Embed Size (px)
Citation preview

receptors in high modification states to keep the activity of thereceptor complex at a moderate level (,0.3). In the MWC model,large Hill coefficients are observed only when values of both N and Lare large, whereas high sensitivity is observed when N is large and Lis small (Box 1). Thus, high sensitivity of wild-type cells can beexplained by combination of receptor interactions and control overreceptor activity. CheB is particularly crucial22 because it generateslower modification states with lower activity and higher sensitivity.Clearly, more remains to be learned about the details of signalprocessing and amplification in chemotaxis, and more complicatedmodels are required to quantitatively account for integration ofsignal between receptors of different types; however, the functionalinteractions between receptors observed here provide a frameworkfor further experimental analysis and modelling. A
MethodsPlasmidsThe genes cheY–eyfp and cheZ–ecfp, encoding CheY–YFP and CheZ–CFP fusions22, wereamplified with polymerase chain reaction (PCR) and cloned in tandem, using PCR-introduced SacI, XbaI and HindIII restriction sites, into pTrc99A (AmpR, Pharmacia)under an isopropyl b-D-thiogalactoside (IPTG)-inducible promoter, yielding pVS88. Allother chemotaxis proteins were expressed under the tight control of a salicylate-induciblepromoter from the pLC112-derived plasmids20. The Tar expression plasmid pLC113,the Tsr expression plasmid pPA114, the CheA expression plasmid pPA113 and thepLC112-derived expression vector pKG110 were obtained from J. S. Parkinson. Plasmidsexpressing CheA and CheW or only CheW (pVS125 and pVS126, respectively) wereconstructed by cloning PCR-amplified cheA and cheW or cheW genes into NdeI and KpnIrestriction sites of pKG110. Plasmid pVS121 that expresses Tar(Q295E/Q309E) (referredto as Tar(EEEE)), pVS120 that expresses Tar(Q309E) (Tar(QEEE)) and pVS122 thatexpresses Tar(E491Q) (Tar(QEQQ)) were constructed by replacing the KpnI/BamHIfragment of pLC113 with a fragment containing corresponding mutations of tar that wereamplified via PCR from strains VS134, VS131 and VS147 (ref. 22), respectively.
Strains and their growthDerivatives of E. coli K12 strain RP437 used in this study are listed in SupplementaryInformation. Ordinarily, proteins were expressed from plasmids in strains deleted for thechromosomal copy of the corresponding gene. Cells were grown as described22 in thepresence of 100 mg ml21 ampicillin, 34 mg ml21 chloramphenicol, 50 mM IPTG andvarying amounts (0–1 mM) of sodium salicylate. Immunoblots, performed as before6, wereused to compare the expression levels of proteins induced by salicylate with their nativeexpression levels. Polyclonal antibody raised against the signalling domain of Tsr13 thatreacts equally well with both Tar and Tsr was obtained from J. S. Parkinson. Polyclonalantibodies raised against CheA and CheW were obtained from R. C. Stewart.
FRET measurementsCells expressing CheY–YFP and CheZ–CFP were attached to a polylysine-coated coverslipand placed in a flow cell. The flow cell was kept under a constant flow (0.5 ml min21) oftethering buffer (10 mM potassium phosphate, 0.1 mM EDTA, 1 mM L-methionine,10 mM sodium lactate, pH 7). Solutions of the same buffer were used to add and removespecified amounts of attractants (a-methyl-D,L-aspartate or L-serine) in a sequence ofsteps of increasing size. FRET, defined as the fractional change in cyan fluorescence due toenergy transfer, was calculated from changes in the ratios of yellow and cyan fluorescencesignals, measured as described before22,28. For CheRþ CheBþ cells, the initial response wasmeasured (before the cells had time to adapt). A field of 300–500 cells was monitoredin each experiment. Under the conditions of our experiments, interaction betweenCheY–YFP and CheZ–CFP provides a measure of the activity of CheA22, so FRET datawere plotted as kinase activity. Zero CheA activity was determined by adding saturatingamounts of serine and MeAsp.
Data analysis and simulationsData analysis and simulations are described in Supplementary Information.
Received 19 November 2003; accepted 5 February 2004; doi:10.1038/nature02406.
1. Gegner, J. A., Graham, D. R., Roth, A. F. & Dahlquist, F. W. Assembly of an MCP receptor, CheW, and
kinase CheA complex in the bacterial chemotaxis signal transduction pathway. Cell 70, 975–982
(1992).
2. Boukhvalova, M. S., Dahlquist, F. W. & Stewart, R. C. CheW binding interactions with CheA and
Tar. Importance for chemotaxis signaling in Escherichia coli. J. Biol. Chem. 277, 22251–22259
(2002).
3. Borkovich, K. A., Kaplan, N., Hess, J. F. & Simon, M. I. Transmembrane signal transduction in
bacterial chemotaxis involves ligand-dependent activation of phosphate group transfer. Proc. Natl
Acad. Sci. USA 86, 1208–1212 (1989).
4. Ninfa, E. G., Stock, A., Mowbray, S. & Stock, J. B. Reconstitution of the bacterial chemotaxis signal
transduction system from purified components. J. Biol. Chem. 266, 9764–9770 (1991).
5. Maddock, J. R. & Shapiro, L. Polar location of the chemoreceptor complex in the Escherichia coli cell.
Science 259, 1717–1723 (1993).
6. Sourjik, V. & Berg, H. C. Localization of components of the chemotaxis machinery of Esherichia coli
using fluorescent protein fusions. Mol. Microbiol. 37, 740–751 (2000).
7. Bray, D., Levin, M. D. & Morton-Firth, C. J. Receptor clustering as a cellular mechanism to control
sensitivity. Nature 393, 85–88 (1998).
8. Shi, Y. & Duke, T. Cooperative model of bacterial sensing. Phys. Rev. E 58, 6399–6406 (1998).
9. Duke, T. A. J. & Bray, D. Heightened sensitivity of a lattice of membrane receptors. Proc. Natl Acad. Sci.
USA 96, 10104–10108 (1999).
10. Shimizu, T. S., Aksenov, S. V. & Bray, D. A spatially extended stochastic model of the bacterial
chemotaxis signalling pathway. J. Mol. Biol. 329, 291–309 (2003).
