Embed Size (px)
Citation preview

(CANCER RESEARCH 49, 1916-1920, April 15, 1989]
Formation of Cigarette Smoke-induced DNA Adducts in the Rat Lungand Nasal Mucosa1
Kämest)C. Gupta, Mohan L. Sopori, and C. G. Gairola2
Department of Pharmacology, Baylor College of Medicine, Houston, Texas 77030 [R. C. G.J; Lovelace BiomédicalFoundation, Albuquerque, New Mexico 87108 [M. L.S.]; and Tobacco and Health Research Institute and the Graduate Center for Toxicology, University of Kentucky, Lexington, Kentucky 40506 [C. G. G.J
ABSTRACT
The formation of DNA adducts in the nasal, lung, and liver tissues ofrats exposed daily to fresh smoke from a University of Kentucky referencecigarette (2R1) for up to 40 weeks was examined. The amount of smoketotal particulate matter (TPM) inhaled and the blood carboxyhemoglobin(COHb) values averaged 5-5.5 mg smoke TPM/day/rat and 5.5%, respectively. The pulmonary AHH activity measured at the termination ofeach experiment showed an average increase of about two- to threefoldin the smoke-exposed groups. These observations suggested that animalseffectively inhaled both gaseous and particulate phase constituents ofcigarette smoke. DNAs from nasal, lung, and liver tissue were extractedand analyzed by an improved 32P-postlabeling procedure. The results
showed that the mainstream cigarette smoke induced a spectrum of atleast four new DNA adducts in the nasal mucosa of the exposed rats andthe magnitude of these adducts increased with the duration of exposure.In the lung tissue, the smoke exposure induced an accumulation of oneDNA adduct, which upon cessation of exposure for 19 weeks was reducedby about 75%. Smoke-related adducts were not detected in the liver, anontarget tissue. Selective chromatography and butanol extractabilitysuggested that the nasal and lung DNA adducts are aromatic and/orhydrophobic in nature and that the smoke-related lung DNA-adduct maycontain polar group(s). These data demonstrate the DNA-damagingpotential of long term fresh cigarette smoke exposure and suggest theability of the tissue to partially recover from such damage followingcessation of the exposure.
INTRODUCTION
Several epidemiological studies have strongly implicated cigarette smoking with higher incidence of respiratory tract cancers, including those of larynx, the oral cavity, and the lung (1,2). In the rat, nose-only chronic exposure to cigarette smokehas been found to induce a variety of hyperplastic and meta-plastic changes in the upper and lower respiratory tract ofanimals (3). Daily exposure of rats to cigarette smoke for 2.5years was found to increase the incidence of tumors in the nasalas well as pulmonary region of the respiratory tract (4). By whatmechanism(s) the cigarette smoke induces such precancerousand cancerous alterations in the respiratory tract is not clearlyunderstood. It is generally believed that the initial event in thechemical carcinogenic process involves an interaction betweenthe reactive chemical species and the cellular DNA to formadducts (5). Cigarette smoke, which is a complex chemicalmixture (6, 7) and is known to contain a number of carcinogenicand mutagenic substances (8-9), may conceivably interact withthe cellular DNA in the respiratory tract of exposed individualsto form adducts which may subsequently initiate a neoplasticprocess. To ascertain whether exposure to cigarette smokeindeed induces qualitative and quantitative changes in the cellular DNA, we have examined the formation of DNA adducts
Received 8/18/88; revised 12/12/88; accepted 1/18/89.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1This work was supported by the EPA Cooperative Agreement CR 813840(R. C. G.), and by grants from USPHS [CA-30606 (R. C. G.). DA 04208-NIHCTR 157 (M. L. S.), and KTRB41031 (C. G. G.)].
! To whom requests for reprints should be addressed, at Tobacco and HealthResearch Institute, University of Kentucky, Lexington, KY 40546-0236.
in the target tissues, i.e., nasal mucosa and the lung, and anontarget tissue, liver, of rats given exposures to mainstreamsmoke by inhalation route. The results demonstrate that chronicexposure to smoke does induce the formation of new DNAadducts in the nasal mucosa and significantly increases themagnitude of already existing DNA adducts in the lungs.
MATERIALS AND METHODS
Animals and Treatment Groups. Male Sprague-Dawley rats, weighingabout 200 g (9-10 weeks old) were purchased from Harlan/Sprague-Dawley and maintained in quarantine rooms for 2 weeks. Animalsshowing no abnormal clinical signs were randomly selected for thestudy and divided into the following three groups: (a) room controls(RC), handled once a week during weekly cage cleaning; (b) shamcontrols (SH), given daily treatments identical to those of smoke-exposed groups but in the absence of smoke; and (c) smoke-exposedgroups (SM), exposed to fresh mainstream smoke from a high tar/highnicotine University of Kentucky reference cigarette (2R1) for up to 40weeks.
