Embed Size (px)
Citation preview

Influence of Economics, Interspecific Competition, and Sexual Dimorphism on Territorialityof Migrant Rufous HummingbirdsAuthor(s): Astrid Kodric-Brown and James H. BrownSource: Ecology, Vol. 59, No. 2 (Mar., 1978), pp. 285-296Published by: Ecological Society of AmericaStable URL: http://www.jstor.org/stable/1936374 .Accessed: 21/06/2011 14:29
Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unlessyou have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and youmay use content in the JSTOR archive only for your personal, non-commercial use.
Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained at .http://www.jstor.org/action/showPublisher?publisherCode=esa. .
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printedpage of such transmission.
JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
Ecological Society of America is collaborating with JSTOR to digitize, preserve and extend access to Ecology.
http://www.jstor.org

Ecology, 59(2), 1978, pp. 285-296 ?) 1978 by the Ecological Society of America
INFLUENCE OF ECONOMICS, INTERSPECIFIC COMPETITION, AND SEXUAL DIMORPHISM ON TERRITORIALITY OF MIGRANT
RUFOUS HUMMINGBIRDS1
ASTRID KODRIC-BROWN AND JAMES H. BROWN Department of Ecology and Evolutionary Biology, University of Arizona, Tucson, Arizona 85721 USA
Abstract. Migrant Rufous Hummingbirds (Selasphorus rufus) arrive in eastern Arizona in late summer and establish feeding territories from which other hummingbirds are excluded. Territories vary 100-fold in area and 5-fold in number of flowers. A simple cost-benefit model accounts for observed variation in territory size and number of flowers defended. Both sexes defend territories, but d d utilize denser flowers than Y Y. These differences appear to be related to sexual dimorphism in wing disc loading. Selasphorus rufus appears to have sacrificed efficient flight for aggressive ability as a strategy for competing with resident hummingbird species during its migration. Comparison of feeding territories of S. rufus and other nectarivorous birds indicate similarities which suggest that these systems may be subject to similar economic constraints.
Key words: Aggression; Aves; behavior; competition; energetics; foraging; hummingbird; mi- gration; nectar; Selasphorus rufus; sexual dimorphism; territoriality; wing disc loading.
INTRODUCTION
Territoriality is utilized by many animals to exploit limited resources such as food, breeding sites and mates. Territoriality is presumed to be adaptive when- ever it is economical; when the benefits of exclusive use of a resource outweigh the costs of its defense (Brown 1964, Brown and Orians 1970). This concept of economic defensibility has stimulated empirical at- tempts to assess the adaptive significance of territo- riality by measuring characteristics of organisms and the resources that determine these costs and benefits.
Characteristics of feeding territories of nectarivo- rous birds have caused them to be frequently used in investigations of the economics of territoriality. Most of these studies have attempted to assess costs and benefits by estimating the energy budgets of territorial (and sometimes nonterritorial) individuals and relating these to energy content of available food (Pearson 1954, Stiles and Wolf 1970, Wolf 1970. 1975, Stiles 1971, Gill and Wolf 1975, Carpenter and MacMillen 1976a, b, see also Lasiewski 1963, Wolf and Hains- worth 1971, Hainsworth and Wolf 1972, Hainsworth 1974). This approach has many advantages. However, several potential errors in extrapolating laboratory measurements of energetic costs to field measure- ments of time budgets make it difficult to determine the precise conditions when territoriality is advanta- geous. An alternative approach is to assume observed territories are economical, to measure variation in the kind and distribution of resources utilized by territorial and nonterritorial individuals, and to infer costs and benefits from these data (e.g., Gass et al. 1976). We have used the latter approach to study feeding terri- tories of Rufous Hummingbirds (Selasphorus rufus).
The present study investigates economics of terri- toriality in this species in the context of intra- and interspecific competition for food. Selasphorus rufus migrates through the mountains of the southwestern United States enroute from its breeding ground in northwestern North America to its wintering ground in southern Mexico (Grant and Grant 1967, Phillips 1975, Gass et al. 1976). During migration, individuals of both sexes temporarily establish territories where nectar-producing flowers are sufficiently abundant. In this paper, we describe and discuss the temporal pat- tern of territory establishment and occupation, cor- relates of variation in territory size and number of flowers defended, and differences in aggressive and foraging behavior between the sexes.
METHODS
The study area
Fieldwork was conducted in the White Mountains of east-central Arizona during the summers of 1973- 1976. All investigations of territoriality at natural flow- ers were made in the immediate vicinity of a large meadow 2,300 m elevation, 6 km NE of Nutrioso, Arizona, from 2 July-10 August 1975 and 5-17 August 1976. At this primary study site, hummingbirds de- fended territories where 3 species of red, tubular flow- ers bloomed in the meadow and on adjacent sparsely wooded slopes. Some supplementary work was done in other areas and habitats within a few kilometres of the primary study site. In 1974, experiments with ar- tificial feeders were performed in open woodland hab- itat at 2 sites: 1,900 m elevation, 25 km SE of Alpine and 2,600 m elevation, 6 km N of Alpine, Arizona.
The birds
Two species of hummingbirds, breeding Selaspho- rus platycercus and migrant S. rufus were abundant
1 Manuscript received 4 April 1977; accepted 2 August 1977.

286 ASTRID KODRIC-BROWN AND JAMES H. BROWN Ecology, Vol. 59, No. 2
in the primary study area. These species were cen- sused during standardized I-h walks at weekly inter- vals during 1975. During all 4 summers, 276 birds were captured in mist nets to sample the pollen they were carrying and to measure their body weight (in 1975 and 1976 only), wing length and bill (culmen) length.
Territories of 20 male and 4 female S. rufus were mapped at the primary study site in 1975; territories of 19 males and 5 females were mapped in the same area in 1976. Territory size was determined by ob- serving defense and foraging behavior of resident birds, marking boundaries with flagging tape, and cal- culating the included area. In most cases, boundaries of territories, and age and sex of residents were de- termined accurately by observing territorial individu- als. In the few cases where the boundaries of adjacent territories overlapped significantly, the area of overlap and included flowers were divided equally between the 2 birds. In 1975, 6 territorial birds (3 males and 3 fe- males) were captured in mist nets and individually marked with small pieces of colored flagging tape glued to the crown. This procedure facilitated mapping of large territories and provided some data on duration of territorial occupancy. In 1976, 7 birds (3 females and 4 juvenile males) were captured to verify age and sex (Stiles 1972).
The flowers
Hummingbirds utilized 3 species of red, tubular flowers, Castilleja integra, Penstemon barbatus and Ipomopsis aggregata as primary food sources. The number of flowers of each species within a territory was counted and flowers were selected arbitrarily for measurement of available nectar. From 1400-1600 h, the entire nectar contents of these flowers were mea- sured with 20-,ul calibrated micropipettes. In 1975, when P. barbatus was the most abundant flower on territories, available nectar was measured in 2 flowers on each of 30 plants of this species on each territory. In 1976, when I. aggregata was more abundant than P. barbatus on most territories. we measured available nectar in I flower on each of 30 plants of each species (if abundances permitted) on each territory.
Other floral characteristics of the 3 species were measured on a large scale to obtain values character- istic of the study area as a whole. These included (1) corolla length, measured with calipers to the nearest 0.5 mm; (2) nectar concentration, measured as equiv- alent percent sucrose using a temperature compensat- ed hand refractometer; (3) 24-h nectar production, de- termined by covering inflorescence with nylon mesh bags, measuring the nectar accumulated after 24 h, and correcting for the number of new flowers opening each day and the amount of nectar in control flowers on adjacent plants sampled at the time of bagging; and (4) diel patterns of nectar secretion, measured as de- scribed above except that bags were left in place for 2-h periods distributed throughout the day. In addi-
tion, seasonal patterns of flower density and nectar availability were determined by sampling at weekly intervals during 1975. Flower densities were ranked on a scale from 1(<0.05 inflorescences/m2) to 5(>5 inflorescences/m2), and 2 flowers from each of 50 plants of each species were sampled for nectar from 1400-1600 h.
Experimental manipulations of territories
In 1975, 2 kinds of perturbations of natural territo- ries were attempted. The number of flowers on 3 mapped territories was reduced by cutting off approx- imately half of the inflorescences. Then, 48 h later, territories were mapped and available nectar in flow- ers was sampled. One artificial hummingbird feeder that supplied a 20% sucrose solution was placed in each of 6 mapped territories. One resident bird began using its feeder almost immediately; its territory was mapped and nectar in flowers was sampled 48 h later. The other feeders never were used, although they re- mained in place for 7 days.
Because of an exceptional drought in 1974, so few flowers were in bloom we were unable to observe sus- tained territorial defense of native flowers. We studied some aspects of territoriality by observing behavior of birds at artificial feeders. At each of the supplemental study sites described above, we placed several glass feeders containing 150 cm3 of 20% sucrose solution, which the birds removed from the bottom through a single glass tube. Feeders were hung in open areas from the lower branches of trees at a height of -=1.5 m and separated by distances of at least 5 m. Most of the hummingbirds using the feeders were captured in mist nets and individually marked as described above. Utilization and defense of feeders was recorded by thorough censuses conducted at least once each day.
