Embed Size (px)
Citation preview

ABSThACT OP THE THESE OP
Gerald Q. Still ror the Maeter oZ Science degree in Cheiiistry.
Dat. thesis le presented ----T -'í?'3.
Titi. TH OCCURRENCE OP THE N-DOUD1OFF PATHWAY IN ATìCT] VINELíNDII
Abstract Approved.
The c*tàcUrn of glucose in proltterating cille of Azotobactsi has been examined with r.epsst to the nature end Uts aticn of the individuai pathways. The radiorespi5lMtric method, as wefl as incorporation end degradation experita, were used in this investigation.
The radiorespirometric data has been interpreted to indicate that the &itner-Doudorot'f and Pentose Phosphate pathwa7s are playing inortent roles in glucose catabolien. The thden-yerhof-Parnas pathway, if' present, does not contribute xmich to the overall utilization of glucose, Incorporation rnd deadation experimente support the findings ob3erved in the radioreaplrnetric expernte and revealed the operation of the Hexose Cycle in these proliferating cuiture3. lt is estinted that the operation of the Pentosa Phornhat. pathy in this organism should be not greater than ZQ% o1 the totaUr eatabolized glucose. Therefore, the Etner-Do*ioroft pathway represents the major glucoac utilizing pathway in opbacter e14I,

THE OCCURRENCE OF THE ENTNER-DOtJDOROFF PATHWAY IN AZOTOBACTER VINELANDII
by
GERALD G. STILL
A THESIS
submitted to
OREGON STATE UNIVERSITY
in partial fulfillment of the requirements for the
degree of
MASTER OF SCIENCE
June 1963

APPROVED:
ernistry
In Charge of Major
rman of Department of Chemistry
Date thesis is presented 1zLLí,3T /3 Typed by Kay J. Stili

THIS THESIS IS DEDICATED TO "MY THREE STILLS" WHO WERE INSTRUMENTAL IN THE CONDENSATION AND COMPLETION OF THIS STUDY.

ACKNOWLEDGEMEiTS
The author wishes to acknowledge Dr. A. C. Zagallo
who introduced this unique microbe to this laboratory.
To Mrs. Julie Krackov who nurtured and studied this
nitrogen-fixing bacterium with sufficient finesse to
yield the basic patterns upon which this thesis was
built, I extend my appreciation.
My deepest gratitude to my major professor
and friend, Dr. Chih H. Wang, for his patience and
unfaltering confidence.
Financial assistance was provided by Federal
grants from the Atomic Energy Commission and. the
National Institute of Health.

TABLE OF CONTENTS
Page
INTRODUCTION. . . . . , . . . . . . . . . . . . . i
MATERIALS AND METHODS , . . . . . . . . . . . . . . 3
Culture Conditions . . . . . . . . e . s i s e 3 Radiochemical Substrates . . . . . . . . . . . :3
Radlorespirometric Experiments . . . . . . . .
Incorporation Experiments . . . . . e . e 14.
Degradation Procedures . . . . . . . s e e e e 6 Determination of Radioactivity . . . . . . . s 7
RESULTS. . . . . . . . . . . . . . . . . . . . . 9
DISCUSSION. . . . . . . . . . . . . . . . . . . .
BIBLIOGRAPHY . . . . . . . . . e e s 22

LIST OF FIGURES
Figure 1. Radiorespirometric Pattern. Azotobacter vinelan&1i metabolizing specifically labeled glucose . . . . . . 10
Figure 2. Rad.iorespirometrlo Pattern. Azptpbpcter vinelandli metabolizing specifically labeled gluconate . . . . . li

LIST OF TABLES
Page
Table I, Utilization of Labeled Substrate by ptobacter vinelandli . . . . . . 12
Table II. Incorporation of G1ucose-C NaHCO3 into Azotobacte vinelandii . . . . . 13
Table III. The Isotopic Distribution Pattcr of Cel1ulr Alanine , , 19

