Embed Size (px)
Citation preview

Hydrobiologia 503: 59–67, 2003.M.B. Jones, A. Ingolfsson, E. Olafsson, G.V. Helgason, K. Gunnarsson &J. Svavarsson (eds), Migrations and Dispersal of Marine Organisms.© 2003 Kluwer Academic Publishers. Printed in the Netherlands.
59
Hitch-hiking on floating marine debris: macrobenthic species in theWestern Mediterranean Sea
Stefano Aliani1 & Anne Molcard1,2
1CNR, Istituto Scienze Marine - Sezione La Spezia, Forte Santa Teresa, 19036 Pozzuolo di Lerici (SP), Italy2 RSMAS-MPO University of Miami, 4600 Rickenbacker Causeway Miami, FL, 33149-1098, U.S.A.E-mail: [email protected]
Key words: dispersal, rafting, flotsam, benthos, Lagrangian model
Abstract
Marine litter has been defined as solid materials of human origin discarded at sea, or reaching the sea throughwaterways. The effect of marine debris on wildlife, tourism and human health is well documented and there isconsiderable scientific literature about plastic litter in the sea and over the seabed, mostly highlighting the possibleimpact on marine mammals and tourism. Dispersal of marine and terrestrial organisms on floating objects hasbiogeographical and ecological interest. For some species, extension of their geographical range is more likely tobe related to transport of mature individuals on floating rafts than to the active or passive dispersal of reproductivepropagules. Variability and variety of rafting materials has increased dramatically in recent years and marine litterhas been used widely as a raft by ‘hitch-hiking’ species. This paper reports on the benthic invertebrates living onmarine debris transported by wind and surface currents over the western Mediterranean Sea. Plastics accountedfor the major item of debris because of poor degradability, however glass, cans, fishing nets and polyurethanecontainers, were also found. Macro-benthos living on raft material comprised mainly molluscs, polychaetes andbryozoans. Large fish were found commonly below large plastic bags. Estimations of the distances that may becovered by hitch-hiking species and the contribution of rafting to the theoretical dispersal of species is provided.
Introduction
Typical marine invertebrates have life histories thatinclude at least one dispersal stage. Tropical andwarm-temperate species (e.g. >150 species of proso-branch gastropods) have a very long planktonic devel-opment and their teleplanic larvae provide evidence ofvery long range dispersal potential (Scheltema, 1988).However, there are exceptions to the traditional hy-pothesis ‘longer larval period – wider distribution’(Thorson, 1950; Mileikovski, 1971) and alternativemodes of dispersal have been considered (Zibrowius,1983; Scheltema, 1986). For example, for littoral gast-ropods along the east coast of North America, thereis no clear relationship between the length of de-velopment reported from laboratory experiments andgeographical range. Apparently, dispersal along con-tinental coastlines may be accomplished by stepwise
progression as long as there are no ecological barrierssuch as ocean basins (Scheltema, 1989).
Brooding species, or species with a short-livedlarval phase, may also be widely distributed and,for some of them (mainly living on hard substrata),extension of geographical range may be achievedthrough transport of mature individuals by floatingrafts, rather than through the active or passive dispersalof reproductive propagules (Highsmith, 1985). Theso-called ‘Rockall paradox’ is one example (Johan-nesson, 1988). Evidence from hydroids showed thatwide oceanic distances are covered more by raftinghydroids than by long-distance dispersal of planulae(Cornelius, 1992; Boero & Bouillon, 1993). Coloniesof the ascidian Botrylloides sp., attached to Zosteraleaves in California, were found 200 times furtheraway, and had greater recruitment success, comparedwith swimming larvae (Worchester, 1994).

60
Theories about the role of rafting in species distri-bution have become refined for several taxa living onhard substrata. Following Guppy’s (1917) monographon trans-Atlantic flotsam, rafting has been invoked as amain way of increasing the range of hydroids, (Ralph,1961) and remote populations of cnidarians (Cor-nelius, 1992). Jackson (1986) proposed that rafting isthe only reasonable explanation for the existence ofthe vast majority of (benthic) clonal species on Indo-Pacific oceanic islands and argued that, as medusal lifewas too short to disperse, species used flotsam (i.e.by rafting). Analysis of surface currents and reporteddrift patterns of pumice were used to estimate raftingof tropical corals (Jokiel, 1989); floating corals, dueto air trapped, wander the sea for months (DeVantier,1992).
The variety and availability of rafting material hasincreased dramatically in recent years with the spreadof human population (Barnes, 2002). Ship hulls, glass,plastic bottles and even items of footwear can be usedas rafts by hitch-hiking species. Prior to the spread ofhumans, available rafts were volcanic pumices, mac-roalgae, seagrasses, trees and seeds. Today marine lit-ter is also available, i.e. solid material of human originthat is discarded at sea, or reach the sea through wa-terways or domestic and industrial outfalls (NationalAcademy of Science, 1975 cited in Rees & Pond,1995). Monitoring marine debris has been undertakenfor several years (Rees & Pond, 1995), including as-sessment of the distribution and abundance of floatingplastic in the Gulf of Mexico by aerial surveys (Lecke-Mitchell & Mullin, 1992), and by ship observers inthe Eastern Mediterranean Sea (McCoy, 1988). Mostpapers focused mainly on the impact of debris on themarine environment in terms of pollution, aestheticdamage to tourism, or risks for turtles, cetaceans andfish.