RESULTS
Seasonal patterns and interspecific interactions
Specialized red, tubular hummingbird flowers were available and used by hummingbirds throughout the summer (Fig. 1). Selasphorus platycercus utilized these flowers early in the season.
Selasphorus platycercus were displaced abruptly by migrant S. rufus in early July (Fig. 1). The date of the first observed S. rufus was remarkably consistent from year to year: 7 July 1973; 3 July 1974; 4 July 1975; not observed in 1976. The migrants appeared to arrive in waves that coincided with the onset of clear weather following cloudy and rainy periods. The first arrivals were exclusively adults, and males appeared to out- number females. Juveniles were rarely seen or cap- tured in mist nets before mid-August, when they began to arrive in numbers. The newly arrived S. rufus were intensely aggressive, both intra- and interspecifically. The first arrivals established territories on the densest patches of flowers. By mid-July, all flowers on the study site had been incorporated into territories of S.
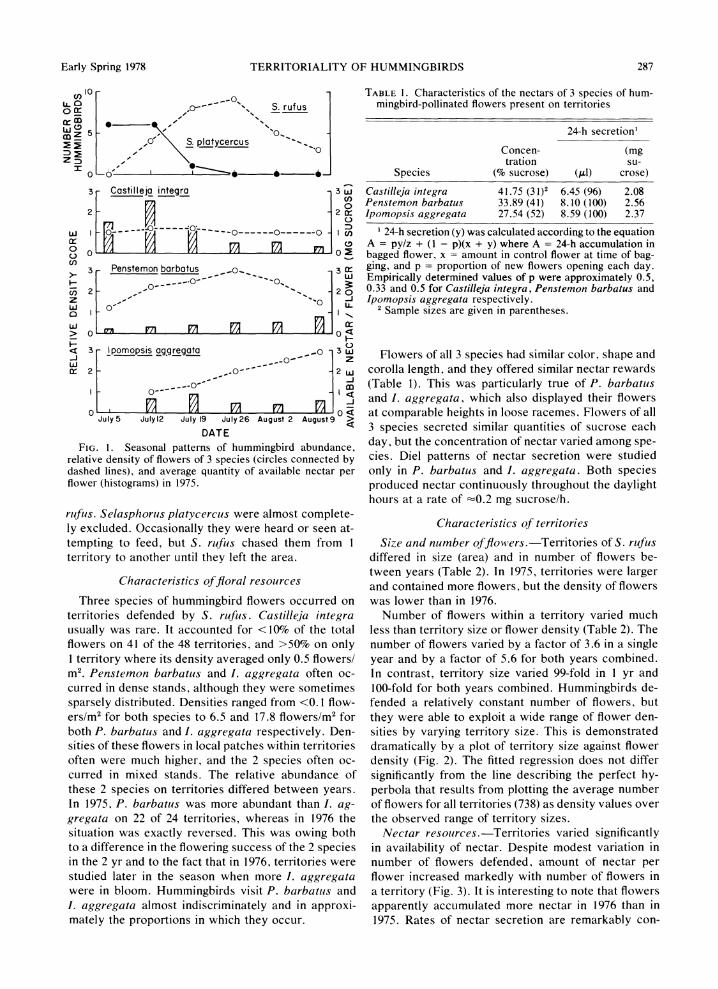
Early Spring 1978 TERRITORIALITY OF HUMMINGBIRDS 287
O_: , A S. rufus
(r ;i- -- xx-2
cr~ * - S. p atycercus 0_ .
_ 0
3 Capoostilia intereat_o- 3 W)
I U) 10
t:2 _,0 2 LL
DAT E ~ ~ ~
dashed in an ------ qu y I a
Glwr ahstol mtgra?-o in w95
te Penstemon barbatus r c t from
z _j~~~~~~
CharacteristicsofJly26l Augurst2Auu
FThre1 esonaepaters of hummingbird abwes cunrdance
rltvdestofflowerson4 of 3h specrrtoies, (circles%connce bny dahe lerinoywes), and averaesuatty ofvavailablel nectaropers
rufus. Selsphmous plratyr e Wregat omte-
lycluded. Ocaesiostnd,allyug they were hado semeniat- teptingl ditoibfeed,.butnS.irufs chasged thmfrom 10flw
teri2 ory toothe until th lef the ae.8
Chh . arbacterstics of floreala resourctiesyDn sthreseies of humingir flowers occua achswthntrredtonie
ths pce nterritories defneib.rfus.e Castilejaentyegars usually. brbtu was raemIcouned foru<10%no then total- flowers on 41 of the48 territories, ande>50% on onlyth
Jurittory where its dly renrsiy a Aguse ol 0ig.5 flo
FIn1.PeStemsona baratuers anfI humigbrdgabuanoftnoc-,
ureti dinfdense snds fower to ug species were sometes
sparshedlyinstributd. Dvraensiatieso rangedlfrom <0.1a flow
el/em2 fr bhistog thrpeie to5atnd 197. flers/ie fore
bothuP. bearbatrus pandyIcagregataereamspectivelyDene-
sities ol these Owerseinall pa whesre Ithin tereriate
ured in toomied, sutaS. Tfres eatsie ambaun anc o hthese species on tertiomines differe occurbee y
Ins97ua.lbrbtu was mae Icorned abundanto than totag- gregata on 22 of 2he48 territories, whereas% in17 they seritutony wasr exatly renstversed.Thi wasy 0fowingbot
in thenst2 obyrtu and totefatta ing97,rerritaories were studied ltrin thne sead,astonghwhen more somggegtame wearsel ditinbbloo.Hummngbirdes visitd P.o barbatusland
btiP aggregataulmst indiscagrimiatarselytandin appox- matielyo thes proportos in whichl ptcheyiti occr. iore
TABLE 1. Characteristics of the nectars of 3 species of hum- mingbird-pollinated flowers present on territories
24-h secretion'
Concen- (mg tration su-
Species (% sucrose) (Al) crose)
Castilleja integra 41.75 (3 1)2 6.45 (96) 2.08 Penstemon barbatus 33.89 (41) 8.10 (100) 2.56 Ipomopsis aggregata 27.54 (52) 8.59 (100) 2.37
1 24-h secretion (y) was calculated according to the equation A = py/z + (1 - p)(x + y) where A = 24-h accumulation in bagged flower, x = amount in control flower at time of bag- ging, and p = proportion of new flowers opening each day. Empirically determined values of p were approximately 0.5, 0.33 and 0.5 for Castilleja integra, Penstemon barbatus and Ipomopsis aggregata respectively.
2 Sample sizes are given in parentheses.
Flowers of all 3 species had similar color, shape and corolla length, and they offered similar nectar rewards (Table 1). This was particularly true of P. barbatus and I. aggregata, which also displayed their flowers at comparable heights in loose racemes. Flowers of all 3 species secreted similar quantities of sucrose each day, but the concentration of nectar varied among spe- cies. Diel patterns of nectar secretion were studied only in P. barbatus and I. aggregata. Both species produced nectar continuously throughout the daylight hours at a rate of =-0.2 mg sucrose/h.
Characteristics of territories
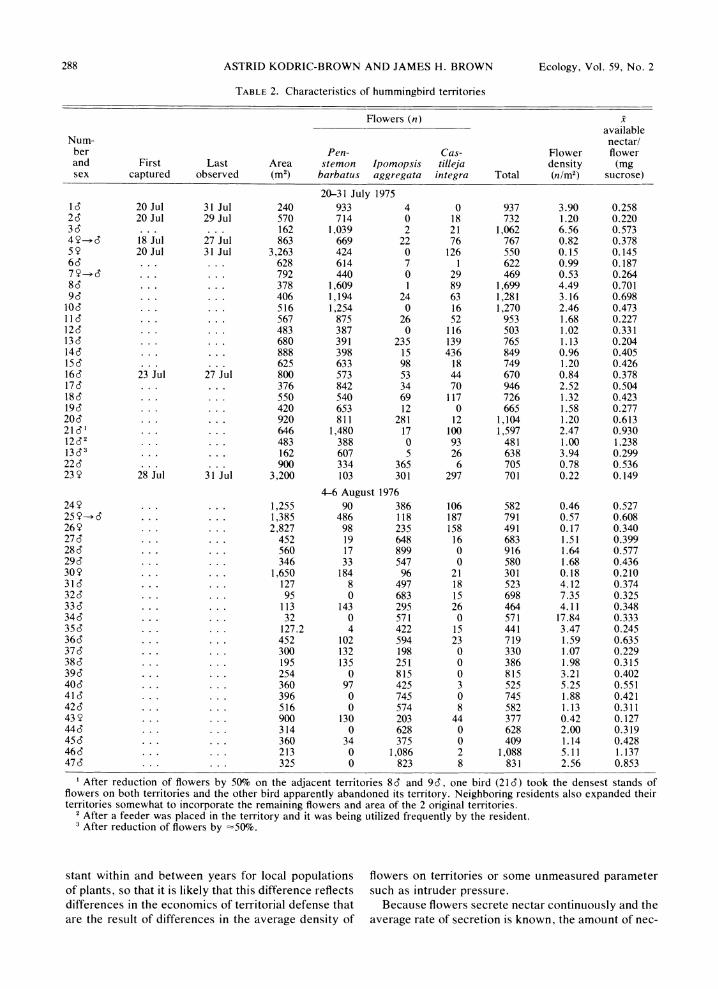
Size and number offlowers.-Territories of S. rufus differed in size (area) and in number of flowers be- tween years (Table 2). In 1975, territories were larger and contained more flowers, but the density of flowers was lower than in 1976.
