Embed Size (px)
Citation preview

ECOLOGY AND TAXONOMY OF POTENTIAL AMNESIC SHELLFISH POISING (ASP) CAUSING DIATOM Pseudo-nitzschia
SPECIES (BACILLARIOPHYCEAE) IN MALAYSIAN WATER
Suriyanti Su Nyun Pau
Master of Science . 2011

,.....
Pu~t KbidmarMaklumat Akademik UNIVERSm MALAY IA SARAWAK
:'
ECOLOGY AND TAXONOMY OF POTENTIAL AMNESIC SHELLFISH POISING (ASP) CAUSING DIATOM Pseudo-nitzschia
SPECIES (BACILLARIOPHYCEAE) IN MALAYSIAN WATER F'.KHIDMAT MAKLUMAT AKADEMIK
111111111 IIi'mi 111111111 1000246372
SURlYANTI SU NYUN PAU
A thesis submitted in fulfillment of the requirement for the Degree of
Master of Science (Aquatic Science)
! Faculty of Resources Sciences and Technology UNIVERSITI MALAYSIA SARAWAK
2011
I'

DECLARATION
I hereby declare that no portion of the work referred to this thesis has been submitted
in support of an application for another degree ofqualification to this or any other
university or institution of higher learning.
(Suriyanti Su Nyun Pau)
Date:
I

ACKNOWLEDGEMENTS
First and foremost, I would like to thank Universiti Malaysia Sarawak for giving me
the opportunities to pursue my postgraduate study and providing the Zamalah
Postgraduate Scholarship. lowe the greatest honors to my supervisor Dr. Lim Po Teen
and Dr. Leaw Chui Pin for their guidance and support throughout this study. Without
them, this thesis would be impossible. Thanks to Mr. Jongkar Grinang from IBEC
UNIMAS for his help in the PCA analysis and Fisheries Department for the
accessibility to the sampling site. Not forgetting Ms. Y ong of the Fisheries
Department for the permission to analyze the samples and Prof Dr. Gires Usup from
UKM for sequencing analyses.
I am indebted to the following individuals for their help of various kinds:
Hartina, Hong Chang, Zubaidah, Fareha, Toh Hii, Tung, Soon, the lab members of
Ecotoxicology Lab and lecturers of Aquatic Science Department. My special
appreciation is due to the Aquatic Lab assistants and FRST science officers especial1y
Mr. Nazri, Mr. Besar and Mdm. Ting for their help and hospitality. Last but not least,
my sincere gratitude is dedicated to my family especially Dzulhelmi for his devotion
and courage given throughout my study.
II

...
ABSTRACT
(Amnesic shellfish poisoning (ASP) is a type of human intoxication due to
consumption of contaminated shellfish with algal origin domoic acid (DA). In this
study, seasonal occurrence of Pseudo-nitzschia species in two estuarine waters was
investigated by plankton and water samplings for 30 months period at fortnightly
interval. The abundance of Pseudo-nitzchia was apparently influenced by water
salinity, pH, silicate concentrations and precipitation rates. Pseudo-nitzschia occurred
mainly between April and September but cells were mostly abundant in May. Highest
cell density recorded in Santubong and Sarnariang was 8.0x 103 cells L-1 and 2. 9x 104
cells L-1 respectively. Based on ultrastructural features of Pseudo-nitzschia under
transmission and scanning electron microscopic (TEM and SEM) observations, the
Pseudo-nitzschia species were identified as Pseudo-nitzschia pungens and P.
brasiliana.}he two species were also common in phytoplankton assemblage collected
from twenty one locations in South China Sea and Malacca straits. None of the clonal
cultures of Pseudo-nitzschia was tested positive with domoic acid production by high
performance liquid chromatography (HPLC) fluorometric method. In this study, LSU
rDNA sequences obtained for both species were used to reconstruct the phylogenetic
inference of Pseudo-nitzschia spp. The tree topologies of Pseudo-nitzschia revealed
close relationship of P. brasiliana and P. americana which is congruent with
morphological data, while P. pungens formed a sister pair with P. multiseries. Based
on LSU rDNA multiple sequence alignment, signature regions of LSU rDNA for both
P. brasiliana and P. pungens were determined and potential species-specific rRNA
probes were designed in silico for both species. Overall, findings from this study may
be useful in assisting country phytoplankton monitoring of harmful algal blooms
(HABs) species in Malaysia.
Key words: Pseudo-nitzschia, domoic acid, cell abundance, morphology,
oligonucleotide probes
1Il