11. Mello, B. A. & Tu, Y. Quantitative modeling of sensitivity in bacterial chemotaxis: the role of coupling
among different chemoreceptor species. Proc. Natl Acad. Sci. USA 100, 8223–8228 (2003).
12. Asakura, S. & Honda, H. Two-state model for bacterial chemoreceptor proteins. The role of multiple
methylation. J. Mol. Biol. 176, 349–367 (1984).
13. Ames, P. & Parkinson, J. S. Constitutively signaling fragments of Tsr, the Escherichia coli serine
chemoreceptor. J. Bacteriol. 176, 6340–6348 (1994).
14. Barkai, N. & Leibler, S. Robustness in simple biochemical networks. Nature 387, 913–917 (1997).
15. Morton-Firth, C. J., Shimizu, T. S. & Bray, D. A. Free-energy-based stochastic simulation of the Tar
receptor complex. J. Mol. Biol. 286, 1059–1074 (1999).
16. Borkovich, K. A., Alex, L. A. & Simon, M. I. Attenuation of sensory receptor signaling by covalent
modification. Proc. Natl Acad. Sci. USA 89, 6756–6760 (1992).
17. Bornhorst, J. A. & Falke, J. J. Attractant regulation of the aspartate receptor-kinase complex: Limited
cooperative interactions between receptors and effects of the receptor modification state. Biochemistry
39, 9486–9493 (2000).
18. Li, G. & Weis, R. M. Covalent modification regulates ligand binding to receptor complexes in the
chemosensory system of Escherichia coli. Cell 100, 357–365 (2000).
19. Gestwicki, J. E. & Kiessling, L. L. Inter-receptor communication through arrays of bacterial
chemoreceptors. Nature 415, 81–84 (2002).
20. Ames, P., Studdert, C. A., Reiser, R. H. & Parkinson, J. S. Collaborative signaling by mixed
chemoreceptor teams in Escherichia coli. Proc. Natl Acad. Sci. USA 99, 7060–7065 (2002).
21. Levit, M. N. & Stock, J. B. Receptor methylation controls the magnitude of stimulus–response
coupling in bacterial chemotaxis. J. Biol. Chem. 277, 36760–36765 (2002).
22. Sourjik, V. & Berg, H. C. Receptor sensitivity in bacterial chemotaxis. Proc. Natl Acad. Sci. USA 99,
123–127 (2002).
23. Shimizu, T. S. et al. Molecular model of a lattice of signalling proteins involved in bacterial
chemotaxis. Nature Cell Biol. 2, 792–796 (2000).
24. Levit, M. N., Grebe, T. W. & Stock, J. B. Organization of the receptor–kinase signaling array that
regulates Escherichia coli chemotaxis. J. Biol. Chem. 277, 36748–36754 (2002).
25. Kim, S. H., Wang, W. & Kim, K. K. Dynamic and clustering model of bacterial chemotaxis receptors:
structural basis for signaling and high sensitivity. Proc. Natl Acad. Sci. USA 99, 11611–11615 (2002).
26. Kim, K. K., Yokota, H. & Kim, S. H. Four-helical-bundle structure of the cytoplasmic domain of a
serine chemotaxis receptor. Nature 400, 787–792 (1999).
27. Monod, J., Wyman, J. & Changeux, J.-P. On the nature of allosteric transitions: a plausible model.
J. Mol. Biol. 12, 88–118 (1965).
28. Sourjik, V. & Berg, H. C. Binding of the Escherichia coli response regulator CheY to its target measured
in vivo by fluorescence resonance energy transfer. Proc. Natl Acad. Sci. USA 99, 12669–12674 (2002).
Supplementary Information accompanies the paper on www.nature.com/nature.
Acknowledgements We thank J. S. Parkinson for providing some of the plasmids, strains and
antibody used in this study, and R. C. Stewart for providing antibody. We also thank D. Bray,
K. A. Fahrner, J. J. Falke, J. S. Parkinson, T. Shimizu and A. Vaknin for comments on the
manuscript. We thank P. Zucchi for technical help. This research was supported by the NIH.
Competing interests statement The authors declare that they have no competing financial
interests.
Correspondence and requests for materials should be addressed to H.C.B.
..............................................................
Enzymic activation and transferof fatty acids as acyl-adenylatesin mycobacteriaOmita A. Trivedi, Pooja Arora, Vijayalakshmi Sridharan, Rashmi Tickoo,Debasisa Mohanty & Rajesh S. Gokhale
National Institute of Immunology, Aruna Asaf Ali Marg, New Delhi 110 067,India.............................................................................................................................................................................
The metabolic repertoire in nature is augmented by generatinghybrid metabolites from a limited set of gene products1–3. Inmycobacteria, several unique complex lipids are produced by thecombined action of fatty acid synthases and polyketide synthases(PKSs)4–6, although it is not clear how the covalently sequestered
letters to nature
NATURE | VOL 428 | 25 MARCH 2004 | www.nature.com/nature 441© 2004 Nature Publishing Group

biosynthetic intermediates are transferred from one enzymaticcomplex to another. Here we show that some of the 36 annotatedfadD genes, located adjacent to the PKS genes in the Mycobacter-ium tuberculosis genome, constitute a new class of long-chainfatty acyl-AMP ligases (FAALs). These proteins activate long-chain fatty acids as acyl-adenylates, which are then transferred tothe multifunctional PKSs for further chain extension. This modeof activation and transfer of fatty acids is contrary to thepreviously described universal mechanism involving the for-mation of acyl-coenzyme A thioesters. Similar mechanismsmay operate in the biosynthesis of other lipid-containing metab-olites and could have implications in engineering novel hybridproducts.
