Embed Size (px)
Citation preview

Homogenization in Cardiac Electrophysiology
andBlow-Up in Bacterial Chemotaxis
by
Paul Earl Hand
A dissertation submitted in partial fulfillment
of the requirements for the degree of
Doctor of Philosophy
Department of Mathematics
New York University
May 2009
Professor Charles Peskin
Professor Nader Masmoudi

c© Paul Earl Hand
All Rights Reserved, 2009


Acknowledgements
I am indebted to many people who made this dissertation possible. First andforemost, I would like to thank my mother, father, and the rest of my familyfor the support and sacrifices they made to allow me to pursue my interests inmathematics. I would also like to thank my advisors, Charlie Peskin and NaderMasmoudi, for giving me the freedom and guidance to pursue these interests asthey developed.
I gratefully acknowledge Boyce Griffith, Yoichiro Mori, Glenn Fishmann, andGreg Morley for many discussions about mathematical and experimental cardiacelectrophysiology.
I am thankful for several years of funding by the United States Department ofDefense through a National Defense Science and Engineering Graduate Fellowship.
Finally, I would like to thank many of my friends for their help and distraction,including Ben Olsen, Saverio Spagnolie, Will Findley, Mike Damron, Murphy Stein,Giulio Trigila, Alex Rubinsteyn, Al Momin, Ross Tulloch, Dan Goldberg, TomAlberts, Jeff Ryan, Kela Lushi, Shilpa Khatri, Jens Jørgensen, and Thomas Fai.
iv

Abstract
In the first part of this dissertation, we investigate three different issues involv-ing homogenization in cardiac electrophysiology.
We present a modification for how heart tissue is typically modeled in orderto derive values for intracellular and extracellular conductivities needed for bido-main simulations. In our model, cardiac myocytes are rectangular prisms and gapjunctions appear in a distributed manner as flux boundary conditions for Laplace’sequation. In other models, gap junctions tend to be explicit geometrical entities.Using directly measurable microproperties such as cellular dimensions and end-to-end and side-to-side gap junction coupling strengths, we inexpensively obtaineffective conductivities close to those given by simulations with a detailed cyto-architecture. This model provides a convenient framework for studying the effecton conductivities of aligned vs. brick-like arrangements of cells and the effect ofdifferent distributions of gap junctions between the sides and ends of myocytes.
We further illustrate this framework by investigating the effect on conductivityof non-uniform distributions of gap junctions within the ends of cells. We showthat uniform distributions are local maximizers of conductivity through analyticalperturbation arguments.
We also derive a homogenized description of an ephaptic communication mech-anism along a single strand of cells. We perform numerical simulations of the fullmodel and its homogenization. We observe that the two descriptions agree whengap junctional coupling is at physiologically normal levels. When gap junctionalcoupling is low, the homogenized description does not capture the behavior thatthe ephaptic mechanism can speed up action potential propagation.
In the second part of this dissertation, we investigate finite-time blow-up andstability of the Keller-Segel model for bacterial chemotaxis. We use a secondmoment calculation to establish finite-time blow-up for the Keller-Segel system ona disk with Dirichlet boundary conditions and a supercritical mass.
We numerically investigate the evolution and stability of the Keller-Segel sys-tem in order to provide a conjecture about the generality of boundary blow-up forsupercritical mass under the Jager-Luckhaus boundary conditions.
Finally, we use the free energy of solutions to Keller-Segel equations to derivea functional inequality that may be helpful for analyzing the stability of steadystates.
v

Contents
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ivAbstract . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . vList of Figures . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . xiiiList of Tables . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . xv
I Homogenization in Cardiac Electrophysiology 1
1 Introduction 2
1.1 Cellular Biophysics . . . . . . . . . . . . . . . . . . . . . . . . . . . 21.1.1 The Hodgkin-Huxley Ionic Model . . . . . . . . . . . . . . . 31.1.2 The Luo-Rudy Ionic Model . . . . . . . . . . . . . . . . . . 4
1.2 The Cable Equation . . . . . . . . . . . . . . . . . . . . . . . . . . 71.3 The Bidomain Equations . . . . . . . . . . . . . . . . . . . . . . . . 81.4 Homogenization of Partial Differential Equations . . . . . . . . . . . 91.5 Outline of Part I . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
2 Homogenization of Cardiac Models that Describe Gap Junctions
Through Boundary Conditions 13
2.1 Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 132.2 Bidomain Equations and Effective Conductivity . . . . . . . . . . . 152.3 Full Cellular Model in an Aligned Arrangement . . . . . . . . . . . 16
2.3.1 Nondimensionlization . . . . . . . . . . . . . . . . . . . . . . 182.3.2 Statement of the Effective Conductivity Problem . . . . . . 18
2.4 Homogenization in an Aligned Arrangement . . . . . . . . . . . . . 192.4.1 Analytical Solution to the Corrector Problem . . . . . . . . 212.4.2 Resulting Effective Conductivities . . . . . . . . . . . . . . . 222.4.3 Equivalent Resistor Network . . . . . . . . . . . . . . . . . . 22
2.5 Full Cellular Model and Homogenization in a Brick-like Arrangement 232.5.1 Resulting Effective Conductivities . . . . . . . . . . . . . . . 25
2.6 Discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 272.6.1 Effective Conductivity Values . . . . . . . . . . . . . . . . . 27
vi

2.6.2 Comparison of PDE and Resistor Network Methods . . . . . 292.6.3 Application to Electromechanical Simulations . . . . . . . . 30
2.7 Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 302.8 Extracellular Conductivities . . . . . . . . . . . . . . . . . . . . . . 312.9 Parameters and Variables . . . . . . . . . . . . . . . . . . . . . . . 33
3 Gap Junction Distributions for Optimal Effective Conductivity 35
3.1 Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 353.2 Model and Statement of the Gap Junction Distribution Problem . . 38
3.2.1 Nondimensionlization and Homogenization . . . . . . . . . . 393.2.2 Statement of the Gap Junction Distribution Problem . . . . 41
3.3 Local optimality of a uniform gap junctional distribution . . . . . . 413.4 Discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44
4 Homogenization of a Model for Ephaptic Cardiac Communication 45
4.1 Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 454.2 The Full Ephaptic Model . . . . . . . . . . . . . . . . . . . . . . . . 47
4.2.1 Nondimensionalization of the Full System . . . . . . . . . . 484.3 The Homogenized Ephaptic Model . . . . . . . . . . . . . . . . . . 50
4.3.1 Derivation of Homogenized System . . . . . . . . . . . . . . 514.4 Numerical Simulation of the Full and Homogenized Models . . . . . 52
4.4.1 Initial Value Problems for Numerical Simulation . . . . . . . 534.4.2 Numerical Results for the Full System . . . . . . . . . . . . 574.4.3 Numerical Results for the Homogenized System . . . . . . . 59
4.5 Discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 614.6 Simulation Parameters . . . . . . . . . . . . . . . . . . . . . . . . . 654.7 Numerical Schemes . . . . . . . . . . . . . . . . . . . . . . . . . . . 65
4.7.1 Full System . . . . . . . . . . . . . . . . . . . . . . . . . . . 654.7.2 Homogenized System . . . . . . . . . . . . . . . . . . . . . . 70
II Blow-Up in Bacterial Chemotaxis 73
5 Introduction 74
5.1 Derivation of Keller-Segel Equations . . . . . . . . . . . . . . . . . 745.2 Finite-Time Blow-Up in the Whole Plane . . . . . . . . . . . . . . . 765.3 Free Energy for Keller-Segel Systems . . . . . . . . . . . . . . . . . 775.4 Outline of Part II . . . . . . . . . . . . . . . . . . . . . . . . . . . . 78
6 Finite-Time Blow-Up of Keller-Segel on a Disk with Dirichlet
Boundary Conditions 79
6.1 Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 79
vii

6.2 Finite-Time Blow-Up under Dirichlet Boundary Conditions for Su-percritical Mass . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 80
7 Numerically Motivated Conjecture on Boundary Blow-Up with
Jager-Luckhaus Boundary Conditions 84
7.1 Introduction and Blow-Up Conjecture . . . . . . . . . . . . . . . . . 847.1.1 Outline of Chapter . . . . . . . . . . . . . . . . . . . . . . . 86
7.2 Numerical Simulation of Keller-Segel Evolution . . . . . . . . . . . 877.2.1 Initial Value Problem . . . . . . . . . . . . . . . . . . . . . . 877.2.2 Numerical Scheme . . . . . . . . . . . . . . . . . . . . . . . 877.2.3 Results . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 89
7.3 Numerical Stability of Keller-Segel Steady States . . . . . . . . . . 897.3.1 Discretization . . . . . . . . . . . . . . . . . . . . . . . . . . 927.3.2 Method . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 937.3.3 Results for Numerical Stability Analysis . . . . . . . . . . . 93
7.4 Discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 937.5 Convergence Study for Evolution Simulation . . . . . . . . . . . . . 95
7.5.1 Spatial Convergence Study . . . . . . . . . . . . . . . . . . . 957.5.2 Temporal Convergence Study . . . . . . . . . . . . . . . . . 96
8 A Free Energy Stability Criterion 98
8.1 Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 988.2 Free Energy Inequality . . . . . . . . . . . . . . . . . . . . . . . . . 99
8.2.1 Motivation and Derivation . . . . . . . . . . . . . . . . . . . 998.3 Application and Discussion . . . . . . . . . . . . . . . . . . . . . . . 100
8.3.1 Stability of Uniform Profiles . . . . . . . . . . . . . . . . . . 1008.3.2 Differences with Linear Stability . . . . . . . . . . . . . . . . 1018.3.3 Does the Free Energy Inequality Aid Analysis? . . . . . . . . 101
Bibliography 102
viii

List of Figures
1.1 The electrical circuit model of an isopotential cell (shaded). Anycurrent injected inside would either charge the plasma membrane asa capacitor or flow across the membrane through ion channels. . . . 2
1.2 A cable of electrically active membrane filled with conducting fluid.A flux balance calculation allows for a derivation of the cable equation. 7
1.3 A depiction of the complicated geometrical structure of the intra-cellular and extracellular space of cardiac tissue. The thick, darkvertical stripes separating cells are known as intercalated disks.Reprinted with permission from Guyton and Hall (Fig. 9-2, p. 108). 8
1.4 In this section, we homogenize Laplace’s equation over the periodicdomain Y ε (a). This domain is composed of translated and scaledversions of the unit cell Y (b). . . . . . . . . . . . . . . . . . . . . . 10
2.1 Instead of modeling gap junctions through complex cellular geome-try (a), we model them through flux boundary conditions on simplegeometry (b). The multiple resistors shown in (b) represent a con-tinuous boundary condition, see equations (2.2) - (2.7). . . . . . . . 15
2.2 The aligned cellular architecture: (a) cells of dimension l×wc ×wc
are arranged in three space with period l × wp × wp; (b) an x1, x2
cross-section. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 162.3 In order to compute effective conductivity, we impose a potential
difference across a large number of cells and calculate the result-ing current density (a). In this figure, extracellular space has beenomitted for clarity. The resulting electric potential is depicted in (b). 19
2.4 The unit cell in the aligned geometry is composed of an intracellularregion Yi and an extracellular region Ye. . . . . . . . . . . . . . . . . 21
2.5 The brick-like cellular architecture. Cells of dimension l × wc × wc
are arranged with a period of l×wp×wp, except that adjacent fibersare offset by half a cell length (a). The arrangement can be viewedas the periodic extension of the region inside the dashed prism. Anx1, x2 cross-section is shown in (b). . . . . . . . . . . . . . . . . . . 23
ix

2.6 Electric potential in several cells in a brick-like arrangement with alongitudinally applied potential gradient using cellular parametersfrom Table 2.4. The second and fourth cells from the left are offsetby half a cell length because they are in a neighboring fiber to thefirst and third cells. The units of the vertical axis are arbitrary. Asthe cellular domain is three dimensional, the plot shows only thex3-averaged potential within a cell. . . . . . . . . . . . . . . . . . . 25
2.7 Effective intracellular transverse and longitudinal conductivities asa function of the fraction of gap junctions expressed on the cell sides,keeping the total conductance constant. As gap junctions are movedfrom the ends to the sides of a cell, intracellular longitudinal con-ductivity decreases (a). With an aligned arrangement it decreases tozero, but with the brick arrangement there is a nonzero intracellu-lar longitudinal conductivity even when all gap junctions are on thesides of the cells. Meanwhile, in the parameter regime given by cel-lular measurements, intracellular transverse conductivity increaseslinearly with the fraction of gap junctions on the cell sides (b). Theside-to-side and end-to-end conductance measurements given in Ta-ble 2.4 correspond to a fraction 0.68 of gap junctions on the sidesof cells. A uniform density of gap junctions between the sides andends of cells with dimensions given in Table 2.4 corresponds to afraction 0.92 of gap junctions on the sides of cells. . . . . . . . . . . 26
2.8 In this cross-section of tissue, the longitudinal direction points outof the page. The dark shaded regions indicate cells, and the remain-ing regions indicate extracellular space. We treat the light shadedregions as insulators for the calculation of transverse conductivityin the x2 direction. . . . . . . . . . . . . . . . . . . . . . . . . . . . 32
3.1 A simplified, two-dimensional depiction of the three-dimensional,two cell model of [8]. Each cell is represented as a cubic lattice.Gap junctions are represented by lattice points connecting the twocells. The gap junctions can be arranged as a single placque ofadjacent vertices (a) or can be scattered randomly (b). . . . . . . . 36
3.2 The cellular model of [29]. Cells in a row are modeled as rectangularregions. Gap junctions are represented as holes in the boundariesbetween them, making intracellular space contiguous. Keeping thetotal length of the gap junctions fixed, few large holes correspondto gap junctions aggregated into placques (a). Many small holesrepresent a scattered gap junctional distribution (b). . . . . . . . . 37
x

3.3 Our model of cells and gap junctions. Cells in a sequence are mod-eled as squares whose interiors are not physically connected. Gapjunctions are represented in a continuous manner as resistive connec-tions between neighboring cells. The multiple resistors shown rep-resent a flux boundary condition between the cells. A non-uniformdistribution of resistance (a) corresponds to a gap junction distri-bution with placques. A uniform distribution of resistance (b) cor-responds to a scattered distribution of gap junctions. . . . . . . . . 37
3.4 The tissue level cellular architecture. Cells are modeled as squaresadjoined without extracellular space. The thick horizonal lines rep-resent membranes that prevent vertical current flow. Hence, thebehavior along each row of cells is identical. An individual cell needonly be identified by its horizontal cell number, i. . . . . . . . . . . 38
4.1 A cartoon of the ephaptic mechanism between two adjacent cells(shaded). Initially, the Na+ channels of both the pre-junctional(left) and post-junctional (right) cells are closed (a). When an actionpotential reaches the left cell, its Na+ channels open, allowing a fluxof Na+ current in from extracellular space via the clefts (b). As perOhm’s law, the potential inside the cleft decreases, resulting in thedepolarization of the post-junctional membrande. If this effect issufficiently strong, this membrance may reach threshold, causingthe Na+ channels to open and allowing current to flow inward (c). . 46
4.2 The geometry and circuit diagram for our model of an ephapticmechanism. We describe cells as active cables coupled through di-rect resistive connections and active membranes involving sharedcleft potentials. We model extracellular space as grounded. Weignore the effects of any changes in ion concentration. . . . . . . . . 48
4.3 A schematic of the domains for the full (a) and homogonized (b)models. The full model is posed over discrete cells of length ε withan equipotential cleft between adjacent cells. The intracellular po-tential within the i-th cell is φi(t, x), and the cleft potential to theright of the i-th cell is φi
c(t). Note that the cleft potential is definedonly over a discrete set of points. The homogenized model is posedover the entire length Ltissue of tissue, as it does not resolve individ-ual cells. The intracellular potential φ0(t, x) and the cleft potentialφc,0(t, x) are defined over the entire domain. . . . . . . . . . . . . . 54
xi

4.4 Conduction velocity under the full ephaptic model as a function ofthe nondimensional cleft-to-ground resistance for gap junction ex-pression ranging from 1% to 100% of normal. For these simulations,ε = 0.01. Na+ channels are distributed such that (a) all are locatedat the intercalated disks, (b) half are located at the intercalateddisks, or (c) channel density is uniform. . . . . . . . . . . . . . . . . 58
4.5 Conduction velocity under the homogenized ephaptic model as afunction of the nondimensional cleft-to-ground resistance for gapjunction expression ranging from 1% to 100% of normal. Na+ chan-nels are distributed such that (a) all are located at the intercalateddisks, (b) half are located at the intercalated disks, or (c) channeldensity is uniform. . . . . . . . . . . . . . . . . . . . . . . . . . . . 60
4.6 An overlay of the conduction speeds computed under the full andhomogenized systems. At normal gap junction expression levels(κ = 1) the systems agree well, but that agreement disappears asthe gap junctional coupling is reduced. . . . . . . . . . . . . . . . . 62
4.7 An overlay of the computed solutions to the full and homogenizedsystems under β = 10−3, κ = 1, fNa = 1 at two different times. Thetop panels show intracellular potentials. Note that individual cellscan be resolved in the upstroke of the full simulations. The bottompanels show cleft potential. For clarity, the intracellular potentialsof the full simulations are plotted for only 5 of the 20 interior nodes. 63
4.8 An overlay of the computed solutions to the full and homogenizedsystems under β = 10−3, κ = 0.01, fNa = 1 at two different times.The top panels show intracellular potentials. Note that individualcells can be resolved in the upstroke of the full simulations. Thebottom panels show cleft potential. For clarity, the intracellularpotentials of the full simulations are plotted for only 5 of the 20interior nodes. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 64
4.9 The spatial discretization of each biological cell in the full model.Each biological cell is broken into n computational nodes and twoghost nodes, represented by hollow dots. Note that there are nocomputational nodes for the clefts as the cleft potential can be de-duced from the potential at the interior and ghost nodes. Also notethat the rightward ghost node of a cell is distinct from the leftwardghost node of its neighbor. The variable φi
k denotes the potential atthe k-th node of the i-th cell, where k = 0 and k = n+1 correspondto the ghost nodes. . . . . . . . . . . . . . . . . . . . . . . . . . . . 67
xii

7.1 Evolution of the Keller-Segel system with (JL) boundary conditionsover the singly periodic square, T × [0, 1] with mass M = 4.5πinitially distributed in a noisy uniform shape. Note that betweentimes t = 0.5 and t = 2.5, the solution has the form of a growingcosinusoidal disturbance from uniform. After that, the solution ap-proaches the steady state of the 1d simulation with the same mass.In this simulation ∆t = 0.005,∆x = ∆y = 1/40. For clarity, theplot only shows a 20 × 20 sampling of the 40 × 40 simulated gridpoints. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 90
7.2 Evolution of the Keller-Segel system with (JL) boundary conditionsover the singly periodic squre, T×[0, 1] with mass M = 4.6π initiallydistributed in a noisy uniform shape. The solution follows the samedescription as the simulation shown in Figure 7.1, but it stays nearthe 1d steady state only until t ≈ 50. At this time, a disturbanceat the boundary grows until the mass concentrates there by finitetime. In this simulation ∆t = 0.005,∆x = ∆y = 1/40. For clarity,the plot only shows a 20×20 sampling of the 40×40 simulated gridpoints. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 91
7.3 Plot of the smallest eigenvalue of the discretization of the Keller-Segel equations over T × [0, 1] with (JL) boundary conditions, lin-earized about the one-dimensional non-uniform steady state withmass M . A negative eigenvalue indicates a linear instability. The1d steady states change from being linearly stable to linearly unsta-ble as mass increased beyond M∗ ≈ 4.57π. . . . . . . . . . . . . . . 94
xiii

List of Tables
1.1 Differential Equations, conductivities, reversal potentials, and rest-ing values used in simulations in Chapter 4, based on the Luo Rudy1991 dynamic [34]. All ordinary differential equations carry unitsms−1. From [34], the resting value of X was unclear. For it, weselected a small, non-zero initial value for our evolution simulationsin Chapter 4. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
1.2 Rate constants and values for gating variables. . . . . . . . . . . . . 6
2.1 Intracellular, longitudinal conductivities σi,l (mS/cm) obtained fromthe aligned and brick-like arrangements for various values of gGJ,end
and gGJ,side. The numbers before the commas are for the alignedarrangement. The numbers after the commas are for the brick-likearrangement. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26
2.2 Conductivity values obtained by fitting macroscopic wavespeed datato solutions of bidomain equations. Values are obtained for ventric-ular tissue from various animals, such as dogs, cows, and sheep. . . 28
2.3 Conductivity values obtained directly from microscopic measure-ments via homogenization in the present work. . . . . . . . . . . . . 28
2.4 The physical parameters, derived parameters, and variables thatenter our cellular model. Measured parameters from [56, 18] corre-spond to mouse ventricular myocytes. . . . . . . . . . . . . . . . . . 34
4.1 The physical and derived parameters that enter our full and homog-enized models. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 66
4.2 Computational parameters that enter our numerical simulations ofthe full and homogenized models. . . . . . . . . . . . . . . . . . . . 66
4.3 Spatial convergence rates for full system with ε = 0.01 computed atnondimensional time 0.2. . . . . . . . . . . . . . . . . . . . . . . . . 69
4.4 Temporal convergence rates for full system computed at nondimen-sional time 0.2. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 70
4.5 Spatial convergence rates for homogenized system computed at nondi-mensional time 0.2. . . . . . . . . . . . . . . . . . . . . . . . . . . . 72
xiv

4.6 Temporal convergence rates for homogenized system computed atnondimensional time 0.2. . . . . . . . . . . . . . . . . . . . . . . . . 72
7.1 Spatial convergence rates for Keller-Segel simulation computed attime 0.25. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 96
7.2 Temporal convergence rates computed at time 0.25. . . . . . . . . . 97
xv

Part I
Homogenization in Cardiac
Electrophysiology
1

Chapter 1
Introduction
1.1 Cellular Biophysics
Many cells require electricity to function. Some, such as epithelial cells, controlthe concentrations of ions like Na+ and K+, primarily for the purpose of control-ling cell volume [29, 24] . Others, such as neurons and cardiac muscle cells, arecalled excitable and control ionic flow for the purpose of electrical communication.If the electric potential of excitable cells is slightly disturbed due to a current in-jection, it relaxes to its resting value. If, instead, the potential is disturbed by alarge enough current, it undergoes a large spike, called an action potential, beforereturning to rest. For a more thorough introduction, see [29, 16]. These actionpotentials can propagate down nerve axons and through cardiac muscle, resultingin communication between distant cells.
In the electric circuit model of cells, the membrane is described by a capacitor
Figure 1.1: The electrical circuit model of an isopotential cell (shaded). Anycurrent injected inside would either charge the plasma membrane as a capacitoror flow across the membrane through ion channels.
2

in parallel with non-ohmic resistors. The phospholipid bilayer acts as a capacitorbecause it is an insulator which can separate charge. Membrane ion channelsare the resistors as they permit current flow in response to electric potentials.Figure 1.1 shows the resulting model circuit for an electrically active isopotentialcell. With it, we can see that current injected into the intracellular region eithercharges the cell as a capacitor or flows through the ion channels. In the absense ofsuch an injection, the potential evolves according to the differential equation
Cdφ
dt+ Iion(φ, w) = 0, (1.1)
where φ is the transmembrane potential, C is the membrane capacitance per unitarea, and Iion is the outward ionic current per unit area at specific values of φ andany relevant gating variables w.
The specific ions and currents relevant for physiology varies between cell typesand animal species. Mathematically, these differences alter the Iion function. Themost famous ionic model is the Hodgkin-Huxley model [23] of the squid giantaxon. A similar model for mammalian ventricular muscle is the Luo-Rudy dynamic[34, 35, 14]. There are many other realistic models, but scientists also study non-physiological ones, such as the Fitzhugh-Nagumo model [39, 19, 43] or McKean’spiecewise linear model [38], because of their mathematical simplicity. We detailthe Hodgkin-Huxley and Luo-Rudy models now.
1.1.1 The Hodgkin-Huxley Ionic Model
The Hodgkin-Huxley model is composed of three ionic currents: a fast-activating,slow-inactivating Na+ current; a slow-rectifying K+ current; and a leak current.The resistances underlying the Na+ and K+ currents are governed by gating vari-ables which evolve in a voltage dependent way.
Precisely, the Hodgkin-Huxley ionic model is given by
w = (n,m, h),
Iion(φ, w) = gNa ·m3 · h · (φ−ENa) + gK · n4 · (φ− EK) + gl · (φ− El),
ds
dt= αs(φ)(1 − s) − βs(φ)s for s = n,m, or h,
where gNa, gK, gl are the maximal conductances for the respective Na+, K+, andleak currents; n,m, h are gating variables between 0 and 1; ENa, EK, El are thereversal potentials for the Na+, K+, and leak currents; and αs(φ), βs(φ) are theexperimentally obtained voltage dependent rate constants governing the openingand closing of gates within the relevant ion channels. The precise form of αs(φ)and βs(φ) can be found in [29]. In the above equations, w is a three component
3

vector, and s stands for any single one of those components.
The reversal potentials depend on the ionic concentrations inside and outsidethe cell through the Nernst equation. Technically, any current through the channelsalters this concentration, but such changes are typically small enough that theconcentrations, and hence the reversal potentials, are constants in time.
1.1.2 The Luo-Rudy Ionic Model
The Luo-Rudy ionic model, also known as the Luo-Rudy dynamic (LRd) is anionic model of mammalian cardiac ventricular cells. The 1991 version of the model[34] consists of six currents, all with separate dynamics. More recent versions areeven more detailed [35, 15, 14].
Precisely, the LRd ionic model is given by
w = (m, h, j, d, f,X, [Ca]i),
Iion(φ, w) = INa + Isi + IK + IK1 + IKp + Ib,
INa = gNa ·m3 · h · j · (φ−ENa),
Isi = gsi · d · f · (φ−Esi([Ca]i)) ,
IK = gK ·X ·Xi · (φ− EK),
IK1 = gK1 ·K1∞ · (φ−EK1),
IKp = gKp ·Kp · (φ− EKp),
Ib = gb · (φ− Eb),
dw
dt= LRD(φ, w),
K1∞ = αK1(φ)/ (αK1(φ) + βK1(φ)) ,
Xi = Xi(φ),
Kp = Kp(φ),
where INa is the fast inward sodium current, Isi is the slow inward current, IKis the time-dependent potassium current, IK1 is the time-independent potassiumcurrent, IKp is the plateau potassium current, Ib is the background current; thetime dependent gating variables arem, h, j, d, f, and X; and the g’s and E’s are themaximal conductances and reversal potentials for their corresponding variables.The K1 variable relaxes quickly, so it is replaced with its steady state value.Further, the Xi and Kp variables have no time dependence and are only a functionof potential. The differential equations for w given by LRD(φ, w), the values forunspecified constants, and the resting values used in our simulations are given inTable 1.1. Table 1.2 presents the voltage dependent rate constants and gatingvariables.
4