In this paper, we identify the floating objects inthe Ligurian Sea from Corsica to the Ligurian Coast(Western Mediterranean) and we asses their potentialas ways of extending dispersal range of the benthic in-vertebrates living on them. Macrobenthic species wereidentified and the time needed for these benthic spe-cies to cover the distance from Corsica to LigurianCoast by hitch hiking on flotsam was inferred from anumerical simulation of a transport model.
Materials and methods
Study area
The Ligurian Sea is situated at the north eastern bor-der of the Western Mediterranean and is connected tothe southern basin (Tyrrhenian Sea) across the Cor-sica Channel. The eastern and northern borders are theTuscan and Ligurian coasts, and the western borderis open toward the Provençal region and the Gulf ofLions. The major large-scale feature of the deep andsurface layers of the Ligurian Sea is a cyclonic cir-culation active all year round, more intense in winterthan in summer. Climatic forcing can greatly changethe intensity of fluxes but the general pattern can beconsidered permanent (Astraldi & Gasparini, 1992).Southern waters, filling the Ligurian Sea, occur in twomain currents running along each side of northern Cor-sica. The West Corsica Current runs along the westernside of Corsica, and the warm and salty TyrrhenianCurrent passes trough the Corsica Channel betweenCapraia and Corsica (Artale et al., 1994). The twowaters merge to the north of Corsica and flow alongthe Ligurian coast toward the Gulf of Lions (Astraldiet al., 1995). Species living offshore in the Tyrrhenianbasin can be carried northward by the Tyrrhenian cur-rent across the Corsica Channel, providing a forcedpassage for the fauna. Changes in the water fluxesdue to climatic variability at this point can controlfaunal exchanges from the warm Tyrrhenian basin tothe colder Ligurian Sea (Astraldi et al., 1995; Aliani& Meloni, 1999).
The oceanographic cruise
Visual sightings of floating objects were made in theLigurian Sea from 10 to 15 July 1997 from the topdeck of the R/V Urania. The type and GPS position ofall objects on the sea surface during favourable lightand sea conditions while underway and at each sta-tion were recorded. Sightings were made close to theLigurian Coast (areas A,B,C, in Fig. 1), and close toCorsica and the coast of Tuscany (areas D,E,F, in Fig.1). Samples of floating objects were collected after15 min of searching by the ship’s tender. Immedi-ately after retrieval, samples were fixed in a bufferedformaldehyde-seawater solution (final concentration:5%) and the macrobenthic species identified in thelaboratory. Cluster analysis was performed on thepresence/absence species × samples matrix.
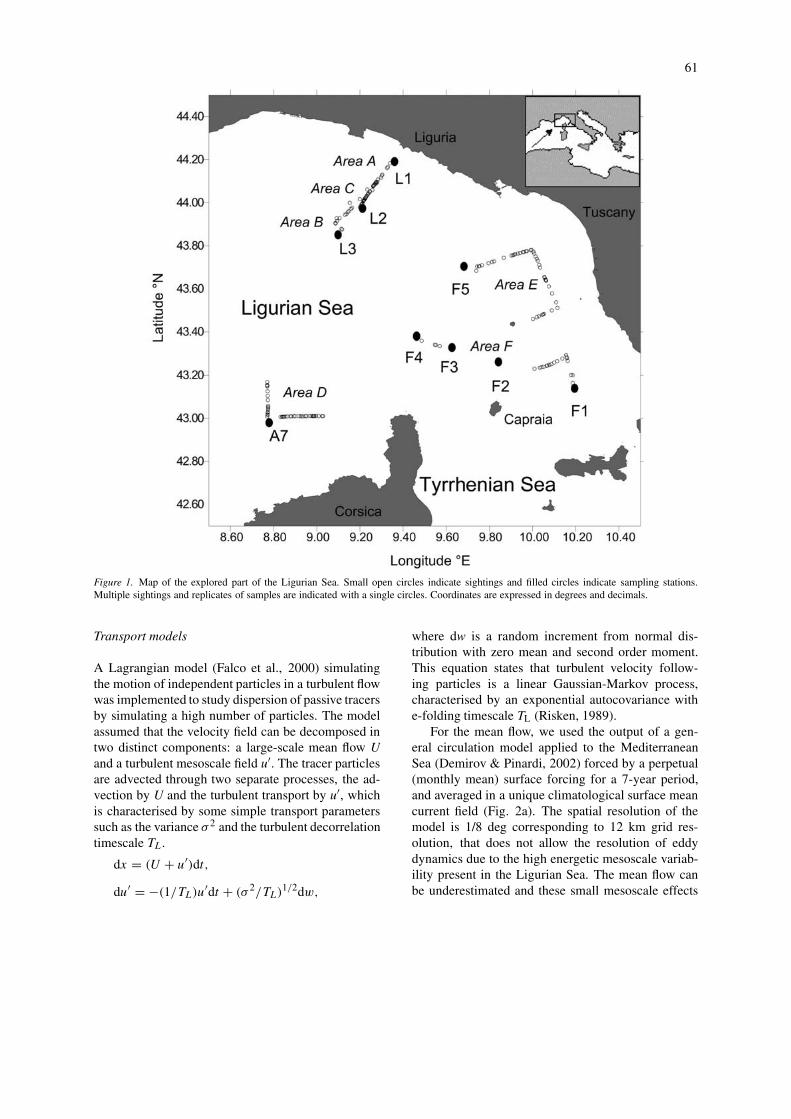
61
Figure 1. Map of the explored part of the Ligurian Sea. Small open circles indicate sightings and filled circles indicate sampling stations.Multiple sightings and replicates of samples are indicated with a single circles. Coordinates are expressed in degrees and decimals.