Number of flowers within a territory varied much less than territory size or flower density (Table 2). The number of flowers varied by a factor of 3.6 in a single year and by a factor of 5.6 for both years combined. In contrast, territory size varied 99-fold in 1 yr and 100-fold for both years combined. Hummingbirds de- fended a relatively constant number of flowers, but they were able to exploit a wide range of flower den- sities by varying territory size. This is demonstrated dramatically by a plot of territory size against flower density (Fig. 2). The fitted regression does not differ significantly from the line describing the perfect hy- perbola that results from plotting the average number of flowers for all territories (738) as density values over the observed range of territory sizes.
Nectar resources. -Territories varied significantly in availability of nectar. Despite modest variation in number of flowers defended, amount of nectar per flower increased markedly with number of flowers in a territory (Fig. 3). It is interesting to note that flowers apparently accumulated more nectar in 1976 than in 1975. Rates of nectar secretion are remarkably con-
288 ASTRID KODRIC-BROWN AND JAMES H. BROWN Ecology, Vol. 59, No. 2
TABLE 2. Characteristics of hummingbird territories
Flowers (n) x available
Num- nectar/ ber Pen- Cas- Flower flower and First Last Area stemon Ipomopsis tilleja density (mg sex captured observed (m2) barbatus aggregata integra Total (n/M2) sucrose)
20-31 July 1975 1c 20Jul 31 Jul 240 933 4 0 937 3.90 0.258 26 20 Jul 29 Jul 570 714 0 18 732 1.20 0.220 36d ... ... 162 1,039 2 21 1,062 6.56 0.573 4 Y ----> 6 18 Jul 27 Jul 863 669 22 76 767 0.82 0.378 5Y 20Jul 31 Jul 3,263 424 0 126 550 0.15 0.145 66 ... ... 628 614 7 1 622 0.99 0.187 7 Y-* ... ... 792 440 0 29 469 0.53 0.264 86 ... ... 378 1,609 1 89 1,699 4.49 0.701 96 ... ... 406 1,194 24 63 1,281 3.16 0.698
106 ... ... 516 1,254 0 16 1,270 2.46 0.473 lid ... ... 567 875 26 52 953 1.68 0.227 126 ... ... 483 387 0 116 503 1.02 0.331 136 ... ... 680 391 235 139 765 1.13 0.204 146 ... ... 888 398 15 436 849 0.96 0.405 156 ... ... 625 633 98 18 749 1.20 0.426 166 23 Jul 27 Jul 800 573 53 44 670 0.84 0.378 176 ... ... 376 842 34 70 946 2.52 0.504 186 ... ... 550 540 69 117 726 1.32 0.423 196 ... ... 420 653 12 0 665 1.58 0.277 206 ... ... 920 811 281 12 1,104 1.20 0.613 21 dI ... ... 646 1,480 17 100 1,597 2.47 0.930 1262 ... ... 483 388 0 93 481 1.00 1.238 13 63 ... ... 162 607 5 26 638 3.94 0.299 226 ... ... 900 334 365 6 705 0.78 0.536 23Y 28 Jul 31 Jul 3,200 103 301 297 701 0.22 0.149
4-6 August 1976 24 Y ... ... . 1,255 90 386 106 582 0.46 0.527 25 Y -- ... ... . 1,385 486 118 187 791 0.57 0.608 26 Y ... ... . 2,827 98 235 158 491 0.17 0.340 276 ... ... . 452 19 648 16 683 1.51 0.399 286 ... ... . 560 17 899 0 916 1.64 0.577 296 ... ... . 346 33 547 0 580 1.68 0.436 30Y ... ... . 1,650 184 96 21 301 0.18 0.210 316 ... ... . 127 8 497 18 523 4.12 0.374 326 ... ... . 95 0 683 15 698 7.35 0.325 33 c ... ... . 113 143 295 26 464 4.11 0.348 346 ... ... . 32 0 571 0 571 17.84 0.333 356 ... ... . 127.2 4 422 15 441 3.47 0.245 366 ... ... . 452 102 594 23 719 1.59 0.635 376 ... ... . 300 132 198 0 330 1.07 0.229 386 ... ... . 195 135 251 0 386 1.98 0.315 396 ... ... . 254 0 815 0 815 3.21 0.402 406 ... ... . 360 97 425 3 525 5.25 0.551 416 . ... . . . 396 0 745 0 745 1.88 0.421 426 . . . . . . 516 0 574 8 582 1.13 0.311 43 Y . . . . . . 900 130 203 44 377 0.42 0.127 446 . . . . . . 314 0 628 0 628 2.00 0.319 456 . . . . . . . 360 34 375 0 409 1.14 0.428 466 . . . . . . 213 0 1,086 2 1,088 5.11 1.137 47. . . . . . . 325 0 823 8 831 2.56 0.853
1 After reduction of flowers by 50% on the adjacent territories 86 and 96, one bird (216) took the densest stands of flowers on both territories and the other bird apparently abandoned its territory. Neighboring residents also expanded their territories somewhat to incorporate the remaining flowers and area of the 2 original territories.
2 After a feeder was placed in the territory and it was being utilized frequently by the resident. 3 After reduction of flowers by -50c.
stant within and between years for local populations of plants, so that it is likely that this difference reflects differences in the economics of territorial defense that are the result of differences in the average density of
flowers on territories or some unmeasured parameter such as intruder pressure.
Because flowers secrete nectar continuously and the average rate of secretion is known, the amount of nec-
Early Spring 1978 TERRITORIALITY OF HUMMINGBIRDS 289
10,000
5,000
- 9 \\ A= 738 D-'O
-1,000
N A=649D082 V /
. V) 500 E 9
cr :cd C1 0
. , , . * . 4
100-
50
01 0.5 10 5 10 50
FLOWER DENSITY (NUMBER/M2
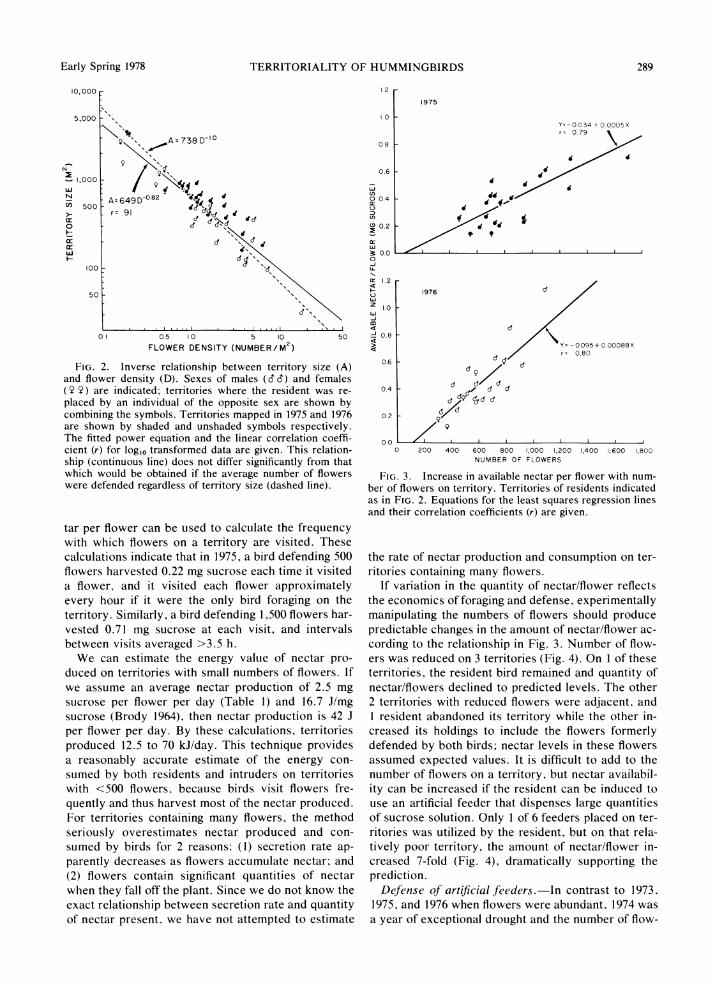
FIG. 2. Inverse relationship between territory size (A) and flower density (D). Sexes of males (cc) and females (Y Y) are indicated; territories where the resident was re- placed by an individual of the opposite sex are shown by combining the symbols. Territories mapped in 1975 and 1976 are shown by shaded and unshaded symbols respectively. The fitted power equation and the linear correlation coeffi- cient (r) for log10 transformed data are given. This relation- ship (continuous line) does not differ significantly from that which would be obtained if the average number of flowers were defended regardless of territory size (dashed line).
tar per flower can be used to calculate the frequency with which flowers on a territory are visited. These calculations indicate that in 1975, a bird defending 500 flowers harvested 0.22 mg sucrose each time it visited a flower, and it visited each flower approximately every hour if it were the only bird foraging on the territory. Similarly, a bird defending 1,500 flowers har- vested 0.71 mg sucrose at each visit, and intervals between visits averaged >3.5 h.
