Embed Size (px)
Citation preview

Climate and local geomorphic interactions drive patterns of riparian forest declinealong a Mediterranean Basin river
John C. Stella a,⁎, Jess Riddle a, Hervé Piégay b, Matthieu Gagnage b, Marie-Laure Trémélo b
a Forest and Natural Resources Management, State University of New York College of Environmental Science and Forestry, One Forestry Drive, Syracuse, NY, 13210, USAb University of Lyon, CNRS, ENS Lyon, ISIG Platform, EVS, F-69007, France
a b s t r a c ta r t i c l e i n f o
Article history:Accepted 11 January 2013Available online xxxx
Keywords:Riparian forest declineMediterraneanDendrochronologyPopulus nigra (Salicaceae)In-channel gravel miningClimate change
Dynamic fluvial processes strongly influence ecological communities and ecosystem health in riverine and riparianecosystems, particularly in drought-prone regions. In these systems, there is a need to develop tools tomeasure im-pacts from local and regional hydrogeomorphic changes on the key biological and physical processes that sustainriparian ecosystem health and potential recovery. We used dendrochronology of Populus nigra, a riparian tree thatis vulnerable to changes in local hydrology, to analyze ecosystem response following channel incision due to gravelmining along the Drôme River, aMediterranean Basin stream in southern France.We cored 55 trees at seven flood-plain sites, measured ring widths, and calculated basal area growth to compare the severity and timing of localgrowth decline along the river. Current basal area increment (BAI) growth per tree ranged almost 10-fold amongsites (7.7±1.3 to 63.9±15.2 cm2 year!1, mean±SE) and these differences were significant. Mean BAI was corre-lated positively with the proportion of healthy trees at a site, and negatively with proportion of dead canopy area.Regime shift analysis of the tree-ring series indicates that tree growth declined significantly at four sites since1978, coincidentwith documented channel incision. In addition, patterns of low growth and crowndieback are con-sistent with stress due to reduced water supply. The most impaired sites were not directly adjacent to local miningpits visible on aerial photographs, nor did the sequence of growth regime shifts suggest a pattern of channel incisionprogressing from these areas. The initiation of site growthdeclineswasmost typically associatedwith drought years,and the most impaired sites were spatially distributed to suggest the influence of local bedrock controls on soildepth. Climate in the Drôme basin and in theMediterranean region is trending significantly toward hotter growingseasons with a decrease in summer river discharge, and this will increase both chronic and acutewater shortage forriparian trees. This study shows that drought-prone riparian forests are vulnerable to hydrogeomorphologicalchanges, but the severity of impacts is conditioned by interactions between drivers at different scales, including re-gional climate variability, reach-based geomorphic alteration, and local lithological controls.
© 2013 Elsevier B.V. All rights reserved.
1. Introduction
Dynamic fluvial processes influence riparian forest and woodlandecosystems in numerous ways that affect the initial habitat condi-tions, resource fluxes, and ecological succession pathways. Impor-tant hydrogeomorphic processes include: (1) creating new growingspace through bank erosion, horizontal channel migration, channelcutoff, and accretion of new floodplain surfaces (Piégay et al., 2005;Florsheim et al., 2008; Stella et al., 2011); (2) influencing the recruit-ment and survival of riparian seedlings through scour, burial anddesiccation, all of which depend on the flooding regime and flood-plain morphodynamics (Mahoney and Rood, 1998; Dufour et al.,
2007; Moggridge and Gurnell, 2009); and (3) influencing groundwa-ter table dynamics via channel bed incision and aggradation, whichin general reduces or increases, respectively, water availability tothe root zone (Scott et al., 2000; Pont et al., 2009).
In drought-prone regions, these hydrogeomorphic processes createriparian ecosystems that are particularly resource-rich, productive,structurally complex, and biodiverse compared with the surroundinglandscape (Patten, 1998; Gasith and Resh, 1999; Underwood et al.,2009; Stella et al., 2012). Phreatophytic poplars and cottonwoods(Populus spp.), which dominate sub-humid riparian communities inmuch of the northern hemisphere (Karrenberg et al., 2002), are impor-tant foundational species (sensu Ellison et al., 2005) in providing orregulating water quality, microclimate, structural habitat for wildlifeand fish, an energy base for the food web, and bank stability (Braatneet al., 1996; Lewis et al., 2009; Young-Mathews et al., 2010).
The distribution, composition and health status of Populus-dominatedriparian forests are directly influencedby current andhistorical hydrology(Braatne et al., 1996; Rood et al., 2003) and geomorphic conditions(e.g., Cooper et al., 2003a; Stella et al., 2011). Over the last century,
Geomorphology xxx (2013) xxx–xxx
⁎ Corresponding author at: Department of Forest and Natural Resources Management,State University of New York College of Environmental Science and Forestry, One ForestryDrive, Syracuse, NY 13210, USA. Tel.: +1 315 380 9759; fax: +1 315 470 6535.
E-mail addresses: [email protected] (J.C. Stella), [email protected] (J. Riddle),[email protected] (H. Piégay), [email protected] (M. Gagnage),[email protected] (M.-L. Trémélo).
GEOMOR-04238; No of Pages 14
0169-555X/$ – see front matter © 2013 Elsevier B.V. All rights reserved.http://dx.doi.org/10.1016/j.geomorph.2013.01.013
Contents lists available at SciVerse ScienceDirect
Geomorphology
j ourna l homepage: www.e lsev ie r .com/ locate /geomorph
Please cite this article as: Stella, J.C., et al., Climate and local geomorphic interactions drive patterns of riparian forest decline along aMediterraneanBasin river, Geomorphology (2013), http://dx.doi.org/10.1016/j.geomorph.2013.01.013

humans have modified the dynamic processes that sustain riparianecosystems in multiple ways, including construction of dams andflow diversions, groundwater pumping, and in-stream gravel mining(Steiger et al., 2005; Palmer et al., 2009; Stromberg et al., 2010). As aresult, pioneer riparian forest stands have proven vulnerable to thesedisturbances and are in decline in many ecosystems (Scott et al., 1999;Cooper et al., 2003b; Dufour et al., 2007). Temporary reductions inwater availability can result in reversible changes such as slower growthand increased water use efficiency (Horton et al., 2001; Amlin and Rood,2003; Rood et al., 2003; Lambs et al., 2006; Stella and Battles, 2010), butmore severe and abrupt water table declines can cause permanent dam-age and high mortality (Tyree et al., 1994; Scott et al., 1999; Rood et al.,2000; Stella et al., 2010). Plant water availability may be limited notonly bymodifyingwater supply, canopy demand and vapor pressure def-icits (e.g., Cooper et al., 2003b), but also by altering geomorphologicalprocesses that affect groundwater depth and capillarity (e.g., Scott et al.,1999, 2000).
Negative impacts to riparian tree survival, growth and function arelikely to continue as increasing human populations compete for water.Climate change will further alter the water balance in many areas(Steiger et al., 2005; Palmer et al., 2008; Perry et al., 2011). Because ofits phreatophytic physiology and low tolerance to drought, suddenand severe damage to Populus stands often occurs when the watertable declines beyond a threshold (Scott et al., 1999; Cooper et al.,2003b). Furthermore, local geomorphic changes in conjunction withwater competition and climate change could act synergistically to injurePopulus forests in areas where their separate impacts would be minor(Albertson and Weaver, 1945; Stromberg et al., 1996). The spatiallyand temporally discontinuous pattern of riparian forest decline aftersome channel incision events suggests that local geomorphicmanipula-tion can moderate the impacts on phreatophytic floodplain trees (Graf,1982; Scott et al., 1999; Amlin and Rood, 2003). However, few investiga-tions of riparian forest decline have examined the simultaneous effects ofhydrogeomorphic alteration, climate change, and local geomorphic fac-tors (Palmer et al., 2009).
In this study, we investigated patterns of riparian tree growth overtime, both for individuals and forwhole stands, in the context of local set-ting, reach-wide geomorphic changes and climate change. The DrômeRiver in southeastern France provides an excellent opportunity to studythese interactions at varying temporal and spatial scales. The Mediterra-nean region, including the Drôme basin, is undergoing climatic dryingthat is projected to increase in the 21st Century (Giorgi and Lionello,2008; García-Ruiz et al., 2011). Along a 5-km reach downstream of thetown of Luc-en-Diois, the river is situated in a narrow valley confinedby marl and limestone deposits where the active channel was mined forgravel beginning in the 1970s and subsequently has incised along itslength (Landon et al., 1998; Kondolf et al., 2002; Toone et al., 2012). Adja-cent to the channel, riparian forest stands dominatedby Populus nigra cur-rently display patchy, local evidence of severe crown dieback (Gagnage,2008; Dunford et al., 2009). This species is the dominant European poplarspecies and, because of its high ecological and economic value, has beenthe focus of international efforts to conserve its habitat and genetic diver-sity (Lefèvre et al., 1998; Hughes and Rood, 2003).
In this context we asked the general question, ‘How are patternsin tree growth and decline consistent with overall climate drivers versuslocal environmental conditions such as valley morphology and human-induced geomorphic changes?’ Specifically, we investigated: (1) relation-ships between tree growth and independent measures of riparian standhealth, and how these vary among sites within the reach; (2) trendsin the spatial extent and timing of P. nigra growth decline within thereach; and (3) regional climate-based versus local site-based environ-mental correlates with growth patterns evident across the tree-ringchronologies. We expected that if gravel mining was the immediatecause of tree mortality, crown dieback and reduced growth, two pre-dicted patterns in the tree-ring record should be evident. First, anygrowth declines should have occurred relatively suddenly and only
after the initiation of gravel mining (approximately 1975–1980 fromaerial photograph evidence; Landon et al., 1998). Secondly, floodplainsites closest to the pits should show the earliest and strongest declines,with more distal sites demonstrating later and/or more subtle declines.These spatio-temporal patterns are consistent with other studies ofwater table decline associated with in-channel mining (Scott et al.,1999; Amlin and Rood, 2003; Rollet et al., 2006) and channel bed inci-sion from sediment starvation after dam construction (Kondolf, 1997).In addition, the presence of both confining slopes on one side of theriver channel and tributary confluences upstream of the reachmay con-tribute to edaphic heterogeneity in the floodplain, with potentially localeffects on riparian forest stands. Conversely, if tree growth along theDrôme River responds primarily to climate rather than local drivers,growth patterns should show amore regional signal that is coordinatedwith either short-term droughts (Singer et al., 2012), or long-term cli-mate change in the Mediterranean basin (Giorgi and Lionello, 2008;García-Ruiz et al., 2011).
2. Materials and methods
2.1. Regional setting and study sites
The Drôme, which is a tributary of the Rhône River in southeast-ern France (Fig. 1), is a fourth-order stream with a mountainous pre-Alpine catchment of 1640 km2 of Cretaceous age (elevation range800–1400 m), that is underlain primarily by marl deposits in its valleywith steep slopes rising to limestone ridges. Throughout the water-shed, the climate follows a weakly Mediterranean pattern of warm,relatively dry summers and cool, moister winters. The study area re-ceives 889 mm average annual precipitation (gauged at Luc-en-Diois;Fig. 1); at lower elevations in the watershed, winter precipitation fallsas rain, but snow accumulates at higher elevations.
The catchment area for the study reach is approximately 225 km2
(194 km2 at the Luc-en-Diois gaging station) and annual discharge aver-ages 2.4 m3 s!1 (1961–2007 for Luc-en-Diois). Mean discharge in spring(March–May) is 3.8 (±1.7 SD) m3 s!1 and exceeds the summerbaseflow by an order of magnitude (0.6±0.4 SD m3 s!1 for July–Sept).Between 1948 and 2009, major floods occurred in 1951 (Q16), 1978(Q17), 1994 (Q100) and 2003 (Q45). Major growing-season droughts,with multiple years of lower-than-average streamflow, occurred in1964–65, 1989–91, and 2003–2006.