I
EKOLOGI DAN TAKSONOMI DIATOM BERPOTENSI PENYEBAB
KERACUNAN KERANG-KERANGAN AMNESIK (ASP) Pseudo-nitzschia
SPESIES (BACILLARIOPHYCEAE) DI PERAIRAN MALAYSIA
ABSTRAK
Keracunan kerang-kerangan amnesik (ASP) adalah sejenis keracunan yang
disebabkan oleh pemakanan kerang-kerangan yang terce mar dengan asid domoik
(DA). Kajian mengenai kehadiran bermusim spesies Pseudo-nitzschia di muara
sungai telah dijalankan dengan persampelan plankton dan air selama 30 bulan
dengan kekerapan setiap dua minggu. Kepadatan Pseudo-nitzschia dipengaruhi oleh
kemasinan air, pH, kepekatan silika dan taburan hujan. Sel kerap dijumpai di antara
bulan April dan September tetapi kepadatan tertinggi adalah pada bulan Mei.
Kepadatan sel tertinggi Pseudo-nitzschia dicatat di Santubong dan Samariang adalah
pada B.Ox]03 sel rl dan 2.9 x lrt sel rl masing-masingnya. Berdasarkan ciri-ciri
ultrastruktur Pseudo-nitzschia di bawah pemerhatian mikroskop transmisi dan
pengimbas (FEM dan SEM), Pseudo-nitzschia tersebut dikenal pasti sebagai Pseudo
nitzschia pungens dan Pseudo-nitzschia brasiliana. Kedua-dua spesies tersebut juga
kerap dijumpai dalam komposisi fitoplankton yang disampel dari dua puluh satu
lokasi dari Laut China Selatan dan Selat Melaka. Tiada penghasilan domoik asid
dalam semua klon berdasarkan analisis jlorometrik kromatografi cecair berprestasi
tinggi (HPLC). Dalam kajian ini, jujukan gen LSU rDNA dari kedua-dua spesies telah
digunakan untuk menentukan hubungan filogeni dalam Pseudo-nitzschia spp.
Topologi pokok filogeni Pseudo-nitzschia menunjukkan perkaitan yang rapat di
antara P. brasiliana dan P. americana yang mana turut serasi dengan data morfologi.
P. pungens pula membentuk kumpulan beradik dengan P. multiseries. Berdasarkan
penyusunan jujukan pelbagai LSU rDNA, jujukan unik LSU rDNA untuk kedua-dua P.
brasiliana dan P. pltngens telah dikenal pasti dan prob rRNA yang spesies-spesijik
telah direkabentuk secara in silico. Secara keseluruhan, penemuan dalam kajian ini
adalah amat berguna dalam pemantauan fitoplankton ledakan algal merbahaya
(HABs) di Malaysia.
Kata kunci: Pseudo-nitzschia, asid domoik, kepadatan sel, mOlfologi, prob
oligonukleotida
IV

Pusat Khidmat Maklumat Akademik Vl'ilVERSm MALAYSIA S.4RAWAK
TABLE OF CONTENTS
DEC LARA TION
AKNOWLEDGEMENTS
ABSTRACT
ABSTRAK
TABLE OF CONTENTS
LIST OF TABLES
LIST OF FIGURES
CHAPTER I INTRODUCTION
1.1 General Introduction
1.2 Taxonomy ofPseudo-nitzschia
1.3 Ecology and bloom dynamics of Pseudo-nitzschia
1.4 Toxicology ofPseudo-nitzschia
1.5 Molecular studies of Pseudo-nitzschia
l.6 Research objectives
CHAPTER II SEASONAL OCCURRENCE OF Pseudo-nitzschia SPECIES IN TWO ESTUARIES OF KUCHING, SARAWAK
2.1 INTRODUCTION
2.2 MATERIALS AND METHODS
2.2.1 Study areas and sampling
2.2.2 Water filtration and macronutrients analysis
2.2.2.1 Nitrate-nitrogen (N03-- N)
2.2.2.2 Orthophosphate (P043
- - P)
2.2.2.3 Silicate (Si02)
2.2.3 Cell enumeration and cell counting
2.3.4 Rainfall distribution
2.3.4 Data analysis
v
Page
11
111
IV
V
IX
XI
2
5
6
10
11
13
15
17
17
17
18
18
18
19

