Embed Size (px)
Citation preview

Cell, Vol. 90, 65–75, July 11, 1997, Copyright 1997 by Cell Press
Identification and Structural Characterizationof the ATP/ADP-Binding Site in the Hsp90Molecular Chaperone
Chrisostomos Prodromou,* S. Mark Roe,*† GP96 or endoplasmin) localized to the endoplasmicreticulum, where it is involved in the assembly of immu-Ronan O’Brien,* John E. Ladbury,*noglobulins (Melnick et al., 1992) and other proteinsPeter W. Piper,* and Laurence H. Pearl*†
destined for secretion or surface presentation. GRP94/*Department of Biochemistry and Molecular BiologyGP96 is also involved in the loading of proteosome-†Joint UCL/LICR X-Ray Crystallography Laboratorygenerated antigenic peptides onto nascent MHC ClassUniversity College LondonI molecules (Li and Srivastava, 1993).Gower Street
Hsp90 alone can prevent protein aggregation and pro-London WC1E 6BTmote refolding in vitro (Weich et al., 1992), but in vivo itUnited Kingdomis functionally associated in multiprotein complexes witha range of accessory proteins. Initially, client proteinsare brought to Hsp90 in a complex involving the ATPaseSummarychaperone Hsp70/DnaK and cochaperones Ydj1/DnaJ(Kimura et al., 1993) and p48/Hip (Hohfeld et al., 1995),Hsp90 molecular chaperones in eukaryotic cells playwhich interact with Hsp90 via p60/Hop/Sti1 (Smith etessential roles in the folding and activation of a rangeal., 1993). These accessory proteins are subsequentlyof client proteins involved in cell cycle regulation, ste-replaced in the Hsp90–client complex by an immuno-roid hormone responsiveness, and signal transduc-philin (FKBP52, Cyp-40, etc.), when the client protein istion. The biochemical mechanism of Hsp90 is poorlya steroid receptor (Smith and Toft, 1993), or p50/CDC37understood, and the involvement of ATP in particular(Stepanova et al., 1996) when the client protein is ais controversial. Crystal structures of complexes be-protein kinase. p60/Hop/Sti1, p50/CDC37, and thetween the N-terminal domain of the yeast Hsp90 chap-various immunophilins compete for binding to Hsp90erone and ADP/ATP unambiguously identify a specific(Owens-Grillo et al., 1996) and are presumed to interactadenine nucleotide binding site homologous to thewith a common site. The final step of conformationalATP-binding site of DNA gyrase B. This site is the samematuration requires the acidic p23 protein, whose bind-as that identified for the antitumor agent geldana-ing to Hsp90 complexes appears to be ATP-dependantmycin, suggesting that geldanamycin acts by blocking(Johnson and Toft, 1994, 1995).thebinding of nucleotides to Hsp90and not the binding
Binding and hydrolysis of ATP is a well-describedof incompletely folded client polypeptides as pre-component of the molecular mechanisms of the Hsp70/viously suggested. These results finally resolve theDnaK (Flynn et al., 1991) and the Hsp60/GroEL (Rose-
question of the direct involvement of ATP in Hsp90man et al., 1996) classes of molecular chaperones. How-
function.ever, the involvement of ATP in the mechanism of actionof Hsp90 has been controversial for several years. Ob-
Introduction servations of apparent ATPase (Nadeau et al., 1992,1993) and autophosphorylation (Csermely and Kahn,