THE OCCURRENCE OF THE ENTNER-DOUDOROFF PATHWAY IN AZOTOBACTER VINELANDII
INTRODUCT ION
Glucose catabolism in &zptpbgcter vinelandl.i has
been of interest to a number of research workers for some
time. These investigations have been stimulated pri-
manly by the ability of this organism to assimilate
molecular nitrogen and to oxidize extensively carbona-
ceous substrates.
It has been shown by Mortenson and Wilson (12, 13)
that glucose can be converted to 6-phosphogluconate
(6-PG) by extracts vinelandit. This
finding implies that either the Pentose Phosphate (P?)
pathway (i1i) or the Entner-Doudoroff (ED) pathway (1i), or
both, are operative in this organism. The operation
of the ED pathway was later suggested by W11on J,.,
(li) on the grounds that 6-PG was found to be cleaved by
coil-free extracts to g3yceraldehyde-3-phosphate (G_3-P)
and pynuvic acid. However, the two key enzymes associated
with the ED pathway, i.e., 6-phosphogluconic dehydrogenase
and 2-keto-3-deoxy-6-phosphogluconic-aldolase, have not
been detected in this organism. In addition, there has
been demonstrated the occurrence of 6-phosphogluconic dehy-
drogenase (13), transaldolase and transketolase (10) in
cell-free extracts of vinelandil which are the key steps

2
in the PP pathway. Al]. of the enzymes of the
Embden-Meyerhof-Parnas (Er'1P) pathway, excluding
phosphohexokinase, have been demonstrated to occur in
cell-free extracts of vinelandli. It was suggested.
by Wilson ., (13) that phosphohexokinase may have
been destroyed in preparation of the cell-free extract.
More recently Sobek and Clifton (7, 17) suggested that
in Azotobacter ai.lis, glucose may have been utilized
via the concurrent operation of the EMP and PP pathways.
The utilization of pyruvate, a key intermediate in
glucose catabolism, has been extensively studied in
vineandi (19, 20, 21). This includes the demon-
stration of the occurrence of the tricarboxylic acid
(TCA) cycle as evidenced by the observed oxidation of
TCA cycle intermediates with cell-free extracts and
the incorporation of acetate-l-C into cellular
constituents.
In the present study the catabolism of glucose
in proliferating cells of vinelandil has been examined
particularly with respect to the nature and the partici.-
pation of the individual pathways. Radiorespirometric
data (26) for the utilization of C specifically labeled
glucose and gluconate are presented along with the experi-
mental findings of the incorporation of specifically
labeled glucose and NaHCO3 into the cellular amino acids.

MATERIALS AND METHODS
Culture Cpndjtjpn
vineland.tl (ATCC 91014.) was maintained on
agar slants. Experimental cells were grown in a medium
containing: K2HPOj4., 0.8 g.; KH2POk, 0.2 g.;
MgSOz. 7H20, 0.2 g.; CaSO. . 2H20, 0.1 g.; NHj4.C1, 0.2 g.;
NHNO3, 0.3 g.; 1.0 cc of' Mo, Fe trace element solution
(No 03, 7.5 mg.; FeSOj., i 7H20, 299.0 mg.; water added to
two volumes of 50 cc.); glucose 12 g.; all dissolved in
one liter of water. Incubation was carried out aerobi-
cally on a rotary shaker at 28° C for 16 hours at which
time the culture was in the first third of the exponential
phase. The cells were harvested by centrifugation and
resuspended in growth medium to a prescribed concentration
prior to experimentation (excluding glucose).
Radiochemical Substrates
The radiochemical substrates used in the present
work were Glucose-1,-2, or 6-Cfl obtained from the National
Bureau of Standards. Glucose-3-C was kindly furnished
by William Sacks of Rockland State Hospital, Orangeburg,
New York.

Glucose-3,k-C was
from rats injected, with Na:
of Wood, Libson and Lorber
6-Ca was prepared in this
Li..
prepared from liver glycogen
according to the method
(28). Gluconate-2,-3,-3,Li or
laboratory from the corre-
spondingly labeled glucose samples according to the
method Oi Moore and Lirk (9). Gluconate-l-C was
obtained from Nuclear Chicago Corporation.
Rpdjprejrpnietrje Exneriments
The radiorespirometric studies on the utilization
of glucose and gluconate by proliferating cells were
carried out according to the method of Wang and Krackov
(26). At the termination of each of the experiments,
the cells and incubation media were separated by centri-
fugation and processed for the analysis of radioactivity
therein.
IncprlDpration ExDerjmpnt$
These experiments were designed to trace the
individual carbon atoms of glucose in biosynthetic
functions. To 62 cc of a coU suspension, containing
50 mg (dry wt.) of cells, were added a given amount of
specifically labeled glucose (Table II). The culture was
incubated at 28° C in the manner similar to that used under