Differential Equations
ds
dt= αs(φ) − (αs(φ) + βs(φ))s for s = m, h, j, d, f,X,
d[Ca]idt
= 10−4Isi + 0.07(10−4 − [Ca]i).
Conductivities and Reversal Potentials
gNa = 23 mS/cm2, ENa = 54.4 mV,
gsi = 0.09 mS/cm2, Esi = 7.7 − 13.0287 · ln([Ca]i) mV,
gK = 0.282 ·√
[K]0/5.4 mS/cm2, EK = −77 mV,
gK1 = 0.6047 ·√
[K]0/5.4 mS/cm2, EK1 = 103RT
Fln
[K]0[K]i
mV,
gKp = 0.0183 mS/cm2, EKp = EK1,
gb = 0.03921 mS/cm2, Eb = −59.87 mV.
Other Constants
[K]0 = 5.4 mM, [K]i = 145 mM,
R = 8.315 J / mol K, F = 9.648 · 104 C / mol,
T = 310.15 K.
Resting Values
φ0 = −88.655 mV, m0 = 0.000838,
h0 = 0.993336, j0 = 0.995484,
d0 = 0.000003, f0 = 0.999745,
X0 = 0.004503, [Ca]i,0 = 0.004503.
Table 1.1: Differential Equations, conductivities, reversal potentials, and restingvalues used in simulations in Chapter 4, based on the Luo Rudy 1991 dynamic[34]. All ordinary differential equations carry units ms−1. From [34], the restingvalue of X was unclear. For it, we selected a small, non-zero initial value for ourevolution simulations in Chapter 4.
5

Rate Constants and Values for Gating Variables
αh =
0 φ < −40 mV,
0.135 · exp[(80 + φ)/ − 6.8] φ ≥ −40 mV,
βh =
1/(0.131 + exp[(V + 10.66)/ − 11.1]) φ < −40 mV,
3.56 · exp(0.079φ) + 3.1 · 105 · exp(0.35φ) φ ≥ −40 mV,
αj =
0 φ < −40 mV,
[−1.2714 · 105 · exp(0.2444φ) − 3.474 · 10−5 · exp(−0.04391φ)]
· (V + 37.78)/1 + exp[0.311 · (φ + 79.23)] φ ≥ −40 mV,
βj =
0.3 · exp(−2.535 · 10−7φ)/1 + exp[−0.1(φ + 32)] φ < −40 mV,
0.1212 · exp(−0.01052φ)/1 + exp[−0.1378(φ + 40.14)] φ ≥ −40 mV,
αm = 0.32(φ + 47.13)/1 − exp[−0.1(φ + 47.13)],βm = 0.08 · exp(−φ/11),
αd = 0.095 · exp[−0.01(φ − 5)]/1 + exp[−0.072(φ − 5)],βd = 0.07 · exp[−0.017(φ + 44)]/1 + exp[0.05(φ + 44)],αf = 0.012 · exp[−0.008(φ + 28)]/1 + exp[0.15(φ + 28)],βf = 0.0065 · exp[−0.02(φ + 30)]/1 + exp[−0.2(φ + 30)],αX = 0.0005 · exp[0.083(φ + 50)]/1 + exp[0.057(φ + 50)],βX = 0.0013 · exp[−0.06(φ + 20)]/1 + exp[−0.04(φ + 20)],
αK1 = 1.02/1 + exp[0.2385 · (φ − EK1 − 59.215)],βK1 = 0.49124 · exp[0.08032 · (φ − EK1 + 5.476)] + exp[0.06175 · (φ − EK1 − 594.31)]
/ 1 + exp[−0.5143 · (φ − EK1 + 4.753)],
Xi =
1 φ ≤ −100 mV,
2.837 · exp[0.04(φ + 77)] − 1/(V + 77) · exp[0.04(φ + 35)] φ > −100 mV,
Kp = 1/1 + exp[(7.488 − φ)/5.98].
Table 1.2: Rate constants and values for gating variables.
6

x1 x2
r
Figure 1.2: A cable of electrically active membrane filled with conducting fluid. Aflux balance calculation allows for a derivation of the cable equation.
1.2 The Cable Equation
As mentioned in Section 1.1, action potentials can propagate down nerve axonsor cardiac muscle. In order to study such propagation mathematically, we mustmodify the differential equation (1.1) to incorporate spatial effets. The resultingpartial differential equation (PDE) is called the cable equation, which we nowderive.
Consider a long cylinder of radius r of electrically active membrane, as inFigure 1.2, filled with conducting cytosolic fluid. Assuming there are no appreciablevariations of the potential within a cross-section of the cable, φ varies only withtime and the x coordinate. As per Ohm’s law,
j(t, x) = −σc∂φ
∂x(t, x) (1.2)
where j is the current density, and σc is the cytoplasmic conductivity. The netcurrent into the region between x = x1 and x = x2 either charges the membraneas a capacitor or passes through the ion channels. Thus
πr2
(
−σc∂φ
∂x(t, x1) + σc
∂φ
∂x(t, x2)
)
=
∫ x2
x1
2πr
(
C∂φ
∂t(t, x) + Iion(φ, w)
)
dx.
(1.3)
Writing the left hand side as an integral, we find
πr2σc
∫ x2
x1
∂2φ
∂x2(t, x)dx =
∫ x2
x1
2πr
(
C∂φ
∂t(t, x) + Iion(φ, w)
)
dx. (1.4)
Since this equation holds for all x1, x2, we obtain the cable equation
σcr
2
∂2φ
∂x2= C
∂φ
∂t+ Iion(φ, w). (1.5)
7

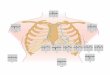
Figure 1.3: A depiction of the complicated geometrical structure of the intracellularand extracellular space of cardiac tissue. The thick, dark vertical stripes separatingcells are known as intercalated disks. Reprinted with permission from Guyton andHall (Fig. 9-2, p. 108).
1.3 The Bidomain Equations
Heart muscle is an irregular, three dimensional arrangement of cells with in-tricate structure. The cells are typically around 100 µ in length and about 20 µin width [29], and are connected to each other at their ends. As can be seen inFigure 1.3, the cells form fibers which can branch. Adjacent cells are connectedthrough the intercalated disks, shown by the thick dark lines in the figure. Thecells are also surrounded by a conducting, irregularly shaped extracellular space.Action potential propagation in cardiac tissue results from ion flow in and out ofcells in this convoluted geometry.
In order to study cardiac action potential propagation mathematically, wemodel the tissue in an averaged way. The resulting partial differential equationsare called the bidomain equations. Their averaged nature avoids the cumbersometask of detailing the complexity of cellular geometry and arrangements at the mi-cron level.
To derive the bidomain equations, we let Ωi denote the intracellular space ofa tissue, Ωe denote the extracellular space, and Ω = Ωi ∪ Ωe. Let φi(x) be theintracellular potential and φe(x) be the extracellular potential. Technically, φi isdefined only over Ωi and φe only over Ωe. Viewing Ω as being a combination ofintracellular and extracellular space for all x, we let φi be defined over all Ω. Weinterpret its value at a point x as the intracellular potential in a region near x.Similarly, we consider φe to be defined over all Ω.
8

We assume that the intracellular and extracellular domains give rise to ananisotropic, ohmic current-voltage relationship,
ji = −σi∇φi, (1.6)
je = −σe∇φe, (1.7)
where ji and je are the intracellular and extracellular current densities, σi and σe
are the intracellular and extracellular conductivity tensors, which could, in princi-ple, vary with spatial position.
Barring outside current injections, there can be no source or sink of current inthe combination of extracellular and intracellular space. Any apparent sink in theintracellular current must then be a source in the extracellular current. Thus
∇ · (−σi∇φi − σe∇φe) = 0. (1.8)
The current flowing from intracellular to extracellular space acts either tocharge the local membrane as a capacitor or to flow through the ion channels.Thus
∇ · (σi∇φi) = β
(
C∂(φi − φe)
∂t+ Iion(φi − φe, w)
)
, (1.9)
where β is the membrane surface area per unit volume of tissue. By combining(1.8) and (1.9), we obtain the bidomain equations
∇ · (σi∇φi) = β
(
C∂(φi − φe)
∂t+ Iion(φi − φe, w)
)
, (1.10)
∇ · (σe∇φe) = −β(
C∂(φi − φe)
∂t+ Iion(φi − φe, w)
)
. (1.11)
These equations are the commonly accepted macroscopic description of of car-diac tissue under normal and pathological conditions [22].
1.4 Homogenization of Partial Differential Equa-
tions
Homogenization is a two-scale asymptotic technique used to describe an av-eraged description of a partial differential equation with periodic structure. Ex-amples of such periodic structure include highly oscillatory conductivity and peri-odic geometry. Such an averaged description allows us to determine the effectiveconductivities of a periodic medium. We now formally demonstrate the homoge-nization of a material with periodic geometry for the purpose of determining its
9

Y ε Y
(a) (b)
Figure 1.4: In this section, we homogenize Laplace’s equation over the periodicdomain Y ε (a). This domain is composed of translated and scaled versions of theunit cell Y (b).
effective conductivities. See [13] for a similar demonstration with periodic conduc-tivity.
Let Y ⊂ T3 have a smooth boundary and a connected periodic extension. Let
Y ε = ∪i,j,k ε · (Y + (i, j, k)), which is depicted in Figure 1.4. Let Ω = [0, 1]3 andΩε = Ω ∩ Y ε.
Consider Laplace’s equation in Ωε,
−σ∆φε(x) = f(x) in Ωε, (1.12)
∂νφε = 0 on ∂Ωε\∂Ω, (1.13)
φε(x) = 0 on ∂Ω. (1.14)
We make the two-scale homogenization ansatz
φε(x) = φ0(x) + εφ1(x; x/ε) + ε2φ2(x; x/ε) + · · · , (1.15)
where φ1 and φ2 are 1 × 1 × 1-periodic in the second variable y = x/ε. Pluggingthe ansatz (1.15) into the boundary value problem (1.12)–(1.14) and extractingthe leading order terms gives
−σ∆yφ1(x; y) = 0 in Y, (1.16)
−∇yφ1 · ν = ∇xφ0 · ν on ∂Y. (1.17)
The next order terms in ε are
∇y (σ∇yφ2 + σ∇xφ1) = −∇x (σ∇xφ0 + σ∇yφ1) in Y, (1.18)
−∇yφ2 · ν = ∇xφ1 · ν on ∂Y. (1.19)
10

The boundary value problem for φ1 can be written in terms of the correctorfunctions wi by
φ1(x; y) = ∂xi(x)wi(y), (1.20)
where wi(y) solves Laplace’s equation over the periodic domain Y . Specifically,
−∆ywi(y) = 0 in Y, (1.21)
−∂νwi(y) = ek · ν on ∂Y. (1.22)
As the macroscopic φ0 does not satisfy the boundary conditions on the fastspatial scale of the periodic domain, the O(ε) term in the ansatz functions to cor-rect the solutions normal derivative on the microscale domain boundary.
Finally, a PDE for φ0 can be obtained by applying the solvability condition forthe φ2 equation. Integrating (1.18) over Ω gives
∫
Ω
∇y · (σ∇yφ2 + σ∇xφ1) + ∇x · (σ∇xφ0) + ∇x · (σ∇yφ1)dx = f(x)|Y |. (1.23)
where |Y | is the volume of Ω. The first two terms cancel by applying the divergencetheorem and the boundary condition (1.19). The equation, which inherits theboundary condition on ∂Ω, then becomes
∂xi
(
σ
(
δik +1
|Y |
∫
Y
∂ykwi(y)dy
)
∂xkφ0(x)
)
= f in Ω, (1.24)
φ0(x) = 0 on ∂Ω, (1.25)
from which we can read off the conductivity tensor.Alternatively, effective conductivity could be determined by placing a potential
difference along one direction of the macroscopic domain Ω and calculating thedrawn current. This approach is used in Chapter 4.
1.5 Outline of Part I
In Chapter 2, we present a modification to the conventional framework fordescribing cardiac myocytes in calculations of effective macroscopic conductivity.Instead of modeling gap junctions as discrete geometrical entities, we model theireffect through Neumann boundary conditions on simple cellular geometry. Wethen derive effective conductivity values based on measured cellular parameters inthe cases of aligned and brick-like cellular arrangements. We compare our conduc-tivity values to those obtained in the literature either (1) by fitting macroscopic
11

wavespeed data to solutions of the bidomain equations or (2) by inference basedon microscopic measurements. We also discuss the applicability of our frameworkto electromechanical simulations.
In Chapter 3, we use this modified framework in order to determine whichdistribution of gap junctions along the ends of cells provides the most macroscopicconductivity. In agreement with random walk and PDE models describing gapjunctions through geometry, we establish that a uniform distribution is a localmaximizer of conductivity when the total number of gap junctions is held fixed.
In Chapter 4, we present a model of ephaptic cardiac communication throughextracellular clefts which are resistively connected to extracellular space. Wenondimensionalize and homogenize the differential equations arising from the bio-physics. We investigate the full and homogenized models numerically and comparethe computed wavespeeds and waveforms over physiologically relevant parameterregimes. We observe that the two models agree when gap junctional coupling isat physiologically normal levels but disagree when gap junction levels are low.
12

Chapter 2
Homogenization of Cardiac
Models that Describe Gap
Junctions Through Boundary
Conditions
2.1 Introduction
Computer simulations have the potential to increase our understanding of nor-mal and pathological cardiac function, and to improve the effectiveness of clinicaltherapies. Although the biophysics at the cellular level is well understood, wholeheart simulations that resolve every cell are computationally infeasible at present.Instead, a more tractable approach is to perform macroscopic simulations basedon the bidomain equations [22], which govern locally averaged potentials insideand outside cells. These equations require physical values of the effective conduc-tivity of intracellular and extracellular regions in both the longitudinal (fiber) andthe transverse (cross-fiber) directions. Ideally, such values should be directly ob-tained from measurable cellular properties, such as geometry and gap junctionalconductivity. Otherwise, the values are free parameters which must be chosen bymatching simulation results to experiments.
There are relatively few studies which attempt to derive the macroscopic pa-rameters required by the bidomain equations directly from measurable microscalequantities. One approach to obtain these values is homogenization of partial dif-ferential equations (PDEs) as in Neu and Krassowska [44]. In this approach, theintracellular region of tissue is modeled as a collection of periodically arrangedcells connected through physical openings that correspond to gap junctions. Theycan compute effective conductivitities by solving Laplace’s equation on a periodicdomain. Although these authors present a detailed derivation involving homoge-
13

nization, they resort to a resistor network model in order to find analytical for-mulae for the effective bidomain conductivities. A shared inconvenience of theirPDE and resistor network models is that they intertwine transverse and longitu-dinal gap junctional connections, making it difficult to assign proper values basedon separate experimental measurements of side-to-side and end-to-end couplingstrengths [56].
An alternative approach to computing passive conductivities is given by Stin-stra et al. [51], who create a detailed tissue model designed to account for re-alistically complex cell shapes with random variability. Sinstra et al. then solveLaplace’s equation over a domain containing several cells to obtain the effectiveconductivities. Although their simulations involve a high level of detail in the cellu-lar microstructure, their calculation for intracellular transverse conductivity yieldsvalues an order of magnitude less than those in the experimental literature. Theypropose two possible explanations of this discrepancy: the total gap junctionalconductivity may be larger than measured, and the gap junctions may be morepreferentially located on the cell sides than is measured. In the present chapter,we explore the feasibility of both explanations.
In this chapter, we follow a homogenization approach similar to that of Neuand Krassowska [44] to derive the effective conductivites of cardiac tissue. Unlikethis earlier homogenization work, however, we employ our microscale PDE modelto obtain the macroscopic bidomain parameters. In particular, we do not resortto a resistor network to obtain the macroscopic parameters from the measuredmicroscale quantities. In our approach, we separate the structure and placementof cells from the gap junctional connections between them. Specifically, we idealizecells as rectangular prisms, inside of which the electric potential satisifies Laplace’sequation. Instead of modeling gap junctions as discrete geometrical entities akin toFigure 2.1a, we include their effect as boundary conditions on each cell membraneas in Figure 2.1b. This approach assumes no more detail than is provided by directmeasurements, such as those in [56]. These modeling decisions make a mathemat-ically natural framework within which to study the effects on conductivity of thearrangement of cells and of the distribution of gap junctions on cell membranes.In some cases, we are able to obtain analytical formulae for the effective conduc-tivitites. In cases where we cannot do so, we need only to solve Laplace’s equationon one cell of fixed geometry.
The remainder of this chapter proceeds as follows: In Section 2.2, we motivatethe calculation of effective conductivities by considering their role in the bidomainequations. Section 2.3 sets up the electrostatic Laplace’s equation in the alignedcellular arrangement. In Section 2.4, we perform the homogenization in the alignedarrangement and calculate the numerical values of the effective conductivities.Similarly, in Section 2.5, we formulate the electrostatic problem in a brick-likecellular arrangement, perform the homogenization, and calculate the corresponding
14

a b
Figure 2.1: Instead of modeling gap junctions through complex cellular geometry(a), we model them through flux boundary conditions on simple geometry (b).The multiple resistors shown in (b) represent a continuous boundary condition,see equations (2.2) - (2.7).
conductivities. In Section 2.6, we compare our conductivity values to those inthe literature and discuss the relevance of our computations to electromechanicalsimulations. In Section 2.7 we present our conclusions. Finally, in a brief Appendix,we present additional details regarding the derivation of the extracellular effectiveconductivities.
2.2 Bidomain Equations and Effective Conduc-
tivity
The bidomain equations provide the most realistic macroscopic description ofthe electrical activity of cardiac tissue under normal and pathological conditions[22]. They govern the intracellular and extracellular electric potential in an aver-aged sense and are given by
∇ · (σi∇φ0i ) = β
(
C∂t(φ0i − φ0
e) + Iion(φ0i − φ0
e, ω))
,
∇ · (σe∇φ0e) = −β
(
C∂t(φ0i − φ0
e) + Iion(φ0i − φ0
e, ω))
,
where σi and σe are the macroscopic conductivity tensors of the intracellular andextracellular spaces, β is the membrane surface area per unit volume of tissue, Cis the membrane capacitance per unit area, ω stands for relevant gating variables,and Iion is the ionic current per unit area of the membrane. Here, φ0
i (t, ~x) andφ0
e(t, ~x) are each defined for all ~x, and are to be interpreted as the locally averagedintracellular or extracellular potential near the point ~x. Note that the intracellularand extracellular potential are each separately averaged, not averaged with eachother.
15

l
wc
wp
φi,j+1,ki φi+1,j+1,k
i
φi,j,ki φi+1,j,k
ix1
x1
x2 x2
x3
a b
Figure 2.2: The aligned cellular architecture: (a) cells of dimension l×wc ×wc arearranged in three space with period l × wp × wp; (b) an x1, x2 cross-section.
We write the superscript ‘0’ because φ0i is the leading order term of the asymp-
totic expansion (2.15) for intracellular potential. Similarly, φ0e is the leading order
term of a corresponding expansion for extracellular potential. If we locally align thecoordinates with the myocardial fibers so that the x1 direction coincides with thefiber direction and assume that the x2 and x3 directions are locally indistinguish-able, then the conductivity tensors are diagonal and involve only the longitudinaland transverse conductivities. That is,
σi =
σi,l 0 00 σi,t 00 0 σi,t
and σe =
σe,l 0 00 σe,t 00 0 σe,t
.
A goal of the present work is to determine σi,l, σi,t, σe,l, σe,t directly from mea-sured microscopic quantities.
2.3 Full Cellular Model in an Aligned Arrange-
ment
We model myocytes as l×wc ×wc prisms arranged periodically in three-spacewith period l × wp × wp, see Figure 2.2. The length of the cells l is assumedto equal the longitudinal period, but the width and height of the cells wc aresmaller than the transverse periods wp in order to provide an extracellular volumefraction α = 1 − (wc/wp)
2. In this model, the intracellular space is not physicallycontiguous. Instead, resistive connections allow current to flow directly between theinteriors of adjacent cells. Note that we do not attempt to account for the branchingof cells in the present work. Additionally, we ignore fiber rotation because ouranalysis is purely local.
To identify the bidomain parameters σi and σe, we consider the degeneratecase of a steady state without transmembrane ionic current. In this situation,
16

the equations for intracellular and extracellular potential decouple. In the follow-ing, we consider only the intracellular potential; the similar calculations for theextracellular potential are described in Appendix 2.8.
The electric potential in the intracellular space satisfies Laplace’s equation,
−∆φi,j,ki (~x) = 0 in Ωi,j,k
i , (2.1)
where Ωi,j,ki = [i · l, (i + 1) · l] × [j · wp, j · wp + wc] × [k · wp, k · wp + wc] is the
region occupied by the (i, j, k)-th cell, and φi,j,ki is the intracellular potential of
the (i, j, k)-th cell, indicated in Figure 2.2b. For ease of notation, we identify thedomain of the function φi,j,k
i with [0, l] × [0, wc] × [0, wc]. We denote position by~x = (x1, x2, x3).
We model the gap junctions between cells in a continuous manner throughboundary conditions on equation (2.1). The current density between two neigh-boring cells is proportional to the potential difference between the positions onopposite sides of the gap junctions:
−σc∂x1φi,j,k
i (l, x2, x3) =gGJ,end
w2c
(
φi,j,ki (l, x2, x3) − φi+1,j,k
i (0, x2, x3))
, (2.2)
−σc∂x2φi,j,k
i (x1, wc, x3) =gGJ,side
l · wc
(
φi,j,ki (x1, wc, x3) − φi,j+1,k
i (x1, 0, x3))
, (2.3)
−σc∂x3φi,j,k
i (x1, x2, wc) =gGJ,side
l · wc
(
φi,j,ki (x1, x2, wc) − φi,j,k+1
i (x1, x2, 0))
, (2.4)
−σc∂x1φi,j,k
i (l, x2, x3) = −σc∂x1φi+1,j,k
i (0, x2, x3), (2.5)
−σc∂x2φi,j,k
i (x1, wc, x3) = −σc∂x2φi,j+1,k
i (x1, 0, x3), (2.6)
−σc∂x3φi,j,k
i (x1, x2, wc) = −σc∂x3φi,j,k+1
i (x1, x2, 0), (2.7)
where σc is the cytoplasmic conductivity (mS/cm), gGJ,end is the total conductance(mS) of all gap junctions on one end of a cell, and gGJ,side is the total conductance(mS) of all gap junctions on one side of a cell. Equations (2.2)–(2.4) balancecytosolic and gap junctional current, whereas equations (2.5)–(2.7) equate thecurrent leaving each cell with the current entering its appropriate neighbor. SeeTable 2.4 for the physical parameters and variables introduced for this model.
In the remainder of the present section, we nondimensionalize equations (2.1)–(2.7) and state the effective conductivity problem in this aligned cellular arrange-ment. Note that the model could easily be generalized to allow for more complexgeometries, such as the ‘jutting’ cells analyzed in [25]. It could also allow gap junc-tional density to be varying within the ends or sides of cells. As an illustration,we explore the effects of a brick-like cellular arrangement in Section 2.5.
17

