic acid 174.9 (26.3) NDb-cis-Bergamotenoic acid 1,968.9 (303.6) NDTotal 5,198.2 (792.3) 93.49 (1.27) 0.0 0.00
Otherc
b-Cubebene 9.7 (1.1) NDUnknown 15.2 (1.6) NDTotal 24.9 (2.7) 0.49 (0.06) 0.0 0.00
Total terpenoids 5,289.0 (802.8) 95.28 (1.50) 652.0 (94.3) 91.00 (1.55)
aCalculation based on the quantification of peak areas of all terpenoids in chromatograms fromchloroform extracts as described in “Materials and Methods.” bCould not be consistently detectedthroughout this study. cHas not been allocated to a specific class of sesquiterpenes. ND, Not detected.
Besser et al.
502 Plant Physiol. Vol. 149, 2009
Dow
nloaded from https://academ
ic.oup.com/plphys/article/149/1/499/6107851 by guest on 27 N
ovember 2021
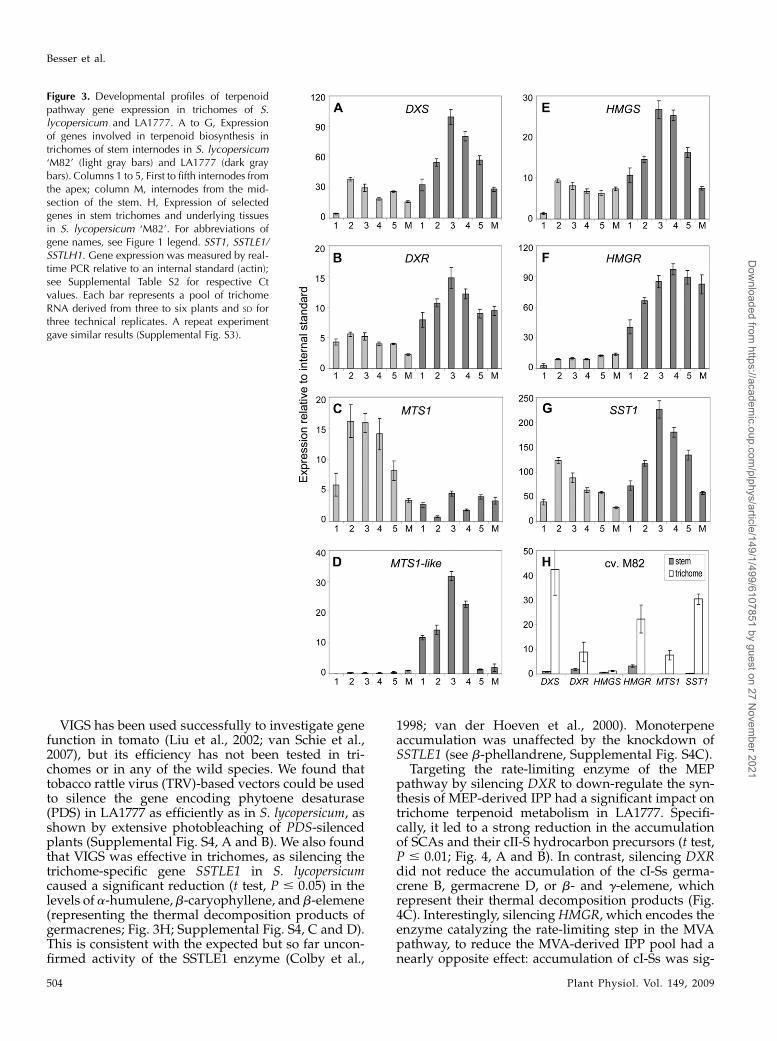
different pathway enzymes in trichomes from succes-sive stem internodes. For this experiment, DXS, DXR,HMGS, andHMGRwere selected to represent theMEPand MVA pathways, as well as several genes encodingterpenoid cyclases that are represented in the ESTcollection (MTS1, MTS1-like, and SSTLH1).A similar pattern of transcript accumulation was
observed for most genes in the two species, with astrong increase in elongating internodes followed by agradual decrease in more mature sections of the stem(Fig. 3, A–G; Supplemental Fig. S3, A–E), which wasless pronounced in S. lycopersicum (HMGR, MTS1-like;Fig. 3, D and F). The expression patterns of DXS, DXR,HMGS, and HMGR also closely matched those of thegenes encoding the SSTLE1 and SSTLH1 sesquiterpenecyclases (SST1) as well as known (MTS1) and putative(MTS1-like) monoterpene cyclases. This indicated thatgenes encoding terpenoid pathway enzymes synthe-sizing isoprenoid precursors and acting farther down-stream may be similarly regulated (Fig. 3, A–G).Interestingly, both monoterpene cyclase genes repre-sented in the EST collection were differentially regu-lated in the two species, a situation that may be relevantto the divergence in monoterpene production betweenS. lycopersicum and LA1777 (Fig. 3, C and D; Table I).There was no major difference in the expression of
genes encoding MVA versus MEP pathway enzymesin the two species, suggesting that both pathways may
be similarly active in S. lycopersicum and LA1777(Fig. 3, A, B, E, and F). While the selected genes weregenerally more expressed in trichomes than in under-lying tissues in S. lycopersicum (Fig. 3H; SupplementalFig. S3F) and LA1777 (data not shown), the expressionlevels of MVA andMEP pathway genes were generallyhigher in LA1777 in comparison with S. lycopersicum,with a maximum (10-fold) difference for HMGR. Fur-thermore, the onset of terpenoid accumulation pre-ceded the observed increase in pathway geneexpression in both species, and the drop in sesquiter-pene production in the first three internodes was notaccompanied by a decrease in transcript levels (Figs. 2,C–F, and 3, A–G). Thus, a correlation between tran-script and metabolite levels, especially in successiveinternodes, could not be detected.
cII-Ss Derive from MEP Pathway Precursors in LA1777
Early MEP pathway genes such as DXS and DXRwere expressed in LA1777 at comparable levels tothose in S. lycopersicum trichomes (Fig. 3, A and B;Supplemental Fig. S3, A and B). As LA1777 in contrastto S. lycopersicum does not accumulate significantlevels of monoterpenes, which are classically associ-ated with the MEP pathway, we investigated the rolesof the two IPP pathways in the trichome terpenoidmetabolism of this species.
Figure 2. Developmental profiles of terpenoidaccumulation and trichome density in S. lycoper-sicum and LA1777. A and B, Trichome density ofthe most abundant types on equivalent steminternodes of S. lycopersicum ‘M82’ type V andVI trichomes (A) and of S. habrochaites LA1777type IV and VI trichomes (B). C to F, Accumulationof terpenoids in trichomes of equivalent steminternodes of cv M82 monoterpenes (C) andsesquiterpenes (E) and of LA1777 cII-SCAs (D)and cI-S and cII-S (F) hydrocarbons. Columns 1 to5, First to fifth internodes from the apex; columnM, internodes from the midsection of the stem. B,Bergamotene; BA, bergamotenoic acid; c, cis;germ, germacrene; S, santalene; SA, santalenoicacid; t, trans. Values represent averages and SD
from nine plants. For metabolite analysis, mate-rial was extracted in three pools of three plantseach. A repeat experiment gave similar results.
Diverged Terpenoid Metabolism in the Trichomes of Tomato Species
Plant Physiol. Vol. 149, 2009 503
Dow
nloaded from https://academ
ic.oup.com/plphys/article/149/1/499/6107851 by guest on 27 N
ovember 2021
VIGS has been used successfully to investigate genefunction in tomato (Liu et al., 2002; van Schie et al.,2007), but its efficiency has not been tested in tri-chomes or in any of the wild species. We found thattobacco rattle virus (TRV)-based vectors could be usedto silence the gene encoding phytoene desaturase(PDS) in LA1777 as efficiently as in S. lycopersicum, asshown by extensive photobleaching of PDS-silencedplants (Supplemental Fig. S4, A and B). We also foundthat VIGS was effective in trichomes, as silencing thetrichome-specific gene SSTLE1 in S. lycopersicumcaused a significant reduction (t test, P # 0.05) in thelevels of a-humulene, b-caryophyllene, and b-elemene(representing the thermal decomposition products ofgermacrenes; Fig. 3H; Supplemental Fig. S4, C and D).This is consistent with the expected but so far uncon-firmed activity of the SSTLE1 enzyme (Colby et al.,
1998; van der Hoeven et al., 2000). Monoterpeneaccumulation was unaffected by the knockdown ofSSTLE1 (see b-phellandrene, Supplemental Fig. S4C).