All animals were housed in hanging stainless-steel wire cages andmaintained under a light cycle of 12 h in environmentally controlledBioclean rooms equipped with HEPA filters and undergoing 40 airchanges per hour to minimize the exposure of animals to extraneousparticulates. Animals had free access to Purina Rodent Chow (RalstonPurina, St. Louis, MO) and water ad libitum.
Exposure System. The smoke generation and exposure system hasbeen described in detail (10, II). Briefly, the mainstream smoke wasgenerated once each minute by a 2-s, 35-ml puff from the cigarette. Anaverage of 10 puffs were generated from each 2R1 cigarette, and thesmoke was transferred to the exposure chamber, into which the animals'
noses were protruding through the ports. In this system, the animalsreceived exposure to fresh mainstream cigarette smoke by nose only inan intermittent fashion analogous to human smoking.
Indicators of Cigarette Smoke Exposure. Exposure of animals tocigarette smoke was ascertained by measuring inhaled TPM3. COHb
levels, and the induction of pulmonary AHH activity.Daily estimates of the inhaled TPM by animals and biweekly meas
urements of blood COHb levels were made as described elsewhere (11).Blood COHb levels were determined spectrophotometrically on bloodsamples obtained from the orbital sinus of a few rats, immediately afterthe completion of sham and smoke-exposure treatments. PulmonaryAHH activity was measured in 9000 x g supernatants of lung homog-enates by a radiometrie procedure as described earlier (12).
Isolation of DNA. DNA was isolated from 0.2 to 0.4 g of individuallung and liver, and from 0.05 to 0.09 g of pooled (from four to fiverats) nasal mucosa, using a procedure in which protein and RNA areremoved by extensive digestions with proteinase K and RNases, respectively, and by solvent extractions (13). DNA yields from lung, liver,and nasal tissues were 1.8 ±0.2, 1.6 ±0.2, and 2.3 ±0.4 mg DNA/gtissue, respectively.
Analysis of Adducts. DNA adducts were analyzed by 32P-postlabeling
assay (14) after enhancement of the sensitivity of detection (15, 16).Briefly, DNA (5-10 ug) was enzymatically hydrolyzed to 3'-mono-
phosphates of normal deoxynucleosides and adducts. Adducts wereenriched by extraction in 1-butanol (15) or by nuclease PI-mediatedprocedure (16), 5'-32P labeled in the presence of molar excess of
' The abbreviations used are: TPM, total particulate matter; COHb, bloodcarboxyhemoglobin; AHH, aryl hydrocarbon hydroxylase; TLC, thin-layer chromatography.
1916
Research. on November 26, 2018. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from
CIGARETTE SMOKE-INDUCED DNA ADDUCTS IN RAT
[y-32P]ATP >3000 Ci/mmol) and polynucleotide kinase, and analyzedby multidirectional PEI-cellulose TLC (see Fig. 2 legends for specificsolvents). To calculate adduct levels, total nucleotides (5 ng) were also32P labeled using the same specific activity [7-32P]ATP as used foradducts and analyzed by one-directional PEI-cellulose TLC (15). Ad-duct levels vere then evaluated by calculating relative adduct labelingas (cpm in a Iduct/cpm in total nucleotides) x (I/dilution factor). Thesevalues were then translated into atto(a)mole of adducts per ^g DNA(15).
RESULTS
Smoke Ãx̄posure. The exposure of animals to cigarette smokeas measured daily by an estimation of the TPM intake indicatedan average value of 1.46 ±0.56 mg/session/rat and suggesteda dose of about 5-5.5 mg/kg body weight. The blood COHblevels of S VI animals averaged 5.5 ±1% over the entire exposure duration as compared to less than 1% in SH group (Fig.1). The pulmonary AHH activity was also significantly elevatedin the SM group and averaged about two- to threefold higherthan the si am group (Fig. 1).