Transport models
A Lagrangian model (Falco et al., 2000) simulatingthe motion of independent particles in a turbulent flowwas implemented to study dispersion of passive tracersby simulating a high number of particles. The modelassumed that the velocity field can be decomposed intwo distinct components: a large-scale mean flow Uand a turbulent mesoscale field u′. The tracer particlesare advected through two separate processes, the ad-vection by U and the turbulent transport by u′, whichis characterised by some simple transport parameterssuch as the variance σ 2 and the turbulent decorrelationtimescale TL.
dx = (U + u′)dt,
du′ = −(1/TL)u′dt + (σ 2/TL)1/2dw,
where dw is a random increment from normal dis-tribution with zero mean and second order moment.This equation states that turbulent velocity follow-ing particles is a linear Gaussian-Markov process,characterised by an exponential autocovariance withe-folding timescale TL (Risken, 1989).
For the mean flow, we used the output of a gen-eral circulation model applied to the MediterraneanSea (Demirov & Pinardi, 2002) forced by a perpetual(monthly mean) surface forcing for a 7-year period,and averaged in a unique climatological surface meancurrent field (Fig. 2a). The spatial resolution of themodel is 1/8 deg corresponding to 12 km grid res-olution, that does not allow the resolution of eddydynamics due to the high energetic mesoscale variab-ility present in the Ligurian Sea. The mean flow canbe underestimated and these small mesoscale effects
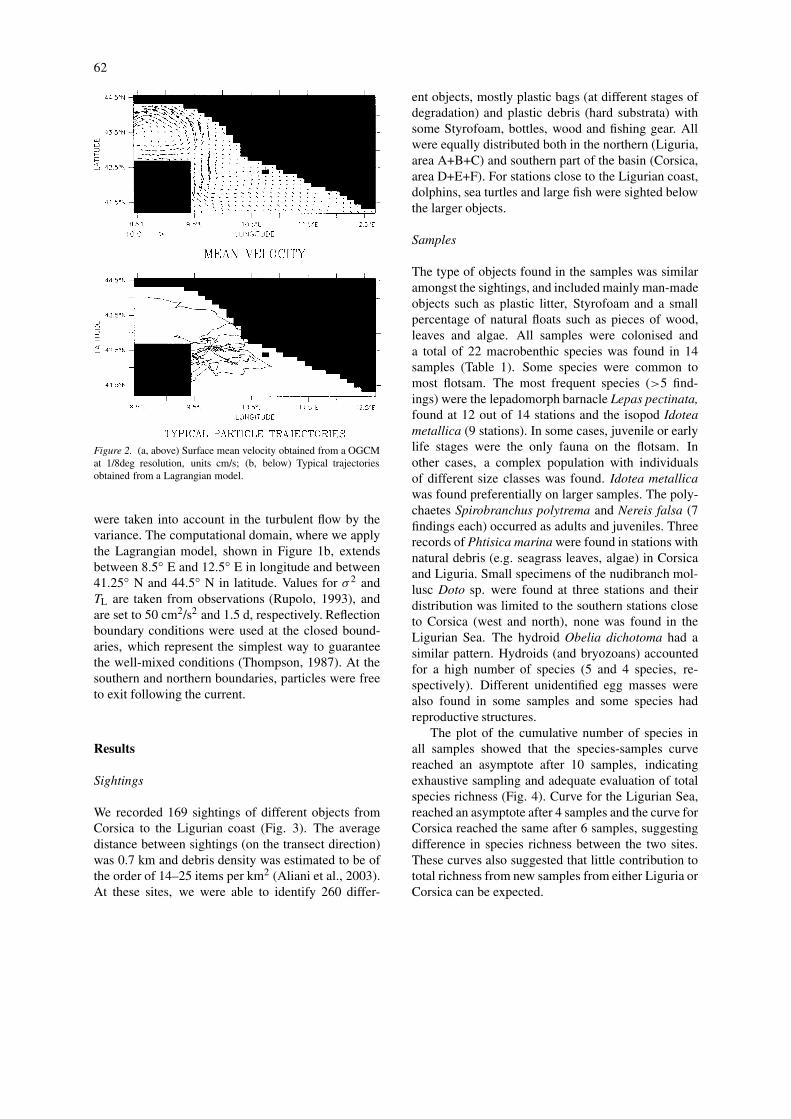
62
Figure 2. (a, above) Surface mean velocity obtained from a OGCMat 1/8deg resolution, units cm/s; (b, below) Typical trajectoriesobtained from a Lagrangian model.
were taken into account in the turbulent flow by thevariance. The computational domain, where we applythe Lagrangian model, shown in Figure 1b, extendsbetween 8.5◦ E and 12.5◦ E in longitude and between41.25◦ N and 44.5◦ N in latitude. Values for σ 2 andTL are taken from observations (Rupolo, 1993), andare set to 50 cm2/s2 and 1.5 d, respectively. Reflectionboundary conditions were used at the closed bound-aries, which represent the simplest way to guaranteethe well-mixed conditions (Thompson, 1987). At thesouthern and northern boundaries, particles were freeto exit following the current.
Results
Sightings
We recorded 169 sightings of different objects fromCorsica to the Ligurian coast (Fig. 3). The averagedistance between sightings (on the transect direction)was 0.7 km and debris density was estimated to be ofthe order of 14–25 items per km2 (Aliani et al., 2003).At these sites, we were able to identify 260 differ-
ent objects, mostly plastic bags (at different stages ofdegradation) and plastic debris (hard substrata) withsome Styrofoam, bottles, wood and fishing gear. Allwere equally distributed both in the northern (Liguria,area A+B+C) and southern part of the basin (Corsica,area D+E+F). For stations close to the Ligurian coast,dolphins, sea turtles and large fish were sighted belowthe larger objects.
