Embed Size (px)
Citation preview

Yeast 15, 507–511 (1999)
Heterologous URA3MX Cassettes for GeneReplacement in Saccharomyces cerevisiae
ALAN L. GOLDSTEIN, XUEWEN PAN AND JOHN H. MCUSKER*
Department of Microbiology, Duke University Medical Center, Durham, North Carolina 27710, USA
Heterologous gene replacement cassettes are powerful tools for dissecting gene function in Saccharomyces cerevisiae.Their primary advantages over homologous gene replacement cassettes include reduced gene conversion (leading toefficient site-specific integration of the cassette) and greater independence of strain background. Perhaps the mostwidely used cassettes are the MX cassettes containing the dominant selectable kanamycin resistance gene (kanr),which confers resistance to G418 (Wach et al., 1994). One limitation of the kanMX cassettes is that they are notcounterselectable and therefore not readily recyclable, which is important when constructing strains with more thanone gene deletion. To address this limitation, and to expand the choices of heterologous markers, we have createdtwo new MX cassettes by replacing the kanr ORF from plasmids pFA6–kanMX3 and pFA6–kanMX4 with theCandida albicans URA3 ORF. These plasmids, pAG60 (CaURA3MX4) and pAG61 (CaURA3MX3) are identical tothe kanMX cassettes in all other respects but have the added advantage of being counterselectable and thereforereadily recyclable in S. cerevisiae. Copyright ? 1999 John Wiley & Sons, Ltd.
— URA3; MX cassette; 5-FOA; heterologous marker; selectable marker
INTRODUCTION
Genetic analysis of Saccharomyces cerevisiae hasbeen aided by the development of heterologousgene replacement cassettes. These cassettes can bePCR-amplified, with primers containing short(30–40 bases) regions of S. cerevisiae gene hom-ology in the 5* ends, and efficiently targeted tospecific S. cerevisiae genes to create insertion–deletion mutations (Baudin et al., 1993; Lorenzet al., 1995). The MX cassettes are probably themost widely used gene targeting cassettes. Thereare currently four heterologous selectable markersin MX cassettes—the bacterial kanamycin (kanr)resistance open reading frame (ORF), theKluyveromyces lactis HIS3 ORF, the Ashbya
Durham, NC, USA
CCC 0749–503X/99/060507–05 $17.50Copyright ? 1999 John Wiley & Sons, Ltd.
gossypii LEU2 ORF (Wach et al., 1994, 1997) andthe Schizosaccharomyces pombe his5 (Longtineet al., 1998)—which allows some flexibility andchoice in selectable markers.
Because many S. cerevisiae strains contain ura3mutations, it would be useful to have a MXcassette containing an heterologous URA3. There-fore, we have created two new MX cassettes thatreplace the kanr ORF of kanMX3 and kanMX4with the Candida albicans URA3 ORF. Like thekanMX3 and kanMX4 cassettes, the CaURA3MXcassettes are heterologous. However, unlike theexisting MX cassettes, the CaURA3MX3 cassetteis counterselectable in 5-fluoro-orotic acid (5-FOA)-containing media and is therefore easilyrecycled for repeated use.
MATERIALS AND METHODS
*Correspondence to: John H. McCusker PhD, Box 3020DUMC, Dept. of Microbiology, Duke University MedicalCenter, Durham, NC 27710, USA. Tel: 919 681-6744; fax: 919684-8735; e-mail: [email protected]/grant sponsor: Duke University Medical Center,MediaYeast extract peptone dextrose (YPD), synthetic
dextrose (SD), synthetic dextrose complete (SDC),
Received 15 May 1998Accepted 12 October 1998

508 . . , . . .
and 5-fluoro-orotic-acid 5-FOA media were pre-pared as previously described (Rose et al., 1989).
Preparation of C. albicans genomic DNA andPCR amplification of the URA3 ORF
Genomic DNA was prepared from a pro-totrophic C. albicans strain ATCC 36801 usingstandard protocols for purifying S. cerevisiaegenomic DNA (Ausubel et al., 1988). The com-plete C. albicans URA3 open reading frame wasPCR-amplified using primers PR59 and PR60.These primers also contain 5*-extensions (seeTable 1; extensions underlined) homologous to thepromoter (PR59) or terminator (PR60) directlyadjacent to the kanamycin resistance ORF ofplasmid pFA6–kanMX4 (Wach et al., 1994).