.~ ,...
3.3.2 Morphology of Pseudo-nitzschia pungen 57
3.3.3 Morphology of Pseudo-nitzschia brasiliana 60
3.3.4 Toxicity analysis 63
3.4 DISCUSSION 64
CHAPTER IV IN SILleo DEVELOPMENT OF Pseudo-nitzschia SPECIES-SPECIFIC OLIGONUCLEOTIDE PROBES FOR RAPID DETECTION
4.1 INTRODUCTION 69
4.2 MA TERIALS AND METHODS
4.2.1 Pseudo-nitzchia culture 71
4.2.2 Genomic DNA extraction 71
4.2.2.1 Conventional DNA extraction 71
4.2.2.2 DNA extraction using Wizard® 72 Genomic DNA Purification Kit
4.2.3 Ribosomal RNA gene amplification and 73 sequencmg
4.2.4 Sequence analysis and taxon sampling 74
4.2.5 Phylogenetic analysis 78
4.2.6 Pairwise distance analysis 79
4.2.7 In silico design of species-specific 79 oligonucleotide probes
4.3 RESULTS
4.3. I Ribosomal RNA gene ~mplification and 80 sequencmg
4.3.2 Phylogenetic analysis 83
4.4.3 Distance analysis of P. brasiliana and P. 85 pungens
4.3.4 Potential signature sequence as species- 88 specific oligonucleotde probes
I4.4.5 Proposed oligonucleotide probes binding sites 90
4.4 DISCUSSION 93
VB

I
CHAPTER V CONCLUSION 96
REFERENCES 99
APPENDICES 114
Vlll

Table
Table 1.1
Table 3.1
Table 3.2
Table 3.3
Table 4.1
Table 4.2
Table 4.3
Table 4.4
Table 4.5
LIST OF TABLES
Page
Pseudo-nitzschia species grouped into the subgroup delicatissima and seriata according to the lengths transapical axis with toxic species in bold.
of of
5
Pseudo-nitzschia spp. used in the study with the numbers, date of isolation and sampling location.
strain 55
Morphometric data of Pseudo-nitzschia pungens isolated from different locations in Santubong, Sarawak (Sb) and Pulau Mamutik, Sabah (Pm).
58
Morphometric data of Pseudo-nitzschia brasiliana isolated from different locations in Samariang (Sm), Santubong, Sarawak (Sb) and Pulau Mamutik, Sabah (Pm).
61
Currently available Pseudo-nitzschia spp. probes with the label, sequence and reference.
70
LSU rONA (01-03) sequences of Pseudo-nitzschia spp. used in this study with the locality, GenBank accession number, and reference.
75
Pairwise distance analysis of P. brasiliana used in this study from different locations. Numbers above diagonal show mean character differences and numbers below diagonal show total character differences. PnSb, strains from Santubong; PnPm, strains from Pulau Mamutik.
86
Pairwise distance analysis of P. pungens used in this study from different locations. Numbers above diagonal show mean character differences and numbers below diagonal show total character differences. PnMt, strains from Muara Tebas.
87
Proposed oligonucleotide probes of P. brasiliana with nucleotide length, melting temperature (TM), and GC content (%). Red nucleotide(s) represents mismatch(s).
91
IX

Table Page
Table 4.6 P. pungens oligonucleotide probe with the nucleotide length, melting temperature (TM) and GC content (%).The red nucleotide(s) represents misrnatch(s) while the blue nucleotides indicates probe published by Miller and Scholin (1998).
92
.'
x