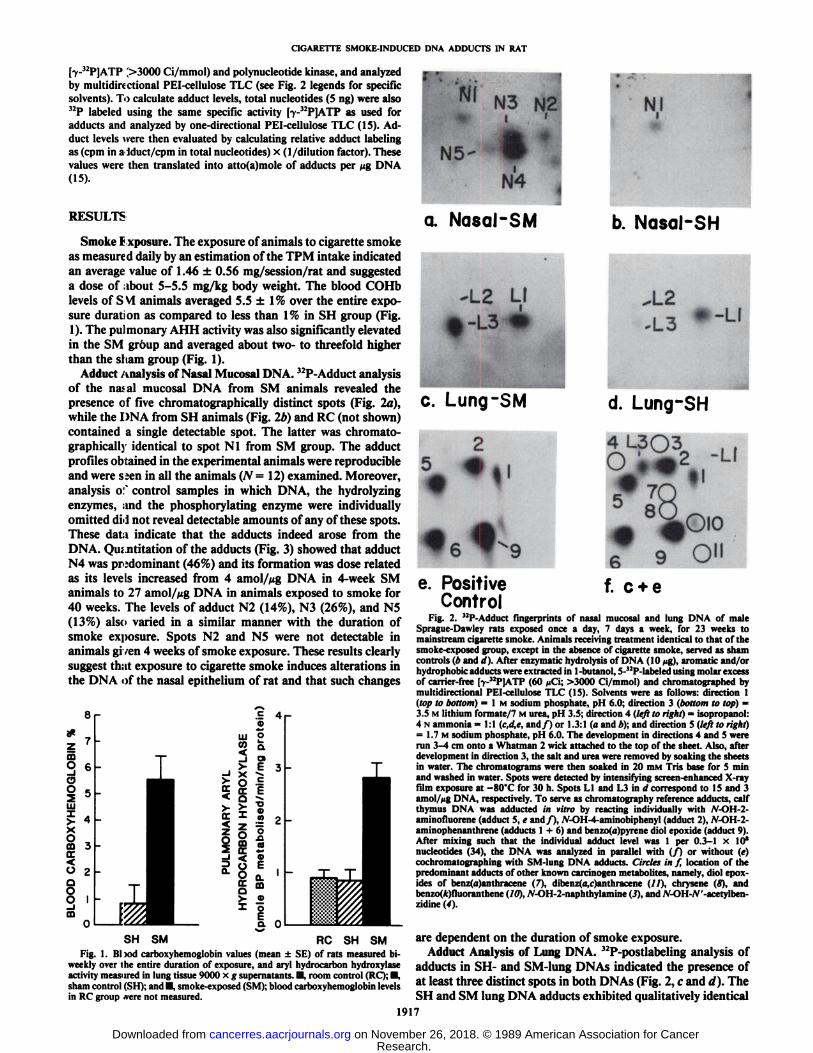
Adduct Analysis of Nasal Mucosa! DNA. 32P-Adduct analysis
of the nasal mucosal DNA from SM animals revealed thepresence of five Chromatograph ¡cally distinct spots (Fig. 2a),while the DNA from SH animals (Fig. 2b) and RC (not shown)contained a single detectable spot. The latter was chromato-graphically identical to spot Nl from SM group. The adductprofiles obtained in the experimental animals were reproducibleand were s;en in all the animals (N= 12) examined. Moreover,analysis o:' control samples in which DNA, the hydrolyzing
enzymes, and the phosphorylating enzyme were individuallyomitted dH not reveal detectable amounts of any of these spots.These data indicate that the adducts indeed arose from theDNA. Qu; nthut ion of the adducts (Fig. 3) showed that adductN4 was predominant (46%) and its formation was dose relatedas its levels increased from 4 amol/Mg DNA in 4-week SManimals to 27 amol/Mg DNA in animals exposed to smoke for40 weeks. The levels of adduct N2 (14%), N3 (26%), and N5(13%) also varied in a similar manner with the duration ofsmoke exposure. Spots N2 and N5 were not detectable inanimals given 4 weeks of smoke exposure. These results clearlysuggest that exposure to cigarette smoke induces alterations inthe DNA of the nasal epithelium of rat and that such changes
a 0SH SM RC SH SM
Fig. 1. Bl >od carboxyhemoglobin values (mean ±SE) of rats measured biweekly over the entire duration of exposure, and aryl hydrocarbon hydroxylaseactivity measured in lung tissue 9000 x g supernatants. H. room control (RC); D,sham control (SH); and •¿�.smoke-exposed (SM); blood carboxyhemoglobin levelsin RC group »erenot measured.
