Embed Size (px)
Citation preview

frbiol$502
Freshwater Biology (1996) 35, 301–309
Hydraulic influences on periphyton and benthicmacroinvertebrates: simulating the effects ofupstream bed roughness
J O H N M . Q U I N N , C H R I S T O P H E R W. H I C K E Y A N D WAY N E L I N K L A T E R *NIWA, National Institute of Water and Atmospheric Research, Box 11115, Hamilton, New Zealand
*Present address: Ecology Department, Massey University, Private Bag 222, Palmerston North, New Zealand
S U M M A R Y
1. Hydraulic conditions, periphyton biomass and invertebrate communities werecompared on artificial substrates exposed to a range of upstream roughness conditionsacross an area of uniform current velocity and depth in a gravel-bedded river. The effectof river bed roughness was simulated by installing roughness elements upstream ofartificial substrates.2. Increasing upstream roughness reduced the average near-bed velocity above thesubstrates and increased short-term variability in velocity (i.e. turbulence).3. Periphyton chlorophyll a density showed a general decline with near-bed velocityand was significantly lower on the substrates exposed to the river bed reference and0 mm roughness treatments than the 110 mm roughness elements. Chlorophyll a wasalso negatively correlated with the abundance of larger collector-browsing invertebrates.This indicates that effects of the changes in hydraulic conditions on invertebrates mayhave contributed to the observed treatment effects on periphyton.4. Invertebrate abundance and diversity declined with increasing upstream roughness.Filter-feeders, collector-browsers and predatory invertebrates all declined in abundancewith increasing upstream roughness, but the effect was strongest for filter-feeders. Eightof the nine most common taxa showed significant treatment effects. The orthocladchironomid, Eukiefferiella sp., was not influenced strongly by upstream roughness, but itsabundance was correlated significantly with periphyton biomass.
Introduction
Hydraulic conditions influence the distribution and gain nutrient and dissolved gases in given hydraulicconditions (Wiley & Kohler, 1980; Stevenson, 1984;abundance of both periphyton (Stevenson, 1984; Poff
et al., 1990; Biggs & Hickey, 1994) and invertebrates Weissenberger et al., 1991). Conditions of flow alsocorrelate with bedload sediment transport, which can(Statzner, Gore & Resh, 1988; Jowett et al., 1991; Quinn
& Hickey, 1994) in rivers. Near-bed velocity and affect biota on exposed surfaces. Furthermore, filter-feeders depend on water movement to feed and showturbulence are expected to be particularly important
(Statzner et al., 1988), and increasing upstream bed strong preferences for areas of high near-bed velocity(Quinn & Hickey, 1994). Thus, upstream roughness isroughness (e.g. due to increasing particle size or
woody debris deposition) is predicted by hydraulic likely to have an important influence on patchdynamics of periphyton and invertebrates in streams.theory to reduce near-bed velocities and increase
turbulence (Carling, 1992). We hypothesized that responses of benthic inverteb-rates to the changes in hydraulic conditions due toPeriphyton and invertebrate taxa differ in their
ability to colonize free surfaces, maintain position, and different upstream roughness would vary between
© 1996 Blackwell Science Ltd 301

302 J.M. Quinn, C.W. Hickey and W. Linklater
different functional feeding groups. Specifically, we unglazed clay tiles glued in the centre of100 3 200 3 7-mm tiles (Fig. 1). Ten substrates werehypothesized that filter-feeders would be more
sensitive to the changes in hydraulic conditions than attached by flat head nails to each plywood sheet onthe downstream margin, centred three pipe diameterscollector-browsers and that their abundance would be
inversely related to the size of the upstream roughness from the last pipe. Refuges, similar to the interstitialspaces between stones, were provided by 2-mm-highelements. We also expected that lower retention of
particulate organic matter and lower near-bed current ‘feet’ on the undersides of the tiles. The feet resultedin a 2-mm gap between the upper and lower tile andvelocities would result in lower densities of the col-
lector-browsers downstream of larger roughness between the lower tile and the platform. Three grooves(3-mm wide, 3.5-mm deep) were cut along the upperelements.
