Embed Size (px)
Citation preview

EBS7 is a plant-specific component of a highlyconserved endoplasmic reticulum-associateddegradation system in ArabidopsisYidan Liua, Congcong Zhangb, Dinghe Wangb, Wei Sua,1, Linchuan Liub, Muyang Wangb, and Jianming Lia,b,2
aDepartment of Molecular, Cellular, and Developmental Biology, University of Michigan, Ann Arbor, MI 48109; and bShanghai Center for Plant StressBiology, Shanghai Institutes for Biological Sciences, The Chinese Academy of Sciences, Shanghai 201602, China
Edited by Natasha V. Raikhel, Center for Plant Cell Biology, Riverside, CA, and approved August 12, 2015 (received for review June 19, 2015)
Endoplasmic reticulum (ER)-associated degradation (ERAD) is anessential part of an ER-localized protein quality-control system foreliminating terminally misfolded proteins. Recent studies havedemonstrated that the ERAD machinery is conserved among yeast,animals, and plants; however, it remains unknown if the plant ERADsystem involves plant-specific components. Here we report that theArabidopsis ethyl methanesulfonate-mutagenized brassinosteroid-insensitive 1 suppressor 7 (EBS7) gene encodes an ER membrane-localized ERAD component that is highly conserved in land plants.Loss-of-function ebs7 mutations prevent ERAD of brassinosteroidinsensitive 1-9 (bri1-9) and bri1-5, two ER-retained mutant variantsof the cell-surface receptor for brassinosteroids (BRs). As a result,the two mutant receptors accumulate in the ER and consequentlyleak to the plasma membrane, resulting in the restoration of BRsensitivity and phenotypic suppression of the bri1-9 and bri1-5 mu-tants. EBS7 accumulates under ER stress, and its mutations lead tohypersensitivity to ER and salt stresses. EBS7 interacts with the ERmembrane-anchored ubiquitin ligase Arabidopsis thaliana HMG-CoA reductase degradation 1a (AtHrd1a), one of the central compo-nents of the Arabidopsis ERAD machinery, and an ebs7 mutationdestabilizes AtHrd1a to reduce polyubiquitination of bri1-9. Takentogether, our results uncover a plant-specific component of a plantERAD pathway and also suggest its likely biochemical function.
brassinosteroid receptor BRI1 | ERAD | EMS-mutagenized bri1 suppressor |ubiquitin ligase E3 | unfolded protein response
Endoplasmic reticulum (ER)-associated degradation (ERAD)is an integral part of an ER-mediated protein quality-control
system in eukaryotes, which permits export of only correctly foldedproteins but retains misfolded proteins in the ER for repair viaadditional folding attempts or removal through ERAD. Geneticand biochemical studies in yeast and mammalian cells have revealedthat the core ERAD machinery is highly conserved between yeastand mammals and that ERAD involves four tightly coupled steps:substrate selection, retrotranslocation through the ER membrane,ubiquitination, and proteasome-mediated degradation (1, 2).Because the great majority of secretory/membrane proteins are
glycosylated in the ER, diversion of most ERAD substrates fromtheir futile folding cycles into ERAD is initiated through pro-gressive mannose trimming of their asparagine-linked glycans(N-glycans) by ER/Golgi-localized class I mannosidases, includinghomologous to α-mannosidase 1 (Htm1) and its mammalianhomologs ER degradation-enhancing α-mannosidase-like proteins(EDEMs) (3). The processed glycoproteins are captured by two ERresident proteins, yeast amplified in osteosarcoma 9 (OS9 inmammals) homolog (Yos9) and HMG-CoA reductase degradation3 (Hrd3) [suppressor/enhancer of Lin-12–like (SEL1L) in mam-mals], which recognize the mannose-trimmed N-glycans and sur-face-exposed hydrophobic amino acid residues, respectively (4, 5).The selected ERAD clients are delivered to an ER membrane-anchored ubiquitin ligase (E3), which is the core component of theERADmachinery (6), for polyubiquitination. Yeast has two knownERAD E3 ligases, Hrd1 and degradation of alpha 10 (Doa10),both containing a catalytically active RING finger domain, whereas
mammals have a large collection of ER membrane-anchored E3ligases, including Hrd1 and gp78 (7). The yeast Hrd1/Doa10-con-taining ERAD complexes target different substrates, with theformer ubiquitinating substrates with misfolded transmembrane orluminal domains and the latter acting on clients with cytosolicstructural lesions (8).Because of the cytosolic location of the E3′s catalytic domain and
proteasome, all ERAD substrates must retrotranslocate throughthe ER membrane. It is well known that the retrotranslocation stepis tightly coupled with substrate ubiquitination and is powered by anAAA-type ATPase, cell division cycle 48 (Cdc48) in yeast and p97in mammals. However, the true identity of the retrotranslocon re-mains controversial. Earlier studies implicated the secretory 61(Sec61) translocon, degradation in the endoplasmic reticulum 1(Der1) [Der1-like proteins (Derlins) in mammals], and Hrd1 in ret-rotranslocating ERAD substrates (9). After retrotranslocation, ubiq-uitinated ERAD clients are delivered to the cytosolic proteasomewith the help of Cdc48/p97 and their associated factors for proteolysis(10). In addition to the above-mentioned proteins, the yeast/mam-malian ERAD systems contain several other components, includingseveral ubiquitin-conjugating enzymes (E2), a membrane-anchoredE2-recruiting factor, Cue1 that has no mammalian homolog, a scaf-fold protein U1-Snp1–associating 1 (Usa1) [homocysteine-inducedER protein (HERP) in mammals] of the E3 ligases, and amembrane-anchored Cdc48-recruiting factor, Ubx2 (Ubxd8 inmammals) (6).
Significance
Endoplasmic reticulum (ER)-associated degradation (ERAD) is awell-studied cellular process in yeast and mammalian systems.Recent molecular and genetic studies in the reference plantArabidopsis have revealed that ERAD also is conserved in plants.Here we report that an Arabidopsis ERAD process for degradingmisfolded/mutant receptor-like kinases requires a plant-specificprotein, ethyl methanesulfonate-mutagenized brassinosteroid-insensitive 1 suppressor 7 (EBS7), that is localized to the ERmembrane and is induced by ER stress. Our biochemical studiessuggest that EBS7 functions as a key regulator of this Arabi-dopsis ERAD process by maintaining the protein stability of itscore component, a membrane-anchored E3 ligase, Arabidopsisthaliana HMG-CoA reductase degradation 1a (AtHrd1a).
Author contributions: Y.L. and J.L. designed research; Y.L., C.Z., D.W., W.S., and L.L. per-formed research; D.W., W.S., and M.W. contributed new reagents/analytic tools; Y.L.,C.Z., D.W., L.L., M.W., and J.L. analyzed data; and Y.L., C.Z., and J.L. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Freely available online through the PNAS open access option.1Present address: The State Key Laboratory of Genetic Engineering and Institute of PlantBiology, School of Life Sciences, Fudan University, Shanghai 200433, China.