Targeting the rate-limiting enzyme of the MEPpathway by silencing DXR to down-regulate the syn-thesis of MEP-derived IPP had a significant impact ontrichome terpenoid metabolism in LA1777. Specifi-cally, it led to a strong reduction in the accumulationof SCAs and their cII-S hydrocarbon precursors (t test,P # 0.01; Fig. 4, A and B). In contrast, silencing DXRdid not reduce the accumulation of the cI-Ss germa-crene B, germacrene D, or b- and g-elemene, whichrepresent their thermal decomposition products (Fig.4C). Interestingly, silencingHMGR, which encodes theenzyme catalyzing the rate-limiting step in the MVApathway, to reduce the MVA-derived IPP pool had anearly opposite effect: accumulation of cI-Ss was sig-
Figure 3. Developmental profiles of terpenoidpathway gene expression in trichomes of S.lycopersicum and LA1777. A to G, Expressionof genes involved in terpenoid biosynthesis intrichomes of stem internodes in S. lycopersicum‘M82’ (light gray bars) and LA1777 (dark graybars). Columns 1 to 5, First to fifth internodes fromthe apex; column M, internodes from the mid-section of the stem. H, Expression of selectedgenes in stem trichomes and underlying tissuesin S. lycopersicum ‘M82’. For abbreviations ofgene names, see Figure 1 legend. SST1, SSTLE1/SSTLH1. Gene expression was measured by real-time PCR relative to an internal standard (actin);see Supplemental Table S2 for respective Ctvalues. Each bar represents a pool of trichomeRNA derived from three to six plants and SD forthree technical replicates. A repeat experimentgave similar results (Supplemental Fig. S3).
Besser et al.
504 Plant Physiol. Vol. 149, 2009
Dow
nloaded from https://academ
ic.oup.com/plphys/article/149/1/499/6107851 by guest on 27 N
ovember 2021
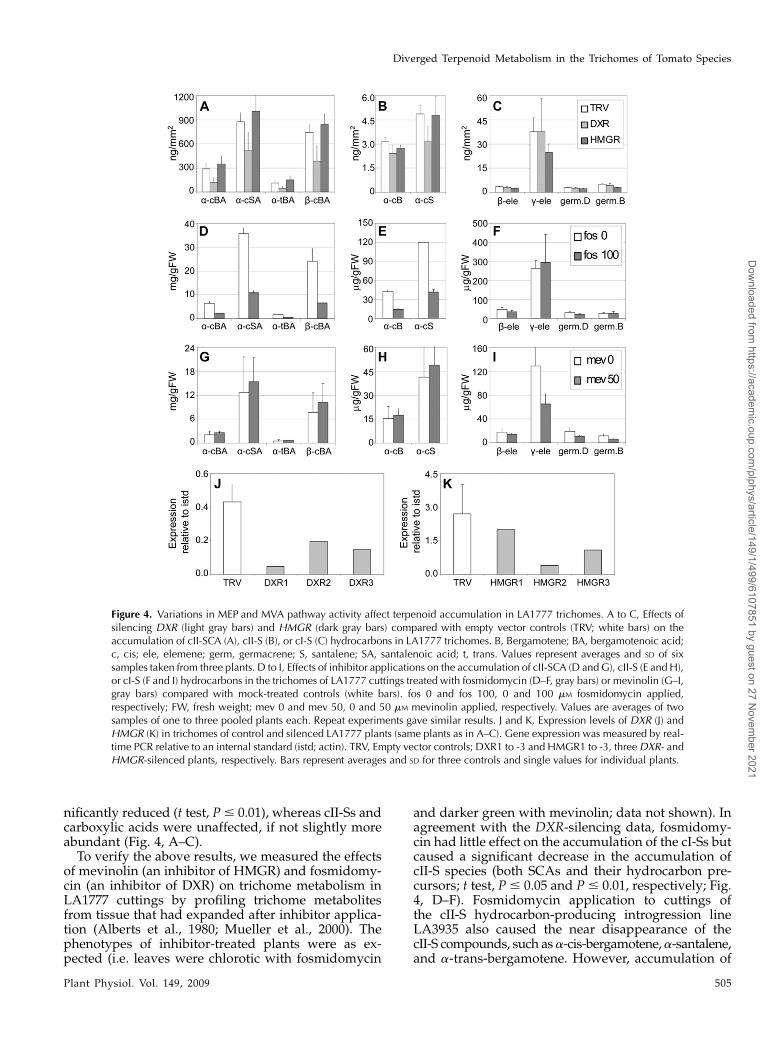
nificantly reduced (t test, P# 0.01), whereas cII-Ss andcarboxylic acids were unaffected, if not slightly moreabundant (Fig. 4, A–C).To verify the above results, we measured the effects
of mevinolin (an inhibitor of HMGR) and fosmidomy-cin (an inhibitor of DXR) on trichome metabolism inLA1777 cuttings by profiling trichome metabolitesfrom tissue that had expanded after inhibitor applica-tion (Alberts et al., 1980; Mueller et al., 2000). Thephenotypes of inhibitor-treated plants were as ex-pected (i.e. leaves were chlorotic with fosmidomycin
and darker green with mevinolin; data not shown). Inagreement with the DXR-silencing data, fosmidomy-cin had little effect on the accumulation of the cI-Ss butcaused a significant decrease in the accumulation ofcII-S species (both SCAs and their hydrocarbon pre-cursors; t test, P # 0.05 and P # 0.01, respectively; Fig.4, D–F). Fosmidomycin application to cuttings ofthe cII-S hydrocarbon-producing introgression lineLA3935 also caused the near disappearance of thecII-S compounds, such asa-cis-bergamotene,a-santalene,and a-trans-bergamotene. However, accumulation of
Figure 4. Variations in MEP and MVA pathway activity affect terpenoid accumulation in LA1777 trichomes. A to C, Effects ofsilencing DXR (light gray bars) and HMGR (dark gray bars) compared with empty vector controls (TRV; white bars) on theaccumulation of cII-SCA (A), cII-S (B), or cI-S (C) hydrocarbons in LA1777 trichomes. B, Bergamotene; BA, bergamotenoic acid;c, cis; ele, elemene; germ, germacrene; S, santalene; SA, santalenoic acid; t, trans. Values represent averages and SD of sixsamples taken from three plants. D to I, Effects of inhibitor applications on the accumulation of cII-SCA (D and G), cII-S (E and H),or cI-S (F and I) hydrocarbons in the trichomes of LA1777 cuttings treated with fosmidomycin (D–F, gray bars) or mevinolin (G–I,gray bars) compared with mock-treated controls (white bars). fos 0 and fos 100, 0 and 100 mM fosmidomycin applied,respectively; FW, fresh weight; mev 0 and mev 50, 0 and 50 mM mevinolin applied, respectively. Values are averages of twosamples of one to three pooled plants each. Repeat experiments gave similar results. J and K, Expression levels of DXR (J) andHMGR (K) in trichomes of control and silenced LA1777 plants (same plants as in A–C). Gene expression was measured by real-time PCR relative to an internal standard (istd; actin). TRV, Empty vector controls; DXR1 to -3 and HMGR1 to -3, threeDXR- andHMGR-silenced plants, respectively. Bars represent averages and SD for three controls and single values for individual plants.
Diverged Terpenoid Metabolism in the Trichomes of Tomato Species
Plant Physiol. Vol. 149, 2009 505
Dow
nloaded from https://academ
ic.oup.com/plphys/article/149/1/499/6107851 by guest on 27 N
ovember 2021

major cI-S b-caryophyllene was less affected, as wasthat of other more minor class I compounds (Supple-mental Fig. S5A). Consistent with the results ofHMGRsilencing, mevinolin application had no impact on theaccumulation of class II terpenoids in LA1777 butcaused a strong decrease in cI-S accumulation (Fig. 4,G–I).
To confirm that silencing and inhibitor applicationstruly target the MEP and MVA pathways, we supplied14C-labeled pyruvate (a MEP precursor) and [14C]MVAto LA1777 cuttings in the presence or absence ofinhibitors. After incubation, trichome metaboliteswere extracted and separated on thin-layer chroma-tography (TLC) plates (Supplemental Fig. S6), andlabel incorporation into sesquiterpene acids and hy-drocarbons was measured. Whereas SCAs werestrongly labeled at 3 h after [14C]pyruvate feeding,only low levels of incorporation (about 50 times less)were detected from [14C]MVA (Table II). The cI-S andcII-S hydrocarbons were not resolved under our TLCconditions. However, as a group, sesquiterpene hy-drocarbons were strongly labeled by [14C]pyruvate, inagreement with cII-S hydrocarbons acting as interme-diates in SCA biosynthesis. Hydrocarbons were alsolabeled from [14C]MVA, although less strongly, con-sistent with the low abundance of cI-S accumulation inLA1777 trichomes.
The above data suggest that the MEP pathwaydirectly contributes to cII-S formation and that thecontribution of the MVA pathway, if it exists, is neg-ligible. In contrast, the incorporation of 14C label fromMVA into sesquiterpene hydrocarbons indicated thatother noncarboxylic sesquiterpenes are likely derivedfrom the MVA pathway.
To show that 14C incorporation into SCAs frompyruvate depends on DXR and not on HMGR activity,LA1777 cuttings were treated with fosmidomycin ormevinolin prior to labeling. Fosmidomycin signifi-cantly inhibited incorporation, whereas mevinolinhad little or no effect (Table II).