r
Figure
Figure 1.1
Figure 1.2
Figure 1.3
Figure 2.1
Figure 2.2
Figure 2.3
Figure 2.4
Figure 2.5
Figure 2.6
Figure 2.7
Figure 3.1
Figure 3.2
Figure 3.3
LIST OF FIGURES
Page
Diagram showing the main ultrastructural morphologies terms used in Pseudo-nitzschia species identifications.
4
Domoic acid and its derivatives (Adopted from F AO 2004). 8
General organization of the genes encoding the ribosomal RNA, ETS, external transcribed spacer; SSU, small subunit; ITS, internal transcribed spacer; LSU, large subunit (Adopted from Coleman 2003).
11
Map showing the two sampling sites, Santubong estuary and Samariang River in Kuching, Sarawak.
16
Pseudo-nitzschia spp. temporal distributions (mean ± SE) at Santubong estuary from 18 July 2007 to 29 March 2010.
21
Pseudo-nitzschia spp. temporal distributions (mean ± SE) at the Samariang estuary from 18 July 2007 to 29 March 2010.
23
In situ data of salinity (a, b), temperature (c, d) and pH (e, f) obtained in this study at Santubong and Samariang estuary from July 2007 until March 2010.
26
Macronutrients (Nitrate-nitrogen, Phosphate, Silicate) reading at Santubong and Samariang estuary from July 2007 until March 2010.
30
Monthly precipitation rate of Kuching Department of Meteorology, Malaysia).
city (Source: 32
Biplot of Principle Component Analysis showing cell density structure and relationships with environmental variables. Solid circles represent Pseudo-nitzschia cell densities of Santubong and Samariang (missing values for silica variables were calculated by regression with salinity).
34
Map showing the sampling locations in this study. 50
A flow diagram of acid wash method in preparation Pseudo-nitzschia cells for SEM and TEM observation.
of 52
Map showing the Malaysian water.
distribution of Pseudo-nitzschia spp. In 56
Xl

Figure Page
Figure 3.4 Micrographs of P. pungens from Sarawak and Sabah. SEM 59 (A-F). A, B, C: Valve view (Santubong), scale= 101!m. 0: Valve view (P. Mamutik), scale= 101!m. E: Girdle band, scale= 101!m. F: Pointed apical end (Pulau Mamutik), scale= ll!m. TEM (G-K). G: Pointed apical end (Santubong), scale= ll!m. H: Tapering valve towards end, scale= 21!m. I: Details of the cingulum, scale= 51!m. 1: Poroids, fibulae and striae, scale= 21!m. K: Absence ofcentral interspace, scale= 21!m.
Figure 3.5 Micrographs of P. brasiliana from Sabah and Sarawak. SEM 63 (A-C). A: Valve view (Santubong), scale= 101!m. B: Valve view (Samariang), scale = 51!m. C: Valve view (Pulau Mamutik), scale= 51!m. TEM (0-1). 0, E: Girdle band, scale= 21!m. F, G, 1: Rounded apical end, scale= ll!m (F & 1), 21!m (G). H: Regularly spaced fibulae and interstriae with absence of central interspace, scale= 21!m. I: Valve surface with poroids, scale= 21!m.
Figure 3.6 HPLC chromatogram of DA analysis using authentic DA as 63 standard (a). The result shows absence of DA in the samples analysed (b).
Figure 4.1 Gel images of PCR products loaded 51!L per lane on 1 % 81 agarose gel with 100 bp DNA ladder (Fermentas, USA).
Figure 4.2 Gel images of purified PCR product loaded 1 I!L per lane on 82 1% agarose gel with lkb DNA ladder.
Figure 4.3 Phylogenetic tree of Pseudo-nitzschia spp. inferred from 84 maximum parsimony (MP) with tree length of 308 evolutionary steps.
Figure 4.4 Sequence logo of signature sequence regions used for probe 89 design for P. brasiliana and P. pungens in this study.
XII

CHAPTER I
INTRODUCTION
1.1 General Introduction
Phytoplankton plays important roles as the base of aquatic food web and generating
atmospheric oxygen through photosynthesis. About 2% of total phytoplankton are
potent phycotoxin producers, thus rapid growth of these microalgae at high
concentration create nuisance and impact to the surrounding environments. Harmful
Algal Bloom (HAB) is a natural phenomenon when drastic increase of phytoplankton
at unusually high density causes adverse effect to other life including humans through
the production of toxins (Andersen 1996), physical damage or degradation of water
quality (Usup et al. 1989). Blooms were usually monospecific and dominated by a
single species.
Malaysia is one of many countries affected by the HABs (Usup et al. 2002).
Blooms of the paralytic shellfish poisoning (PSP) causative dinoflagellates in Sabah
have been reported almost annually (Usup and Azanza 1998). The perception that PSP
was only confined to Sabah changed following the cases of human intoxications in
1991 when human consumers of farmed mussels were hospitalized due to the paralytic
shellfish toxin, PST (Usup et al. 2002).
HABs can create severe threats to the development of fisheries resources
especially in coastal aquaculture industries (Usup et al. 2002). The HAB monitoring