2.3.1 Nondimensionlization
We rescale space so that cells are of length ε by letting
~x =ε
l~x.
Dropping the tildes, equations (2.1)–(2.7) become
−∆φi,j,ki (~x) = 0 in [0, ε] × [0, εhc] × [0, εhc], (2.8)
−∂x1φi,j,k
i (ε, x2, x3) =1
εκend
(
φi,j,ki (ε, x2, x3) − φi+1,j,k
i (0, x2, x3)
)
, (2.9)
−∂x2φi,j,k
i (x1, εhc, x3) =1
εκside
(
φi,j,ki (x1, εhc, x3) − φi,j+1,k
i (x1, 0, x3)
)
, (2.10)
−∂x3φi,j,k
i (x1, x2, εhc) =1
εκside
(
φi,j,ki (x1, x2, εhc) − φi,j,k+1
i (x1, x2, 0)
)
, (2.11)
−∂x1φi,j,k
i (ε, x2, x3) = −∂x1φi+1,j,k
i (0, x2, x3), (2.12)
−∂x2φi,j,k
i (x1, εhc, x3) = −∂x2φi,j+1,k
i (x1, 0, x3), (2.13)
−∂x3φi,j,k
i (x1, x2, εhc) = −∂x3φi,j,k+1
i (x1, x2, 0), (2.14)
where κend =gGJ,endl
σcw2c
is the nondimensional end-to-end gap junctional conductance,
κside =gGJ,side
σcwcis the nondimensional side-to-side gap junctional conductance, the
intracellular domain of the (i, j, k)-th cell is of size [0, ε]×[0, εhc]×[0, εhc], and hc =wc/l. Letting hp = wp/l, the tissue microstructure now has period ε× εhp × εhp.The small parameter ε can be interpreted as the ratio of the length of a myocyteto the length scale of significant variations of electrical potential within the tissue;see [44] for further details.
2.3.2 Statement of the Effective Conductivity Problem
Consider a cube of tissue of dimensionless size 1×1×1, composed of cells of sizeε×εhc×εhc arranged with period ε×εhp×εhp. The index notation distinguishingthe (i, j, k)-th cell is cumbersome, and we introduce φi(~x) as a single functiondefined over Ωi = ∪i,j,kΩ
i,j,ki . The function φi satisfies the PDE (2.8) within each
cell, and microscopic boundary conditions akin to (2.9)–(2.14) on the boundary ofeach cell. As depicted in Figure 2.3a, it will also satisfy the macroscopic boundary
18

replacemen
εεhc
φi = 0 φi = V
φi
φ0i (~x)
φ0i (~x) + εφ1
i (~x; ~x/ε)a b
x1
x1
x2
Figure 2.3: In order to compute effective conductivity, we impose a potentialdifference across a large number of cells and calculate the resulting current density(a). In this figure, extracellular space has been omitted for clarity. The resultingelectric potential is depicted in (b).
conditions
φi(0, x2, x3) = 0,
φi(1, x2, x3) = V,
∂x2φi(x1, 0, x3) = 0,
∂x2φi(x1, 1, x3) = 0,
∂x3φi(x1, x2, 0) = 0,
∂x3φi(x1, x2, 1) = 0.
The dimensional intracellular longitudinal conductivity is then given by
σi,l = σc ·V
1A
∫∫
Ωi∩x1=c ∂x1φi dx2dx3
,
where A is the cross-sectional area of the tissue, the domain of integration is theset of interior points across a cross-section, and the integral is independent of theconstant c.
Intracellular transverse conductivity can be determined analogously, providedthe macroscopic boundary conditions are altered to impose the potential differenceV in the x2 direction.
2.4 Homogenization in an Aligned Arrangement
In a homogeneous medium with no microstructure, we expect the potential tochange linearly in the direction of the applied potential difference. Such a φi will
19

not satisfy the cellular boundary conditions, and hence we add a small amplitudecorrection that varies on the length scale of a cell. We are led to the homogenizationansatz
φi(~x) = φ0i (~x) + εφ1
i (~x; ~x/ε) + · · · , (2.15)
where φ1i is 1 × hp × hp periodic in its second variable and is only defined when
that variable, ~x/ε, is in a 1 × hc × hc subregion of that period. We will denotethe second variable by ~y = (y1, y2, y3). Both φ0
i and φ1i are defined for all values of
the first variable ~x. Note that an equation similar to (2.15) could be obtained byguessing that the electric potential behaves like φ0
i (~x) plus a periodic disturbance.We present the homogenization ansatz instead because it parallels the derivationof the bidomain equations in [44], and also because it is applicable to more generalmacroscopic boundary conditions. For a more general treatment of homogenizationin the context of bidomain equations, see [44] or [29].
Applying the ansatz (2.15) to the PDE (2.8)–(2.14) and extracting the leadingorder terms in ε, we arrive at
−∆yφ1i (~x; ~y) = 0 in Yi = [0, 1] × [0, hc] × [0, hc],
−∂y1φ1
i (x; 1, y2, y3) − κend(φ1i (x; 1, y2, y3) − φ1
i (x; 0, y2, y3)) = ∂x1φ0
i (x),
−∂y2φ1
i (x; y1, hc, y3) − κside(φ1i (x; y1, hc, y3) − φ1
i (x; y1, 0, y3)) = ∂x2φ0
i (x),
−∂y3φ1
i (x; y1, y2, hc) − κside(φ1i (x; y1, y2, hc) − φ1
i (x; y1, y2, 0)) = ∂x3φ0
i (x),
−∂y1φ1
i (x; 0, y2, y3) = −∂y1φ1
i (x; 1, y2, y3),
−∂y2φ1
i (x; y1, 0, y3) = −∂y2φ1
i (x; y1, hc, y3),
−∂y3φ1
i (x; y1, y2, 0) = −∂y3φ1
i (x; y1, y2, hc),
where ∆y is the Laplacian in the variable ~y = (y1, y2, y3). By linearity, φ1i can be
expressed in terms of φ0i through
φ1i (~x; ~y) =
3∑
k=1
∂xkφ0
i (~x)wk(~y),
where wk solves the corrector problem
−∆ywk(~y) = 0 in [0, 1] × [0, hc] × [0, hc], (2.16)
−∂y1wk(1, y2, y3) − κend(wk(1, y2, y3) − wk(0, y2, y3)) = δk1, (2.17)
−∂y2wk(y1, hc, y3) − κside(wk(y1, hc, y3) − wk(y1, 0, y3)) = δk2, (2.18)
−∂y3wk(y1, y2, hc) − κside(wk(y1, y2, hc) − wk(y1, y2, 0)) = δk3, (2.19)
−∂y1wk(0, y2, y3) = −∂y1
wk(1, y2, y3), (2.20)
−∂y2wk(y1, 0, y3) = −∂y2
wk(y1, hc, y3), (2.21)
−∂y3wk(y1, y2, 0) = −∂y2
wk(y1, y2, hc). (2.22)
20

1
hp
hc
YiYe
Figure 2.4: The unit cell in the aligned geometry is composed of an intracellularregion Yi and an extracellular region Ye.
Here, δkj is 1 if k = j and 0 otherwise. Notice that finding each of the wk onlyrequires solving Laplace’s equation over one cell, depicted in Figure 2.4.
We can compute the effective conductivities in the longitudinal and transversediretions by applying a potential and determining the average current density. Fora voltage applied in the x1 direction, the nondimensional current within cells is
−∂x1φi(x) = −∂x1
φ0i (x) − ∂y1
φ1i (x; x/ε) +O(ε).
The average longitudinal current density across a period near ~x is thus
iavg = −∂x1φ0
i (x)1
h2p
∫ hc
0
∫ hc
0
(
1 + ∂y1w1(y1, y2, y3)
)
dy2dy3,
which is independent of y1. Hence, the effective conductivity in an aligned cellarrangement is
σi,l =σc
h2p
∫ hc
0
∫ hc
0
(
1 + ∂y1w1(y1, y2, y3)
)
dy2dy3. (2.23)
Similarly, the transverse conductivity in an aligned cell arrangement is
σi,t =σc
1 · hp
∫ hc
0
∫ 1
0
(
1 + ∂y2w2(y1, y2, y3)
)
dy1dy3 (2.24)
2.4.1 Analytical Solution to the Corrector Problem
Equations (2.16)–(2.22), which define the corrector problems for aligned geom-etry, can be solved exactly. The problems are effectively one-dimensional and their
21

solutions are
w1(~y) =1
1 + κend
(
1
2− y1
)
, (2.25)
w2(~y) =1
1 + κsideh
(
h
2− y2
)
, (2.26)
w3(~y) =1
1 + κsideh
(
h
2− y3
)
, (2.27)
of which the first is depicted in Figure 2.3b.
2.4.2 Resulting Effective Conductivities
The effective longitudinal and transverse conductivities derived from (2.23)–(2.27) are
σi,l = σc(1 − α)
(
1 − 1
1 + κend
)
, (2.28)
σi,t = σc
√1 − α
(
1 − 1
1 + κsideh
)
. (2.29)
Using the cellular parameters in Table 2.4, these become
σi,l = 1.01 mS/cm (2.30)
σi,t = 0.03 mS/cm (2.31)
2.4.3 Equivalent Resistor Network
For the aligned arrangement, the effective conductivities in the longitudinal andtransverse directions could be more easily computed by considering the cytoplasmsand gap junctions as two resistors in series. For example, the total cytosolic lon-gitudinal resistance for a tissue of length L and cross-sectional area A is 1
σc
L(1−α)A
.
The total resistance of gap junctions is Lℓ
(
1gGJ,end
w2p
A
)
. Combining these in series
and computing the effective conductivity, one can obtain (2.28). A similar calcu-lation applies for obtaining (2.29). No similar simplification will end up providingsuch an immediate calculation of longitudinal and transverse conductivities for thebrick-like arrangement we detail next.
22

l
wc
wp
φi,j+2,ki φi+1,j+2,k
i
φi,j+1,ki
φi+1,j+1,ki
φi,j,ki φi+1,j,k
i
a b
x1
x1
x2 x2
x3
Figure 2.5: The brick-like cellular architecture. Cells of dimension l×wc ×wc arearranged with a period of l×wp×wp, except that adjacent fibers are offset by halfa cell length (a). The arrangement can be viewed as the periodic extension of theregion inside the dashed prism. An x1, x2 cross-section is shown in (b).
2.5 Full Cellular Model and Homogenization in
a Brick-like Arrangement
The histology of the myocardium shows that cells in adjacent fibers are notaligned. We model this by assuming a half-cell offset between neighboring fibers,resulting in the brick-like arrangement shown in Figure 2.5.
The nondimensional analogs of (2.8)–(2.14) are tedius to write down becauseeach cell in brick geometry is adjacent to 10 cells. Using the notation ‘nbh’ todenote the correct neighbor to the (i, j, k)-th cell, (2.8)–(2.14) become
−∆φi,j,ki (~x) = 0,
−σc∂x1φi,j,k
i (1, x2, x3) =gGJ,end
w2c
(φi,j,ki (1, x2, x3) − φnbh
i (0, x2, x3)),
−σc∂x2φi,j,k
i (x1, hc, x3) =gGJ,side
lwc(φi,j,k
i (x1, hc, x3) − φnbhi (x1 + 1/2, 0, x3)),
−σc∂x3φi,j,k
i (x1, x2, hc) =gGJ,side
lwc(φi,j,k
i (x1, x2, hc) − φnbhi (x1 + 1/2, x2, 0)),
−σc∂x1φi,j,k
i (1, x2, x3) = −σc∂x1φnbh
i (0, x2, x3),
−σc∂x2φi,j,k
i (x1, hc, x3) = −σc∂x2φnbh
i (x1 + 1/2, 0, x3),
−σc∂x3φi,j,k
i (x1, x2, hc) = −σc∂x3φnbh
i (x1 + 1/2, x2, 0).
Notice that the boundary conditions in the transverse directions, y2 and y3, involvethe values of φi at the neighboring cell, offset by half a cell length. As in Section 2.3,we rescale space so that each cell is of length ε and consider a single function φi(~x)
23

defined over the interiors of all cells. The ansatz is the same as (2.15),
φi(~x) = φ0i (~x) + εφ1
i (~x; ~x/ε) + · · · ,
but now φ1i is periodic in its second variable with period ε × 2εhp × 2εhp. This
period is depicted by the dashed prism in Figure 2.5a. As before,
φ1i (~x; ~y) =
3∑
k=1
∂xkφ0
i (~x)wk(~y).
Due to a symmetry between the four cells within a period of wk,
wk(y1 + 1, y2, y3) = wk(y1, y2, y3),
wk(y1 + 1/2, y2 + hp, y3) = wk(y1, y2, y3),
wk(y1 + 1/2, y2, y3 + hp) = wk(y1, y2, y3),
where
−∆wk(~y) = 0 in [0, 1] × [0, hc] × [0, hc], (2.32)
−∂y1wk(1, y2, y3) − κend(wk(1, y2, y3) − wk(0, y2, y3)) = δk1, (2.33)
−∂y2wk(y1, hc, y3) − κside(wk(y1, hc, y3) − wk(y1 + 1/2, 0, y3)) = δk2, (2.34)
−∂y3wk(y1, y2, hc) − κside(wk(y1, y2, hc) − wk(y1 + 1/2, y2, 0)) = δk3, (2.35)
−∂y1wk(1, y2, y3) = −∂y1
wk(0, y2, y3), (2.36)
−∂y2wk(y1, hc, y3) = −∂y2
wk(y1, 0, y3), (2.37)
−∂y3wk(y1, y2, hc) = −∂y3
wk(y1, y2, 0). (2.38)
The procedure for computing effective conductivities with a brick-like arrange-ment is slightly different than that for the uniform arrangement. As a cross-sectionnormal to the x1 direction intersects cells at two different values of the local y1
coordinate, intracellular longitudinal conductivity is now
σi,l =σc
2h2p
∫ hc
0
∫ hc
0
(
2 + ∂y1w1(y1, y2, y3) + ∂y1
w1(y1 + 1/2, y2, y3))
dy2dy3. (2.39)
The solutions to equations (2.32)–(2.38) for w2 and w3 in the brick-like arrange-ment are identical to those of equations (2.16)–(2.22) in the aligned arrangement.Hence, formula (2.29) holds for the computation of σi,t in the brick-like cellulararrangement.
24

00.5
11.5
2
00.10.2
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
x1
Pot
entia
lx
2
Figure 2.6: Electric potential in several cells in a brick-like arrangement with alongitudinally applied potential gradient using cellular parameters from Table 2.4.The second and fourth cells from the left are offset by half a cell length becausethey are in a neighboring fiber to the first and third cells. The units of the verticalaxis are arbitrary. As the cellular domain is three dimensional, the plot shows onlythe x3-averaged potential within a cell.
2.5.1 Resulting Effective Conductivities
We solve (2.32)–(2.38) numerically, using a standard second-order accurate 7-point finite difference discretization of the Laplacian on a 100×10×10 cell-centeredgrid. We enforce Neumann boundary conditions by introducing ghost points andsolve the resulting linear system by Gaussian elimination. Using numerical valuesin Table 2.4, the effective intracellular conductivities are
σi,l = 1.40 mS/cm, (2.40)
σi,t = 0.034 mS/cm. (2.41)
The numerical value of σi,l agrees with that obtained on a 200× 20× 20 grid up tofive digits of accuracy. The value for σi,t is the same as formula (2.31), obtainedfrom (2.29).
Figure 2.6 shows the response in intracellular potential to a voltage applied tocells in a brick-like arrangement.
To give a sense for the conductivities that can arise from aligned and brick-likearrangements, Table 2.1 provides the values for σi,l computed for various values ofgGJ,end and gGJ,side.
25

0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 10
0.5
1
1.5
2
2.5
Fraction of Gap Junctions on Sides
Intr
acel
lula
r Lo
ngitu
dina
l Con
duct
ivity
(m
S/c
m)
Brick ArrangementAligned Arrangement
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 10
0.01
0.02
0.03
0.04
0.05
0.06
Fraction of Gap Junctions on Sides
Intr
acel
lula
r T
rans
vers
e C
ondu
ctiv
ity (
mS
/cm
)
Brick or Aligned Arrangement
Figure 2.7: Effective intracellular transverse and longitudinal conductivities asa function of the fraction of gap junctions expressed on the cell sides, keepingthe total conductance constant. As gap junctions are moved from the ends tothe sides of a cell, intracellular longitudinal conductivity decreases (a). With analigned arrangement it decreases to zero, but with the brick arrangement there is anonzero intracellular longitudinal conductivity even when all gap junctions are onthe sides of the cells. Meanwhile, in the parameter regime given by cellular mea-surements, intracellular transverse conductivity increases linearly with the fractionof gap junctions on the cell sides (b). The side-to-side and end-to-end conductancemeasurements given in Table 2.4 correspond to a fraction 0.68 of gap junctions onthe sides of cells. A uniform density of gap junctions between the sides and endsof cells with dimensions given in Table 2.4 corresponds to a fraction 0.92 of gapjunctions on the sides of cells.
σi,l gGJ,side
2.94 · 10−4 mS 5.88 · 10−4 mS 1.18 · 10−3 mS2.79 · 10−4 mS 0.55, 0.80 0.55, 1.02 0.55, 1.38
gGJ,end 5.58 · 10−4 mS 1.01, 1.22 1.01, 1.40 1.01, 1.701.12 · 10−3 mS 1.72, 1.87 1.72, 2.00 1.72, 2.23
Table 2.1: Intracellular, longitudinal conductivities σi,l (mS/cm) obtained from thealigned and brick-like arrangements for various values of gGJ,end and gGJ,side. Thenumbers before the commas are for the aligned arrangement. The numbers afterthe commas are for the brick-like arrangement.
26

2.6 Discussion
We remark that our solutions to Laplace’s equation in the aligned and brick-like arrangements have the saw-tooth nature that has been characteristic of manystudies in modeling cardiac tissue [44, 32, 27]. In the aligned arrangement withpotential applied in one of the principal directions, the profile is linear in cells,as given by the corrector functions (2.25)–(2.27). In the brick-like arrangement,the solution shown in Figure 2.6 has a piecewise nonlinear shape due to the cur-rent through the side-to-side gap junctions, even though potential is only appliedlongitudinally. We remark that the primary difference between our model andthose of Neu and Krassowska [44] and Krassowska et al. [32] is that we model gapjunctions through boundary conditions, and not as discrete geometrical entities.Recall Figure 2.1.
2.6.1 Effective Conductivity Values
Table 2.2 presents values from the literature for effective conductivities obtainedby fitting macroscopic measurements of conduction velocity to solutions of thecable or bidomain equations. Table 2.3 presents the corresponding values of ourcalculations, obtained from microscopic measurements via homogenization. Ourvalues for σi,l, σe,l, and σe,t lie within the reported ranges in Table 2.2. Our valuefor σi,t, however, is about an order of magnitude smaller than the values from thisliterature based on macroscopic measurements.
Our calculation of intracellular and extracellular effective conductivities com-pare favorably to those obtained by Stinstra et al. [51] using a much more detailedcellular architecture. Our values for σi,l, σi,t, σe,l and σe,t are within a factor of 3of those of Stinstra et. al. Such differences should be expected as Stinstra et. aluse different values for myocyte dimensions, conductivities, and other microscopicproperties.
We will now explore several possible explanations of the discrepancy in trans-verse conductivities between calculations based in macroscopic and microscopicmeasurements. Stinstra et al. propose two possible explanations for the discrep-ancy in σi,t. These are: (1) the total measured gap junctional conductivity of singlecells may be too low, and (2) the measured fraction of gap junctions located onthe sides of myocytes may be too low.
To address the first possible explanation, we use (2.29) to ask what the micro-scopic measurement of gGJ,side would have to be in order for the effective transverseintracellular conductivity to be in the range reported in Table 2.2. For a typicalvalue like σi,t = 0.2 mS/cm, the microscopic measurement of gGJ,side would have tobe 3.5 ·10−3 mS/cm, which is an order of magnitude larger than the value reportedin [56].
27

Conductivity Type Value (mS/cm) Referenceσi,l 1.6 [55]
1.7 [11]2.8 [46]3.4 [48]4.5 [30]
σi,t 0.2 [11]0.3 [46]0.6 [48]
σe,l 1.2 [48]2.2 [46]4.0 [30]5.3 [55]6.3 [11]
σe,t 0.8 [48]1.3 [46]2.4 [11]
Table 2.2: Conductivity values obtained by fitting macroscopic wavespeed data tosolutions of bidomain equations. Values are obtained for ventricular tissue fromvarious animals, such as dogs, cows, and sheep.
Conductity Type Value (mS/cm) Arrangement Equationσi,l 1.01 Aligned (2.30)
1.40 Brick-like (2.40)σi,t 0.03 Aligned or brick-like (2.31)σe,l 3.00 Aligned or brick-like (2.49)σe,t 1.56 Aligned or brick-like (2.50)
Table 2.3: Conductivity values obtained directly from microscopic measurementsvia homogenization in the present work.
28

To address the second possible explanation, we study the effects of gap junc-tional distribution on transverse extracellular conductivity. We assume that thetotal conductance of all gap junctions within a cell is 4 · gGJ,side + 2 · gGJ,end. Ifall this conductance is localized exclusively to the sides of each cell, σi,t becomes5.0 · 10−2 mS/cm, which is still much smaller than the range of values reported inTable 2.2. Thus, gap junctional distribution is not the primary explanation of thediscrepancy between σi,t calculated from microscopic parameters and the valuesfrom experimental measurements.
An additional possibility is that our microscopic model omits an importantcellular mechanism that aides transverse conductivity. Two such mechanisms in-clude ephaptic propagation [36] and electrical coupling via gap junctions betweenmyocytes and fibroblasts [31, 6]. We leave the incorporation of such mechanismsinto our model as future work.
As noted in Neu and Krassowska [44], another possible explanation of the dis-crepancy is that the experimental measurements are unable to completely distin-guish longitudinal and transverse conductivity. For example, even though Robertset al. [46] performed in situ surface recordings, the propagation pattern is influ-enced by transmural fiber rotation. The inferred value of intracellular transverseconductivity will therefore be an overestimate.
Our model of cellular architecture allows us to compare the effects of a brick-likecellular arrangement on intracellular conductivities. As mentioned in Section 2.5,the change from an aligned architecture to a brick-like one does not influence thetransverse intracellular conductivity under our model. Comparing (2.40) to (2.30),we see that a brick-like arrangement has 40% more intracellular longitudinal con-ductivity than the corresponding aligned arrangement. Note that this increase isparameter-dependent; specifically, it depends on the distribution of gap junctionsbetween the ends and the sides of the cells. As shown in Figure 2.7, if all thegap junctions are located on the sides of the cells, the intracellular longitudinalconductivity is zero in the aligned architecture but is non-zero in the brick-likearchitecture. If all gap junctions are located on the ends of the cells, there isno transverse conductivity and no difference between the brick-like and alignedarrangements. A brick-like arrangement of cells allows side-to-side coupling tocontribute to both longitudinal and transverse conductivity. In an aligned ar-rangement of cells, however, side-to-side coupling contributes only to transverseconductivity.
2.6.2 Comparison of PDE and Resistor Network Methods
Resistor network models of cardiac tissue sometimes allows analytical computa-tions of conductivity that are more elementary than those from PDE models. Thecalculation in Section 2.4.3 and the hexagonal network from Neu and Krassowska
29

[44] are both examples. To be useful, the PDE approach should provide some ben-efit. This benefit is that it allows one to investigate effects of different cell shapes,cell arrangements, and distributions of gap junctions within the ends and sides ofcells. The simple equivalent resistor network from Section 2.4.3 is only possiblebecause of the one-dimensional nature of solutions to Laplace’s equation in thealigned arrangement when voltage is applied along one of the principal directions.
Alternate cell shapes, arrangements, and gap junction distributions introduce anon-trivial dependence on multiple spatial dimensions and would make equivalentresistor networks complicated at best. One way to obtain such a network, albeitonly approximately, would be to introduce a discretization of Laplace’s equation(2.8)–(2.14). To be accurate, it would need to involve many resistors per cell.Possibly, the corrector functions for the brick-like arrangement could be adequatelyand analytically computed under a very coarse grid, but we do not pursue this ideahere.
2.6.3 Application to Electromechanical Simulations
A goal of this work is to strengthen the connection between measured cellularparameters and the effective conductivities used in whole-heart bidomain simula-tions. This connection may be particularly useful in the context of simulationscoupling cardiac electrophysiology with muscle mechanics [45]. As a region of car-diac muscle contracts, the length, width, and cross-sectional area fraction of cellswill vary. Because the effective conductivities depend on these parameters, theymay need to be recalculated at each time step in such a simulation. In Panfilov etal. [45], the authors demonstrate nontrivial electro-mechanical interactions in thesetting of fixed macroscopic conductivity tensors. Similar simulations may revealnew interactions resulting from deformation-dependent conductivities.
One complication in recomputing conductivities based on deformation is thatvolume preserving contractions of rectangular prisms do not preserve surface area.As a result, cardiac myocyte membranes presumably wrinkle during contraction.Under our model, we could view a rectangularly shaped cell with surface wrinklesas a perfect rectangular prism with a different gap junctional density. Our modelaccounts for this change in density automatically because it would distribute theprescribed conductance of gap junctions over the ends and sides of the deformedrectangular prism. Thus, our approach for computing effective conductivity tensorsis well suited for bidomain simulations which include muscle deformation.
2.7 Conclusion
This chapter attempts to find the right level of detail of cyto-architecture andgap junctional connectivity in order to produce effective conductivity values for
30

bidomain simulations based on directly measurable quantities. We propose thatthis right level of detail is obtained by modeling cells as simple prisms and gapjunctions as continuous Neumann boundary conditions to Laplace’s equation.
The strengths of our approach are that (1) the physical parameters it requiresare directly measureable; (2) it distinguishes end-to-end and side-to-side conduc-tivities, which can be separately measured; and (3) it is a mathematically naturalframework within which to view the effects of cell arrangements, geometrical pa-rameters, and the distribution of gap junctions within cells. For the cases whereit is not possible to obtain analytical solutions, numerical solutions are obtainedby solving Laplace’s equation on simple domains and with standard boundaryconditions.
The ease of computation afforded by our approach makes it attractive for use aspart of an electromechanical simulation. As cardiac muscle deforms, the effectiveconductivity tensors change and would need to be recalculated at each timestep.For such purposes, it is important to have inexpensive calculations, such as ana-lytical formulae or solutions to Laplace’s equation on simple domains.
A weakness of our approach is its inabililty to deal with random variation incyto-architecture. Although the corrector problem could be phrased over a domainconsisting of several randomly generated cells, there would be little benefit to thehomogenization point of view. The simulations of Stinstra et al. [51] indicatethat variability in cellular properties has a relatively small impact on macroscopicconductivities. Consequently, the inability to handle randomness does not severelylimit the applicability of our approach. Although, the levels of complexity of ourmodel and that of Stinstra et al. [51] are quite different, the effective conductivityvalues they predict are quite similar.
2.8 Extracellular Conductivities
In our model of myocardial geometry, the extracellular domain Ωe is transla-tionally symmetric in the longitudinal direction but is periodically perforated inthe transverse directions. Figure 2.8 shows an x2, x3 cross-section of it. Extracel-lular potential satisfies Laplace’s equation in Ωe with no-flux boundary conditions.
31

x2
x3
Figure 2.8: In this cross-section of tissue, the longitudinal direction points outof the page. The dark shaded regions indicate cells, and the remaining regionsindicate extracellular space. We treat the light shaded regions as insulators for thecalculation of transverse conductivity in the x2 direction.
Hence, effective extracellular longitudinal conductivity is found by solving
∆φe(~x) = 0 in Ωe, (2.42)
φe(0, x2, x3) = 0, (2.43)
φe(1, x2, x3) = V, (2.44)
∂x2φe(x1, 0, x3) = 0, (2.45)
∂x2φe(x1, 1, x3) = 0, (2.46)
∂x3φe(x1, x2, 0) = 0, (2.47)
∂x3φe(x1, x2, 1) = 0, (2.48)
and computing
σe,l = σextV
1A
∫∫
Ωe∩x1=c ∂x1φi dx2dx3
,
where A is the cross-sectional area of the tissue, the domain of integration is the setof exterior points of a cross-section, and the integral is independent of the constantc.
The extracellular transverse conductivity can be found by altering the boundaryconditions in (2.42)–(2.48) so that voltage is imposed and current is measured inthe x2 direction.
The response to a longitudinally applied potential gradient is linear, and hencethe effective conductivity is the extracellular conductivity scaled by the cross-sectional area fraction of extracellular space α.
32