2To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1511724112/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1511724112 PNAS | September 29, 2015 | vol. 112 | no. 39 | 12205–12210
PLANTBIOLO
GY

For many years ERAD has been known to operate in plants (11),but the research on the plant ERAD pathway lagged far behindsimilar studies in yeast and mammalian systems. Recent molecularand genetic studies in the reference plant Arabidopsis, especially twoArabidopsis dwarf mutants, brassinosteroid-insensitive 1-5 (bri1-5)and bri1-9, carrying ER-retained mutant variants of the brassinos-teroid receptor (BR) BRASSINOSTEROID-INSENSITIVE 1(BRI1) (12–14), revealed that the ERAD system also is conservedin plants (reviewed in refs. 15 and 16). For example, the ERADN-glycan signal to mark misfolded glycoproteins in Arabidopsis wasfound to be the same as that in yeast/mammalian cells (17, 18). Bothforward and reverse genetic studies have shown that Arabidopsishomologs of the yeast/mammalian ERAD components, includingYos9/OS9 (19, 20), Hrd3/Sel1L (21, 22), Hrd1 (21), EDEMs (23),and a membrane-anchored E2 (24), are involved in degradingmisfolded glycoproteins. However, it remains unknown if the plantERAD requires one or more plant-specific components to degradeterminally misfolded proteins efficiently. In this study, we took aforward genetic approach to identify a novel Arabidopsis ERADmutant, ethyl methanesulfonate-mutagenized bri1 suppressor 7 (ebs7),and subsequently cloned the corresponding EBS7 gene. We dis-covered that EBS7 encodes an ER-localized membrane protein thatis highly conserved in land plants but lacks a homolog in yeast ormammals. Our biochemical studies strongly suggested that EBS7plays a key role in an Arabidopsis ERAD process by regulating theprotein stability of the Arabidopsis thaliana HRD1a (AtHrd1a).
ResultsThe ebs7-1 Mutation Restores BR Sensitivity to bri1-9 by BlockingDegradation of Its Mutant BR Receptor. We previously showed thatthe dwarf phenotype of a BR-insensitive mutant, bri1-9, was causedby ER retention and subsequent ERAD of a structurally defectivebut biochemically competent BR receptor, bri1-9 (12, 13, 17). Agenetic screen for extragenic bri1-9 suppressors coupled with asecondary screen for bri1-9–accumulating mutants revealed aconserved ERAD N-glycan signal and also identified several con-served components of an Arabidopsis ERAD system (15, 19, 21).This study is focused on a new mutant, ebs7-1. As shown in Fig. 1A,ebs7-1 bri1-9 accumulated a higher level of bri1-9 than the parentalbri1-9 mutant (Fig. 1A). A simple cycloheximide (CHX)-chase ex-periment revealed that the observed increase in bri1-9 abundancewas caused largely by attenuated degradation rather than by in-creased biosynthesis of bri1-9 (Fig. 1B). Similar to observations inother known Arabidopsis ERAD mutants (17–19, 21), the accu-mulation of bri1-9 in the ER likely saturates its ER retention sys-tems, leading to detection of a small percentage of bri1-9 proteinscarrying the complex type (CT) N-glycans (indicative of ER es-cape) that are resistant to hydrolysis by endoglycosidase H(Endo H), which is capable of cleaving high-mannose-type (HM)N-glycans of ER-localized proteins but not Golgi-processed CTN-glycans (Fig. 1A). Consistent with the Endo H result, a con-focal microscopic analysis of the root fluorescent signals of theGFP-tagged bri1-9 revealed increased bri1-9–GFP abundanceand enhanced fluorescent signal on the plasma membrane (PM)in a pBRI1:bri1-9–GFP/bri1-9 ebs7-1 transgenic line as com-pared with a pBRI1:bri1-9–GFP/bri1-9 EBS7+ line generatedby crossing the pBRI1:bri1-9-GFP/bri1-9 ebs7-1 line with bri1-9(Fig. S1 A and B).Morphologically, ebs7-1 bri1-9 has a larger rosette with much
bigger rosette leaves than bri1-9 (Fig. 1C). The double mutant alsohas a longer hypocotyl when grown in the dark and taller in-florescence stems at maturity than bri1-9 (Fig. 1D andE). Consistentwith the detection of a small pool of ER-escaping bri1-9, ebs7-1 bri1-9partially regains the BR sensitivity. As shown in Fig. 1F, exogenouslyapplied brassinolide (BL) inhibited the root growth of wild-type andebs7-1 bri1-9 plants in a dose-dependent manner but had a muchweaker impact (especially at concentrations <10 nM) on bri1-9roots. The regained BR sensitivity in ebs7-1 bri1-9 was con-firmed by a widely used biochemical assay that measured theBR-induced dephosphorylation of bri1-EMS-suppressor 1 (BES1),a key transcription factor of BR signaling (25). Fig. 1G shows that
BL had a marginal effect on the BES1 phosphorylation status inbri1-9 but efficiently and partially dephosphorylated BES1 in wild-type and ebs7-1 bri1-9 seedlings, respectively. Importantly, over-expression of EBS2, a limiting component of the bri1-9’s ER-retention systems (12), fully retained bri1-9 in the ER (Fig. S1D)and nullified the suppressor activity of ebs7-1 on bri1-9 (Fig. S1C).