a. Nasal-SM
"'b. Nasal-SH
-LI
c. Lung-SM d. Lung-SH
Q>«VU
9 O"
e. PositiveControl
f. c + eFig. 2. •¿�'•!'-\ddiici fingerprints of nasal mucosal and lung DNA of male
Sprague-Dawley rats exposed once a day, 7 days a week, for 23 weeks tomainstream cigarette smoke. Animals receiving treatment identical to that of thesmoke-exposed group, except in the absence of cigarette smoke, served as shamcontrols (b and d). After enzymatic hydrolysis of DNA (10 >ig),aromatic and/orhydrophobic adducts were extracted in 1-butanol, 5-"P-labeled using molar excessof carrier-free [y-"P]ATP (60 ¿iCi;>3000 Ci/mmol) and chromatographed bymultidirectional PEI-cellulose TLC (15). Solvents were as follows: direction 1(top to bottom) = l M sodium phosphate, pH 6.0; direction 3 (bottom to top) =3.5 M lithium formate/7 M urea, pH 3.5; direction 4 (left to right) = isopropanol:4 N ammonia = 1:1 (c,d,e, and/) or 1.3:1 (a and *); and direction 5 (left to right)= 1.7 M sodium phosphate, pi I 6.0. The development in directions 4 and 5 wererun 3-4 cm onto a Whatman 2 wick attached to the top of the sheet. Also, afterdevelopment in direction 3, the salt and urea were removed by soaking the sheetsin water. The chromatograms were then soaked in 20 HIMTris base for 5 minand washed in water. Spots were detected by intensifying screen-enhanced X-rayfilm exposure at —¿�80"Cfor 30 h. Spots LI and L3 in d correspond to 15 and 3
amol/Mg DNA, respectively. To serve as chromatography reference adducts, calfthymus DNA was adducted in vitro by reacting individually with .V-OI1 2aminofluorene (adduct 5, e and/), JV-OH-4-aminobiphenyl (adduct 2), N-OH-2-aminophenanthrene (adducts 1 + 6) and benzo(a)pyrene diol epoxide (adduct 9).After mixing such that the individual adduct level was 1 per 0.3-1 x 10'
nucleotides (34), the DNA was analyzed in parallel with (/) or without (e)cochromatographing with SM-lung DNA adducts. Circles in f, location of thepredominant adducts of other known carcinogen metabolites, namely, diol epox-ides of benz(a)anthracene (7), dibenz(a,r)anthracene (//), chrysene (8), andbenzo(*)fluoranthene (10), Ar-OH-2-naphthylamine (3), and A'-OH-/V'-acetylben-
zidine (4).
are dependent on the duration of smoke exposure.Adduct Analysis of Lung DNA. 32P-postlabeling analysis of
adducts in SH- and SM-lung DNAs indicated the presence ofat least three distinct spots in both DNAs (Fig. 2, c and d). TheSH and SM lung DNA adducts exhibited qualitatively identical
1917
Research. on November 26, 2018. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from
CIGARETTE SMOKE-INDUCED DNA ADDUCTS IN RAT
24
16 24
Time (Weeks)32 40
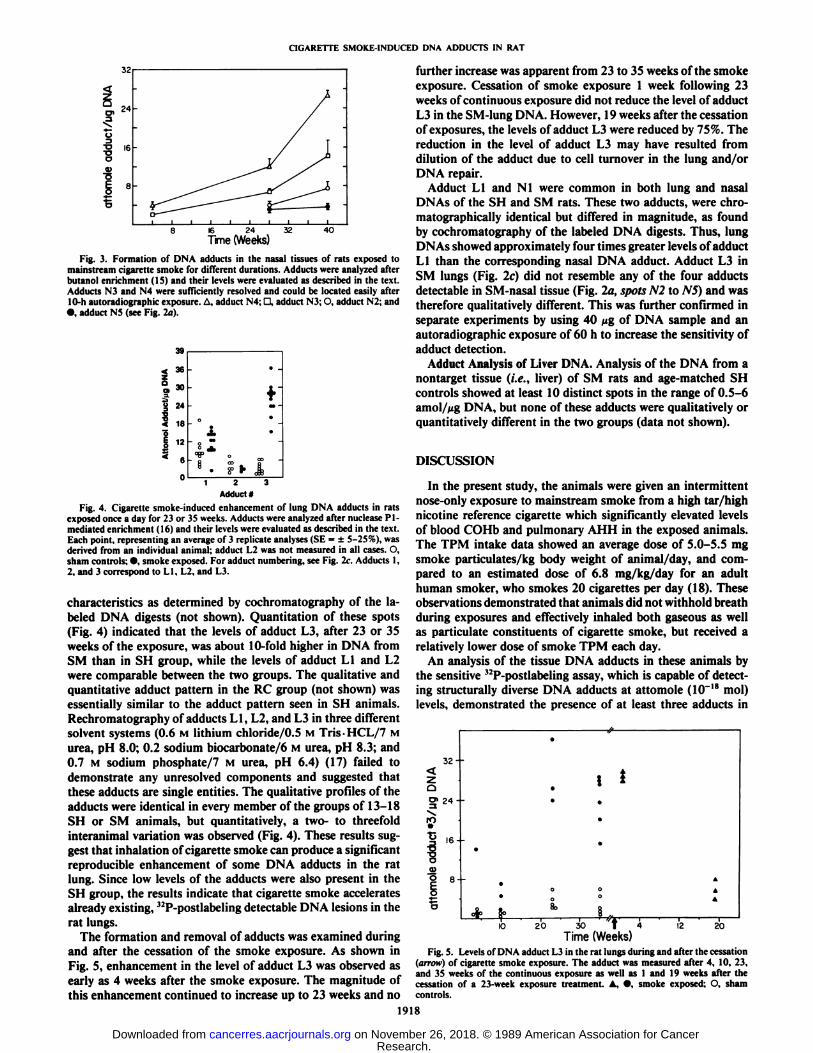
Fig. 3. Formation of DNA adducts in the nasal tissues of rats exposed tomainstream cigarette smoke for different durations. Adducts were analyzed afterbutanol enrichment (15) and their levels were evaluated as described in the text.Adducts N3 and N4 were sufficiently resolved and could be located easily afterI0-h autoradiographic exposure. A, adduci N4: D, adduci N3; O. adduci N2; and•¿�.adduci N5 (see Fig. 2a).