Taken together, the results from VIGS experiments,inhibitor applications, and labeled precursor feedingstudies were consistent and suggested that pyruvate,but not MVA, is a direct precursor in the biosynthesis
of cII-Ss in LA1777. On the other hand, these data alsoindicate that cI-Ss are produced mainly from MVAviathe MVA pathway in LA1777.
The MVA and MEP Pathways Contribute toSesquiterpene Biosynthesis in S. lycopersicum
To evaluate the relative contributions of the MVAand MEP pathways to terpenoid biosynthesis in S.lycopersicum trichomes, we performed VIGS and in-hibitor studies in cv M82 plants.
Silencing of DXR, leading to the reduction of MEP-derived precursor accumulation, caused a significantdrop in monoterpene accumulation (t test, P # 0.01)but had no effect on sesquiterpene levels (Fig. 5, A andB). In contrast to the effects of DXR silencing, down-regulation of HMGR and MVA-derived precursorsynthesis did not affect monoterpene production(Fig. 5A) but caused a small and consistent decreasein cI-S accumulation (Fig. 5B). A similar effect was ob-served when DXR and HMGR were down-regulatedin the cvMicroTom plants (Supplemental Fig. S7, A–D).However, the down-regulation of transcript levels ofDXR and HMGR was variable among silenced plants(Fig. 5, E and F).
To assess further the contributions of the MVA andMEP pathways to the production of different trichometerpenoid classes in S. lycopersicum, cv M82 cuttingswere treated independently with mevinolin (MVA)and fosmidomycin (MEP) and trichome metaboliteswere profiled from the youngest tissue. The pheno-types of inhibitor-treated plants were as observed forLA1777. Consistent with the results of VIGS, fosmido-mycin nearly abolished monoterpene accumulation(Fig. 5C) but also caused a strong decline in theproduction of sesquiterpenes (t test, P # 0.01; Fig.5D). This effect was not obvious from VIGS experi-ments, which generally show a weaker phenotypethan inhibitor applications, as they reduce rather thaneliminate transcript levels and show some “patchi-ness” due to nonuniform spread of the virus (Fig. 5, Eand F; Supplemental Fig. S4A). Applying differentconcentrations of fosmidomycin (from 10 to 100 mM)showed that even the lowest concentration caused the
Table II. Incorporation of radiolabeled MVA and MEP pathway-derived precursors into sesquiterpenesin LA1777
Values are averages and SE (in parentheses) for four replicates. * Significant and ns no significantdifference of treatment versus control in t test at P # 0.05. fos, 100 mM fosmidomycin; mev, 100 mM
mevinolin.
Substrate Inhibitor Sesquiterpene Hydrocarbonsa SCAs
pmol g21 fresh weight h21
[14C]MVA (75 nmol, 2.5 mCi) 1.2 (0.3) 1.7 (0.3)[14C]Pyruvate (152.5 nmol, 2.5 mCi) Control 21.4 (4.8) 67.5 (13.2)
fos 6.8* (0.9) 20.2* (2.4)Control 13.5 (3.8) 41.5 (5.5)mev 16.5ns (3.7) 71.9ns (14.5)
aClass I and II sesquiterpene hydrocarbons were not resolved in this experiment.
Besser et al.
506 Plant Physiol. Vol. 149, 2009
Dow
nloaded from https://academ
ic.oup.com/plphys/article/149/1/499/6107851 by guest on 27 N
ovember 2021
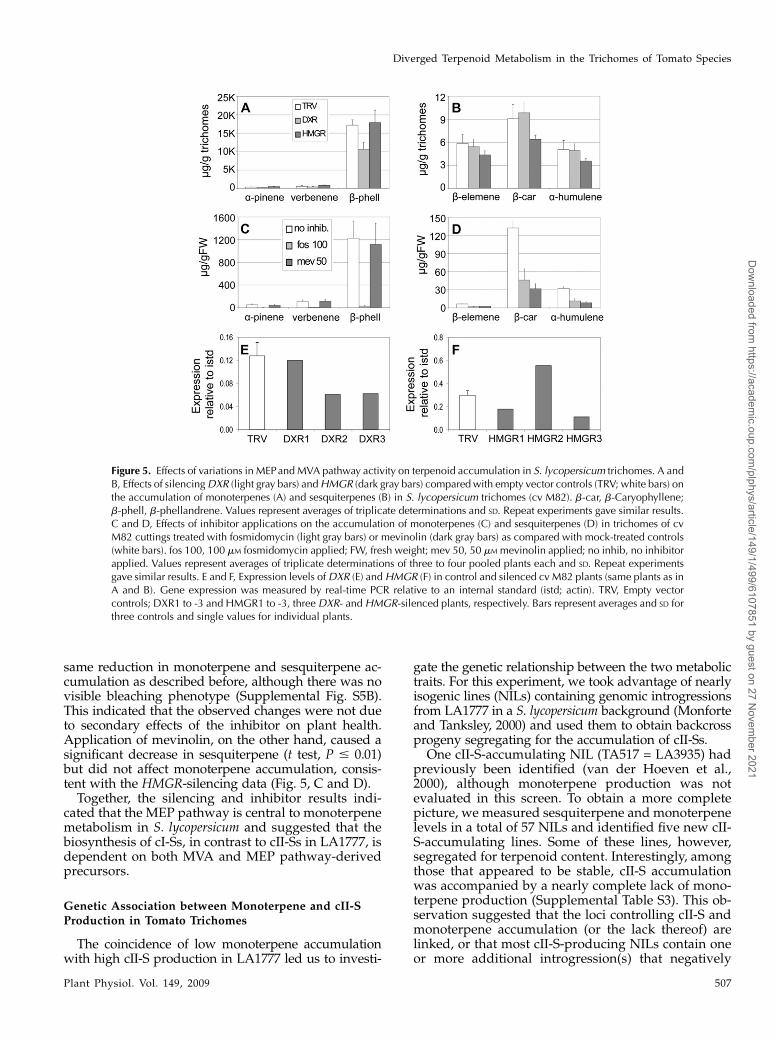
same reduction in monoterpene and sesquiterpene ac-cumulation as described before, although there was novisible bleaching phenotype (Supplemental Fig. S5B).This indicated that the observed changes were not dueto secondary effects of the inhibitor on plant health.Application of mevinolin, on the other hand, caused asignificant decrease in sesquiterpene (t test, P # 0.01)but did not affect monoterpene accumulation, consis-tent with the HMGR-silencing data (Fig. 5, C and D).Together, the silencing and inhibitor results indi-
cated that the MEP pathway is central to monoterpenemetabolism in S. lycopersicum and suggested that thebiosynthesis of cI-Ss, in contrast to cII-Ss in LA1777, isdependent on both MVA and MEP pathway-derivedprecursors.
Genetic Association between Monoterpene and cII-SProduction in Tomato Trichomes
The coincidence of low monoterpene accumulationwith high cII-S production in LA1777 led us to investi-
gate the genetic relationship between the two metabolictraits. For this experiment, we took advantage of nearlyisogenic lines (NILs) containing genomic introgressionsfrom LA1777 in a S. lycopersicum background (Monforteand Tanksley, 2000) and used them to obtain backcrossprogeny segregating for the accumulation of cII-Ss.
One cII-S-accumulating NIL (TA517 = LA3935) hadpreviously been identified (van der Hoeven et al.,2000), although monoterpene production was notevaluated in this screen. To obtain a more completepicture, we measured sesquiterpene and monoterpenelevels in a total of 57 NILs and identified five new cII-S-accumulating lines. Some of these lines, however,segregated for terpenoid content. Interestingly, amongthose that appeared to be stable, cII-S accumulationwas accompanied by a nearly complete lack of mono-terpene production (Supplemental Table S3). This ob-servation suggested that the loci controlling cII-S andmonoterpene accumulation (or the lack thereof) arelinked, or that most cII-S-producing NILs contain oneor more additional introgression(s) that negatively
Figure 5. Effects of variations in MEPand MVA pathway activity on terpenoid accumulation in S. lycopersicum trichomes. A andB, Effects of silencingDXR (light gray bars) andHMGR (dark gray bars) compared with empty vector controls (TRV; white bars) onthe accumulation of monoterpenes (A) and sesquiterpenes (B) in S. lycopersicum trichomes (cv M82). b-car, b-Caryophyllene;b-phell, b-phellandrene. Values represent averages of triplicate determinations and SD. Repeat experiments gave similar results.C and D, Effects of inhibitor applications on the accumulation of monoterpenes (C) and sesquiterpenes (D) in trichomes of cvM82 cuttings treated with fosmidomycin (light gray bars) or mevinolin (dark gray bars) as compared with mock-treated controls(white bars). fos 100, 100 mM fosmidomycin applied; FW, fresh weight; mev 50, 50 mM mevinolin applied; no inhib, no inhibitorapplied. Values represent averages of triplicate determinations of three to four pooled plants each and SD. Repeat experimentsgave similar results. E and F, Expression levels ofDXR (E) andHMGR (F) in control and silenced cv M82 plants (same plants as inA and B). Gene expression was measured by real-time PCR relative to an internal standard (istd; actin). TRV, Empty vectorcontrols; DXR1 to -3 and HMGR1 to -3, three DXR- and HMGR-silenced plants, respectively. Bars represent averages and SD forthree controls and single values for individual plants.