ebs7-1 Inhibits the Degradation of Other ERAD Substrates. To de-termine if ebs7-1 inhibits the degradation of other known Ara-bidopsis ERAD clients, we first crossed ebs7-1 into bri1-5 thatcarries a different ER-retained mutant BR receptor, bri1-5 (14).As shown in Fig. 2A, ebs7-1 suppressed the dwarfism of bri1-5(Fig. 2 A and B and Fig. S2A), restored its BR sensitivity (Fig. 2C and D), and greatly elevated the bri1-5 level (Fig. 2E). Thecrucial role of EBS7 in ERAD of bri1-5 was confirmed further bythe identification of two allelic ebs7 mutants in an independentgenetic screen for ERAD mutants of bri1-5 (Fig. S2 B and C).We also examined the effect of ebs7-1 on ERAD of the ER-
retained conformer of EF-Tu receptor (EFR), an Arabidopsis PM-localized immunity receptor that recognizes and binds the bacterialtranslation elongation factor EF-Tu (26). Previous studies demon-strated that the proper folding of EFR requires an Arabidopsis ERquality-control system consisting of EBS1 and EBS2 [also known as“priority in sweet life2 (PSL2)/elf18-insensitive21” (PSL2/ELFIN21)
bri1-9ebs7-1 bri1-9
- + - + - +
bri1-9HMbri1-9CT
Endo H
RbcS
WT
ebs7-1 bri1-9
bri1-9 WT
ebs7-1 bri1-9
bri1-9 WT
ebs7-1 bri1-9 bri1-9 WT
WT
P-BES1 BES1RbcS
BLbri1-9
ebs7-1 bri1-9
- + - + - +
CHX(h)
bri1
-9
ebs7
-1
bri1
-9
bri1-9
bri1-9RbcS
RbcS
0 3 6 12 16 24
0 1 2 3 4 5 6 7
WT
bri1-9 ebs7-1 bri1-9
bri1-9 WT
100 80 60 40 20
120
Roo
t Inh
ibiti
on %
0 10-3 10-2 10-1 1 101 102 103
BL Concentration (nM)
0
A
B
C
D
E
F
G
Fig. 1. The ebs7-1mutation suppresses bri1-9 and restores its BR sensitivity byblocking bri1-9 degradation. (A) Immunoblot analysis of BRI1/bri1-9. Totalproteins of 2-wk-old seedlings were treated with or without Endo H, separatedby SDS/PAGE, and analyzed by immunoblot with a BRI1 antibody. bri1-9HM andbri1-9CT denote BRI1/bri1-9 proteins carrying HM and CT N-glycans, respec-tively. (B) Immunoblot analysis of the bri1-9 stability. Two-week-old seedlingswere treated with 180 μM CHX for the indicated hours, and extracted proteinswere separated by SDS/PAGE and analyzed by immunoblot with the anti-BRI1antibody. (C–E) Pictures of 4-wk-old light-grown (C) and 5-d-old dark-grown(D) seedlings, and 2-mo-old soil-grown plants (E). (F) The BR-induced root in-hibition assay. Root lengths of ∼40 7-d-old seedlings grown on BL-containing1/2 Murashiga and Skoog (MS) medium were measured, and measurementswere converted into percentage values relative to that of the same genotypegrown on medium without BL. Error bars represent ± SE of three independentassays. (G) The BR-induced BES1 dephosphorylation assay. Protein extracts of2-wk-old seedlings treated with or without 1 μM BL were separated by SDS/PAGE and analyzed by immunoblot with an anti-BES1 antibody. In A, B, and G,Coomassie blue staining of the small subunit of the ribulose-1,5-bisphosphatecarboxylase/oxygenase (RuBisCo, hereafter RbcS) enzyme on a duplicate gelwas used as a loading control.
12206 | www.pnas.org/cgi/doi/10.1073/pnas.1511724112 Liu et al.

and PSL1/ELFIN5 (27, 28), respectively] and that their loss-of-function mutations cause misfolding, ER retention, and ERAD ofEFR (29). We crossed ebs7-1 into ebs1-3 bri1-9 and analyzed theEFR abundance in both ebs1-3 bri1-9 and ebs7-1 ebs1-3 bri1-9 mu-tants using an anti-EFR antibody. As shown in Fig. 2F, although noEFR was detected in ebs1-3 bri1-9, the EFR abundance in ebs1-3ebs7-1 bri1-9mutants was similar to that in wild type, bri1-9, or ebs7-1bri1-9 plants, indicating that EBS7 also is required for degradingmisfolded EFR. Taken together, our data demonstrated that EBS7is required for the degradation of both mutant BR receptors andmisfolded EFR.
Molecular Cloning and Characterization of the EBS7 Gene. To understandhow ebs7-1 affects the Arabidopsis ERAD system, we positionallycloned the EBS7 gene. PCR-based genetic mapping delimited theEBS7 locus within an 850-kb region on chromosome 4 (Fig. 3A),which includes two candidate ERAD genes. One, At4g29330, en-codes a homolog of the yeast Der1 known to be involved in ERAD(30–32), and the other, At4g29960, is coexpressed with many known/predicted ER proteins including three ERAD genes, EBS5(At1g18260), EBS6 (At5g35080), and DERLIN-2.1 (At4g21810) (Fig.S3A) (33). We PCR-amplified and sequenced the two candidategenes from ebs7-1 bri1-9. Comparison of the resulting sequences withthe published Arabidopsis wild-type sequences detected no mutationin At4g29330 but identified a G→A mutation in At4g29960 thatcauses a missense mutation of Ala131 to Thr (Fig. 3B). Its identity asthe EBS7 gene was supported by sequencing At4g29960 in two allelicebs7 mutants (ebs7-2 and ebs7-3) discovered in an independent ge-netic screen for bri1-5 suppressors (Fig. S2 B and C). In both mu-tants, a G→A mutation was detected that changes Gly74 to Glu inebs7-2 and disrupts the correct splicing of the At4g29960 mRNA inebs7-3 (Fig. 3B). Further support for At4g29660 being the EBS7 genecame from our complementation experiment showing that a genomic
At4G29960 transgene rescued the morphological and biochemicalphenotypes of the ebs7-1 mutation (Fig. 3 C–E and Fig. S2D).
EBS7 Is a Plant-Specific Protein Conserved in Land Plants. The EBS7gene consists of five exons plus four introns (Fig. 3A), encodes apredicted polypeptide of 291 amino acids (Fig. S4A), and is ubiq-uitously expressed in Arabidopsis tissues/organs (Fig. S3B) (34).BLAST searches failed to identify any known protein motif inEBS7 or to discover EBS7 homologs in fungi or animals but didfind EBS7 homologs in land plants, including Physcomitrella andSelaginella (Fig. S4). Sequence alignment between EBS7 and itsplant homologs identified two major conserved regions, one in themiddle and the other at the C terminus (Fig. S4A). The annotatedAt4g29960 protein was predicted to contain three transmembranesegments [aramemnon.botanik.uni-koeln.de/tm_sub.ep?GeneID=31811&ModelID=0 (35)], with the first two corresponding to theconserved middle region and the third overlapping with the con-served C terminus (Fig. S4A), suggesting that EBS7 is likely amembrane protein. It is interesting that Ala131, which is mutated toThr in ebs7-1, is not conserved among the plant EBS7 homologs,whereas Gly74 mutated to Glu in ebs7-2 is absolutely conservedbetween EBS7 and its plant homologs (Fig. S4A). It is also worthmentioning that both the Ala131-to-Thr and Gly74-to-Glu muta-tions greatly reduce the abundance of ebs7 protein, likely because ofpotential misfolding of the mutant ebs7 proteins and their subsequentdegradation (Fig. 3D and Fig. S2 C and D).
EBS7 Is an ER-Localized Membrane Protein That Accumulates UnderER Stress. In line with the presence of three predicted transmembranesegments, EBS7/At4g29960 was identified previously as a putativePM protein by two independent studies (36, 37). Both studies usedthe aqueous two-phase partitioning method to enrich the PM
ebs7-1 bri1-5 bri1-5 WT
ebs7-1 bri1-5 bri1-5 WT
!"
#!"
$!"
%!"
&!"
'!!"
'#!"