5
radioresplron!etric experiments. Utilization of the
respective substrates was followed by examining the
iLl. rate of respiratory C 02 production. Upon termina-
tion of each of the experiments the cells were
harvested, washed, and dried over P205 j vacuo.
In a separate experiment the incorporation of
metabolic CO2 into cellular constituents was examined
by the use of NaHCO3 (2OO''c; specific activity
233 /(c/mg) as co-substrate to unlabeled glucose in an
enclosed system. After four hours of incubation the
flask was chilled to 00C, the p11 of the media adjusted
to 5 and the residual CO2 recovered from the system. Thereafter, the cells were processed as described under
Radiorespirometric Experiments.
The cell samples obtained from the incorporation
experiments were hydrolyzed individually with 20% 11Cl
(in sealed tubes) at C for 19 hours. The hydroly-
sates were evaporated to dryness j and taken up
in a defined amount of water. These hydrolysate
solutions were then chromatograhed on large sheets of
Schleicher and Schuell 14.70-A chromatograph paper employing
several solvent systems.

6
The solvent, 80% phuo1-water, was used for the
separctio. of alauinc, aspartic acid and glutamlo acid
(is). TI-ic a1mine fraction which wai slightly contami-
nated waE; purified by paper ohroruatography using
2butano1-3 &q, (15). The aspartic acid arid gluta-
mie acid fractions were individually purified with
butariol:acetic acid:water, O:1O :50 paper chromatography.
Specific activities of' each of the amino acids were
calculated from the radioactivity counting data using
liquid scintillation counting and the chemical analysis
according to the method of lemm and cocking (29).
For degradation studies each of the amino acids
was diluted with carrier to a suitable quantity.
Dezradation Procedures
lnthe:
The distribution of C in the samples of cellular
alanine was determined by means of the following chemical
operations (A) The total radiochemical assay of alanine
was carried out according to the combustion method
described by Schöniger (8); (B) The ninhydrin decarboxy-
latiori reaction (23) was used to convert C-1 of alanine to
CO2 for radiochemical assay; (C) C-2 and C-3 of alanine

7
were trapped j. HS03 as acetaldohyde, the product of
the nirthydrin reaction, and further degraded by means
of the iodoforni reaction (16) forming CHI from C-3,
CHI3 was combusted. to CO2 by the Van S1ykeFo1ch wet
combustion as described by Calvin (2); (D) The
radioactivity of C-2 was calculated by difference.
Determination Rpdjopctvjty
The respiratory CO2 obtained in radiorespiro-
metric experiments was absorbed in 10 cc of ethanolic
0.25 ?4 hyamne hydroxide [(p-diisobutylcresoxyethyl)
dimethylbenzyl ammonium hydroxide] and placed lU a
radiorespirometric CO2 trap (26). The trap solution
was replaced hourly and its radioactivity assayed by
liquid scintillation counting. Usually 5 cc aliquote
of the trapped solution were mixed with 10 cc of
toluene containing terphenyl (3g/l.) and l,14-bis-2
(5-phenyloxazolyl)-benzene (30 mg./l), in a 20 cc glass
counting vial. Countings were carried out with a
tri-carb liquid scintillation counter in the manner
described by Wang (26). The radioactivity of the cell
samples was also counted by means of liquid scintillation
counting as described by White and Helf (27). The
efficiency of radioactivity detectim for each type of

[;:1
counting sample was determined by the use of internal
standards.
The samples obtained in the degradation
experiments were converted into BaCO and mounted on
aluminum planchets by means of the centrifugation
technique (6). Determination of radioactivity was
carried out on a Geiger-Muller counter. Corrections
of counting data for background and self-absorption
were applied in the usual manner. Ali counting operations
were carried out to a relative standard deviation no
greater than 2%.