Analysis of the mycobacterial genome has revealed an array ofgenes involved in the lipid metabolism, including a large family offadD genes7. These mycobacterial FadD proteins show homologyto acyl-CoA synthetases that convert free fatty acids into acyl-coenzyme A (CoA) thioesters, the first step in fatty acid degra-dation. The functional importance of fadD genes in the biology ofmycobacteria has recently been demonstrated by gene inactivationexperiments8–11. However, the precise biochemical functions ofFadD proteins have not been investigated thus far. The compu-tational analysis of mycobacterial FadD proteins revealed poorsequence homology with other acyl-activating enzymes such asfirefly luciferases12, adenylation domains of non-ribosomal peptidesynthetases (NRPSs)13 and acetyl CoA synthetases14. The dendro-gram obtained from multiple sequence alignments showed twodistinct protein clusters (see Supplementary Fig. 1). The smallercluster consisted of 12 FadD proteins with distinctly high sequencesimilarity. Interestingly, nine of these 12 fadD genes were locatedadjacent to PKS and NRPS gene clusters. In contrast, only two fadDgenes (fadD22 and fadD10) from the other cluster were in proximitywith PKS and NRPS genes. On careful analysis of the FadD proteins
from the smaller cluster, it was observed that a number of conservedpositions identified from multiple alignments corresponded withthe active site residues of the adenylation domain of NRPS proteins.We therefore set out to investigate whether FadD proteins from thesmaller cluster would activate fatty acids as acyl-adenylates and thentransfer these acyl chains on to PKSs, in a mechanism analogous tothe adenylation domains.
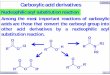
The small cluster includes the fadD26 and fadD28 gene products,which have been demonstrated to be essential for the biosynthesis ofa virulent phthiocerol dimycocerosate lipid. It is interesting thatdespite the presence of other functional fadD genes and plentifulpools of acyl-CoA in the bacteria, the fadD26 and fadD28 mutantsof mycobacteria lacked phthiocerol dimycocerosate on the cellenvelope8–10,15. The basis for this crucial non-redundant metabolicrole was examined by analysing the enzymatic function of FadD26and FadD28 proteins. Our computational analysis predicted twoputative translational start sites for the FadD26 protein. We there-fore cloned and expressed this protein from two independent startsites in Escherichia coli. The functional FadD26 protein wasobtained from the downstream start codon. The enzymatic activityof the FadD26 and FadD28 proteins was monitored by using radio-labelled C3 to C18 fatty acids. Analysis of the reaction products usingradio thin layer chromatography (TLC) showed identical retentionfactor (Rf) both in the presence and absence of reduced coenzyme A(CoASH) (Fig. 1a). The maximal activity for these proteins wasobtained for long-chain fatty acids. These metabolites were thenanalysed on high-performance liquid chromatography (HPLC)(Fig. 1b, peak 1) and were characterized as fatty acyl-adenylate(AMP) by using electrospray ionization (ESI) mass spectrometry(MS) (Fig. 1c). The identity of the product was also confirmed bychemical synthesis of an authentic reference. This data establishesthat both FadD26 and FadD28 proteins do not catalyse acyl-CoAformation as suggested in earlier studies8–10,15, but instead synthesize
CoASH – + – + – + – + – + – + – + – +
Fatty acid
Acyl-CoA
Origin
Acyl-AMP
FadD15 FadD17 FadD19 FadD26 FadD28 FadD29 FadD30 FadD32
2 4 6 8 10 12 14 16 18 20
mA
U
0
50
100
150
200
1
2
Retention time (min)
420 440 460 480 500 520 540 560 580 6000
20
40
60
80
100
Rel
ativ
e in
tens
ity (%
)
NH2
NN
N N
O
O
O OO
O
OP
PH H
H
HO
10
m/z (a.m.u) m/z (a.m.u)
1 Lauroyl-AMP
[M-H]–1
528.24
c
a b
d
500 550 600 650 700 750 800 850 900 9500
20
40
60
80
100N
N N
HH
H
N
NH2
O OOOH
OH
O
PPOH
O
OOH
HN
O
HN
O
S
O
PHO OO
OH
10
Rel
ativ
e in
tens
ity (%
)
484.61
473.60
[M-2H]–2/2
[M-H]–1
948.22
2 Lauroyl-CoA
OH
OH
OH
Figure 1 Chemical characterization of the FadD products. a, Radio-TLCs showing
activity of various FadDs in the presence and absence of CoASH. The fatty acid
migrates on the TLC with an Rf of 0.95. The acyl-AMP and acyl-CoA has an Rf value of 0.5
and 0.4 respectively. b, Reverse-phase HPLC chromatograms of reaction products
synthesized by FadD28 (solid line) and FadD19 (dotted line) recorded at 259 nm. FadD28
catalysed the biosynthesis of lauroyl-AMP (structure 1) while FadD19 formed
lauroyl-CoA (structure 2). c, ESI-MS spectra for FadD28 product (Lauroyl-AMP). The
mass/charge (m/z ) peak at 528.24 corresponds to the molecular ion peak [M-H]21.
d, ESI-MS spectra for the FadD19 product (Lauroyl-CoA). The peak at 948.22
corresponds to the molecular ion peak, whereas the peak at 473.60 corresponds to
the [M-2H]22/2 ion.
letters to nature
NATURE | VOL 428 | 25 MARCH 2004 | www.nature.com/nature442 © 2004 Nature Publishing Group

acyl-adenylates of long-chain fatty acids. In an earlier report, thepurified FadD28 protein from Mycobacterium bovis BCG (whichdiffers in 2/580 amino acids from M. tuberculosis FadD28 proteinused in this study) was shown to catalyse formation of acyl-CoAsfrom their corresponding acids16. This may be because themetabolites were not chemically characterized and the enzymaticactivity was inferred from radio-scintillation counting assays thatcannot distinguish between the acyl-AMP and acyl-CoA. Wetherefore suggest that these two proteins should be annotated asFAALs.
Because the FadD26 and FadD28 proteins belong to the group ofhighly conserved 12 FadDs, we hypothesized that this small clustermight constitute a FAAL family of proteins. To test this hypothesis,three more fadD genes (fadD29, fadD30 and fadD32) from thiscluster were cloned and expressed in E. coli. As controls, three fadDgenes from the larger cluster (fadD15, fadD17 and fadD19) were alsoexpressed in the heterologous host. All six of these purified FadDproteins catalysed the formation of acyl-adenylate from long-chainfatty acids in the absence of CoASH. The addition of CoASH to the
reaction mixture did not alter the product formation with FadD29,FadD30 and FadD32 (Fig. 1a). In contrast, a new product withlower Rf was observed for FadD15, FadD17 and FadD19 proteins(Fig. 1a), which also eluted at a different time on a reverse-phaseHPLC (Fig. 1b, peak 2). This new product was established to be fattyacyl-CoA by using ESI-MS (Fig. 1d).