'" #" (" $" )" %" *" &"
+,"
-./'0)"
()0'"
WT
100 80 60 40 20
120
Roo
t Inh
ibiti
on %
0 10-310-210-1 1 101 102 103
BL Concentration (nM)
bri1-5
0
ebs7-1 bri1-5
- +
ebs7-1 bri1-5 WT
P-BES1 BES1RbcS
BLbri1-5
- + - +
- + - + - +
bri1-5HMbri1-5CT
Endo H
RbcS
WT bri1-5ebs7-1 bri1-5
*
EFR
A
B
C
D
E
F
Fig. 2. ebs7-1 inhibits the degradation of bri1-5 and misfolded EFR. (A and B)Pictures of 4-wk-old light-grown (A) and 5-d-old dark-grown (B) seedlings. In(B), three individual images of two seedlings (separated by white lines) wereassembled together. (C) The root-growth inhibition assay of wild-type, bri1-5,and ebs7-1 bri1-5 seedlings. (D) The BES1 dephosphorylation assay. (E) Im-munoblot analysis of BRI1/bri1-5 abundance. bri1-5HM and bri1-5CT denote theBRI1/bri1-5 proteins carrying the HM and CT N-glycans, respectively. (F) Im-munoblot analysis of EFR. Protein extracts of 2-wk-old seedlings were sepa-rated by SDS/PAGE and analyzed by immunoblot with an anti-EFR antibody. InD and E, Coomassie blue staining of the RbcS band on a duplicate gel serves asa loading control, and in F, a nonspecific cross-reacting band indicated by theasterisk serves as a loading control.
A
B
C
D
E
GF
Fig. 3. EBS7 encodes an ER-localized membrane protein that is highly con-served in land plants. (A) Map-based cloning of EBS7. EBS7 was mapped to an∼850-kb region on chromosome 4. Marker names are shown above the line,and recombinant numbers are shown below the line. The EBS7 gene structureis shown with bars denoting exons and lines indicating introns. Arrowheadsshow the positions of three ebs7 mutations. (B) The nucleotide changes andpredicted molecular defects of the three ebs7 alleles. (C) Pictures of 4-wk-oldwild-type, bri1-9, and two transgenic ebs7-1 bri1-9 mutant plants. (D) Immu-noblot analysis of EBS7. Total proteins of 2-wk-old seedlings were separated bySDS/PAGE and analyzed by immunoblot using an anti-EBS7 antibody. (E) Im-munoblot analysis of BRI1/bri1-9. (F) The confocal images of tobacco leafepidermal cells transiently expressing GFP–EBS7 (Left), RFP–HDEL (Center), andsuperimposition of GFP and RFP signals (Right). (G) Immunoblot analysis ofEBS7. Total (T), membrane (M), and soluble (S) proteins extracted from 2-wk-old seedlings were separated by SDS/PAGE and analyzed by immunoblot withantibodies against EBS2, EBS5, and EBS7. In D and G, the asterisk indicates anonspecific band serving as a loading control.
Liu et al. PNAS | September 29, 2015 | vol. 112 | no. 39 | 12207
PLANTBIOLO
GY
fraction, which often is contaminated with organellar membraneproteins such as ER proteins. To determine where EBS7 localizes,we generated two GFP fusion constructs (with GFP fused at the N orC terminus of EBS7) driven by the constitutively active 35S promoterand transformed the two fusion constructs into ebs7-1 bri1-9. Only theGFP–EBS7, but not EBS7–GFP, transgene rescued the ebs7-1 mu-tation (Fig. S5A), indicating that GFP–EBS7 was physiologicallyfunctional. When GFP–EBS7 was transiently expressed in tobaccoleaf epidermal cells, its subcellular localization pattern overlappedwith that of a known ER maker, His-Asp-Glu-Leu–tagged red fluo-rescent protein (RFP–HDEL) (Fig. 3F) (38). Consistently, confocalmicroscopic analysis of the root tips of rescued GFP–EBS7/ebs7-1bri1-9 transgenic seedlings revealed colocalization of GFP–EBS7 withanother ER marker, ER Tracker Red dye (Fig. S5B). We also per-formed a subcellular fractionation experiment that separated solubleproteins from membrane proteins of wild-type Arabidopsis seedlings,which were analyzed further by immunoblot using antibodies againstEBS2 [an ER luminal protein (12)], EBS5 [an ER membrane-anchored protein (21)], and EBS7. As shown in Fig. 3G, EBS2 wasdetected exclusively in the soluble fraction, but both EBS5 and EBS7were detected only in the membrane fraction. Taken together, theseexperiments indicated that EBS7 is an ER membrane protein.Consistent with its pattern of coexpression with genes of ER
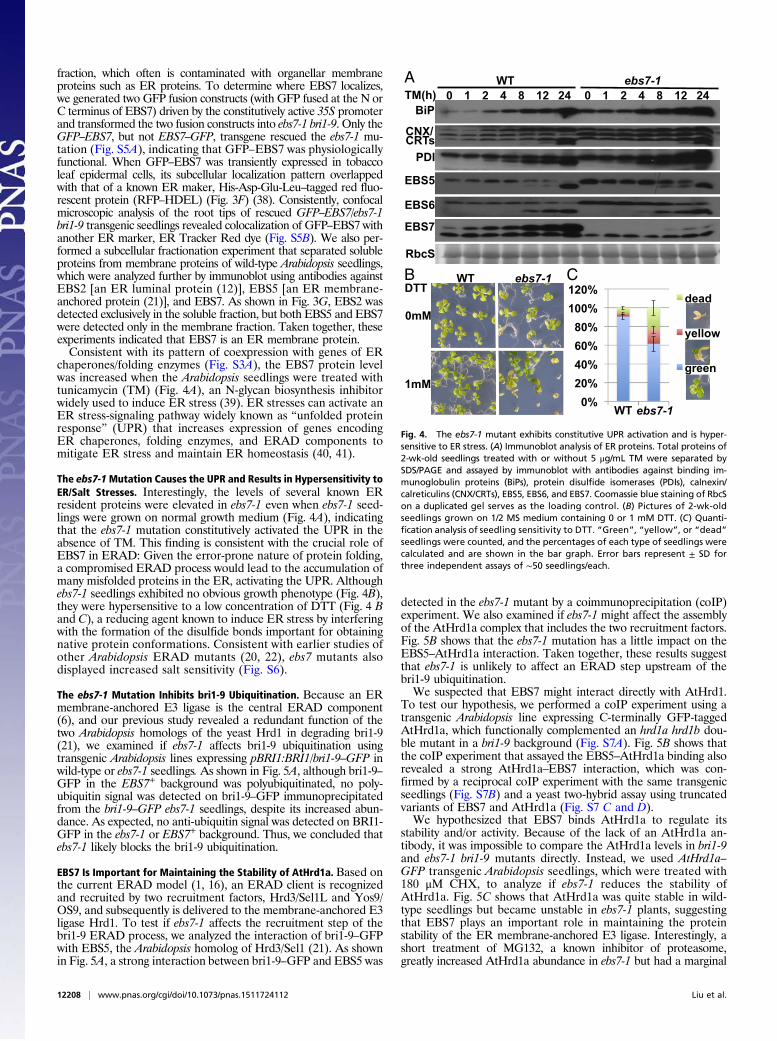
chaperones/folding enzymes (Fig. S3A), the EBS7 protein levelwas increased when the Arabidopsis seedlings were treated withtunicamycin (TM) (Fig. 4A), an N-glycan biosynthesis inhibitorwidely used to induce ER stress (39). ER stresses can activate anER stress-signaling pathway widely known as “unfolded proteinresponse” (UPR) that increases expression of genes encodingER chaperones, folding enzymes, and ERAD components tomitigate ER stress and maintain ER homeostasis (40, 41).