programs facilities done by the Malaysian Department of Fisheries has been mainly
focusing on the PSP with no attention given to other potential HABs. No confirmed
ciguatera shellfish poisoning (CFP), diarrheatic shellfish poisoning (DSP) and
amnesic shellfish poisoning (ASP) events have been reported in the country to date,
however organisms that known to responsible for these poisoning are commonly
found in the phytoplankton assemblages ofour waters.
1.2 Taxonomy of Pseudo-nitzschia
The genus Pseudo-nitzschia is a group of pennate marine diatom that causes ASP by
producing the neurotoxin domoic acid (DA). This pennate diatom was previously
established in a section of the genus Nitzschia and remained until Hasle re-c1assified
the species as a separate genus (Has Ie 1994). The feature which distinguished the
genus was the unique stepped chain-forming of the Pseudo-nitzschia. However, some
of the Pseudo-nitzschia species may be observed as solitary cells (Orlova and
Shevchenko 2002). Furthermore, Pseudo-nitzschia cells may be registered in paired
cells when cell divisions were taking part (Orlova and Shevchenko 2002). Pseudo
nitzschia is widely accepted as planktonic pennate diatom, though some authors have
reported the epiphytic behavior of the species, e.g. P. americana as epiphyte in other
diatom (Hasle and Syvertsen 1997; Orlova and Shevchenko 2002).
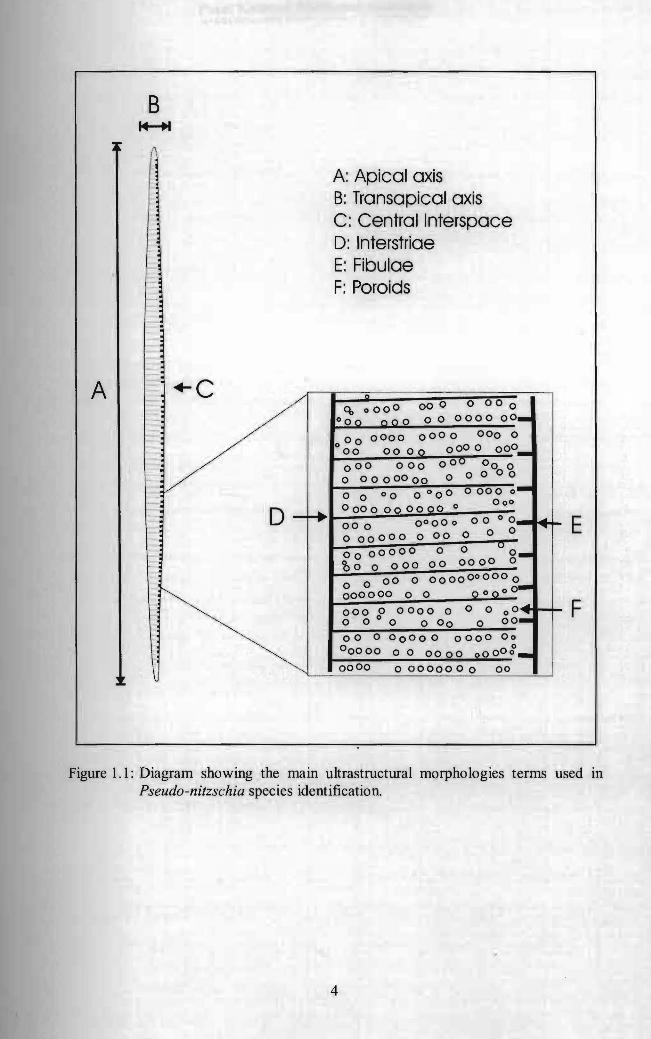
Under light microscope, cells of Pseudo -nitzschia are lanceolate in shape and
characterized by the siliceous cell walls with two chloroplasts located at both polar
ends. Pseudo-nitzschia species has generic features of strongly elongate cells with
rounded or pointed ends (Hasle and Syvertsen 1997) (Figure 1.1). The valve of
Pseudo-nitzschia is weakly silicified and the raphe slits (fibulae) are not elevated
2