Diverged Terpenoid Metabolism in the Trichomes of Tomato Species
Plant Physiol. Vol. 149, 2009 507
Dow
nloaded from https://academ
ic.oup.com/plphys/article/149/1/499/6107851 by guest on 27 N
ovember 2021

affect(s) monoterpene production. To distinguish be-tween these two possibilities, we generated two BC1S1segregating populations by selfing backcross lines ob-tained from crosses between two cII-S-producing NILs(LA3935 and LA3940) with the cultivated parent cvE6203. About 40 progeny for each BC1S1 populationwere grown and analyzed for terpenoid production.
Three classes of progeny, which shared similar ter-penoid profiles, were identified in each population.The first class, which comprised the larger number ofprogeny in each population, produced monoterpenesthat were similar to those found in cv E6203 plants andalso accumulated cII-Ss. The second class accumulatedcII-Ss but little or no monoterpenes, and the third classproduced monoterpenes but no cII-Ss. The propor-tions of plants in each phenotypic category weresimilar in the two BC1S1 populations and suggestedthat cII-S and E6203 monoterpene accumulation areboth under the control of single dominant loci (TableIII). The sizes of the phenotypic groups and the lackof a fourth class of progeny, which produced neithercII-Ss nor E6203-type monoterpenes, were more com-patible with linkage between these two loci than withindependent assortment (Table III).
All plants producing E6203-type monoterpenesaccumulated a-pinene, verbenene, d-2-carene, andb-phellandrene, and these compounds always accu-mulated in nearly identical proportions (approxi-mately 3%, 5%, 20%, and 72%, respectively). Thisfinding and the segregation ratios of monoterpeneaccumulation strongly suggest that the accumulationof the different monoterpenes is controlled either by asingle gene or by closely linked genes.
The cII-S production was lower in plants that pro-duced monoterpenes than in plants that did not. Inboth populations, cII-S production was only half whenaccompanied by monoterpenes than that observed intheir absence, and monoterpene production appearedto be inversely affected (Table III). This suggests eitheran effect of semidominant alleles or competition forprecursors such as IPP between the cII-S and themonoterpene synthases.
Taken together, the above results suggested stronggenetic and metabolic associations between cII-S andmonoterpene production in the wild and cultivatedspecies.
The Role of Sesquiterpene Cyclases in cII-SAccumulation in LA1777
The molecular mechanisms controlling cII-S accu-mulation in LA1777 have not been defined, althoughgenetic analysis has ruled out the involvement of thecI-S synthase genes SSTLH1 and SSTLH2 or any genethat is highly similar in sequence to them (van derHoeven et al., 2000).
To examine the possible role of other, more divergedsesquiterpene cyclases in this process, we screened theEST collection and identified eight unigenes with sim-ilarity to known terpenoid synthases. Among these,four had highest similarity to sesquiterpene cyclases(Supplemental Table S4). The most highly expressed ofthese sequences corresponded to SSTLH1 (92 ESTs), andthe sequences of two other unigenes matched nonover-lapping sections of SSTLH2 (five and one EST, respec-tively). A fourth sequence, represented by only one EST(corresponding to GenBank accession no. AW616373)with little to no DNA sequence similarity to eitherSSTLH1 or SSTLH2, was associated with a predictedproduct most similar to sesquiterpene synthases, whichwe termed SSTLH3 (Supplemental Table S4).
Using a primer pair that could amplify an insertionin the LA1777 allele of SSTLH3, a smaller fragmentdiagnostic of the S. lycopersicum allele was found to bepresent in each introgression line tested, whereas noneof the lines contained the fragment for the LA1777allele (Supplemental Fig. S8A). This pattern is incom-patible with the presence of an introgressed fragmentat the SSTLH3 locus. In addition, the expression levelof SSTLH3 was much higher in S. lycopersicum than inLA1777 trichomes, and its developmental expressionpattern was different from those of the other terpenoidpathway genes tested in LA1777 (Fig. 3, A–G; Supple-mental Fig. S8, B and C).
Table III. Genetic analysis of S. lycopersicum monoterpene and LA1777 cII-S accumulation
Observed and expected numbers of BC1S1 progeny from isogenic lines accumulating S. lycopersicum monoterpenes (M) or LA1777 cII-Ss underfour different hypotheses: monoterpene (1) and cII-S accumulation (2) are under the control of single dominant loci; (3) the loci are completelylinked; (4) the loci are unlinked. The progeny are the F2s of two backcrosses to S. lycopersicum E6203, involving LA3935 (39 plants) and LA3940(40 plants). The data from both populations were pooled.
PhenotypeObserved No.
of Individuals
Expected No.
of Individuals Monoterpenes cII-Ss
M One Locus (1) cII-S One Locus (2) M/cII-S Linked (3) M/cII-S Unlinked (4)
ng cm22
M/2 19 19.75 19.75 14.81 2,034.5 0.0All M 62 59.25M/cII-S 43 39.50 44.44 1,457.8 272.8All cII-S 60 59.252/cII-S 17 19.75 19.75 14.81 13.5 486.92/2 0 0.00 4.94x2 probability 0.48 0.85 0.70 0.09
Besser et al.
508 Plant Physiol. Vol. 149, 2009
Dow
nloaded from https://academ
ic.oup.com/plphys/article/149/1/499/6107851 by guest on 27 N
ovember 2021

On the basis of these and previously reported re-sults, we conclude that cII-S production is unlikely toinvolve a highly trichome-expressed gene encoding atypical sesquiterpene cyclase.
DISCUSSION
In this report, we have investigated the molecularbasis of terpenoid accumulation and the relative con-tributions of precursor pathways in the glandulartrichomes of two related tomato species, S. lycopersi-cum and its wild relative S. habrochaites LA1777. Bothspecies have greatly diverged in the types andamounts of terpenoids they accumulate in trichomes:S. lycopersicum mainly produces monoterpenes andtraces of cI-Ss, whereas LA1777 produces mainly cII-Scarboxylic acids, small amounts of cI-Ss, and traces ofmonoterpenes (Table I). The monoterpenes and cI-Ssand cII-Ss analyzed in this study all derive from typeVI trichomes (Supplemental Fig. S1; Frelichowski andJuvik, 2001; Li et al., 2004). Despite a similar density oftype VI trichomes in both species, LA1777 accumu-lated much higher levels of measured terpenoids persurface area compared with S. lycopersicum. This couldbe related to the difference in volatility between mono-terpenes and SCAs and/or to differences in the chem-ical forms in which terpenoids accumulate in the twospecies (glycosylated, etc.). The different terpenoidlevels measured in both species could also result fromdifferences in trichome productivity. Determiningwhich is themost likely scenario would require furtheranalysis. The regulation of monoterpene and sesqui-terpene accumulation differs during development, asdecreasing levels of sequiterpenes and constant levelsof monoterpenes were measured in aging internodes.This might reflect different functions of these terpe-noids in the plant. However, the regulation patterns ofterpenoid synthesis seem to be conserved in bothspecies.
Developmental Regulation of Terpenoid Metabolism
Does Not Occur at the Transcript Level
Analysis of a public EST collection from LA1777trichomes showed that most ESTs represent geneproducts associated with metabolism, and about 9%of the ESTs representing the 100 most highly expressedsequences represent genes involved in terpenoid me-tabolism. In comparison with the representation ofother metabolic pathways in the trichomes of LA1777(e.g. flavonoid [3%] or lipid [6%] metabolism), therelatively higher EST representation of the terpenoidmetabolism reflects its high activity in the glandulartrichomes and is consistent with the high level ofterpenoid accumulation in LA1777 trichomes. It hasbeen shown that up to 1.2% of leaf fresh weightconsists of SCAs in LA1777 (Frelichowski and Juvik,2001), when trichome exudate levels of 3% to 30% ofleaf dry weight are considered to indicate high-levelaccumulators (Wagner, 1991).
The comparable EST representation of MEP to MVApathway genes in the LA1777 trichome library as wellas similar (to higher) expression levels of MEP path-way genes in LA1777 compared with S. lycopersicummay hint at the involvement of the MEP pathway inthe production of terpenoids in LA1777. Transcriptlevels of MVA pathway genes appeared to be up to 10-fold higher in LA1777 than in S. lycopersicum, and thecI-S synthase encoding SSTLH1 was the most highlyrepresented gene in the ESTcollection. This may reflectthe clearly higher levels of cI-S accumulation inLA1777 compared with S. lycopersicum but does notcorrelate with the main components of the terpenoidprofile in LA1777, namely the cII-SCAs. This discrep-ancy could be due to differences in turnover of cI-Ssand cII-Ss and/or in the activity and stability of theenzymes involved in their synthesis.