The ebs7-1Mutation Causes the UPR and Results in Hypersensitivity toER/Salt Stresses. Interestingly, the levels of several known ERresident proteins were elevated in ebs7-1 even when ebs7-1 seed-lings were grown on normal growth medium (Fig. 4A), indicatingthat the ebs7-1 mutation constitutively activated the UPR in theabsence of TM. This finding is consistent with the crucial role ofEBS7 in ERAD: Given the error-prone nature of protein folding,a compromised ERAD process would lead to the accumulation ofmany misfolded proteins in the ER, activating the UPR. Althoughebs7-1 seedlings exhibited no obvious growth phenotype (Fig. 4B),they were hypersensitive to a low concentration of DTT (Fig. 4 Band C), a reducing agent known to induce ER stress by interferingwith the formation of the disulfide bonds important for obtainingnative protein conformations. Consistent with earlier studies ofother Arabidopsis ERAD mutants (20, 22), ebs7 mutants alsodisplayed increased salt sensitivity (Fig. S6).
The ebs7-1 Mutation Inhibits bri1-9 Ubiquitination. Because an ERmembrane-anchored E3 ligase is the central ERAD component(6), and our previous study revealed a redundant function of thetwo Arabidopsis homologs of the yeast Hrd1 in degrading bri1-9(21), we examined if ebs7-1 affects bri1-9 ubiquitination usingtransgenic Arabidopsis lines expressing pBRI1:BRI1/bri1-9–GFP inwild-type or ebs7-1 seedlings.As shown in Fig. 5A, although bri1-9–GFP in the EBS7+ background was polyubiquitinated, no poly-ubiquitin signal was detected on bri1-9–GFP immunoprecipitatedfrom the bri1-9–GFP ebs7-1 seedlings, despite its increased abun-dance. As expected, no anti-ubiquitin signal was detected on BRI1-GFP in the ebs7-1 or EBS7+ background. Thus, we concluded thatebs7-1 likely blocks the bri1-9 ubiquitination.
EBS7 Is Important for Maintaining the Stability of AtHrd1a. Based onthe current ERAD model (1, 16), an ERAD client is recognizedand recruited by two recruitment factors, Hrd3/Sel1L and Yos9/OS9, and subsequently is delivered to the membrane-anchored E3ligase Hrd1. To test if ebs7-1 affects the recruitment step of thebri1-9 ERAD process, we analyzed the interaction of bri1-9–GFPwith EBS5, the Arabidopsis homolog of Hrd3/Sel1 (21). As shownin Fig. 5A, a strong interaction between bri1-9–GFP and EBS5 was
detected in the ebs7-1 mutant by a coimmunoprecipitation (coIP)experiment. We also examined if ebs7-1 might affect the assemblyof the AtHrd1a complex that includes the two recruitment factors.Fig. 5B shows that the ebs7-1 mutation has a little impact on theEBS5–AtHrd1a interaction. Taken together, these results suggestthat ebs7-1 is unlikely to affect an ERAD step upstream of thebri1-9 ubiquitination.We suspected that EBS7 might interact directly with AtHrd1.
To test our hypothesis, we performed a coIP experiment using atransgenic Arabidopsis line expressing C-terminally GFP-taggedAtHrd1a, which functionally complemented an hrd1a hrd1b dou-ble mutant in a bri1-9 background (Fig. S7A). Fig. 5B shows thatthe coIP experiment that assayed the EBS5–AtHrd1a binding alsorevealed a strong AtHrd1a–EBS7 interaction, which was con-firmed by a reciprocal coIP experiment with the same transgenicseedlings (Fig. S7B) and a yeast two-hybrid assay using truncatedvariants of EBS7 and AtHrd1a (Fig. S7 C and D).We hypothesized that EBS7 binds AtHrd1a to regulate its
stability and/or activity. Because of the lack of an AtHrd1a an-tibody, it was impossible to compare the AtHrd1a levels in bri1-9and ebs7-1 bri1-9 mutants directly. Instead, we used AtHrd1a–GFP transgenic Arabidopsis seedlings, which were treated with180 μM CHX, to analyze if ebs7-1 reduces the stability ofAtHrd1a. Fig. 5C shows that AtHrd1a was quite stable in wild-type seedlings but became unstable in ebs7-1 plants, suggestingthat EBS7 plays an important role in maintaining the proteinstability of the ER membrane-anchored E3 ligase. Interestingly, ashort treatment of MG132, a known inhibitor of proteasome,greatly increased AtHrd1a abundance in ebs7-1 but had a marginal
WT ebs7-1 DTT
0mM
1mM
0 1 2 12 24 4 8 0 1 2 12 24 4 8 WT ebs7-1
BiP
CNX/CRTs
PDI
EBS5
EBS6
EBS7
RbcS
TM(h)
0% 20% 40% 60% 80%
100% 120%
dead
yellow
green
WT ebs7-1
A
B C
Fig. 4. The ebs7-1 mutant exhibits constitutive UPR activation and is hyper-sensitive to ER stress. (A) Immunoblot analysis of ER proteins. Total proteins of2-wk-old seedlings treated with or without 5 μg/mL TM were separated bySDS/PAGE and assayed by immunoblot with antibodies against binding im-munoglobulin proteins (BiPs), protein disulfide isomerases (PDIs), calnexin/calreticulins (CNX/CRTs), EBS5, EBS6, and EBS7. Coomassie blue staining of RbcSon a duplicated gel serves as the loading control. (B) Pictures of 2-wk-oldseedlings grown on 1/2 MS medium containing 0 or 1 mM DTT. (C) Quanti-fication analysis of seedling sensitivity to DTT. “Green”, “yellow”, or “dead”seedlings were counted, and the percentages of each type of seedlings werecalculated and are shown in the bar graph. Error bars represent ± SD forthree independent assays of ∼50 seedlings/each.
12208 | www.pnas.org/cgi/doi/10.1073/pnas.1511724112 Liu et al.