RESt'LTS
The radiorespirometric data on the utilization of
specifically labeled glucose and gluconate by
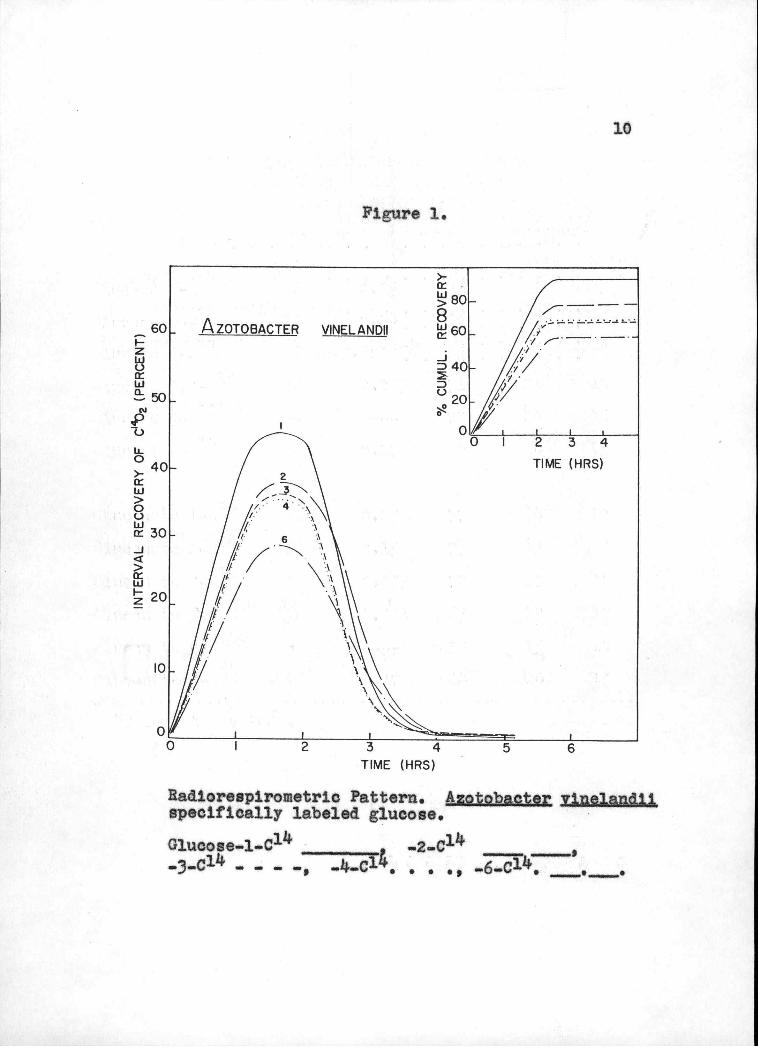
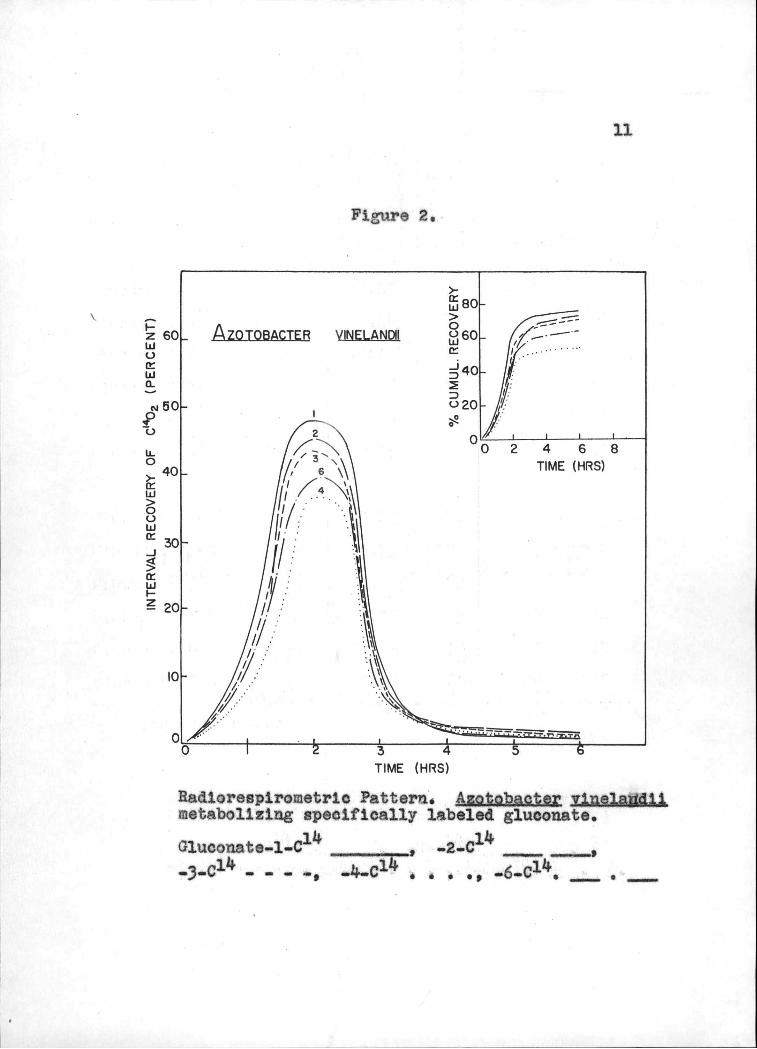
vinelandil are given in Figuren I and 2 respectively.
The experimental conditions as well as the distribution
of the substrate activity in cells, i.ncubation media and
respiratory CO2 at the end of the respective radio-
respirometric experiment are given in Table I.
In Table II is given the essential findings
obtained in the incorporation experiment including the
isotope distribution of cellular alanine samples derived
from the respective labeled substrates.
H z w C)
w ;5o N
o
4o
w > o o
3o -J
> n: w H z
Elli
lo
Fjure 1.
__80L . AZOTOBACTER VNELANDII 60 . //
4O- /ì'Ï
.
»\ - 7 \ TIME (HRS)
f 2
I iì
.
/i: 6 //
/ì2
/1/ II
Ì 5
TIME (HRS)
Radlorespi.rometric Pattern. Azotobacter vthelandii. specifically labeled glucose.
Glucosel-C _________ -2..0 _____ ____ 3-C11 _ _ - -, .J4-C1.
, _6_Cl'.
I- z w o w Q-
('J
o
IL o >-
w > o o w
-J
w H z
u
FIgure 2,
>-
w -
60 AZOTOBACTER VNELANDII
50 -
_\ 02:
' I t
I/::::::N\ o 2 4 6 8
40 - 1/e'
6 \\\\ TIME (HRS)
II //i:
30-
/,y.:
20-
/ Io- ii
ooJI TIME (HRS)
Radlorespirornetric Pattern. Aztpbacer vne].adl1 netabo11zing specifically labeled gluconate.
G1uconato_1_C14 ________ _2_C14 ____ (I1l h r1 ¿ n14 - _ - .-, --r-%.. I S I L)'.l $