above the general valve level (HasIe and Syvertsen 1997) (Figure 1.1). The
ultrastructural features considered in the diatom taxonomy included the number of
interstriae and fibulae in 1 0 ~m, poroid density in each micron, row of poroid and
presence of central interspace (Has Ie and Syvertsen 1997) (Figure 1.1). Hasle (1994)
defined two subgroups of Pseudo-nitzschia according to their valve widths, namely
seriata subgroup (valve width> 3 )lm) and delicatissima subgroup (valve width < 3
)lm) (Table 1.1).
3
B ~
A: Apical axis B: Transapical axis
11 C: Central Interspace D: Interstriae E: Fibulae F: Poroids
+C ./A
000 000q, 000 0 0
/ 0000 00_00°00 000
000 000000 00 0000 000 0 000_I 00 0000
o Ov 00 0000 000 o 0 0000 0000000
0 0 00 0 0 00 o 000 ~-000
0000 00 0000 0 ...00 00D - f+ 0°000 ...000 E00o 00 0o 00000 0 o °00000000 d-COo 0 000 00 0000
00 0 000000 0000o 0 ,_ 000000 0 0 a 0 0 0 0
0 0 0 o~000 0 0000 0 F o 0 0 0 00o 00 0
00 0 00 0 00 0000 00 000000 o 0 00 000 _0000
.~ 0000 o 0000000 00
Figure 1.1: Diagram showing the main ultrastructural morphologies terms used in Pseudo-nitzschia species identification.
4

Pusat Khidmat Maklumat Akademik UNlVERSm MALAYSIA SARAWAK
Table 1.1: Pseudo-nitzschia species in the subgroup delicatissima and seriata according to the lengths oftransapical axis with toxic species in bold.
Delicatissima subgroup Seriata subgroup
P. americana
P. antartica
P. brasiliana
P. caciantha
P. caOiantha
P. cuspidate
P. delicatissima
P. galaxie
P. granii
P. injlatula
P. lineola
P. micropora
P. multistriata
P. prolongotoides
P. pseudodelicatissima
P. sinica
P. subcurvata
P. turgidula
P. turgiduloides
P. australis
P. fraudulenta
P. heimii
P. multiseries
P. pungens
P. pungiformis
P. seriata
P. subfraudulenta
P. subpacifica
1.3 Ecology and bloom dynamics of Pseudo-nitzschia
Pseudo-nitzschia spp. has been reported to present in the water column whole year
round in some areas (Fitrianti 2004; Yap-Dejeto 2008; Klein et al. 2010). In some
bloom event, Pseudo-nitzschia cells account for high percentage of total
phytoplankton biomass (e.g. Dortch et al. 1997; Fryxell et al. 1997; Trainer et al.
1998). Blooms of Pseudo-nitzschia in some temperate regions showed seasonal shifts
with the fluctuations in water temperature while in other regions occur occasionally in
the years. In west coast of United States, P. australis is common in the water column
5

while P. pungens and P. delicatissima appeared in high abundant in summer and
autumn (Fryxell et al. 1997). Follow through the winter, the seasonal succession is
replaced by P. subpacifica, P. pseudodelicatissima, P. heimii and P. multiseries
(Fryxell et al. 1997). Abundance of Pseudo-nitzschia was also observed occasionally
in the Antarctic region and contributed significantly to the total diatom biomass
(Almandoz et al. 2008). In the South China Sea, the P. pungens was dominant in the
whole year (Qi et al. 1996).
The bloom dynamics of Pseudo-nitzschia was associated with environmental
parameters (Fehling et al. 2005). Nutrients availability was shown to be the main
contributing factor to the occurrence of Pseudo-nitzschia. Pseudo-nitzschia blooms
were observed in nutrient rich water contributed by the upwelling events and
increased river discharge following the sustained precipitation (Trainer et al. 1998).
On the other hand, high concentration of P. australis was found in US west coast after
upwelling ended when inorganic nutrients were lower (Buck et al. 1992). Optimal
growth of Pseudo-nitzschia was attained in nutrient rich waters and other favorable
surrounding conditions. However, selective nature of the condition preferences in
different species or ecotypes of Pseudo-nitzschia species has cause complexity in
understanding the blooms dynamic ofPseudo-nitzschia species.
1.4 Toxicology of Pseudo-nitzschia
Amnesic shellfish poisoning (ASP) is the only shellfish poisoning caused by
. diatoms (Van Dolah 2000). The toxin that responsible for the poisoning is domoic
acid (DA), which is produced by several species of the pennate diatom Pseudo
nitzschia. In recent years, DA was also reported in other diatom species such as
6