The expression levels of most of the genes of theMVA or MEP pathway and monoterpene or sesqui-terpene synthesis follow the same profile during de-velopment, suggesting a coordinated regulation ofterpenoid biosynthesis at the gene level. However,although expression of these genes is relatively tri-chome specific, the expression profiles do not mirrormetabolite accumulation during development, sug-gesting that terpenoid synthesis is not regulated at thetranscript level in tomato trichomes but rather in-volves other (posttranscriptional) regulatory mecha-nisms. Similarly, only a loose correlation betweenterpenoid pathway gene expression and enzyme ac-tivity has been found in basil (Ocimum basilicum)terpenoid metabolism (Iijima et al., 2004). This is incontrast to the regulation of monoterpene synthesis inmint, in which transcript, protein, and metaboliteaccumulation have been shown to follow the samedevelopmental pattern (McConkey et al., 2000). There-fore, terpenoid metabolism in glandular trichomesseems to be regulated at different levels dependingon the species involved.
A Functional Genomics Strategy for Dissecting
Terpenoid Metabolism in Trichomes of Wild andCultivated Tomato Species
Our results show that TRV-mediated VIGS can beused as an effective approach to silence trichome-expressed genes in cultivated as well as wild tomatospecies. By silencing SSTLE1, we have demonstrated,to our knowledge for the first time, the function of thisgene, confirming its expected activity in vivo (van derHoeven et al., 2000).
The effectiveness of VIGS in tomato trichomes willfacilitate future comparative genomic studies thatwould otherwise be impractical using stable transfor-mation. The validity of candidate gene approaches hasbeen demonstrated for studying the regulation ofepidermal metabolism in Arabidopsis (Broun et al.,2004), and VIGS should enable similar strategies intomato. However, since TRV does not induce completeand uniform silencing in tomato plants (including
Diverged Terpenoid Metabolism in the Trichomes of Tomato Species
Plant Physiol. Vol. 149, 2009 509
Dow
nloaded from https://academ
ic.oup.com/plphys/article/149/1/499/6107851 by guest on 27 N
ovember 2021

trichomes), VIGS will likely be most suitable for thestudy of processes that are particularly sensitive torelatively small variations in target gene expression.
Precursor Origin Influences Terpenoid AccumulationPatterns in Trichomes of Wild and Cultivated Tomato
Using a combination of strategies from VIGS toinhibitor application and labeled precursor feeding,we have demonstrated that the production of cII-Ss inLA1777 is, similar to monoterpene biosynthesis in S.lycopersicum, very sensitive to fluctuations in the MEPpathway but seemingly insensitive to variations in thesupply of MVA. cI-S production, in contrast, isstrongly influenced by changes in MVA pathwayactivity. The large difference in the levels of cI-Ss andcII-Ss in LA1777, therefore, may be as much due todifferences in precursor supply through the MEP andMVA pathways as to differences in catalytic activitiesof cI-S and cII-S cyclases and/or turnover of thecompounds. In S. lycopersicum, however, the biosyn-thesis of cI-Ss appears to rely partially also on non-MVA precursors, since inhibition of DXR activity byfosmidomycin is detrimental to their accumulation.This implies an exchange of precursors between com-partments from plastids to the cytosol (Adam et al.,1999; Hemmerlin et al., 2003; Laule et al., 2003;Dudareva et al., 2005), which is more pronounced inS. lycopersicum than in LA1777.
Our findings suggest the existence, in the wildspecies, of a mechanism controlling the productionof two distinct groups of sesquiterpenes from differentprecursor pools. Such a partition in sesquiterpenebiosynthesis could be the result of metabolite chan-neling, for example through two distinct farnesyldiphosphate (FPP) synthases in the cytoplasm thatare associated either with cI-S synthases or with plas-tidial transporters and the cII-S synthases. However, itseems unlikely that MVA precursors are available forthe biosynthesis of other terpenoid classes, such assterols, but not for cII-Ss, or that MEP precursorsexported into the cytoplasm are only available for cII-Ssynthases but not for other cytosolic enzymes. Aphysical separation of cI-S and cII-S synthases indifferent subcellular compartments would be a sim-pler explanation.
Since terpenoids in the mitochondria are known toderive from the MVA pathway (Disch et al., 1998), aplastidial site of synthesis would be most logical (Fig.6). Such localization would be highly unusual, assesquiterpene production is thought to be limited tothe cytoplasmic or mitochondrial compartments inplant cells. There are a few examples of sesquiterpenesderiving solely from MEP pathway precursors. Forexample, the biosynthesis of the phytohormone ab-scisic acid ultimately involves MEP precursors and isinitiated in chloroplasts. Interestingly, however, asabscisic acid derives from carotenoids, it is not pro-duced from FPP but from gernaylgeranyl diphosphate(Cutler and Krochko, 1999). Two other irregular ses-
quiterpenes, anthecotuloide from Anthemis cotula andhodgsonox from a liverwort (Marchantia polymorpha),are formed exclusively via the MEP pathway (Barlowet al., 2003; van Klink et al., 2003). Anthecotuloide hasbeen found to derive from a non-FPP precursor, whichhas been proposed to be synthesized via a “head-to-head” linkage of geranyl diphosphate to DMAPP (vanKlink et al., 2003). Another irregular sesquiterpene,artemone from Artemisia pallens, has been reported tooriginate from an unusual non-FPP precursor, pro-duced by condensation of IPP and dimethylvinylcar-binylpyrophosphate (Akhila et al., 1986). These reportsled us to speculate that if the cII-S synthase is localizedin the plastids, MEP-derived IPP and DMAPP mightbe used to produce a non-FPP precursor that is thenutilized to form the cII-Ss, as there are no reportsdescribing a substantial FPP synthase activity in plas-tids (Fig. 6). A localization in plastids would also easilyexplain the metabolic linkage between monoterpeneand cII-S accumulation in the NILs containing S.habrochaites introgressions. MEP-derived precursorsthat are available for monoterpene cyclases in theplastids of S. lycopersicum are also used by the cII-Scyclase in cII-S-producing introgression lines, leadingto a competition for precursors and a reduction in S.lycopersicum-type monoterpene synthesis. However,the ultimate discrimination between cytoplasmic and
Figure 6. Proposed terpenoid pathway in the trichomes of S. habro-chaites f. typicum LA1777 and S. lycopersicum.
Besser et al.
510 Plant Physiol. Vol. 149, 2009
Dow
nloaded from https://academ
ic.oup.com/plphys/article/149/1/499/6107851 by guest on 27 N
ovember 2021

plastidial localization requires the identification of thecII-S cyclase and its subcellular location.
cII-S Cyclase
The nature of the cII-S cyclase remains unclear, ascII-S and cI-S cyclases are thought to have substan-tially diverged in sequence (van der Hoeven et al.,2000). We confirm in this study that none of the highlyexpressed trichome-specific typical sesquiterpenecyclases are likely to produce cII-Ss. Our analysis ofcII-S-accumulating NILs suggests that LA1777 cII-SandE2603monoterpene accumulation are both inheritedas single-locus dominant traits. The backcross datasuggest that these two traits represent either alleles atthe same locus or two closely linked loci. Recent workby Kampranis et al. (2007) supports the possible evo-lutionarily simple interconversion of terpene syn-thases that are involved in the biosynthesis ofdifferent size classes of isoprenoids. Especially syn-thesis of monoterpenes and sesquiterpenes, includingthe cII-S trans-a-bergamotene, has been shown to beinterconverted by a single amino acid alteration of aterpene synthase. If the cII-S synthase has evolvedfrom a monoterpene synthase in tomato, it would belikely that it is still localized in the plastids, corrobo-rating our results on precursor origin. It remains to beshown if sufficient FPP supply is available in theplastids or if another non-FPP precursor is used as astarting point for cII-S synthesis.There are two highly expressed terpene synthase
homologs other than monoterpene or sesquiterpenesynthases represented in the trichome EST database,an ent-kaurene synthase and a prenyl transferase(Supplemental Table S1; TC116365, now TC174632,and TC124549). The close relationship of ent-kaurenesynthases with linalool synthases, which appear tohave evolved from a recombination event betweenmonoterpene and ent-kaurene synthases (Cseke et al.,1998; Aubourg et al., 2002), suggests this homolog as acandidate monoterpene/sesquiterpene synthase gene.Further investigation is necessary to show that themajor accumulated monoterpenes in S. lycopersicumand the cII-Ss in S. habrochaites are derived from thisgene product. The prenyl transferase homolog mayrepresent a candidate gene for non-FPP precursorproduction in the chloroplasts, but its function andputative involvement in the synthesis of cII-Ss in thewild tomato species has to be shown.The extensive differences in the terpenoid profiles of
LA1777 and S. lycopersicum trichomes underline theremarkable metabolic flexibility of glandular trichomes,the highly productive cellular factories that provide theplants with a huge variety of chemicals. Such metabolicshifts also reflect the staggering capacity of plants toevolve in their production of defense-related com-pounds in a relatively short evolutionary time. Ourdata suggest that the metabolic divergence from acommon ancestor required few evolutionary steps af-fecting catalytic mechanisms and precursor availability.