We can estimate the energy value of nectar pro- duced on territories with small numbers of flowers. If we assume an average nectar production of 2.5 mg sucrose per flower per day (Table 1) and 16.7 J/mg sucrose (Brody 1964), then nectar production is 42 J per flower per day. By these calculations, territories produced 12.5 to 70 kJ/day. This technique provides a reasonably accurate estimate of the energy con- sumed by both residents and intruders on territories with <500 flowers, because birds visit flowers fre- quently and thus harvest most of the nectar produced. For territories containing many flowers, the method seriously overestimates nectar produced and con- sumed by birds for 2 reasons: (1) secretion rate ap- parently decreases as flowers accumulate nectar; and (2) flowers contain significant quantities of nectar when they fall off the plant. Since we do not know the exact relationship between secretion rate and quantity of nectar present, we have not attempted to estimate
12 -
1975
10 O Y=-0 034 + 0 0005X r= 0.79
0.6 _
0 0.4 _
(0.2
3.10.0 0 _J
< 1.2
1976 dr
Z 1.0 _
< /l
J 0.8
Y = -0.095 + 0.00088 X r= 0.80
0.6
0.4 _d0 o d
0.2 - ?
/9
0.0 LI I
0 200 400 600 800 1,000 1,200 1,400 1,600 1,800 NUMBER OF FLOWERS
FIG. 3. Increase in available nectar per flower with num- ber of flowers on territory. Territories of residents indicated as in FIG. 2. Equations for the least squares regression lines and their correlation coefficients (r) are given.
the rate of nectar production and consumption on ter- ritories containing many flowers.
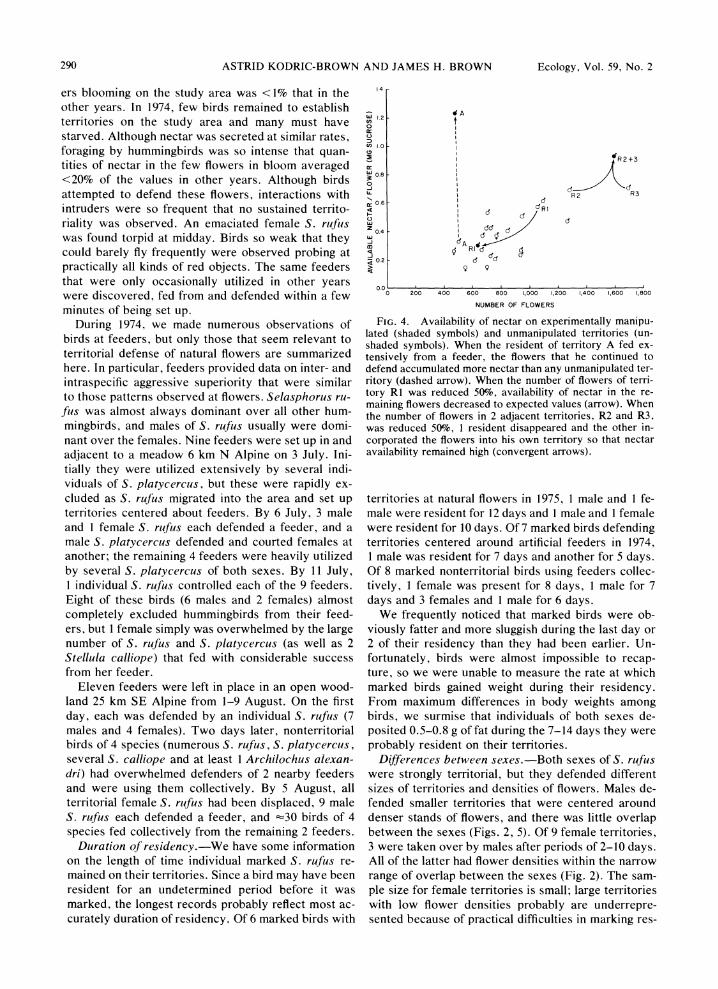
If variation in the quantity of nectar/flower reflects the economics of foraging and defense, experimentally manipulating the numbers of flowers should produce predictable changes in the amount of nectar/flower ac- cording to the relationship in Fig. 3. Number of flow- ers was reduced on 3 territories (Fig. 4). On I of these territories, the resident bird remained and quantity of nectar/flowers declined to predicted levels. The other 2 territories with reduced flowers were adjacent, and I resident abandoned its territory while the other in- creased its holdings to include the flowers formerly defended by both birds; nectar levels in these flowers assumed expected values. It is difficult to add to the number of flowers on a territory, but nectar availabil- ity can be increased if the resident can be induced to use an artificial feeder that dispenses large quantities of sucrose solution. Only I of 6 feeders placed on ter- ritories was utilized by the resident, but on that rela- tively poor territory, the amount of nectar/flower in- creased 7-fold (Fig. 4), dramatically supporting the prediction.
Defense of artificial feeders.-In contrast to 1973, 1975, and 1976 when flowers were abundant, 1974 was a year of exceptional drought and the number of flow-
290 ASTRID KODRIC-BROWN AND JAMES H. BROWN Ecology, Vol. 59, No. 2
ers blooming on the study area was <1% that in the other years. In 1974, few birds remained to establish territories on the study area and many must have starved. Although nectar was secreted at similar rates, foraging by hummingbirds was so intense that quan- tities of nectar in the few flowers in bloom averaged <20% of the values in other years. Although birds attempted to defend these flowers, interactions with intruders were so frequent that no sustained territo- riality was observed. An emaciated female S. rufus was found torpid at midday. Birds so weak that they could barely fly frequently were observed probing at practically all kinds of red objects. The same feeders that were only occasionally utilized in other years were discovered, fed from and defended within a few minutes of being set up.
During 1974, we made numerous observations of birds at feeders, but only those that seem relevant to territorial defense of natural flowers are summarized here. In particular, feeders provided data on inter- and intraspecific aggressive superiority that were similar to those patterns observed at flowers. Selasphorus ru- fus was almost always dominant over all other hum- mingbirds, and males of S. rufus usually were domi- nant over the females. Nine feeders were set up in and adjacent to a meadow 6 km N Alpine on 3 July. Ini- tially they were utilized extensively by several indi- viduals of S. platycercus, but these were rapidly ex- cluded as S. rufus migrated into the area and set up territories centered about feeders. By 6 July, 3 male and I female S. rufus each defended a feeder, and a male S. platycercus defended and courted females at another; the remaining 4 feeders were heavily utilized by several S. platycercus of both sexes. By 11 July, I individual S. rufus controlled each of the 9 feeders. Eight of these birds (6 males and 2 females) almost completely excluded hummingbirds from their feed- ers, but I female simply was overwhelmed by the large number of S. rufus and S. platycercus (as well as 2 Stellula calliope) that fed with considerable success from her feeder.
Eleven feeders were left in place in an open wood- land 25 km SE Alpine from 1-9 August. On the first day, each was defended by an individual S. rufus (7 males and 4 females). Two days later, nonterritorial birds of 4 species (numerous S. rufus, S. platycercus, several S. calliope and at least I Archilochus alexan- dri) had overwhelmed defenders of 2 nearby feeders and were using them collectively. By 5 August, all territorial female S. rufus had been displaced, 9 male S. rufus each defended a feeder, and -30 birds of 4 species fed collectively from the remaining 2 feeders.
Duration of residency.-We have some information on the length of time individual marked S. rufus re- mained on their territories. Since a bird may have been resident for an undetermined period before it was marked, the longest records probably reflect most ac- curately duration of residency. Of 6 marked birds with
1.4
1.2 A
o t
( 1.0 I
'R2+3
w 0.8
o
,J I
tz d dR A R LL ~ ~~~~~~~~d R2 R3 tr0,6 -R
I d~~~~~CR
w d~~~~~ 4 RI d 6 <
0.2 v l f
4 ~~~~~~9 9
0.c 0 200 400 600 800 1,000 1,200 1,400 1,600 1,800
NUMBER OF FLOWERS
FIG. 4. Availability of nectar on experimentally manipu- lated (shaded symbols) and unmanipulated territories (un- shaded symbols). When the resident of territory A fed ex- tensively from a feeder, the flowers that he continued to defend accumulated more nectar than any unmanipulated ter- ritory (dashed arrow). When the number of flowers of terri- tory RI was reduced 50%, availability of nectar in the re- maining flowers decreased to expected values (arrow). When the number of flowers in 2 adjacent territories, R2 and R3, was reduced 50%, 1 resident disappeared and the other in- corporated the flowers into his own territory so that nectar availability remained high (convergent arrows).
territories at natural flowers in 1975, 1 male and I fe- male were resident for 12 days and I male and I female were resident for 10 days. Of 7 marked birds defending territories centered around artificial feeders in 1974, 1 male was resident for 7 days and another for 5 days. Of 8 marked nonterritorial birds using feeders collec- tively, 1 female was present for 8 days, I male for 7 days and 3 females and I male for 6 days.