Construction of pAG60 and pAG61Plasmids based upon pFA6–kanMX3 and
pFA6–kanMX4 but containing the C. albicansURA3 ORF instead of the kanr ORF were con-structed in three steps. First, the kanMX cassetteswere subcloned into a yeast/E. coli shuttle vector.Second, the kanamycin resistance ORF wasreplaced by the C. albicans URA3 ORF by in vivoPCR-directed recombination (Oldenburg et al.,1997). In this method the DNA to be cloned isPCR-amplified with 30–40 bp of homology justupstream and downstream to the cloning site.Co-transformation with the PCR product and thelinearized target vector results in recombinationbetween the homologous ends of the PCR productand the target vector. Finally, the CaURA3MXcassettes were subcloned from the shuttle vectors
into the pFA6 vector.Copyright ? 1999 John Wiley & Sons, Ltd.
To subclone the kanMX cassettes into theyeast/E. coli shuttle vector pRS314 (Sikorski andHieter, 1989) (TRP1 CEN ARS), plasmids pFa6–kanMX3 and pFA6–kanMX4 were NotI-digested,releasing the entire cassettes. Gel purified NotI-digested kanMX cassettes were then subclonedinto NotI-digested pRS314, resulting in plasmidspAG24 (kanMX4) and pAG50 (kanMX3). Toreplace the kanMX ORFs with the CaURA3MXORFs by in vivo PCR-directed recombinationalcloning, pAG24 and pAG50 were linearized withinthe kanr ORF by NruI digestion. Approximately100 ng of each plasmid was co-transformed with100 ng of C. albicans URA3 ORF PCR product(amplification described above) into S. cerevisiaestrain YJM395 (MATá ade2–101his3Ä300 leu2Ä1trpÄ1 ura3–52) by the lithium acetate procedure.Trp+ Ura+ transformants were selected onsynthetic dextrose complete medium lackingtryptophan and uracil. Plasmids pXP45(CaURA3MX4) and pAG51 (CaURA3MX3)were recovered from Trp+ Ura+ yeast transform-ants and shuttled into E. coli for amplification andverification (Rose et al., 1989). Finally theCaURA3MX4 and CaURA3MX3 cassettes weresubcloned into the BamHI and SpeI sites of pFA6to create plasmids pAG60 (pFA6–CaURA3MX4)and pAG61 (pFA6–CaURA3MX3).
Table 1. Oligonucleotides.
Primer Sequence 5* to 3*
JM7 CTCATAAGCAGCAATCAATTCCATCTATACTTTAAAGCATAGGCCACTJM8 CTTTTATTACATACAACTTTTTAAACTAATATACACATTTCAGCTGAAGCTTCGTACGCJM37 CCTCGACATCATCTGCCCPH7 CGCAAGTCCTGTTTCTATGCPH8 ACTCGTTGCCTCCATCGPH14 GTATTGAGAAGGGCAACGGPH15 GGAGTACACCTTATCGGCPR59 TTCTCACATCACATCCGAACATAAACAACCATGACAGTCAACACTAAGACCTATAGTGPR60 AAACAACAATCTTTTTATTGTCAGTACTGATTTTAATTGGCCAGTCTTTTTCAAATAAGPR78 CCTTGACAGTCTTGACGTGCPR79 GCGACTTAACTTCGCATCTG
Underlined portion of oligonucleotides are 5* extension targeting sequences.
Gene disruptionTo integrate the CaURA3MX cassettes into the
S. cerevisiae genome, the cassettes were PCR-amplified in reactions containing 10 m KCl,10 m (NH4)2SO4, 20 m Tris–HCl (pH 8·8),
2 m MgSO4, 0·1% Triton X-100, 25 ng plasmidYeast 15, 507–511 (1999)

509 3
DNA, 0·2 ì each dNTP, 0·5 ì each primerand 1 unit Vent DNA polymerase (New EnglandBioLabs, Inc.). Template amplification (30 cyclesof 94)C for 30 s, 55)C for 15 s, and 72)C for 3 min)was modified from the conditions previouslydescribed by shortening the annealing time. Thisshort annealing time eliminates most of the un-wanted PCR products observed when amplifyingthe direct repeat containing MX3 cassettes.CaURA3MX PCR products were gel purified andapproximately 0·1–1·0 ìg of each product wastransformed into S. cerevisiae strains by lithiumacetate (Geitz et al., 1995). Transformants wereselected on SD plates. Homologous integration ofCaURA3MX cassettes was verified in Ura+ trans-formants by colony PCR (Niedenthal et al., 1996).