effect on that of the AtHrd1a–GFP EBS7+ transgenic line (Fig. 5D).TheMG132 result not only confirmed the role of EBS7 in regulatingthe AtHrd1a stability but also suggested that AtHrd1a in ebs7-1might be degraded via a proteasome-mediated pathway.
DiscussionEBS7 Is a Plant-Specific ER Membrane-Anchored Component of anArabidopsis ERAD System. Despite rapid progress in understandingthe plant ERAD process, it remains unknown if plant ERADinvolves a plant-specific component, because all previous studiesidentified homologs of known components of the yeast/mammalianERAD pathways (see review in ref. 16). In this study, we showthat EBS7 is a plant-specific component of the Arabidopsis ERADmachinery. First, all three ebs7 mutants were isolated in two in-dependent genetic screens for potential ERADmutants defective indegrading bri1-9 or bri1-5, and our subsequent biochemical exper-iments confirmed that ebs7 mutations prevented the degradation oftwo ER-retained mutant variants of BRI1 (Figs. 1 and 2). Our studyshowed that ebs7-1 also blocked ERAD of a misfolded conformerof an Arabidopsis immunity receptor, EFR (Fig. 2F). Second,multiple previous microarray experiments revealed that EBS7/At4g29960 is coexpressed with genes encoding ERAD componentsand other ER-localized chaperones/folding catalysts (Fig. S3A); thiscoexpression was the main reason for its being considered a primecandidate gene for map-based cloning. We also showed that EBS7is an ER-localized membrane protein that accumulates in responseto ER stress (Figs. 3F and 4A). Third, our biochemical assaysdemonstrated that EBS7 interacted with at least two known ERADcomponents, EBS5 and AtHrd1a (Fig. 5B and Fig. S7), and thatebs7-1 inhibited the ubiquitination of the bri1-9 protein (Fig. 5A).Fourth, in line with previous observations of other known Arabi-dopsis ERAD components (20, 22), we found that loss-of-functionebs7 mutations constitutively activated the UPR and led to in-creased sensitivity to ER/salt stresses (Fig. 4 and Fig. S6). Last,BLAST searches against protein databases failed to identify EBS7homologs in fungi or animals but discovered highly similar proteinsin land plants, including Selaginella and Physcomitrella (Fig. S4).
EBS7 Plays an Important Role in Regulating AtHrd1a Stability.Althoughour data do not exclude the possibility that EBS7 is necessary forthe ubiquitin ligase activity of AtHrd1a, our study strongly suggeststhat EBS7 regulates the stability of the AtHrd1a protein. OurCHX-chasing assay revealed that ebs7-1 greatly increased thedegradation rate of AtHrd1a (Fig. 5C), and our MG132 assay in-dicated that a brief treatment with the proteasome inhibitor sig-nificantly increased the AtHrd1a abundance (Fig. 5D). It is wellknown that yeast and mammalian ERAD E3 ligases are regulatedby self-/heterologous ubiquitination (42), and our observed EBS7–AtHrd1a interaction suggested that EBS7 binding could preventthe self-/heterologous ubiquitination activity and subsequentdegradation of the Arabidopsis ERAD E3 ligase. Alternatively,EBS7 might be involved in the assembly of an AtHrd1a-containingERAD complex because EBS7 also interacts with EBS5 (Fig. S7E),the Arabidopsis homolog of Hrd3/SEl1L, which functions as one ofthe two recruitment factors for a committed ERAD client. Al-though the ebs7-1 mutation had little effect on the EBS5–AtHrd1ainteraction (Fig. 5B), it could interfere the binding of AtHrd1a withother components of the Arabidopsis ERAD machinery, includinganother ERAD client-recruitment factor, EBS6 (19, 20), an ERmembrane-anchored E2, UBC32 (24), and/or one or more Arabi-dopsis homologs of the yeast Der1/mammalian Derlins. It also ispossible that ebs7 mutations compromise the AtHrd1a oligomeri-zation. It is well known that the ER-mediated protein quality-control system retains and degrades not only misfolded proteins,such as bri1-5, bri1-9, and misfolded EFR, but also improperlyassembled polypeptides. An improperly assembled AtHrd1a thuscould be degraded by an AtHrd1a-independent ERAD pathway. Itis interesting that both the yeast Usa1 and mammalian HERP areknown to interact directly with their corresponding ER membrane-anchored E3 ligases and are thought to regulate the stability and/oroligomerization of their interacting E3 ligases (43–45). Despite theabsence of sequence homology with Usa1 and HERP, both ofwhich carry an ubiquitin-like domain at the N terminus, EBS7could be a functional homolog of Usa1/HERP. However, ourcomplementation experiments revealed that EBS7 failed tocomplement a yeast Δusa1 mutant and that a HERP transgenedriven by the strong 35S promoter was unable to rescue the ebs7-1mutation (Fig. S8). We hypothesize that the biochemical mechanismby which EBS7 regulates the stability and/or oligomerization ofthe Arabidopsis Hrd1 homologs might be different from themechanism by which Usa1/HERP regulate their interactingERAD E3 ligases. Further investigation is needed to understandfully the biochemical function of this plant-specific componentin regulating a highly conserved plant ERAD pathway.
Materials and MethodsDetails of map-based cloning of EBS7, plasmid construction, generation oftransgenic plants, expression of fusion proteins in Escherichia coli, generationof antibodies, and immunoblot, membrane fractionation, and coIP experi-ments are given in SI Materials and Methods. Oligonucleotides used in thisstudy are given in Table S1.
Plant Materials and Growth Conditions. Most of the Arabidopsis wild-type,mutant, and transgenic plants used in the study are in the Columbia-0 (Col-0)ecotype, except for bri1-5, its two allelic ebs7 suppressors (ebs7-2 and ebs7-3),and a different bri1-9 mutant (used for mapping the EBS7 locus), which areall in the Wassilewskija-2 (Ws-2) ecotype. Methods of seed sterilizationand plant growth conditions were described previously (46), and the root-growth inhibition assay on BL-containing medium was performed accordingto a previously described protocol (47).
Tobacco Transient Expression and Confocal Microscopic Analysis of EBS7–GFPFusion Proteins. The Agrobacterium strains carrying the p35S:EBS7–GFP orp35S:RFP–HDEL plasmid were mixed and coinfiltrated into leaves of 5-wk-old tobacco (Nicotiana benthamiana) plants by the agro-infiltration method(48). The subcellular localization patterns of EBS7–GFP and RFP–HDEL pro-teins were examined using a Leica confocal laser-scanning microscope (TCSSP5 DM6000B) and LAS AF software (Leica Microsystems). GFP and RFP wereexcited by using 488- and 543-nm laser lights, respectively.