We frequently noticed that marked birds were ob- viously fatter and more sluggish during the last day or 2 of their residency than they had been earlier. Un- fortunately, birds were almost impossible to recap- ture, so we were unable to measure the rate at which marked birds gained weight during their residency. From maximum differences in body weights among birds, we surmise that individuals of both sexes de- posited 0.5-0.8 g of fat during the 7-14 days they were probably resident on their territories.
Differences between sexes.-Both sexes of S. rufus were strongly territorial, but they defended different sizes of territories and densities of flowers. Males de- fended smaller territories that were centered around denser stands of flowers, and there was little overlap between the sexes (Figs. 2, 5). Of 9 female territories, 3 were taken over by males after periods of 2-10 days. All of the latter had flower densities within the narrow range of overlap between the sexes (Fig. 2). The sam- ple size for female territories is small; large territories with low flower densities probably are underrepre- sented because of practical difficulties in marking res-
Early Spring 1978 TERRITORIALITY OF HUMMINGBIRDS 291
0.70F -Z
1,500 COST OF DEFENDING ADDITIONAL RESOURCES EXCEEDS BENEFIT ;,,.v,-,,-
ZU 5 00 d?e33dO
1 14 Z 7',
O1,000 0 550 0.2350 Li- 0.264r~ 02w -' k ~ , Li- 0,85 Cdo058
o d~~~~~C 043 O.41
m 0.4063~00 .805 043 0.54
Q 3Q03Cf 0.24 40o19 0.5 0.15 D d033 d0O44 0.31_ - z d d0 37 0.26 0.34 500 - o5100339
0.25 c d0.43 0.13 032
0.23 _0.21 ''
;;>A?7,~~Ay?IN SUFFICIENT RESOURCES TO SUPPORT TERRITORIAL INDIVIDUAL_'\'7\,\
0 500 1,000 1,500 2,000 2,500 3,000 3,500 TERRITORY SIZE (M2)
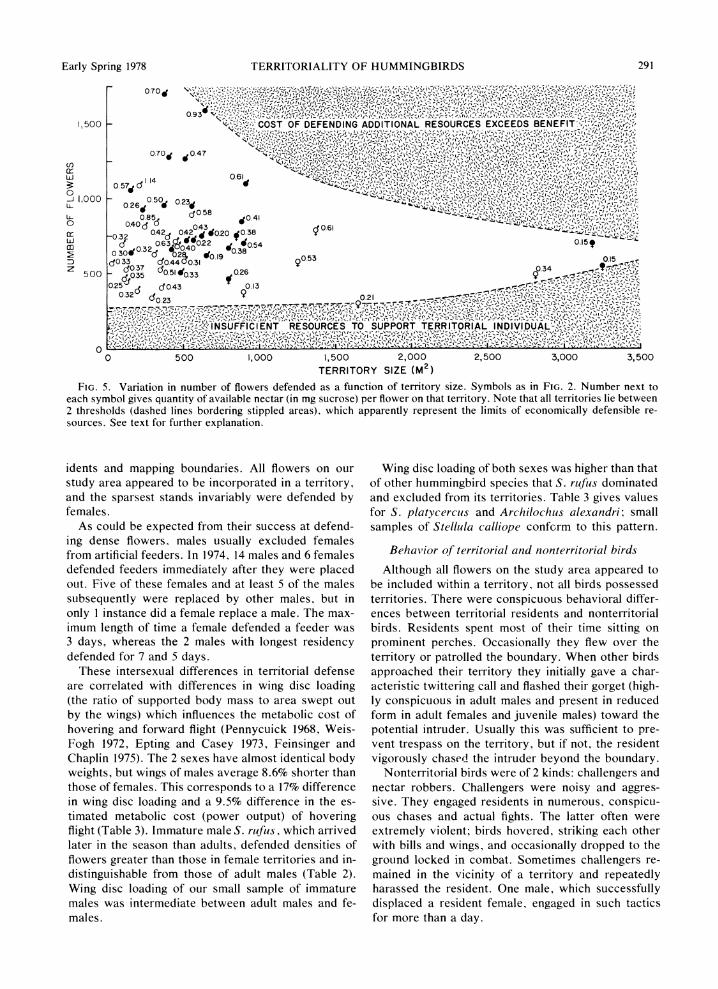
FIG. 5. Variation in number of flowers defended as a function of territory size. Symbols as in FIG. 2. Number next to each symbol gives quantity of available nectar (in mg sucrose) per flower on that territory. Note that all territories lie between 2 thresholds (dashed lines bordering stippled areas), which apparently represent the limits of economically defensible re- sources. See text for further explanation.
idents and mapping boundaries. All flowers on our study area appeared to be incorporated in a territory, and the sparsest stands invariably were defended by females.
As could be expected from their success at defend- ing dense flowers, males usually excluded females from artificial feeders. In 1974, 14 males and 6 females defended feeders immediately after they were placed out. Five of these females and at least 5 of the males subsequently were replaced by other males, but in only I instance did a female replace a male. The max- imum length of time a female defended a feeder was 3 days, whereas the 2 males with longest residency defended for 7 and 5 days.
These intersexual differences in territorial defense are correlated with differences in wing disc loading (the ratio of supported body mass to area swept out by the wings) which influences the metabolic cost of hovering and forward flight (Pennycuick 1968, Weis- Fogh 1972, Epting and Casey 1973, Feinsinger and Chaplin 1975). The 2 sexes have almost identical body weights, but wings of males average 8.6% shorter than those of females. This corresponds to a 17% difference in wing disc loading and a 9.5% difference in the es- timated metabolic cost (power output) of hovering flight (Table 3). Immature male S. rufus, which arrived later in the season than adults, defended densities of flowers greater than those in female territories and in- distinguishable from those of adult males (Table 2). Wing disc loading of our small sample of immature males was intermediate between adult males and fe- males.
Wing disc loading of both sexes was higher than that of other hummingbird species that S. rufus dominated and excluded from its territories. Table 3 gives values for S. platycercus and Archilochus alexandri; small samples of Stellula calliope conform to this pattern.
Behavior of territorial and nonterritorial birds
Although all flowers on the study area appeared to be included within a territory, not all birds possessed territories. There were conspicuous behavioral differ- ences between territorial residents and nonterritorial birds. Residents spent most of their time sitting on prominent perches. Occasionally they flew over the territory or patrolled the boundary. When other birds approached their territory they initially gave a char- acteristic twittering call and flashed their gorget (high- ly conspicuous in adult males and present in reduced form in adult females and juvenile males) toward the potential intruder. Usually this was sufficient to pre- vent trespass on the territory, but if not, the resident vigorously chased the intruder beyond the boundary.
Nonterritorial birds were of 2 kinds: challengers and nectar robbers. Challengers were noisy and aggres- sive. They engaged residents in numerous, conspicu- ous chases and actual fights. The latter often were extremely violent; birds hovered, striking each other with bills and wings, and occasionally dropped to the ground locked in combat. Sometimes challengers re- mained in the vicinity of a territory and repeatedly harassed the resident. One male, which successfully displaced a resident female, engaged in such tactics for more than a day.

292 ASTRID KODRIC-BROWN AND JAMES H. BROWN Ecology, Vol. 59, No. 2
TABLE 3. Measurements of wing length and body weight and calculated wing disc loading and power output (in milliwatts) for hovering flight for 3 species of hummingbirds
Wing disc Wing length Body wt loading1 Power output2
Species Sex (cm) (g) (g/mm2) (J/h)
Selasphorus rufus 4.07 (118)3 3.5 (13) .044 407 [= 113.1 mW] Y Y 4.42 (97) 3.5 (13) .038 372 [= 103.3 mW]
Selasphorus platycercus4 4.91 (12) 3.4 (39) .031 335 [= 93.1 mW] Y Y 5.15 (10) 3.8 (21) .031 367 [ 101.9 mW]
Archilochus alexandri5 4.25 (27) 3.1 (15) .037 332 [= 98.2 mW] Y Y 4.59 (22) 3.3 (16) .034 342 [= 95.0 mW]
Wing disc loading (LWD) calculated from wing length (1) and body weight (W) using the equation LWD W/[l7r(l + 0.404 106)2](Feinsinger et al. in press; Greenewalt 1975).
2 Power output for hovering flight (PHOV) calculated from wing disc loading (LwD), body weight (W), and air density (p) using the equation PHoV = 19.44 W(LwD/p)05, where p was assumed to equal 0.0122 (Feinsinger et al. in press).
3Sample sizes are given in parentheses. 4Includes body wt of 39 d d and 21 Y Y from N. M. Waser (personal communication). 5 Includes wing lengths for 20 d d and 20 Y Y and body wt for 15 d d and 16 Y Y from Stiles (1973).