1zo3iB1•j38302418126a.*»-0
«1-
o""4>*
o"fi00 rP
Adduci*Fig. 4. Cigarette smoke-induced enhancemenl of lung DNA adducls in rats
exposed once a day for 23 or 35 weeks. Adducls were analyzed after nuclease Pl-medialed enrichmenl (16) and iheir levels were evalualed as described in ihe lexl.Each poinl, represenling an average of 3 replicale analyses (SE = ±5-25%), wasderived from an individual animal; adduci L2 was not measured in all cases. O,sham controls: •¿�.smoke exposed. For adduct numbering, see Fig. 2c. Adducts 1.2, and 3 correspond to LI. L2. and L3.
characteristics as determined by cochromatography of the labeled DNA digests (not shown). Quantitation of these spots(Fig. 4) indicated that the levels of adduct L3, after 23 or 35weeks of the exposure, was about 10-fold higher in DNA fromSM than in SH group, while the levels of adduct LI and L2were comparable between the two groups. The qualitative andquantitative adduct pattern in the RC group (not shown) wasessentially similar to the adduct pattern seen in SH animals.Rechromatography of adducts LI, L2, and L3 in three differentsolvent systems (0.6 M lithium chloride/0.5 M Tris-HCL/7 Murea, pH 8.0; 0.2 sodium biocarbonate/6 M urea, pH 8.3; and0.7 M sodium phosphate/7 M urea, pH 6.4) (17) failed todemonstrate any unresolved components and suggested thatthese adducts are single entities. The qualitative profiles of theadducts were identical in every member of the groups of 13-18SH or SM animals, but quantitatively, a two- to threefoldinteranimal variation was observed (Fig. 4). These results suggest that inhalation of cigarette smoke can produce a significantreproducible enhancement of some DNA adducts in the ratlung. Since low levels of the adducts were also present in theSH group, the results indicate that cigarette smoke acceleratesalready existing, 12P-postlabeling detectable DNA lesions in the
rat lungs.The formation and removal of adducts was examined during
and after the cessation of the smoke exposure. As shown inFig. 5, enhancement in the level of adduct L3 was observed asearly as 4 weeks after the smoke exposure. The magnitude ofthis enhancement continued to increase up to 23 weeks and no
further increase was apparent from 23 to 35 weeks of the smokeexposure. Cessation of smoke exposure 1 week following 23weeks of continuous exposure did not reduce the level of adductL3 in the SM-lung DNA. However, 19 weeks after the cessationof exposures, the levels of adduct L3 were reduced by 75%. Thereduction in the level of adduct L3 may have resulted fromdilution of the adduct due to cell turnover in the lung and/orDNA repair.
Adduct LI and Nl were common in both lung and nasalDNAs of the SH and SM rats. These two adducts, were chro-matographically identical but differed in magnitude, as foundby cochromatography of the labeled DNA digests. Thus, lungDNAs showed approximately four times greater levels of adductLI than the corresponding nasal DNA adduct. Adduct L3 inSM lungs (Fig. 2c) did not resemble any of the four adductsdetectable in SM-nasal tissue (Fig. la, spots N2 to NS) and wastherefore qualitatively different. This was further confirmed inseparate experiments by using 40 ^g of DNA sample and anautoradiographic exposure of 60 h to increase the sensitivity ofadduct detection.
Adduct Analysis of Liver DNA. Analysis of the DNA from anontarget tissue (i.e., liver) of SM rats and age-matched SHcontrols showed at least 10 distinct spots in the range of 0.5-6amol/^g DNA, but none of these adducts were qualitatively orquantitatively different in the two groups (data not shown).
DISCUSSION
In the present study, the animals were given an intermittentnose-only exposure to mainstream smoke from a high tar/highnicotine reference cigarette which significantly elevated levelsof blood COHb and pulmonary AHH in the exposed animals.The TPM intake data showed an average dose of 5.0-5.5 mgsmoke particulates/kg body weight of animal/day, and compared to an estimated dose of 6.8 mg/kg/day for an adulthuman smoker, who smokes 20 cigarettes per day (18). Theseobservations demonstrated that animals did not withhold breathduring exposures and effectively inhaled both gaseous as wellas paniculate constituents of cigarette smoke, but received arelatively lower dose of smoke TPM each day.