CounterselectabilityTo select for pop-out of the CaURA3MX3
cassette in S. cerevisiae, strain YAG60 (MATáura3::CaURA3MX3) was grown overnight inYPD medium containing 0·01% uracil and thenspread on to plates containing 5-FOA (Rose et al.,1989).
RESULTS AND DISCUSSION
MX insertion-deletion cassettes containing thekanamycin resistance gene confer resistance to theaminoglycoside G418; there are also MX cassettescontaining the K. lactis HIS3, A. gossypii LEU2(Wach et al., 1994, 1997), and S. pombe his5 ORFs(Longtine et al., 1998) as selectable markers. Thesesophisticated and widely used gene targeting cas-settes contain transcriptional control sequencesfrom the A. gossypii translation elongation factor 1alpha (TEF) gene which promotes strong tran-scription in S. cerevisiae (Steiner and Phillipsen,1994).
Since heterologous markers do not homolo-gously recombine with genomic DNA, directedsite-specific integration efficiencies are higher thancassettes containing selectable S. cerevisiae genes.This is of particular concern when transformingwith PCR products containing short regions ofgene homology. For example, Langle-Rouault andJacobs (1995) compared disruption of the PMS1locus using the S. cerevisiae URA3 and K. lactisURA3 genes as selectable markers; these ORFs areapproximately 68% identical at the nucleotidelevel. They found that when the S. cerevisiaeURA3 gene was used as the selectable marker,
Copyright ? 1999 John Wiley & Sons, Ltd.
none of the 17 Ura+ clones analysed contained apms1::ScURA3 disruption; in contrast, when theK. lactis URA3 gene was used as a selectablemarker, 10 out of 12 Ura+ clones analysed werecorrectly targeted by the K. lactis URA3 cassette togenerate pms1::KlURA3 mutants (Langle-Rouaultand Jacobs, 1995). In some instances this limita-tion of homologous integration cassettes has beenreduced by the creation of specialized designerstrains carrying deletions of commonly used S.cerevisiae auxotrophic markers (Brachmann et al.,1998).
One limitation of the MX cassettes is that noneare readily recyclable, which is important whenconstructing strains with more than one genedeletion, as when examining gene families (e.g.Dolinski et al., 1997). Although the MX3 cassettescontain 466 bp direct repeats flanking theselectable marker, allowing the cassette to be‘popped-out’ and recycled, loss of the cassettes isinefficient and tedious as there is no selection formarker loss. Since S. cerevisiae strains bearing theURA3 gene are sensitive to 5-fluoro-orotic acid(Boeke et al., 1987), an MX cassette based upon aURA3 ORF would be both selectable in uracildeficient medium and counterselectable in 5-FOA-containing medium.
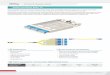
To address these limitations we have created twonew MX cassettes by replacing the kanr ORF fromplasmids pFA6–kanMX3 and pFA6–kanMX4with the C. albicans positively selectable markerURA3 (66·9% identical at nucleotide level with S.cerevisiae URA3 ORF). The construction of theseplasmids, pAG60 (CaURA3MX4) and pAG61(CaURA3MX3) is described in the Materialsand Methods section. Since the C. albicans URA3ORF replaces the kanr ORF from START toSTOP, pAG60 and pAG61 are identical in allother respects to plasmids pFA6–kanMX4 andpFA6–kanMX3, respectively (Figure 1).
The frequency of homologous integration of theCaURA3MX4 cassette by site-directed integrationwas tested by targeting the cassette to the ho locusof S. cerevisiae strain YJM74 (MATá ura3–52).The CaURA3MX4 cassette was PCR amplifiedusing primers JM7 and JM8. The 5* end of JM7contains 38 nt of targeting sequence homologousdirectly upstream of the ho ATG codon followedby 22 nt amplification sequence homologous tothe pFA6 multi-cloning site (MCS). The 5* end ofJM8 contains 40 nt of targeting sequence homolo-gous immediately downstream of the ho TAAcodon, followed by 19 nt of targeting sequence
Yeast 15, 507–511 (1999)

510 . . , . . .