In the case that α is small, the response to a transversely applied potentialgradient in the x2 direction is approximately linear because of the the high resis-tance of the regions indicated with light shading in Figure 2.8. In that case, theextracellular conductivity is scaled by the area fraction of extracellular space inthe transverse direction, 1 −
√1 − α. This rough calculation agrees up to O(α2)
with the derivation in [44].With the values of parameters given in Table 2.4, the extracellular longitudinal
and transverse conductivities are
σe,l = σextα = 3.00 mS/cm, (2.49)
σe,t = σext(1 −√
1 − α) = 1.56 mS/cm. (2.50)
Alternatively, σe,t could have been computed by homogenization. The extra-cellular domain Ωe is periodic, and the extracellular potential satisifes Laplacesequation with no-flux boundary conditions on the cell membranes. The calcula-tion would proceed exactly as in Section 2.4 but with the corrector
∆ywk(x; y) = 0 in Ye,
−∂νwk = ek · ν on ∂Ye,
where Ye is the periodic unit of the extracellular space depicted in Figure 2.4, ek
is the unit vector in the xk direction, and ν is the unit outer normal vector to theextracellular space.
2.9 Parameters and Variables
33

Measured ParametersSymbol Parameter Value Referenceσc conductivity of cytoplasm 6.7 mS/cm [49]σext conductivity of extracellular space 20 mS/cm [44]gGJ,end total conductance of all gap
junctions on one end of cell5.58 · 10−4 mS [56]
gGJ,side total conductance of all gapjunctions one one side of cell
5.88 · 10−4 mS [56]
l length of cell 1.58 · 10−2 cm [18]wc width of cell 2.47 · 10−3 cm [18]α volume fraction of extracellular space 0.15 [44]
Derived Parameters and VariablesSymbol Parameter Valuewp transverse period wc√
1−α
hc aspect ratio of cells wc
l
hp aspect ratio of period wp
l
κend nondimensional parameterof total end-to-end gapjunctional conductance
gGJ,endl
σcw2c
κside nondimensional parameterof total side-to-side gapjunctional conductance
gGJ,side
σcwc
~x macroscale spatial coordinates~y microscale spatial coordinatesε nondimensional cell length
relative to significant electricpotential variations in the tissue
Table 2.4: The physical parameters, derived parameters, and variables that en-ter our cellular model. Measured parameters from [56, 18] correspond to mouseventricular myocytes.
34

Chapter 3
Gap Junction Distributions for
Optimal Effective Conductivity
3.1 Introduction
Gap junctions play an important role in many kinds of cell-to-cell communi-cation. For example, they facilitate action potential propagation in both neuronsand cardiac muscle, and they allow passive diffusion between other cells. Presum-ably for the purpose of modulating communication, gap junctions can assembleinto aggregates called junctional placques. As it is difficult to distinguish individ-ual gap junctions from other membrane proteins, they are usually observed whenin placques. While quantitative correlations between observed gap junctions andmembrane permeability have been difficult to obtain (see the discussion in [8]),some simulations and analytical work suggest a resolution to this difficulty: gapjunctions aggregated into junctional placques result in a lower membrane perme-ability than when scattered across the cell membrane. Gap junctions would thusbe most influential when they are least observable.
Chen and Meng [8] study the effects of junctional placques by simulating adiffusion between two cells via a random walk. They model cells asN×N×N cubiclattices and gap junctions through special lattice points adjoining the shared face ofthe cubes, see Figure 3.1. The random walkers can move betweeen the cells only ifthey reach one of the lattice points corresponding to the gap junctions. Fixing thenumber of gap junctional lattice points, they find that scattered distributions ofgap junctions allow a group of initially localized random walkers to cross betweenthe two cells much faster than tightly packed distributions of gap junctions.
Keener and Sneyd [29] study a similar problem analytically. They describea line of cells, where each is modeled as a rectangular region. Gap junctionsare modeled as missing segments in the boundaries between neighboring cells, seeFigure 3.2. They find the density of a diffusing chemical in response to an ap-
35

gap junctionscell
a b
Figure 3.1: A simplified, two-dimensional depiction of the three-dimensional, twocell model of [8]. Each cell is represented as a cubic lattice. Gap junctions arerepresented by lattice points connecting the two cells. The gap junctions can bearranged as a single placque of adjacent vertices (a) or can be scattered randomly(b).
plied concentration difference by solving Laplace’s equation with no-flux boundaryconditions on the membrane. Due to the complexity of the domain, they writea piecewise solution involving two different eigenfunction expansions. After anextensive algebraic and numerical calculation matching the solution on the bound-aries between the subdomains, they conclude that few, large gap junctions resultin a lower effective diffusivity than many, small gap junctions.
In this chapter, we apply the modeling framework of Chapter 2 toward de-termining which microscopic distributions of gap junctions result in the largesteffective conductivity over a fiber of many cells. Our approach is similar to [29],as we model the problem by partial differential equations. Unlike [29] and [8], wedescribe gap junctions through boundary conditions, instead of as discrete geo-metrical entities. Figures 3.1, 3.2, 3.3 highlight the differences in the modelingapproaches of [8], [29], and this chapter. In our model, a non-uniform distributioncorresponds to the presence of junctional placques, whereas a uniform distributionof resistance corresponds to a scattered arrangement of gap junctions. Throughhomogenization, we find the effective conductivity of a tissue of cells with a givendistribution of gap junctions. We determine that the uniform distribution of gapjunctions provides a higher effective conductivity than distributions perturbed fromuniform.
The remainder of this chapter proceeds as follows: in Section 3.2, we provideour model for cells and gap junctional distributions. We then use homogenizationto state the optimization problem for effective conductivity. In Section 3.3, weapply formal perturbation methods to demonstrate the local optimality of theuniform gap junctional distribution. In Section 3.4, we briefly discuss a biologicalquestion raised by the optimal distribution result of this chapter.
36

gap junctionscell
a b
Figure 3.2: The cellular model of [29]. Cells in a row are modeled as rectangularregions. Gap junctions are represented as holes in the boundaries between them,making intracellular space contiguous. Keeping the total length of the gap junc-tions fixed, few large holes correspond to gap junctions aggregated into placques(a). Many small holes represent a scattered gap junctional distribution (b).
gap junctionscell
a b
Figure 3.3: Our model of cells and gap junctions. Cells in a sequence are modeledas squares whose interiors are not physically connected. Gap junctions are repre-sented in a continuous manner as resistive connections between neighboring cells.The multiple resistors shown represent a flux boundary condition between the cells.A non-uniform distribution of resistance (a) corresponds to a gap junction distri-bution with placques. A uniform distribution of resistance (b) corresponds to ascattered distribution of gap junctions.
37

ℓ
ℓ
φi−1(~x) φi(~x) φi+1(~x)
Figure 3.4: The tissue level cellular architecture. Cells are modeled as squares ad-joined without extracellular space. The thick horizonal lines represent membranesthat prevent vertical current flow. Hence, the behavior along each row of cells isidentical. An individual cell need only be identified by its horizontal cell number,i.
3.2 Model and Statement of the Gap Junction
Distribution Problem
We model cells as ℓ× ℓ squares arranged in a two-dimensional grid and ignoreextracellular space. Intracellular space is not physically contiguous. Instead, re-sistive connections allow current to flow directly between the interiors of adjacentcells. For simplicity, we assume there are no gap junctions on the top and bottomsides of cells, allowing us to focus on only a single row of cells, see Figure 3.4.
The electric potential inside cells satisfies
−∆φi(~x) = 0 in Ωi, (3.1)
where Ωi = [i · ℓ, (i + 1) · ℓ] × [0, ℓ] is the region occupied by the i-th cell, and φi
is the intracellular potential of the i-th cell. For ease of notation, we identify thedomain of the function φi with [0, ℓ]× [0, ℓ]. We denote position in a tissue of manycells by ~x = (x1, x2).
We model the gap junctions between cells in a continuous manner throughboundary conditions on equation (3.1). The current density between two neigh-boring cells is proportional to the potential difference between the positions onopposite sides of the gap junctions. We assume that the top and bottom sidesof cells are perfect insulators, as our model includes no gap junctions there. Asopposed to the work in Chapter 2, the conductivity between cells can now vary as
38

a function of position along the cell ends. Consequently, the boundary conditionsfor (3.1) are
−σc∂x1φi(ℓ, x2) = gGJ,end
1
ℓf(x2/ℓ)
(
φi(ℓ, x2) − φi+1(0, x2))
, (3.2)
−σc∂x1φi(ℓ, x2) = −σc∂x1
φi+1(0, x2), (3.3)
−σc∂x2φi(x1, ℓ) = 0, (3.4)
−σc∂x2φi(x1, 0) = 0, (3.5)
where σc is the two-dimensional cytoplasmic conductivity (mS), gGJ,end is the con-ductance (mS) of all gap junctions on one end of a cell, and f is a non-negativefunction specifying the distribution of that conductance along the cell ends. We re-quire
∫ 1
0f(x2)dx2 = 1 to enforce a fair comparison between different distributions
of gap junctions.
3.2.1 Nondimensionlization and Homogenization
We rescale space so that cells are of length ε by letting
~x =ε
l~x.
Dropping the tildes, equations (3.1)–(3.5) become
−∆φi(~x) = 0 in [0, ε] × [0, ε], (3.6)
−∂x1φi(ε, x2) =
f(x2/ε)
εκ
(
φi(ε, x2) − φi+1(0, x2)
)
, (3.7)
−∂x1φi(0, x2) = −∂x1
φi(ε, x2), (3.8)
−∂x2φi(x1, ε) = 0, (3.9)
−∂x2φi(x1, 0) = 0, (3.10)
where κ =gGJ,end
σcis the nondimensional, total end-to-end gap junctional conduc-
tance, and the intracellular domain of the i-th cell is [0, ε] × [0, ε].Consider a cube of tissue of dimensionless size 1×1, composed of cells arranged
with period ε× ε. The index notation distinguishing the i-th cell is cumbersome,and we introduce φ(~x) as a single function defined over Ω = ∪iΩ
i. The function φsatisfies the PDE (3.6) within each cell, and microscopic boundary conditions akinto (3.7)–(3.10) on the boundary of each cell. With an applied potential, φ will also
39

satisfy that macroscopic boundary conditions
φ(0, x2) = 0,
φ(1, x2) = V,
∂x2φ(x1, 0) = 0,
∂x2φ(x1, 1) = 0.
The nondimensional intracellular effective conductivity is given by
σnondimeff =
1L
∫
Ω∩x1=c ∂x1φ dx2
V, (3.11)
where L = 1 is the cross-sectional length of the tissue, the domain of integrationis the set of interior points across a cross-section, and the integral is independentof the constant c.
Similarly to the derivation in Chapter 2, we derive the effective conductivityby making the homogenization ansatz
φ(~x) = φ0(~x) + εφ1(~x; ~x/ε) + · · · , (3.12)
where φ1 is 1 × 1 periodic in its second variable, which we call ~y = (y1, y2). Bothφ0 and φ1 are defined for all values of the first variable, ~x.
Considering a potential imposed purely in the x1 direction, applying the ansatz(3.12) to the PDE system (3.6)–(3.10), and proceeding as in Chapter 2, we obtain
φ1(~x; ~y) = ∂x1φ0(~x)w(~y),
where w solves the so-called corrector problem
−∆yw(~y) = 0 in [0, 1] × [0, 1], (3.13)
−∂y1w(1, y2) − κf(y2)(w(1, y2) − w(0, y2)) = 1, (3.14)
−∂y1w(0, y2) = −∂y1
w(1, y2), (3.15)
−∂y2w(y1, 1) = 0, (3.16)
−∂y2w(y1, 0) = 0. (3.17)
The effective nondimensional conductivity is given by
σnondimeff =
∫ 1
0
(
1 + ∂y1w(y1, y2)
)
dy2,
which is independent of y1.
40

3.2.2 Statement of the Gap Junction Distribution Problem
The gap junction distribution problem can now be stated as follows: over theset of sufficiently regular f ’s such that
∫ 1
0f(y2)dy2 = 1, which one maximizes
∫ 1
0
(
1 + ∂y1w(y1, y2)
)
dy2, where w is determined from f by solving (3.13)-(3.17)?In the following section, we partially answer the question by using perturba-
tion and Fourier analysis to show that the uniform distribution f ≡ 1 is a localmaximizer of conductivity. This result is consistent with the work of [8] and [29].
3.3 Local optimality of a uniform gap junctional
distribution
In this section, we establish for all sufficiently regular g 6≡ 0 satisfying theconstraint
∫ 1
0g(y2)dy2 = 0, that
d
dǫ
∫ 1
0
(
1 + ∂y1wǫ
g(y1, y2))
dy2
∣
∣
∣
ǫ=0= 0, (3.18)
d2
dǫ2
∫ 1
0
(
1 + ∂y1wǫ
g(y1, y2))
dy2
∣
∣
∣
ǫ=0< 0, (3.19)
where wǫg is the corrector function for the problem with gap junctional density
f(y2) = 1+ǫg(y2). Note that the variable ǫ is a measure of perturbation magnitudeand should not be confused with ε, the dimensionless size of cells in Section 3.2.1.Precisely, wǫ
g satisfies
−∆ywǫg(~y) = 0 in [0, 1] × [0, 1], (3.20)
−∂y1wǫ
g(1, y2) − κ(1 + ǫg(y))(wǫg(1, y2) − wǫ
g(0, y2)) = 1, (3.21)
−∂y1wǫ
g(0, y2) = −∂y1wǫ
g(1, y2), (3.22)
−∂y2wǫ
g(y1, 1) = 0, (3.23)
−∂y2wǫ
g(y1, 0) = 0. (3.24)
To establish (3.18)–(3.19), we consider the perturbation expansion
wǫg(~y) = w0(~y) + ǫw1
g(~y) + ǫ2w2g(~y) + · · · . (3.25)
It suffices to show that∫ 1
0
∂y1w1
g(y1, y2)dy2 = 0, (3.26)
∫ 1
0
∂y1w2
g(y1, y2)dy2 < 0. (3.27)
41

Plugging the expansion (3.25) into (3.20)-(3.24) and equating like powers of ǫ, wearrive at the following systems. The O(1) system for w0 is
−∆yw0(~y) = 0 in [0, 1] × [0, 1], (3.28)
−∂y1w0(1, y2) − κ(w0(1, y2) − w0(0, y2)) = 1, (3.29)
−∂y1w0(0, y2) = −∂y1
w0(1, y2), (3.30)
−∂y2w0(y1, 1) = 0, (3.31)
−∂y2w0(y1, 0) = 0. (3.32)
The O(ǫ) system for w1g is
−∆yw1g(~y) = 0 in [0, 1] × [0, 1], (3.33)
−∂y1w1
g(1, y2) − κ(w1g(1, y2) − w1
g(0, y2)) = κg(y2)(
w0(1, y2) − w0(0, y2))
, (3.34)
−∂y1w1
g(0, y2) = −∂y1w1
g(1, y2), (3.35)
−∂y2w1
g(y1, 1) = 0, (3.36)
−∂y2w1
g(y1, 0) = 0. (3.37)
Finally, the O(ǫ2) system for w2g is
−∆yw2g(~y) = 0 in [0, 1] × [0, 1], (3.38)
−∂y1w2
g(1, y2) − κ(w2g(1, y2) − w2
g(0, y2)) = κg(y2)(
w1g(1, y2) − w1
g(0, y2))
, (3.39)
−∂y1w2
g(0, y2) = −∂y1w2
g(1, y2), (3.40)
−∂y2w2
g(y1, 1) = 0, (3.41)
−∂y2w2
g(y1, 0) = 0. (3.42)
As all three boundary value problems have the same form, we pause for aproposition providing the solution to such equations.
Proposition 3.1. Let u(y1, y2) solve
−∆yu(~y) = 0 in [0, 1] × [0, 1], (3.43)
−∂y1u(1, y2) − κ(u(1, y2) − u(0, y2)) = h(y2), (3.44)
−∂y1u(0, y2) = −∂y1
u(1, y2), (3.45)
−∂y2u(y1, 1) = 0, (3.46)
−∂y2u(y1, 0) = 0. (3.47)
Then, u can be written as
u(y1, y2) =1
1 + κh(0)
(
1
2− y1
)
+ c0 +
∞∑
n=1
cn(y1) cos(nπy2), (3.48)
cn(y1) = an sinh(nπy1) + bn cosh(nπy2), (3.49)
42

where h(y2) =∑∞
n=0 h(n) cos(nπy2), and an, bn satisfy
an(−nπ − κ sinh(nπ)) + bn(−κ cosh(nπ) + κ) = h(n), (3.50)
an(1 − cosh(nπ)) − bn sinh(nπ) = 0. (3.51)
We note that the additive constant c0 in (3.48) is arbitrary and irrelevant tothe effective conductivity calculation, so we choose it to be zero.
We now show (3.26) holds. We remark that the only component of h in (3.44)that effects the calculation of effective conductivity is its average, h(0). Hence, forestablishing (3.26) it suffices to show
∫ 1
0
κg(y2)(
w0(1, y2) − w0(0, y2))
= 0, (3.52)
which holds by plugging the exact solution for w0,
w0(~y) =1
1 + κ
(
1
2− y1
)
, (3.53)
into (3.28)-(3.32).We now show (3.27) holds. It suffices to show that the right hand side in (3.39)
has positive average. We let h2(y2) be this right hand side,
h2(y2) = κg(y2)(w1g(1, y2) − w1
g(0, y2)). (3.54)
Applying Proposition 3.1 to (3.33)–(3.37), we write w1g as
w1g(y1, y2) =
∞∑
n=1
cn(y1) cos(nπy2). (3.55)
Finally, we compute
∫ 1
0
h2(y2)dy2 = κ
∞∑
n=1
(
cn(1) − cn(0))
g(y2) cos(nπy2) (3.56)
= κ∞
∑
n=1
1
2g(n)
(
cn(1) − cn(0))
(3.57)
=κ2
1 + κ
∞∑
n=1
g(n)2 sinh(nπ/2)
nπ cosh(nπ/2) + 2κ sinh(nπ/2)(3.58)
> 0. (3.59)
The value of cn(1) − cn(0) that permits the last equality comes from the explicitform of w1
g given by Proposition 3.1.
43

3.4 Discussion
While the optimal distribution result we obtain is only local in nature, weexpect the uniform distribution to be a global optimizer for conductivity under afixed number of gap junctions. A natural question, then, is why do gap junctionsform placques at all? As Chen and Meng [8] suggest, it is conceivable that cellsmodulate their conductivity by forming or breaking these placques. That is, cellscould place gap junctions together precisely when they seek to minimize theireffect. This idea is intriguing as isolated gap junctions are difficult to detect, and,hence, they would be least observable when they are most influential.
The mathematics in this chapter is woefully blind to the biological complica-tions of gap junction production. In addition to synthesizing connexin proteins,hemi-channels on the plasma membrane of adjacent cells need to find each otherin order to form a gap junction. Further mathematical modeling could be usefulin ascertaining whether placques aid such pairing. If they did, cells may be willingto give up some effective conductivity due to gap junctional distribution in orderto maximize their hemi-channel pairing into full gap junctions.
44

Chapter 4
Homogenization of a Model for
Ephaptic Cardiac Communication
4.1 Introduction
Heart cells communicate electrically to coordinate the muscular contractionthat pumps blood. Ions, such as Na+ and K+, pass in and out of cells throughspecial channels, eventually causing a release of Ca2+ that initations this contrac-tion. It is well accepted that gap junctions provide the primary conduit for thiscommunication between myocytes [47].
There is some evidence, though, that gap junctions do not tell the completestory of cellular coupling during action potential propagation in the heart. Forexample, hearts of lower vertebrates lack gap junctions yet still exhibit chainsof electrical excitation [50]. Additionally, a gene knockout study revealed thatConnexin-43 deficient mice still have cardiac conduction, albeit at a reduced speed[20]. A final observation suggesting that gap junctions are not the exclusive mech-anism of cardiac electrical communication is the preferential localization to inter-calated disks of Na+ channels in rat ventricular myocytes [33]. Considering thenarrow cleft between cells, it seems risky for the major depolarizing current to beplaced there. These clefts are much more likely than extracellular space to exhibitlarge fluctuations in ion concentrations, altering the reversal potentials that driveionic current. Further, any ion channels connecting to a cleft have their effectreduced by the high cleft-to-extracellular-space resistance.
Each of these observations suggests the existence of an electrical communicationmechanism not mediated by gap junctions. Sperelakis and McConnell [50] detailseveral possibilities including capacitive coupling, the electric field mechanism, andion accumulation. We focus on the electric field mechanism, also known as ephapticcommunication, which is depicted in Figure 4.1. In this mechanism, the inwardNa+ current of the activated pre-junctional membrane produces a cleft potential
45

(a) (b) (c)
φ = 0
Na+ Channels Na+ Current Na+ Current
φ < 0 φ < 0
Figure 4.1: A cartoon of the ephaptic mechanism between two adjacent cells(shaded). Initially, the Na+ channels of both the pre-junctional (left) and post-junctional (right) cells are closed (a). When an action potential reaches the leftcell, its Na+ channels open, allowing a flux of Na+ current in from extracellularspace via the clefts (b). As per Ohm’s law, the potential inside the cleft decreases,resulting in the depolarization of the post-junctional membrande. If this effect issufficiently strong, this membrance may reach threshold, causing the Na+ channelsto open and allowing current to flow inward (c).
below that of extracellular space. This lowered cleft potential can then depolarizethe post-junctional membrane, continuing the action potential propagation.
Kucera et al. [33] study action potential propagation along a strand of discretecells with active membranes, coupled by gap junctions and a cleft potential. Intheir model, current can flow into and out of the cleft by a resistive connection togrounded extracellular space. Their model does not take into account effects ofchanges in ion concentration. Their numerical simulations show that the ephapticmechanism can act to speed up conduction velocity relative to analogous non-ephaptic models. In order to see this effect, though, Na+ channels must be localizedto the intercalated disks at the ends of cells, and gap junction expression levels mustbe significantly reduced from their physiologically normal values. Their simulationsalso show that under normal gap junctional coupling, the ephaptic mechanism actsto slow conduction.
Mori et al. [36] show similar results with three dimensional models accountingfor ion concentration effects within clefts. With normal gap junction expression,relocating Na+ channels to the intercalated disks can slow conduction if the cleftsare sufficiently narrow. These authors also describe a new mode of cardiac com-munication where the ephaptic and gap junctional mechanisms alternate as theprincipal means of communication across successive junctions.
In this chapter, we derive a homogenized partial differential equation for anephaptic model almost identical to that in [33]. We then investigate numericallyhow well it captures the ephaptic effects described in that paper. We detail thedependence of conduction velocity on gap junctional coupling, Na+ channel lo-
46

calization, and cleft-to-ground resistance. We observe that the homogenized andfull descriptions of the ephaptic effect agree well when gap junctions are at phys-iologically normal levels. They disagree, however, when gap junction levels aresubstantially below normal.
The remainder of the chapter is organized as follows. In Section 4.2, we presentour microscopic model for ephaptic conduction, write down the correspondingPDEs from the biophysics, and nondimensionalize them. In Section 4.3, we de-rive an effective, macroscale PDE for electric potential by homogenization. InSection 4.4, we present results from numerical simulations, and, in Section 4.5,we discuss the parameter regimes where the homogenization does and does notcapture the important dynamics of the full system. In Section 4.6, we present themeasured and derived parameters that enter our model. Finally, in Section 4.7, wepresent the details of the numerical schemes used in Section 4.4.
4.2 The Full Ephaptic Model
We model cells as one-dimensional cables, each corresponding to a cylinder ofradius r and length ℓ. We model extracellular space as grounded. We describe theelectric potential inside cells through the cable equation
Aσc∂2φi
∂x2(t, x) = S
(
Cm∂φi
∂t(t, x) + Iion
(
φi(t, x), wiside(t, x)
)
)
in Ωi, (4.1)
where Ωi = [iℓ, (i + 1)ℓ] is the space occupied by the i-th cell, φi(t, x) is theintracellular potential within the i-th cell, A = πr2 is the cross-sectional area ofthe cell, σc is the conductivity of the cytoplasm, S = 2πr is the perimeter of a crosssection, Cm is the capacitance per unit area of membrane, Iion is the outward ioniccurrent per unit area of membrane, and wi are the relevant gating variables alongthe i-th cell. For ease of notation, we identify the domain of φi with [0, ℓ]. We willalso omit the reference to gating variables like wi whenever convenient. Resistorsrepresenting gap junctions allow current to flow directly between adjacent cells.We model the cleft between cells by a single potential with a resistive connectionto the extracellular space, which we refer to as the cleft resistance. This choice ofa one noded cleft potential is the primary structural difference between our modeland that of [33]. We account for the capacitive and ionic current from the cell endsinto the cleft, see Figure 4.2. Note that we ignore any effects of ion concentrationschanging within the clefts.
The current balance into the cleft provides the following equation for the cleftpotential and boundary conditions for (4.1):
47