Nitzschia navis-varingica (Kotaki et al. 2000; Lundholm and Moestrup 2000; Kotaki
et al. 2005).
DA and its derivatives (Figure 1.2) are water soluble and heat stable tri
carboxylic amino acid (Rue and Bruland 2001; Van Dolah 2000). In its pure form, this
excitatory amino acid appears as colorless crystal needles. Filter feeders shellfish
mollusks are the main vector of DA, even though other vectors such as anchovies
(Scholin et al. 2000), planktivorous fish (Lefebvre et al. 2002), metazoans (Buck et al.
1992) and cuttlefish (Costa et al. 2005) were also documented. DA accumulated
mainly in the digestive system of vector organisms and present in the flesh in small
concentration. The chemical structure resembles kainic acid in which it binds to the
kainite receptor in the brain hippocampus and misinterpreted by the brain as the
natural neurotransmitter, the L-glutamate (Biscoe et al. 1976; Bates 2004). The
mechanism causes continuous stimulation and over-excitement of signals and finally
leads to irreversible brain damage. The gastro-intestinal symptoms of intoxication
include nausea and disorientation within 3-5 hours (Hallegraeff 2003). More severe
poisoning involved neurological symptoms of breathing difficulties, seizure,
irreversible lost of short-term memory and even death (Bates 1998; Hallegraeff 2003).
Ideally, the intoxication effects depend on the health condition, while patients with
age and renal dysfunction are at the higher risk. The diagnosis was mainly based on
the bivalve ingestion history. Treatment for the poisoning was symptomatic as there is
so far no anti-dote for ASP poisoning (Hallegraeff 2003). In the case of first aid
treatment, withdrawal of stomach content was usually done and activated charcoals
were left inside the stomach.
7
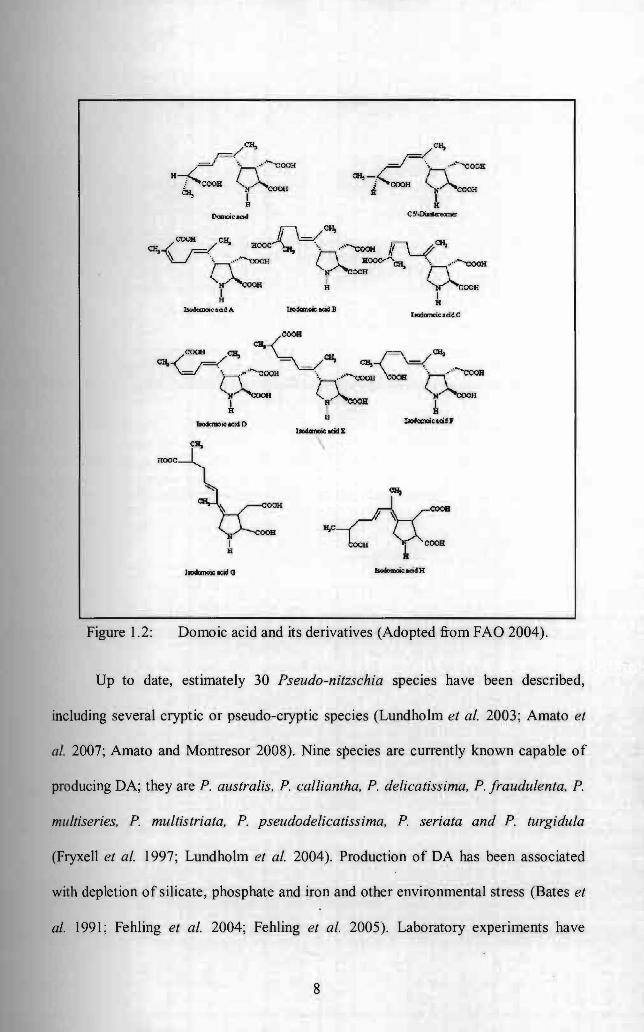
Figure 1.2: Domoic acid and its derivatives (Adopted from FAO 2004).
Up to date, estimately 30 Pseudo-nitzschia speCIes have been described,
including several cryptic or pseudo-cryptic species (Lundholm et al. 2003; Amato et
ai. 2007; Amato and Montresor 2008). Nine species are currently known capable of
producing DA; they are P. australis, P. calliantha, P. delicatissima, P. fraudulenta, P.
multiseries, P. multistriata, P. pseudodelicatissima, P. seriata and P. turgidula
(Fryxell et al. 1997; Lundholm et al. 2004). Production of DA has been associated
with depletion of silicate, phosphate and iron and other environmental stress (Bates et
al. 1991; Fehling et al. 2004; Fehling et al. 2005). Laboratory experiments have
8