Samples
The type of objects found in the samples was similaramongst the sightings, and included mainly man-madeobjects such as plastic litter, Styrofoam and a smallpercentage of natural floats such as pieces of wood,leaves and algae. All samples were colonised anda total of 22 macrobenthic species was found in 14samples (Table 1). Some species were common tomost flotsam. The most frequent species (>5 find-ings) were the lepadomorph barnacle Lepas pectinata,found at 12 out of 14 stations and the isopod Idoteametallica (9 stations). In some cases, juvenile or earlylife stages were the only fauna on the flotsam. Inother cases, a complex population with individualsof different size classes was found. Idotea metallicawas found preferentially on larger samples. The poly-chaetes Spirobranchus polytrema and Nereis falsa (7findings each) occurred as adults and juveniles. Threerecords of Phtisica marina were found in stations withnatural debris (e.g. seagrass leaves, algae) in Corsicaand Liguria. Small specimens of the nudibranch mol-lusc Doto sp. were found at three stations and theirdistribution was limited to the southern stations closeto Corsica (west and north), none was found in theLigurian Sea. The hydroid Obelia dichotoma had asimilar pattern. Hydroids (and bryozoans) accountedfor a high number of species (5 and 4 species, re-spectively). Different unidentified egg masses werealso found in some samples and some species hadreproductive structures.
The plot of the cumulative number of species inall samples showed that the species-samples curvereached an asymptote after 10 samples, indicatingexhaustive sampling and adequate evaluation of totalspecies richness (Fig. 4). Curve for the Ligurian Sea,reached an asymptote after 4 samples and the curve forCorsica reached the same after 6 samples, suggestingdifference in species richness between the two sites.These curves also suggested that little contribution tototal richness from new samples from either Liguria orCorsica can be expected.
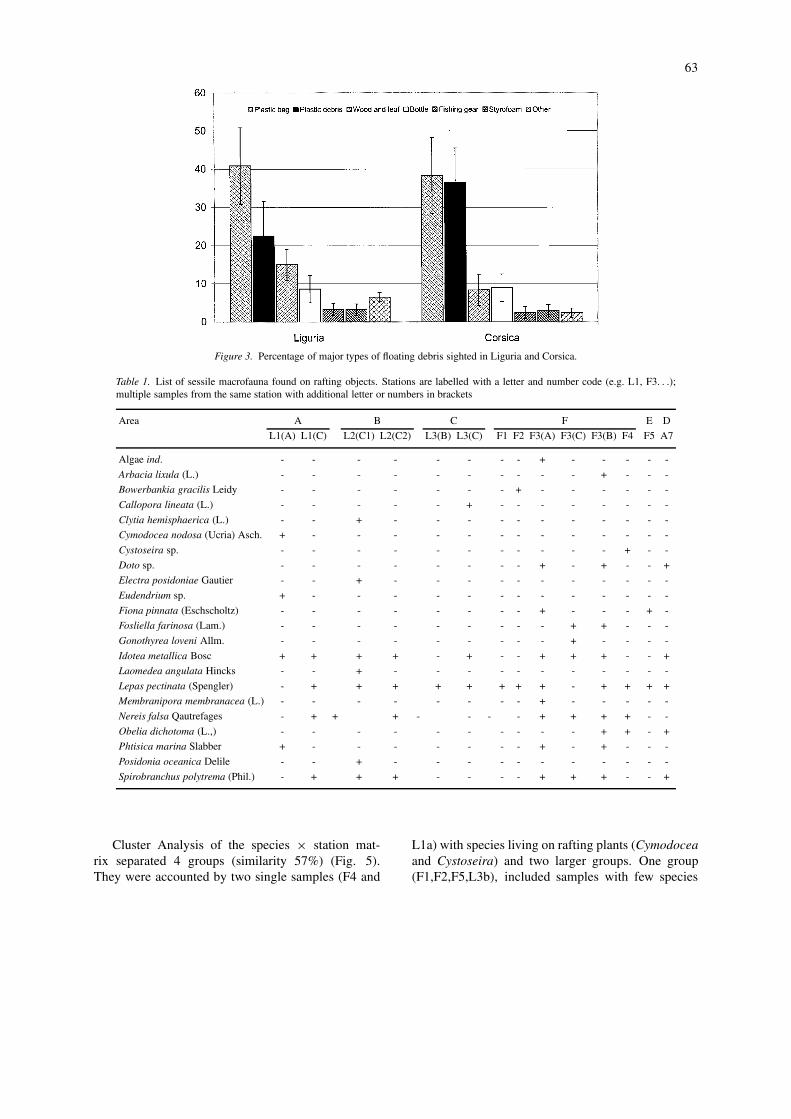
63
Figure 3. Percentage of major types of floating debris sighted in Liguria and Corsica.