In this study, we investigated the effects of upstream tiles to provide potential sites for colonization by filter-feeding hydropsychid caddis (Downes & Jordan, 1993).roughness on periphyton biomass and invertebrate
community structure in an experimental manipulation The platforms were placed across a 18-m-wide areaof uniform current velocity and depth (see Table 1) inacross a run of relatively uniform current velocity
and depth in the gravel-bedded Mohaka River. The a 120-m-wide run downstream of a 350-m-long pool.Plywood treatment locations were allocated randomlyupstream roughness was altered by attaching pipes of
varying diameters (26–160 mm) to platforms, and the to avoid systematic bias. The sequence of treatmentsacross the river from right to left was 160, 60, 26, 0effects on the hydraulic conditions, periphyton bio-
mass and invertebrate communities that colonized and 110 mm. The natural river bed substrates (‘R’treatment) were mounted with the same orientationsubstrates placed downstream were measured.on a 0.15 3 1.2 m plywood platform held across thecurrent so that the upstream edges of the tiles rested
Materials and methodson the river bed. This treatment did not affect thedownstream hydraulic conditions and was placedThe experiments were carried out in the Mohaka River
(mean annual flow 30 m3 s–1; 176°389 E, 39°129 S). The upstream of the other treatments. The depth (D) andmean velocity (U, mean over 20 s at six-tenths ofMohaka drains the Kaimanawa Ranges east of Lake
Taupo in the central North Island of New Zealand. The depth) were measured 2 m upstream of the treatmentplatforms using an electromagnetic current metercatchment is 92% native forest, scrub and undeveloped
grassland, but land use around the study site is (Montedoro-Whitney Model PVM–2 A) attached to agauging rod. These measurements were used to calcu-pastoral agriculture. At base flow, the river water is
typically clear (black disc visibility 2.5–6.0 m) and late the upstream Froude numbers [Fr 5 U/(gD)0.5,where g 5 acceleration due to gravity (cm s–2)].unenriched (dissolved reactive phosphorus 4–8 mg
m–3 and NO3-N 82–163 mg m–3; Close & Davies- The effects of the differences in upstream roughnesson the hydraulic habitat at the tiles were investigatedColley, 1990).
Communities were compared on artificial substrates by measuring vertical velocity profiles and short-termvariations in velocity (turbulence) at the bed. Profilesexposed to contrasting hydraulic environments. Levels
of upstream roughness were generated by mounting were measured at five depths, 2–25 cm above thecentre of the middle tile of each treatment at thedifferent sized PVC pipes on 2.4 m long 3 1.2 m wide
sheets of untreated plywood that were secured to the beginning of the experiment to provide an indicationof treatment effects. Velocities were averaged overstream bed with the pipes perpendicular to the flow
(Fig. 1). A flat plywood platform, without pipes, 20 s. Indicative shear velocities (Uf 5 b/5.75, where b 5
the slope of the logarithmic velocity profile [velocity vreduced the upstream roughness, whereas pipes withouter diameters of 26, 60, 110 and 160 mm provided log(depth); Smith, 1975)] were calculated from the
near-bed observations at 2, 5 and 10 cm above thea gradation of increasing upstream roughness condi-tions. The pipe spacing was six times the pipe diameter tiles. Treatment effects on turbulence were assessed
by measuring variations of velocity at 1-s intervals forto produce isolated roughness flows, generating con-siderable turbulence and near-maximal frictional res- a period of 3 min (Campbell CR-10 logger) at the
position of the middle of the centre tile, after firstistance (see Fig. 3 in Young, 1992).Artificial substrates comprised 100 3 100 3 7-mm removing the tile. The current meter head was placed
© 1996 Blackwell Science Ltd, Freshwater Biology, 35, 301–309

Hydraulic influences on periphyton and benthic macroinvertebrates 303
Table 1 Characterization of hydraulic conditions upstream of the platforms and shear velocity above the centre file. Treatmentcodes correspond to river reference (R) and the size of the upstream roughness elements
Initial conditions Final conditions
Shear velocity Velocity Depth Velocity DepthTreatment (cm s–1) (cm s–1) (cm) Froude (cm s–1) (cm) Froude
R 2.1 51 62 0.21 32 50 0.150 mm 3.0 64 60 0.26 41 48 0.1926 mm 3.8 60 60 0.25 38 47 0.1860 mm 6.7 52 60 0.21 33 46 0.16110 mm 10.8 58 60 0.24 39 48 0.18160 mm 1.8 48 67 0.19 26 54 0.11
at the height of the surface of the upper tile, 2 cmabove the platform. Because the sensors are on theunderside of the instrument head, this was invertedso that the sensors experienced similar down-wellingturbulence to the top of the tiles.