MATERIALS AND METHODS
Plant Materials and Growth Conditions
Tomatoes (Solanum lycopersicum ‘M82’, ‘Moneymaker’, ‘MicroTom’, and
‘E6203’ and Solanum habrochaites f. typicum accession LA1777 and backcross
recombinant inbred lines [introgression lines] representing the genome of
LA1777 in the background of S. lycopersicum E6203 [a core set of 57 NILs,
LA3913–LA3969; Monforte and Tanksley, 2000]) were obtained from the
Tomato Genetics Resource Center. Additional LA1777 seeds were generously
provided by Steven Tanksley (Cornell University). Prior to germination, seeds
were soaked in 10% (w/v) trisodium phosphate dodecahydrate for 30 min
and then for another 2 h at room temperature in fresh solution. After washing
five times in sterile water, seeds were incubated at 50�C in water (30 min),
placed in 2.7% sodium hypochlorite (30 min), and washed a further five times
in sterile water before being placed in soil. Plants were grown in the
greenhouse (16-h/8-h photoperiod) or in a growth chamber for silencing
experiments (16-h/8-h photoperiod, 20�C day, 18�C night) in 3.5-inch pots
(one plant per pot) using F2+S Levington modular compost.
VIGS
Conditions for VIGS were adapted from Liu et al. (2002). Plants were
grown in the greenhouse to the two-leaf stage, at which time they were
submerged upside down in Agrobacterium tumefaciens suspensions (GV3101;
optical density at 600 nm of approximately 0.4) containing the TRV RNA1-
and RNA2-derived pTRV1 and pTRV2 VIGS vectors (ratio 1:1) for vacuum
infiltration. Bacterial suspensions were obtained by inoculating Luria-Bertani
medium (50 mL, pH 7, with 10 mM MES, pH 5.5, 20 mM acetosyringone, and
antibiotic selection) with precultures (5 mL) that were incubated at 28�Covernight. Cells were washed with 10 mM MgCl2 solution, resuspended in 5
mL of infiltration medium (10 mM MgCl2, 10 mM MES, pH 5.5, and 200 mM
acetosyringone), and incubated for 2 h at room temperature before TRV1 and
TRV2 cultures were mixed and used for vacuum infiltration with Silwet L-77
(0.004%). The plants were then transferred to a growth chamber for 4 to 5
weeks prior to harvest.
For silencing experiments, gene-specific fragments were PCR amplified
from S. lycopersicum or S. habrochaites cDNA and verified by sequencing before
being cloned in antisense orientation into pTRV2 (pYL156; Liu et al., 2002) by
restriction digest. The following primers were designed from sequences in
the Tomato Gene Index databases (http://compbio.dfci.harvard.edu/tgi/):
SSTLE1 (TC162858): 5#-ATGTGTTCAAGCAATTCA-3# and 5#-ATCCAAA-
TCTTTCCACCA-3#; DXR (TC163170): 5#-TCCATTTGTCCTTCCTCCTG-3#and 5#-TGAATCCTGTGTTTCCACCA-3#; HMGR (TC155027): 5#-CCTA-
CACGCTTCCAAACACA-3# and 5#-AACCACCACATCGTCCTCTA-3#. A
pTRV2-PDS construct containing a genomic fragment of tomato PDS (X71023;
1,545 bp spanning exon 8 to exon 11; kindly provided by Sophia K. Ekengren,
Cornell University) was used as a control for silencing experiments.
RNA Extraction, Real-Time Reverse Transcription-PCR,and Genomic PCR Analyses
Trichome glands were harvested by abrasion of frozen stem tissue (three to
four internodes) and represented amixture of all trichome types. Other tissues
(approximately 100 mg) were ground in liquid nitrogen. The resulting
powders were dispersed in RNA extraction buffer (50 mM Tris base, pH 9,
0.8 mM NaCl, 10 mM EDTA, 0.5% [w/v] cetyl-trimethyl-ammonium bromide,
2% [w/v] polyvinylpolypyrrolidone, and 1% [v/v] b-mercaptoethanol). Fol-
lowing extractions with phenol:chloroform (1:1, v/v) and chloroform, the
RNA was recovered by ethanol precipitation and centrifugation in the cold,
before being resuspended in diethyl pyrocarbonate-treated water. After
treatment with DNase I (DNA-free; Ambion), cDNA was prepared using
SuperScript II reverse transcriptase (Invitrogen) according to the manufac-
turer’s protocol. Three microliters of a diluted reverse transcriptase reaction
was used for real-time PCR analysis. Amplification was performed in tripli-
cate reactions using the fluorescent dye SYBRGreen (Applied Biosystems) in a
thermocycler (AB7000 Sequence Detection System; Applied Biosystems).
Expression of a tomato LeACTIN gene (TC116322; http://compbio.dfci.
harvard.edu/tgi/) was used for normalization. Calibration curves were pro-
duced for each of the primer pairs, and quantification was performed using the
AB7000 Sequence Detection Software version 1.2.3 (Applied Biosystems).
Diverged Terpenoid Metabolism in the Trichomes of Tomato Species
Plant Physiol. Vol. 149, 2009 511
Dow
nloaded from https://academ
ic.oup.com/plphys/article/149/1/499/6107851 by guest on 27 N
ovember 2021

The following primers were designed from sequences in the Tomato Gene
Index databases (http://compbio.dfci.harvard.edu/tgi/) or fromGenBank and
were used for measuring transcript levels by real-time reverse transcription-
PCR: DXR (TC163170): 5#-ACTACCTTTTCGGAGCTGAGTATGA-3# and
5#-CCACCCTAGCTGTGCCAATACT-3#; DXS (TC154799): 5#-CAGAACT-
AAGAGCAGAAATTGTGTATTCA-3# and 5#-CCACAGTTAAATCCACAA-
CACCTAAA-3#; HMGR (TC155027): 5#-TCCGGTGGCGCTACGA-3# and
5#-TGCGCTGCCGAACCTAA-3#; HMGS (TC153567): 5#-CCTCTTCCCG-
AACTTGTAGGATT-3# and 5#-CACCGACGACGTTTATTTCCTT-3#; SSTLE1(TC162858): 5#-AGCAAACCTTAGAACAAACAAGCAA-3# and 5#-CCAAA-
CAGATGGGTGAAAATTAGC-3#; SSTLH1 (TC162148): 5#-GCAAACCTTA-
GAACAAACAAGCAATGG-3# and 5#-TAATTGTCTCTTTGTACTCATCAA
CTTCAAC-3#; SSTLH3 (AW616373): 5#-GAACTCATCAACACAATCCA-
ATGTC-3# and 5#-ATAGCATGAAGATCACCAATCGAA-3#; MTS1 (TC166486):
5#-GTAACATAGGGATGATGATTGTCACCTT-3# and 5#-CTGAACGCCTTGT-
GGTGGAAAT-3#; MTS1-like (AW617523): 5#-CCTCTCCACTGGACAGC-
CACTT-3# and 5#-CCACATGGTAGGCTCGTAATTCC-3#; ACTIN (TC116322):
5#-AAATTGTCAGGGACGTGAAAGAA-3# and 5#-TCTCAACAGAAGAGC-
TGGTCTTTG-3#. Each amplicon derived using these primers was cloned from
S. lycopersicum and S. habrochaites cDNA and sequenced to confirm amplification
of the target sequence.
The following primers were used to PCR amplify a diagnostic sequence
for SSTLH3 (AW616373) from genomic DNA, which was extracted using
standard methods: 5#-CTCCTTGTGAAAATGGAGTTGTGTA-3# and 5#-CCT-TGAAATTTCCTTGGTCATTAGT-3#. Amplicons derived from both tomato
species were cloned and sequenced to verify amplification of the target.
Extraction and Analysis of Trichome Essential Oils
Trichome essential oils were extracted from target tissues by immersion
of the tissue in chloroform for 2 to 3 h at room temperature. Alternatively,
the trichomes were first separated by abrasion of frozen tissue before being
added to the solvent (200 mL), which gave comparable terpenoid profiles to
the previous method (data not shown). The resulting extracts were dried
over Na2SO4 and filtered through Whatman paper before being concen-
trated, if necessary, to approximately 200 mL under nitrogen (a 5-fold
concentration). The content of individual type VI glands was collected from
leaflets with a capillary containing a small volume of solvent (50 mL) and
compared with leaf extracts from the opposite leaflet of the same leaf.
Tetradecane (10 ng mL21; Fluka) was added to the solvent as an internal
standard before extractions. For gas chromatography-mass spectrometry
analyses of sesquiterpene carboxylic acids, a separate fraction of the
chloroform extracts was derivatized in BSTFA-TMCS (99:1; Supelco) at
70�C for 1 h after complete drying of the original extracts under nitrogen.