Table 1. List of sessile macrofauna found on rafting objects. Stations are labelled with a letter and number code (e.g. L1, F3. . .);multiple samples from the same station with additional letter or numbers in brackets
Area A B C F E D
L1(A) L1(C) L2(C1) L2(C2) L3(B) L3(C) F1 F2 F3(A) F3(C) F3(B) F4 F5 A7
Algae ind. - - - - - - - - + - - - - -
Arbacia lixula (L.) - - - - - - - - - - + - - -
Bowerbankia gracilis Leidy - - - - - - - + - - - - - -
Callopora lineata (L.) - - - - - + - - - - - - - -
Clytia hemisphaerica (L.) - - + - - - - - - - - - - -
Cymodocea nodosa (Ucria) Asch. + - - - - - - - - - - - - -
Cystoseira sp. - - - - - - - - - - - + - -
Doto sp. - - - - - - - - + - + - - +
Electra posidoniae Gautier - - + - - - - - - - - - - -
Eudendrium sp. + - - - - - - - - - - - - -
Fiona pinnata (Eschscholtz) - - - - - - - - + - - - + -
Fosliella farinosa (Lam.) - - - - - - - - - + + - - -
Gonothyrea loveni Allm. - - - - - - - - - + - - - -
Idotea metallica Bosc + + + + - + - - + + + - - +
Laomedea angulata Hincks - - + - - - - - - - - - - -
Lepas pectinata (Spengler) - + + + + + + + + - + + + +
Membranipora membranacea (L.) - - - - - - - - + - - - - -
Nereis falsa Qautrefages - + + + - - - - + + + + - -
Obelia dichotoma (L.,) - - - - - - - - - - + + - +
Phtisica marina Slabber + - - - - - - - + - + - - -
Posidonia oceanica Delile - - + - - - - - - - - - - -
Spirobranchus polytrema (Phil.) - + + + - - - - + + + - - +
Cluster Analysis of the species × station mat-rix separated 4 groups (similarity 57%) (Fig. 5).They were accounted by two single samples (F4 and
L1a) with species living on rafting plants (Cymodoceaand Cystoseira) and two larger groups. One group(F1,F2,F5,L3b), included samples with few species
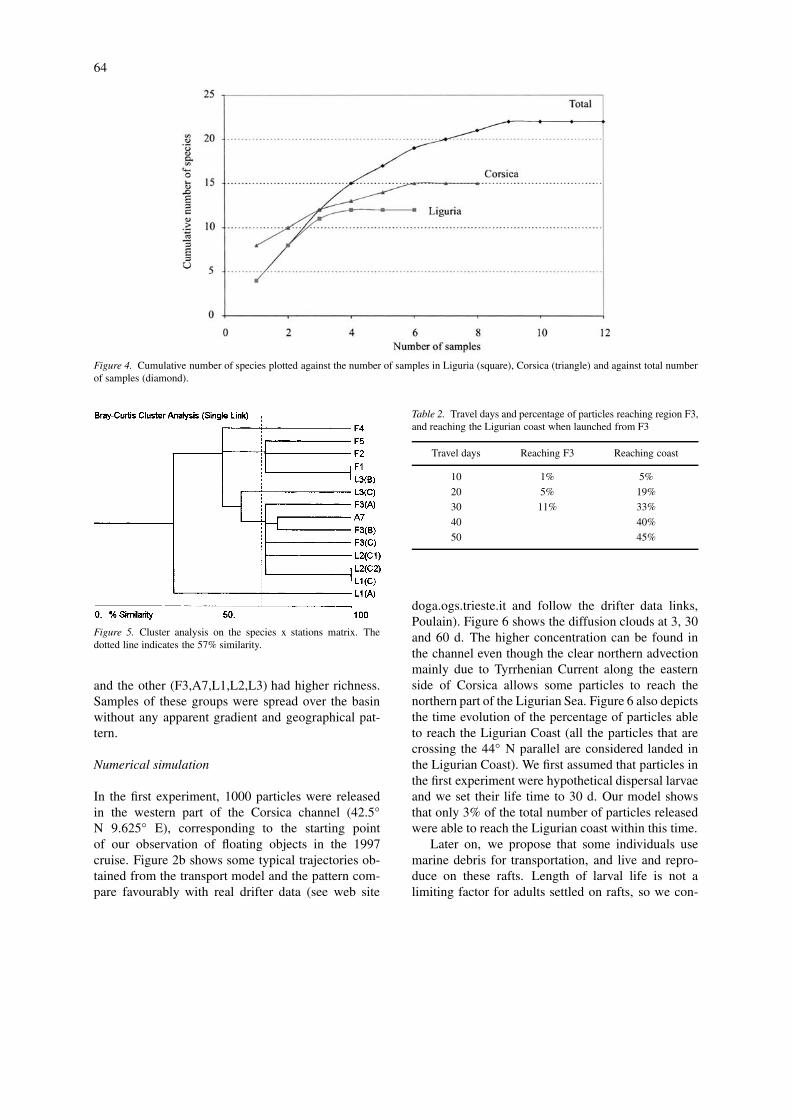
64
Figure 4. Cumulative number of species plotted against the number of samples in Liguria (square), Corsica (triangle) and against total numberof samples (diamond).
Figure 5. Cluster analysis on the species x stations matrix. Thedotted line indicates the 57% similarity.
and the other (F3,A7,L1,L2,L3) had higher richness.Samples of these groups were spread over the basinwithout any apparent gradient and geographical pat-tern.