The tiles and associated periphyton and inverteb-rates were sampled by lifting them into a 500-µm-mesh net. Preliminary processing was carried outimmediately by rinsing and scrubbing the periphytonand invertebrates into a large white sorting tray usinga brush with stiff nylon bristles. All periphyton wasremoved from each of the upper and lower tiles offive substrates per treatment (i.e. every second tile) Fig. 2 Mohaka River hydrograph during the experiment.and stored separately on ice in the dark in 120-mlpolyethylene containers for transport to the laboratory
Statistical analyseswhere they were stored at 4 °C. Within 4 days ofcollection, these samples were homogenized in a The effects of the upstream roughness on periphytonblender, subsampled, concentrated onto GF/C filter biomass and the invertebrate community character-papers and analysed for chlorophyll a (tissue grinding istics were investigated using one-way ANOVA, withand 90% acetone extraction), adjusted for phaeophyton post-hoc Bonferroni multiple inference tests where(APHA, 1989). Periphyton densities were calculated ANOVAs were significant at P , 0.05 (Day & Quinn,per unit exposed surface area, including the upper 1989). The data were normalized by log transforma-surface and sides of tiles and allowing for the addi- tions where necessary. All statistical analyses weretional surface area due to the grooves in the upper tiles. carried out using Datadesk 4.2 (Data Description Inc.).
The invertebrates on each tile were sorted livefrom the periphyton in the field and, where possible,
Resultsidentified (using hand lenses with 310 magnification)and counted into vials containing 70% isopropyl alco- The experiment ran from 3 March until 24 March 1993.hol. Field identifications were checked and additional A small spate occurred on 5 March, when the flowidentifications were made in the laboratory under a increased by 2.4 times, but flows returned to levelsmicroscope (3100). All abundances were calculated similar to those on 3 March within 3 days and wereper 200 cm2 planar surface area of substrate. fairly steady for the remainder of the experiment
Taxon richness was calculated as the number of taxa (Fig. 2). The lower flows at the end of the experimentrecorded per substrate and community diversity was resulted in lower depths and velocities than at the
beginning (Table 1).calculated using the Shannon Index (Magurran, 1988).
© 1996 Blackwell Science Ltd, Freshwater Biology, 35, 301–309

304 J.M. Quinn, C.W. Hickey and W. Linklater
Fig. 4 Boxplots showing the variations in current velocityabove the centre tile of each treatment over 3 min at the end ofthe experiment. Bar represents the median, box theinterquartile range, and whiskers extend to highest and lowestdatum within 1.5 3 the interquartile range. Outliers andextreme outliers (.1.5 3 and .3 3 the interquartile range,respectively) are denoted as open circles and starbursts,
Fig. 3 Velocity profiles above the centre tile of each treatment respectively. See Table 1 for treatment codes.at the beginning of the experiment. See Table 1 for treatmentcodes.
Table 2 Characterization of hydraulic conditions at the surfaceof the centre tile using an inverted velocity sensor (1 sTreatment effects on benthic hydraulic habitatmeasurements for 3 min) at the end of the experiment
All experimental treatments were established withVelocity Coefficient ofsimilar upstream water velocity (mean 55 cm s–1),
Treatment (cm s–1) variation (%)depth (mean 62 cm) and Froude number (mean 0.23)(Table 1). The Froude numbers all corresponded to R 24 16.2
0 mm 28 11.7run habitat (Fr 0.18–0.41; Jowett, 1993). Water velocities26 mm 17 31.125 cm above the platforms were on average 30% higher60 mm 9.4 50.7
than the upstream flow velocity. During the 20-day 110 mm –4.2 155experimental period the flow declined from 19 to 160 mm –0.6 95315.5 m3 s–1 (Fig. 2), resulting in reductions in velocity(by 37%), depth (by 20%), and Froude number (by32%). The water depth above the platform was greater ive reduction in average current velocity near the bed
and an increase in the turbulence (Fig. 4 and Table 2).than three times the pipe diameter for all treatments,implying non-chaotic flow conditions (Davis & Downstream of the two largest roughness treatments
(110 and 160 mm) both the magnitude and directionBarmuta, 1989). Small surface waves were presentabove the 110- and 160-mm treatments. of velocity were highly variable, with strong upstream
current at times, resulting in average velocities in theThe treatments produced different velocity gradi-ents above the tiles (Fig. 3). Shear velocities increased upstream direction (Fig. 4 and Table 2). Instantaneous
velocity data showed periodic bursts of turbulencewith the size of the roughness elements up to 110 mm.However, downstream of the 160 mm roughness ele- occurring at 40–90-s intervals for all of the treatments.