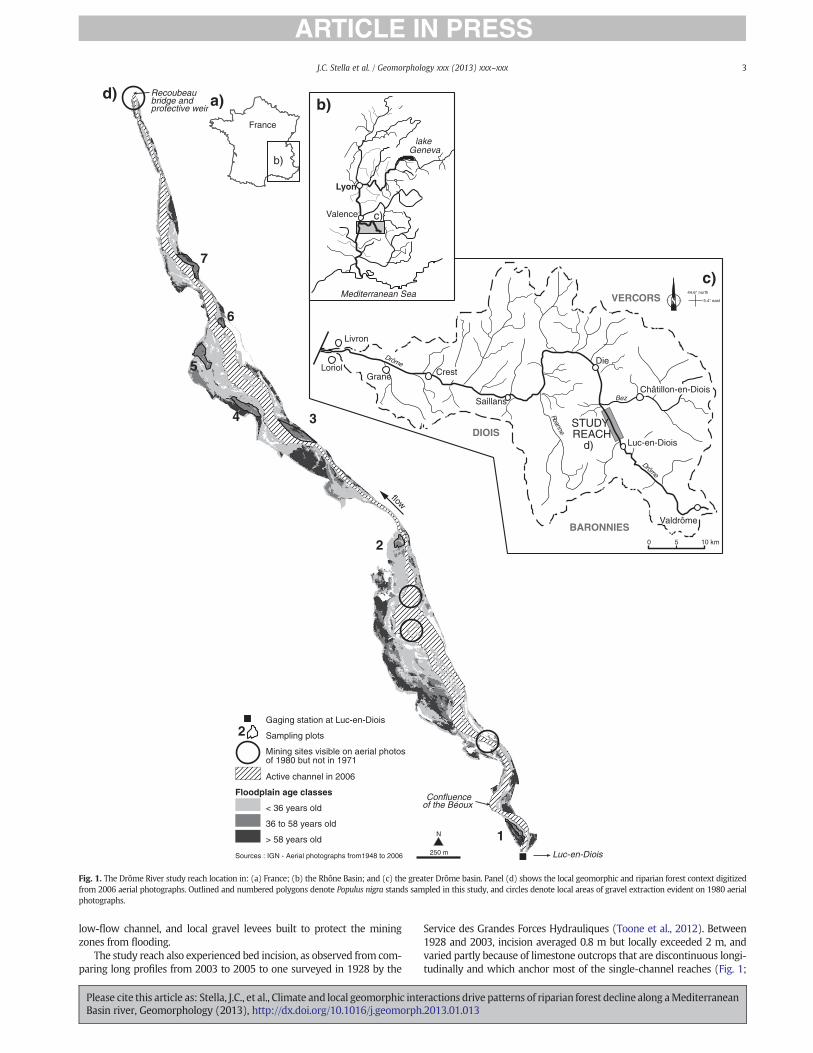
The study was undertaken within a 5-km study reach downstream ofthe town of Luc-en_Diois (Fig. 1). The channel in this reach is character-ized by alternating straight and braided sections, an average longitudinalslope of 0.1%, an active width ranging from 10 to >200 m, and no signif-icant bank protection (Toone et al., 2012). As a result, unconstrainedchannel migration occurs over the floodplain, which supports an exten-sive riparian forest that developed since the 1950s and is currently dom-inated by poplar (P. nigra), willow (Salix alba), ash (Fraxinus excelsior),and alder (Alnus glutinosa) (Piégay and Landon, 1997; Gagnage, 2008).The channel still experiences significant flood events (Landon et al.,1998; Kondolf et al., 2002), including one in December 2003 thatresulted in failure of the bridge at Die.
The Drôme channel and floodplain were subjected to intensive gravelmining beginning in the 1970s. Gravel extraction through channel dredg-ing and strip mining occurred at a rate of 250,000 m3 yr!1 within anddownstream of the study reach during this period, dwarfing the Drôme'snatural annual bedload flux, estimated at 35,000 m3 yr!1 (Landon et al.,1998). Though the most concentrated activity was downstream of the5-km study reach, several areas of intensive extraction within the studyreach are evident on 1980 aerial photographs, but not on the previous se-ries flown in 1971 nor the subsequent one in 1991 (Fig. 1; Toone et al.,2012). This activity was centered downstream of the Béoux confluenceand downstream of the Recoubeau bridge. Geomorphic evidence ofin-streammining includes pits in the gravel bars, sections of straightened
2 J.C. Stella et al. / Geomorphology xxx (2013) xxx–xxx
Please cite this article as: Stella, J.C., et al., Climate and local geomorphic interactions drive patterns of riparian forest decline along aMediterraneanBasin river, Geomorphology (2013), http://dx.doi.org/10.1016/j.geomorph.2013.01.013
low-flow channel, and local gravel levees built to protect the miningzones from flooding.
The study reach also experienced bed incision, as observed from com-paring long profiles from 2003 to 2005 to one surveyed in 1928 by the
Service des Grandes Forces Hydrauliques (Toone et al., 2012). Between1928 and 2003, incision averaged 0.8 m but locally exceeded 2 m, andvaried partly because of limestone outcrops that are discontinuous longi-tudinally and which anchor most of the single-channel reaches (Fig. 1;
1
2
34
5
6
7
250 m
> 58 years old
Sources : IGN - Aerial photographs from1948 to 2006
36 to 58 years old
< 36 years old
Floodplain age classes
d)
Active channel in 2006
Mining sites visible on aerial photos of 1980 but not in 1971
Sampling plots
Gaging station at Luc-en-Diois
N
2
wofl
Recoubeaubridge and protective weir
Luc-en-Diois
Confluenceof the Béoux
44.6° north
5.4° east
STUDY REACH
d)
Livron
LoriolGrane
Saillans
Die
Châtillon-en-Diois
Valdrôme
Luc-en-Diois
emôrD
Crest
DIOIS
VERCORS
BARONNIES
enna
oR
Bez
emôrD
0 5 10 km
c)
b)
Lyon
Valence
lakeGeneva
Mediterranean Sea
c)
a)
b)
France
Fig. 1. The Drôme River study reach location in: (a) France; (b) the Rhône Basin; and (c) the greater Drôme basin. Panel (d) shows the local geomorphic and riparian forest context digitizedfrom 2006 aerial photographs. Outlined and numbered polygons denote Populus nigra stands sampled in this study, and circles denote local areas of gravel extraction evident on 1980 aerialphotographs.
3J.C. Stella et al. / Geomorphology xxx (2013) xxx–xxx
Please cite this article as: Stella, J.C., et al., Climate and local geomorphic interactions drive patterns of riparian forest decline along aMediterraneanBasin river, Geomorphology (2013), http://dx.doi.org/10.1016/j.geomorph.2013.01.013

Toone et al., 2012). Though the period 1928–2003 is too long to assesswhether the recent gravel mining initiated channel bed incision, someof the most incised areas are associated with local mining sites, includingthe Recoubeau bridge, which was undermined during the mining periodand subsequently protected by a concrete weir. Since themid-1970s, thisarea experienced both channel incision that propagated upstream, andlocal aggradation (+0.33 cm between 1928 and 2003) near the weirafter its installation in the early 1990s (Toone et al., 2012).
2.2. Field sampling
We sampled trees in seven floodplain stands located throughout thestudy reach (Fig. 1). These stands were selected by first examining achronosequence of aerial photograph series to locate floodplain sur-faces that had been created prior to 1971 but were still young enoughto be dominated by P. nigra and other pioneer riparian trees. Conifer-dominated patches, which indicated xeric, high-elevation areas outsidethe riparian zone, were filtered out using anNDVI index on recent aerialphotographs (Quickbird image flown 7 Dec. 2001). This process identi-fied 11 mid-aged sites that ranged from 36 to 58 years old. From thisgroup we selected seven sites for field sampling that were relativelywell-distributed within the reach and that supported deciduous riparianforest stands dominated by P. nigra (Fig. 1; Gagnage, 2008; Lejot et al.,2011).
In each stand we surveyed the relative elevation of its floodplain sur-face above the current baseflow elevation using DGPS (Trimble 5800 Dif-ferential Global Positioning System) with a vertical resolution of b10 cm,at 1–7 points (mean 3.4) per stand. Surveys were conducted fromMay toJuly 2008 on days when the river discharge was b2 m3 s!1. Using a soilcorer, we measured the fine sediment thickness at 6 to 14 locations dis-tributed across the stand.
At the center of each stand, we surveyed a circular plot of 500 m2 tosample tree composition, density and crownhealth. Canopy coverwas es-timated visually for the entire stand using a range of values to representthe high heterogeneity in cover present. All canopy trees within the plotweremeasured and the species compositionwas recorded. From the cen-ter of each plot, we selected 20 dominant trees (i.e., those structuring thecanopy) forwhichwe estimated the proportion of live crown; thesemea-surements were aggregated across the sample to assess the health of thestand (Scott et al., 1999).
The estimates of crown integrity were complemented using low-elevation aerial photograph analysis of the sites to identify the areaof dead branches within the canopy (Gagnage, 2008; Dunford et al.,2009). In this method, digital photographs taken from an unmannedaerial vehicle were orthorectified to the project basemap and deadbranches were identified using the object-oriented classification ap-proach detailed in Dunford et al. (2009). Dead branch area wassummed and divided by the sampling area for each of the stands toestimate the proportion dead canopy.
2.3. Dendrochronological analysis
Of the live trees in each stand, we selected the largest dominantP. nigra individuals for coring, regardless of health status. A total of55 trees were cored across the seven sites; samples ranged 6–9 treesper site. Cores were extracted at breast height (1.4 m) using a 5-mmincrement borer, with two cores collected per tree wherever possible(N=114 cores).
Following standard dendrochronological procedures, we mountedcores and clarified ring anatomyby sanding each corewith a series of pro-gressively finer sand papers (Stokes and Smiley, 1968). Ringwidthsweremeasured to 0.001 mm resolution using a sliding stage micrometer(Velmex Inc., Bloomfield, NY) and Acu-Rite encoder (Heidenhain Corp.,Schaumberg, IL). Potentiallymisdated coreswere identifiedwith the soft-ware program COFECHA (Holmes, 1983) and checked for false rings,missing rings, and other sources of dating errors. All ages reported are
minimumages, as theydonot account for time for the tree to reach coringheight or the time between the innermost ring on the core and the pith.
To examine long term trends in tree growth, we converted the rawring widths to basal area increments (BAI) using the formula
BAI ! ! rn2!rn!1
2! "
"1#
where r is the radius at the end of year n. BAI typically increases as trees'crowns expand or understory trees are exposed to higher light levels,and stabilizes or continues to increase in healthy, undamaged, maturetrees (Biondi and Qeadan, 2008). The BAI series for multiple cores pertree were averaged, and then site chronologies were constructed bytaking the arithmetic mean of all BAI series from the site for each year.
We analyzed the BAI chronologies using Rodionov's regime shift de-tection (RSD; Rodionov, 2004) to explore the possibility of rapid andsustained shifts in the site mean growth rates, and to determine thetiming of any shifts. The algorithmuses sequential t-tests to identify shiftsin the mean without an a priori hypothesis of timing, and uses an indexbased on normalized anomalies to determine if the shifts aremaintained.We applied the algorithm with the RSD add-in for Microsoft Excel 2002(Rodionov, 2006a) with significance level=0.05, cut-off=10 years,Huber's weight parameter=2, and autocorrelation estimated by the in-verse proportionalitymethod on 7 year subsamples (Rodionov, 2006a,b).
To examine correlations with growing season climatic and hydrolog-ical variables, we used a data-adaptive detrending procedure within theprogram ARSTAN (Cook, 1985) to produce a chronology that emphasizeshigh-frequency variation (Cook and Peters, 1981). This series was moreappropriate to use than the BAI series, because lower-frequency signalssuch as decadal growth trends due to site conditions would be preservedin the BAI chronologies and would obscure the high-frequency climatecorrelations (Cook et al., 1990). Nine of the 114 total cores were re-moved from this analysis due to poor cross-dating; in some cases,short series may have compromised the ability of statistical correla-tions to identify misdated cores (Grissino-Mayer, 2001). A Friedmanvariable span smoother (alpha=5) was fit to each remaining series(Friedman, 1984), which was then divided by its fit curve to producea time series of dimensionless index values; the autocorrelation wasthen estimated and removed from each series. All residual series wereaveraged by calendar year using a robust bi-weight mean to generatethe final residual chronology. The chronology was truncated where theexpressed population signal, ameasure of chronology strengthwithmax-imum of 1, fell below 0.85 (Wigley et al., 1984).