Numerical simulation
In the first experiment, 1000 particles were releasedin the western part of the Corsica channel (42.5◦N 9.625◦ E), corresponding to the starting pointof our observation of floating objects in the 1997cruise. Figure 2b shows some typical trajectories ob-tained from the transport model and the pattern com-pare favourably with real drifter data (see web site
Table 2. Travel days and percentage of particles reaching region F3,and reaching the Ligurian coast when launched from F3
Travel days Reaching F3 Reaching coast
10 1% 5%
20 5% 19%
30 11% 33%
40 40%
50 45%
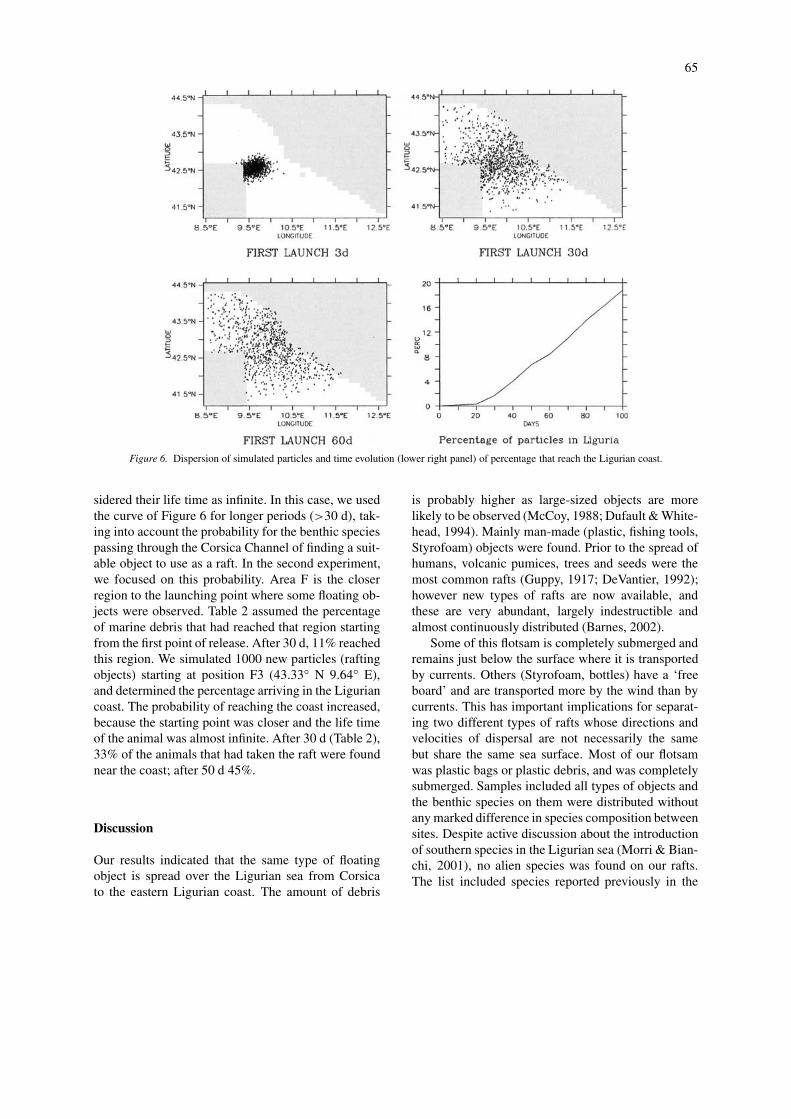
doga.ogs.trieste.it and follow the drifter data links,Poulain). Figure 6 shows the diffusion clouds at 3, 30and 60 d. The higher concentration can be found inthe channel even though the clear northern advectionmainly due to Tyrrhenian Current along the easternside of Corsica allows some particles to reach thenorthern part of the Ligurian Sea. Figure 6 also depictsthe time evolution of the percentage of particles ableto reach the Ligurian Coast (all the particles that arecrossing the 44◦ N parallel are considered landed inthe Ligurian Coast). We first assumed that particles inthe first experiment were hypothetical dispersal larvaeand we set their life time to 30 d. Our model showsthat only 3% of the total number of particles releasedwere able to reach the Ligurian coast within this time.
Later on, we propose that some individuals usemarine debris for transportation, and live and repro-duce on these rafts. Length of larval life is not alimiting factor for adults settled on rafts, so we con-
65
Figure 6. Dispersion of simulated particles and time evolution (lower right panel) of percentage that reach the Ligurian coast.
sidered their life time as infinite. In this case, we usedthe curve of Figure 6 for longer periods (>30 d), tak-ing into account the probability for the benthic speciespassing through the Corsica Channel of finding a suit-able object to use as a raft. In the second experiment,we focused on this probability. Area F is the closerregion to the launching point where some floating ob-jects were observed. Table 2 assumed the percentageof marine debris that had reached that region startingfrom the first point of release. After 30 d, 11% reachedthis region. We simulated 1000 new particles (raftingobjects) starting at position F3 (43.33◦ N 9.64◦ E),and determined the percentage arriving in the Liguriancoast. The probability of reaching the coast increased,because the starting point was closer and the life timeof the animal was almost infinite. After 30 d (Table 2),33% of the animals that had taken the raft were foundnear the coast; after 50 d 45%.
Discussion
Our results indicated that the same type of floatingobject is spread over the Ligurian sea from Corsicato the eastern Ligurian coast. The amount of debris
is probably higher as large-sized objects are morelikely to be observed (McCoy, 1988; Dufault & White-head, 1994). Mainly man-made (plastic, fishing tools,Styrofoam) objects were found. Prior to the spread ofhumans, volcanic pumices, trees and seeds were themost common rafts (Guppy, 1917; DeVantier, 1992);however new types of rafts are now available, andthese are very abundant, largely indestructible andalmost continuously distributed (Barnes, 2002).