The outlier values (Fig. 4) were all associated with thisments a zone of low velocity occurred up to 10 cmabove the substrates, resulting in low shear velocities bursting phenomenon which occurred on only a few
occasions during the velocity monitoring period.(Table 1). The medium–large gravels upstream of theriver bed treatment (R) produced a velocity profile Mean near-bed velocity and the coefficient of vari-
ation of near-bed velocity (Table 2) were stronglysimilar to the 0-mm treatment.Variation in current velocities measured at centre tile negatively correlated (Spearman correlation, rs 5
–0.94, P 5 0.02, n 5 6), but neither variable was correl-of each treatment provided a measure of turbulence.Increasing upstream roughness resulted in a progress- ated with the shear velocity measured at the beginning
© 1996 Blackwell Science Ltd, Freshwater Biology, 35, 301–309

Hydraulic influences on periphyton and benthic macroinvertebrates 305
dwellers. These counts undoubtedly underestimatedthe numbers of small chironomids hidden within theperiphyton. However, they indicate that during theday most of the larger invertebrates occupied the areasbetween and beneath the tiles that made up thesubstrates.
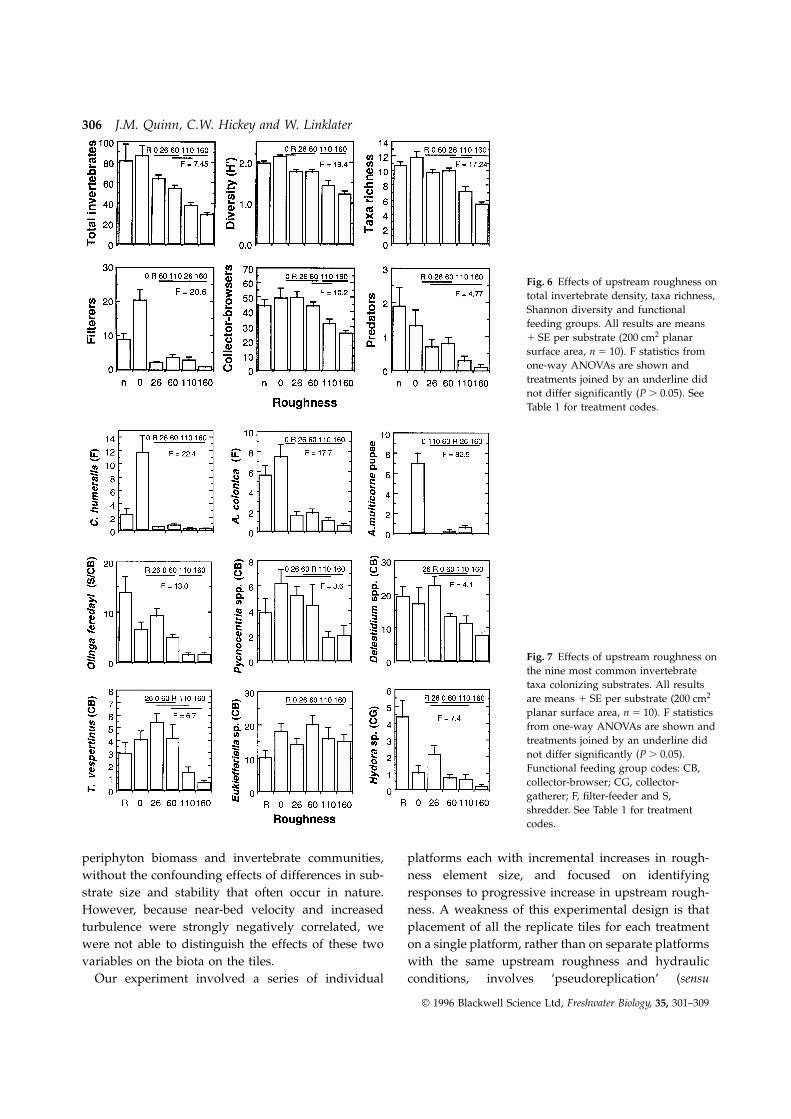
Upstream roughness had significant effects on thetotal density, taxon richness and diversity of the inver-tebrate community and the density of invertebratesbelonging to the main functional feeding groups(Fig. 6) and eight of the nine most common taxa(Fig. 7). Significant positive correlations (rs ù 0.829,P , 0.05, n 5 6) were found between the near-bedvelocity at the end of the experiment and the averagetreatment values of total invertebrate density, taxon
Fig. 5 Effects of upstream roughness on periphyton algal richness, Shannon diversity, and the densities of thebiomass on substrates (mean 1 SE, n 5 5). The treatments two dominant filter-feeders Coloburiscus humeralisjoined by an underline did not differ significantly in one-way (Walker) (Oligoneuriidae) and Aoteapsyche colonicaANOVAs (P . 0.05). See Table 1 for treatment codes.
(McLachlan) (Hydropsychidae). These invertebrateparameters were also significantly negatively correl-ated (rs ù –0.829, P , 0.05, n 5 6) with the coefficientof the experiment (Table 1; rs 5 –0.37 and 0.09, respect-
ively, n 5 6). of variation in velocity, but no significant relationshipswere found between the invertebrate variables andshear velocity.
Upstream roughness effects on periphyton