Figure 1. CaURA3MX plasmids pAG60 and pAG61 differonly by the inclusion of direct repeats in pAG61. With theexception of the CaURA3 ORF, these plasmids are identical topFA6–kanMX4 and pFA6–kanMX3, respectively. See Wachet al., 1994, for further details.
homologous to the other side of the pFA6 MCS.As described in Materials and Methods, approxi-mately 1 ìg of gel-purified PCR product wastransformed into strain YJM74 and Ura+ trans-formants were selected on SD plates. To confirmintegration of the cassette into the ho locus,nine colonies were subjected to colony PCR usingprimers JM37 (within the terminator region ofcassette) and PH7 (approximately 500 nt upstreamof ho ATG). Six of the nine colonies producedbands of approximately 600 nt expected for hom-ologous integration of the CaURA3MX4 cassetteinto the ho locus (data not shown). This targetingefficiency is similar to the 50–100% frequencyobserved when the kanMX4 cassette was directedto 11 different S. cerevisiae genes (Shoemakeret al., 1996).
An additional benefit of placing the URA3 ORFin the pFA6-based cassette is that there areapproximately 200–400 bp of homology betweenthe cassettes in the promoter and terminatorregions making cassette exchange relatively easy.For example, a kanr cassette can easily be replacedwith a CaURA3 cassette by transforming thekanMX containing strain with a CaURA3 cassettePCR amplified with primers homologous to thetranscriptional control region common to bothcassettes.
Copyright ? 1999 John Wiley & Sons, Ltd.
To test the facility of cassette swapping,CaURA3MX4 was PCR-amplified using primersPR78 and PR79 which are homologous tosequences within the TEF promoter and termi-nator. Approximately 100 ng of gel-purified PCRproduct was transformed into strain S105 (MATáura3::kanMX3) and Ura+ transformants wereselected on SD plates. From the dozens of colonieson the SD plate, eight were selected for verificationof cassette replacement by colony PCR usingprimers PH7 (2500 nt upstream of ho ATG) andpH8 (2500 nt downstream of ho TAA). Since thekanr and URA3 ORFs are nearly the same size, thePCR products were digested with NruI (only foundin kanr ORF) to discriminate the markers. Alleight colonies contained the CaURA3MX cassetteat the ho locus (data not shown). One strain keptfor further analysis was named YAG60 (MATáura3::CaURA3MX3).
To determine whether the heterologous C.albicans URA3 gene is counterselectable on5-FOA medium in S. cerevisiae, 5-FOA resistantcolonies of S. cerevisiae strain YAG60 (MATáura3::CaURA3MX3) were selected as described inMaterials and Methods. To verify the loss of theCaURA3MX3 marker, colony PCR using primersPH14 and PH15 (500 nt distal to the S. cerevisiaeURA3 ORF), was performed on YAG60 and three5-FOA resistant YAG60 colonies. PCR products,including 1·0 kb of distal genomic DNA, ofapproximately 3·5 and 1·5 kb, were expectedfrom YAG60 and 5-FOA resistant colonies,respectively. The approximate 1·5 kb size forCaURA3MX3 pop-out assumes that recombi-nation between the direct repeats will leave one466 bp direct repeat at the URA3 locus. PCRamplification of the URA3 locus from threeindependent YAG60 5-FOA resistant coloniesproduced products approximately 1·5 kb in length,demonstrating that the CaURA3 marker was lostfrom these cells (data not shown). These resultsshow that the C. albicans URA3 ORF confersboth uracil prototrophy and 5-FOA sensitivity inS. cerevisiae.
In summary, we have created two new MXcassettes by replacing the kanr ORF with the C.albicans URA3 ORF. We have done this in partbecause the heterologous CaURA3MX cassettesprovide another positively selectable marker andtherefore more experimental flexibility when usingMX cassettes. Additionally we have shown thatthe C. albicans URA3 marker is counterselectablein S. cerevisiae cells grown in the presence of
Yeast 15, 507–511 (1999)

511 3
5-FOA, enabling the CaURA3MX3 cassette tofunction as a recyclable heterologous genereplacement marker.