These studies corroborate our earlier computational analysis ofthe presence of two distinct groups of FadD proteins, one of whichactivates metabolic carboxylates as acyl-adenylates, while the othersactivate them as acyl-CoAs. The CoA analogues are used by the hostin lipid degradation as well as for providing precursors in thebiosynthesis of metabolites. We have recently characterized a newfamily of type III PKSs from M. tuberculosis, which uses such long-chain acyl-CoA substrates17.
To investigate the ability of acyl-AMP ligases to transfer theactivated fatty acids specifically to the cognate PKSs, we cloned andoverexpressed three PKS genes, ppsA, pks13 and pks6 in E. coli. Thedomain organization, as predicted by PKSDB18,19, indicated thepresence of two acyl carrier protein (ACP)-like domains in these
Figure 2 Activation and transfer of fatty acids on to the cognate PKS proteins.
a, Autoradiogram showing radioactivity associated with PKS proteins. The Coomassie-
stained PKS and FadD proteins are present in lanes 1, 4 and 7. Lanes 2, 5 and 8 show
radio-labelled bands associated with PpsA, PKS13 and PKS6 proteins in the presence of
their cognate FadD proteins and ATP. No radioactivity is observed in the absence of ATP
(lanes 3, 6 and 9). b, The mechanism of fatty acid activation and transfer, mediated by
FAAL proteins. The long-chain fatty acids are activated as acyl-adenylates and are
transferred on to the P-pant arm of the N-terminal PCP domain of PKS proteins. This acyl
group is subsequently loaded at the active-site cysteine of the ketosynthase (KS) domain.
The acyl transferase (AT) domain transfers the extender unit malonyl-CoA on to the ACP
P-pant arm, which is used for a two-carbon chain extension through a decarboxylative
condensation reaction. The ACP-bound intermediates are reduced by ketoreductase (KR)
domain.
Figure 3 Structural analysis of FAAL and FACL proteins. a, Limited proteolysis of FAAL
proteins with trypsin. b, Limited trypsin cleavage of FACL proteins. Each panel in a and b
shows two lanes for each FadD protein, when incubated in the absence and the presence
of trypsin. c, Three-dimensional model for FAAL proteins. The larger N-terminal domain is
shown in green and the smaller C-terminal domain is shown in blue. The flexible hinge
region is drawn in yellow. The site of trypsin cleavage is indicated by an arrow. d, Multiple
sequence alignment of the hinge region of representative members of FAAL and FACL
proteins. The trypsin cleavage site on FadD28 is indicated by an arrow. Invariant residues
are shown in red. The conserved amino-acid residues of FACL proteins are shown in blue.
letters to nature
NATURE | VOL 428 | 25 MARCH 2004 | www.nature.com/nature 443© 2004 Nature Publishing Group

PKS proteins. The amino-terminal ACP-like domain also showedhomology to the peptidyl carrier protein (PCP) domain of NRPSproteins. The ACP and PCP are small homologous domains presentin the multifunctional PKSs and NRPSs respectively, which arepost-translationally modified at the active site serine residue. Thisphosphopantetheinylation (P-pant) modification of both PCP andACP domains was achieved by co-expressing surfactin P-panttransferase (sfp) in E. coli host20,21. The loading of activated acylchains on to the N-terminal PCP domains of the cognate PKSproteins was monitored by incubating the proximal FadD–PKSprotein pairs (FadD26–PpsA, FadD32–PKS13 and FadD30–PKS6)along with 14C-long-chain fatty acids and ATP. The proteins wereresolved on SDS–polyacrylamide gel electrophoresis (PAGE) andanalysed using autoradiography. Radioactive bands were associ-ated with the PKS proteins in the presence (Fig. 2a; lanes 2, 5 and8), but not in the absence (Fig. 2a; lanes 3, 6 and 9) of ATP. Thetransfer of fatty acids on to the PKS proteins and its subsequentextension is illustrated in Fig. 2b. This biosynthetic mechanism ofincorporation of long-chain fatty acids is unprecedented and isreminiscent of adenylation domains of NRPS modules (see alsoSupplementary Fig. 2). Several lipopeptide biosynthetic geneclusters also contain open reading frames (ORFs) homologousto fadD22,23 and it is plausible that activation of lipid moiety mightproceed through acyl-adenylate formation. Indeed, the loadingdomain of rifamycin synthetase initiates biosynthesis in a similarmanner24.
Structural investigations of the adenylate-forming family ofenzymes have revealed conformational flexibility in the relativeorientations of their two major domains12–14. Our modelling studiesof mycobacterial FadD proteins suggested that these proteins wouldbe composed of a large N-terminal domain and a smaller carboxy-terminal domain. Because mycobacterial FadD proteins could beclassified in two distinct families of FAAL and fatty acyl-CoA ligase(FACL) proteins, we examined whether they would exhibit differentconformations by using limited proteolysis25. Despite several poten-tial tryptic cleavage sites, trypsin makes a single cut on theFAAL proteins (Fig. 3a). The smaller fragment of approximately12 kDa was purified and characterized using mass spectrometry andN-terminal sequencing. The proteolytic cleavage site was mapped toa region between 470 and 471 residues of the FadD28 protein, whichis predicted to be present in linker region between the two structuralsubdomains (Fig. 3c). Interestingly, this tryptic digestion pattern issimilar to the adenylation domain of tyrocidine synthetase26. TheFACL proteins in contrast were much more susceptible to trypsincleavage, and did not yield any particularly stable protein fragment(Fig. 3b). The comparison of the sequences of the hinge regionbetween FAAL and FACL proteins also reveals differences in con-served amino-acid residues (Fig. 3d), suggesting that orientation ofthe C-terminal domain may be different between these two proteinfamilies.