Mean periphyton chlorophyll a on the upper, groovedPeriphyton–invertebrate interactions
tiles of the substrates was 2.8-fold higher than onexposed areas of the lower, smooth tiles (F 5 53.2, Periphyton biomass (chlorophyll a) on individual sub-
strates was positively correlated with densities of theP , 0.0001 in a two-way ANOVA comparing upperand lower tiles and upstream roughness). Chlorophyll chironomid Eukiefferiella sp. (rs 5 0.67, P , 0.001) and
negatively correlated with taxon richness (rs 5 –0.53,a on the whole substrates (upper and lower portionscombined) showed a significant treatment effect P , 0.005), Shannon diversity (rs 5 –0.65, P , 0.001),
and densities of large collector-browsers (excluding(Fig. 5). Chlorophyll a was least on the substratesdownstream of the natural bed roughness and chironomids) (rs 5 –0.50, P , 0.01) and filterers (rs 5
–0.61, P , 0.001).increased with upstream roughness up to 110 mm, butdeclined at 160 mm. When the 160-mm data wereexcluded, chlorophyll a was significantly correlated
Discussion(P , 0.05, n 5 5) with each of near-bed velocity (rs 5
–0.90), shear velocity (rs 5 0.90) and coefficient of The velocity and depth conditions upstream and over-lying the experimental roughness treatments werevariation in velocity (rs 5 0.90).comparable with a run environment (Jowett, 1993)and environmental conditions with which diverse and
Effects of upstream roughness on invertebrateabundant invertebrate assemblages would be expected
colonization of tiles(Jowett et al., 1991; Quinn & Hickey, 1994). The experi-mental manipulations simulated isolated roughnessInvertebrates observed on the upper surfaces of the
tiles prior to sampling only accounted for 1–7% of the conditions, across an area of relatively uniformupstream hydraulic conditions. This experimentaltotal numbers recovered per substrate, with larvae
and pupae of the simuliid Austrosimilium multicorne design enabled investigation of the effects of changingnear-bed current velocity and turbulence regime onmulticorne Tonnoir comprising 60–100% of the surface-
© 1996 Blackwell Science Ltd, Freshwater Biology, 35, 301–309
306 J.M. Quinn, C.W. Hickey and W. Linklater
Fig. 6 Effects of upstream roughness ontotal invertebrate density, taxa richness,Shannon diversity and functionalfeeding groups. All results are means1 SE per substrate (200 cm2 planarsurface area, n 5 10). F statistics fromone-way ANOVAs are shown andtreatments joined by an underline didnot differ significantly (P . 0.05). SeeTable 1 for treatment codes.