We tested differences in recent growth among sites using analysis ofcovariance (ANCOVA). Recent growth, the response variable, was calcu-lated by summing the BAI for themost recent four years (2004–07); thismetricwas log-transformed in the ANCOVAmodel to satisfy residual as-sumptions. Predictor variables in addition to site included tree age andearly tree size, which was represented as basal area in 1980. This lattervariable was included in order to control for differences in tree size atthe start of the growth decline period, when larger trees would beexpected to have amore extensive root system and therefore potential-ly greater access to water. To estimate individual tree basal areas at thetime when growth declines might have begun, we summed for eachcore all ring widths prior to 1980, then averaged the radius estimatesfor all cores within trees, and converted these to basal areas.
2.4. Climate data analysis
Climate data for the period 1961–2007 were obtained from severalsources in order to evaluate the climate signal within the tree-ring chro-nologies. Monthly maximum and minimum temperature data (i.e., dailyhighs and lows averaged by month) were extracted from the E-OBSdataset (Haylock et al., 2008), which are gridded (0.25° cell) interpola-tions from regional station data withmissing data estimated and stationswith >20% days with missing data excluded. Daily precipitation totals
4 J.C. Stella et al. / Geomorphology xxx (2013) xxx–xxx
Please cite this article as: Stella, J.C., et al., Climate and local geomorphic interactions drive patterns of riparian forest decline along aMediterraneanBasin river, Geomorphology (2013), http://dx.doi.org/10.1016/j.geomorph.2013.01.013

were obtained for the Luc-en-Diois station (Météo France station#26167001) and daily river discharge values from the Luc-en-Dioisgauging station (#V4214010, French National Office for Water andthe Aquatic Environment). Monthlymetrics were calculated by averag-ing the daily discharge values and summing the daily precipitation totals.
We used Pearson correlations to assess the relationship between an-nual radial growth, as indicated by the high-frequency chronology, andboth monthly and growing season climate and discharge variables. Be-cause conditions during the year prior to ring formation often influencering width (Fritts, 1976), we used a 19-month window beginning in theprevious April and extending to the current October to assess annualgrowth correlations with monthly values for discharge, precipitation,and minimum and maximum temperatures. The best-correlated monthswere further aggregated as a single period (i.e., growing season) to com-pare the sensitivity of tree-ring growth among climate metrics. Themonthly discharge values had substantially unequal variances, thereforewe transformed all data metrics to standard scores (i.e., z-scores) priorto averaging months together for the growing season. Standardizationwas necessary because the unequal variances could lead to biased grow-ing season discharge estimates whereby only one or two months domi-nated the seasonal signal. The discharge standard scores were calculated
z ! x–μ" #=σ "2#
where μ and σ are the overall mean and standard deviation, respective-ly, for a particular month throughout the period of record and x is themonthly value in a given year. Following transformation, monthly scoreswere then averaged for the growing season, June through September.
We used amodifiedMann–Kendall test proposed by Yue et al. (2002)to estimate the significance of trends in monthly and seasonal climaticand discharge variables over the period 1961–2007. In that procedure,trend is first estimated as the median slope of all possible point pairs(Theil, 1950a–c; Sen, 1968) then removed from the time series. Next, au-tocorrelation is modeled as a lag-one autoregressive process and also re-moved from the time series. Finally, the trend ismixed back into the timeseries, and significance of the trend is assessed by theMann–Kendall test(Mann, 1945; Kendall, 1975). We completed all trend analysis with thezyp package in R (version 2.12.1, R Development Core Team).
Table1
Characteristicsof
thePo
pulusnigrastan
dsstud
iedalon
gtheDrômeRive
r.
Site
Floo
dplain
elev
ationab
ove
chan
nel(
m)
Fine
sedimen
tde
pth(cm,
mea
n±
1SE)
Max
imum
incision
betw
een19
28&20
03long
profi
les(m
)
Cano
pyco
ver
Den
sity
ofcano
pytree
s(trees
ha!
1)
Prop
ortion
oftree
swith>50
%bran
ches
dead
Dea
dcano
pypo
rtionof
total
stan
darea
No.
tree
sco
red
Diameter
ofcoredtrees
(cm,m
ean±1S
E)Age
ofco
redtree
s(yea
rs,m
edian,
min
andmax
)
Basala
reaincrem
ent
ofco
redtree
s(cm
2 ,mea
n±
1SE)
11.55
70.3
(±14
.5)
2.2
25%
740
6%1.4%
847
.1(±
5.4)
58(3
8,71
)47
.4(±
10.4)
22.09
31.9
(±10
.5)
1.8
25%–
50%
1040
0%2.0%
649
.7(±
5.1)
35.5
(31,
37)
63.9
(±15
.2)
31.45
46.5
(±6.9)
1.7
0%–25
%24
065
%2.5%
937
.6(±
4.1)
47(4
4,64
)7.7(±
1.3)
42.79
54.2
(±11
.2)
1.0
80%
1340
11%
2.5%
954
.5(±
5.6)
37(2
4,62
)27
.3(±
6.8)
50.73
54.2
(±10
.9)
0.3
90%
740
50%
0.5%
850
.2(±
4.7)
36.5
(24,
43)
51.3
(±10
.3)
61.87
25.0
(±5.8)
1.2
0%–25
%32
061
%2.4%
744
.0(±
4.3)
48(2
6,61
)12
.5(±
3.7)
71.09
15.5
(±3.7)
1.1
25%
540
26%
1.6%
831
.5(±
2.9)
43.5
(32,
50)
48.5
(±16
.4)
Site Number
Rec
ent B
AI (
cm2
year
-1)
c c a bc c ab bc
1 2 3 4 5 6 7
1
5
10
50
100
150
Fig. 2. Boxplots of recent average growth (basal area increment means for 2004–07)for Populus nigra trees at the seven Drôme River floodplain sites. Thick solid lines indi-cate site means, boxes indicate 25th and 75th quartiles, and whiskers span the range ofthe data. Different letters at the top denote significant differences between sites basedon the Tukey Honestly Significant Difference test and a threshold pb0.05.
5J.C. Stella et al. / Geomorphology xxx (2013) xxx–xxx
Please cite this article as: Stella, J.C., et al., Climate and local geomorphic interactions drive patterns of riparian forest decline along aMediterraneanBasin river, Geomorphology (2013), http://dx.doi.org/10.1016/j.geomorph.2013.01.013
3. Results
3.1. Site conditions, tree health, and recent growth
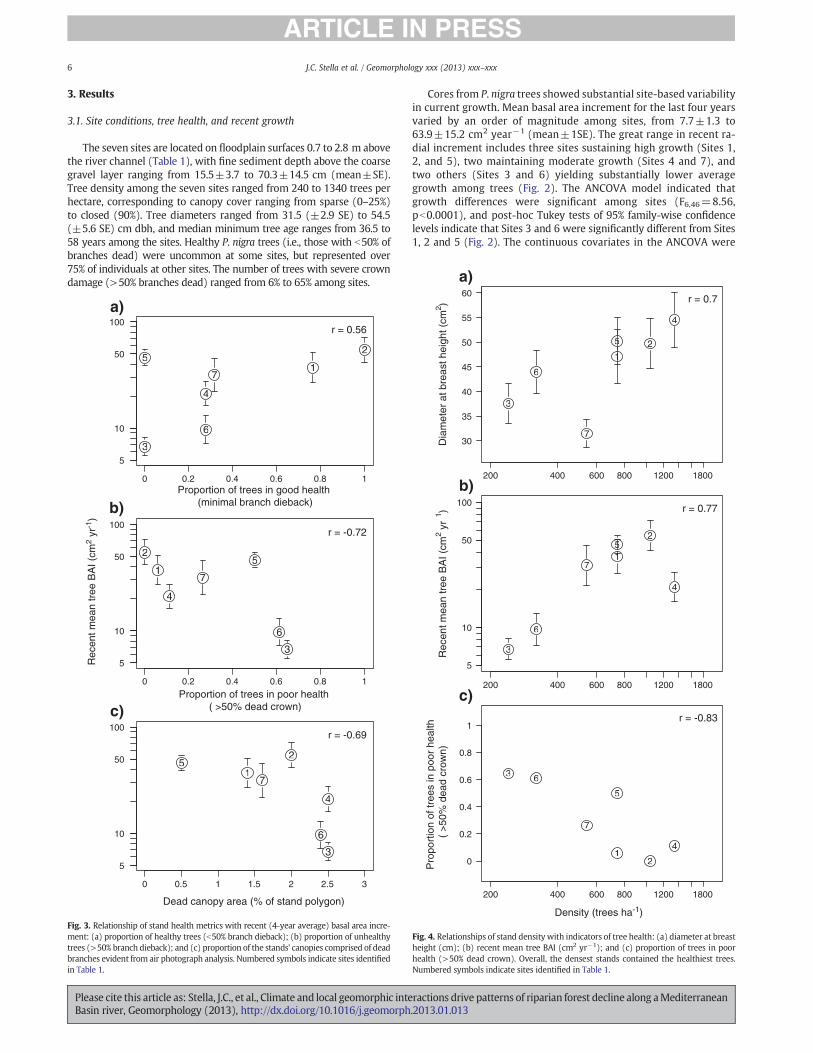
The seven sites are located on floodplain surfaces 0.7 to 2.8 m abovethe river channel (Table 1), with fine sediment depth above the coarsegravel layer ranging from 15.5±3.7 to 70.3±14.5 cm (mean±SE).Tree density among the seven sites ranged from 240 to 1340 trees perhectare, corresponding to canopy cover ranging from sparse (0–25%)to closed (90%). Tree diameters ranged from 31.5 (±2.9 SE) to 54.5(±5.6 SE) cm dbh, and median minimum tree age ranges from 36.5 to58 years among the sites. Healthy P. nigra trees (i.e., those with b50% ofbranches dead) were uncommon at some sites, but represented over75% of individuals at other sites. The number of trees with severe crowndamage (>50% branches dead) ranged from 6% to 65% among sites.
Cores from P. nigra trees showed substantial site-based variabilityin current growth. Mean basal area increment for the last four yearsvaried by an order of magnitude among sites, from 7.7±1.3 to63.9±15.2 cm2 year!1 (mean±1SE). The great range in recent ra-dial increment includes three sites sustaining high growth (Sites 1,2, and 5), two maintaining moderate growth (Sites 4 and 7), andtwo others (Sites 3 and 6) yielding substantially lower averagegrowth among trees (Fig. 2). The ANCOVA model indicated thatgrowth differences were significant among sites (F6,46=8.56,pb0.0001), and post-hoc Tukey tests of 95% family-wise confidencelevels indicate that Sites 3 and 6 were significantly different from Sites1, 2 and 5 (Fig. 2). The continuous covariates in the ANCOVA were
1
2
3
4
5
6
7
Proportion of trees in good health (minimal branch dieback)
1
2
3
4
5
6
7
0 0.2 0.4 0.6 0.8 1
5
10
50
100a)
b)
c)
r = 0.56
1
2
3
4
5
6
7
Proportion of trees in poor health ( >50% dead crown)
1
2
3
4
5
6
7
0 0.2 0.4 0.6 0.8 1
5
10
50
100r = -0.72
1
2
3
4
5
6
7
Dead canopy area (% of stand polygon)
1
2
3
4
5
6
7
0 0.5 1 1.5 2 2.5 3
5
10
50
100r = -0.69
Rec
ent m
ean
tree
BA
I (cm
2 yr
-1)
Fig. 3. Relationship of stand health metrics with recent (4-year average) basal area incre-ment: (a) proportion of healthy trees (b50% branch dieback); (b) proportion of unhealthytrees (>50% branch dieback); and (c) proportion of the stands' canopies comprised of deadbranches evident from air photograph analysis. Numbered symbols indicate sites identifiedin Table 1.