12
Table I
Utilization of C- Labeled Substrate by Azotobacter viaelandii
Subsi.rate % Recovçry of C1
Amount mg. CO2 Cells Mediwn
G1ucose-1-C 16 0.25 93 5 3
G1ucose_2_C14 16 O.21 80 18 6
Giucose-3-C 16 9.8x103 69 22 8
Glucose _3,L4,_C1 16 0.063 71 21 12
Glucose-4-C1 -- --- 73 20 16
Glucose 6_C1k 16 0.27 59 2
Gluconate_l_C1 16 0.23 77 1 10
Giuconate_2_Chl 16 0.15 73 13 8
G1uoonate_3_Chi 16 0.055 72 23 10
Giuconate_3,L_C1 16 0.081 63 19 12
G1uconate4_C1* -- --- 5 1l,
G1uconate_6_C1 16 0.17 6i 24 15
*Clculated values.

Table II
Incorporation of Glucose-Ca NaHC'O3 into Azotobacter vinelandil
Substrate G1ucose-l-C- Glucose_2Cl Glucose-6Cl C1Ll.02
Glucose mg 125 125 125 125 G1ucoseA4- e lo 10 10 NaECO31'+'&c 200 Initial cell
(dry) wt. mg 50 50 50 50 % increase of cell mass 23 17 19 32
Specific Activity*
Fies, CO2 (DPM x i7) 1,93 1.53 1.Li5 32,5 Alanine DPM x 1O1'/miM 1.21 1.81 1.i6 1.81 Aspartic Acid DPN x
106/mM 0.27 1,79 1.12 5.91 Gluttnjo Acid DPM x
106/mN 0.12 1.91 0.66 5.09
Alanjue Carbon Distribution (1) COOM 98 11 32 57 (2) CFINH2 1 85 16 31 (3) CH3 1 52 12 *The amino acid specific activity represents the label in isolated cellular amino acid.
I-J
A)

DIS CUSS ION
Prevous1y, enzymological findings wore presented
by Wilson ., (19, 20, 21) to demonstrate the
operation of the TCA cycle in cell-free extracts of
vinelandii. These extrct oxUized pyruvic acid,
acetic acid. and the acids of the TCA cycle. Upon
addition of acetate_1_C1, radio carbon was rapidly
incorporated into the cellular TCA cycle intermediates.
Radiorespirometric experiments with proliferating cells
of vinelandti fed specifically labeled pyruvate and
acetate have added further proof that these substrates
are routed through the TCA cycle to CO2. With respect
to this information lt is desirable to elucidate the
pathways involved in the production of pyruvate from
substrate glucose. Previous work with cell-free extracts
by Wilson L l) suggested. strongly the
occurrence of the ED pathway and the PP pathway. The
operation of the EMP pathway was uncertain since these
workers failed to detect the presence of phosphohexo-
kinase. However, recent work of Sobek and Clifton (17)
with intact cells tends to support the concurrent
operation of the E1P and the PP pathways for glucose
utilization. Inasmuch as glucose can be catabolizec3. by