In contrast to residents and challengers, nectar rob- bers were submissive and inconspicuous. When tres- passing they rarely vocalized, consistently remained near the ground and vegetative cover, and usually fed on flowers far from the perch of the resident. Although robbers sometimes perched for long periods and vis- ited many flowers during several feeding bouts, they fled immediately when discovered and chased by the resident. Although adult males occasionally behaved in this manner, they seldom were successful robbers, probably because of their conspicuous color and dis- tinctive sound while in flight. Most robbers were fe- males and (later in the season) juveniles. Because of their secretive behavior, robbers were difficult to ob- serve and unfortunately we have no estimates of their abundance or their impact on nectar resources.
DISCUSSION
Economics of territoriality
In recent years, much emphasis has been placed on the economics of territoriality, and nectar-feeding birds frequently have been used for empirical studies because of the relative ease with which the costs and benefits of their territorial defense can be quantified. Most previous workers have attempted to evaluate the economics of territoriality by determining time-energy budgets for territorial (and sometimes nonterritorial) individuals (e.g. Stiles 1971, 1973, Wolf and Hains- worth 1971, Gill and Wolf 1975, Wolf 1975, Carpenter and MacMillen 1976a, b). We have used a different approach; it is more indirect, but has certain advan- tages. We assume that the hummingbird territories we observe are profitable for their owners. This is rea- sonable because the small size and high metabolic rate of hummingbirds prevents them from surviving for long if energy expenditure exceeds income; birds with uneconomical territories should abandon them and either become nonterritorial or move to another area
where resources can be defended economically. Therefore, variability in size of territory and number and density of flowers is assumed to indicate the range of conditions permitting economical defense. Analysis of variation in these parameters and in the quantity of nectar accumulated in flowers should help elucidate the adaptive strategies by which individuals econom- ically defend diverse resources.
Territories vary greatly in size and number of flow- ers, but this variation is confined within definite limits that appear to be determined by economic constraints (Fig. 5). A minimum of :300 flowers is required to support a territorial bird when flowers are dense. The threshold number of flowers to support a resident in- creases with territory size, so that the largest territo- ries contain a minimum of :500 flowers. This increase presumably reflects increasing costs for the resident as territory size increases. Distances flown while for- aging and repelling intruders should increase with ter- ritory size. The fact that the largest territories are oc- cupied exclusively by females, which have less costly flight but inferior aggressive ability in comparison to males, suggests that the costs of foraging become of primary importance when flower densities are low. There is also an upper threshold number of flowers above which cost of defense apparently exceeds ben- efit. This threshold value decreases rapidly with ter- ritory size, presumably because distance of foraging and aggressive flights (and perhaps also the frequency of successful nectar robbing by intruders) increases rapidly with area.
Between the limits defined by the thresholds is a range of territory sizes and number of flowers that is economically defensible (Fig. 5). There is great vari- ation in the number of flowers in small territories, but this variability decreases rapidly with increasing ter- ritory size. When flowers are dense, birds can obtain the benefits of increased amounts of nectar per flower by increasing the number of flowers defended with
Early Spring 1978 TERRITORIALITY OF HUMMINGBIRDS 293
,CL CH
., - ,,'B z w m
0
0
X,,
0
a7RL>
/ -)RH
NUMBER OF FLOWERS DEFENDED
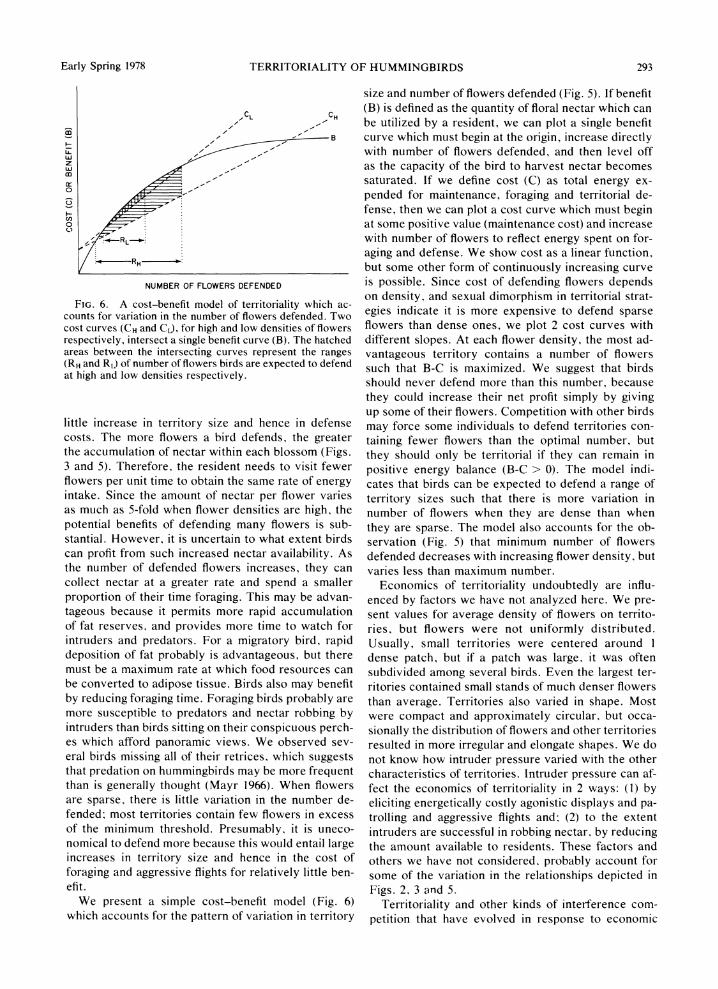
FIG. 6. A cost-benefit model of territoriality which ac- counts for variation in the number of flowers defended. Two cost curves (CH and CL). for high and low densities of flowers respectively, intersect a single benefit curve (B). The hatched areas between the intersecting curves represent the ranges (RH and RL) of number of flowers birds are expected to defend at high and low densities respectively.
little increase in territory size and hence in defense costs. The more flowers a bird defends, the greater the accumulation of nectar within each blossom (Figs. 3 and 5). Therefore, the resident needs to visit fewer flowers per unit time to obtain the same rate of energy intake. Since the amount of nectar per flower varies as much as 5-fold when flower densities are high, the potential benefits of defending many flowers is sub- stantial. However, it is uncertain to what extent birds can profit from such increased nectar availability. As the number of defended flowers increases, they can collect nectar at a greater rate and spend a smaller proportion of their time foraging. This may be advan- tageous because it permits more rapid accumulation of fat reserves, and provides more time to watch for intruders and predators. For a migratory bird, rapid deposition of fat probably is advantageous, but there must be a maximum rate at which food resources can be converted to adipose tissue. Birds also may benefit by reducing foraging time. Foraging birds probably are more susceptible to predators and nectar robbing by intruders than birds sitting on their conspicuous perch- es which afford panoramic views. We observed sev- eral birds missing all of their retrices, which suggests that predation on hummingbirds may be more frequent than is generally thought (Mayr 1966). When flowers are sparse, there is little variation in the number de- fended; most territories contain few flowers in excess of the minimum threshold. Presumably, it is uneco- nomical to defend more because this would entail large increases in territory size and hence in the cost of foraging and aggressive flights for relatively little ben- efit.
We present a simple cost-benefit model (Fig. 6) which accounts for the pattern of variation in territory
size and number of flowers defended (Fig. 5). If benefit (B) is defined as the quantity of floral nectar which can be utilized by a resident, we can plot a single benefit curve which must begin at the origin, increase directly with number of flowers defended, and then level off as the capacity of the bird to harvest nectar becomes saturated. If we define cost (C) as total energy ex- pended for maintenance, foraging and territorial de- fense, then we can plot a cost curve which must begin at some positive value (maintenance cost) and increase with number of flowers to reflect energy spent on for- aging and defense. We show cost as a linear function, but some other form of continuously increasing curve is possible. Since cost of defending flowers depends on density, and sexual dimorphism in territorial strat- egies indicate it is more expensive to defend sparse flowers than dense ones, we plot 2 cost curves with different slopes. At each flower density, the most ad- vantageous territory contains a number of flowers such that B-C is maximized. We suggest that birds should never defend more than this number, because they could increase their net profit simply by giving up some of their flowers. Competition with other birds may force some individuals to defend territories con- taining fewer flowers than the optimal number, but they should only be territorial if they can remain in positive energy balance (B-C > 0). The model indi- cates that birds can be expected to defend a range of territory sizes such that there is more variation in number of flowers when they are dense than when they are sparse. The model also accounts for the ob- servation (Fig. 5) that minimum number of flowers defended decreases with increasing flower density, but varies less than maximum number.