12
3
4
5
6
7D
iam
eter
at b
reas
t hei
ght (
cm2 )
12
3
4
5
6
7
200 400 600 800 1200 1800
30
35
40
45
50
55
60 r = 0.7
1
2
3
4
5
6
7
Rec
ent m
ean
tree
BA
I (cm
2yr
1 )
1
2
3
4
5
6
7
200 400 600 800 1200 1800
5
10
50
100 r = 0.77
12
3
4
5
6
7
Pro
port
ion
of tr
ees
in p
oor
heal
th
( >
50%
dea
d cr
own)
12
3
4
5
6
7
200 400 600 800 1200 1800
0
0.2
0.4
0.6
0.8
1r = -0.83
Density (trees ha-1)
a)
b)
c)
Fig. 4. Relationships of stand density with indicators of tree health: (a) diameter at breastheight (cm); (b) recent mean tree BAI (cm2 yr!1); and (c) proportion of trees in poorhealth (>50% dead crown). Overall, the densest stands contained the healthiest trees.Numbered symbols indicate sites identified in Table 1.
6 J.C. Stella et al. / Geomorphology xxx (2013) xxx–xxx
Please cite this article as: Stella, J.C., et al., Climate and local geomorphic interactions drive patterns of riparian forest decline along aMediterraneanBasin river, Geomorphology (2013), http://dx.doi.org/10.1016/j.geomorph.2013.01.013
not significant, including tree age (F1,46=0.35, p=0.56) and treesize normalized to estimated 1980 basal area (F1,46=2.36, p=0.13); nor were any variable interactions significant. Thereforesite-based growth differences were not influenced by variation in theintrinsic biotic factors of tree age and size early in stand development.
At the site level, recent radial growth was positively correlated withthe proportion of healthy (i.e., minimal branch mortality) P. nigra trees(r=0.56; Fig. 3a), and negatively correlated with the proportion oftrees with >50% dead crown (r=!0.72; Fig. 3b), and the proportionof the stand's aerial image comprising dead branches in the upper can-opy (r=!0.69; Fig. 3c).Metrics of standvigorwere positively correlatedwith tree density, with the sparsest stands supporting the least produc-tive and healthy trees. For example, stand density was positively cor-related with tree diameter (r=0.70; Fig. 4a) and recent growth(r=0.77; Fig. 4b), and negatively correlated with the proportion oftrees with severe crown damage (r=!0.83; Fig. 4c).
Of the geomorphic variables measured, elevation above the channelwas correlated positively with areal proportion dead canopy in thestand (r=0.74), indicating greater damage in higher stands. However,therewas no relationship evident between recent growth and elevationabove the channel (r=!0.16), nor between recent growth and maxi-mum bed incision in the adjacent channel calculated from the 1928and 2003 long profiles (r=!0.09, t=!0.21, df=5, p=0.84).
3.2. Dendrochronological and growth results
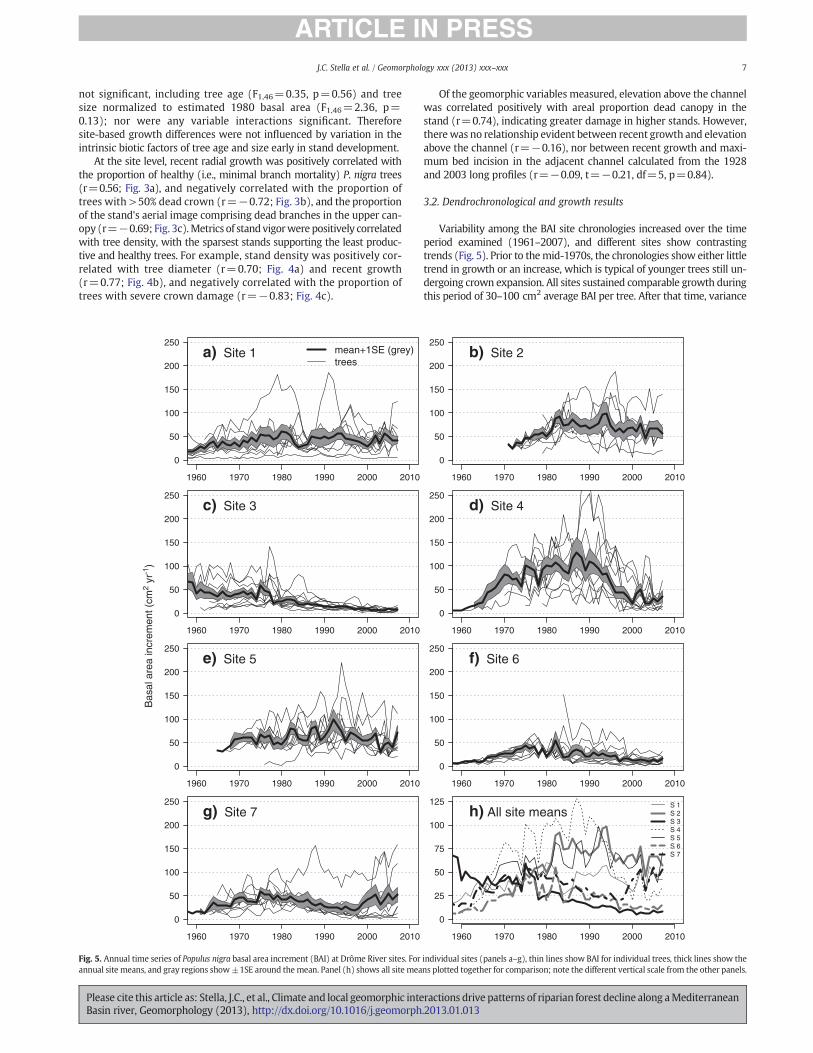
Variability among the BAI site chronologies increased over the timeperiod examined (1961–2007), and different sites show contrastingtrends (Fig. 5). Prior to themid-1970s, the chronologies show either littletrend in growth or an increase, which is typical of younger trees still un-dergoing crown expansion. All sites sustained comparable growth duringthis period of 30–100 cm2 average BAI per tree. After that time, variance
1960 1970 1980 1990 2000 2010
0
50
100
150
200
250a) Site 1 mean+1SE (grey)
trees
1960 1970 1980 1990 2000 2010
0
50
100
150
200
250b) Site 2
1960 1970 1980 1990 2000 2010
0
50
100
150
200
250c) Site 3
1960 1970 1980 1990 2000 2010
0
50
100
150
200
250d) Site 4
1960 1970 1980 1990 2000 2010
0
50
100
150
200
250e) Site 5
1960 1970 1980 1990 2000 2010
0
50
100
150
200
250f) Site 6
1960 1970 1980 1990 2000 2010
0
50
100
150
200
250g) Site 7
S 1S 2S 3S 4S 5S 6S 7
1960 1970 1980 1990 2000 2010
0
25
50
75
100
125h) All site means
Bas
al a
rea
incr
emen
t (cm
2 yr
-1)
Fig. 5. Annual time series of Populus nigra basal area increment (BAI) at Drôme River sites. For individual sites (panels a–g), thin lines show BAI for individual trees, thick lines show theannual site means, and gray regions show±1SE around the mean. Panel (h) shows all site means plotted together for comparison; note the different vertical scale from the other panels.
7J.C. Stella et al. / Geomorphology xxx (2013) xxx–xxx
Please cite this article as: Stella, J.C., et al., Climate and local geomorphic interactions drive patterns of riparian forest decline along aMediterraneanBasin river, Geomorphology (2013), http://dx.doi.org/10.1016/j.geomorph.2013.01.013
among chronologies increased and sites diverged steeply in averagegrowth, with Sites 1, 2, and 5 maintaining 50–100 cm2 BAI per tree tothe present, and the other sites showing growth declines at various times.
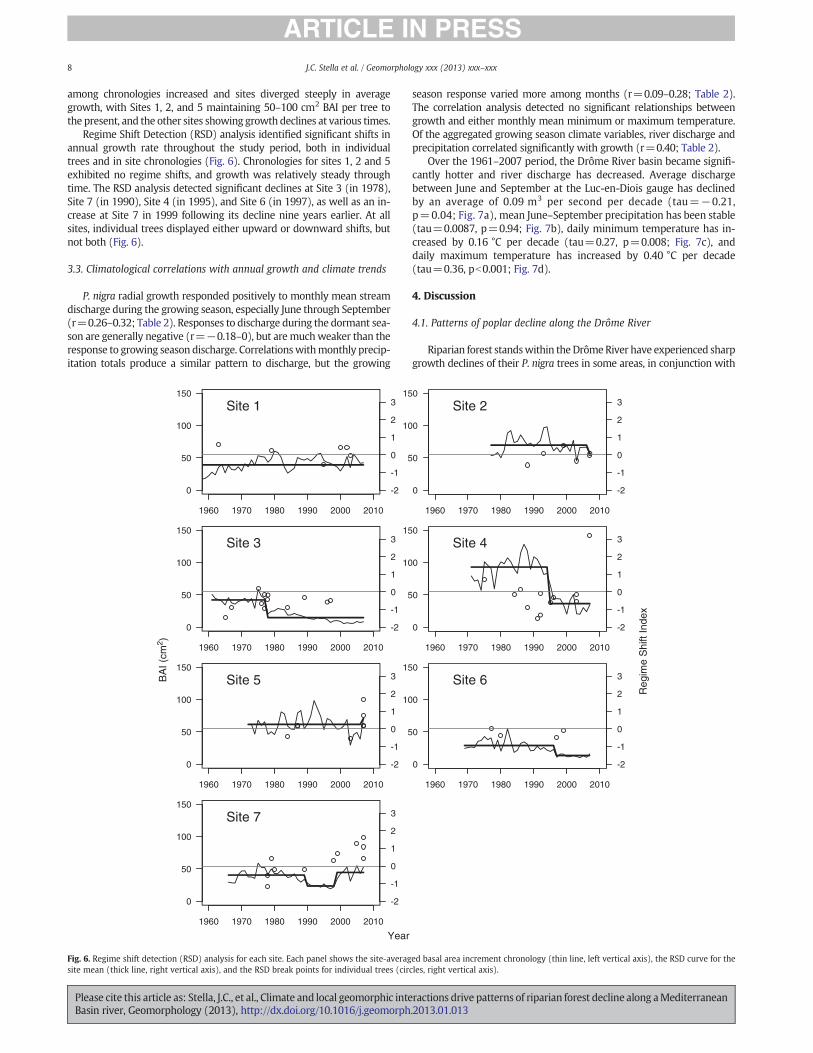
Regime Shift Detection (RSD) analysis identified significant shifts inannual growth rate throughout the study period, both in individualtrees and in site chronologies (Fig. 6). Chronologies for sites 1, 2 and 5exhibited no regime shifts, and growth was relatively steady throughtime. The RSD analysis detected significant declines at Site 3 (in 1978),Site 7 (in 1990), Site 4 (in 1995), and Site 6 (in 1997), as well as an in-crease at Site 7 in 1999 following its decline nine years earlier. At allsites, individual trees displayed either upward or downward shifts, butnot both (Fig. 6).
3.3. Climatological correlations with annual growth and climate trends
P. nigra radial growth responded positively to monthly mean streamdischarge during the growing season, especially June through September(r=0.26–0.32; Table 2). Responses to discharge during the dormant sea-son are generally negative (r=!0.18–0), but aremuchweaker than theresponse to growing season discharge. Correlationswithmonthly precip-itation totals produce a similar pattern to discharge, but the growing
season response varied more among months (r=0.09–0.28; Table 2).The correlation analysis detected no significant relationships betweengrowth and either monthly mean minimum or maximum temperature.Of the aggregated growing season climate variables, river discharge andprecipitation correlated significantly with growth (r=0.40; Table 2).