The results presented here identify novel biochemical functionsfor the FadD proteins of mycobacteria and dissect mycobacterialcarboxylic acid activation metabolism revealing new potential drugtargets. More generally, the identification of a new class of FAALsand their role in the biosynthesis of complex hybrid metabolitesalso reveals a previously unknown molecular mechanism withwhich a limited number of genes can generate metabolitediversity. A
MethodsMaterialsThe bacterial artificial chromosome (BAC) library of M. tuberculosis H37Rv was obtainedfrom the Pasteur Research Institute, France. Radioactive materials were purchased fromAmersham Biosciences, Perkin Elmer Life Sciences and American RadiolabelledChemicals, Inc. All other chemicals used were of analytical grade.
Cloning, expression and purificationM. tuberculosis fadD and pks genes were cloned by PCR amplification using gene-specific
primers from the BAC genomic library. The two ORFs of fadD26 gene were of 1,881 and1,752 base pairs (bp), which correspond to the start-codon positions of 3,243,697 bp and3,243,568 bp respectively on the M. tuberculosis H37Rv genome. To minimize polymerasechain reaction (PCR) errors in larger PKS genes, shotgun libraries of BAC genomic DNAwere constructed. The fadD and pks genes were expressed as hexahistidine-tagged proteinsin the BL21-DE3 strain of E. coli using pET vector systems. Proteins were obtained in thesoluble fraction by inducing cultures at low temperatures and isopropylb-D-thiogalactopyranoside concentrations. These proteins were purified by Ni2þ-NTAaffinity chromatography followed by anion exchange chromatography using AKTA(Pharmacia) chromatographic system (Supplementary Fig. 3).
Enzyme assaysFatty acid activation by various FadD proteins was analysed by radio-TLC and HPLC. Thereaction mixtures included 50 mM Tris-HCl (pH 8.0), 100 mM 14C-fatty acid, 2 mM ATP,8 mM MgCl2 and 10 mM proteins in a 15 ml reaction volume and, wherever applicable,CoASH concentration was 2 mM. After incubation at 30 8C for 5 min, the reactions werequenched with 5% acetic acid and directly spotted on silica gel TLC plates. Products wereresolved on TLCs in n-butanol/acetic acid/water (80:25:40) solvent system at 4 8C. Theradioactive bands on TLC were detected using the phosphoimager (Fuji BAS5000). Thereactions were scaled up for HPLC (Shimadzu) analysis. After incubation, the reactionswere quenched and the products resolved on reverse phase (C8) column (Phenomenex)using an isocratic gradient at 50% B. Solvent A was composed of 20 mM ammoniumacetate (pH 5.4) and solvent B was acetonitrile-methanol (85:15, v/v). Products wereanalysed using ESI-MS (API QSTAR Pulsar i MS/MS, Applied Biosystems).
Synthesis of fatty acyl-AMP standardFatty acyl-AMP was synthesized by modifying the procedure used in ref. 27. The acyl-AMPwas characterized using ESI-MS.
Transfer of activated fatty acids to PKS proteinsFadD and its cognate PKS proteins were incubated along with 2 mM ATP, 8 mMMgCl2 and 100 mM 14C-fatty acid at 30 8C for 15 min. Reactions were quenched andproteins were separated using SDS–PAGE. The gels were stained with Coomassiebrilliant blue R250 and soaked in Amplify (Amersham) for 15 min. These were driedand exposed to phosphoimager. In the control reactions ATP was excluded from thereaction mixture.
Partial trypsin digestionVarious FadD proteins were incubated with 0.25 mg ml21 or 0.05 mg ml21 of trypsin at30 8C for different time periods. The reactions were quenched and analysed on 12%SDS–PAGE gels.
Computer analysisPrimary sequences of the genes were obtained from TubercuList (http://genolist.pasteur.fr/TubercuList/). Multiple sequence alignments of FadD proteins werecarried out by using the SeqWeb (http://www.gcg.com). The INSIGHT II package wasused for depiction of structural models.
Received 29 November 2003; accepted 2 February 2004; doi:10.1038/nature02384.
1. Moore, B. S. & Hertweck, C. Biosynthesis and attachment of novel bacterial polyketide synthase
starter units. Nat. Prod. Rep. 19, 70–99 (2002).
2. Hopwood, D. A. Genetic contributions to understanding polyketide synthases. Chem. Rev. 97,
2465–2498 (1997).
3. Spaink, H. P. et al. A novel highly unsaturated fatty acid moiety of lipo-oligosaccharide signals
determines host specificity of Rhizobium. Nature 354, 125–130 (1991).
4. Gokhale, R. S. & Tuteja, D. in Biotechnology (eds Rehm, H.-J. & Reed, G.) 341–372 (Wiley, Weinheim,
2001).
5. Kolattukudy, P. E., Fernandes, N. D., Azad, A. K., Fitzmaurice, A. M. & Sirakova, T. D. Biochemistry
and molecular genetics of cell-wall lipid biosynthesis in mycobacteria. Mol. Microbiol. 24, 263–270
(1997).
6. Sirakova, T. D., Thirumala, A. K., Dubey, V. S., Sprecher, H. & Kolattukudy, P. E. The Mycobacterium
tuberculosis pks2 gene encodes the synthase for the hepta- and octamethyl-branched fatty acids
required for sulfolipid synthesis. J. Biol. Chem. 276, 16833–16839 (2001).
7. Cole, S. T. et al. Deciphering the biology of Mycobacterium tuberculosis from the complete genome
sequence. Nature 393, 537–544 (1998).
8. Azad, A. K., Sirakova, T. D., Fernandes, N. D. & Kolattukudy, P. E. Gene knockout reveals a novel gene
cluster for the synthesis of a class of cell wall lipids unique to pathogenic mycobacteria. J. Biol. Chem.
272, 16741–16745 (1997).
9. Cox, J. S., Chen, B., McNeil, M. & Jacobs, W. R. Jr Complex lipid determines tissue-specific replication
of Mycobacterium tuberculosis in mice. Nature 402, 79–83 (1999).
10. Camacho, L. R. et al. Analysis of the phthiocerol dimycocerosate locus of Mycobacterium tuberculosis.
Evidence that this lipid is involved in the cell wall permeability barrier. J. Biol. Chem. 276,
19845–19854 (2001).