Fig. 7 Effects of upstream roughness onthe nine most common invertebratetaxa colonizing substrates. All resultsare means 1 SE per substrate (200 cm2
planar surface area, n 5 10). F statisticsfrom one-way ANOVAs are shown andtreatments joined by an underline didnot differ significantly (P . 0.05).Functional feeding group codes: CB,collector-browser; CG, collector-gatherer; F, filter-feeder and S,shredder. See Table 1 for treatmentcodes.
periphyton biomass and invertebrate communities, platforms each with incremental increases in rough-ness element size, and focused on identifyingwithout the confounding effects of differences in sub-
strate size and stability that often occur in nature. responses to progressive increase in upstream rough-ness. A weakness of this experimental design is thatHowever, because near-bed velocity and increased
turbulence were strongly negatively correlated, we placement of all the replicate tiles for each treatmenton a single platform, rather than on separate platformswere not able to distinguish the effects of these two
variables on the biota on the tiles. with the same upstream roughness and hydraulicconditions, involves ‘pseudoreplication’ (sensuOur experiment involved a series of individual
© 1996 Blackwell Science Ltd, Freshwater Biology, 35, 301–309

Hydraulic influences on periphyton and benthic macroinvertebrates 307
Hurlbert, 1984). We were constrained in our ability to between chlorophyll a and near-bed velocity. Thesignificant increase in invertebrate densities with near-undertake a fully replicated experiment by two main
factors: (i) we had limited a priori knowledge of the bed velocity may have counteracted any stimulationof periphyton growth. These patterns in invertebratesize of upstream roughness element likely to produce
marked effects on downstream biota; and (ii) we had abundance, and potential effects of the treatments onbedload scour, make it difficult to identify directa limited area of uniform upstream velocity, depth
and particle size across the run at our site. We endeav- hydraulic effects on periphyton in this experiment.The differences in hydraulic conditions producedoured to reduce the risk of systematic variations in
conditions across the apparently uniform run influen- by the treatments had significant effects on the benthicinvertebrate community, with total invertebratecing our results by randomizing the placement of the
platforms. abundance and diversity, measured as taxon richnessand Shannon diversity, declining with increasingThe higher periphyton biomass (2.8-fold) on the
upper compared with the lower tiles of the substrates upstream roughness conditions. The three main func-tional feeding groups all declined with upstreamindicates that periphyton biomass was influenced
strongly by the small-scale roughness effects of the roughness. As predicted, this effect was strongest forfilterers (highest F statistics). Average densities ofsurface grooves that were present on the upper tiles.
Two factors that probably contributed to the enhanced predators and total invertebrates were strongly correl-ated, indicating that both direct hydraulic effects andgrowth on the grooved portions of the substrates
were the lower velocities expected within the grooves, indirect influences on prey abundance probably con-tributed to the treatment effects on predators.which can increase algal colonization rates (Stevenson,
1984), and the rough texture of the tile surface within Eight of the nine most common taxa also declined indensity downstream of the high roughness treatments,the grooves, which can also increase periphyton
accrual (Clifford, Casey & Saffran, 1992). with particularly strong effects observed for the dom-inant filter-feeding species Coloburiscus humeralis andThe presence of gravels on the platform around the
substrates exposed to the river bed roughness suggests Aoteapsyche colonica. The former two species have beenshown to avoid areas of low current velocity ingreater abrasion of these substrates by bedload may
have contributed to their low algal biomass. The previous studies in the Mohaka and other New Zea-land rivers (Jowett et al., 1991; Quinn & Hickey, 1994).presence of the pipes and the slight elevation off the
bed of the platforms would have restricted abrasion The high level of turbulent variations in velocity,with large pulses in velocity in both upstream andon the other treatments. Periphyton was also much
more apparent on platforms than on the surrounding downstream directions, probably made the substrateareas downstream of the larger roughness treatmentsriver bed gravels, which appeared bare, at the end
of the experiment. These observations suggest that particularly unsuitable for filter-feeders by ‘back-washing’ filtered materials from the nets of the hydro-bedload movement and abrasion may be an important
factor controlling periphyton in the Mohaka River. psychid A. colonica and captured on the leg hairs usedto filter seston by C. humeralis (Wisely, 1961).The low level of periphyton visible on platforms
around the substrates suggests that these may have Pupae of the simuliid Austrosimulium multicornemulticorne Tonnoir, which occupy fixed cocoons on theprovided a refuge for grazing invertebrates. The
abundance of periphyton on the large platforms may upper surfaces of the substrates, showed a particularlystrong preference for the tiles on the 0 mm roughnesshave also been favoured by the large distance from
refuges from hydraulic and/or predator disturbances. treatment (smooth plywood). The greater level ofoxygen transfer to the developing pupae, due to theThis hypothesis is being investigated in a current study.