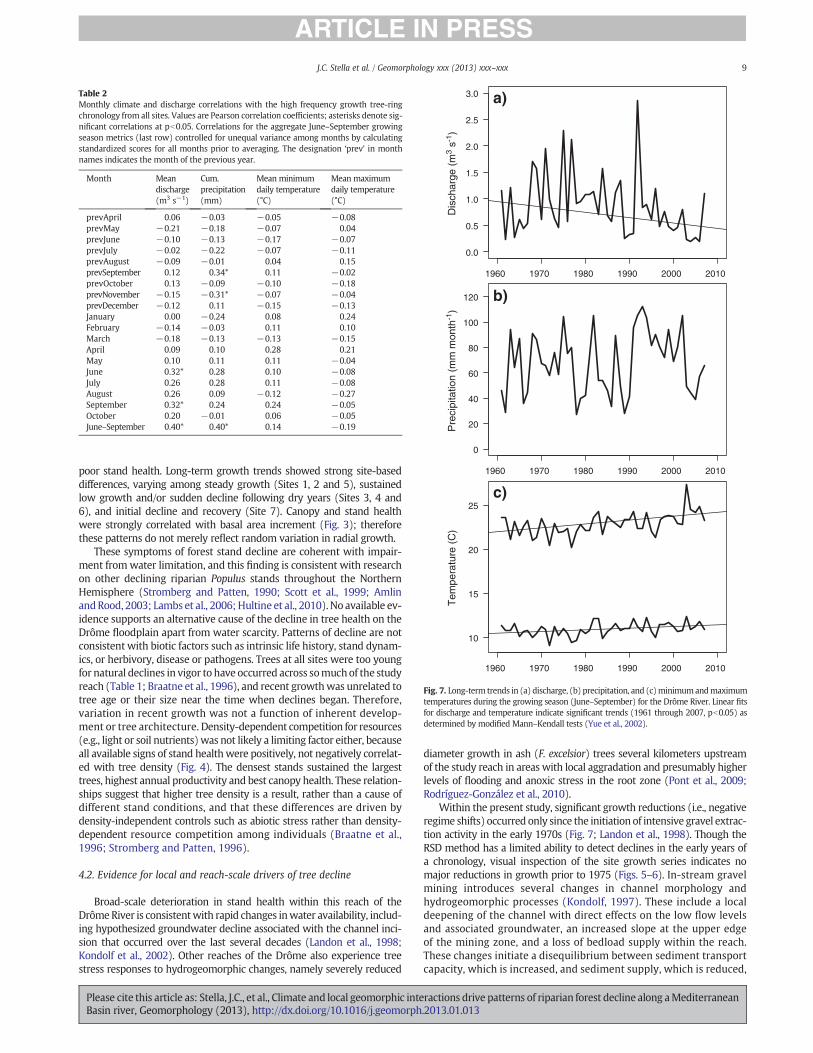
Over the 1961–2007 period, the Drôme River basin became signifi-cantly hotter and river discharge has decreased. Average dischargebetween June and September at the Luc-en-Diois gauge has declinedby an average of 0.09 m3 per second per decade (tau=!0.21,p=0.04; Fig. 7a), mean June–September precipitation has been stable(tau=0.0087, p=0.94; Fig. 7b), daily minimum temperature has in-creased by 0.16 °C per decade (tau=0.27, p=0.008; Fig. 7c), anddaily maximum temperature has increased by 0.40 °C per decade(tau=0.36, pb0.001; Fig. 7d).
4. Discussion
4.1. Patterns of poplar decline along the Drôme River
Riparian forest standswithin theDrômeRiver have experienced sharpgrowth declines of their P. nigra trees in some areas, in conjunction with
1960 1970 1980 1990 2000 2010
0
50
100
150
-2
-1
0
1
2
3Site 1
1960 1970 1980 1990 2000 2010
0
50
100
150
-2
-1
0
1
2
3Site 2
1960 1970 1980 1990 2000 2010
0
50
100
150
-2
-1
0
1
2
3Site 3
1960 1970 1980 1990 2000 2010
0
50
100
150
-2
-1
0
1
2
3Site 4
1960 1970 1980 1990 2000 2010
0
50
100
150
-2
-1
0
1
2
3Site 5
1960 1970 1980 1990 2000 2010
0
50
100
150
-2
-1
0
1
2
3Site 6
1960 1970 1980 1990 2000 2010
0
50
100
150
-2
-1
0
1
2
3Site 7
BA
I (cm
2 )
Year
Reg
ime
Shi
ft In
dex
Fig. 6. Regime shift detection (RSD) analysis for each site. Each panel shows the site-averaged basal area increment chronology (thin line, left vertical axis), the RSD curve for thesite mean (thick line, right vertical axis), and the RSD break points for individual trees (circles, right vertical axis).
8 J.C. Stella et al. / Geomorphology xxx (2013) xxx–xxx
Please cite this article as: Stella, J.C., et al., Climate and local geomorphic interactions drive patterns of riparian forest decline along aMediterraneanBasin river, Geomorphology (2013), http://dx.doi.org/10.1016/j.geomorph.2013.01.013
poor stand health. Long-term growth trends showed strong site-baseddifferences, varying among steady growth (Sites 1, 2 and 5), sustainedlow growth and/or sudden decline following dry years (Sites 3, 4 and6), and initial decline and recovery (Site 7). Canopy and stand healthwere strongly correlated with basal area increment (Fig. 3); thereforethese patterns do not merely reflect random variation in radial growth.
These symptoms of forest stand decline are coherent with impair-ment fromwater limitation, and this finding is consistent with researchon other declining riparian Populus stands throughout the NorthernHemisphere (Stromberg and Patten, 1990; Scott et al., 1999; AmlinandRood, 2003; Lambs et al., 2006; Hultine et al., 2010). No available ev-idence supports an alternative cause of the decline in tree health on theDrôme floodplain apart from water scarcity. Patterns of decline are notconsistent with biotic factors such as intrinsic life history, stand dynam-ics, or herbivory, disease or pathogens. Trees at all sites were too youngfor natural declines in vigor to have occurred across somuchof the studyreach (Table 1; Braatne et al., 1996), and recent growthwas unrelated totree age or their size near the time when declines began. Therefore,variation in recent growth was not a function of inherent develop-ment or tree architecture. Density-dependent competition for resources(e.g., light or soil nutrients)was not likely a limiting factor either, becauseall available signs of stand healthwere positively, not negatively correlat-ed with tree density (Fig. 4). The densest stands sustained the largesttrees, highest annual productivity and best canopy health. These relation-ships suggest that higher tree density is a result, rather than a cause ofdifferent stand conditions, and that these differences are driven bydensity-independent controls such as abiotic stress rather than density-dependent resource competition among individuals (Braatne et al.,1996; Stromberg and Patten, 1996).
4.2. Evidence for local and reach-scale drivers of tree decline
Broad-scale deterioration in stand health within this reach of theDrômeRiver is consistentwith rapid changes inwater availability, includ-ing hypothesized groundwater decline associated with the channel inci-sion that occurred over the last several decades (Landon et al., 1998;Kondolf et al., 2002). Other reaches of the Drôme also experience treestress responses to hydrogeomorphic changes, namely severely reduced
diameter growth in ash (F. excelsior) trees several kilometers upstreamof the study reach in areas with local aggradation and presumably higherlevels of flooding and anoxic stress in the root zone (Pont et al., 2009;Rodríguez-González et al., 2010).
Within the present study, significant growth reductions (i.e., negativeregime shifts) occurred only since the initiation of intensive gravel extrac-tion activity in the early 1970s (Fig. 7; Landon et al., 1998). Though theRSD method has a limited ability to detect declines in the early years ofa chronology, visual inspection of the site growth series indicates nomajor reductions in growth prior to 1975 (Figs. 5–6). In-stream gravelmining introduces several changes in channel morphology andhydrogeomorphic processes (Kondolf, 1997). These include a localdeepening of the channel with direct effects on the low flow levelsand associated groundwater, an increased slope at the upper edgeof the mining zone, and a loss of bedload supply within the reach.These changes initiate a disequilibrium between sediment transportcapacity, which is increased, and sediment supply, which is reduced,
Table 2Monthly climate and discharge correlations with the high frequency growth tree-ringchronology from all sites. Values are Pearson correlation coefficients; asterisks denote sig-nificant correlations at pb0.05. Correlations for the aggregate June–September growingseason metrics (last row) controlled for unequal variance among months by calculatingstandardized scores for all months prior to averaging. The designation ‘prev’ in monthnames indicates the month of the previous year.
Month Meandischarge(m3 s!1)
Cum.precipitation(mm)
Mean minimumdaily temperature(°C)
Mean maximumdaily temperature(°C)
prevApril 0.06 !0.03 !0.05 !0.08prevMay !0.21 !0.18 !0.07 0.04prevJune !0.10 !0.13 !0.17 !0.07prevJuly !0.02 !0.22 !0.07 !0.11prevAugust !0.09 !0.01 0.04 0.15prevSeptember 0.12 0.34* 0.11 !0.02prevOctober 0.13 !0.09 !0.10 !0.18prevNovember !0.15 !0.31* !0.07 !0.04prevDecember !0.12 0.11 !0.15 !0.13January 0.00 !0.24 0.08 0.24February !0.14 !0.03 0.11 0.10March !0.18 !0.13 !0.13 !0.15April 0.09 0.10 0.28 0.21May 0.10 0.11 0.11 !0.04June 0.32* 0.28 0.10 !0.08July 0.26 0.28 0.11 !0.08August 0.26 0.09 !0.12 !0.27September 0.32* 0.24 0.24 !0.05October 0.20 !0.01 0.06 !0.05June–September 0.40* 0.40* 0.14 !0.19
1960 1970 1980 1990 2000 2010
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Dis
char
ge (
m3
s-1)
a)
1960 1970 1980 1990 2000 2010
0
20
40
60
80
100
120
Pre
cipi
tatio
n (m
m m
onth
-1)
b)
1960 1970 1980 1990 2000 2010
10
15
20
25
Tem
pera
ture
(C
)
c)
Fig. 7. Long-term trends in (a) discharge, (b) precipitation, and (c)minimumandmaximumtemperatures during the growing season (June–September) for the Drôme River. Linear fitsfor discharge and temperature indicate significant trends (1961 through 2007, pb0.05) asdetermined by modified Mann–Kendall tests (Yue et al., 2002).
9J.C. Stella et al. / Geomorphology xxx (2013) xxx–xxx
Please cite this article as: Stella, J.C., et al., Climate and local geomorphic interactions drive patterns of riparian forest decline along aMediterraneanBasin river, Geomorphology (2013), http://dx.doi.org/10.1016/j.geomorph.2013.01.013
and typically results in local channel bed incision that propagatesdownstream of the mining site, and a regressive erosion upstreamthat eventually stabilizes the local channel slope. Cascading effectsof channel incision on floodplain habitats due to mining has beenalso showed by Rollet et al. (2006) on the Doubs River, in which in-cision propagated both upstream and downstream of mining sites.Channel bed incision also tends to propagate downstream locallyfrom dams for similar reasons (Kondolf, 1997).
Though channel incision and associated groundwater decline wouldbe expected to have system-wide effects (i.e., affect all sites), they arealso expected to generate coherent spatial and temporal patterns in therelative response of individual sites. For example,we expectedfloodplainstands adjacent to gravel extraction areas on the Drôme to decline moststrongly and earlier as they experience the effective deepening. Scott etal. (1999) documented that widespread cottonwood mortality andcrown damage occurred within two years of intensive sand mining inthe adjacent channel, and that effects of depressed groundwater andtree decline were most severe closest to the excavation site. Amlin andRood (2003) observed extensive leaf senescence, abscission, and re-duced stomatal conductance in a stand of cottonwoods adjacent to anexcavated gravel pit that depressed the water table by 2.5 m, but sawno effect in a reference stand farther away. On the Drôme, we expectednot only this proximity effect, but also that the sequence of impactswould progress from the near-pit sites to farther ones upstream anddownstream over timewith propagation of channel bed incision and as-sociate water table lowering.