φi φi+1φic
Figure 4.2: The geometry and circuit diagram for our model of an ephaptic mecha-nism. We describe cells as active cables coupled through direct resistive connectionsand active membranes involving shared cleft potentials. We model extracellularspace as grounded. We ignore the effects of any changes in ion concentration.
−Aσc∂φi
∂x(t, ℓ) + Aσc
∂φi+1
∂x(t, 0) =
1
Rcφi
c(t), (4.2)
−Aσc∂φi
∂x(t, ℓ) −AgGJ,end
(
φi(t, ℓ) − φi+1(t, 0))
= A
(
Cm∂
∂t(φi(t, ℓ) − φi
c) + Iion(
φi(t, ℓ) − φic(t), w
iright(t)
)
)
,
(4.3)
Aσc∂φi+1
∂x(t, 0) + AgGJ,end
(
φi(t, ℓ) − φi+1(t, 0))
= A
(
Cm∂
∂t(φi+1(t, 0) − φi
c) + Iion(
φi+1(t, 0) − φic(t), w
ileft(t)
)
)
,
(4.4)
where Rc is the cleft resistance, φi is the potential of the cleft to the right of thei-th cell, gGJ,end is the conductance per unit area of the gap junctions, and wi
left andwi
right are gating variables for the left and right sides of the i-th cell, respectively.
4.2.1 Nondimensionalization of the Full System
As discussed in [44], significant variations of electric potential in a tissue occuron the length scale of the so called liminal length,
√Rmσcℓ, where Rm is the
sheet resistance of the cell membrane. We introduce the dimensionless variable
48

ε =√
ℓRmσc
and rescale space so that the liminal length is 1. That is, we let
x =x√
Rmσcℓ=
x
ℓ/ε.
Individual cells are now of length ε. We rescale time by the membrane relaxationconstant,
t =t
RmCm
,
and potential by a typical voltage,
φ =φ
φtypical.
Finally, we rescale the ionic current by
Iion =Rm
φtypicalIion.
Introducing the nondimensional parameters
α =A
Sd,
β = ε2
(
Aσc
d
)
Rc,
κ = ε2gGJ,endRm,
and dropping the tildes, (4.1)–(4.4) become
α∂2φi
∂x2(t, x) =
∂φi
∂t(t, x) + Iion
(
φi(t, x), wiside(t, x)
)
, (4.5)
β
ε
(
−∂φi
∂x(t, ε) +
∂φi+1
∂x(t, 0)
)
= φic, (4.6)
−∂φi
∂x(t, ε) − κ
ε
(
φi(t, ε) − φi+1(t, 0))
= ε( ∂
∂t
(
φi(t, ε) − φic(t)
)
+ Iion(
φi(t, ε) − φic, w
iright(t)
)
)
,
(4.7)
∂φi+1
∂x(t, 0) +
κ
ε
(
φi(t, ε) − φi+1(t, 0))
= ε( ∂
∂t
(
φi+1(t, 0) − φic(t)
)
+ Iion(
φi+1(t, 0) − φic, w
ileft(t)
)
)
.
(4.8)
Note in particular that for β to remain of order 1 as ε→ 0, Rc must scale like ε−2.We subsequently refer to (4.5)-(4.8) as the full ephaptic model, or as the full
ephaptic system.
49

4.3 The Homogenized Ephaptic Model
We now perform homogenization to derive effective, macroscale evolution equa-tions for the intracellular and cleft potentials. The index notation distinguishingthe i-th cell is cumbersome, and we introduce φ(t, x) and wside(t, x) as single func-tions defined over Ω = ∪iΩ
i. We also introduce the functions φc(t, x), wleft(t, x),and wright(t, x), whose discrete samplings provide the cleft potentials and the gatingvariables for the left and right sides of cells.
We make the homogenization ansatz
φ(t, x) = φ0(t, x) + εφ1(t, x, x/ε) + ε2φ2(t, x, x/ε) +O(ε3). (4.9)
We let y denote the third argument of φ1 and φ2, x/ε, known as the fast spatialvariable. For cleft potentials, we assume φc is a function of the slow variable x.That is,
φc(t, x) = φc,0(t, x) + · · · , (4.10)
where higher order terms in ε will not enter our derivation. Similarly, we assumethat gating variables are only a function of time and of the slow spatial variable.That is,
wside(t, x) = w0,side(t, x) +O(ε), (4.11)
wleft(t, x) = w0,end(t, x) +O(ε), (4.12)
wright(t, x) = w0,end(t, x) +O(ε). (4.13)
Note that the gating variables on the left and right ends of a cell are assumed tobe equal. These assumptions of slow spatial variation in cleft potential and gatingvariables are strong and likely to be violated in the case of gap junction deficientmice. Nonetheless, they provide a starting point for mathematical analysis andcould conceivably be weakened in future work.
In this section, we show that the effective equations satisfied by φ0(t, x), φc,0(t, x),w0,side(t, x), and w0,end(t, x) are
α
(
1 − 1
1 + κ
)
∂2φ0
∂x2=∂φ0
∂t+ Iion
(
φ0, w0,side
)
+ 2α∂
∂t
(
φ0 − φc,0
)
+ 2αIion(
φ0 − φc,0, w0,end
)
, (4.14)
φc,0 = β
(
2∂
∂t
(
φ0 − φc,0
)
+ 2Iion(
φ0 − φc,0, w0,end
)
)
. (4.15)
We subsequently refer to (4.14)-(4.15) as the homogenized ephaptic model, or asthe homogenized system.
50

4.3.1 Derivation of Homogenized System
We now derive the effective equations (4.14)-(4.15). Applying the ansatz (4.9)to the PDE (4.5), (4.7), (4.8) and extracting the leading terms, of order ε−1, weobtain
∂2φ1
∂y2(t, x, y) = 0 in [0,1],
−∂φ1
∂y(t, x, 1) − κ
(
φ1(t, x, 1) − φ1(t, x, 0))
= −∂φ0
∂x(t, x),
∂φ1
∂y(t, x, 0) + κ
(
φ1(t, x, 1) − φ1(t, x, 0))
=∂φ0
∂x(t, x).
Solving, we obtain
φ1(t, x, y) =1
1 + κ
(
1
2− y
)
∂φ0
∂x(t, x). (4.16)
The next terms, of order ε0, are
α
(
∂2φ0
∂x2(t, x) + 2
∂2φ1
∂x∂y(t, x, y) +
∂2φ2
∂y2(t, x, y)
)
=∂φ0
∂t(t, x) + Iion
(
φ0(t, x), w0,side(t, x))
, (4.17)
− ∂φ2
∂y(t, x, 1) − κ
(
φ2(t, x, 1) − φ2(t, x, 0))
=∂φ1
∂x(t, x, 1) +
∂
∂t
(
φ0(t, x) − φc,0
)
+ Iion(
φ0(t, x) − φc,0(t, x), w0,end(t, x))
, (4.18)
∂φ2
∂y(t, x, 0) + κ
(
φ2(t, x, 1) − φ2(t, x, 0))
= −∂φ1
∂x(t, x, 0) +
∂
∂t
(
φ0(t, x) − φc,0
)
+ Iion(
φ0(t, x) − φc,0(t, x), w0,end(t, x))
. (4.19)
The solvability condition for φ2 provides the effective dynamics for φ0. We obtainit by integrating the differential equation (4.17) from y = 0 to y = 1. Thus,
α∂2φ0
∂x2+ 2α
∂φ1
∂x
∣
∣
∣
y=1
y=0+ α
∂φ2
∂y(t, x, y)
∣
∣
∣
y=1
y=0=∂φ0
∂t(t, x) + Iion(φ0(t, x), w0,side(t, x)).
(4.20)
51

From the boundary conditions (4.18)–(4.19), we observe
∂φ2
∂y(t, x, y)
∣
∣
∣
y=1
y=0= −∂φ1
∂x(t, x, 1) +
∂φ1
∂x(t, x, 0)
− 2∂
∂t
(
φ0(t, x) − φc,0(t, x))
− 2Iion(
φ0(t, x) − φc,0(t, x), w0,end(t, x))
(4.21)
=1
1 + κ
∂2φ0
∂x2− 2
∂
∂t
(
φ0(t, x) − φc,0(t, x))
− 2Iion(
φ0(t, x) − φc,0(t, x), w0,end(t, x))
. (4.22)
To get the homogenized equation (4.14) for the dynamics of φ0, we combine (4.20),(4.16), and (4.22).
All that remains is to obtain the governing equation for φc,0. Plugging theansatz (4.9) into (4.6) and extracting the leading order terms, of size, ε0, we obtain
β
(
−∂φ1
∂x(t, x, 1) +
∂φ1
∂x(t, x, 0) − ∂φ2
∂y(t, x, 1) +
∂φ2
∂y(t, x, 0)
)
= φc,0. (4.23)
Applying (4.16) and (4.22), this equation simplifies to (4.15), which indicates anontrivial cleft contribution to the dynamics.
4.4 Numerical Simulation of the Full and Ho-
mogenized Models
We now compare the full ephaptic model to its homogenization via numericalsimulation. We would like (1) to replicate the effects of the ephaptic mechanism re-ported in [33], and (2) to determine the conditions under which the homogenizedmodel accurately captures these effects. Specifically, we investigate the depen-dence of the conduction speed on the cleft-to-ground resistance, the degree of gapjunctional coupling, and the degree of localization of Na+ channels to the interca-lated disks. For physiological relevance, we investigate these questions using theLuo-Rudy 1991 ion channel dynamic [34].
See Section 4.6 for the values of the nondimensional parameters that correspondto the physical parameters in the simulations from [33]. In our simulation for thefull system, we use ε = 0.01 instead of 0.0087 for geometric convenience. As thevalue of κ corresponding to normal expression levels of gap junctions is almostunity, we may interpret κ as the fraction of gap junctions expressed relative tophysiologically normal conditions. We run simulations for κ = 1 to κ = 0.01 andrefer to κ as the gap junctional coupling strength.
52

To determine the effect of the Na+ channel distribution within a cell, we fix thetotal number of Na+ channels in a cell at its normal value but distribute them suchthat either 100% are located at the cell ends, such that 50% are at the ends, orsuch that the density is uniform. We denote the fraction of Na+ channels locatedat the cell ends as fNa. These three cases correspond to fNa = 1, 0.5, or 2α
1+2α,
respectively, where α = 0.055 is the ratio of the ratio of a cell’s cross sectional andside areas. Sometimes we refer to the last case as fNa = uniform.
The last parameter we investigate is β, the nondimensional coupling the cleftto the extracellular space. We run simultions from β = 10−4 to β = 5 ·10−3, whichcorrespond to clefts of width 114 nm to 2 nm, see Section 4.6.
We define the position of a traveling action potential as the x-coordinate wherethe potential crosses −40 mV and compute conduction velocity by finding thedistance the front travels between nondimensional times 1 and 1.5. If the travelingwave reaches position x = 1.25 at a time t < 1.5, we coupute conduction velocityby finding the distance the front travels between nondimensional times 2
3t and t.
4.4.1 Initial Value Problems for Numerical Simulation
In this section, we detail the initial value problems (IVPs) used to simulate theephaptic effect for the full and homogenized models.
4.4.1.1 Initial Value Problem for the Full System
We simulate the full model of the ephaptic mechanism on a non-periodic domainof length L composed of cells of length ε, see Figure 4.3.
The initial value problem is
α∂2φi
∂x2(t, x) =
∂φi
∂t(t, x) + Iion, side
(
φi(t, x), wiside(t, x)
)
in Ωiε, (4.24)
−∂φi
∂x(t, ε) − κ
ε
(
φi(t, ε) − φi+1(t, 0))
= ε
(
∂
∂t
(
φi(t, ε) − φic(t)
)
+ Iion, end
(
φi(t, ε) − φic, w
iright(t)
)
)
, (4.25)
∂φi
∂x(t, 0) +
κ
ε
(
φi−1(t, ε) − φi(t, 0))
= ε
(
∂
∂t
(
φi(t, 0) − φi−1c (t)
)
+ Iion, end
(
φi(t, 0) − φi−1c , wi
left(t))
)
,
(4.26)
β
ε
(
−∂φi
∂x(t, ε) +
∂φi+1
∂x(t, 0)
)
= φic, (4.27)
53

φi(t, x) φic(t) φi+1(t, x)
(a)
(b)
φ0(t, x), φc,0(t, x)
L
ε
Figure 4.3: A schematic of the domains for the full (a) and homogonized (b) models.The full model is posed over discrete cells of length ε with an equipotential cleftbetween adjacent cells. The intracellular potential within the i-th cell is φi(t, x),and the cleft potential to the right of the i-th cell is φi
c(t). Note that the cleftpotential is defined only over a discrete set of points. The homogenized model isposed over the entire length Ltissue of tissue, as it does not resolve individual cells.The intracellular potential φ0(t, x) and the cleft potential φc,0(t, x) are defined overthe entire domain.
Iion, side(φ, w) = (1 − fNa)(1 + 2α) · INa(φ, w) + Isi(φ, w)
+ IK(φ, w) + IK1(φ, w) + IKp(φ, w) + Ib(φ, w), (4.28)
Iion, end(φ, w) = fNa(1 +1
2α) · INa(φ, w) + Isi(φ, w)
+ IK(φ, w) + IK1(φ, w) + IKp(φ, w) + Ib(φ, w), (4.29)
dwis
dt= LRD(φtrans,s, w
is) for s = side, left,right, (4.30)
φi(t = 0, x) =
φinit 1 ≤ i ≤ linit/ε,
φrest o’wise,(4.31)
φic(t = 0) = 0, (4.32)
wis(t = 0) = wrest for s = side,left,right (4.33)
where Ωiε = [(i−1)ε, iε] represents the i-th cell; φi(t, x) is the intracellular potential
at time t and position x within the i-th cell; φc(t) is the cleft potential to the rightof the i-th cell; wi
left(t), wiright(t), and wi
side(t, x) are the Luo-Rudy gating variablesfor the i-th cell at the left end, the right end, and at position x along the sideof the cell, respectively; and fNa is the fraction of a cell’s Na+ channels that arelocalized to the intercalated disks. The function LRD specifies the evolution of the
54

gating variables and is defined in Section 1.1.2. In (4.28) and (4.29), INa, IK, IK1,IKp, and Ib are the six currents in the Luo-Rudy 1991 dynamic. For convenience,in (4.30), we denote the transmembrane potential jump at the left end, the rightend, and at a position x along the sides of the i-th cell by
φitrans,s =
φi(t, x) if s = side,
φi(t, ε) − φic if s = right,
φi(t, 0) − φi−1c if s = left.
(4.34)
The initial values correspond to a region of length linit beginning at the superthresh-old potential φinit in order to initiate a traveling action potential. All clefts areinitially grounded. See Section 4.6 for the parameters we use in the simulationsand the dimensional values they correspond to.
We discretize the spatial derivatives in (4.24)–(4.27) over a cell-centered gridwith spacing ∆x, introducing two ghost points per biological cell, one at each end,to facility the numerical treatment of the boundary conditions. At each timestep∆t, we compute φ using a second order Runge-Kutta scheme, treating the Iionterms explicitly and the remaining terms implicitly. We solve the linear systemsat each timestep by Gaussian Elimination. See Section 4.7 for the details of thediscretized system and for an empirical convergence study verifying second orderaccuracy in space and first order accuracy in time. For specific parameter choices,such as the spatial grid size ∆x and the timestep ∆t, see Table 4.2 in Section 4.6.
4.4.1.2 Initial Value Problem for the Homogenized System
We simulate the homogenized model of the ephaptic mechanism on a non-periodic domain of length L. With this model, there is no longer a notion of celllength, as the equation was drived for the limit ε → 0. We solve the homogenizedequations over the domain [0, L], as depicted in Figure 4.3b.
Instead of phrasing the initial value problem in terms of φ0 and φc,0, we in-troduce ψ0(t, x) = φ0(t, x) − φc,0(t, x), the intracellular-to-cleft transmembranepotential. Rewriting the homogenized equations in terms of φ0 and ψ0, the initial
55

value problem corresponding to the full system (4.24)–(4.33) is
∂φ0
∂t= α
(
1 − 1
1 + κ
)
∂2φ0
∂x2− Iion, side(φ0, wside) −
α
βφ0 +
α
βψ0 in [0, L],
(4.35)
∂ψ0
∂t= −Iion, end(ψ0, wend, fNa
α
1 + 2α) +
1
2βφ0 −
1
2βψ0 in [0, L], (4.36)
∂xφ0(t, 0) = 0, (4.37)
∂xφ0(t, L) = 0, (4.38)
dws
dt(t, x) = LRD(φtrans,s(t, x), ws(t, x)) for s = side, end, (4.39)
Iion, side(φ, w) = (1 − fNa)(1 + 2α) · INa(φ, w) + Isi(φ, w)
+ IK(φ, w) + IK1(φ, w) + IKp(φ, w) + Ib(φ, w), (4.40)
Iion, end(φ, w) = fNa(1 +1
2α) · INa(φ, w) + Isi(φ, w)
+ IK(φ, w) + IK1(φ, w) + IKp(φ, w) + Ib(φ, w), (4.41)
φ0(t = 0, x) =
φinit 1 ≤ x ≤ linit,
φrest o’wise,(4.42)
ψ0(t = 0, x) = 0, (4.43)
where wside(t, x) and wend(t, x) are the Luo-Rudy gating variables for the sides andends of cells around position x. The transmembrane potential φtrans is given by
φtrans,s =
φ0(t, x) if s = side,
ψ0(t, x) if s = end.(4.44)
As with the IVP for the full system, fNa is the fraction of Na+ channels localizedto the intercalated disks, and LRD specifies the evolution of the gating variablesunder the Luo-Rudy 1991 dynamic. As with the IVP for the full system, the intialvalues correspond to a region of linit being at a superthreshold potential, initiatinga traveling action potential. See Section 4.6 for the parameters we use in thesimulations and the dimensional values they correspond to.
Note that a consequence of the homogenization is that all dependent variables,φ0, ψ0, wside and wend are defined for all spatial gridpoints. Their values are to beinterpreted as local spatial averages. In particular, the homogenization assumedthat gating variables on opposite sides of a cleft must be equal.
We discretize the only spatial derivative, in (4.35), to second order by thestandard 3-point stencil. As with the simulation of the full system, we used asecond order Runge-Kutta scheme, treating the Iion terms explicitly and the other
56

terms implicitly. We solve the corresponding linear systems at each time step byGaussian Elimination. See Section 4.6 for details of the discretized system and foran empirical convergence study verifying second order accuracy in time and firstorder accuracy in space. For specific parameter choices, such as the spatial gridsize ∆x and the timestep ∆t, see Table 4.2 in Section 4.6.
4.4.2 Numerical Results for the Full System
Figure 4.4 shows the conduction speed of the full ephaptic model for selectedvalues of the cleft-to-ground resistance, gap junctional coupling strength, and de-gree of Na+ channels localized to the intercalated disks. Table 4.2 in Section 4.6presents specific values of the simulation parameters. Conduction speeds computedfrom simulations with half of that spatial resolution and a quarter of that temporalresolution resulted in conduction velocities off by no more than 5% from those inFigure 4.4.
We first remark on conduction speed for small cleft-to-ground resistance, β.Conduction speeds for β = 10−4 (dcleft = 115 nm) range from about 1.5 (86 cm/s)under normal gap junction levels to about 0.1 (5.7 cm/s) under 1% of normalgap junction expression levels. See Appendix 4.6 for the relation between β andthe cleft width dcleft. The conduction speeds are only weakly influenced by Na+
channel localization. This lack of dependence on fNa is reasonable as the β → 0limit corresponds to clefts that stay grounded, hence acting like extracellular space.
4.4.2.1 Little Effect of Cleft-to-Ground Resistance on Conduction Speed
Under Uniform Na+ Channel Density
Figure 4.4c shows that the physiologically plausible cleft-to-ground resistanceshave little effect on conduction speed if Na+ channel density is uniform.
4.4.2.2 Reduced Conduction Speed with Normal Gap Junctional Cou-
pling and Localized Na+ Channels
As shown in the κ = 1 curves in Figure 4.4ab, conduction speed decreasesmonotonically over the range of β surveyed. With 100% Na+ channel localization,the clefts can act to slow down conduction by a third before propagaion fails.With only 50% Na+ channel localization, the conduction speed decreases onlyslightly over the cleft-to-ground resistances investigated. These decreases over βare reasonable as the localization of Na+ channels to the intercalated disks forcescurrent to go through the extra cleft-to-ground resistance. When such resistanceis larger, and when more current is forced through the clefts, conduction shouldbe slower.
57

0 1 2 3 4 5
x 10−3
0
0.5
1
1.5
β (cleft resistance)
fNa
= 1
k = 1 eps = 0.01
k = 0.3 eps = 0.01
k = 0.1 eps = 0.01
k = 0.03 eps = 0.01
k = 0.01 eps = 0.01
0 1 2 3 4 5
x 10−3
0
0.5
1
1.5
β (cleft resistance)
Con
duct
ion
spee
d (n
ondi
men
sion
al)
fNa
= 0.5
0 1 2 3 4 5
x 10−3
0
0.5
1
1.5
β (cleft resistance)
fNa
= Uniform
Figure 4.4: Conduction velocity under the full ephaptic model as a function ofthe nondimensional cleft-to-ground resistance for gap junction expression rangingfrom 1% to 100% of normal. For these simulations, ε = 0.01. Na+ channels aredistributed such that (a) all are located at the intercalated disks, (b) half arelocated at the intercalated disks, or (c) channel density is uniform.
58

4.4.2.3 Enhanced Conduction Speed with Low Gap Junctional Cou-
pling and Localized Na+ Channels
As shown in the κ = 0.01 and κ = 0.03 curves in Figure 4.4a, counduction speedis not necessarily monotonically decreasing in β. For example, the conductionspeed with gap junctional coupling at 1% of normal is about 0.1 (5.7 cm/s) forβ = 10−4 (dcleft = 115 nm), increasing to about 0.4 (22.8 cm/s) for β = 10−3
(dcleft = 11 nm), and then decreases for larger cleft-to-ground resistances. Toemphasize this point, note that in the case of β = 10−3 conduction is faster thanin the analogous non-clefted model.
This effect reproduces the corresponding qualitative result in [33]. In thatpaper, the cleft width for maximal conduction speed under 1% of normal gapjunction expression levels is dcleft ≈ 40 nm. That maximal speed is ≈ 15 cm/s.These values are quantitatively different than the results of our work, presented inFigure 4.4a.
4.4.3 Numerical Results for the Homogenized System
Figure 4.5 shows the conduction speed of the homogenized ephaptic system forselected values of cleft-to-ground resistance, gap junctional coupling strength, anddegree of Na+ channel localization to the intercalated disks. Table 4.2 in Section 4.6presents specific values of the simulation parameters. Conduction speeds computedfrom simulations with half of that spatial resolution and a quarter of that temporalresolution resulted in conduction velocities off by no more than 2% from those inFigure 4.4.
As before, we first remark that conduction speed for small β ranges from about1.5 (86 cm/s) under normal gap junction expression levels to about 0.1 (5.7 cm/s)under 1% of normal gap junction levels. Conduction speed for small β is notstrongly affected by Na+ channel localization.
4.4.3.1 Little Effect of Cleft-to-Ground Resistance on Conduction Speed
Under Uniform Na+ Channel Density
As with the full system, Figure 4.5c shows that the physiologically plausiblecleft-to-ground resistances have little effect on conduction speed when Na+ channeldensity is uniform.
4.4.3.2 Reduced Conduction Speed with Normal Gap Junctional Cou-
pling
As shown in the κ = 1 curves in Figure 4.5ab, conduction speed decreasesmonotonically over the range of β surveyed. Just as with the full system, under
59

0 1 2 3 4 5
x 10−3
0
0.5
1
1.5
β (cleft resistance)
fNa
= 1
k = 1 eps = 0
k = 0.3 eps = 0
k = 0.1 eps = 0
k = 0.03 eps = 0
k = 0.01 eps = 0
0 1 2 3 4 5
x 10−3
0
0.5
1
1.5
β (cleft resistance)
Con
duct
ion
spee
d (n
ondi
men
sion
al)
fNa
= 0.5
0 1 2 3 4 5
x 10−3
0
0.5
1
1.5
β (cleft resistance)
fNa
= Uniform
Figure 4.5: Conduction velocity under the homogenized ephaptic model as a func-tion of the nondimensional cleft-to-ground resistance for gap junction expressionranging from 1% to 100% of normal. Na+ channels are distributed such that (a)all are located at the intercalated disks, (b) half are located at the intercalateddisks, or (c) channel density is uniform.
60