Economics of territoriality undoubtedly are influ- enced by factors we have not analyzed here. We pre- sent values for average density of flowers on territo- ries, but flowers were not uniformly distributed. Usually, small territories were centered around I dense patch, but if a patch was large, it was often subdivided among several birds. Even the largest ter- ritories contained small stands of much denser flowers than average. Territories also varied in shape. Most were compact and approximately circular, but occa- sionally the distribution of flowers and other territories resulted in more irregular and elongate shapes. We do not know how intruder pressure varied with the other characteristics of territories. Intruder pressure can af- fect the economics of territoriality in 2 ways: (1) by eliciting energetically costly agonistic displays and pa- trolling and aggressive flights and; (2) to the extent intruders are successful in robbing nectar, by reducing the amount available to residents. These factors and others we have not considered, probably account for some of the variation in the relationships depicted in Figs. 2, 3 and 5.
Territoriality and other kinds of interference com- petition that have evolved in response to economic

294 ASTRID KODRIC-BROWN AND JAMES H. BROWN Ecology, Vol. 59, No. 2
tradeoffs in resource utilization can result in food lim- iting populations such that some individuals can starve while others have access to food in excess of their needs. This probably happens in our study area, at least in years of average to low flower abundance. Many individuals of S. rufus are unable to secure ter- ritories, while others defend territories containing sev- eral times the amount of nectar required to support a resident and enable it to deposit sufficient fat to con- tinue migration. For territorial organisms, it probably is unnecessary to invoke group selection (Wynne-Ed- wards 1962) or special mechanisms other than selec- tion for individual economic benefit to account for the apparent "self regulation" of populations below the limit determined by available resources.
Wing disc loading, sexual dimorphism and the strategy of migration
Territoriality enables migrant individuals of S. rufus to gain access to floral resources and replenish lipid reserves. This species has the longest migratory route of any North American hummingbird. Its migration follows the blooming of hummingbird-specialized flowers (Grant and Grant 1967, Gass et al. 1976). From its winter range in southern Mexico, it migrates north along the Pacific Coast where lowland and foothill flowers bloom in response to winter rains. It breeds from Oregon and Idaho to southern Alaska and the Yukon. Its southward migration in late summer fol- lows inland mountain ranges where flowers bloom at intermediate to high elevations. The pattern of migra- tion appears to consist of a series of long flights inter- rupted by intense foraging that usually involves terri- toral defenses of dense flowers for periods of several days (Armitage 1955, Grant and Grant 1967, 1968, Dunford and Dunford 1972, Gass et al. 1976). Long- distance migratory flights may be a common charac- teristic of temperate-zone hummingbirds. Archilochus colubris is known to fly =1,000 km across the Gulf of Mexico (Lasiewski 1962, 1963), and the exhausted, emaciated appearance that we observed in newly ar- rived S. rufus (and also Stellula calliope) suggests that other species may fly comparable distances.
Migrant hummingbirds face the problem of compet- ing for floral resources not only with other members of their own species but also with breeding and resi- dent hummingbird species during migration and on the wintering ground. Such competition may be particu- larly severe during migration because at each stop the energy-depleted individual must rapidly and effective- ly compete with established birds which are utilizing the local flowers. This may account for the apparent paradox that males of S. rufus, the species with the longest migratory route, have the highest wing disc loading and hence, the energetically most costly flight reported for any North American hummingbird (Fein- singer and Chaplin 1975, present study: Feinsinger and Chaplin [1975] give a higher value of wing disc loading
for male Selasphorsus sasin [0.0466 g/cm2] than for male S. rufus. However, they used body-weight data for captive individuals of S. sasin, and such birds tend to be more obese than free-living ones. Recalculating using data from Stiles [1973] gives a value of 0.0403 g/cm2 for male S. sasin, which is significantly lower than our value [0.0428] for male S. rufus).
Several authors have noted that metabolic cost of both hovering and forward flight varies directly with wing disc loading (Pennycuick 1968, Hainsworth and Wolf 1972, Weis-Fogh 1972, Epting and Casey 1973, Greenewalt 1975). Feinsinger and Chaplin (1975), however, point out that there is a tradeoff between energetically efficient flight and aggressive ability. They note that territorial hummingbird species have higher wing disc loading than traplining species, and they suggest that selection for speed and maneuver- ability in aerial encounters has favored the evolution of relatively short wings in territorial hummingbirds. Our observations support this pattern. Both sexes of S. rufus have higher wing disc loading than the 2 spe- cies of breeding hummingbird (S. platycercus and A. alexandri) that are displaced from flower patches and feeding stations when aggressively territorial S. rufus migrated in our study area. We suggest that S. rufus has sacrificed efficient flight for aggressive ability, en- abling it to compete successfully with resident hum- mingbirds during migration and perhaps also on the wintering ground.
It is interesting that the virtually absolute domi- nance of S. rufus over resident hummingbird species observed at our study area is not characteristic of its interspecific competitive ability in other habitats. Our study area was located at intermediate elevation and contained relatively dense flowers. Observations in other habitats indicate that S. platycercus is not com- pletely excluded by S. rufus at high elevations (>2,500 m) nor is A. alexandri completely excluded at low el- evations (>1,500 m). We suggest that at high eleva- tions, low air density (which increases flight costs; Feinsinger et al., in press) and low temperature (which increases total metabolic costs) increase the advantages of efficient flight and enable S. platycercus to compete successfully with S. rufus. In the desert habitats at low elevations, flowers tend to be sparsely distributed which favors their exploitation by A. al- exandri with its low wing disc loading. It appears that differences in aggressive ability and foraging behavior, which are related to differences in wing disc loading, play a major role in mediating competition among tem- perate North American hummingbird species, and these may be particularly important during migration. It is during migration that these species, which have largely nonoverlapping breeding ranges but are all about the same size and able to utilize the same flower species, come into greatest contact and have greatest potential for direct competition.
The sexes of S. rufus differ conspicuously in wing

Early Spring 1978 TERRITORIALITY OF HUMMINGBIRDS 295
disc loading and in size of territory and density of flowers defended. Males apparently use the aerial agility conferred by their short wings to aggressively defend flowers that are sufficiently dense to pay their high foraging and defense costs. Females, because their longer wings permit more efficient flight, are able to forage and defend flowers that are too sparse to be defended economically by males. The result is subdi- vision of floral resources on the basis of density with only a narrow overlap between the sexes (Figs. 2, 5). Most of the successful nectar robbers which we ob- served were females. Feinsinger and Chaplin (1975) have reported other examples of intersexual differ- ences in foraging and territorial behavior, in hum- mingbirds, that are correlated with differences in wing disc loading.
The adaptive basis of sexual dimorphism in wing disc loading in S. rufus is not clear. Although these differences provide the basis for resource subdivision and consequent reduction of competition during mi- gration, it is likely that selective pressures related to the different roles of the sexes during the breeding season are primarily responsible for the evolution and maintenance of sexual dimorphism. We suggest 2 pos- sibilities. First, males may be selected for high wing disc loading because they aggressively defend terri- tories of dense flower patches that are used for court- ship and mating. Superiority in aerial aggressive en- counters conferred by relatively short wings may increase reproductive success by affecting the out- come of intrasexual competition for mates. Evidence against this hypothesis is the observation that the re- lated species, S. platycercus, has similar territorial breeding behavior, but low wing disc loading in both sexes (Table 3). We favor the alternative hypothesis that low wing disc loading is advantageous to female S. rufus on the breeding grounds. For a period of sev- eral weeks, activity of females must be centered around the nest. Because the abundance of flowers in nesting habitats of S. rufus may be low (W.A. Calder, personal communication), energetically efficient flight should be particularly advantageous to females while they are nesting and feeding young. We suggest that wing disc loading of females reflects a compromise between selection for efficient flight during the breed- ing season and selection for aggressive territoriality during migration. Because males do not participate in incubation or care of young, they are not subject to this compromise and can respond to a much greater extent to selection for aggressive ability.
Feeding territories of nectarivorous birds
Selasphorus rufus resembles many other humming- birds, African sunbirds, and Hawaiian honeycreepers in defending feeding territories during the nonbreeding season (Pitelka 1942, Grant and Grant 1968, Wolf 1969, 1970, Stiles and Wolf 1970, Wolf and Hainsworth 1971, Stiles 1973, Gill and Wolf 1975, Feinsinger and
Chaplin 1975, Carpenter 1976, Feinsinger 1976, Car- penter and MacMillen 1976a,b). These feeding terri- tories have been investigated intensively because they provide excellent systems for field studies of avian behavior, territorial economics, and plant-pollinator interaction. Results of these studies invite searches for common patterns and mechanisms.
Although some hummingbirds use traplines to ex- ploit sparse floral resources and apparently are not strongly territorial, many species aggressively defend feeding territories. Feinsinger and Chaplin (1975; see also Feinsinger et al., in press) pointed out the rela- tionship between wing disc loading and foraging and territorial behavior that appears to be widespread in hummingbirds and to account for the interspecific and intersexual interactions of S. rufus. Several authors (e.g., Pitelka 1942, Cody 1968, Stiles 1973, Feinsinger 1976) have discussed the prominent role of interspe- cific aggression and territoriality in subdividing food resources among coexisiting hummingbird species. Gass et al. (1976) used an approach similar to ours to study territoriality of migrant S. rufus. Although they did not comment on many of the patterns reported in this paper, they observed precise regulation of territo- rial size in response to flower density (Fig. 2) virtually identical to that shown in Fig. 4 of their paper. It is interesting that only females and juvenile S. rufus uti- lized their study area in northwest California during July and August, but these birds defended small ter- ritories with high densities of flowers comparable to those defended by adult males on our study site.