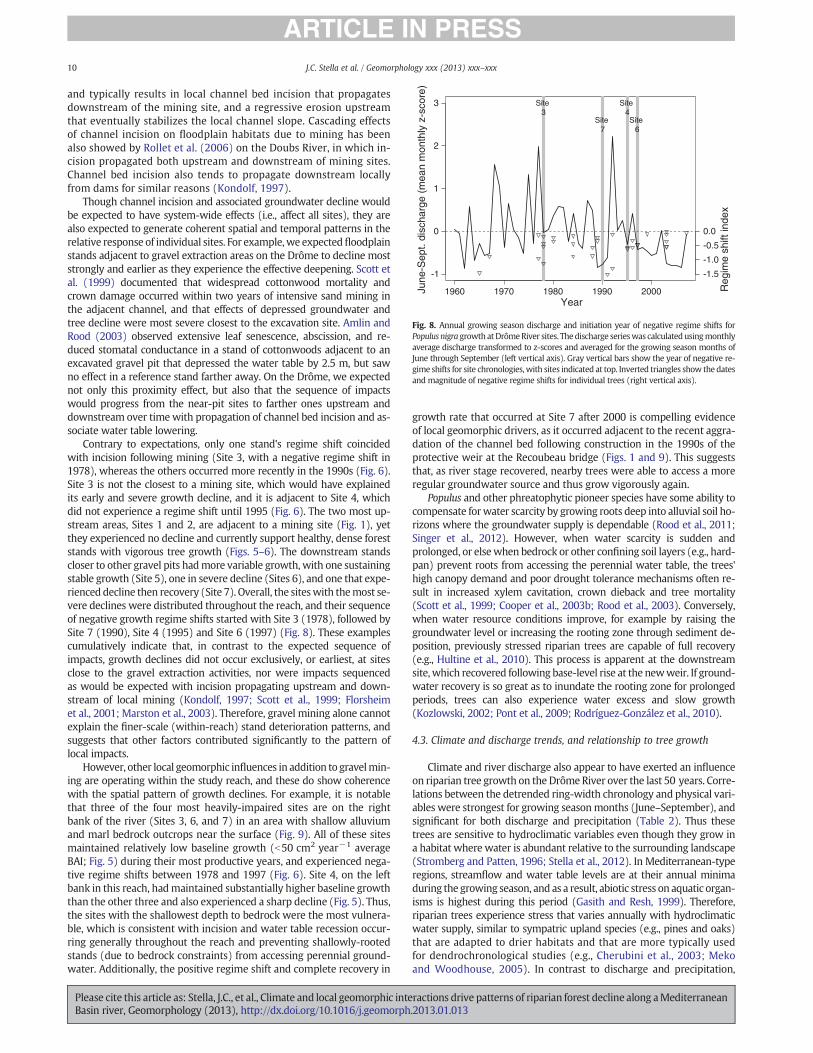
Contrary to expectations, only one stand's regime shift coincidedwith incision following mining (Site 3, with a negative regime shift in1978), whereas the others occurred more recently in the 1990s (Fig. 6).Site 3 is not the closest to a mining site, which would have explainedits early and severe growth decline, and it is adjacent to Site 4, whichdid not experience a regime shift until 1995 (Fig. 6). The two most up-stream areas, Sites 1 and 2, are adjacent to a mining site (Fig. 1), yetthey experienced no decline and currently support healthy, dense foreststands with vigorous tree growth (Figs. 5–6). The downstream standscloser to other gravel pits hadmore variable growth, with one sustainingstable growth (Site 5), one in severe decline (Sites 6), and one that expe-rienced decline then recovery (Site 7). Overall, the siteswith themost se-vere declines were distributed throughout the reach, and their sequenceof negative growth regime shifts started with Site 3 (1978), followed bySite 7 (1990), Site 4 (1995) and Site 6 (1997) (Fig. 8). These examplescumulatively indicate that, in contrast to the expected sequence ofimpacts, growth declines did not occur exclusively, or earliest, at sitesclose to the gravel extraction activities, nor were impacts sequencedas would be expected with incision propagating upstream and down-stream of local mining (Kondolf, 1997; Scott et al., 1999; Florsheimet al., 2001; Marston et al., 2003). Therefore, gravel mining alone cannotexplain the finer-scale (within-reach) stand deterioration patterns, andsuggests that other factors contributed significantly to the pattern oflocal impacts.
However, other local geomorphic influences in addition to gravelmin-ing are operating within the study reach, and these do show coherencewith the spatial pattern of growth declines. For example, it is notablethat three of the four most heavily-impaired sites are on the rightbank of the river (Sites 3, 6, and 7) in an area with shallow alluviumand marl bedrock outcrops near the surface (Fig. 9). All of these sitesmaintained relatively low baseline growth (b50 cm2 year!1 averageBAI; Fig. 5) during their most productive years, and experienced nega-tive regime shifts between 1978 and 1997 (Fig. 6). Site 4, on the leftbank in this reach, hadmaintained substantially higher baseline growththan the other three and also experienced a sharp decline (Fig. 5). Thus,the sites with the shallowest depth to bedrock were the most vulnera-ble, which is consistent with incision and water table recession occur-ring generally throughout the reach and preventing shallowly-rootedstands (due to bedrock constraints) from accessing perennial ground-water. Additionally, the positive regime shift and complete recovery in
growth rate that occurred at Site 7 after 2000 is compelling evidenceof local geomorphic drivers, as it occurred adjacent to the recent aggra-dation of the channel bed following construction in the 1990s of theprotective weir at the Recoubeau bridge (Figs. 1 and 9). This suggeststhat, as river stage recovered, nearby trees were able to access a moreregular groundwater source and thus grow vigorously again.
Populus and other phreatophytic pioneer species have some ability tocompensate forwater scarcity by growing roots deep into alluvial soil ho-rizons where the groundwater supply is dependable (Rood et al., 2011;Singer et al., 2012). However, when water scarcity is sudden andprolonged, or elsewhen bedrock or other confining soil layers (e.g., hard-pan) prevent roots from accessing the perennial water table, the trees'high canopy demand and poor drought tolerance mechanisms often re-sult in increased xylem cavitation, crown dieback and tree mortality(Scott et al., 1999; Cooper et al., 2003b; Rood et al., 2003). Conversely,when water resource conditions improve, for example by raising thegroundwater level or increasing the rooting zone through sediment de-position, previously stressed riparian trees are capable of full recovery(e.g., Hultine et al., 2010). This process is apparent at the downstreamsite,which recovered following base-level rise at the newweir. If ground-water recovery is so great as to inundate the rooting zone for prolongedperiods, trees can also experience water excess and slow growth(Kozlowski, 2002; Pont et al., 2009; Rodríguez-González et al., 2010).
4.3. Climate and discharge trends, and relationship to tree growth
Climate and river discharge also appear to have exerted an influenceon riparian tree growth on the DrômeRiver over the last 50 years. Corre-lations between the detrended ring-width chronology and physical vari-ables were strongest for growing seasonmonths (June–September), andsignificant for both discharge and precipitation (Table 2). Thus thesetrees are sensitive to hydroclimatic variables even though they grow ina habitat where water is abundant relative to the surrounding landscape(Stromberg and Patten, 1996; Stella et al., 2012). In Mediterranean-typeregions, streamflow and water table levels are at their annual minimaduring the growing season, and as a result, abiotic stress on aquatic organ-isms is highest during this period (Gasith and Resh, 1999). Therefore,riparian trees experience stress that varies annually with hydroclimaticwater supply, similar to sympatric upland species (e.g., pines and oaks)that are adapted to drier habitats and that are more typically usedfor dendrochronological studies (e.g., Cherubini et al., 2003; Mekoand Woodhouse, 2005). In contrast to discharge and precipitation,
1960 1970 1980 1990 2000
-1
0
1
2
3
June
-Sep
t. di
scha
rge
(mea
n m
onth
ly z
-sco
re)
Site 3
Site 7
Site 4Site
6
-1.5
-1.0
-0.5
0.0
Year
Reg
ime
shift
inde
x
Fig. 8. Annual growing season discharge and initiation year of negative regime shifts forPopulus nigra growthatDrômeRiver sites. Thedischarge serieswas calculatedusingmonthlyaverage discharge transformed to z-scores and averaged for the growing season months ofJune through September (left vertical axis). Gray vertical bars show the year of negative re-gime shifts for site chronologies,with sites indicated at top. Inverted triangles show the datesand magnitude of negative regime shifts for individual trees (right vertical axis).
10 J.C. Stella et al. / Geomorphology xxx (2013) xxx–xxx
Please cite this article as: Stella, J.C., et al., Climate and local geomorphic interactions drive patterns of riparian forest decline along aMediterraneanBasin river, Geomorphology (2013), http://dx.doi.org/10.1016/j.geomorph.2013.01.013

growth correlations with temperature were poorer and not significant(Table 2). Though riparian tree phenology inMediterranean-type regionsis sensitive to seasonal temperature differences (Stella et al., 2006), tem-perature is not typically limiting to growth for most of the leaf-on period,
except for high daytime vapor pressure deficits in mid-summer thatinduce stomatal closure (Lambs et al., 2006; Hultine et al., 2010). There-fore, temperature is likely limiting primarily through its effects on waterdemand.
Bedrock outcrop
Weir
flow
7
6
3
4
5
2
1
Luc- en-
Diois
Recoubeau
!
Bedrock contact
a)
b) c)
Fig. 9. Panel (a), topographic map of the Drôme River study reach showing the river channel alignment (dashed line), steep valleywalls on the right bank, and location of Populus nigra stands(numbered sites). Sites 3, 4, 6, and 7 suffered negative growth regime shifts. Panel (b), ground-level photograph ofmarl outcrop and shallow alluviumdepth on theDrôme's right bank at Site 3.Panel (c), aerial photograph of the Recoubeau bridge, protective weir, and bedrock outcrop on the right bank; these are just downstream of Site 7, where tree growth declined in 1990 andrecovered in 2000. Map data from SCAN 25®; copyright© IGN, 2008.
11J.C. Stella et al. / Geomorphology xxx (2013) xxx–xxx
Please cite this article as: Stella, J.C., et al., Climate and local geomorphic interactions drive patterns of riparian forest decline along aMediterraneanBasin river, Geomorphology (2013), http://dx.doi.org/10.1016/j.geomorph.2013.01.013

Despite a somewhat limited sample size, the temporal patternof growth declines in individual trees and sites suggests a generalcorrespondence with severely dry years or longer droughts (Fig. 8).The trees at Site 3 initiated a growth decline in 1978, a year with averagesummer discharge, but this followed severe droughts in 1974 and 1976and very high annual variability throughout the 1970s (Fig. 8). This peri-od also coincideswith the intensive gravelmining activity and, assumingthat channel incision had occurred, trees would have been predisposedto water stress. Negative regime shifts for individual trees also tendedto start in drought years, though in some cases they began the yearafter, and in other cases do not appear related to discharge.
Factors other than climate and geomorphic change may also haveinfluenced the stand-level pattern of growth regime shifts. Onemethod-ological issue is that the regime shift detection method recognizes stepchanges in time series, which can mask the timing of a more gradualand sustained trend. Most trees and sites show fairly sudden rapid shiftsin growth rate (e.g., Sites 3, 4, and Site 7 recovery in 1999), but others(Sites 6 and 7 declines) show some evidence of a more gradual declinethat initiated in the late 1970s or early 1980s (Fig. 5). Another reasonfor the asynchrony in timing of site growth declines with climate andgeomorphic change may have to do with the differential response of in-dividual trees at a site. There is great variability in both early growth rateand onset of slow growth among trees at a site (Fig. 5), suggesting thatbackground site conditions were quite varied spatially, and that follow-ing the onset of multiple environmental stressors, site conditions fortrees reached negative thresholds for growth at different times.
4.4. Synergistic drivers and global change
Though neither channel and groundwater alteration from gravelmin-ing nor the sequence of short-term droughts is sufficient alone to explainthe fine-scale patterns of forest stand decline along the Drôme River,there is compelling evidence that these factors acted jointly to influencethe timing and location of the strongest impacts. First, the recentness ofall stand-level declines (i.e., since the early 1970s) suggests that channelbed incision and associated groundwater table decline in adjustment tomining contributed to overall forest vulnerability throughout the reach.Secondly, dry years following the early 1970s appeared to provide envi-ronmental triggers that induced major growth declines. For example,the first decline at Site 3 (1978) occurred after several major dry years(1974 and 1976), and the declines at Sites 4, 6 and 7 were associatedwith individual dry years that occurred subsequent to the mining period(Fig. 8). Finally, the positive regime shift at Site 7 in 2000 following con-struction nearby of the Recoubeau bridge weir (Figs. 1 and 9c) suggeststhat local geomorphic manipulation can reverse growth declines by im-proving sediment/groundwater conditions. Together, these examplesdemonstrate the high sensitivity of forest stands to resource availabilityfrom a combination of drought and local geomorphic factors that includereach-wide incision, locally-induced aggradation, and bedrock controlson soil depth.