11. Rindi, L. et al. Involvement of the fadD33 gene in the growth of Mycobacterium tuberculosis in the liver
of BALB/c mice. Microbiol. 148, 3873–3880 (2002).
12. Conti, E., Franks, N. P. & Brick, P. Crystal structure of firefly luciferase throws light on a superfamily of
adenylate-forming enzymes. Structure 4, 287–298 (1996).
13. May, J. J., Kessler, N., Marahiel, M. A. & Stubbs, M. T. Crystal structure of DhbE, an archetype for aryl
acid activating domains of modular nonribosomal peptide synthetases. Proc. Natl Acad. Sci. USA 99,
12120–12125 (2002).
14. Gulick, A. M., Starai, V. J., Horswill, A. R., Homick, K. M. & Escalante-Semerena, J. C. The 1.75 A
crystal structure of acetyl-CoA synthetase bound to adenosine-5 0 -propylphosphate and coenzyme A.
Biochemistry 42, 2866–2873 (2003).
letters to nature
NATURE | VOL 428 | 25 MARCH 2004 | www.nature.com/nature444 © 2004 Nature Publishing Group

15. Fitzmaurice, A. M. & Kolattukudy, P. E. An acyl-CoA synthase (acoas) gene adjacent to the
mycocerosic acid synthase (mas) locus is necessary for mycocerosyl lipid synthesis in Mycobacterium
tuberculosis var. bovis BCG. J. Biol. Chem. 273, 8033–8039 (1998).
16. Fitzmaurice, A. M. & Kolattukudy, P. E. Open reading frame 3, which is adjacent to the mycocerosic
acid synthase gene, is expressed as an acyl coenzyme A synthase in Mycobacterium bovis BCG.
J. Bacteriol. 179, 2608–2615 (1997).
17. Saxena, P., Yadav, G., Mohanty, D. & Gokhale, R. S. A new family of type III polyketide synthases in
Mycobacterium tuberculosis. J. Biol. Chem. 278, 44780–44790 (2003).
18. Yadav, G., Gokhale, R. S. & Mohanty, D. SEARCHPKS: A program for detection and analysis of
polyketide synthase domains. Nucleic Acids Res. 31, 3654–3658 (2003).
19. Yadav, G., Gokhale, R. S. & Mohanty, D. Computational approach for prediction of domain
organization and substrate specificity of modular polyketide synthases. J. Mol. Biol. 328, 335–363
(2003).
20. Lambalot, R. H. et al. A new enzyme superfamily—the phosphopantetheinyl transferases. Chem. Biol.
3, 923–936 (1996).
21. Gokhale, R. S., Tsuji, S. Y., Cane, D. E. & Khosla, C. Dissecting and exploiting intermodular
communication in polyketide synthases. Science 284, 482–485 (1999).
22. Duitman, E. H. et al. The mycosubtilin synthetase of Bacillus subtilis ATCC6633: a multifunctional
hybrid between a peptide synthetase, an amino transferase, and a fatty acid synthase. Proc. Natl Acad.
Sci. USA 96, 13294–13299 (1999).
23. Tosato, V., Albertini, A. M., Zotti, M., Sonda, S. & Bruschi, C. V. Sequence completion,
identification and definition of the fengycin operon in Bacillus subtilis 168. Microbiology 143,
3443–3450 (1997).
24. Admiraal, S. J., Walsh, C. T. & Khosla, C. The loading module of rifamycin synthetase is an
adenylation-thiolation didomain with substrate tolerance for substituted benzoates. Biochemistry 40,
6116–6123 (2001).
25. Polverino de Laureto, P. et al. Protein aggregation and amyloid fibril formation by an SH3 domain
probed by limited proteolysis. J. Mol. Biol. 334, 129–141 (2003).
26. Dieckmann, R., Pavela-Vrancic, M., von Dohren, H. & Kleinkauf, H. Probing the domain structure
and ligand-induced conformational changes by limited proteolysis of tyrocidine synthetase 1. J. Mol.
Biol. 288, 129–140 (1999).
27. Keller, U., Kleinkauf, H. & Zocher, R. 4-Methyl-3-hydroxyanthranilic acid activating enzyme from
actinomycin-producing Streptomyces chrysomallus. Biochemistry 23, 1479–1484 (1984).
Supplementary Information accompanies the paper on www.nature.com/nature.
Acknowledgements The authors thank S. K. Basu for discussions. P.A. is a Senior Research
Fellow of CSIR, India. R.S.G is a Wellcome Trust International Senior Research Fellow in
India. This work was also supported by grants to the National Institute of Immunology by
DBT, India.
Competing interests statement The authors declare that they have no competing financial
interests.
Correspondence and requests for materials should be addressed to R.S.G. ([email protected]).
..............................................................
corrigendum
Delta-promoted filopodiamediate long-range lateralinhibition in Drosophila
Cyrille de Joussineau, Jonathan Soule, Marianne Martin,Christelle Anguille, Philippe Montcourrier & Daniel Alexandre
Nature 426, 555–559 (2003)..............................................................................................................................................................................
In the authors’ published address, ‘UMR 5539’ should read ‘CNRSUMR 5539’. An additional affiliation for the first and last authors(C.d.J. and D.A.) was omitted, who are also at the Laboratoire deNeurogenetique, Unite INSERM 432, Universite Montpellier II. A
..............................................................
erratum
Folding proteins in fatal ways
Dennis J. Selkoe
Nature 426, 900–904 (2003)..............................................................................................................................................................................
In this Insight Review Article, a competing interest was declared bythe author that was accidentally omitted. This statement is: “D.J.S. isa founding scientist of Athena Neurosciences, now Elan plc.” A
letters to nature
NATURE | VOL 428 | 25 MARCH 2004 | www.nature.com/nature 445© 2004 Nature Publishing Group

23. Gehring, N. H., Neu-Yilik, G., Schell, T., Hentze, M. W. & Kulozik, A. E. Y14 and hUpf3b form an
NMD-activating complex. Mol. Cell 11, 939–949 (2003).
24. Macchi, P. et al. Barentsz, a new component of the Staufen-containing ribonucleoprotein particles in
mammalian cells, interacts with Staufen in an RNA-dependent manner. J. Neurosci. 23, 5778–5788
(2003).