ACKNOWLEDGEMENTS
We thank Drs. Joseph Heitman and PhillipHartzog for comments and reagents. We alsothank Duke University Medical Center forfinancial assistance.
REFERENCES
Ausubel, F. M., Brent, R., Kingston, R. E., Moore,D. D., Seidman, J. G. and Smith, J. A. (1988). CurrentProtocols in Molecular Biology. Wiley, New York.
Baudin, A., Ozier-Kalogeropoulos, O., Denouel, A.,Lacroute, F. and Cullin, C. (1993). A simple andefficient method for direct gene deletion in Sac-charomyces cerevisiae. Nucl. Acids Res. 21, 3329–3330.
Boeke, J. D., Trueheart, J., Natsoulis, G. and Fink,G. R. (1987). 5-fluoro-orotic acid as a selective agentin yeast molecular genetics. Meth. Enzymol. 154,164–175.
Brachmann, C. B., Davies, A., Cost, G. J., Caputo, E.,Li, J., Hieter, P. and Boeke, J. D. (1998). Designerdeletion strains derived from Saccharomyces cerevi-siae S288C: a useful set of strains and plasmidsfor PCR-mediated gene disruption and otherapplications. Yeast 14, 115–132.
Dolinski, K., Muir, S., Cardenas, M. and Heitman, J.(1997). All cyclophilins and FK506 binding proteinsare, individually and collectively, dispensable forviability in Saccharomyces cerevisiae. Proc. Natl.Acad. Sci. U S A 94, 13 093–13 098.
Geitz, R. D., Scheistl, R. H., Wellems, A. R. andWoods, R. A. (1995). Studies on the transformationof intact yeast cells by the LiAc/SS–DNA/PEGprocedure. Yeast 11, 355–360.
Langle-Rouault, F. and Jacobs, E. (1995). A method forperforming precise alterations in the yeast genomeusing a recyclable selectable marker. Nucleic AcidsRes. 23, 3079–3081.
Copyright ? 1999 John Wiley & Sons, Ltd.
Longtine, M. S., McKenzie, A., Demarini, A. J., Shah,N. G., Wach, A., Brachat, A., Phillippsen, P. andPringle, J. R. (1998). Additional modules for versatileand economical PCR-based gene deletion andmodification in Saccharomyces cerevisiae. Yeast 14,953–961.
Lorenz, M., Muir, R. S., Lim, E., McElver, J., Weber,S. C. and Heitman, J. (1995). Gene disruption withPCR products in Saccharomyces cerevisiae. Gene 158,113–117.
Niedenthal, R. K., Riles, L., Johnston, M. andHegemann, J. H. (1996). Green fluorescent proteinas a marker for gene expression and subcellularlocalization in budding yeast. Yeast 12, 773–786.
Oldenburg, K. R., Vo, K. T., Michaelis, S. and Paddon,C. (1997). Recombination-mediated PCR-directedplasmid construction in vivo in yeast. Nucleic AcidsRes. 25, 451–452.
Rose, M. D., Winston, F. and Hieter, P. (1989).Methods in Yeast Genetics. Cold Spring HarborLaboratory Press, New York.
Shoemaker, D. D., Lashkari, D. A., Morris, D.,Mittman, M. and Davis, R. W. (1996). Quantitativephenotypic analysis of yeast deletion mutants using ahighly parallel molecular bar-coding strategy. NatureGenet. 14, 450–456.
Sikorski, R. S. and Hieter, P. (1989). A system of shuttlevectors and yeast host strains designed for efficientmanipulation of DNA in Saccharomyces cerevisiae.Genetics 122, 19–27.
Steiner, S. and Phillipsen, P. (1994). Sequence andpromoter analysis of the highly expressed TEF gene ofthe filamentous fungus Ashbya gossypii. Mol. Gen.Genet. 242, 263–271.
Wach, A., Brachat, A., Alberti-Segui, C., Rebischung,C. and Philippsen, P. (1997). Heterologous HIS3marker and GFP reporter modules for PCR-targetingin Saccharomyces cerevisiae. Yeast 13, 1065–1075.
Wach, A., Brachat, A., Pohlmann, R. and Philippsen, P.(1994). New heterologous modules for classicalor PCR-based gene disruptions in Saccharomycescerevisiae. Yeast 10, 1793–1808.
Yeast 15, 507–511 (1999)