shown that prolonged light hours and urea-grown Pseudo-nitzschia tend to produce
DA at higher concentration (Fehling et al. 2005; Howard et al. 2007).
The first incidence which involved human intoxication was reported in Prince
Edward Island, Canada in 1987 (Bates et al. 1989). Three fatalities were reported and
over hundreds of people were taken ill after consuming cultivated blue mussels
contaminated with high level of domoic acid in the shellfish meat (Bates et al. 1989).
Even though no human intoxication of DA was reported after the first incident (Van
Dolah 2000), Pseudo-nitzschia has been associated with DA contamination in filter
reeders and other vectors. Mass mortality of pelicans and connorants that feeds on
anchovies were reported in Monterey Bay, California in 1991 (Fritz et al. 1992). Again
three years after the death of seabirds in Monterey Bay, deaths of sea lions occurred in
the same location (Scholin et al. 2000). Following the numerous incidence of ASP,
precautious measure had been taken in shellfish areas in Bay of Fundy by extracting
the shellfish tissue and revealed unacceptable content of DA (Martin et al. 1990).
Elevated DA exceeding the regulatory limit was detected in king scallop in Scottish
shellfish fanns resulted in termination of harvesting in 1999 (Campbell et al. 2001). P.
delicatissima, P. australis, P. multiseries and P. seriata were among the well-known
Pseudo-nitzschia species which responsible for the DA contamination in many events
(e.g. Bates et al. 1989; Martin et al. 1990; Sch~lin et al. 2000). However, toxicity
characteristics of diatom Pseudo-nitzschia are dissimilar among species, in term of
their toxin concentration and toxin producing ability.
9

1.5 Molecular studies of Pseudo-nitzschia
In Pseudo-nitzschia taxonomy, specIes can only distinguishable through
details observation and measurement of ultrastructural features of frustules under
electron microscopy (Hasle and Syvertsen 1997; Fryxell and Hasle 2003).
Furthermore, samples treatment for electron microscopy identification is a time
consuming process and required expertise in diatom taxonomy. Due to the constraints
of the morphological identification, molecular approaches have been developed for
detection of several HAB species (e.g. Adachi et al. 1996; Anderson et al. 1999;
Miller and Scholin 1998) from natural phytoplankton samples in a rapid and precise
manner. Some detection methods of HAB species included molecular probes, consist
of lectin or antibody biding assays, as well the DNA probes (Scholin et al. 2003).
Application of DNA probes have been thus far relied on the basis of whole-cell
hybridization and cells homogenate (Scholin et al. 2003). Fluorescently labeled probes
bind specifically to the target species can be easily revealed through epifluorescence
microscopy or detected by flow cytometry (Scholin et al. 2003).
Ribosomal genes including the internal transcribed spacer (ITS 1 and ITS2)
region, small subunit ribosomal genes (SSU rDNA) and large subunit ribosomal gene
(LSU rDNA) have been widely used in molecular characterizations of HABs species
(Figure 1.3), with numerous amounts of data available in the database. In molecular
analyses of diatom, the LSU rDNA region is frequently applied in inference of the
relationship among the HABs species, especially in Pseudo-nitzschia (Lundholm et al.
2002).
I
10

Figure 1.3: General organization of the genes encoding the ribosomal RNA, ETS,
external transcribed spacer; SSU, small subunit; ITS, internal transcribed
spacer; LSU, large subunit (Adopted from Coleman 2003).
Several species specific probes have been developed for Pseudo-nitzschia spp.
(Miller and Scholin 1996, 1998). However, the probes designed are not applicable for
Pseudo-nitzschia species from different geographical regions. For example, the lectin
binding assays seemed promising in discriminating different Pseudo-nitzschia spp.
from the same location, but could not differentiate between strains with different
geographical origin (Rhodes 1998; Skov et al. 2004). Hence, it is important to develop
molecular probe exclusive for Pseudo-nitzschia species and strains for a specific
region.
1.6 Research objectives
Studies on Pseudo-nitzschia in the country are lacking and little attention paid to it.
Therefore, combination of field and laboratory studies were carried out to gain better
understanding of seasonal occurrence of Pseudo-nitzschia and the environmental
conditions that most likely affect cell abundance in natural environment. Nonetheless,
11





