25. Rørth, P. Gal4 in the Drosophila female germline. Mech. Dev. 78, 113–118 (1998).
26. Horton, R. M., Cai, Z. L., Ho, S. N. & Pease, L. R. Gene splicing by overlap extension: tailor-made
genes using the polymerase chain reaction. Biotechniques 5, 528–535 (1991).
27. Ausubel, F. M. et al. (eds) Current Protocols in Molecular Biology vol. 1 (Wiley, New York, 1997).
28. Riechmann, V., Gutierrez, G. J., Filardo, P., Nebreda, A. R. & Ephrussi, A. Par-1 regulates stability of
the posterior determinant Oskar by phosphorylation. Nature Cell Biol. 4, 337–342 (2002).
29. Wilkie, G. S., Shermoen, A. W., O’Farrell, P. H. & Davis, I. Transcribed genes are localized according to
chromosomal position within polarized Drosophila embryonic nuclei. Curr. Biol. 9, 1263–1266
(1999).
30. Lehmann, R. & Nusslein-Volhard, C. Abdominal segmentation, pole cell formation, and
embryonic polarity require the localized activity of oskar, a maternal gene in Drosophila. Cell 47,
141–152 (1986).
Supplementary Information accompanies the paper on www.nature.com/nature.
Acknowledgements We thank A.-M. Voie for embryo injections for transgenesis, S. Curado for
the oskA87, Nanos-Gal4:VP16 recombinant stock, A. Cyrklaff for the oskar in situ probe, P. Rørth
for the pCog-Gal4:VP16 driver, D. St Johnston for anti-Staufen antibody, and members of the
Ephrussi laboratory, V. Hachet and E. Izaurralde for advice and comments on the manuscript.
O.H. was supported in part by a predoctoral ‘allocation de recherche’ from the French
government.
Competing interests statement The authors declare that they have no competing financial
interests.
Correspondence and requests for materials should be addressed to A.E.
..............................................................
erratum
Enzymic activation and transferof fatty acids as acyl-adenylatesin mycobacteria
Omita A. Trivedi, Pooja Arora, Vijayalakshmi Sridharan, Rashmi Tickoo,Debasisa Mohanty & Rajesh S. Gokhale
Nature 428, 441–445 (2004)..............................................................................................................................................................................
In this Letter to Nature, lanes FadD17, -19, and -28 of Fig. 1a did notappear. Figure 1a should have appeared as shown. A
letters to nature
NATURE | VOL 428 | 29 APRIL 2004 | www.nature.com/nature 963© 2004 Nature Publishing Group

7. Horn, R. Coupled movements in voltage-gated ion channels. J. Gen. Physiol. 120, 449–453 (2002).
8. Frank, J. Single-particle imaging of macromolecules by cryo-electron microscopy. Annu. Rev. Biophys.
Biomol. Struct. 31, 303–319 (2002).
9. Henderson, R. The potential and limitations of neutrons, electrons and X-rays for atomic resolution
microscopy of unstained biological molecules. Q. Rev. Biophys. 28, 171–193 (1995).
10. Adrian, M., Dubochet, J., Fuller, S. D. & Harris, J. R. Cryo-negative staining. Micron 29, 145–160 (1998).
11. Golas, M. M., Sander, B., Will, C. L., Luhrmann, R. & Stark, H. Molecular architecture of the
multiprotein splicing factor SF3b. Science 300, 980–984 (2003).
12. Saxton, W. O. & Baumeister, W. The correlation averaging of a regularly arranged bacterial cell
envelope protein. J. Microsc. 127, 127–138 (1982).
13. van Heel, M. Similarity measures between images. Ultramicroscopy 21, 95–100 (1987).
14. Conway, J. F. et al. Characterization of a conformational epitope on hepatitis B virus core antigen and
quasiequivalent variations in antibody binding. J. Virol. 77, 6466–6473 (2003).
15. Rosenthal, P. B. & Henderson, R. Optimal determination of particle orientation, absolute hand, and
contrast loss in single-particle electron cryomicroscopy. J. Mol. Biol. 333, 721–745 (2003).
16. Laine, M. et al. Atomic proximity between S4 segment and pore domain in Shaker potassium
channels. Neuron 39, 467–481 (2003).
17. Gandhi, C. S., Clark, E., Loots, E., Pralle, A. & Isacoff, E. Y. The orientation and molecular movement
of a Kþ channel voltage-sensing domain. Neuron 40, 515–525 (2003).
18. Neale, E. J., Elliott, D. J., Hunter, M. & Sivaprasadarao, A. Evidence for intersubunit interactions
between S4 and S5 transmembrane segments of the Shaker potassium channel. J. Biol. Chem. 278,
29079–29085 (2003).
19. Chandy, K. G. & Gutman, G. A. in Ligand and Voltage-Gated Channels (ed. North, R. A.) 1–72 (CRC,
Boca Raton, 1995).
20. Seoh, S. A., Sigg, D., Papazian, D. M. & Bezanilla, F. Voltage-sensing residues in the S2 and S4 segments
of the Shaker Kþ channel. Neuron 16, 1159–1167 (1996).
21. Aggarwal, S. K. & MacKinnon, R. Contribution of the S4 segment to gating charge in the Shaker Kþ
channel. Neuron 16, 1169–1177 (1996).
22. Orlova, E. V. et al. Structure of keyhole limpet hemocyanin type 1 (KLH1) at 15 A resolution by
electron cryomicroscopy and angular reconstitution. J. Mol. Biol. 271, 417–437 (1997).
23. Gabashvili, I. S. et al. Solution structure of the E. coli 70S ribosome at 11.5 A resolution. Cell 100,
537–549 (2000).
24. Ludtke, S. J., Baldwin, P. R. & Chiu, W. EMAN: semiautomated software for high-resolution single-
particle reconstructions. J. Struct. Biol. 128, 82–97 (1999).
25. van Heel, M., Harauz, G., Orlova, E. V., Schmidt, R. & Schatz, M. A new generation of the IMAGIC
image processing system. J. Struct. Biol. 116, 17–24 (1996).
26. van Heel, M. Angular reconstitution: a posteriori assignment of projection directions for 3D
reconstruction. Ultramicroscopy 21, 111–123 (1987).