IP:
GFP
Inpu
tebs7-1
BRI1 -GFP
bri1-9 -GFP
WT
BRI1/bri1-9
BRI1/bri1-9
WB
:-U
b
EBS5
EBS5
BRI1 -GFP
bri1-9 -GFP
Hrd1a-GFP
IP:
GFP
WT WT ebs7-1
EBS5
EBS7
Hrd1a-GFP
Inpu
t
EBS5
EBS7
Hrd1a-GFP
*
Hrd1a-GFP
Hrd1a-GFP
MG132(min)0 45 90
RbcS
RbcSEBS7
+ eb
s7-1
CHX(h) 0 3 6 12 16 24
EBS7
+ eb
s7-1
Hrd1a-GFP
Hrd1a-GFP
RbcS
RbcS
A B
C D
Fig. 5. EBS7 interacts with AtHrd1a and affects its stability. (A) CoIP of bri1-9and EBS5. Total proteins and anti-GFP immunoprecipitates obtained fromwild-type or transgenic plants were separated by SDS/PAGE and analyzed byimmunoblot with antibodies against GFP, ubiquitin, or EBS5. (B) CoIP ofEBS7, AtHrd1a, and EBS5. Total proteins and anti-GFP immunoprecipitatesobtained from wild-type or transgenic Arabidopsis plants were separated bySDS/PAGE and analyzed by immunoblot with anti-GFP, anti-EBS7, and anti-EBS5 antibodies. (C and D) Immunoblot analysis of Hrd1a–GFP. Total proteinsextracted from 2-wk-old seedlings treated with 180 μM CHX (C) or 80 μMMG132 (D) for varying durations were separated by SDS/PAGE and assayedby immunoblot with an anti-GFP antibody. Coomassie blue staining of RbcSon a duplicate gel serves as a loading control.
Liu et al. PNAS | September 29, 2015 | vol. 112 | no. 39 | 12209
PLANTBIOLO
GY

ACKNOWLEDGMENTS.We thank the Arabidopsis Biological Resource Centerat Ohio State University for supplying the T-DNA insertional mutant ofAtHrd1a and AtHrd1b; F. Tax (University of Arizona) for seeds of bri1-5(Ws-2) and bri1-9 (Ws-2); J. Chory (Salk Institute) for anti-BRI1 antibody; Y. Yin(Iowa State University) for anti-BES1 antibody; R. Boston (North Carolina
State University) for anti-maize CRT antibody; T. Tzfira for the p35S:RFP–HDEL plasmid; and members of the J.L. laboratory for helpful discussions.This work was supported in part by Grant OVPRE-U032381 from the Univer-sity of Michigan and Grant 2012CSP004 from the Chinese Academyof Sciences.
1. Ruggiano A, Foresti O, Carvalho P (2014) Quality control: ER-associated degradation:Protein quality control and beyond. J Cell Biol 204(6):869–879.
2. Vembar SS, Brodsky JL (2008) One step at a time: Endoplasmic reticulum-associateddegradation. Nat Rev Mol Cell Biol 9(12):944–957.
3. Hebert DN, Molinari M (2012) Flagging and docking: Dual roles for N-glycans inprotein quality control and cellular proteostasis. Trends Biochem Sci 37(10):404–410.
4. Denic V, Quan EM, Weissman JS (2006) A luminal surveillance complex that selectsmisfolded glycoproteins for ER-associated degradation. Cell 126(2):349–359.
5. Gauss R, Jarosch E, Sommer T, Hirsch C (2006) A complex of Yos9p and the HRD ligaseintegrates endoplasmic reticulum quality control into the degradation machinery. NatCell Biol 8(8):849–854.
6. Kostova Z, Tsai YC, Weissman AM (2007) Ubiquitin ligases, critical mediators of en-doplasmic reticulum-associated degradation. Semin Cell Dev Biol 18(6):770–779.
7. Olzmann JA, Kopito RR, Christianson JC (2013) The mammalian endoplasmic re-ticulum-associated degradation system. Cold Spring Harb Perspect Biol 5(9):a013185.
8. Xie W, Ng DT (2010) ERAD substrate recognition in budding yeast. Semin Cell Dev Biol21(5):533–539.
9. Bagola K, Mehnert M, Jarosch E, Sommer T (2011) Protein dislocation from the ER.Biochim Biophys Acta 1808(3):925–936.
10. Wolf DH, Stolz A (2012) The Cdc48 machine in endoplasmic reticulum associatedprotein degradation. Biochim Biophys Acta 1823(1):117–124.
11. Ceriotti A, Roberts LM (2006) Endoplasmic reticulum-associated protein degradationin plant cells. The Plant Endoplasmic Reticulum, ed Robinson DG (Springer, Berlin), pp75–98.
12. Jin H, Hong Z, Su W, Li J (2009) A plant-specific calreticulin is a key retention factor fora defective brassinosteroid receptor in the endoplasmic reticulum. Proc Natl Acad SciUSA 106(32):13612–13617.
13. Jin H, Yan Z, Nam KH, Li J (2007) Allele-specific suppression of a defective brassi-nosteroid receptor reveals a physiological role of UGGT in ER quality control. Mol Cell26(6):821–830.
14. Hong Z, Jin H, Tzfira T, Li J (2008) Multiple mechanism-mediated retention of a de-fective brassinosteroid receptor in the endoplasmic reticulum of Arabidopsis. PlantCell 20(12):3418–3429.
15. Hong Z, Li J (2012) The protein quality control of plant receptor-like kinases in theendoplasmic reticulum. Receptor-Like Kinases in Plants: From Development to De-fense. Signaling and Communication in Plants, eds. Tax F, Kemmerling B. (Springer,Berlin), Vol. 13, pp 275–307.
16. Liu Y, Li J (2014) Endoplasmic reticulum-mediated protein quality control in Arabi-dopsis. Front Plant Sci 5:162.
17. Hong Z, et al. (2012) Evolutionarily conserved glycan signal to degrade aberrant bras-sinosteroid receptors in Arabidopsis. Proc Natl Acad Sci USA 109(28):11437–11442.
18. Hong Z, et al. (2009) Mutations of an alpha1,6 mannosyltransferase inhibit endo-plasmic reticulum-associated degradation of defective brassinosteroid receptors inArabidopsis. Plant Cell 21(12):3792–3802.
19. Su W, Liu Y, Xia Y, Hong Z, Li J (2012) The Arabidopsis homolog of the mammalianOS-9 protein plays a key role in the endoplasmic reticulum-associated degradation ofmisfolded receptor-like kinases. Mol Plant 5(4):929–940.
20. Hüttner S, Veit C, Schoberer J, Grass J, Strasser R (2012) Unraveling the function ofArabidopsis thaliana OS9 in the endoplasmic reticulum-associated degradation ofglycoproteins. Plant Mol Biol 79(1-2):21–33.
21. Su W, Liu Y, Xia Y, Hong Z, Li J (2011) Conserved endoplasmic reticulum-associateddegradation system to eliminate mutated receptor-like kinases in Arabidopsis. ProcNatl Acad Sci USA 108(2):870–875.