An analysis of the tissue DNA adducts in these animals bythe sensitive "P-postlabeling assay, which is capable of detecting structurally diverse DNA adducts at attomole (10~18 mol)
levels, demonstrated the presence of at least three adducts in
32--
Cl0«24--
ro•¿�c
I6--
9 8--
10 20 30 t 4
Time (Weeks)12 20
Fig. 5. Levels of DNA adduct L3 in the ral lungs during and after Ihe cessalion(arrow) of cigarelle smoke exposure. The adduci was measured after 4. 10, 23,and 35 weeks of Ihe conlinuous exposure as well as 1 and 19 weeks after Ihecessalion of a 23-week exposure Irealmenl. A, •¿�,smoke exposed: O. shamcontrols.
1918
Research. on November 26, 2018. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from

CIGARETTE SMOKE-INDUCED DNA ADDUCTS IN RAT
the lungs o ' adult control rats. Prolonged exposure of animals
to mainstream cigarette smoke significantly enhanced one ofthe lung DNA adducts by several fold. In contrast, four out offive nasal DNA adducts formed in the smoke-exposed rats wereundetectable in control rats. Since the composition of cigarettesmoke is complex (6, 7), the differences between the DNAadducts of nasal and lung tissues may reflect the diversity ofreactive constituents and their differential absorption in different regions of the respiratory tract.
The smoke-related nasal and lung DNA adducts were aromatic and/Dr hydrophobic in nature, as reflected by their ex-tractability in 1-butanol (15) and selective chromatography (14,15). In comparison to the lung DNA adducts, the adducts inthe nasal DNA were less hydrophobic as suggested by theirfaster Chromatographie mobility and requirement for less polardeveloping solvent in direction 4 (Fig. 2). Identity of the lungDNA addi et L3 was investigated by its extractability in 1-butanol in the presence and absence of tetrabutylammoniumchloride, an ion-pairing agent known to increase extractabilityof polar compounds into organic solvents (15, 19). This adductwas found to be almost completely (95%) lost in the absence ofthe ion-pairing agent, suggesting the presence of polar group(s)in this adcuct structure. Identity of this adduct was furtherinvestigated by comparison with several reference DNA adducts. It wjs found that adduct L3 did not cochromatographwith any o *the reference DNA adducts prepared by reacting
calf thymus DNA in vitro with active metabolites of 4-amino-biphenyl, :!-naphthylamine, TV'-acetylbenzidine, 2-aminoflu-
orene, 2-atninophenanthrene, benzo(a)pyrene, benz(u)anthra-cene, dib;nz(a,c)anthracene, benzo(A)fluoranthene, andchrysene (Fig. 2/). Since some of these chemicals are presentin cigarette smoke, our results suggest that these constituentsof cigarette smoke may not be directly responsible for theformation of the DNA adducts in the lungs of smoke-exposedrats. Simili r cochromatography experiments indicated that thesmoke-induced adducts in nasal DNA might also be unrelatedto the above carcinogens in the cigarette smoke. It is welldocumented that cigarette smoke contains many other DNA-reactive constituents than those examined above. The enhancement of lung DNA adduct and the formation of new nasalDNA adducts in the smoke-exposed animals may, therefore,occur as a result of an interaction of these other presumablymore polar smoke constituents with cellular DNA. Inability ofthe 32P-postlabeling assay to detect low levels (<1 adduct/105-106 nucleo) ides) of certain types of DNA damage, known to be
caused by t le tobacco specific nitrosamines, aldehydes or highlyreactive rac icals in smoke (20-24), may suggest that the adductsdetected in this study possibly represent only a fraction of thetotal DNA-damaging potential of the cigarette smoke. However, it is liso possible that some of the smoke carcinogens,when present with other fresh smoke constituents, e.g., CO,may escape interaction with cellular DNA because of the inability of Cyt-P-450 enzymes to activate them to reactive speciesor inactiva ion of the reactive metabolites before their interaction with DNA.
In order to characterize chemically the smoke-induced DNAadducts by mass spectroscopic technique, 0.1-1 Mgof specificadduct wo'ild be required. This amount of individual adductcorresponds to approximately 10-100 g of total cellular DNAor 5-50 kg of tissue. Obviously, these constraints make the useof current spectroscopic methods impractical and, therefore,alternative approaches are being explored to provide someinsight into the chemical nature of these adducts.
Randera ehand coworkers (25, 26) have reported the presence
of one major and several minor adducts, in addition to adiagonal radioactive zone containing presumably unresolvedDNA adducts in the bronchus and larynx of heavy smokers,and the skin, lung, and heart of mice treated topically withcigarette-smoke condensate. Although we have observed discrete adduct spots reproducibly, no diagonal zone of adductradioactivity was observed in our studies. This could be relatedto the use of different animal species, dose and/or route oftreatments. Interestingly, however, when we employed similarchromatography solvents as specified by these investigators,our major adduct L3 in the lung DNA of SM rats migrated toa similar location of the chromatogram (not shown) as theirmajor adduct, namely, adduct 1.