Uptake of nutrients by periphyton is known to high current velocity near the bed, could account forthis preference.increase with current velocity (Lock & John, 1979;
Riber & Wetzel, 1987), and mass transfer of nutrients The collector-browsers Deleatidium spp. (Leptophle-biidae), Pycnocentria spp. (Conesucidae) and Tanytarsusmight be expected to limit growth rate in the relatively
oligotrophic Mohaka River. Thus an increase in chloro- vespertinus Hutton (Chironomidae), the facultativeshredder Olinga feredayi (McLachlin) (Conesucidae),phyll a with near-bed velocity might have been
expected. However, we observed a negative correlation and collector-gathering Hydora sp. larvae (Elmidae), all
© 1996 Blackwell Science Ltd, Freshwater Biology, 35, 301–309

308 J.M. Quinn, C.W. Hickey and W. Linklater
to a hydraulic gradient in a regulated river in Newshowed a general decline in abundance as roughnessZealand. Freshwater Biology, 32, 49–59.treatment increased above 26 mm. The low abundance
Carling P. (1992) The nature of the fluid boundary layerof these taxa downstream of the larger roughnessand the selection of parameters for benthic ecology.treatments is consistent with previous findings fromFreshwater Biology, 28, 273–284.surveys of distribution patterns within stream reaches
Clifford H.F., Casey R.J. & Saffran K.A. (1992) Short-termthat these taxa avoid areas of low current velocitycolonization of rough and smooth tiles by benthic(Jowett et al., 1991; Collier, 1993; Quinn & Hickey,macroinvertebrates and algae (chlorophyll a) in 2
1994). The additional stress of the turbulent currentsstreams. Journal of the North American Benthological
downstream of the larger roughness elements would Society, 11, 304–315.be expected to have contributed to their low densities Close M.E. & Davies-Colley R.J. (1990) Baseflow waterby making it difficult to hold position in the rapidly chemistry in New Zealand rivers. 1. Characterisation.changing flow environment and reducing the retention New Zealand Journal of Marine and Freshwater Research,of particulate organic matter relative to the lower 24, 319–341.roughness treatments where unidirectional currents Collier K.J. (1993) Flow preferences of larvaloccurred. Chironomidae (Diptera) in Tongariro River, New
Zealand. New Zealand Journal of Marine and FreshwaterThe abundance of the other common collector-Research, 27, 219–226.browser Eukiefferiella sp. did not appear to be related
Davis J.A. & Barmuta L.A. (1989) An ecologically usefuldirectly to the upstream roughness conditions, butclassification of mean and near-bed flows in streamswas strongly correlated with biomass of periphytonand rivers. Freshwater Biology, 21, 271–282.within which it lives. This suggests that the periphyton
Day R.W. & Quinn G.P. (1989) Comparisons of treatmentsgrowths provide a hydraulic refuge for this species.after an analysis of variance in ecology. EcologicalThis experiment has demonstrated that, for hydraul-Monographs, 59, 433–463.ically defined run habitats (Froude number 0.18–0.41;
Downes B.J. & Jordan J. (1993) Effects of stone topographyJowett, 1993), feeding groups of invertebrates differ
on abundance of net-building caddisfly larvae andin their sensitivity to the hydraulic effects of increased
arthropod diversity in an upland stream. Hydrobiologia,upstream bed roughness. Filter-feeders were sensitive 252, 163–174.to small increases in upstream roughness, whereas Hurlbert S.H. (1984) Pseudoreplication and the design ofcollector-browsers only declined significantly down- ecological field experiments. Ecological Monographs, 54,stream of larger roughness elements that induced 187–211.marked reductions in near-bed velocity and formation Jowett I.G. (1993) A method for objectively identifyingof turbulent eddies. pool, run, and riffle habitats from physical
measurements. New Zealand Journal of Marine andFreshwater Research, 27, 241–248.