The amount of input material was determined either by weighing or, in the
case of stems, by measuring their surface areas.
One to 2 mL of extract was used for gas chromatography-mass spectrom-
etry (GCQplus; ThermoQuest Finnigan) or gas chromatography-flame ioni-
zation detection (GC8000top; ThermoQuest Finnigan). The essential oil
constituents were separated on a ZB-1 column (30 m, 0.25 mm i.d., 0.50-mm
film thickness; Zebron-Phenomenex) using the following temperature profile:
50�C to 250�C, 5�C min21; 250�C to 320�C, 70�C min21; 320�C, 3 min; 320�C to
50�C, 70�C min21. Temperature of the injector was 250�C, that of the transfer
line was 320�C, and that of the ion source was 200�C.The essential oil constituents were identified by comparing their mass
spectra with those of true standards whenever possible. When standards were
not available, the mass spectra of the compounds were matched to published
information (Coates et al., 1988; van der Hoeven et al., 2000). In the latter case,
the Kovats index of the compounds was used in combination with spectral
information (Adams, 2004). Sesquiterpene carboxylic acids were identified
on the basis of their mass spectra, relative abundance, and similarity with
cII-S hydrocarbons (Coates et al., 1988). Standard curves of a-pinene and
a-humulene were used as a reference for the quantification of monoterpenes
and sesquiterpenes, respectively, after normalization of peak areas to the
internal standard and surface area or weight of input material.
Scanning Electron Microscopy Analysis
Stem segments were mounted on stubs and immersed in liquid nitrogen
before examination on the cold stage of a Hitachi S-5000 scanning electron
microscope.
Inhibitor Treatments and Radioactive Precursor Feeding
Treatments with fosmidomycin (Molecular Probes) and mevinolin (Sigma-
Aldrich) were performed on 3-week-old S. lycopersicum seedlings or side
shoots of mature S. habrochaites plants after severing them with a razor blade
under water. Inhibitor or control solutions were replaced weekly. After 19 to
20 d, the youngest shoot tissue was harvested for metabolite analysis.
For radiolabeled precursor feeding experiments, cuttings were placed in 50
mM phosphate buffer (pH 7.2) to which 2.5 mCi of RS-[2-14C]mevalonic acid
(dibenzyl ethylenediamine salt, specific activity of 67 mCi mmol21; Amer-
sham Bioscience) or [2-14C]pyruvic acid (sodium salt, specific activity of 16.4
mCi mmol21; Perkin-Elmer) had been added, until the solution was taken up.
The cuttings were then transferred to water (or inhibitor solutions) and
incubated for 3 to 5.5 h at room temperature. When appropriate, cuttings were
grown for 3 d in the presence of inhibitor (100 mM) before being fed
radiolabeled substrates.
Labeled terpenoids were concentrated and run on TLC plates (SILGUR-25
UV254; Macherey-Nagel) using hexane:diethylether:formic acid (80:20:2, v/v/v)
as the mobile phase. The plates were then exposed to phosphorimager screens,
and incorporation into the different terpenoids was measured using Quantity
One software (Bio-Rad).
Supplemental Data
The following materials are available in the online version of this
article.
Supplemental Figure S1. Terpenoid accumulation in type VI trichomes of
S. habrochaites LA1777.
Supplemental Figure S2. Functional classification of the 100 most highly
represented genes in an EST collection derived from LA1777 trichomes.
Supplemental Figure S3. Developmental profile of terpenoid pathway
gene expression in trichomes of S. lycopersicum and LA1777: repeat
experiment.
Supplemental Figure S4. VIGS as a tool for the analysis of gene function in
tomato species and trichomes.
Supplemental Figure S5. Effects of fosmidomycin applications on terpe-
noid profiles from cII-S-accumulating introgression line LA3935 (TA517)
and S. lycopersicum ‘M82’.
Supplemental Figure S6. TLC analysis of LA1777 sesquiterpenes.
Supplemental Figure S7. Effects of variations in MEP and MVA pathway
activity on terpenoid accumulation in S. lycopersicum ‘MicroTom’ tri-
chomes.
Supplemental Figure S8. Evaluation of a putative role for SSTLH3 in cII-S
production by genetic and gene expression analyses.
Supplemental Table S1. Unigenes represented in the LA1777 trichome
EST library annotated according to the results of BLASTX searches
against protein sequences deposited in GenBank.
Supplemental Table S2. Ct values for quantitative PCR amplifications for
developmental profiles of terpenoid pathway gene expression in tri-
chomes of S. lycopersicum (L) and S. habrochaites (H).
Supplemental Table S3. Terpenoid accumulation of cII-S-producing NILs
of LA1777 introgressions in cv E6203 background.
Supplemental Table S4. Terpene synthase-like genes represented in the
LA1777 trichome EST library.
ACKNOWLEDGMENTS
We thank Meg Stark for help with scanning electron microscopy analysis,
Tony Larson and Stuart Graham for assistance with metabolite profiling, and
Savithramma Dinesh-Kumar and Sophia Ekengren for providing pTRV1 and
pTRV2 vectors.
Received July 14, 2008; accepted November 4, 2008; published November 7,
2008.
Besser et al.
512 Plant Physiol. Vol. 149, 2009
Dow
nloaded from https://academ
ic.oup.com/plphys/article/149/1/499/6107851 by guest on 27 N
ovember 2021

LITERATURE CITED
Adam KP, Thiel R, Zapp J (1999) Incorporation of 1-[1-(13)C]deoxy-
D-xylulose in chamomile sesquiterpenes. Arch Biochem Biophys 369:
127–132
Adams RP (2004) Identification of Essential Oil Components by Gas
Chromatography/Quadrupole Mass Spectroscopy. Allured Publishing,
Carol Stream, IL
Akhila A, Sharma PK, Thakur RS (1986) A novel biosynthesis of irregu-
lar sesquiterpene artemone in Artemisia pallens. Tetrahedron Lett 27:
5885–5888
Alberts AW, Chen J, Kuron G, Hunt V, Huff J, Hoffman C, Rothrock J,
Lopez M, Joshua H, Harris E, et al (1980) Mevinolin: a highly potent
competitive inhibitor of hydroxymethylglutaryl-coenzyme A reductase
and a cholesterol-lowering agent. Proc Natl Acad Sci USA 77: 3957–3961
Aubourg S, Lecharny A, Bohlmann J (2002) Genomic analysis of the
terpenoid synthase (AtTPS) gene family of Arabidopsis thaliana. Mol
Genet Genomics 267: 730–745
Barlow AJ, Lorimer SD, Morgan ER, Weavers RT (2003) Biosynthesis of the
sesquiterpene hodgsonox from the New Zealand liverwort Lepidolaena
hodgsoniae. Phytochemistry 63: 25–29
Bartram S, Jux A, Gleixner G, Boland W (2006) Dynamic pathway
allocation in early terpenoid biosynthesis of stress-induced lima bean
leaves. Phytochemistry 67: 1661–1672
Bick JA, Lange BM (2003) Metabolic cross talk between cytosolic and
plastidial pathways of isoprenoid biosynthesis: unidirectional transport
of intermediates across the chloroplast envelope membrane. Arch
Biochem Biophys 415: 146–154
Broun P, Poindexter P, Osborne E, Jiang CZ, Riechmann JL (2004) WIN1, a
transcriptional activator of epidermal wax accumulation in Arabidop-
sis. Proc Natl Acad Sci USA 101: 4706–4711
Burch-Smith TM, Anderson JC, Martin GB, Dinesh-Kumar SP (2004)
Applications and advantages of virus-induced gene silencing for gene
function studies in plants. Plant J 39: 734–746
Chen JC, Jiang CZ, Gookin TE, Hunter DA, Clark DG, Reid MS (2004)
Chalcone synthase as a reporter in virus-induced gene silencing studies
of flower senescence. Plant Mol Biol 55: 521–530
Coates RM, Denissen JF, Juvik JA, Babka BA (1988) Identification of
alpha-santalenoic and endo-beta bergamotenoic acids as moth oviposi-
tion stimulants from wild tomato leaves. J Org Chem 53: 2186–2192
Colby SM, Crock J, Dowdle-Rizzo B, Lemaux PG, Croteau R (1998)
Germacrene C synthase from Lycopersicon esculentum cv. VFNT cherry
tomato: cDNA isolation, characterization, and bacterial expression of
the multiple product sesquiterpene cyclase. Proc Natl Acad Sci USA 95:
2216–2221
Cseke L, Dudareva N, Pichersky E (1998) Structure and evolution of
linalool synthase. Mol Biol Evol 15: 1491–1498
Cutler AJ, Krochko JE (1999) Formation and breakdown of ABA. Trends
Plant Sci 4: 472–478
Disch A, Hemmerlin A, Bach TJ, Rohmer M (1998) Mevalonate-derived
isopentenyl diphosphate is the biosynthetic precursor of ubiquinone
prenyl side chain in tobacco BY-2 cells. Biochem J 331: 615–621
Dudareva N, Andersson S, Orlova I, Gatto N, Reichelt M, Rhodes D,
Boland W, Gershenzon J (2005) The nonmevalonate pathway supports
both monoterpene and sesquiterpene formation in snapdragon flowers.