Aging has been shown to induce structural changes in DNA(reviewed in Refs. 27, 28), accumulation of chromosomal mutations (28, 29), and generation of DNA adducts (30-32). Lungcancer is primarily a disease of aging, and it is conceivable thatcigarette smoke enhances the age-related DNA damage, therebypredisposing smokers to cancer. It has been suggested that theeffects of chronic smoking may represent, to some degree, anacceleration of the aging process (33). Our data is consistentwith such a hypothesis.
ACKNOWLEDGMENTS
We thank Drs. J. A. Miller and L. I. Rothblum for critical commentson the manuscript, and Karen Earley and Judy Glass for expert technical assistance. Gift of adducted DNA references from Drs. F. F.Kadlubar, F. A. Beland, and C. C. Harris is appreciated.
REFERENCES
1. Fielding, J. E. Smoking-health effects and control (Pt 1). New Engl. J. Med.5/3:491-498, 1985.
2. Fielding, J. E. Smoking-health effects and control (Pt 2). New Engl. J. Med.3/3:555-561, 1985.
3. Wehner, A. P., Dagle, G. E., Milliman, E. M., Phelps, D. W., Carr, D. B.,Decker, J. R., and Filipy, R. E. Inhalation bioassay of cigarette smoke inrats. Toxicol. Appi. Pharmol., 61: 1-17, 1981.
4. Dalbey, W. E., Nettesheim, P., Griesemer, R., Catón,J. E., and Guerin, M.R. Chronic inhalation of cigarette smoke by F344 rats. J. Nati. Cancer Inst.,64: 383-390, 1980.
5. Miller, E. C. Some current perspective on chemical carcinogenesis in humansand experimental animals. Presidential address. Cancer Res., 38:1479-1496,1978.
6. Stedman, R. L. The chemical composition of tobacco and tobacco smoke.Chemical Rev., 68: 153-207, 1968.
7. Schmeltz, I., and Hoffmann, D. Nitrogen-containing compounds in tobaccoand tobacco smoke. Chemical Rev., 77: 295-311, 1977.
8. ( .airóla. C. Genetic effects of fresh cigarette smoke in Saccharomyces cere-risiae. Mutât.Res., 102: 123-136, 1982.
9. Demarini, D. M. Genotoxicity of tobacco smoke and tobacco smoke condensate. Mutât.Res., 114: 59-89, 1983.
10. Griffith, R. B., and Hancock, R. Simultaneous mainstream-sidestream smokeexposure systems I. Equipment and procedures. Toxicology, 34: 123-138,1985.
11. Griffith, R. B., and Standafer, S. Simultaneous mainstream-sidestreamsmoke exposure systems II. The rat exposure system. Toxicology 35: 13-24,1985.
12. Gairola, C. Pulmonary aryl hydrocarbon hydroxylase activity of mice, rats,and guinea pigs following long-term exposure to mainstream and sidestreamcigarette smoke. Toxicology, 45: 177-184, 1987.
13. Gupta, R. C. Nonrandom binding of the carcinogen iV-hydroxy-2-acetylamino-fluorene to repetitive sequences of rat liver DNA in vivo. Proc. Nati.Acad. Sci. USA, 89:6943-6947, 1984.
14. Gupta, R. C., Reddy, M. V., and Randerath, K. "P-postlabeling analysis ofnon-radioactive aromatic carcinogen-DNA adducts. Carcinogenesis (Lond.),3:1081-1092, 1982.
15. Gupta, R. C. Enhanced sensitivity of 32P-postlabeling analysis of aromaticcarcinogen-DNA adducts. Cancer Res., 45: 5656-5662, 1985.
16. Reddy, M. V., and Randerath, K. Nuclease P|-mediated enhancement ofsensitivity of 3!P-postlabeling test for structurally diverse DNA adducts.Carcinogenesis (Lond.), 7: 1543-1551, 1986.
17. Gupta, R. C., and Randerath, K. Analysis of DNA adducts by 33P-labeling
and thin layer chromatography. In: E. C. Friedberg and P. C. Hanawalt
1919
Research. on November 26, 2018. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from

CIGARETTE SMOKE-INDUCED DNA ADDUCTS IN RAT
(eds.), DNA Repair, Vol. 3, pp. 401-420. New York: Marcel-Dekker, Inc.,1988.
18. Binns, R. Inhalation toxicity studies on cigarette smoke IV. Expression ofthe dose of smoke paniculate material to the lungs of experimental animals.Toxicology, 7: 189-195, 1977.