27. Frank, J. et al. SPIDER and WEB: processing and visualization of images in 3D electron microscopy
and related fields. J. Struct. Biol. 116, 190–199 (1996).
28. Sander, B., Golas, M. M. & Stark, H. Automatic CTF correction for single particles based upon
multivariate statistical analysis of individual power spectra. J. Struct. Biol. 142, 392–401 (2003).
29. Hawkes, P. W. in Computer Processing of Electron Microscopic Images (ed. Hawkes, P. W.) 1–33
(Springer, Berlin, 1980).
30. Jones, T. A., Zou, J. Y., Cowan, S. W. & Kjeldgaard Improved methods for building protein models in
electron density maps and the location of errors in these models. Acta Crystallogr. A 47, 110–119 (1991).
Supplementary Information accompanies the paper on www.nature.com/nature.
Acknowledgements We thank members of the MacKinnon lab, S. Darst, N. Opalka and D. Stokes
for helpful discussions. This work was supported by grants from the NIH to D.N.W. and R.M.
R.M is an investigator in the Howard Hughes Medical Institute.
Competing interests statement The authors declare that they have no competing financial
interests.
Correspondence and requests for materials should be addressed to R.M.
([email protected]). The electron microscopy map has been deposited with European
Bioinformatics Institute (www.ebi.ac.uk/msd/emd-2771.map).
..............................................................
corrigenda
Enzymic activation and transferof fatty acids and acyl-adenylatesin mycobacteriaOmita A. Trivedi, Pooja Arora, Vijayalakshmi Sridharan, Rashmi Tickoo,Debasisa Mohanty & Rajesh S. Gokhale
Nature 428, 441–445 (2004)..............................................................................................................................................................................
The structures of lauroyl–AMP and lauroyl–CoA are incorrectlyrepresented in Fig. 1c, d of this Letter, which shows the L-form ratherthan the natural D-form of adenosine; there is no phosphate groupat the 2 0-position of the ribosyl moiety of lauroyl–AMP (Fig. 1c)and the phosphate group in lauroyl–CoA (Fig. 1d) should be at the3 0-position. These errors do not affect our conclusions. A
..............................................................
corrigendum
Reduction of hysteresis losses inthe magnetic refrigerant Gd5Ge2Si2by the addition of ironVirgil Provenzano, Alexander J. Shapiro & Robert D. Shull
Nature 429, 853–857 (2004)..............................................................................................................................................................................
It has come to our attention that we failed to cite in our Letter toNature a key publication1 that describes changes in the magneto-caloric capabilities of Gd5(Si2Ge2) as a result of alloying thecompound with small amounts of various metals, including iron.That paper supports the findings presented in our Letter. However,we also show that it is important to consider the magnetic hysteresisin addition to the magnetocaloric effect when assessing the useful-ness of a material as a magnetic refrigerant. A
1. Pecharsky, V. K. & Gschneidner, K. A. Effect of alloying on the giant magnetocaloric effect of
Gd5(Si2Ge2). J. Magn. Magn. Mater. Res. 167, L179–L184 (1997).
letters to nature
NATURE | VOL 430 | 12 AUGUST 2004 | www.nature.com/nature810 © 2004 Nature Publishing Group

23. Gehring, N. H., Neu-Yilik, G., Schell, T., Hentze, M. W. & Kulozik, A. E. Y14 and hUpf3b form an
NMD-activating complex. Mol. Cell 11, 939–949 (2003).
24. Macchi, P. et al. Barentsz, a new component of the Staufen-containing ribonucleoprotein particles in
mammalian cells, interacts with Staufen in an RNA-dependent manner. J. Neurosci. 23, 5778–5788
(2003).
25. Rørth, P. Gal4 in the Drosophila female germline. Mech. Dev. 78, 113–118 (1998).
26. Horton, R. M., Cai, Z. L., Ho, S. N. & Pease, L. R. Gene splicing by overlap extension: tailor-made
genes using the polymerase chain reaction. Biotechniques 5, 528–535 (1991).
27. Ausubel, F. M. et al. (eds) Current Protocols in Molecular Biology vol. 1 (Wiley, New York, 1997).
28. Riechmann, V., Gutierrez, G. J., Filardo, P., Nebreda, A. R. & Ephrussi, A. Par-1 regulates stability of
the posterior determinant Oskar by phosphorylation. Nature Cell Biol. 4, 337–342 (2002).
29. Wilkie, G. S., Shermoen, A. W., O’Farrell, P. H. & Davis, I. Transcribed genes are localized according to
chromosomal position within polarized Drosophila embryonic nuclei. Curr. Biol. 9, 1263–1266
(1999).
30. Lehmann, R. & Nusslein-Volhard, C. Abdominal segmentation, pole cell formation, and
embryonic polarity require the localized activity of oskar, a maternal gene in Drosophila. Cell 47,
141–152 (1986).
Supplementary Information accompanies the paper on www.nature.com/nature.
Acknowledgements We thank A.-M. Voie for embryo injections for transgenesis, S. Curado for
the oskA87, Nanos-Gal4:VP16 recombinant stock, A. Cyrklaff for the oskar in situ probe, P. Rørth
for the pCog-Gal4:VP16 driver, D. St Johnston for anti-Staufen antibody, and members of the
Ephrussi laboratory, V. Hachet and E. Izaurralde for advice and comments on the manuscript.
O.H. was supported in part by a predoctoral ‘allocation de recherche’ from the French
government.
Competing interests statement The authors declare that they have no competing financial
interests.
Correspondence and requests for materials should be addressed to A.E.
..............................................................
erratum
Enzymic activation and transferof fatty acids as acyl-adenylatesin mycobacteria
Omita A. Trivedi, Pooja Arora, Vijayalakshmi Sridharan, Rashmi Tickoo,Debasisa Mohanty & Rajesh S. Gokhale
Nature 428, 441–445 (2004)..............................................................................................................................................................................
In this Letter to Nature, lanes FadD17, -19, and -28 of Fig. 1a did notappear. Figure 1a should have appeared as shown. A
letters to nature
NATURE | VOL 428 | 29 APRIL 2004 | www.nature.com/nature 963© 2004 Nature Publishing Group