The 90 kDa heat shock protein family (Hsp90) are ubiqui- 1991) activities may have been due to trace amounts oftous molecular chaperones with essential roles in stress the various protein kinases for which Hsp90 is the spe-tolerance (Borkovich et al., 1989) and protein folding cific chaperone and/or contamination with the true(Freeman and Morimoto, 1996). In eukaryotes, the cyto- ATPase chaperone, Hsp70/DnaK, with which Hsp90 isplasmic Hsp90s act as specific chaperones for a wide functionally associated (Czar et al., 1994). Consistentrange of client proteins involved in signal transduction, with this, reports of ATPase and autophosphorylationcell cycle regulation, and hormone responsiveness. activities have been very variable between laboratoriesSpecific clients in mammalian cells include tyrosine ki- and appear to depend on the protein source and thenases such as pp60v-src (Opperman et al., 1981) and degree of purification attained (Weich et al., 1993).Sevenless (Cutforth and Rubin, 1994); serine/threonine Other work has suggested that ATP triggers substantialkinases such as Wee1 (Aligue et al., 1994), c-Raf (Stan- conformational change in Hsp90 (Csermely et al., 1993)cato et al., 1993), and Cdk4 (Dai et al., 1996); helix- and that ATP binding converts Hsp90 to a conforma-loop-helix transcription factors (Wilhelmson et al., 1990); tional form required for binding of the p23 maturationtumor suppressers such as Rb (Chen et al., 1996) and factor to Hsp90–client protein–immunophilin complexesp53 (Sepehrnia et al., 1996); and the cytoplasmic re- (Sullivan et al., 1997). However, a comparative study ofceptors for steroid hormones such as estrogen, proges- highly purified Hsp90 and Hsp70/DnaK (Jakob et al.,terone, and glucocorticoid (Joab et al., 1984). The 1996) has clearly demonstrated the absence of a signifi-involvement of several of these client proteins in cell cant inherent ATPase activity in Hsp90 compared withproliferation and tumor progression has prompted inter- Hsp70/DnaK. The apparent inability of Hsp90 to bindest in Hsp90 as a target for antitumor drugs. One such adenine nucleotides was also demonstrated by the fail-compound, geldanamycin, has been shown to interact ure to photoaffinity label Hsp90 with 8-azido-ATP, todirectly with Hsp90 (Whitesell and Cook, 1996) and bind Hsp90 to an ATP–agarose affinity resin, or to ob-appears to promote the degradation of client proteins serve enhancement of the fluorescence of MABA-ADPbefore they are fully activated (Schneider et al., 1996). (N-8-[4-N9-methylanthranylaminobutyl]-8-amino adeno-
sine diphosphate) by Hsp90, all of which occur withHigher eukaryotes have a distinct form of Hsp90 (GRP94/
Cell66

Structure of ADP/ATP–Hsp90 N-Domain Complex67
Hsp70/DnaK. Although the apparent involvement of ATP N-domain (residues 1–220) of the yeast Hsp90 chaper-one, providing definitive evidence for the involvementin p23 binding remains unexplained, it has become
widely accepted that unlike Hsp70/DnaK and Hsp60/ of ATP binding in the molecular mechanism of Hsp90.GroEL, Hsp90 does not bind ATP.
Although the structure of the intact Hsp90 molecule Results and Discussionhas not yet been determined, crystal structures of anamino-terminal domain identified by limited proteolysis Cocrystals of yeast Hsp90 N-domain and ADP, ATP, or
ATPgS were grown under conditions previously de-have been determined for yeast (Prodromou et al., 1997)and human (Stebbins et al., 1997) proteins. Consistent scribed for native crystals (Prodromou et al., 1996), but