22. Liu L, et al. (2011) The endoplasmic reticulum-associated degradation is necessary forplant salt tolerance. Cell Res 21(6):957–969.
23. Hüttner S, et al. (2014) Arabidopsis Class I α-Mannosidases MNS4 and MNS5 Are In-volved in Endoplasmic Reticulum-Associated Degradation of Misfolded Glycoproteins.Plant Cell 26(4):1712–1728.
24. Cui F, et al. (2012) Arabidopsis ubiquitin conjugase UBC32 is an ERAD component thatfunctions in brassinosteroid-mediated salt stress tolerance. Plant Cell 24(1):233–244.
25. Yin Y, et al. (2002) BES1 accumulates in the nucleus in response to brassinosteroids toregulate gene expression and promote stem elongation. Cell 109(2):181–191.
26. Zipfel C, et al. (2006) Perception of the bacterial PAMP EF-Tu by the receptor EFRrestricts Agrobacterium-mediated transformation. Cell 125(4):749–760.
27. Li J, et al. (2009) Specific ER quality control components required for biogenesis of theplant innate immune receptor EFR. Proc Natl Acad Sci USA 106(37):15973–15978.
28. Saijo Y, et al. (2009) Receptor quality control in the endoplasmic reticulum for plantinnate immunity. EMBO J 28(21):3439–3449.
29. Saijo Y (2010) ER quality control of immune receptors and regulators in plants. CellMicrobiol 12(6):716–724.
30. KnopM, Finger A, Braun T, Hellmuth K,Wolf DH (1996) Der1, a novel protein specificallyrequired for endoplasmic reticulum degradation in yeast. EMBO J 15(4):753–763.
31. Lilley BN, Ploegh HL (2004) A membrane protein required for dislocation of misfoldedproteins from the ER. Nature 429(6994):834–840.
32. Ye Y, Shibata Y, Yun C, Ron D, Rapoport TA (2004) A membrane protein complex me-diates retro-translocation from the ER lumen into the cytosol. Nature 429(6994):841–847.
33. Obayashi T, Hayashi S, Saeki M, Ohta H, Kinoshita K (2009) ATTED-II providescoexpressed gene networks for Arabidopsis. Nucleic Acids Res 37(Database issue):D987–D991.
34. Winter D, et al. (2007) An “Electronic Fluorescent Pictograph” browser for exploringand analyzing large-scale biological data sets. PLoS One 2(8):e718.
35. Schwacke R, et al. (2003) ARAMEMNON, a novel database for Arabidopsis integralmembrane proteins. Plant Physiol 131(1):16–26.
36. Mitra SK, Walters BT, Clouse SD, Goshe MB (2009) An efficient organic solvent basedextraction method for the proteomic analysis of Arabidopsis plasma membranes.J Proteome Res 8(6):2752–2767.
37. Benschop JJ, et al. (2007) Quantitative phosphoproteomics of early elicitor signalingin Arabidopsis. Mol Cell Proteomics 6(7):1198–1214.
38. Liu CJ, Dixon RA (2001) Elicitor-induced association of isoflavone O-methyltransferasewith endomembranes prevents the formation and 7-O-methylation of daidzeinduring isoflavonoid phytoalexin biosynthesis. Plant Cell 13(12):2643–2658.
39. Elbein AD (1981) The tunicamycins - useful tools for studies on glycoproteins. TrendsBiochem Sci 6(8):219–221.
40. Hetz C (2012) The unfolded protein response: Controlling cell fate decisions under ERstress and beyond. Nat Rev Mol Cell Biol 13(2):89–102.
41. Howell SH (2013) Endoplasmic reticulum stress responses in plants. Annu Rev PlantBiol 64:477–499.
42. Weissman AM, Shabek N, Ciechanover A (2011) The predator becomes the prey:Regulating the ubiquitin system by ubiquitylation and degradation. Nat Rev Mol CellBiol 12(9):605–620.
43. Carroll SM, Hampton RY (2010) Usa1p is required for optimal function and regulationof the Hrd1p endoplasmic reticulum-associated degradation ubiquitin ligase. J BiolChem 285(8):5146–5156.
44. Kim I, Li Y, Muniz P, Rao H (2009) Usa1 protein facilitates substrate ubiquitylationthrough two separate domains. PLoS One 4(10):e7604.
45. Leitman J, et al. (2014) Herp coordinates compartmentalization and recruitment ofHRD1 and misfolded proteins for ERAD. Mol Biol Cell 25(7):1050–1060.
46. Li J, Nam KH, Vafeados D, Chory J (2001) BIN2, a new brassinosteroid-insensitive locusin Arabidopsis. Plant Physiol 127(1):14–22.
47. Clouse SD, Langford M, McMorris TC (1996) A brassinosteroid-insensitive mutant inArabidopsis thaliana exhibits multiple defects in growth and development. PlantPhysiol 111(3):671–678.
48. Voinnet O, Rivas S, Mestre P, Baulcombe D (2003) An enhanced transient expressionsystem in plants based on suppression of gene silencing by the p19 protein of tomatobushy stunt virus. Plant J 33(5):949–956.
49. Friedrichsen DM, Joazeiro CA, Li J, Hunter T, Chory J (2000) Brassinosteroid-insensitive-1 is a ubiquitously expressed leucine-rich repeat receptor serine/threoninekinase. Plant Physiol 123(4):1247–1256.
50. Fankhauser C, et al. (1999) PKS1, a substrate phosphorylated by phytochrome thatmodulates light signaling in Arabidopsis. Science 284(5419):1539–1541.
51. Bechtold N, Pelletier G (1998) In planta Agrobacterium-mediated transformation ofadult Arabidopsis thaliana plants by vacuum infiltration. Methods Mol Biol 82:259–266.
52. Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W: Improving the sensitivity ofprogressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res 22(22):4673–4680.
53. Chevenet F, Brun C, Bañuls AL, Jacq B, Christen R (2006) TreeDyn: Towards dynamicgraphics and annotations for analyses of trees. BMC Bioinformatics 7:439.
54. Mora-García S, et al. (2004) Nuclear protein phosphatases with Kelch-repeat domainsmodulate the response to brassinosteroids in Arabidopsis. Genes Dev 18(4):448–460.
55. Wang ZY, Seto H, Fujioka S, Yoshida S, Chory J (2001) BRI1 is a critical component of aplasma-membrane receptor for plant steroids. Nature 410(6826):380–383.
56. Persson S, et al. (2003) Phylogenetic analyses and expression studies reveal two dis-tinct groups of calreticulin isoforms in higher plants. Plant Physiol 133(3):1385–1396.
57. Gietz RD, Woods RA (2002) Transformation of yeast by lithium acetate/single-stranded carrier DNA/polyethylene glycol method. Methods Enzymol 350:87–96.
12210 | www.pnas.org/cgi/doi/10.1073/pnas.1511724112 Liu et al.