15
either the ED the EMP pathway giving rise to
un.ts, and the fact that the concurrent operation of
these two pathways in a given organism has never been
demonstrated, one is therefore interested in which
one of these two pathways is operating in vinelandli. Much is learned by an examination of the radiorespiro-
metric data for the utilization of glucose (Figure 1 and
Table I) by vin,elandii. The C1Z4O2 production patterns
for individual carbon atoms of glucose are basically
similar to those observed with Pseudotnonas fluorescens
KBl (18). There is strong evidence that the latter
organism relies on the concurrent operation of the PP
and ED pathways for the utilization of glucose and
gluconate. In comparing the findings with these two
organisris, a noticeable difference is that with
L2 vinelandU the conversion of C-Lf of glucose to CO2
is significantly less than that of C-2. Nevertheless,
the radiorespirometric data (Figure 1) can be interpreted
to indicate that: (A) the ED and PP pathways are playing
important roles in glucose catabolism; (B) the EM? pathway,
if present, does not contribute much to the overall utili-
zation of glucose. The latter conclusion is drawn from
the fact that a C_3 and Ck of glucose were converted to
CO2 to a baser extent as compared to that of C-2 and

16
particularly C-1. This observation is not in line with
the degradation of glucose via the EMP-TCA sequence
which calls for the preferential oxidation of C3 and C13 of glucose to CO2.
The foregoing analysis is further and more
clearly supported by the radiorespirornetric data on
gluconate utilization (Figure 2, Table I). Here again, the conversion of either C3 or C-11 of gluconate to CO2
is lower in rate and extent as compared to that of C-].
arid C2. A striking observation in the gluconate
experiments is the extraordinary low yield of CO2 from
C-J4 of gluconate even in comparison with C-6. This fact
cannot be explained on the basis of the PP scheme or the
EDTCA route. Some other mechanism must be operative to
account for this finding which may involve either a
preferential combustion of C-6 to CO2 or the retarded
combustion of C-k to CO2.
To elucidate further the mechanism of glucose and
gluconate assimilation in proliferating cells of
vinelandil, the results obtained in the incorporation experiment are helpful. These experiments were designed
to trace the fate of the individual carbon atoms of
glucose in biosynthetic functions, particularly those
responsible for the formation of the carbon skeleton of

'7
key amino acids such as alanine, aspartic acid and
glutamlo acid. An understanding of the or1gin of the
carbon skeletons of these amino acids, which are closely
related to the TCA intermediates, should also provide
information on the nature of the primary pathways of
carbohydrate breakdown (25). This is particularly
true when studying the metabolism of microorganisms
which do not excrete detectable amounts of carbohydrate
intermediates into the incubation medium. To account
for the res-entry of metabolic CO2 into the skeleton of
these amino acids, a separate experiment was carried
out in which unlabeled glucose and NaHCO3 were used
as co-substrates. Insofar as the specific activities
of these amino acidsare concerned, one finds that the
extent of labeling from C-1, C-2, or C-6 of glucose,
in alanine is approximately the same in magnitude.
However, preferential incorporation of C2 and C-6 of
glucose over that of C-1 into either aspartic acid or
glutamic acid was noted. This latter fact is not in
line with the operation of the EP pathway but is
compatible with the operation of the ED scheme for glucose
catabolism. This is true since the precursor of alanine,
pyruvic acid, is one of the key intermediates of glucose

degradation. Prior to the analysis of these patterns,
it was recognized that proliferating cells are capable
of incorporating metabolic CO2 into cellular const-
tuents. This was evidenced by the observed labeling
of amino acids from adnilnistered NaHC14O3 (Table II).
Consequently, the isotopic distribution pattern of
cellular alanine samples, corrected for the re-entry
of metabolic CO2 given in Table III, is much more
meaningful. The corrections were made from the iso-
topic distribution pattern of cellular alanine observed
In the NaHCO3 experiment as described by Wang and
:eaa (25).
The exclusive role played by the ED pathway in
the conversion of glucose to pyruvate is Indicated by
analysis of these isotopic distribution patterns. Thus,
C-1 of glucose was found to be exclusively incorporated
into C-1 of alanine and C-2 of glucose was heavily
incorporated into C-2 of alanlne. The anticipated isotopic
distribution pattern of alanine, or pyruvate, from a
number of possible pathways is summarized in Table III.
The minor labeling of C-1 and C-3 of alanine from C-2
of glucose may reflect the randomization of the glucose
skeleton via the operation of the PP pathway (i, 3).