It is clear that, in addition to these stressors, the Drôme basin isundergoing long-term climate change to a hotter, drier growing sea-son and discharge regime (Fig. 7). Though precipitation trends werenot significant at the Luc-en-Diois station (Fig. 7b), precipitation hasdeclined significantly over the basin as a whole (E-OBS gridded datanot shown). These trends are expected to continue and accelerate inthe future throughout the Mediterranean Basin (Giorgi and Lionello,2008; García-Ruiz et al., 2011). For mature riparian forest stands, itis likely that local water availability will decrease as the river stageand groundwater table recede farther, faster, and more frequentlyduring the growing season. These processes will stress mature treesthat have developed root distributions, crown size and other adaptivemorphological traits in response to wetter conditions such as higherwater tables and less variability (Rood et al., 2011). Climate changeimpacts can be expected to be most severe at sites with local geomor-phic and/or geologic controls. Existing pioneer stands may decline or
transition to other vegetation types faster and irreversibly under theseconditions, and new stands may have trouble establishing (Shafrothet al., 2000). Riparian studies in other water-limited ecosystems show acommon response of forest decline following rapid hydrologic change(Stromberg and Patten, 1996; Scott et al., 1999; Dufour and Piégay,2008), and in some cases an irreversible shift to alternative system states(Stromberg, 2001; Palmer et al., 2008). This process has occurred local-ly on the Drôme (Sites 3, 4, and 6 in particular), in which stands thatsustained high and variable growth initially switched to permanent-ly suppressed growth with little variation annually or amongst trees.Though there remains some uncertainty as to the specific ecologicaltrigger(s) that would precipitate widespread mortality or growth de-clines in any one location (Groffman et al., 2006), it is clear that the cli-mate sensitivity and accelerating hydroclimatic change, combined withmultiple local stressors due to geologic setting and human-inducedchanges, will make drought-prone riparian zones exceedingly vulnera-ble in the future (Rood et al., 2008; Perry et al., 2011).
5. Conclusions
In the case of Drôme River poplars, major declines in growth werecoincident with a period of reach-based geomorphic alteration frominstream gravel mining, and these impacts were most severe on flood-plain surfaces with thin alluvial substrates overlaying shallow bedrock.Furthermore, these declines were triggered for many trees in droughtyears, and the climate in the region is trending significantly toward hot-ter growing seasons with a decrease in summer river discharge. Thesetrends, which are projected to continue, will increase both chronic andacute water shortage for riparian trees in drought-prone ecosystems,with phreatophytic species such as P. nigra particularly vulnerable.Chronic stress from climate change may induce growth declines soonerand more severely in the future.
This study has shown that tree-ring growth is a sensitive indicatorof water stress for riparian trees, and is a useful tool to understand theeffects of hydrogeomorphic controls and human modifications on riverecosystems. Riparian forests in semi-arid systems are sensitive to manyphysical drivers and can demonstrate threshold responses to multiplestressors. Pioneer trees respond to decreased water availability throughgreater mortality and crown loss, and decreased radial growth. Howev-er, the severity of these impacts depends on interactions with physicaldrivers atmultiple scales, and not all floodplain sites are affected equal-ly. Site-based variation in environmental conditions needs to be takeninto account to understand specific vulnerability to hydrogeomorphicalteration.
Acknowledgments
This work was supported financially by a grant from the Interna-tional Collaborative Scientific Project (PICS) and a seed grant fromthe State University of New York Research Foundation. We gratefullyacknowledge the E-OBS dataset from the EU-FP6 project ENSEMBLES(http://ensembles-eu.metoffice.com) and the data providers in theEuropean Climate Assessment and Dataset project (http://eca.knmi.nl).Finally, we thank Alex Fremier and Maya Hayden for their assistance inthe field, and three anonymous reviewers and Adrian Harvey for im-provements to the manuscript.
References
Albertson, F.W., Weaver, J.E., 1945. Injury and death or recovery of trees in prairie climate.Ecological Monographs 15, 393–433.
Amlin, N.M., Rood, S.B., 2003. Drought stress and recovery of riparian cottonwoods due towater table alteration along Willow Creek, Alberta. Trees-Structure and Function 17,351–358.
Biondi, F., Qeadan, F., 2008. A theory-driven approach to tree-ring standardization: de-fining the biological trend from expected basal area increment. Tree-Ring Research64, 81–96.
12 J.C. Stella et al. / Geomorphology xxx (2013) xxx–xxx
Please cite this article as: Stella, J.C., et al., Climate and local geomorphic interactions drive patterns of riparian forest decline along aMediterraneanBasin river, Geomorphology (2013), http://dx.doi.org/10.1016/j.geomorph.2013.01.013

Braatne, J.H., Rood, S.B., Heilman, P.E., 1996. Life history, ecology, and conservation of ripariancottonwoods in North America. In: Stettler, R.F., Bradshaw Jr., H.D., Heilman, P.E.,Hinckley, T.M. (Eds.), Biology of Populus and Its Implication for Management and Con-servation. NRC Research Press, National Research Council of Canada, Ottawa, pp. 57–86.
Cherubini, P., Gartner, B.L., Tognetti, R., Bräker, O.U., Schoch, W., Innes, J.L., 2003. Identifica-tion, measurement and interpretation of tree rings in woody species from Mediterra-nean climates. Biological Reviews of the Cambridge Philosophical Society 78, 119–148.
Cook, E.R. 1985. A time series analysis approach to tree ring standardization. PhD, Universityof Arizona, Tucson. 171 pp.
Cook, E.R., Peters, K., 1981. The smoothing spline: a new approach to standardizing forestinterior tree-ring width series for dendroclimatic studies. Tree-Ring Bulletin 41, 45–55.
Cook, E.R., Briffa, K.R., Shiyatov, S.G., Mazepa, V., 1990. Tree-ring standardization andgrowth-trend estimation. In: Cook, E.R., Kairiukstis, L. (Eds.),Methods of Dendrochronol-ogy: Applications in the Environmental Sciences. Kluwer Academic Publishers, Boston,pp. 104–122.
Cooper, D.J., Andersen, D.C., Chimner, R.A., 2003a. Multiple pathways for woody plant estab-lishment on floodplains at local to regional scales. Journal of Ecology 91, 182–196.
Cooper, D.J., D'Amico, D.R., Scott, M.L., 2003b. Physiological andmorphological response pat-terns of Populus deltoides to alluvial groundwater pumping. Environmental Manage-ment 31, 215–226.
Dufour, S., Piégay, H., 2008. Geomorphological controls of Fraxinus excelsior growth and re-generation in floodplain forests. Ecology 89, 205–215.
Dufour, S., Barsoum, N., Muller, E., Piégay, H., 2007. Effects of channel confinementon pioneer woody vegetation structure, composition and diversity along theRiver Drôme (SE France). Earth Surface Processes and Landforms 32, 1244–1256.
Dunford, R., Michel, K., Gagnage,M., Piégay, H., Trémelo, M.L., 2009. Potential and constraintsof Unmanned Aerial Vehicle technology for the characterization ofMediterranean ripar-ian forest. International Journal of Remote Sensing 30, 4915–4935.
Ellison, A.M., Bank, M.S., Clinton, B.D., Colburn, E.A., Elliott, K., Ford, C.R., Foster, D.R., Kloeppel,B.D., Knoepp, J.D., Lovett, G.M., Mohan, J., Orwig, D.A., Rodenhouse, N.L., Sobczak, W.V.,Stinson, K.A., Stone, J.K., Swan, C.M., Thompson, J., Von Holle, B., Webster, J.R., 2005.Loss of foundation species: consequences for the structure and dynamics of forestedecosystems. Frontiers in Ecology and the Environment 3, 479–486.
Florsheim, J.L., Mount, J.F., Rutten, L.T., 2001. Effect of baselevel change on floodplain andfan sediment storage and ephemeral tributary channel morphology, Navarro River,California. Earth Surface Processes and Landforms 26, 219–232.
Florsheim, J.L., Mount, J.F., Chin, A., 2008. Bank erosion as a desirable attribute of rivers.Bioscience 58, 519–529.
Friedman, J.H., 1984. A variable span smoother. Technical Report No. 5, Laboratory for Com-putational Statistics. Department of Statistics Stanford University, Palo Alto, CA (32 pp.).
Fritts, H.C., 1976. Tree Rings and Climate. Academic Press, New York (567 pp.).Gagnage, M., 2008. Evaluation des effets des changements morphologiques fluviaux sur la
santé des peupliers M.S. Université Lyon 2 Lumière, Lyon, FR (101 pp.).García-Ruiz, J.M., López-Moreno, J.I., Vicente-Serrano, S.M., Lasanta-Martínez, T., Beguería,
S., 2011. Mediterranean water resources in a global change scenario. Earth-ScienceReviews 105, 121–139.
Gasith, A., Resh, V.H., 1999. Streams in Mediterranean climate regions: abiotic influencesand biotic responses to predictable seasonal events. Annual Review of Ecology andSystematics 30, 51–81.
Giorgi, F., Lionello, P., 2008. Climate change projections for the Mediterranean region.Global and Planetary Change 63, 90–104.
Graf, W.L., 1982. Tamarisk and river-channel management. Environmental Management6, 283–296.
Grissino-Mayer, H.D., 2001. Evaluating crossdating accuracy: a manual and tutorial forthe computer program COFECHA. Tree-Ring Research 57, 205–221.
Groffman, P., Baron, J., Blett, T., Gold, A., Goodman, I., Gunderson, L., Levinson, B., Palmer, M.,Paerl, H., Peterson, G., Poff, N., Rejeski, D., Reynolds, J., Turner, M., Weathers, K., Wiens, J.,2006. Ecological thresholds: the key to successful environmental management or animportant concept with no practical application? Ecosystems 9, 1–13.
Haylock,M.R., Hofstra, N., Klein Tank, A.M.G., Klok, E.J., Jones, P.D., New,M., 2008. A Europeandaily high-resolution gridded dataset of surface temperature and precipitation. Journalof Geophysical Research-Atmospheres 113, D20119. http://dx.doi.org/10.1029/2008JD10201.
Holmes, R., 1983. Computer-assisted quality control in tree-ring dating and measurement.Tree-Ring Bulletin 44, 69–75.
Horton, J.L., Kolb, T.E., Hart, S.C., 2001. Physiological response to groundwater depth variesamong species and with river flow regulation. Ecological Applications 11, 1046–1059.
Hughes, F.M.R., Rood, S.B., 2003. Allocation of river flows for restoration of floodplain forestecosystems: a review of approaches and their applicability in Europe. EnvironmentalManagement 32, 12–33.
Hultine, K.R., Bush, S.E., Ehleringer, J.R., 2010. Ecophysiology of riparian cottonwood andwil-low before, during, and after two years of soil water removal. Ecological Applications 20,347–361.
Karrenberg, S., Edwards, P.J., Kollmann, J., 2002. The life history of Salicaceae living inthe active zone of floodplains. Freshwater Biology 47, 733–748.
Kendall, M.G., 1975. Rank Correlation Methods. Griffin, London (202 pp.).Kondolf, G.M., 1997. Hungry water: effects of dams and gravel mining on river channels.
Environmental Management 21, 533–551.Kondolf, G.M., Piegay, H., Landon, N., 2002. Channel response to increased and decreased
bedload supply from land use change: contrasts between two catchments. Geomor-phology 45, 35–51.
Kozlowski, T.T., 2002. Physiological-ecological impacts of flooding on riparian forest ecosys-tems. Wetlands 22, 550–561.