with the addition of 5 mM nucleotide and 5 mM Mg21.with the high homology among all Hsp90 sequences(69% identity, yeast to human) the tertiary structure of Electron density maps for Hsp90 complexed with Mg21-
ATP, Mg21-ADP, and Mg21-ATPgS were obtained at 1.8,these two domains is extremely similar, consisting of ahighly twisted eight-stranded b sheet covered on one 2.0, and 2.5 A resolution, respectively.face by a helices. The quaternary structures of the hu-man and yeast N-domains observed in the crystals are, Location of the ATP/ADP-Binding Site
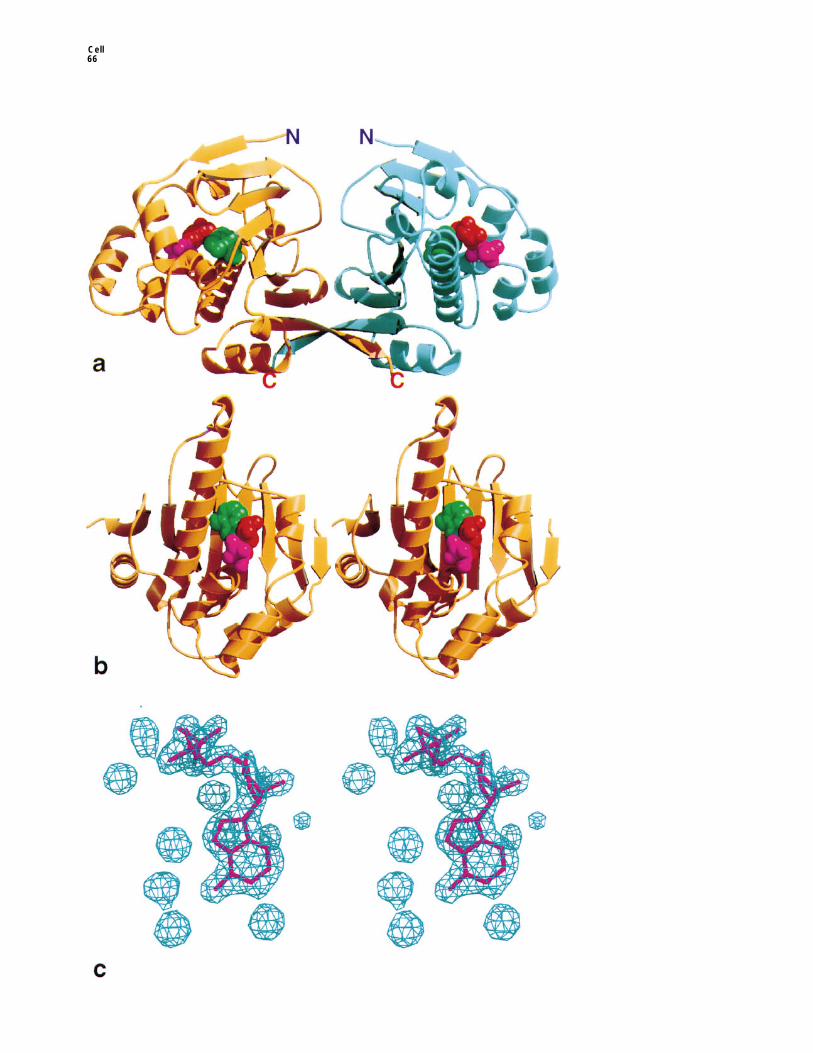
Difference Fourier maps showed clear positive featureshowever, quite different. The yeast N-domain crystal-lizes as a dimer in which the C-terminal b strands of corresponding to the bound nucleotides, lying in a deep
pocket on the helical face of the N-domain (Figures 1athe sheets in each monomer make an antiparallel inter-action, generating a continuously hydrogen-bonded and 1b). This pocket is bounded by the helices from
28–50 and from 85–94 on two sides, and the end of the16-stranded sheet in the dimer. This dimeric sheet foldsback on itself, forming a roughly cylindrical channel be- helix and loop from 117–124 and the loop from 81–85
on the other two sides. The base of the pocket is formedtween the two monomers, whose size and shape sug-gest that it could function as a molecular clamp, capable by residues Ile-77, Asp-79, Val-136, Ser-138, Thr-171,
and Ile-173, whose side chains project up from the bur-of accommodating 8–10 residues of polypeptide chainin an extended conformation (Prodromou et al., 1997). ied face of the b sheet. The electron density for the base,
sugar, and a phosphate groups, which make extensiveIn contrast, thehuman N-domain crystallized as a mono-mer, the b strand that forms the dimer interface being contacts within the pocket, is extremely clear in all the
complexes (Figure 1c). The electron density for the bdisordered, and the potential peptide-binding channelwas not observed (Stebbins et al., 1997). The outside phosphate, which lies higher up in the pocketand makes
fewer contacts, is somewhat weaker, but no significanthelical faces of the monomers in the yeast N-domaindimer are formed by residues, many of which are very electron density is present for the g phosphate in com-
plexes with ATP or with the nonhydrolyzable analog,highly conserved in all Hsp90-family sequences. Atthe center of this helical face, a deep pocket penetrates ATPgS. This suggests that the phosphate has not been
lost by hydrolysis of ATP over the time scale of theto the surface of the buried b sheet and is the bindingsite for the antitumor agent geldanamycin in a complex crystallization experiment but is truly disordered in the
crystals. Analysis of the binding of ATP and ADP towith the human N-domain (Stebbins et al., 1997). On thebasis of this complex, it has been postulated that this the Hsp90 N-domain in solution by isothermal titration
calorimetry indicates dissociation constants of 132 6pocket is a binding site for segments of polypeptidechain from incompletely folded client proteins. Geldana- 47 and 29 6 3 mM, respectively, with binding stoichio-
metries close to 1 nt per N-domain monomer.mycin is therefore proposed to act as a competitiveinhibitor of client–protein binding.