Acknowledgments Jowett I.G., Richardson J., Biggs B.J.F., Hickey C.W. &Quinn J.M. (1991) Microhabitat preferences of benthicThanks to Morag Stroud and Mike Martin for assist-invertebrates and the development of generalisedance in the field. Peter Arnold provided flow data forDeleatidium spp. habitat suitability curves, applied to
the Mohaka at Glenfalls. The paper was improved byfour New Zealand rivers. New Zealand Journal of
constructive review comments from Ian Jowett andMarine and Freshwater Research, 25, 187–200.
two anonymous referees. This study was funded by Lock M.A. & John P.H. (1979) The effect of flowthe New Zealand Foundation for Research Science patterns on uptake of phosphorus by river periphyton.and Technology. Limnology and Oceanography, 24, 376–383.
Magurran A.E. (1988) Ecological Diversity and itsMeasurement. Croom Helm, London.
ReferencesPoff N.L., Voelz N.J., Ward J.V. & Lee R.E. (1990) Algal
colonization under four experimentally controlledAPHA (1989) Standard methods for the examination of watercurrent regimes in a high mountain stream. Journaland wastewater. American Public Health Association,of the North American Benthological Society, 9, 303–318.American Water Works Association, Water Pollution
Quinn J.M. & Hickey C.W. (1994) Hydraulic parametersControl Federation, Washington, DC.Biggs B.J.F. & Hickey C.W. (1994) Periphyton responses and benthic invertebrate distributions in two gravel-
© 1996 Blackwell Science Ltd, Freshwater Biology, 35, 301–309

Hydraulic influences on periphyton and benthic macroinvertebrates 309
bed New Zealand rivers. Freshwater Biology, 32, Weissenberger J., Spatz H.-Ch., Emanns A. &Schwoerbel J. (1991) Measurement of lift and drag489–500.forces in the mN range experienced by benthicRiber H.H. & Wetzel R.G. (1987) Boundary-layer andarthropods at flow velocities below 1.2 m s–1.internal diffusion effects on phosphorus fluxes inFreshwater Biology, 25, 21–31.lake periphyton. Limnology and Oceanography, 32,
Wiley M.J. & Kohler S.L. (1980) Position changes of1181–1194.mayfly nymphs due to behavioural regulation ofSmith I.R. (1975) Turbulence in lakes and rivers. Freshwateroxygen consumption. Canadian Journal of Zoology, 58,Biological Association Scientific Publication no., 29,618–622.
79 pp.Wisely B. (1961) Studies on Ephemeroptera. 1.
Statzner B., Gore J.A. & Resh V.H. (1988) Hydraulic Coloburiscus humeralis (Walker); early life history andstream ecology: observed patterns and potential nymph. Transactions of the Royal Society of New Zealandapplications. Journal of the North American Benthological Zoology, 1, 249–257.Society, 7, 307–360. Young W.J. (1992) Clarification of the criteria used to
Stevenson R.J. (1984) How currents on different sides identify near-bed flow regimes. Freshwater Biology, 28,of substrates in streams affect mechanisms of benthic 383–391.algae accumulation. Internationale Revue der Gesamten
(Manuscript accepted 5 January 1996)Hydrobiologie, 69, 241–262.
© 1996 Blackwell Science Ltd, Freshwater Biology, 35, 301–309

Fig. 1 Photographs showing the layout of the platforms across the Mohaka River and underwater upstream views of the sixtreatments at the end of the experiment. See Table 1 for treatment codes.





![Biological Monitoring two.ppt [Read-Only] - Lycoming … Monitoring-Benthic Macroinvertebrates-Periphyton, ... let critters take a ride on your net from one site to another ... (the](https://img.pdfslide.net/doc/110x75/5ab704197f8b9a86428e4add/biological-monitoring-twoppt-read-only-lycoming-monitoring-benthic-macroinvertebrates-periphyton.jpg)