100% Na+ channel localization to the ends of the cells, the clefts can act to slowdown conduction by a third before propagation fails.
4.4.3.3 Reduced conduction speed with low gap junctional coupling
and Na+ channel localization
As shown in all the curves in Figure 4.5ab, conduction speed is apparentlymonotonically decreasing in β for all gap junction expression levels under thehomogenized system. The simulations show no parameter range where more cleftresistance can increase conductionspeed. This behavior is qualitatively differentfrom that of the full system.
4.4.3.4 Comparisons of the Full and Homogenized Systems
Figure 4.6 overlays the conduction speeds of the full and homogenized systemsfor various values of gap junctional coupling, cleft-to-ground resistance, and Na+
channel localization. Figures 4.7 and 4.8 show the intracellular and cleft potentialsof the full and homogenized systems under high and low gap junctional coupling,respectively. We observe a good agreement between the models when κ = 1. Thetwo systems disagree quantitatively for low gap junctional coupling levels underuniform Na+ channel density. Further, the systems diagree qualitatively when gapjunction expression is low and Na+ channels are localized to the intercalated disks.
4.5 Discussion
The intent of this chapter is to provide a homogenized description of the ephap-tic effect in cardiac tissue for the purpose of easing analysis and numerical sim-ulation that delineates when the effect is relevant. To be most useful, such adescription must be more convenient than, yet capture the behavior of the fulldescription. Indeed, the homogenized system (4.14)–(4.15) is nicer than the fullsystem because it simplifies the spatial domain and boundary conditions. It alsoavoids the requirement of resolving individual cells. Our primary question is, then,does the homogenized system adequately capture the dynamics of the full system?We investigate this question empirically in its intended context, instead of throughrigorous convergence results.
Our simulations show that the homogenized system appears to capture thewaveform and wavespeed of the full system when gap junctions are present at theirnormal levels. In this case, if Na+ channels are localized to the intercalated disks,the ephaptic mechanism can slow propagation substantially. The homogenizedmodel provides a convenient description for studying any ephaptic effect in tissuewithout gap junction pathologies.
61

0 1 2 3 4 5
x 10−3
0
0.5
1
1.5
β (cleft resistance)
fNa
= 1
k = 1 eps = 0.01
k = 1 eps = 0
k = 0.1 eps = 0.01
k = 0.1 eps = 0
k = 0.01 eps = 0.01
k = 0.01 eps = 0
0 1 2 3 4 5
x 10−3
0
0.5
1
1.5
β (cleft resistance)
Con
duct
ion
spee
d (n
ondi
men
sion
al)
fNa
= 0.5
0 1 2 3 4 5
x 10−3
0
0.5
1
1.5
β (cleft resistance)
fNa
= Uniform
Figure 4.6: An overlay of the conduction speeds computed under the full andhomogenized systems. At normal gap junction expression levels (κ = 1) the sys-tems agree well, but that agreement disappears as the gap junctional coupling isreduced.
62

0 0.5 1 1.5−100
−80
−60
−40
−20
0
Intracellular Potential at t=0.5
x (nondimensional)
Pot
entia
l (m
V)
Full
Homogenized
0 0.5 1 1.5−100
−80
−60
−40
−20
0
Cleft Potential at t=0.5
x (nondimensional)
Pot
entia
l (m
V)
0 0.5 1 1.5−100
−80
−60
−40
−20
0
Intracellular Potential at t=1
x (nondimensional)P
oten
tial (
mV
)
0 0.5 1 1.5−100
−80
−60
−40
−20
0
Cleft Potential at t=1
x (nondimensional)
Pot
entia
l (m
V)
Figure 4.7: An overlay of the computed solutions to the full and homogenizedsystems under β = 10−3, κ = 1, fNa = 1 at two different times. The top panelsshow intracellular potentials. Note that individual cells can be resolved in theupstroke of the full simulations. The bottom panels show cleft potential. Forclarity, the intracellular potentials of the full simulations are plotted for only 5 ofthe 20 interior nodes.
63

0 0.5 1 1.5−100
−80
−60
−40
−20
0
Intracellular Potential at t=0.5
x (nondimensional)
Pot
entia
l (m
V)
FullHomogenized
0 0.5 1 1.5−100
−80
−60
−40
−20
0
Cleft Potential at t=0.5
x (nondimensional)
Pot
entia
l (m
V)
0 0.5 1 1.5−100
−80
−60
−40
−20
0
Intracellular Potential at t=1
x (nondimensional)P
oten
tial (
mV
)
0 0.5 1 1.5−100
−80
−60
−40
−20
0
Cleft Potential at t=1
x (nondimensional)
Pot
entia
l (m
V)
Figure 4.8: An overlay of the computed solutions to the full and homogenizedsystems under β = 10−3, κ = 0.01, fNa = 1 at two different times. The toppanels show intracellular potentials. Note that individual cells can be resolved inthe upstroke of the full simulations. The bottom panels show cleft potential. Forclarity, the intracellular potentials of the full simulations are plotted for only 5 ofthe 20 interior nodes.
64

Unfortunately, the full and homogenized systems disagree when gap junctionlevels are significantly reduced. Hence, the homogenized equations in their presentform are unlikely to contribute toward understanding cardiac propagation in gap-junction deficient mice.
We now conjecture about why the homogenized and full systems disagree un-der low gap junctional coupling yet agree under high coupling. Most likely, thediscrepancy arises because of a failure of the ansatzes (4.10)–(4.13). The homog-enized equations assumed that gating variables have O(1) variations only on theslow, macroscopic spatial scale. In particular, gating variables on opposing sides ofa cleft are assumed to be equal. Under the view of ephaptic mechanism presentedin Figure 4.1 there should be substantial differences in the gating values across acleft, and it is not surprising that the homogenization fails.
We suggest that the agreement of the full and homogenized systems undernormal gap junction levels arises because the ansatz is met in that case. Highcoupling means potential differences between neighboring cells result in a largecorrecting current. With closer potentials of neighboring cells, the gating valuesshould be similar, affording an accurate approximation by the ansatz.
4.6 Simulation Parameters
We run the simulations of the full and homogenized systems with nondimen-sional parameters corresponding to the dimensional values used in [33]. The cleft-to-ground resistance in the models would be very difficult to measure experimen-tally. Instead, we relate it to the cleft width, as in [33], through Rc = ρmyo
8πw, where
w is the cleft width. Plausible values for w are on the order of 10–100 nm.
4.7 Numerical Schemes
4.7.1 Full System
4.7.1.1 Spatial Discretization
To discretize the spatial derivatives in (4.24)–(4.27) to second order, we intro-duce a cell-centered grid with n interior and 2 ghost points per biological cell ofwidth ε, see Figure 4.9. The values at the ghost potential will be such that theboundary conditions are enforced to second order spatial accuracy.
We let φij denote the j-th node within the i-th cell. Note that j = 0 and
j = n + 1 correspond to the i-th cell’s ghost points. Further, the rightward ghostpoint of one cell is not identical to the leftward ghost point of its neighboring cell.Let wi
j be the gating variables at for the i-th cell, for 0 ≤ j ≤ n+ 1. The variables
65

Measurable ParametersSymbol Parameter Value Referencer radius of cell 11 · 10−4 cm [33]ℓ length of cell 10−2 cm [33]σc cytosolic conductivity 6.67 mΩ−1cm−1 [33]gGJ,end gap junctional conductance
per area6.66 · 102 mΩ−1cm−2 [33]
Rm membrane sheet resistance 20 kΩcm2 [44]Cm membrance capacitance
per area1 µF cm2 [33]
w cleft width 2 nm – 115 nm
Derived ParametersSymbol Parameter Value ExpressionRc cleft-to-ground resistance 2.98 · 104 kΩ
– 5.19 · 102 kΩ
18πσcw
α aspect ratio of cells 0.055 πr2
2πrℓ
ε nondimensional length of cells 0.0087√
ℓRmσc
κ nondimensional gap junctionalconductivity
0.9985 ε2gGJ,endRm
β nondimensional cleft resistance 5.7 · 10−3–10−4 ε2πr2 σc
ℓRc
Table 4.1: The physical and derived parameters that enter our full and homoge-nized models.
Simulation ParametersSymbol Parameter Value (nondim) Value∆t time step 1.25 · 10−5 2.5 · 10−4 ms∆x node spacing 5 · 10−4 5 · 10−4 cmL domain length 1.5 1.5 cmTend time of simulation 1.5 30 mslinit width of initial stimulation 0.125 0.125 cmφinit potential of initial stimulation 0 0 mV
Table 4.2: Computational parameters that enter our numerical simulations of thefull and homogenized models.
66

φi0 φi
1 φin φi
n+1 φi+10 φi+1
1 φi+1n φi+1
n+1
ε = n∆x
Figure 4.9: The spatial discretization of each biological cell in the full model.Each biological cell is broken into n computational nodes and two ghost nodes,represented by hollow dots. Note that there are no computational nodes for theclefts as the cleft potential can be deduced from the potential at the interior andghost nodes. Also note that the rightward ghost node of a cell is distinct from theleftward ghost node of its neighbor. The variable φi
k denotes the potential at thek-th node of the i-th cell, where k = 0 and k = n + 1 correspond to the ghostnodes.
wi0 and wi
n+1 correspond to gating values for the left and right ends of the i-th cell.Equation (4.24) can be discretized immediately
∂φik
∂t= α
(
1
∆x2,− 2
∆x2,
1
∆x
)
·(
φik−1, φ
ik, φ
ik+1
)
− Iion, side(φik, w
iside, k)
for 1 ≤ k ≤ n. Cleft potentials need not be directly computed, as they are givenin terms of intracellular potentials by (4.27). Discretizing, we obtain
φic =
(
β
ε∆x,− β
ε∆x,− β
ε∆x,β
ε∆x
)
·(
φin, φ
in, φ
in+1, φ
i+10 , φi+1
1
)
.
Now, we may discretize (4.25) and (4.26) as
∂
∂t
(
− β
ε∆x,β
ε∆x,1
2+
β
ε∆x,1
2− β
ε∆x
)
·(
φi−1n , φi−1
n+1, φi0, φ
i1
)
=
(
1
ε
κ
2ε,1
ε
κ
2ε,1
ε(− 1
∆x− κ
2ε),
1
ε(
1
∆x− κ
2ε)
)
·(
φi−1n , φi−1
n+1, φi0, phi
i1
)
− Iion, end(φi
0 + φi1
2, wi
left),
67

∂
∂t
(
1
2− β
ε∆x,1
2+
β
ε∆x,β
ε∆x,− β
ε∆x
)
·(
φin, φ
in+1, φ
i+10 , φi+1
1
)
=
(
1
ε(
1
∆x− κ
2ε),
1
ε(− 1
∆x− κ
2ε)
)
,1
ε
κ
2ε,1
ε
κ
2ε·(
φi−1n , φi−1
n+1, φi0, φ
i1
)
− Iion, end(φi
0 + φi1
2, wi
left).
At the left and right boundaries of the strand of cells, the boundary conditions(4.26) or (4.25) refer to a neighboring cell that does not exist. We complete the IVP
by defining φ0n+1
def= φ1
0, φ0n
def= φ1
1, and similarly for the right end of the rightmostcell. This boundary condition is equivalent to a domain of length L coupled to theoriginal domain by an even reflection.
Equations (4.24)–(4.27) can be phrased as a single matrix equation of the form
∂tLtransu = Ldxu− Iion(Ltransu, w), (4.45)
where u is a column vector of the potential at intracellular and ghost nodes; Ltransugives the transmembrane potential at each node; Ldxu is the discretization of thespatial derivatives and differences; and w is a column vector of all gating vari-ables at intracellular and ghost nodes. The transmembrane potential and gatingvariables at the ghost nodes correspond to the cell-to-cleft potential and gatingvalues.
4.7.1.2 Time Discretization
We discretize (4.45) by treating the Ldxu term implicitly via Crank-Nicholsonand the Iion term explicitly through a first order Euler method. Letting um andwm be the vector of potentials and gating variables at the time m∆t, we evolve uby
Ltransum+1 − um
∆t= Ldx
um+1 + um
2− Iion(Ltransu
m, wm),
wm+1 − wm
∆t= LRD(Ltransu
m, wm).
4.7.1.3 Spatial Convergence Study
To verify the second order spatial accuracy we expect from our numericalscheme, we run several simulations for an empirical convergence study.
Let φ∆xfull be the computed solution to the full system with
ε = 0.01,∆t = 5 · 10−5, n = 5 · 2k,∆x =ε
n, L = 0.5,
68

Parameters Lp rspacep [φ; 0.01/5] rspace
p [φ; 0.01/10] rspacep [φ; 0.01/20]
κ = 1 L1 1.95 1.99 2.00β = 10−3 L2 1.91 1.97 2.00fNa = 1 L∞ 1.88 1.96 2.00κ = 0.01 L1 2.00 2.00 2.00β = 10−3 L2 2.00 2.00 2.00fNa = 1 L∞ 1.99 2.00 2.00κ = 0.01 L1 2.00 2.00 1.99β = 10−3 L2 2.02 1.96 2.07fNa = uniform L∞ 2.07 1.84 2.31
Table 4.3: Spatial convergence rates for full system with ε = 0.01 computed atnondimensional time 0.2.
where k = 0, · · · , 4. Note that in increase in k by 1 corresponds to a doubling ofthe spatial resolution.
We define the error between the k-th and the k + 1-st solutions as
espacep [φ; ∆x] = ||φ∆x
full − I∆x/2→∆xφ∆x/2full ||p,
where I∆x/2→∆x is the interpolation operator between the finer to the coarser gridobtained by averaging adjacent interior nodes, and where || · ||p is the discretizationof the p-norm. We also define an empirical spatial convergence rate as
rspacep [φ; ∆x] = log2
espacep [φ; ∆x]
espacep [φ; ∆x/2]
.
Table 4.3 presents the empirical rates of convergence for intracellular potential inthe full system under the L1, L2, and L∞ norms. We observe that the convergenceappears to be second order in space for all three parameter choices.
4.7.1.4 Temporal Convergence Study
To verify the first order temporal accuracy we expect from our numericalscheme, we run several simulations for an empirical convergence study.
Let φ∆tfull be the computed solution to the full system with
ε = 0.01,∆t = 2 · 10−4 · 2−k, n = 20,∆x =ε
n, L = 0.5,
where k = 0, · · · , 4. Note that in increase in k by 1 corresponds to a doubling ofthe spatial resolution.
69

Parameters Lp rtimep [φ; 2 · 10−4] rtime
p [φ; 1 · 10−4] rtimep [φ; 5 · 10−5]
κ = 1 L1 1.03 1.02 1.00β = 10−3 L2 1.06 1.03 1.00fNa = 1 L∞ 1.06 1.02 1.00κ = 0.01 L1 1.02 0.99 1.00β = 10−3 L2 1.02 1.02 1.01fNa = 1 L∞ 1.06 1.03 1.02κ = 0.01 L1 1.06 1.03 1.01β = 10−3 L2 1.05 1.02 1.01fNa = uniform L∞ 0.98 0.99 1.00
Table 4.4: Temporal convergence rates for full system computed at nondimensionaltime 0.2.
We define the error between the k-th and the k + 1-st solutions as
etimep [φ; ∆t] = ||φ∆t
full − φ∆t/2full ||p,
where ||·||p is the discretization of the p-norm. We also define an empirical temporalconvergence rate as
rtimep [φ; ∆t] = log2
etimep [φ; ∆t]
etimep [φ; ∆t/2]
.
Table 4.4 presents the empirical rates of convergence for intracellular potential inthe full system under the L1, L2, and L∞ norms. We observe that the convergenceappears to be first order in time for all three parameter choices.
4.7.2 Homogenized System
4.7.2.1 Discretization in Space
The only spatial derivative in the homogenized system (4.35)-(4.43) is ∂2
∂x2 ,which we discretize according to the standard 3-point stencil.Unlike the full system,the cell-to-cleft transmembrane potentials must be computed directly. Equations(4.35)-(4.36) can be written in the form
∂tu = Ldxu− Iion(u, w),
where u is a column vector of the cell-to-ground and cell-to-cleft transmembranepotentials at all nodes.
70

4.7.2.2 Discretization in Time
The temporal discretization is the same as with the full system.
4.7.2.3 Spatial Convergence Study
To verify the second order spatial accuracy we expect from our numericalscheme, we run several simulations for an empirical convergence study.
Let φ∆xhomog be the computed solution to the homogenized system with
∆t = 5 · 10−5, n = 5 · 2k,∆x =ε
n, L = 0.5,
where k = 0, · · · , 4. Note that in increase in k by 1 corresponds to a doubling ofthe spatial resolution.
We define the error between the k-th and the k + 1-st solutions as
espacep [φ; ∆x] = ||φ∆x
homog − I∆x/2→∆xφ∆x/2homog||p,
where I∆x/2→∆x is the interpolation operator between the finer to the coarser gridobtained by averaging adjacent interior nodes, and where || · ||p is the discretizationof the p-norm. We also define an empirical spatial convergence rate as
rspacep [φ; ∆x] = log2
espacep [φ; ∆x]
espacep [φ; ∆x/2]
.
Table 4.5 presents the empirical rates of convergence for intracellular potential inthe full system under the L1, L2, and L∞ norms. We observe that the convergenceappears to be second order in space for all three parameter choices.
4.7.2.4 Temporal Convergence Study
To verify the second order temporal accuracy we expect from our numericalscheme, we run several simulations for an empirical convergence study.
Let φ∆thomog be the computed solution to the homogenized system with
∆t = 2 · 10−4 · 2−k, n = 20,∆x =ε
n, L = 0.5,
where k = 0, · · · , 4. Note that in increase in k by 1 corresponds to a doubling ofthe spatial resolution.
We define the error between the k-th and the k + 1-st solutions as
etimep [φ; ∆t] = ||φ∆t
homog − φ∆t/2homog||p,
71

Parameters Lp rspacep [φ; 0.01/5] rspace
p [φ; 0.01/10] rspacep [φ; 0.01/20]
κ = 1 L1 5.65 1.98 2.01β = 10−3 L2 5.34 1.99 2.02fNa = 1 L∞ 5.15 1.99 2.01κ = 0.01 L1 1.99 2.05 2.00β = 10−3 L2 1.27 2.09 2.01fNa = 1 L∞ 0.66 2.04 1.99κ = 0.01 L1 0.70 2.01 2.02β = 10−3 L2 0.29 2.02 2.02fNa = uniform L∞ 0.19 2.03 2.02
Table 4.5: Spatial convergence rates for homogenized system computed at nondi-mensional time 0.2.
Parameters Lp rtimep [φ; 2 · 10−4] rtime
p [φ; 1 · 10−4] rtimep [φ; 5 · 10−5]
κ = 1 L1 1.03 1.02 1.00β = 10−3 L2 1.06 1.03 1.00fNa = 1 L∞ 1.06 1.02 1.00κ = 0.01 L1 1.02 0.99 1.00β = 10−3 L2 1.02 1.02 1.01fNa = 1 L∞ 1.07 1.03 1.02κ = 0.01 L1 1.06 1.03 1.01β = 10−3 L2 1.05 1.02 1.01fNa = uniform L∞ 0.98 0.99 1.00
Table 4.6: Temporal convergence rates for homogenized system computed at nondi-mensional time 0.2.
where ||·||p is the discretization of the p-norm. We also define an empirical temporalconvergence rate as
rtimep [φ; ∆t] = log2
etimep [φ; ∆t]
etimep [φ; ∆t/2]
.
Table 4.6 presents the empirical rates of convergence for intracellular potential inthe homogenized system under the L1, L2, and L∞ norms. We observe that theconvergence appears to be first order in time for all three parameter choices.
72

Part II
Blow-Up in Bacterial Chemotaxis
73

Chapter 5
Introduction
Many bacteria can spontaneously organize themselves into patterns of remark-able regularity, complexity, and beauty. For example, the bacteria Escherichiacoli and Salmonella typhimurium can form patterns like concentric rings, spirals,and radial stripes and spots when in the presence of chemicals such as succinate[39, 40]. The bacteria achieve this cell-cell communication through the productionand release of a chemo-attractant, which is a chemical that they preferentiallymove toward. Such chemically driven motion of cells is known as chemotaxis andis an important mechanism in biology, for example in the body’s response to abacterial infection [40].
A well studied model of chemotaxis is that for Dictyostelium discoideum, whosechemo-attractant is cyclic-AMP. See [52] for a collection of many mathematical re-sults. With these bacteria, waves and pattern formation are observed experimen-tally. The model of Keller and Segel [28] describes the bacteria, chemo-attractantsystem as a continuum through coupled reaction diffusion equations, and it is thesubject of our analysis.
5.1 Derivation of Keller-Segel Equations
The Keller-Segel equations describe the dynamics of a population of bacteriaunder the following modeling assumptions:
1. The bacteria undergo Fickian diffusion.
2. The bacteria are attracted up the concentration of a chemo-attractant.
3. The bacteria produce this chemo-attractant at a steady rate.
4. The chemo-attractant undergoes Fickian diffusion and decays.
74

Let u(t, x) and c(t, x) be the density of bacteria and chemo-attractant, respec-tively, at time t and position x ∈ R
d. Assumptions 1 and 2 provide
j = −D∇u+ χu∇c, (5.1)
where j is the flux density of bacteria, D is the diffusion coefficient of the bacteria,and χ is a scalar known as the sensitivity. Note that the flux of bacteria up theconcentration gradient of c is proportional to the local density of bacteria.
The evolution equation corresponding to the flux (5.1) coupled with no-fluxboundary conditions over a bounded domain Ω is
∂tu+ ∇ · (−D∇u+ χu∇c) = 0, (5.2)
−D∂νu+ χu∂νc = 0 on ∂Ω. (5.3)
The behavior of c can be modeled through the reaction-diffusion equation
∂tc− γ∆c + µc = βu, (5.4)
with suitable boundary conditions, where γ, µ, and β are constants. Often (5.4) issimplified by assuming the diffusion of c is fast. Then, the evolution PDE becomeselliptic. Three popular simplifications are the Dirichlet field, the Neumann field,and the Jager-Luckhaus field, which we subsequently refer to as (D), (N), and (JL)boundary conditions to the Keller-Segel system. The Dirichlet field is given by
−∆c = αu in Ω,
c = 0 on ∂Ω,(D)
and is derived by assuming γ is large and µ is small. The Neumann field is givenby
−∆c + ac = αu in Ω,
∂νc = 0 on ∂Ω,(N)
and is derived by assuming γ, µ, and β are large and of the same order. The JLfield is given by
−∆c = α(u− u) in Ω,
∂νc = 0 on ∂Ω,(JL)
and is derived by assuming γ and β are large and of the same order [26]. Here,u represents the avarage of u and must be subtracted in order for (JL) to admitsolutions. The Keller-Segel system is (5.2)– (5.3) coupled with either (D), (N), or(JL).
By an appropriate rescaling, we let D = 1. We also let the other parameters αand χ be unity. Their value does not substantively effect the arguments we present.Sometimes, we will remark on the specific effect of other values of χ. Except forliterature reviews, we do not remark on (N).
75

5.2 Finite-Time Blow-Up in the Whole Plane
The Keller-Segel equations in two dimensions exhibit a critical mass effect. Forthe equations posed over the whole plane,
∂tu+ ∇ · (−∇u+ u∇c) = 0 in R2, (5.5)
−∆c = u in R2, (5.6)
solutions with initial mass M =∫
R2 u0(x)dx > 8π blow up in finite time by con-centrating into a Dirac mass. A common tool for investigating this blow-up ofclassical solutions is the second moment of the bacteria density [12]. Equations(5.5)-(5.6) provide simple dynamics for the second moment of u, as detailed in thefollowing lemma.
Lemma 5.1. Let u(t, x) be a classical solution to (5.5)-(5.6) which decays suffi-ciently fast as |x| → ∞. Then
∂t
∫
R2
|x|2u(t, x)dx = 4M
(
1 − M
8π
)
. (5.7)
Proof of Lemma 5.1. Multiplying (5.5) by |x|2 and integrating by parts, we get
∂t
∫
R2
|x|2u(t, x)dx = −∫
R
|x|2∇ · (−∇u+ u∇c)
=
∫
R2
∇|x|2 · (−∇u+ u∇c)dx (5.8)
=
∫
R2
∆|x|2 u+
∫
R2
2x · u∇c dx (5.9)
= 4M + 2
∫
R2
x · u∇c dx. (5.10)
From (5.6), we can write c explicitly in terms of u:
c(t, x) = − 1
2π
∫
R2
log |x− y|u(t, y)dy, (5.11)
∇c(t, x) = − 1
2π
∫
R2
x− y
|x− y|2u(t, y)dy (5.12)
76

Now, plugging (5.12) into (5.10), we get
∂t
∫
R2
|x|2u(t, x)dx = 4M − 1
π
∫∫
R2×R2
x · x− y
|x− y|2u(t, x)u(t, y)dxdy (5.13)
= 4M − 1
2π
∫∫
R2×R2
(x− y) · x− y
|x− y|2u(t, x)u(t, y)dxdy (5.14)
= 4M − 1
2π
∫∫
R2×R2
u(t, x)u(t, y)dxdy (5.15)
= 4M
(
1 − 1
8πM
)
. (5.16)
The second equality comes from a symmetry of x and y in the integrand from(5.13).
This lemma allows us to conclude finite time blow-up of classical solutions withmass larger than 8π, as the second moment must remain positive for as long asthe solution exists. If χ 6= 1, the critical mass for blow-up would be 8π/χ.
5.3 Free Energy for Keller-Segel Systems
One nice property of Keller-Segel equations is that solutions have a decreasingfree energy, which we demonstrate in a bounded domain Ω with smooth boundaryand (JL) boundary conditions.
Lemma 5.2. Let u(t, x), c(t, x) be a classical solution to
∂tu+ ∇ · (−∇u+ u∇c) = 0 in Ω, (5.17)
−∆c = u− u in Ω, (5.18)
∂νc = ∂νu = 0 on ∂Ω. (5.19)
Then
∂t
∫
Ω
(
u log u− 1
2|∇c|2
)
dx = −∫
Ω
u|∇(log u− c)|2dx. (5.20)
We denote the free energy of a solution by
F [u] =
∫
Ω
(
u log u− 1
2|∇c|2
)
dx (5.21)
Proof of Lemma 5.2. We rewrite (5.17) as
∂tu = ∇ ·(
u∇(log u− c))
. (5.22)
77