19. Sjoberg, K. PTC in practice. Aldrichimica Acta, 13: 55-58, 1980.20. Belinsky, S. A., Walker, V. E., Maronpot, R. R., Swenberg, J. A., and
Anderson, M. W. Molecular dosimetry of DNA adduci formation and celltoxicity in rat nasal mucosa following exposure to the tobacco specificnitrosamine 4-(Ar-methyl-iV-nitrosamino)-l-(3-puridyl)-l-butanone and theirrelationship to induction of neoplasia. Cancer Res., 47:6058-6065, 1987.
21. Hoffman, D., and Hecht, S. S. Nicotine-derived JV-nitrosamines and tobaccorelated cancer: current status and future directions. Cancer Res., 45: 935-944, 1985.
22. Grafstrom, R. C, Curren, R. D., Yang, L. L., and Harris, C. C. Genotoxicityof formaldehyde in cultured human bronchial fibroblasts. Science (Wash.DC), 228: 89-91, 1985.
23. Nakayama, T., Kaneko, M., Kodama, M., and Nagata, C. Cigarette smokeinduces DNA single-strand breaks in human cells. Nature (Lund.). 314:462-464, 1985.
24. Borish, E. T., Cosgrove, J. P., Church, D. F., Deutsch, W. A., and Pryor, W.A. Cigarette tar causes single-strand breaks in DNA. Biochem. Biophys. Res.Commun., 133: 780-786, 1985.
25. Everson, R. B., Randerath, E., Santella, R. M., Cefalo, R. C., Avitts, T., andRanderath, K. Detection of smoking-related covalent DNA adducts in humanplacenta. Science (Wash. DC), 231: 54-57, 1986.
26. Randerath, E., Avitts. T. A., Reddy, M. V., Miller, R. H., Everson, R. B.,and Randerath, K. Comparative 32P-analysis of cigarette smoke-induced
DNA damage in human tissues and mouse skin. Cancer Res., 46: 5869-5877, 1986.
27. Scmookler-Reiss, R. J., and Goldstein, S. Genetic diversification and theclonality of senescence. In: M. Rothstein (ed.), Review of Biological Researchin Aging, Vol. 2, pp. 115-139. New York: Alan R. Liss, 1985.
28. Schneider, E. L. Cytogenetics of aging. In: C. E. Finch and E. L. Schneider(eds.), Handbook of the Biology of Aging, pp. 357-373. New York: VanNostrand Reinhold, 1985.
29. Martin, G. M., Smith, A. C., Ketterer, D. J., Ogburn, C. E., and Disteche,C. M. Increased chromosomal aberrations in first metaphases of cells isolatedfrom the kidneys of aged mice. Israel J. Med. Sci., 21: 296-301, 1985.
30. Randerath, K., Reddy, M. V., and Disher, R. M. Age- and tissue-relatedDNA modifications in untreated rats: detection by 32P-postlabeling assay andpossible significance for spontaneous tumor induction and aging, (arduogenesis (Lond.), 7: 1615-1617, 1986.
31. Gupta, R. C., Earley, K., Locker, J., and Lombard!, B. 32P-postlabelinganalysis of liver DNA adducts in rats chronically fed a choline-devoid diet.Carcinogenesis (Lond.), 8: 187-189, 1987.
32. Garg, A., and Gupta, R. C. Tissue-specific DNA modifications in untreated,aged rats as detected by 32P-adduct assay. Proc. Am. Assoc. Cancer Res., 29:
104, 1988.33. Holt, P. G., and Keast, D. Environmentally induced change in immunological
function: acute and chronic effects of inhalation of tobacco smoke and otheratmospheric contaminant in man and experimental animals. Bacterio!. Rev.,41: 205-216, 1977.
34. Gupta, R. C., Sharma, S., Earley, K., Mohapatra, N., Nesnow, S., andChacko, M. 32P-adduct assay: principle and application to carcinogen-exposed animal and human DNA. In: S. S. Sandhu, D. M. DeMarini, M. S.Mars, M. M. Moore, and J. L. Mumford (eds.). Short-term Bioassays in theAnalysis of Complex Environmental Mixtures V, Vol. 36, pp. 127-142. NewYork: Plenum, 1988.
1920
Research. on November 26, 2018. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from

1989;49:1916-1920. Cancer Res Ramesh C. Gupta, Mohan L. Sopori and C. G. Gairola Lung and Nasal MucosaFormation of Cigarette Smoke-induced DNA Adducts in the Rat
Updated version
http://cancerres.aacrjournals.org/content/49/8/1916
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/49/8/1916To request permission to re-use all or part of this article, use this link
Research. on November 26, 2018. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from





