Lambs, L., Loubiat, M., Girel, J., Tissier, J., Peltier, J.P., Marigo, G., 2006. Survival andacclimatation of Populus nigra to drier conditions after damming of an alpineriver, southeast France. Annals of Forest Science 63, 377–385.
Landon, N., Piégay, H., Bravard, J.P., 1998. The Drôme river incision (France): from as-sessment to management. Landscape and Urban Planning 43, 119–131.
Lefèvre, F., Légionnet, A., Sven de Vries, S., Turok, J., 1998. Strategies for the conserva-tion of a pioneer tree species, Populus nigra L., in Europe. Genetics Selection Evolu-tion 30 (Suppl. 1), S181–S196.
Lejot, J., Piégay, H., Hunter, P.D., Moulin, B., Gagnage, M., 2011. Utilisation de latélédétection pour la caractérisation des corridors fluviaux : exemples d’applicationset enjeux actuels. Géomorphologie: Relief, Processus, Environnement 2, 37–52.
Lewis, D.B., Harms, T.K., Schade, J.D., Grimm, N.B., 2009. Biogeochemical function andheterogeneity in arid-region riparian zones. In: Stromberg, J.C., Tellman, J. (Eds.),Ecology and Conservation of the San Pedro River. University of Arizona Press, Tucson,pp. 323–341.
Mahoney, J.M., Rood, S.B., 1998. Streamflow requirements for cottonwood seedlingrecruitment — an integrative model. Wetlands 18, 634–645.
Mann, H.B., 1945. Nonparametric tests against trend. Econometrica 13, 245–259.Marston, R.A., Bravard, J.P., Green, T., 2003. Impacts of reforestation and gravel mining
on the Malnant River, Haute-Savoie, French Alps. Geomorphology 55, 65–74.Meko, D.M., Woodhouse, C.A., 2005. Tree-ring footprint of joint hydrologic drought in
Sacramento and Upper Colorado river basins, western USA. Journal of Hydrology308, 196–213.
Moggridge, H.L., Gurnell, A.M., 2009. Controls on the sexual and asexual regenerationof Salicaceae along a highly dynamic, braided river system. Aquatic Sciences 71,305–317.
Palmer, M.A., Reidy Liermann, C.A., Nilsson, C., Flörke, M., Alcamo, J., Lake, P.S., Bond, N.,2008. Climate change and the world's river basins: anticipating management op-tions. Frontiers in Ecology and the Environment 6, 81–89.
Palmer, M.A., Lettenmaier, D.P., Poff, N.L., Postel, S.L., Richter, B., Warner, R., 2009. Climatechange and river ecosystems: protection and adaptation options. Environmental Man-agement 44, 1053–1068.
Patten, D.T., 1998. Riparian ecosystems of semi-arid North America: diversity andhuman impacts. Wetlands 18, 498–512.
Perry, L.G., Andersen, D.C., Reynolds, L.V., Nelson, S.M., Shafroth, P.B., 2011. Vulnerabil-ity of riparian ecosystems to elevated CO2 and climate change in arid and semiaridwestern North America. Global Change Biology 18, 821–842.
Piégay, H., Landon, N., 1997. Promoting ecological management of riparian forests onthe Drome River, France. Aquatic Conservation: Marine and Freshwater Ecosys-tems 7, 287–304.
Piégay, H., Darby, S.E., Mosselman, E., Surian, N., 2005. A review of techniques availablefor delimiting the erodible river corridor: a sustainable approach to managing bankerosion. River Research and Applications 21, 773–789.
Pont, D., Piégay, H., Farinetti, A., Allain, S., Landon, N., Liébault, F., Dumont, B., Richard-Mazet, A., 2009. Conceptual framework and interdisciplinary approach for the sus-tainable management of gravel-bed rivers: the case of the Drôme River basin (S.E.France). Aquatic Sciences 71, 356–370.
Rodionov, S.N., 2004. A sequential algorithm for testing climate regime shifts. Geophys-ical Research Letters 31, L09204. http://dx.doi.org/10.1029/2004GL019448.
Rodionov, S.N., 2006a. Use of prewhitening in climate regime shift detection. Geophys-ical Research Letters 33, L12707. http://dx.doi.org/10.1029/2006GL025904.
Rodionov, S.N., 2006b. Sequential Regime Shift Detection, Version 3.2 (software add-in to Excel 2002). NOAA Pacific Marine Environmental Laboratory, Seattle, WA(http://www.beringclimate.noaa.gov/regimes/).
Rodríguez-González, P.M., Stella, J.C., Campelo, F., Ferreira, M.T., Albuquerque, A., 2010.Subsidy or stress? Tree structure and growth in wetland forests along a hydrologicalgradient in Southern Europe. Forest Ecology and Management 259, 2015–2025.
Rollet, A.J., Piégay, H., Citterio, A., 2006. Impact des extractions de graviers dans le litmineur sur la géométrie des zones aquatiques périfluviales du Doubs (France).Géographie physique et Quaternaire 60, 253–270.
Rood, S.B., Patino, S., Coombs, K., Tyree, M.T., 2000. Branch sacrifice: cavitation-associateddrought adaptation of riparian cottonwoods. Trees-Structure and Function 14, 248–257.
Rood, S.B., Braatne, J.H., Hughes, F.M.R., 2003. Ecophysiology of riparian cotton-woods: stream flow dependency, water relations and restoration. Tree Physiolo-gy 23, 1113–1124.
Rood, S.B., Pan, J., Gill, K.M., Franks, C.G., Samuelson, G.M., Shepherd, A., 2008. Decliningsummer flows of Rocky Mountain rivers: changing seasonal hydrology and proba-ble impacts on floodplain forests. Journal of Hydrology 349, 397–410.
Rood, S.B., Bigelow, S.G., Hall, A.A., 2011. Root architecture of riparian trees: river cut-banks provide natural hydraulic excavation, revealing that cottonwoods are facul-tative phreatophytes. Trees 25, 907–917.
Scott, M.L., Shafroth, P.B., Auble, G.T., 1999. Responses of riparian cottonwoods to allu-vial water table declines. Environmental Management 23, 347–358.
Scott, M.L., Lines, G.C., Auble, G.T., 2000. Channel incision and patterns of cottonwoodstress and mortality along the Mojave River, California. Journal of Arid Environments44, 399–414.
Sen, P.K., 1968. Estimates of the regression coefficient based on Kendall's tau. Journal ofthe American Statistical Association 63, 1379–1389.
Shafroth, P.B., Stromberg, J.C., Patten, D.T., 2000. Woody riparian vegetation responseto different alluvial water table regimes. Western North American Naturalist 60,66–76.
Singer, M.B., Stella, J.C., Dufour, S., Johnstone, L.B., Piégay, H., Wilson, R.J.S., 2012. Con-trasting water-uptake and growth responses to drought in co-occurring ripariantree species. Ecohydrology. http://dx.doi.org/10.1002/eco.1283.
Steiger, J., Tabacchi, E., Dufour, S., Corenblit, D., Peiry, J.L., 2005. Hydrogeomorphic process-es affecting riparian habitatwithin alluvial channel-floodplain river systems: a reviewfor the temperate zone. River Research and Applications 21, 719–737.
Stella, J.C., Battles, J.J., 2010. How do riparian woody seedlings survive seasonal drought?Oecologia 164, 579–590.
13J.C. Stella et al. / Geomorphology xxx (2013) xxx–xxx
Please cite this article as: Stella, J.C., et al., Climate and local geomorphic interactions drive patterns of riparian forest decline along aMediterraneanBasin river, Geomorphology (2013), http://dx.doi.org/10.1016/j.geomorph.2013.01.013

Stella, J.C., Battles, J.J., Orr, B.K., McBride, J.R., 2006. Synchrony of seed dispersal, hydrologyand local climate in a semi-arid river reach in California. Ecosystems 9, 1200–1214.
Stella, J.C., Battles, J.J., McBride, J.R., Orr, B.K., 2010. Riparian seedling mortality from sim-ulatedwater table recession, and the design of sustainable flow regimes on regulatedrivers. Restoration Ecology 18, 284–294.
Stella, J.C., Hayden, M.K., Battles, J.J., Piégay, H., Dufour, S., Fremier, A.K., 2011. The role ofabandoned channels as refugia for sustaining pioneer riparian forest ecosystems.Ecosystems 14, 776–790.
Stella, J.C., Rodríguez-González, P.M., Dufour, S., Bendix, J., 2012. Riparian vegetationresearch in Mediterranean-climate regions: common patterns, ecological process-es, and considerations for management. Hydrobiologia. http://dx.doi.org/10.1007/s10750-012-1304-9.
Stokes, M.A., Smiley, T.L., 1968. An Introduction to Tree-Ring Dating. The University ofChicago Press, Chicago (73 pp.).
Stromberg, J.C., 2001. Restoration of riparian vegetation in the south-western UnitedStates: importance of flow regimes and fluvial dynamism. Journal of Arid Environ-ments 49, 17–34.
Stromberg, J.C., Patten, D.T., 1990. Riparian vegetation in-stream flow requirements: acase study from a diverted stream in the eastern Sierra Nevada, California, USA.Environmental Management 14, 185–194.
Stromberg, J.C., Patten, D.T., 1996. Instream flow and cottonwood growth in the easternSierra Nevada of California, USA. Regulated Rivers: Research & Management 12, 1–12.
Stromberg, J.C., Tiller, R., Richter, B., 1996. Effects of ground water decline on riparian vege-tation of semiarid regions: the San Pedro, Arizona. Ecological Applications 6, 113–131.
Stromberg, J.C., Lite, S.J., Dixon, M.D., 2010. Effects of stream flow patterns on riparianvegetation of a semiarid river: implications for a changing climate. River Researchand Applications 26, 712–729.
Theil, H., 1950a. A rank-invariant method of linear and polynomial regression analysis, I.Koninklijke Nederlandse Akademie van Wetenschappen, Proceedings 53, 386–392.
Theil, H., 1950b. A rank-invariant method of linear and polynomial regression analysis, II.Koninklijke Nederlandse Akademie van Wetenschappen, Proceedings 53, 521–525.
Theil, H., 1950c. A rank-invariant method of linear and polynomial regression analysis, III.Koninklijke Nederlandse Akademie van Wetenschappen, Proceedings 53, 1397–1412.
Toone, J., Rice, S.P., Piégay, H., 2012. Spatial discontinuity and temporal evolution ofchannel morphology along a mixed bedrock-alluvial river, upper Drôme River,southeast France: contingent responses to external and internal controls. Geomor-phology. http://dx.doi.org/10.1016/j.geomorph.2012.05.033.
Tyree, M.T., Kolb, K.J., Rood, S.B., Patino, S., 1994. Vulnerability to drought-induced cavita-tion of riparian cottonwoods in Alberta — a possible factor in the decline of the eco-system. Tree Physiology 14, 455–466.
Underwood, E.C., Viers, J.H., Klausmeyer, K.R., Cox, R.L., Shaw, M.R., 2009. Threats andbiodiversity in the mediterranean biome. Diversity and Distributions 15, 188–197.
Wigley, T.M.L., Briffa, K.R., Jones, P.D., 1984. On the average value of correlated time series,with applications in dendroclimatology and hydrometeorology. Journal of Climateand Applied Meteorology 23, 201–213.
Young-Mathews, A., Culman, S.W., Sánchez-Moreno, S., O'Geen, A.T., Ferris, H., Hollander,A.D., Jackson, L.E., 2010. Plant-soil biodiversity relationships and nutrient retention inagricultural riparian zones of the Sacramento Valley, California. Agroforestry Systems80, 41–60.
Yue, S., Pilon, P., Phinney, B., Cavadias, G., 2002. The influence of autocorrelation on theability to detect trend in hydrological series. Hydrological Processes 16, 1807–1829.
14 J.C. Stella et al. / Geomorphology xxx (2013) xxx–xxx
Please cite this article as: Stella, J.C., et al., Climate and local geomorphic interactions drive patterns of riparian forest decline along aMediterraneanBasin river, Geomorphology (2013), http://dx.doi.org/10.1016/j.geomorph.2013.01.013





