In light of recent observations that the structure of the Nucleotide–Protein InteractionsThe bound nucleotides make extensive interactions withHsp90 N-domain has a similar topology to an N-terminal
ATP-binding domain of the bacterial type II topoisomer- the protein and bound solvent in the pocket (Figure 2).The adenine base penetrates into the pocket, makingase, DNA gyrase (Dunbrack et al., 1997), the involve-
ment of ADP/ATP in the function of Hsp90 must again only a single direct hydrogen bond to the protein, fromthe exocyclic N6 amino group of the adenine base tobe reexamined. Sequence motifs comprising the ATP-
binding sites within the N-terminal domain of DNA gy- the carboxyl side chain of Asp-79 at the bottom of thepocket. All the other hydrogen-bonding possibilities ofrase, and conserved in other type II topoisomerases,
are also conserved in Hsp90 sequences (Bergerat et al., the adenine base are fulfilled by water molecules boundby protein groups within the pocket. Thus, the second1997), raising the possibility that these sequences might
also constitute an ATP-binding site in Hsp90. Here, we hydrogen bond to adenine N6 is made by a water mole-cule bound by the peptide carbonyl of Leu-34; adeninereport the high resolution crystal structures of specific
complexes of Mg21-ATP and of Mg21-ADP with the N1 is hydrogen bonded to a water molecule bound by
Figure 1. Nucleotide Binding by the Hsp90 N-Terminal Domain
(a) Secondary structure cartoon of the yeast Hsp90 N-domain dimer showing the position of the bound ADP/ATP. The base, sugar, andphosphates of the bound nucleotide are colored green, red, and magenta, respectively.(b) Stereo view of the ADP/ATP-binding site in one monomer.(c) Stereo view of electron density for the bound nucleotide and associated solvent from the Hsp90 N-domain–ATP complex. The electrondensity is from an Fo-Fc Fourier, phased from protein coordinates only, refined against the data at 2.0 A using simulated annealing. Contoursare at 3.0s.
Cell68
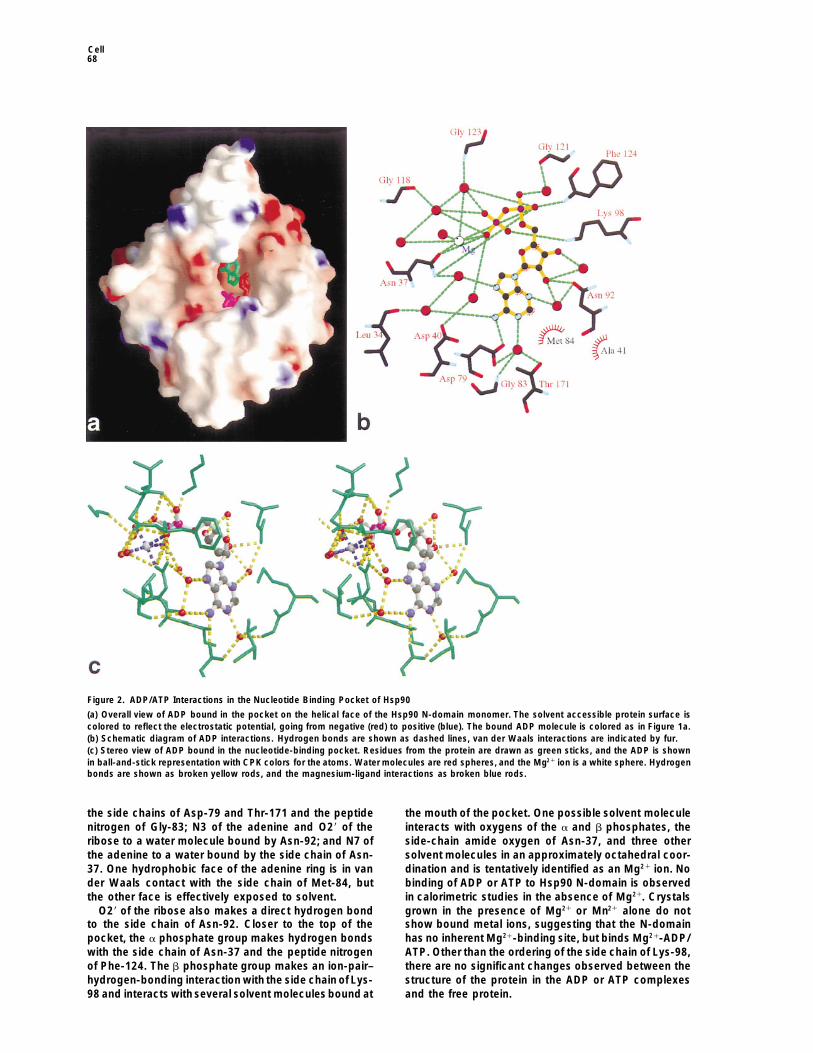
Figure 2. ADP/ATP Interactions in the Nucleotide Binding Pocket of Hsp90
(a) Overall view of ADP bound in the pocket on the helical face of the Hsp90 N-domain monomer. The solvent accessible protein surface iscolored to reflect the electrostatic potential, going from negative (red) to positive (blue). The bound ADP molecule is colored as in Figure 1a.(b) Schematic diagram of ADP interactions. Hydrogen bonds are shown as dashed lines, van der Waals interactions are indicated by fur.(c) Stereo view of ADP bound in the nucleotide-binding pocket. Residues from the protein are drawn as green sticks, and the ADP is shownin ball-and-stick representation with CPK colors for the atoms. Water molecules are red spheres, and the Mg21 ion is a white sphere. Hydrogenbonds are shown as broken yellow rods, and the magnesium-ligand interactions as broken blue rods.
the side chains of Asp-79 and Thr-171 and the peptide the mouth of the pocket. One possible solvent moleculeinteracts with oxygens of the a and b phosphates, thenitrogen of Gly-83; N3 of the adenine and O29 of the