19 Table III
The Isotopic Distribution Pattern of Cellular Alanine
Pathway Followed by the Expected and Observed Labeled Carbon Atoms Isotopic Distribution
Pattern %
G1ucose-l-C COOH CH(NH2) CH3
Via ED route lOO 0 0
Via EMP route O O loo
Via PP rQute O O O Observed* loo O O
Glucose_2_C14
Via ED route o loo O
Via EMP route 0 lOO O
VIa PP and ED routes (a) 66 o 33 Via PP and EMP routes (a) 33 O 66 Observed* lo 86
Gluco s_6_C
Via ED route o O 100 Via EMP route o O 100 Via liC and ED routes (b) 50 0 50
Via BC and ENP routes (b) O O 100 Observed* 31 16 53
'J 02
Via CO2 fixation (C3 plus C1) lOO 0 0
Observed 57 31 12
a) PP pathway cycles to reform hexose phosphate which is again cleaved to yield triose (i, 26).
b) Triose may be supplied from ED, PP, or EMP pathway. The Hexose Cycle (HC) wifl result in the formation of hoxose phosphate which may then be cleaved as shown.
* Corrected for the re-entry of metabolic C1L102.

ç'
The observed isotopic dlstrlbuti.on. of alanine in the
g1uoose_6-C1' experiment provides additional lnfor-.
matlon on the glucose catabolism of this organism.
The heavy labeling of C-3 of alanine (52% of the total
ratio in the alanine) is in line with the operation of
the ED mechanism. It is surprising to note that signi-
ficant labeling also ocoured. at C-2 and particularly
C-1 of this atnino acia. As shown in Table III, this
finding may have been the consequence of the operation
of the Hexose Cycle (FTC) pathway. Such a pathway
involves the isomerization of G3P to dihydroxyacetone phosphate, condensation of these trioses to fructose
l,6-.diphosphate, followed by dephosphorylization to
fructose 6-P and isornerization to glucose 6-P. This
has been demonstrated by Hochster and Katznelson
(28, -29). It is recalled that earlier Wilson
failed to detect the presence of phosphohexokinase in
this organism. This may account for the observed
operation of the HC in this organism (28). The findings
given in the present work indicate strongly that glucose
in vinelandii is primarily metabolized via the ED
pathway. The extent of participation of the ED pathway
in overall glucose catabolism cannot be quantitatively

21
ascertained without additional information. The method
of estimation previously given in connection with the
studies on Pseudornonas cannot be directly applied to
the present case. This is true since the occurrence
of the NC pathway results in the shifting of a fraction
of the C-4 of glucose to the C-3 position, thereby
reducing the extent of conversion of this carbon atom to
co2. This consideration implies that at the end of the
radiorespirometric experiment the fraction of C..Lf of
glucose detected in CO2 should be greater than that
given in Table I (i.e., 73% of the total radioactivity
in the glucose molecule). Therefore, it can be
concluded that the operation of the PP pathway of this
organism should not exceed 20% of the total catabolized
glucose, I.e., yielded in the glucose_1_C1U1
experiment minus CO2 yield in the g1ucose_1_C1 experiment (calculated) (25).

, ' '-
B IBLIOGRAPHY
1. Beevers, H, Intermediates of the pentose phosphate pathway es respiretory substrates. Plant Physiology 31 :3393k7, 1956.
2. Calvin, Nelvth. ]. Isotopic Carbon. New York, John Wiley, 199. 93 p.
:3. Dawos, E. A, and W. H, Ha13. On the Quantitative evaluation of routes of glucose metabolism by the use of radioactive glucose. Biochimica et Biophysica Acta 21:1711._175, 1960.
¿j, Entner, Nathan and Michael Doudoroff. Glucose and gluconic acid oxidation of Pseudomonas saccharobt1a. Journal of Biological Chemistry 196:853-862. 1952.
.5. Hochster, R. M. and H. Katznelson. On the mechanism of glucose-6-phosphate oxidation in cell-free extracts of Xanthpmonas Dhaseoli (XP8). Canadian Journal of Biochemistry and Physiology 36:669-689. 1958.
6. Hutchens, Tyra, Cecil K, Claycomb, Wi11ian J, Cathey anì John T, Van Bruggen. Techniques in the use of Cl as a tracer II. Preparation of BaCO3 plates by centrifugation. Nuoleonics 7:Li1_LiLi, 1950.
7. Johnson, Emmett J., J. M. Sobek and C. E. Clifton. Oxidative assimilation by Azotobacter azilis. Journal of Bacteriology 76:658-661. 1958.
8. Kelly, R. C., E, A, Peets, S. Gordon and D, A. Buyske. Determination of Cl'4 and 113 in biological samples by Schonl.ger combustion and liquid scintillation techniques. Analytical Biochemistry 2:267-273. 1961.
9. Moore, Standford and Karl P. Link. Carbohydrate Characterization,I, The oxidation of aldoses by hypolodite in methanol, II. The identification of seven aldomonosacoharides as benzimidazole derivatives. Journal of Biological Chemistry 133:293-311. l9I10,