Multiplying by log u− c and integrating by parts, we get
∫
Ω
∂tu(
log u− c)
= −∫
Ω
u|∇(log u− c)|2dx. (5.23)
At the expense of some extra terms, we can pull the time derivative outside theintegral of the left hand side:
∂t
∫
Ω
(
u log u− u− (u− u)c)
dx+
∫
Ω
(u− u)∂tcdx = −∫
Ω
u|∇(log u− c)|2dx.(5.24)
Noting that
∫
Ω
(u− u)∂tc dx =
∫
Ω
−∆c ∂tc dx =1
2∂t
∫
Ω
|∇c|2 dx, (5.25)
we conclude
∂t
∫
Ω
(
u logu− u− (u− u)c+1
2|∇c|2
)
dx = −∫
Ω
u|∇(log u− c)|2dx, (5.26)
which can be simplified to (5.20).
5.4 Outline of Part II
In Chapter 6 we establish finite-time blow-up for the Keller-Segel equations withDirichlet boundary conditions over a disk when the mass is above the thresholdvalue. We do not assume radial symmetry. We determine the evolution of thesecond moment of the bacterial density by using an explicit representation of theGreen’s Function for Laplace’s equation over the disk.
In Chapter 7 we present a conjecture about how generally chemotactic collapseoccurs on the domain boundary under (JL) boundary conditions. We presentnumerical evidence based on simulations of the evolution equations and on a nu-merical linear stability analysis of the steady states. We consider only the domainT × [0, 1].
In Chapter 8, we propose a tool for assessing stability of steady states to Keller-Segel equations. We derive a functional inequality that determines whether thereare nearby functions with lower free energy than the steady state. We demonstratean application of the tool by using it to determine at what mass the uniform profilechanges stability.
78

Chapter 6
Finite-Time Blow-Up of
Keller-Segel on a Disk with
Dirichlet Boundary Conditions
6.1 Introduction
The Keller-Segel equations are a simple model of bacterial chemotaxis. Theydescribe a population of bacteria undergoing Fickian diffusion yet moving up theconcentration gradient of a chemo-attractant that they produce. They are mathe-matically interesting because of the tension between the spreading due to diffusionand the coalescence due to the chemo-attraction. Under some conditions, solutionssettle into a steady state. Under others, solutions can develop Dirac-delta singu-larities in finite or infinite time, a phenomenon known as chemotactic collapse.Whether the diffusion or the chemoattraction ‘wins’ can depend on the number ofspatial dimensions, specifics of the domain shape and boundary conditions, andon the total amount of bacteria present.
In an early mathematical analysis of the Keller-Segel equations, [9] and [10],Childress and Percus conjectured that chemotactic collapse can not occur in onespatial dimension, but can in two dimensions if there is more than a certain thresh-old of bacteria present. Their primary tools were differential inequalities and dom-inant balance asymptotics.
About a decade later, Jager and Luckhaus [26] provided a proof for existenceof a critical mass effect in two dimensions. They proved that below some value ofmass, solutions exist globally in time. They also showed that solutions with massabove some value can exhibit chemotactic collapse. Their results were posed overbounded domains and under the now-called JL boundary conditions.
Subsequent work by Nagai [41] showed that in bounded domains with JLand other boundary conditions, there can be finite-time blow-up. Specifically,
79

he showed that if the mass M > 8π and if ∃q ∈ Ω such that∫
Ωu0(x)|x − q|dx is
sufficiently small, then finite-time blow-up occurs. Here α and χ from (5.2)–(5.3)and (JL) in Section 5.1 are assumed to be 1. Under JL boundary conditions, solu-tions can blow up on the boundary, and they do so if M > 4π and ∃q ∈ ∂Ω suchthat
∫
Ωu0(x)|x − q|2dx is sufficiently small. Nagai [42] also showed that radial
solutions with subthreshold mass exist globally in time.Analogous work by [12] for the Keller-Segel eqautions in the whole plane, (5.5)–
(5.6), revealed a similar story. They showed existence for some solutions withinitial mass less than 8π. They also showed that classical solutions blow up infinite time if M > 8π. Their primary tool for establishing finite-time blow-up wasthe evolution of the second moment.
In this chapter, we provide an analogous argument to [12] in the case of theKeller-Segel equations posed over the disk with Dirichlet boundary conditions. Weshow the finite-time blow-up for supercritical mass.
6.2 Finite-Time Blow-Up under Dirichlet Bound-
ary Conditions for Supercritical Mass
The Keller-Segel problem on a disk with Dirichlet boundary conditions is
∂tu+ ∇ · (−∇u+ u∇c) = 0 in Ω = B1(0), (6.1)
−∆c = u− u in Ω, (6.2)
−∂νu+ u∂νc = 0 on ∂Ω, (6.3)
c = 0 on ∂Ω. (6.4)
In this chapter, we establish the following theorem.
Theorem 6.1. Let (u(t, x), c(t, x)) be a non-negative, classical solution to (6.1)–(6.4). Let u0(x) = u(0, x) and T
maxbe the maximal time of existence of the solution
(u, c). If∫
Ωu0(x)dx > 8π, then T
max<∞.
That is to say, any classical solution with a supercritical mass will blow up infinite time. Were the equations to involve the sensitivity χ, the result would holdfor initial mass M > 8π/χ.
We now provide some qualitative insight into why the theorem is true. Asshown in Section 5.2 and [12], finite-time blow-up occurs when M > 8π for theKeller-Segel system posed over the whole plane. In a sense, c is clamped to zeroat infinity for such a system. In the case of Dirichlet boundary conditions on adisk, c is clamped to zero on |x| = 1. Roughly speaking, the gradient of c should
80

be steeper as a result, and the chemotactic effect should be stronger than in thewhole place. Hence we expect blow-up for M > 8π.
To establish this blow-up theorem, we use the explicit form of the Green’sfunction to Laplace’s equation on the disk with Dirichlet boundary conditions inorder to derive a differential inequality for the second moment of u. Lemma 6.2establishes the evolution of the second moment of u using the explicit form of theGreen’s function. Lemmas 6.3 and 6.4 establish the negativity of the last term inthe second moment evolution equation. Finally, Lemma 6.5 states the differentialinequality for
∫
Ω|x|2udx.
Lemma 6.2. Let u(t, x) be a classical solution to (6.1)–(6.4). Define the quantityM =
∫
Ωu(t = 0, x)dx. Then
∂t
∫
Ω
|x|2u(t, x)dx = 4M
(
1 − M
8π
)
− 2
∫
∂Ω
udσ
+1
π
∫∫
Ω×Ω
|x|2|y|2 − x · y|x|2|y|2 − 2x · y + 1
u(x)u(y)dxdy. (6.5)
Proof of Lemma 6.2. We compute
∂t
∫
Ω
|x|2u(t, x) dx = −∫
Ω
|x|2∇ · (∇u− u∇c)dx (6.6)
= 4M − 2
∫
∂Ω
u dσ + 2
∫
Ω
x · u∇c dx. (6.7)
We now write c explicitly in terms of u by the Green’s function for −∆ on Ω withDirichlet boundary conditions. That is,
c(x) =
∫
Ω
G(x; y)u(y)dy,
where
G(x; y) = − 1
4πlog |x− y|2 +
1
4πlog |x− y|2 +
1
4πlog |y|2,
and y = y/|y|2 is the reflection of y through the unit circle. We can now compute
u(x) · ∇xc(x) = − 1
2π
∫
Ω
x− y
|x− y|2u(x)u(y)dy +1
2π
∫
Ω
x− y
|x− y|2u(x)u(y)dy. (6.8)
Combining (6.7) and (6.8) and simplifying, we obtain (6.5).
Lemma 6.3. For any u ∈ L2(Ω)
∫∫
Ω×Ω
|x|2|y|2 − x · y|x|2|y|2 − 2x · y + 1
u(x)u(y)dxdy ≤ 0
81

Proof of Lemma 6.3.Let Pr(θ) = 1−r2
r2−2r cos θ+1be the Poisson kernel, and let
Kr(θ) = −1
2Pr(θ) +
1
2=
r2 − r cos θ
r2 − 2r cos θ + 1.
Expressing x and y in polar coordinates as (r1, θ1) and (r2, θ2), we can now compute
∫∫
Ω×Ω
|x|2|y|2 − x · y|x|2|y|2 − 2x · y + 1
u(x)u(y)dxdy
=
∫ 1
0
∫ 1
0
∫ 2π
0
∫ 2π
0
Kr1r2(θ2 − θ1)u(r1, θ1)u(r2, θ2)r1r2dθ2dθ1dr2dr1
=
∫ 1
0
∫ 1
0
−2π2∑
n 6=0
u(r1, n)u(r2, n)(r1r2)|n|+1dr1dr2
= −2π2∑
n 6=0
∫ 1
r1=0
u(r1, n)r|n|+11 dr1
∫ 1
r2=0
u(r2, n)r|n|+12 dr2
≤ 0,
where u(r, n) = 12π
∫ 2π
0u(r, θ)e−inθdθ and the second equality follows from Lemma 6.4.
Lemma 6.4. Let Kr(θ) = r2−r cos θr2−2r cos θ+1
, and v1, v2 ∈ L2([0, 2π]). Let us define
v(n) = 12π
∫ 2π
0v(θ)e−inθdθ. Then
∫ 2π
θ1=0
∫ 2π
θ2=0
Kr(θ2 − θ1)v1(θ1)v2(θ2)dθ1dθ2 = −2π2∑
n 6=0
r|n|v1(n)v2(n).
Proof of Lemma 6.4.Recall that Pr(θ) =
∑∞n=−∞ r|n|einθ and Kr(θ) = −1
2Pr(θ) + 1
2. So,
Kr(θ) = −1
2
∑
n 6=0
r|n|einθ
The θ1 integral becomes
∫ 2π
θ1=0
Kr(θ2 − θ1)v1(θ1)dθ1 = −1
2
∫ 2π
0
∑
n 6=0
r|n|einθ2v1(θ1)e−inθ1dθ1
= −π∑
n 6=0
r|n|einθ2 v1(n).
82

Thus, the double integral becomes
∫ 2π
θ1=0
∫ 2π
θ2=0
Kr(θ2 − θ1)v1(θ1)v2(θ2)dθ1dθ2 = −π∑
n 6=0
r|n|v1(n)
∫ 2π
0
v2(θ2)einθ2dθ2
= −2π2∑
n 6=0
r|n|v1(n)v2(n).
Lemma 6.5. Let u be a classical solution to (6.1)–(6.4). Then
∂t
∫
Ω
|x|2u(t, x)dx ≤ 4M
(
1 − M
8π
)
.
Proof of Lemma 6.5. Combine Lemmas 6.2 and 6.3.
We may now prove the theorem.
Proof of Theorem 6.1. Assume Tmax = ∞. If M > 8π, Lemma 6.5 implies that∃T0 ∈ (0,∞) such that
∫
Ω|x|2u(t, x)dx→ 0 as t → T0. Hence, a classical solution
eventually ceases to exist and Tmax < T0 <∞.
83

Chapter 7
Numerically Motivated
Conjecture on Boundary Blow-Up
with Jager-Luckhaus Boundary
Conditions
7.1 Introduction and Blow-Up Conjecture
As mentioned before, the Keller-Segel equations are mathematically interestingbecause of the tension between diffusion and chemo-attraction. A particularly re-markable property of Keller-Segel is that the specific domain, boundary conditions,initial bacterial distribution, and dimensionality of the problem strongly effect thequalitative behavior of the system. Typically, the baceria either concentrate intoa Dirac mass or settle into a smooth steady state. In two-dimensions, which ofthese can occur is governed by a critical mass effect. That is, for large enoughmass, solutions con exhibit blow-up, while for small mass, solutions exist globallyin time.
Effect of Domain Boundary and Symmetry on Critical Mass and Blow-Up
We begin by illustrating the dependence of the blow-up behavior of two- di-mensional Keller-Segel on the domain boundary. As we noted before, when posedover the whole plane, solutions with initial mass M > 8π concentrate to a Diracdelta in finite time [12]. When initial mass M < 8π, solutions exist for all time[12]. When M = 8π, solutions concentrate to Dirac delta in infinite time [4, 3, 5].
In a bounded domain with smooth boundary and (N) or (JL) boundary con-ditions, the critical mass is, instead, 4π. Global existence for mass less than thisvalue is proven in [52] and uses a version of the Trudinger-Moser inequality [7].
84

This threshold value is sharp, as per the following theorem [54, 52] guaranteeingblow-up when mass is sufficiently localized about a boundary point.
Theorem 7.1 ([54, 52]). If Ω ⊂ R2 is bounded with smooth boundary, the initial
data satisfies∫
Ω∩B(x0,R)u0(x)dx > 4π for some x0 ∈ ∂Ω, R > 0, and
1
R2
∫
Ω∩B(x0,4R)
|x− x0|2u0(x)dx
is sufficiently small, then the solution to the Keller-Segel equations with (N) or(JL) boundary conditions blows up in finite time.
See [41] for a similar theorem.In bounded domains with non-smooth boundary, the critical mass can decrease
futher. For example, in a square, the critical mass becomes M = 2π, and concen-tration can occur at a corner [52, 7].
Symmetry and dimensionality also effect the blow-up behavior of Keller-Segelsystems. For example, one-dimensional domain do not exhibit blow-up [9]. Hence,neither will x-invariant solutions to two-dimensional Keller-Segel on the domainT × [0, 1], which we consider in this chapter.
Additionally, radial symmetry has a big effect on the behavior of Keller-Segelover a disk. Without symmetry, solutions with mass 4π < M < 8π initiallyconcentrated about a boundary point result in blow up, as mentioned above. Withradial symmetry and (N) or (JL) boundary conditions, M < 8π results in globalexistence. Blow up can still occur, if, for example M > 8π and the second momentis small enough [42].
Conjecture on Boundary Blow-Up
The intent of this chapter is to put more context around the boundary blow-uptheorem for localized initial data. For M > 4π and with (JL) boundary conditions,we would like to understand whether boundary blow-up occurs, even when initialmass is not localized about a boundary point. We do this in two ways. First, wesimulate the evolution equations over the domain Ω = T× [0, 1] with noisy uniforminitial data. This initial condition should illuminate the opposite extreme from theassumptions of the localization theorem. From the simulations, we observe a phaseof the evolution where the solution is very close to a one-dimensional steady state,which may or may not give rise to an instability that results in boundary blow-up.This observation motivates the study of stability of these 1d steady states, whichis our second approach for numerical insight. Based on our results, we pose thefollowing conjecture.
85

Conjecture 7.2. Let u, c solve the Keller-Segel system on Ω = T× [0, 1] with (JL)boundary conditions:
∂tu+ ∇ · (−∇u+ u∇c) = 0 in Ω = T × [0, 1], (7.1)
−∆c = u− u in Ω, (7.2)
∂νu = ∂νc = 0 on ∂Ω, (7.3)
u|t=0 = u0(x, y). (7.4)
Then there exists an M∗ ∈ (4π, 8π) such that
• if 4π < M < M∗ and the initial data is not symmetric, then boundary blow-up can fail to occur,
• if M∗ < M < 8π and the initial data is not symmetric, then boundary blow-up will occur.
To emphasize, the second part of the conjecture says that blow-up will neces-sarily occur for any initial data, as long as there is no symmetry in the initial data.We leave the relevant symmetries vague, but they include mirror symmetry, andcontinuous and discrete translational symmetries.
Given our simulations, we estimate M∗ ≈ 4.57π in the case of Ω = T × [0, 1].Presumably, the claim should hold for other domains Ω with smooth boundary,
but we present it only over T× [0, 1] because our numerical investigation is limitedto this case.
Other Related Work
We now remark on some related work in [53]. In it, Senba and Suzuki studystability of steady states under (N) boundary conditions over bounded domainswith smooth boundary. Under the assumtion that a, from equation (N) is small,they show ∃δ such that for M ∈ (4π, 4π+δ), steady solutions are linearly unstable.Note the contrast with our conjecture for (JL) boundary conditions.
As we perform numerical simulations in this chapter, we note that there havebeen several numerical studies of Keller-Segel. For example, Marrocco [37] uses amixed finite element method and demonstrates numerical solutions with supercrit-ical mass that concentrate toward multiple Dirac masses. In [17], Filbet analyzeda finite volume scheme for Keller-Segel and proved convergence of the numeri-cal solutions to the actual solution when initial mass is small enough. He alsodemonstrates boundary blow-up when mass is between 4π and 8π.
7.1.1 Outline of Chapter
This chapter is organized as follows. In Section 7.2, we simulate the evolu-tion equations. We present the numerical scheme and results for simulations with
86

masses just above and below the conjectured threshold value M∗. In Section 7.3,we discretize the linearization about one-dimensional steady states in T× [0, 1] anddetermine the stability of such states as a function of mass. In Section 7.4, we usethese results to motivate the conjecture. In Section 7.5 we present empirical spa-tial and temporal convergence tables for the numerical simulation of the evolutionequations.
7.2 Numerical Simulation of Keller-Segel Evolu-
tion
7.2.1 Initial Value Problem
We consider the Keller-Segel system with JL boundary conditions on either asingly periodic square with noisy uniform initial data. The corresponding initialvalue problem is
∂tu+ ∇ · (−∇u+ u∇c) = 0 in Ω = T × [0, 1], (7.5)
−∆c = u− u in Ω, (7.6)
∂νu = ∂νc = 0 on ∂Ω, (7.7)
u|t=0 = M + noise. (7.8)
In a discretization of this system, the noise in the initial data comes from aUnif[0, 1] distribution sampled at each grid point, but with the average subtractedoff in order to ensure the solution has mass M .
We run simulations for M = 4.5π and M = 4.6π. The motivation for thesespecific cases will be clear from Section 7.3 and the discussion in Section 7.4.
7.2.2 Numerical Scheme
7.2.2.1 Spatial Discretization
We now present a finite volume approach to discretizing (7.5)–(7.8). We dis-cretize the domain into square cells of width and height ∆x = ∆y. Let ui,j and ci,jdenote the bacterial and chemo-attractant density, respectively, at the center of the(i, j)-th cell. We remark that a spatially second order discretization of the flux intothe (i, j)-th cell from the (i + 1, j)-th cell is
(ui+1,j−ui,j
∆x− ui+1,j+ui,j
2
ci+1,j−ci,j
∆x
)
∆x,and similarly for the flux in from other neighboring cells.
The semi-discretization of (7.5)–(7.8) is thus
87

∂tui,j =ui+1,j − ui,j
∆x2− ui+1,j + ui,j
2
ci+1,j − ci,j∆x2
+
ui−1,j − ui,j
∆x2− ui−1,j + ui,j
2
ci−1,j − ci,j∆x2
+
ui,j+1 − ui,j
∆x2− ui,j+1 + ui,j
2
ci,j+1 − ci,j∆x2
+
ui,j−1 − ui,j
∆x2− ui,j−1 + ui,j
2
ci,j−1 − ci,j∆x2
. (7.9)
At cells on the top and bottom boundary, the third or fourth row in (7.9) isomitted, as per the no-flux boundary conditions on u and c. Due to the horizontalperiodicity, each cell has a leftward and rightward neighbor, and the first two linesin (7.9) are understood accordingly.
Letting u and c be column vectors corresponding to the values of u and cat each of the cells in the domain discretization, we can rewrite (7.9) and thecorresponding discretization of (7.6) more simply as
∂tu = Llapu + Ltax(c)u, (7.10)
−Llapc = u − u, (7.11)
where Llap is the constant coefficient matrix due to the Laplacian terms in (7.9),Ltax is the c-dependent coefficient matrix due to chemotaxis terms, and u is theaverage of the elements of u.
7.2.2.2 Temporal Discretization
We discretize (7.10) in time by a second order Runge-Kutta method, treatingLtax(c) explicitly and u implicitly through Crank-Nicholson. Specifically, lettingun and cn be the simulated solution at time n∆t, we evolve u according to
u(n+1/2) − un
∆t/2= (Llap + Ltax(c
n))u(n+1/2) + un
2, (7.12)
−Llapc(n+1/2) = u(n+1/2) − u(n+1/2), (7.13)
un+1 − un
∆t=
(
Llap + Ltax(c(n+1/2))
) un+1 + un
2, (7.14)
−Llapcn+1 = un+1 − un+1, (7.15)
See Section 7.5 for a spatial and temporal convergence study.
88

7.2.3 Results
Figure 7.1 shows a sequence of profiles revealing the simulated evolution ofu for the system (7.5)–(7.8) over the singly periodic square T × [0, 1] with mass4.5π. We observe that the initial jaggedness gets smoothed out, becomes a cosine-shaped disturbance from uniform, which starts to grow exponentially. Eventually,the solution settles to an x-invariant solution which corresponds to a steady stateof the 1d problem over [0, 1].
Figure 7.2 shows the sequence of profiles for mass 4.6π. The evolution beginsjust like in the prior case, but eventually the non-uniform 1d profile gains anexponentially increasing disturbance of sinusoidal shape in x. The solution showsnumerical blow-up by concentrating on just a few boundary nodes. By finite time,the computed u ceases to be positive, which we interpret as blow-up.
7.3 Numerical Stability of Keller-Segel Steady
States
The simulations in Section 7.2 suggest that the generality of boundary blow-upin T × [0, 1] may hinge on the stability of the 1d steady states. Thus, we nowconsider the linearized stability of
∂tu+ ∇ · (−∇u+ u∇c) = 0 in Ω = T × [0, 1], (7.16)
−∆c = u− u in Ω, (7.17)
∂νu = ∂νc = 0 on ∂Ω, (7.18)
around an x-independent steady state u0(y), c0(y). Consider the disturbance fromthe steady state
u(t, x, y) = u0(y) + εu1(t, x, y), (7.19)
c(t, x, y) = c0(y) + εc1(t, x, y), (7.20)
where u1 and c1 have zero average over space for all t. Linearizing (7.16)–(7.18)about u0, c0 yields
∂tu1 − ∆u1 + ∇ · (u1∇c0 + u0∇c1) = 0 in Ω = T × [0, 1], (7.21)
−∆c1 = u1 in Ω, (7.22)
∂νu1 = ∂νc1 = 0 on ∂Ω. (7.23)
Using separation of variables, this system’s stability depends on the eigenvalues to
−∆u1 + ∇ · (u1∇c0 + u0∇c1) = λu1 in Ω, (7.24)
−∆c1 = u1 in Ω, (7.25)
∂νu1 = ∂νc1 = 0 on ∂Ω. (7.26)
89

00.5
1
00.5
114
14.1
14.2
x
t = 0
y 00.5
1
00.5
114.135
14.14
14.145
x
t = 0.25
y
00.5
1
00.5
114.136
14.138
14.14
x
t = 0.5
y 00.5
1
00.5
114.13
14.135
14.14
x
t = 0.75
y
00.5
1
00.5
110
15
20
x
t = 2.5
y 00.5
1
00.5
10
20
40
x
t = 2.75
y
00.5
1
00.5
10
50
100
x
t = 5
y 00.5
1
00.5
10
50
100
x
t = 300
y
Figure 7.1: Evolution of the Keller-Segel system with (JL) boundary conditionsover the singly periodic square, T× [0, 1] with mass M = 4.5π initially distributedin a noisy uniform shape. Note that between times t = 0.5 and t = 2.5, the solutionhas the form of a growing cosinusoidal disturbance from uniform. After that, thesolution approaches the steady state of the 1d simulation with the same mass. Inthis simulation ∆t = 0.005,∆x = ∆y = 1/40. For clarity, the plot only shows a20 × 20 sampling of the 40 × 40 simulated grid points.
90

00.5
10
0.51
14.4
14.5
14.6
x
t = 0
y 00.5
10
0.51
14.445
14.45
14.455
x
t = 0.25
y
00.5
10
0.51
10
15
20
x
t = 2.25
y 00.5
10
0.510
50
100
x
t = 4.5
y
00.5
10
0.510
50
100
x
t = 45
y 00.5
10
0.510
100
200
x
t = 50
y
00.5
10
0.510
200
400
x
t = 50.725
y 00.5
10
0.510
1000
2000
x
t = 50.9
y
Figure 7.2: Evolution of the Keller-Segel system with (JL) boundary conditionsover the singly periodic squre, T×[0, 1] with mass M = 4.6π initially distributed ina noisy uniform shape. The solution follows the same description as the simulationshown in Figure 7.1, but it stays near the 1d steady state only until t ≈ 50. Atthis time, a disturbance at the boundary grows until the mass concentrates thereby finite time. In this simulation ∆t = 0.005,∆x = ∆y = 1/40. For clarity, theplot only shows a 20 × 20 sampling of the 40 × 40 simulated grid points.
91