ribose to a water molecule bound by Asn-92; and N7 of side-chain amide oxygen of Asn-37, and three othersolvent molecules in an approximately octahedral coor-the adenine to a water bound by the side chain of Asn-
37. One hydrophobic face of the adenine ring is in van dination and is tentatively identified as an Mg21 ion. Nobinding of ADP or ATP to Hsp90 N-domain is observedder Waals contact with the side chain of Met-84, but
the other face is effectively exposed to solvent. in calorimetric studies in the absence of Mg21. Crystalsgrown in the presence of Mg21 or Mn21 alone do notO29 of the ribose also makes a direct hydrogen bond
to the side chain of Asn-92. Closer to the top of the show bound metal ions, suggesting that the N-domainhas no inherent Mg21-binding site, but binds Mg21-ADP/pocket, the a phosphate group makes hydrogen bonds
with the side chain of Asn-37 and the peptide nitrogen ATP. Other than the ordering of the side chain of Lys-98,there are no significant changes observed between theof Phe-124. The b phosphate group makes an ion-pair–
hydrogen-bonding interaction with the side chain of Lys- structure of the protein in the ADP or ATP complexesand the free protein.98 and interacts with several solvent molecules bound at

Structure of ADP/ATP–Hsp90 N-Domain Complex69
Our observation of specific ADP/ATP binding toHsp90 completely contradicts the careful and widelyaccepted biochemical analysis of Jakob et al. (1996),who demonstrated clearly that Hsp90 could not bespecifically photoaffinity labeled with 8-azido-ATP, wasnot retained on C8-ATP-agarose, and did not enhancethe fluorescence of MABA-ADP. In contrast, all thesereagents gave positiveresults with Hsp70/DnaK, leadingto the reasonable conclusion that Hsp90 does not bindadenine nucleotides. We can understand these negativeobservations by consideration of the different confor-mation of the adenine nucleotides bound to the twodifferent chaperones. In an Hsc70–ADP complex (Fla-herty et al., 1994), the bound nucleotide adopts a fullyextended conformation with a 29-endo sugar pucker andthe g torsion angle in the ap conformation (for conforma-tional definitions, see Moodie and Thornton, 1993). Inthis conformation, the 8 position of the adenine is un-hindered, and 8-substituted analogs would have littledifficulty binding to Hsc70. The nucleotide in the Hsp90–ATP/ADP complex, however, has a much more com-pacted conformation, with a 39-endo sugar pucker anda 1sc g torsion angle. In this conformation, the 8 positionis substantially hindered by the 5 carbon and a phos-
Figure 3. Comparison of Bound Nucleotide Conformations in Hsc70phate, so that this conformation cannot be adopted by and Hsp908-substituted ADP/ATP analogs (Figure 3). We suggest Conformations of ADP bound to (a) Hsc70 (Flaherty et al., 1994) andthat the negative results for nucleotide binding by Hsp90 (b) Hsp90. The N1, N6, which make specific contacts in the ADP/obtained by Jakob et al. (1996) result from their use of ATP-binding pocket, are indicated, as is the adenine base C8 atom,
which is unhindered in the Hsc70-bound conformation but hinderedC8 adenine–substituted reagents, which are not able toin the Hsp90-bound conformation.adopt the idiosyncratic conformation required for ATP/
ADP binding to Hsp90.