23
10. Nortenson, L. E. and P, W. Wilson. Metabolism of r1bose-5-phophate by Azotobgoter vine1andi.. Journal of Biological Chemistry 213:713-721. 1955.
li. Nortenson, L. E., P. B. Hamilton and P. W. Wilson. Dissimilation of 6-phosphogluconate by Azotobacter v1rxelruili. Biochimica et Biophysica Acta 16: 238-24h. 1955.
12. Mortenson, L. E. and P. W, Wilson, Initial steps in breakdown of glucose by the Azotobacter. Bacteriological Proceedings of the 514th General Neeting of the Society of American Bacteriologists 54:108. 195L$.
13. Mortenson, L, E, and P. W. Wilson. Initial stages Iti the breakdown of carbohydrates by the Azotobacter vinelandlì. Archives of Biochemistry and Biophysics 53:)+25-1435, l95.
Racker, Efrain, Alternate pathways of glucose and fructose metabolism. Advances in Enzymology 15: 141-178. l95.
15. Roland, J. F. Jr., and A. N. Gross. Quantitative determination of amino acids using monodiniensional paper chromatography. Analytical Chemistry 26: 502-505. l95.
16. Seelye, R. N. and T. A, Turney. The lodoform reaction. Journal of Chemical Education 36:572_5714., 1959.
17. Sobek, J. N, nd C, F, Clifton. Oxidative assirni- lation and 1+ distribution in Azotobacter agilts. Proceedings of the Society for Experimental Biology and Medicine 109:k09-kfl. 1962,
18. Stern, Ivan J., C, H. Wang and C. M, Gilmour, Comparative catabolism of carbohydrates in Pseudomonas species. Journal of Bacteriology 79:601-611. 1960.
19. Stone, R, W, and P, W. Wilson, The effect of oxalacetate on the oxidation of suocinate by Azotobacter extracts. Journal of Bacteriology 63: 619-622. 1952.

21
20. Stone, R. W. and P. W. Wilson. Incorporation of acetate in acids of the citric acid cycle by Azptobpcter extracts. Journal of Biological Chemistry 196:221-225. 1952.
21. Stone, R. W and P. w. Wi1on, Respiratory activity of cell-free extracts from Azotobacter. Journal of Bacteriology 63:605-617. 1952.
22. Vandanis, A. and R. M. Hochster. On the mechanism of glucose metabolism In the plant tumor-inducing organism Arobacterium tumefaciens. Canadian Journal of Biochemistry and Physiology 39:1165-1182. 1961.
23. Van Slyke, Donald D., Douglas A. MacFad.yen and Paul B, Hamilton. The gasometric determination of amino acids in urine by the ninhydrin-carbon dioxide method. Journal of Biological Chemistry 150:251-258, 193.
2L1. Wang, Chih H. Metabolism studies by radiorespiro- metry, in: Advances in Tracer Methodology, ed.
by Seymour Rothchi].d, Vol. 1. Plenuni Press, New York, 1963. p. 27L1_290.
25. Wang, C. H,, and G. J. Ikeda. Biosynthesis of C acids in Pseudomonas fLuorescens KB1. Biochemical Journal 79:61k-620. 1961.
26. Wang, Chih H,, and Julia K. Krackov. The catabolic fate of glucose in Bacillus subtilis. Journal of'
Biological Chemistry 237:361k-3622. 1962.
27. White, C. G.,and Samuel Helf. Suspension counting in scintillation gels. Nuoleonios lk:k6-k8. 1956.
28. Wood, H, G., N. Lifson and V. 1
of fixed carbon in glucose from Journal of Biologloal Chemistry
29. Yemm, E. W,, and E. C. Cocking. of amino acids with ninhydrin. 1955.
rber. rphe position rat liver glycogen. l59:k75-k89, l9Ll.5.
The determination Analyst 80:209-213,





