Proc Natl Acad Sci USA 102: 933–938
Estevez JM, Cantero A, Romero C, Kawaide H, Jimenez LF, Kuzuyama T,
Seto H, Kamiya Y, Leon P (2000) Analysis of the expression of CLA1,
a gene that encodes the 1-deoxyxylulose 5-phosphate synthase of the
2-C-methyl-D-erythritol-4-phosphate pathway in Arabidopsis. Plant
Physiol 124: 95–104
Flugge U-I, Gao W (2005) Transport of isoprenoid intermediates across
chloroplast envelope membranes. Plant Biol 7: 91–97
Frelichowski JE, Juvik JA (2001) Sesquiterpene carboxylic acids from a
wild tomato species affect larval feeding behavior and survival of
Helicoverpa zea and Spodoptera exigua (Lepidoptera: Noctuidae). J Econ
Entomol 94: 1249–1259
Fridman E, Wang J, Iijima Y, Froehlich JE, Gang DR, Ohlrogge J,
Pichersky E (2005) Metabolic, genomic, and biochemical analyses of
glandular trichomes from the wild tomato species Lycopersicon hirsutum
identify a key enzyme in the biosynthesis of methylketones. Plant Cell
17: 1252–1267
Gianfagna TJ, Carter CD, Sacalis JN (1992) Temperature and photoperiod
influence trichome density and sesquiterpene content of Lycopersicon
hirsutum f. hirsutum. Plant Physiol 100: 1403–1405
Hampel D, Mosandl A, Wust M (2005) Biosynthesis of mono- and sesqui-
terpenes in carrot roots and leaves (Daucus carota L.): metabolic cross
talk of cytosolic mevalonate and plastidial methylerythritol phosphate
pathways. Phytochemistry 66: 305–311
Hampel D, Mosandl A, Wust M (2006) Biosynthesis of mono- and sesqui-
terpenes in strawberry fruits and foliage: 2H labeling studies. J Agric
Food Chem 54: 1473–1478
Hemmerlin A, Hoeffler JF, Meyer O, Tritsch D, Kagan IA, Grosdemange-
Billiard C, Rohmer M, Bach TJ (2003) Cross-talk between the cytosolic
mevalonate and the plastidial methylerythritol phosphate pathways in
tobacco Bright Yellow-2 cells. J Biol Chem 278: 26666–26676
Iijima Y, Davidovich-Rikanati R, Fridman E, Gang DR, Bar E, Lewinsohn E,
Pichersky E (2004) The biochemical and molecular basis for the divergent
patterns in the biosynthesis of terpenes and phenylpropenes in the peltate
glands of three cultivars of basil. Plant Physiol 136: 3724–3736
Jux A, Gleixner G, Boland W (2001) Classification of terpenoids according
to the methylerythritolphosphate or the mevalonate pathway with
natural 12C/13C isotope ratios: dynamic allocation of resources in
induced plants. Angew Chem Int Ed 40: 2091–2093
Kampranis SC, Ioannidis D, Purvis A, Mahrez W, Ninga E, Katerelos NA,
Anssour S, Dunwell JM, Degenhardt J, Makris AM, et al (2007)
Rational conversion of substrate and product specificity in a Salvia
monoterpene synthase: structural insights into the evolution of terpene
synthase function. Plant Cell 19: 1994–2005
Lange BM, Wildung MR, Stauber EJ, Sanchez C, Pouchnik D, Croteau R
(2000) Probing essential oil biosynthesis and secretion by functional
evaluation of expressed sequence tags from mint glandular trichomes.
Proc Natl Acad Sci USA 97: 2934–2939
Laule O, Furholz A, Chang HS, Zhu T, Wang X, Heifetz PB, Gruissem W,
Lange M (2003) Crosstalk between cytosolic and plastidial pathways of
isoprenoid biosynthesis in Arabidopsis thaliana. Proc Natl Acad Sci USA
100: 6866–6871
Li L, Zhao Y, McCaig BC, Wingerd BA, Wang J, Whalon ME, Pichersky E,
HoweGA (2004) The tomato homolog of CORONATINE-INSENSITIVE1
is required for the maternal control of seed maturation, jasmonate-signaled
defense responses, and glandular trichome development. Plant Cell 16:
126–143
Lichtenthaler HK (1999) The 1-deoxy-D-xylulose-5-phosphate pathway of
isoprenoid biosynthesis in plants. Annu Rev Plant Physiol Plant Mol
Biol 50: 47–65
Liu Y, Schiff M, Dinesh-Kumar SP (2002) Virus-induced gene silencing in
tomato. Plant J 31: 777–786
Luckwill LC (1943) The genus Lycopersicon. Aberdeen University Studies
120: 5–44
McCaskill D, Croteau R (1995)Monoterpene and sesquiterpene biosynthesis in
glandular trichomes of peppermint (Mentha x piperita) rely exclusively on
plastid-derived isopentenyl diphosphate. Planta 197: 49–56
McCaskill D, Croteau R (1999) Strategies for bioengineering the develop-
ment and metabolism of glandular tissues in plants. Nat Biotechnol 17:
31–36
McConkey ME, Gershenzon J, Croteau RB (2000) Developmental regula-
tion of monoterpene biosynthesis in the glandular trichomes of pep-
permint. Plant Physiol 122: 215–224
Monforte AJ, Tanksley SD (2000) Development of a set of near isogenic
and backcross recombinant inbred lines containing most of the Lycoper-
sicon hirsutum genome in a L. esculentum genetic background: a tool for
gene mapping and gene discovery. Genome 43: 803–813
Mueller C, Schwender J, Zeidler J, Lichtenthaler HK (2000) Properties
and inhibition of the first two enzymes of the non-mevalonate pathway
of isoprenoid biosynthesis. Biochem Soc Trans 28: 792–793
Newman JD, Chappell J (1999) Isoprenoid biosynthesis in plants: carbon
partitioning within the cytoplasmic pathway. Crit Rev BiochemMol Biol
34: 95–106
Page JE, Hause G, Raschke M, Gao W, Schmidt J, Zenk MH, Kutchan TM
(2004) Functional analysis of the final steps of the 1-deoxy-D-xylulose
5-phosphate (DXP) pathway to isoprenoids in plants using virus-
induced gene silencing. Plant Physiol 134: 1401–1413
Park JA, Kim TW, Kim SK, Kim WT, Pai HS (2005) Silencing of NbECR
encoding a putative enoyl-CoA reductase results in disorganized mem-
brane structures and epidermal cell ablation in Nicotiana benthamiana.
FEBS Lett 579: 4459–4464
Diverged Terpenoid Metabolism in the Trichomes of Tomato Species
Plant Physiol. Vol. 149, 2009 513
Dow
nloaded from https://academ
ic.oup.com/plphys/article/149/1/499/6107851 by guest on 27 N
ovember 2021

Piel J, Donath J, Bandemer K, Boland W (1998) Mevalonate-independent
biosynthesis of terpenoid volatiles in plants: induced and constitutive
emission of volatiles. Angew Chem Int Ed 37: 2478–2481
Robertson D (2004) VIGS vectors for gene silencing: many targets, many
tools. Annu Rev Plant Biol 55: 495–519
Steliopoulos P, Wust M, Adam K-P, Mosandl A (2002) Biosynthesis of
sesquiterpene germacrene D in Solidago canadensis: 13C and 2H labeling
studies. Phytochemistry 60: 13–20
van der Hoeven RS, Monforte AJ, Breeden D, Tanksley SD, Steffens JC
(2000) Genetic control and evolution of sesquiterpene biosynthesis in
Lycopersicon esculentum and L. hirsutum. Plant Cell 12: 2283–2294
van Klink J, Becker H, Andersson S, Boland W (2003) Biosynthesis of
anthecotuloide, an irregular sesquiterpene lactone from Anthemis cotula
L. (Asteraceae) via a non-farnesyl diphosphate route. Org Biomol Chem
1: 1503–1508
van Schie CCN, Ament K, Schmidt A, Lange T, Haring MA, Schuurink
RC (2007) Geranyl diphosphate synthase is required for biosynthesis of
gibberellins. Plant J 52: 752–762
Wagner GJ (1991) Secreting glandular trichomes: more than just hairs.
Plant Physiol 96: 675–679
Werker E (2000) Trichome diversity and development. InDLHallahan, JC Gray,
eds, Advances in Botanical Research. Elsevier Science, Amsterdam, pp 1–35
Besser et al.
514 Plant Physiol. Vol. 149, 2009
Dow
nloaded from https://academ
ic.oup.com/plphys/article/149/1/499/6107851 by guest on 27 N
ovember 2021





