Some of this flotsam is completely submerged andremains just below the surface where it is transportedby currents. Others (Styrofoam, bottles) have a ‘freeboard’ and are transported more by the wind than bycurrents. This has important implications for separat-ing two different types of rafts whose directions andvelocities of dispersal are not necessarily the samebut share the same sea surface. Most of our flotsamwas plastic bags or plastic debris, and was completelysubmerged. Samples included all types of objects andthe benthic species on them were distributed withoutany marked difference in species composition betweensites. Despite active discussion about the introductionof southern species in the Ligurian sea (Morri & Bian-chi, 2001), no alien species was found on our rafts.The list included species reported previously in the

66
Mediterranean Sea as offshore fouling species (BellanSantini,1970; Relini 1976; Relini et al., 1977; Aliani& Meloni, 1999; Relini et al., 2000). Most raftedspecies may be included in the ‘list of superwander-ers’ (Cornelius, 1992; Aliani et al., 1998) that userafts as their major means of dispersion (Cornelius,1992). As Barnes (2002) pointed out, the distancefrom the mainland does not seem to influence theproportion of debris colonised and these species arewidely distributed offshore in the Mediterranean Sea.
In contrast to these common widespread species,we encountered also single individuals. For these spe-cies, hitch hiking on rafts may offer a final opportunityto survive for a larvae close to the end of its planktonicperiod (Toonen & Pawlik, 1994) or just an alternativeartificial substratum (Tursi et al., 1985). Their contri-bution to global biodiversity is not well understood.Another group of species, separated by Cluster Ana-lysis, included those found rarely on plastic objectsbut commonly on seagrass leaves and on wood. Theyare typically reported as epibionts of seagrass and wefound them as reproductive adults much further thanthe distance covered by short-living larvae. Rafting onleaves is a very effective means of dispersal for thesespecies.
Species can jump to new rafts in the open seaas some unidentified eggs found on the rafts weretoo young to suggest a long dispersal history. Ac-cumulation of floating objects in strips by Langmuircirculation (Nimmo Smith & Thorpe, 1999) may helpin transferring to a new raft or exchanges of gametesbetween close rafts. A more complex community willprobably evolve from the unidentified eggs and sea-sonal fluctuations in community composition may alsooccur.
Our numerical experiments showed that there is alimited chance for benthic invertebrates to travel longdistances alone without mortality due to ecologicalconstraints (Cowen et al., 2000). Using floating ob-jects as a means of transportation (where they can alsoreproduce), increases their probability of survival. Asplastic litter is everywhere, the probability of finding araft is higher than hypothezised.
In conclusion, a continuum of species and raftsexist in the Ligurian sea. Rafts have different agesaccording to the dispersal processes that brought themoffshore or according to recruitment processes occur-ring in far or nearby rafts. For these individuals, raftingobjects extend their potential dispersal range beyondtheir theoretical extension.
Acknowledgements
We thank Annalisa Griffa for useful discussion andsuggestions, and Encho Demirov for supplying theOGCM output. We are also grateful to S. Geraci,coordinator of the cruise, and to M. Faimali, F. Gara-venta, M. Andrenacci, E. Canepa, E. Schiano forvaluable help in the field work. Special thanks to theCaptain and the crew of the R/V Urania. This paperforms part of the framework of the Italian ResearchProject SINAPSI. We thank two anonymous refer-ees for greatly improving the manuscript with theirsuggestions.
References
Aliani, S., C. De Asmundis, R. Meloni, M. Borghini & G. P.Gasparini, 1998. Transport of benthic species in the Sicilychannel: preliminary observations. In Piccazzo, M. (ed.), AttiXII Congresso Associazione Italiana Oceanologia e Limnologia:173–182.
Aliani, S. & R. Meloni, 1999. Dispersal strategies of benthic speciesand water current variability in the Corsica Channel (WesternMediterranean) Sci. mar. 63: 137–145.
Aliani, S., A. Griffa & A.Molcard, 2003. Floating debris in theLigurian Sea, North Western Mediterranean. Mar. Poll. Bull. 46:1142–1149.
Artale, V., M. Astraldi, G. Buffoni & G. P. Gasparini, 1994. Sea-sonal variability of the gyre-scale circulation in the NorthernTyrrhenian sea. J. Geophys. Res. 99: 14127–14137.
Astraldi, M. & G. P. Gasparini, 1992. The seasonal characteristicsof the circulation in the North Mediterranean Basin and theirrelationships with atmospheric climatic conditions J. Geophys.Res. 97: 9531–9540.
Astraldi, M., C. N. Bianchi, G. P. Gasparini & C. Morri, 1995.Climatic fluctuations, current variability and marine species dis-tribution: a case study in the Ligurian Sea (North WesternMediterranean). Oceanol. Acta 18: 139–149.
Barnes, D. K. A., 2002. Invasions by marine life on plastic debris.Nature 416: 808–809.
Bellan Santini, D., 1970. Salissures biologiques de substrats viergesartificiels immergés en eau pure, durant 26 mois, dans la règionde Marseille Mediterranée Nord occidentale) I. Etude qualitative.Tethys 2: 335–356.
Boero, F. & J. Bouillon, 1993. Zoogeography and life cycle patternsof Mediterranean hydromedusae (Cnidaria). Biol. J. linn. Soc.48: 239–266.
Cornelius, P. F., 1992. Medusa loss in leptolid Hydrozoa (Cnidaria),hydroid rafting and abbreviated life cycles among their remote-island faunae: an interim review. Sci. mar. 56: 245–261.
Cowen, R. K., K. M. M. Lwiza, S. Sponaugle, C. B. Paris & D.B. Olson, 2000. Connectivity of marine populations: open orclosed? Science 287: 857–859.
Demirov, E. & Pinardi, N., 2002. Simulation of the MediterraneanSea circulation from 1979 to 1993: Part I. The interannualvariability. J. mar. Syst. (33–34): 23–50.