Sunbirds in the Old World and honeycreepers in Hawaii have coevolved mutualistic relationships with specialized flowers that they pollinate while foraging for nectar. Convergent similarities between these sys- tems and the hummingbird-plant associations of the New World include frequent defense of feeding terri- tories (Gill and Wolf 1975, Carpenter and MacMillen 1976a,b). Thus, Gill and Wolf (1975) report that the African sunbird Nectarinia reichenowi defended feed- ing territories varying in size from 6.7 to 2,300 m2 and containing 1,000 to 5,000 flowers of Leonotis nepeti- folia; these flowers produced -1 mg of sucrose/(flow- er-day) or 17 to 85 kJ/(territory-day). These values can be compared to those we obtained for S. rufus, which defended territories varying in size from 32 to 3,200 m2 and containing from 300 to 1,700 flowers of 3 spe- cies; these flowers secreted -2.5 mg of sucrose/(flow- er-day) or 12.5 to 70 kJ/(territory-day). Although these 2 coevolved mutualistic systems of nectar-feeding, ter- ritorial birds and bird-pollinated flowers appear to be convergent in many respects, several differences are apparent. Nectarinia reichenowi (body weight, 15g) is -4x larger than S. rufus and, like other sunbirds, it differs from hummingbirds in rarely hovering while foraging.
The striking similarity between the pattern of terri- tory sizes and number of flowers defended by S. rufus

296 ASTRID KODRIC-BROWN AND JAMES H. BROWN Ecology, Vol. 59, No. 2
(Fig. 5) and comparable data for African sunbirds (Gill and Wolf 1975; see Fig. 4 which, however, is not dis- cussed in the same context) suggests that these evo- lutionarily convergent territorial systems are governed by similar economic constraints. These similarities and others mentioned above indicate that it may be possible to develop general models of territorial eco- nomics which are widely applicable to nectarivorous birds and perhaps to other organisms as well.
ACKNOWLEDGMENTS
We thank Kevin and Karen Brown for assistance and com- panionship in the field, R. Vestal for help with data analysis, S. B. Chaplin, P. Feinsinger, C. L. Gass, F. B. Gill, F. R. Hainsworth and L. L. Wolf for critically reading the manu- script, and numerous colleagues including Z. Abramsky, T. C. Gibson, H. R. Pulliam and N. M. Waser for valuable discussions. The study was supported in large part by the National Science Foundation (Grants GB 39260 and DEB 76- 09499).
LITERATURE CITED
Armitage, K. 1955. Territorial behavior in fall migrant Ru- fous Hummingbirds. Condor 57:239-240.
Brody, S. 1964. Bioenergetics and growth. Hafner, New York, New York, USA.
Brown, J. L. 1964. The evolution of diversity in avian ter- ritorial systems. Wilson Bulletin 76:160-169.
Brown, J. L., and G. Orians. 1970. Spacing patterns in mo- bile animals. Annual Review of Ecology and Systematics 1:239-262.
Carpenter, F. L. 1976. Ecology and evolution of Andean hummingbirds. University of California Publications in Zo- ology 106.
Carpenter, F. L., and R. E. MacMillen. 1976a. Threshold model of feeding territoriality and test with a Hawaiian Honeycreeper. Science 194:639-642.
- 1976b. Energetic cost of feeding territories in a Hawaiian Honeycreeper. Oecologia 26:213-223.
Cody, M. L. 1968. Interspecific territoriality among hum- mingbird species. Condor 70:270-271.
Dunford, C., and E. Dunford. 1972. Interspecific aggression of resident Broad-tailed and migrant Rufous Humming- birds. Condor 74:479.
Epting, R. J., and T. M. Casey. 1973. Power output and wing disc loading in hovering hummingbirds. American Naturalist 107:761-765.
Feinsinger, P. 1976. Organization of a tropical guild of nec- tarivorous birds. Ecological Monographs 46:257-291.
Feinsinger, P., and S. B. Chaplin. 1975. On the relationship between wing disc loading and foraging strategy in hum- mingbirds. American Naturalist 109:217-224.
Feirsinger, P., R. K. Colwell, J. Terborgh, and S. B. Chap- lin. In press. Elevation and the morphology, flight ener- getics, and foraging ecology of tropical hummingbirds. American Naturalist.
Gass, C. L., G. Angeher, and J. Centa. 1976. Regulation of food supply by feeding territoriality in the Rufous Hum- mingbird. Canadian Journal of Zoology 54:2046-2054.
Gill, F. B., and L. L. Wolf. 1975. Economics of feeding territoriality in the Golden-Winged Sunbird. Ecology 56:333-345.
Grant, K. A., and V. Grant. 1967. Effects of hummingbird migration on plant speciation in the California flora. Evo- lution 21:457-465.
. 1968. Hummingbirds and their flowers. Columbia University Press, New York, New York, USA.
Greenewalt, C. H. 1975. The flight of birds. Transactions of the American Philosophical Society 65:1-67.
Hainsworth, F. R. 1974. Food quality and foraging efficien- cy: the efficiency of sugar assimilation by hummingbirds. Journal of Comparative Physiology 88:425-431.
Hainsworth, F. R., and L. L. Wolf. 1972. Power for hov- ering flight in relation to body size in hummingbirds. Amer- ican Naturalist 106:589-596.
Lasiewski, R. C. 1962. The capture and maintenance of hummingbirds for experimental purposes. Avicultural Magazine 68:59-64.
1963. Oxygen consumption of torpid, resting, active and flying hummingbirds. Physiological Zoology 36:122- 140.
Mayr, E. 1966. Hummingbird caught by Sparrowhawk. Auk 83:664.
Pearson, 0. P. 1954. The daily energy requirements of a wild Anna's Hummingbird. Condor 56:317-322.
Pennycuick, C. J. 1968. Power requirements for horizontal flight in the pigeon. Columba livia. Journal of Experimen- tal Biology 49:527-555.
Phillips, A. R. 1975. The migrations of Allen's and other hummingbirds. Condor 77:196-205.
Pitelka, F. A. 1942. Territoriality and related problems in North American hummingbirds. Condor 44:189-204.
Stiles, F. G. 1971. Time, energy and territoriality of the Anna's Hummingbird (Calypte anna). Science 173:818- 821.
. 1972. Age and sex determination in Rufous and Al- len's hummingbirds. Condor 74:25-32.
1973. Food supply and the annual cycle of the Anna's Hummingbird. University of California Publications in Zoology 97: 1-109.
Stiles, F. G., and L. L. Wolf. 1970. Hummingbird territo- riality at a tropical flowering tree. Auk 87:467-491.
Weis-Fogh, T. 1972. Energetics of hovering flight in hum- mingbirds and in Drosophila. Journal of Experimental Bi- ology 56:79-104.
Wolf, L. L. 1969. Female territoriality in a tropical hum- mingbird. Auk 86:490-504.
. 1970. The impact of seasonal flowering on the bi- ology of some tropical hummingbirds. Condor 72:1-14.
. 1975. Energy intake and expenditures in a nectar- feeding sunbird. Ecology 56:92-104.
Wolf, L. L., and F. R. Hainsworth. 1971. Time and energy budgets of territorial hummingbirds. Ecology 52:980-988.
Wynne-Edwards, C. V. 1962. Animal dispersion in relation to social behavior. Oliver and Boyd, Edinburgh, Scotland.
![leg.wa.govleg.wa.gov/CodeReviser/WACArchive/Documents/2012/WAC-296-826... · (2/17/09) [Ch. 296-826 WAC—p. 1] Chapter 296-826 Chapter 296-826 WAC ANHYDROUS AMMONIA WAC 296-826-100](https://img.pdfslide.net/doc/110x75/5b2b78217f8b9ae6278b475f/legwa-21709-ch-296-826-wacp-1-chapter-296-826-chapter-296-826-wac.jpg)



![Chapter 296-52 WAC - leg.wa.govleg.wa.gov/CodeReviser/WACArchive/Documents/2018/WAC 296 - 52 CHAPTER.… · (8/1/17) [ch. 296-52 wac p. 1] chapter 296-52 chapter 296-52 wac safety](https://img.pdfslide.net/doc/110x75/5e1c55c238ed802015030b5e/chapter-296-52-wac-legwa-296-52-chapter-8117-ch-296-52-wac-p-1.jpg)




![Chapter 296-19A Chapter 296-19A WAC VOCATIONAL …lawfilesext.leg.wa.gov/law/WACArchive/2012/WAC-296... · 296-19A-010 Vocational Rehabilitation [Ch. 296-19A WAC—p. 2] (11/15/11)](https://img.pdfslide.net/doc/110x75/6013de7783e2d5485a5626ed/chapter-296-19a-chapter-296-19a-wac-vocational-296-19a-010-vocational-rehabilitation.jpg)