Precisely, if there is a negative eigenvalue λ, then the system (7.16)–(7.18) is lin-early unstable. If all eigenvalues are positive, then the system is linearly stable. As(7.24) has non-constant coefficients that depend only on y, we consider an arbitaryFourier mode in x. We let
u1(x, y) = u1(n, y)e2πinx, (7.27)
c1(x, y) = c1(n, y)e2πinx, (7.28)
and occasionally write u1(y) and c1(y) when n is understood. Equations (7.24)–(7.26) now become
∂yyu1 + 4π2n2u1 + ∂y(u1∂yc0) − 4π2n2u0c1 + ∂y(u0∂y c1) = λu1 on [0, 1], (7.29)
−∂yy c1 + 4π2n2c1 = u1 on [0, 1], (7.30)
∂yu1 = 0 for y = 0, 1,(7.31)
∂y c1 = 0 for y = 0, 1.(7.32)
7.3.1 Discretization
We now discretize (7.29)–(7.32). To do so, we introduce a cell centered gridover [0, 1] with spacing ∆y. Let N = 1/∆y. We denote the values of u1(y) andc1(y) at the grid points yk = (k − 1
2)∆y by u1,k and c1,k, respectively. Equations
(7.30) and (7.32) become
c1,k+1 − c1,k
∆y+c1,k−1 − c1,k
∆y+ 4π2n2c1,k∆y = u1,k∆y. (7.33)
We incorporate the no-flux boundary conditions (7.32) by dropping either thefirst or second term for the right- and left-most nodes, respectively. If we writeu, c as the column vectors corresponding the values u1,k and c1,k for k = 1, · · · , N .We write (7.33) compactly as
Lcc = u. (7.34)
Denoting the i-th row of the matrix L−1c as Lc−1,i, we now discretize (7.29) as
u1,i+1 − u1,i
∆y+u1,i+1 + u1,i
2
c0,i − c0,i+1
∆y+u0,i+1 + u0,i
2
L−1c,i − L−1
c,i+1
∆yu
+u1,i−1 − u1,i
∆y+u1,i−1 + u1,i
2
c0,i − c0,i−1
∆y+u0,i−1 + u0,i
2
L−1c,i − L−1
c,i−1
∆yu
+ 4πn2(ui − u0L−1c,i u)∆y = λui∆y. (7.35)
92

Note that the use of the explicit inverse to Lc makes this linear system non-sparse.We rewrite (7.35) as
Luu = λu, (7.36)
and let λ1 be the smallest eigenvalue of Lu.
7.3.2 Method
To ascertain the linear stability of the non-uniform x-invariant steady state,we first compute this steady state by evolving the one-dimensional Keller-Segelequation over [0, 1] analogously to the procedure in Section 7.2 with noisy uniforminitial data. For masses above π2, the solution settles onto a non-uniform profile.With this steady u0(y), c0(y), we compute the smallest eigenvalue of the Lu matrixfrom (7.36) by the ‘eig’ function in MATLAB 7.6.0 (R2008a). Motivated by theobservation from the simulation in Figure 7.2, we only consider modes with n = 1.As expected, simulations not presented showed that for n > 1, the least eigenvalueof the system is higher than for n = 1.
7.3.3 Results for Numerical Stability Analysis
Figure 7.3 plots the dependence of the smallest eigenvalue λ1 of the discretiza-tion to the linearized system (7.29)–(7.32) as a function of the system mass. Re-call that the mass enters the system by setting the one-dimensional steady stateu0(y), c0(y). As shown, the eigenvalues cross below zero around mass M∗ ≈ 4.57π.Hence, we expect the one-dimensional steady profile to be linearly stable for massesbelow M∗ and linearly unstable for masses above M∗.
7.4 Discussion
We now discuss the simulation results as motivation for the boundary blow-up conjecture. Our first line of evidence in support of the conjecture is the pairof simulations of the Keller-Segel evolution for mass M = 4.5π and M = 4.6π,initially distributed in a noisy uniform shape. Recall that the M = 4.5π simula-tion settled to a non-uniform x-invariant state, whereas the M = 4.6π simulationapproached such a state but eventually formed an x-dependent disturbance thatgrew until blow-up on the boundary. Our second line of evidence is that thesenon-uniform x-invariant steady states are linearly unstable for M > M∗ ≈ 4.57πand are linearly stable for M < M∗.
We now justify the first part of the conjecture, that boundary blow-up canfail to occur when 4π < M < M∗. The M = 4.5π simulation strongly suggests
93

4 4.2 4.4 4.6 4.8 5−10
−5
0
5
10
15
Mass (multiples of π)
Smallest Eigenvalue vs. Mass
Figure 7.3: Plot of the smallest eigenvalue of the discretization of the Keller-Segel equations over T× [0, 1] with (JL) boundary conditions, linearized about theone-dimensional non-uniform steady state with mass M . A negative eigenvalueindicates a linear instability. The 1d steady states change from being linearlystable to linearly unstable as mass increased beyond M∗ ≈ 4.57π.
94

that its solution converges to the one-dimensional state. The possibility that thesolution would have resulted in blow-up if the simulation were run for a longertime is unlikely, as the steady state was numerically linearly stable.
We now rationalize the second part of the conjecture, that boundary blow-upwill occur when M > M∗, provided the initial data is not symmetric. We makethis claim based only off simulations with noisy uniform initial data. Two reasonsfor such a broad generalization over initial data are (1) noisy uniform data isthe opposite extreme from the localization hypothesis of the theorem presented inSection 7.1; and (2) if the solution fails to blow up, it will settle to a steady state,which the simulations approach anyway. The numerical stability analysis andnumerical evolution both show that the relevant steady state becomes unstablefor M > M∗. We note that the asymmetry assumption is required because, forexample, an x-invariant solution stays x-invariant. Blow-up is not possible in thisone-dimensional problem [9].
We argue that domains like T × [0, 1] are a potentially fruitful arena for un-derstanding boundary blow-up of the Keller-Segel equations with (JL) boundaryconditions. The simulations suggest that one-dimensional steady states play an im-portant role in blow up. Their simple description should aid analysis, as comparedto their analogues in other domains.
7.5 Convergence Study for Evolution Simulation
We now show the spatial and convergence of the evolution simulation in Sec-tion 7.2.
7.5.1 Spatial Convergence Study
To verify the second order spatial accuracy we expect from our numericalscheme (7.12)–(7.15), we run several simulations for an empirical convergencestudy. Let u∆x be the computed solution to the (7.5)–(7.8) with
∆t = 0.005,∆x = ∆y = 0.1 · 2−k,
where k = 0, · · · , 4. Note that in increase in k by 1 corresponds to a doubling ofthe spatial resolution in both directions. We let the initial data for the simulationsbe
u|t=0 = C
(
1 +1√
2πσ2exp
(
−(x− .45)2 + (y − .45)2
2σ2
))
, (7.37)
σ = 0.125, (7.38)
where C is the constant that makes the mass equal to M .
95

Parameters Lp rspacep [u; 0.1] rspace
p [u; 0.05] rspacep [u; 0.025]
L1 2.01 2.00 1.99M = 5π L2 2.01 2.00 1.98
L∞ 1.99 1.99 1.59
Table 7.1: Spatial convergence rates for Keller-Segel simulation computed at time0.25.
We define the error between the k-th and the k + 1-st solutions as
espacep [u; ∆x] = ||u∆x − I∆x/2→∆xu∆x/2||p,
where I∆x/2→∆x is the interpolation operator between the finer to the coarser two-dimensional grid obtained by averaging adjacent interior nodes, and where || · ||p isthe discretization of the p-norm. We also define an empirical spatial convergencerate as
rspacep [u; ∆x] = log2
espacep [u; ∆x]
espacep [u; ∆x/2]
.
Table 7.1 presents the empirical rates of convergence for the L1, L2, and L∞ norms.We observe that the convergence appears to be second order in space.
7.5.2 Temporal Convergence Study
To verify the second order temporal accuracy of our numerical scheme, we runseveral simulations for an empirical convergence study. Let u∆t be the computedsolution to the homogenized system with
∆t = 0.01 · 2−k,∆x = ∆y = 0.025,
where k = 0, · · · , 4. Note that in increase in k by 1 corresponds to a doubling ofthe spatial resolution. We provide the simulations with the same initial conditions,(7.37)–(7.38).
We define the error between the k-th and the k + 1-st solutions as
etimep [u; ∆t] = ||u∆t − u∆t/2||p,
where ||·||p is the discretization of the p-norm. We also define an empirical temporalconvergence rate as
rtimep [u; ∆t] = log2
etimep [u; ∆t]
etimep [u; ∆t/2]
.
96

Parameters Lp rtimep [φ; 0.01] rtime
p [φ; 0.005] rtimep [φ; 0.0025]
L1 1.89 1.94 1.97M = 5π L2 1.94 1.94 1.97
L∞ 1.89 1.94 1.97
Table 7.2: Temporal convergence rates computed at time 0.25.
Table 7.2 presents the empirical rates of convergence for intracellular potential inthe homogenized system under the L1, L2, and L∞ norms. We observe that theconvergence appears to be second order in time. We remark that given the blow-up behavior of the system, any fixed value of ∆x and ∆t eventually becomes toosmall. As we are most interested in the instabilitie that lead to blow-up, we use thevalues of ∆t and ∆x that showed second order temporal and spatial convergencewhen initial data is not particularly localized.
97

Chapter 8
A Free Energy Stability Criterion
8.1 Introduction
The Keller-Segel system for chemotaxis over a bounded domain with (JL)boundary conditions is
∂tu+ ∇ · (−∇u+ χu∇c) = 0 in Ω, (8.1)
−∆c = u− u in Ω, (8.2)
∂νu = ∂νc = 0 on ∂Ω, (8.3)
where Ω is a subset of R2 with smooth boundary, u(t, x) corresponds to the density
of bacteria, c(t, x) corresponds to the density of the chemo-attractant, χ is a scalarsensitivity, and u = 1
|Ω|∫
Ωudx.
As detailed in Section 5.3, solutions to Keller-Segel equations have a free energythat is decreasing in time. That is,
∂t
∫
Ω
(
u logu− 1
2|∇c|2
)
dx = −∫
Ω
u|∇(log u− c)|2. (8.4)
We let F [u] =∫
Ωu log u − 1
2|∇c|2dx be the free energy. Note that c is not an
argument for F as it is given in terms of u by (8.2).The free energy functional has been an important tool in mathematical analysis
of Keller-Segel equations. For example, equation (8.4) gives a description of steadystates, namely those such that log u−c = constant. Senba and Suzuki [53] providean extensive study and bifurcation diagram of the structure of steady solutionsover the disk for Keller-Segel with Neumann boundary conditions. They also studylinearized stability of steady states at slightly superthreshold masses by analyzingeigenvalues of the linearized differential operator.
The free energy also provides motivation for viewing the Keller-Segel equationsas a gradient flow under the Wasserstein metric [1].
98

In this chapter, we use the free energy to derive an inequality that could beused to assess the stability of steady states to the system (8.1)–(8.3). The chapterproceeds as follows. In Section 8.2, we present the inequality and its derivation.In Section 8.3, we demonstrate its application in determining the critical massbeyond which uniform steady states become unstable. We also discuss benefitsand problems with the inequality as a tool for analysis.
8.2 Free Energy Inequality
We put forth the following tool for ascertaining a certain kind of stability ofsteady states to the Keller-Segel system (8.1)–(8.3). Let u0, c0 be such a steadysolution. We call it free-energy unstable if ∃u1 with u1 = 0 such that
∫
Ω
u21
u0dx <
∫
Ω
|∇∆−1u1|2dx. (8.5)
We call the solution free-energy stable if ∀ sufficiently regular u1 such that u1 = 0,
∫
Ω
u21
u0
dx >
∫
Ω
|∇∆−1u1|2dx. (8.6)
8.2.1 Motivation and Derivation
We begin by repeating that the free energy of solutions to the Keller segelsystem is decreasing. Thus, if nearby functions to u0 have smaller free energy thanu0, then some particular small perturbations must evolve away from u0. On theother hand, if all nearby functions to u0 give a larger free energy than u0, that isa suggestion of some sort of stability of the solution.
To derive the free energy inequality, we study perturbations to F due to smallchanges in u. Let
u(x) = u0(x) + εu1(x), u1 = 0, (8.7)
c(x) = c0(x) + εc1(x), −∆c1 = u1, c1 = 0. (8.8)
We now expand out F [u] in powers of ε:
F [u] = F [u0] + ε
∫
Ω
u1(log u0 − c0)dx+ ε2
∫
Ω
1
2
u21
u0− 1
2|∇c1|2 +O(ε3). (8.9)
If (u0(x), c0(x)) is a steady state, then log u0 − c0 = constant, and the order ε
term drops out. Hence, the sign of∫
Ω
u21
u0− |∇∆−1u1|2 determines whether or not
perturbations in the direction of u1 result in lower free energies than u0.
99

8.3 Application and Discussion
8.3.1 Stability of Uniform Profiles
The free energy inequality can be used to ascertain at which masses the uni-form solution to the Keller-Segel equations with (JL) boundary conditions becomesunstable, as given in the following proposition.
Proposition 8.1. If M < |Ω| · λ21 then the uniform profile u0 ≡ M
|Ω| is free energy
unstable. If M > |Ω| · λ21, then the uniform profile is free energy stable.
Proof of Proposition 8.1. Let φn∞n=0 form an orthonormal basis of eigenfunctinosof −∆ under homogeneous Neumann boundary conditions, with eigenvalues λ2
n.We expand u1 as
u1(x) =∞
∑
n=1
u1(n)φn(x), (8.10)
and we write the left hand side of the inequality as
∫
Ω
u21
u0
dx =1
u0
∫
Ω
u21dx =
1
u0
∞∑
n=1
|u1(n)|2. (8.11)
The right hand side of the inequality is
∫
Ω
|∇∆−1u1|2 =
∫
Ω
∇(−∆)−1u1 · ∇(−∆)−1u1dx (8.12)
=
∫
Ω
(−∆)−1u1 · u1dx (8.13)
=
∞∑
n=1
u1(n)φn
λ2n
·∞
∑
m=1
u1(m)φm (8.14)
=
∞∑
n=1
u1(n)2
λ2n
. (8.15)
If λ21 is the smallest non-zero eigenvalue, then u0 < λ2
1 ensures∫
Ω
u21
u0<
∫
Ω|∇∆−1u1|2.
If u0 > λ21, u1 = φ1 provides an example where
∫
Ω
u21
u0>
∫
Ω|∇∆−1u1|2. The propo-
sition is completed by noting that u0 = M/|Ω|.
We note that the inequality is easy to analyze about a uniform profile becausethe u0 is constant. We also note that the same result can be showed by linearizingthe Keller-Segel equations about a constant u0.
100

Applying Proposition 8.1 or a linearization argument, we remark that theuniform profile loses stability at mass π2 for the domain T × [0, 1] consideredin Chapter 7. For the disk B1(0), the uniform profile loses stability at a massα2
1,1π ≈ 3.39π < 4π, where α1,1 ≈ 1.8412 is the smallest non-zero root of J ′1(r),
where J1(r) is the Bessel function of the first kind of order 1.
8.3.2 Differences with Linear Stability
It is likely, but not obvious, that free-energy stability and linearized stabilityare the same notion. While the uniform profile is easy to analyze in both interpre-tations, perturbations about non-uniform profiles are not.
Linearizing the Keller-Segel equations about (u0, c0) gives rise to the system
∂tu1 − ∆u1 + ∇ · (u1∇c0 + u0∇c1) = 0 in Ω, (8.16)
−∆c1 = u1 in Ω, (8.17)
∂νu1 = ∂νc1 = 0 on ∂Ω, (8.18)
u1 = 0. (8.19)
The prime candidate for the u1 satisfying the free energy inequality (8.5) is theeigenfunction of the differential operator in (8.16)–(8.17) corresponding to theunsteady modes of the system. However, we do not have a calculation showing(8.5) is satisfied for such a function.
8.3.3 Does the Free Energy Inequality Aid Analysis?
There is some hope that the free energy inequality can aid analysis of steadystates to the Keller-Segel systems. Its form as a functional inequality is markedlydifferent than the evolution problem or its linearization. An especially nice featureof the inequality is how it does not involve c0 = (−∆−1)(u0 − u0) explicitly, and itonly involves u0 in one of the terms.
Unfortunately, without an explicit form for the steady solution u0, it maybe difficult to investigate the inequality analytically. The outlook for numericalinvestigation is not much better, as the inequality gives no guidance for whichfunctions u1 to test.
101

Bibliography
[1] Blanchet, A., Calvez, B., Carrillo, J., 2008. Convergence of the mass-transportsteepest descent scheme for the subcritical Patlak-Keller-Segel model. SIAMJ. Numer. Anal. 46(2), 691-721.
[2] Biler, P., Nadzieja, T., 1994. Existence and nonexistence of solutions for amodel of gravitational interaction of particles I, Colloq. Math. 66, 319-334.
[3] Biler, P., Karch, G., Laurencot, P., Nadzieja, T., 2006. The 8π-problem forradially symmetric solutions of a chemotaxis model in the plane. Math. Meth-ods. Appl. Sci. 29(13), 1563-1583.
[4] Biler, P., Karch, G., Laurencot, P., Nadzieja, T., 2006 The 8π-problem forradially symmetric solutions of a chemotaxis model in a disc. Topol. MethodsNonlinear Anal. 27 (1), 133-147.
[5] Blanchet, A., Carrillo, J., Masmoudi, N., 2007. Infinite Time Aggregationfor the Critical Patlak-Keller-Segel model in R
2. Comm. Pure Appl. Math.61(10), 1449-1481.
[6] Camelliti,P., Green, C., Kohl, P., 2006. Structural and functional coupling ofcardiac myocytes and fibroblasts. Dhein, S. (ed): Cardiovascular Gap Junc-tions, Adv Cardiol. Basel, Karger, 42. 132–149.
[7] Chang, S., Yang, P., 1988. Conformal deformation of metrics on S2. J. Differ-ential Geometry 27, 259-296.
[8] Chen,L., Meng, M., 1995. Compact and scattered gap junctions in diffusionmediated cell-cell communication. J. Theor. Biol. 176, 39-45.
[9] Childress, S., Percus, J., 1981. Nonlinear aspects of chemotaxis. Math. Biosci.56, 217-237.
[10] Childress, S., 1984. Chemotactic collapse in two dimensions. Lecture Notes inBiomath., 55, Springer, Berlin, 61-66.
102

[11] Clerc,L., 1976. Directional differences of impulse spread in trabecular musclefrom mammalian heart. J. Physiol. 255, 335–346.
[12] Dolbeault, J., Perthame, B., 2004. Optimal critical mass in the two-dimensional Keller-Segel model in R
2. C.R. Math. Acad. Sci. Paris, 339, 611-616.
[13] Evans, L., 2000. Partial Differential Equations. American Mathematical Soci-ety. Providence.
[14] Faber, G., 2008. Cardiac Bioelectricity Research and Train-ing Center (CBRTC). The Luo-Rudy dynamic (LRd) modelof the mammalian ventricular action potential. Available at:http://www.cwru.edu/med/CBRTC/LRdOnline/. Accessed December2008.
[15] Faber, G., Rudy, Y., 2000. Action potential and contractility changes in [Na+]ioverloaded cardiac myocytes: a simulation study. Biophys. J. 78, 2392-2404.
[16] Fall, C., Marland, E., Wagner, J., Tyson, J. (eds.), 2002. Computational CellBiology. Springer Verlag, New York.
[17] Filbet, F., 2006. A finite volume scheme for the Patlak-Keller-Segel chemotaxismodel. Numer. Math. 104, 457-488.
[18] Fishman,G., 2008. Personal Communication.
[19] FitzHugh, R., 1961. Impulses and physiological states in theoretical modelsof nerve membrane. Biophys. J., 1, 445-466.
[20] Gutstein, D., Morley, G., Tamaddon, H., Vaidya, D., Schneider, M., Chen, J.,Chien, K., Stuhlmann, H., Rishman, G., 2001. Conduction slowing and suddenarrhythmic death in mice with cardiac-restricted inactivation of connexin43.Circ. Res. 88. 333-339.
[21] Guyton, A., Hall, J., 1996. Textbook of Medical Physiology: W.B. Saunders.Philadelphia.
[22] Henriquez,C., 1993. Simulating the electrical behavior of cardiac tissue usingthe bidomain model. Crit. Rev. Biomed. Eng. 21, 1–77.
[23] Hodgkin, A., Huxley, A., 1952. A quantitative description of membrane cur-rent and its application to conduction and excitation in nerve. J. Physiol.(Lond.). 117, 500-544.
103

[24] Hoppensteadt, F., Peskin, C., 1992. Mathematics in Medicine and the LifeSciences. Springer-Verlag, New York.
[25] Hubbard,M., Ying,W. Henriquez,C.,2007. Effect of gap junction distributionon impulse propagation in a monolayer of myocytes: a model study. Europace9, vi20–vi28.
[26] Jager, W., Luckhaus, S., 1992. On explosions of solutions to a system ofpartial differential equations modelling chemotaxis. Trans. Amer. Math. Soc.329, 819-824.
[27] Keener, J., 1996. Direct Activation and Defibrillation of Cardiac Tissue. J.Theor. Biol, 178, 313-324.
[28] Keller, E., Segel, L., 1970. Initiation of slime mold aggregation viewed as aninstability. J. Theor. Biol. 26, 399-415.
[29] Keener,J., Sneyd,J., 1998. Mathematical Physiology. Springer.
[30] Kleber,A, Riegger,C., 1987. Electrical constants of arterially perfused rabbitpapillary muscle. J. Physiol. 385, 307–324.
[31] Kohl, P., Camelliti, P., Vurton, F., Smith, G., 2005. Electrical coupling offibroblasts and myocytes: relevance for cardiac propagation. J. Electrocard,38, 45–50.
[32] Krassowska, W., Pilkington, T., Ideker, R., 1990. Potential Distribution inThree-Dimensional Periodic Myocardium - Part I: Solution with Two-ScaleAsymptotic Analysis. IEEE Trans. Biomed. Eng. 37(3), 252–266.
[33] Kucera, J., Rorh, S., Rudy, Y., 2002. Localization of Sodium Channels inIntercalated Disks Modulates Cardiac Conduction. Circ. Res. 91, 1176-1182.
[34] Luo, C., Rudy, Y., 1991. A model of the ventricular cardiac action potential.Depolarization, repolarization, and their interaction. Circ. Res. 68, 1501-1526.
[35] Luo, C., Rudy, Y., 1994. A dynamic model of the cardiac ventricular actionpotential, I: simulations of ionic currents and concentration changes. Circ.Res.74, 1071-1096.
[36] Mori, Y., Fishman, G., Peskin, C., 2008. Ephaptic conduction in a cardiacstrand model with 3D eletrodiffusion. Proc. Nat. Acad. Sci. 105, 6463–6468.
[37] Marrocco, A., 2003. Numerical Simulation of Chemotactic Bacteria Aggre-gation via Mixed Finite Elements. ESAIM: Math. Model. Num. Anal. 37(4),617-630.
104

[38] McKean, H., 1970. Nagumo’s equation. Adv. in Math. 4, 209-223.
[39] Murray, J., 2002. Mathematical Biology I: An Introduction. Springer. NewYork.
[40] Murray, J. , 2002. Mathematical Biology II: Spatial Models and BiomedicalApplications. Springer. New York.
[41] Nagai, T., 2001. Blowup of Nonradial Solutions to Parabolic-Elliptic SystemsModeling Chemotaxis in Two-dimensional Domains. J. Inequal. Appl, 6, 37–55.
[42] Nagai, T., 1995. Blow-up of radially symmetric solutions to a chemotaxissystem. Adv. Math. Sci. Appl., 5, 581–601.
[43] Nagumo, J., Arimoto, S., Yoshizawa, S., 1962. An active pulse transmissionline simulating nerve axon. Proc. IRE, 50, 2061-2071.
[44] Neu,J., Krassowska, W., 1993. Homogenization of Syncytial Tissues. Crit.Rev. Biomed. Eng. 21, 137–199.
[45] Panfilov,A., Keldermann,R., Nash,M., 2007. Drift and breakup of spiral wavesin reaction-diffusion-mechanics systems. Proc. Nat. Acad. Sci. 104, 7922–7926.
[46] Roberts,D., Hersh,L., Scher, A., 1979. Influence of cardiac fiber orientation onwavefront voltage, conduction velocity, and tissue resistivity in the dog. Circ.Res. 44, 701–712.
[47] Rorh, S., 2004. Role of gap junctions in the propagation of the cardiac actionpotential. Cardiovasc. Res. 62, 309-322.
[48] Roberts,D., Scher,A., 1982. Effect of tissue anisotropy on extracellular poten-tial fields in canine myocardium in situe. Circ. Res. 50, 342–351.
[49] Shaw,R., Rudy,Y., 1997. Ionic mechanisms of propagation in cardiac tissue -Roles of the sodium and L-type calcium currents during reduced excitabilityand decreased gap junction coupling. Circ. Res. 81, 727–741.
[50] Sperelakis, N. McConnell, K., 2002. Electric Field Interactions BetweenClosely Abutting Excitable Cells. IEEE Eng. Med. Biol. Mag. 21. 77-89.
[51] Stinstra, J., Hopenfeld, B., Macleod, R., 2005. On the Passive Cardiac Con-ductivity. Ann. Biomed. Eng. 33, 1743–1751.
[52] Suzuki, T., 2005. Free Energy and Self-Interacting Particles. Birkhauser.Boston.
105

[53] Senba, T., Suzuki, T., 2000. Some Structures of the Solution Set for a Sta-tionary System of Chemotaxis. Adv. Math. Sci. Appl. 10 (1), 191-224.
[54] Senba, T., Suzuki, T., 2001. Parabolic system of chemotaxis: blowup in afinite and the infinite time. Meth. Appl. Anal. 8, 349-368.
[55] Weidmann,S., 1970. Electrical constants of trabecular muscle from mam-malian heart. J. Physiol. 210, 1041–1054.
[56] Yao, J., Gutstein, D., Liu, F., Fishman, G., Wit, A., 2003. Cell Coupling Be-tween Ventricular Myocyte Pairs From Connexin43-Deficient Murine Hearts.Circ. Res. 93, 736–743.
106