De Vantier, L. M., 1992. Rafting of tropical marine organisms onbuoyant coralla. Mar Ecol. Prog. Ser. 86: 301–302.

67
Dufault, S. & H. Whitehead, 1994. Floating marine pollution in “theGully’ on the continental slope, Nova Scotia, Canada. Mar. Poll.Bull. 28: 489–493.
Falco, P., A. Griffa, P. M. Poulain & E. Zambianchi, 2000. Transportproperties in the Adriatic Sea as deduced from drifter data. J.Phys. Oceanogr. 30: 2055–2071.
Guppy, H. B., 1917. Plants, seeds and currents in the West Indiesand Azores. Williams and Norgate. London 1: 531.
Highsmith, R. C., 1985. Floating and algal rafting as potential dis-persal mechanisms in brooding invertebrates. Mar. Ecol. Prog.Ser. 25: 169–179.
Jackson, J. B. C., 1986. Modes of dispersal of clonal benthic in-vertebrates: consequences for species’ distribution and geneticstructure of local populations. Bull. mar. Sci. 39: 588–606.
Johannesson, K., 1988. The paradox of Rockhall, why is a broodinggastropod (Littorina saxatilis) more widespread than one havinga planktonic larval dispersal stage (L. littorea)? Mar. Biol. 99:507–513.
Jokiel, P. L., 1989. Rafting of reef corals and other organisms atKwajalein Atoll. Mar. Biol. 101: 483–493.
Lecke-Mitchell, K. & K. Mullin, 1992. Distribution and abund-ance of large floating marine plastic in the north-central gulf ofMexico. Mar. Poll. Bull. 24: 598–601.
Mc Coy, F., 1988. Floating megalitter in the Eastern Mediterranean.Mar. Poll. Bull. 19: 25–28.
Mileikovski, S. A., 1971. Types of larval development in marinebottom invertebrates, their distribution and ecological signific-ance, a re-evaluation. Mar. Biol. 10: 193–213.
Morri, C. & C. N. Bianchi, 2001. Recent changes in biodiversityin the Ligurian Sea (NW Mediterranean): is there a climaticforcing? In Faranda, F. M., L. Guglielmo & G. Spezie (eds),Mediterranean Ecosystems: Structures and Processes. SpringerVerlag: 375–385.
Nimmo Smith, W. A. M. & S. A. Thorpe, 1999. Dispersion of buoy-ant material by Langmuir circulation and a tidal current. Mar.Poll. Bull. 9: 824–829.
Ralph, P. M., 1961. New Zealand Thecate hydroids. Part V. Thedistribution of the New Zealand Thecate hydroids. Trans. Roy.Soc. N. Z. (Zool. ser.). 1: 103–111.
Rees, G. & K. Pond, 1995. Marine litter monitoring programmes –a review of methods with special reference to national surveys.Mar. Poll. Bull. 30: 103–108.
Relini, G., 1976. Fouling on different material immersed at a depthof 200 m in the Ligurian Sea. Proc. 4th Int. Congr. On MarineCorrosion. Juan les Pins: 431–443.
Relini, G., C. N. Bianchi, G. Diviacco & R. Rosso, 1977. Foulingdi alcune piattaforme offshore dei mari italiani VI: Anfipodi ePolicheti. Boll. Mus. Ist. Biol. Univ. Genova 45: 105–121.
Relini, G., M. Relini & M. Montanari, 2000. An offshore buoy as asmall artificial island and a fish-aggregating device (FAD) in theMediterranean. Hydrobiologia 40: 65–80.
Risken, H., 1989. The Fokker-Planck Equation: Methods of Solu-tions and Applications. Springer-Verlag 1: 472.
Rupolo, V., 1993. Studio delle caratteristiche principali della cir-colazione del mar Tirreno attraverso l’analisi di dati lagrangianie la formulazione di un modello. Tesi di Laurea Università LaSapienza, Roma.
Scheltema, R. S., 1986. Alternative modes of dispersal. Bull. mar.Sci. 39: 310–312.
Scheltema, R. S., 1988. Initial evidence for the transport of tele-planic larvae of benthic invertebrates across the East Pacificbarrier. Biol. Bull. 174: 145–152.
Scheltema, R. S., 1989. In Reproduction, genetics and distributionof marine organisms. In Ryland, J. S. & P. A. Tyler (eds), Olsenand Olsen Holdstedvj, Denmark: 186–187.
Thompson, D. J., 1987: Criteria for the selection of stochastic mod-els of particle trajectories in turbulent flows. J. Fluid Mech. 180:529–556.
Thorson, G., 1950. Reproduction and larval ecology of marinebottom invertebrates. Biol. Rev. 25: 1–45.
Toonen, R. J. & J. R. Pawlik, 1994. Foundations of gregariousness.Nature 370: 511–512.
Tursi, A., A. Matarrese, L. Scalera Liaci, G. Costantino, R. Cavallo& E. Cecere, 1985. Colonizzazione di substrati duri artificiali im-mersi in una biocenosi coralligena ed in un posidonieto. Oebalia11: 401–416.
Worchester, S. E., 1994. Adults vs larval swimming: dispersal andrecruitment of a botryllid ascidian on eelgrass. Mar. Biol. 121:309–317.
Zibrowius, H., 1983. Extension de l’aire de repartition favorisee parl’homme chez les invertebres marins. Oceanis 9: 337–353.





