oxygen of geldanamycin and the buried water boundby Asp-93, Gly-97, and Thr-184 in human Hsp90 corre-Implications for Geldanamycin Binding
The ATP/ADP-binding site we have identified has pre- sponds to the interaction between the adenine N1 andthe buried water bound by Asp-79, Gly-83, and Thr-171viously been shown to be the binding site for the antitu-
mor agent geldanamycin on the N-terminal domain of in yeast Hsp90. Further up the pocket, the hydrogenbond observed between the amide carbonyl of geldana-the human Hsp90 (Stebbins et al., 1997). Geldanamycin
consists of an ansa ring closed by an embedded benzo- mycin and the main-chain nitrogen of Phe-138 in humanHsp90 corresponds to the hydrogen bond between anquinone, with a pendant carbamate group approxi-
mately halfway around the ansa ring. On the basis of oxygen of the a phosphate of ADP/ATP and the main-chain nitrogen of Phe-124 in yeast Hsp90. Similarly, thethe interactions observed between geldanamycin and
Hsp90, Stebbins et al. (1997) suggest that the ansa ring hydrogen bond from the e amino of Lys-112 toa geldana-mycin benzoquinone oxygen corresponds to the hydro-of geldanamycin imitates a pentapeptide in a turn con-
formation and therefore propose that the biological role gen bond/ion pair between Lys-98 and the b phosphateof ADP/ATP. The hydrogen bonds made by the e aminoof the geldanamycin-binding site is in binding segments
of polypeptide chain from incompletely folded client of Lys-58 to methoxy and carbonyl oxygens on geldana-mycin have no direct equivalent in the yeast ADP/ATPproteins. Our results clearly contradict this suggestion
and rather indicate that geldanamycin is acting as an complex but may correspond to interactions betweenthe side chain of Lys-44 in yeast Hsp90 and solventADP/ATP mimetic, specific to the idiosyncratic set of
interactions offered by the nucleotide-binding site of molecules bound to the O29 and O39 oxygens of theribose sugar in ADP/ATP. The recognition that geldana-Hsp90. In support of this idea, we note that almost all
of the polar interactions described between geldana- mycin imitates the binding of ADP/ATP to Hsp90, ratherthan peptides as previously suggested (Stebbins et al.,mycin and human Hsp90 have precise equivalents in
the specific interactions between yeast Hsp90 and 1997), will be of considerable value in the further devel-opment of this and other compounds as antichaperoneADP/ATP (compare Figure 2c of this paper with Figure
6a in Stebbins et al., 1997). Most significantly, at the agents, with potential applications in the treatment ofmany cancers.bottom of the pocket, the direct hydrogen bond ob-
served between the carbamate nitrogen of geldana-mycin and the carboxyl side chain of Asp-93 in human Structural and Functional Similarity of Hsp90
and DNA Gyrase B N-Terminal DomainsHsp90 corresponds to the direct hydrogen bond be-tween the adenine N6 and Asp-79 in yeast Hsp90. In The overall tertiary fold of the yeast and human Hsp90
N-domains has a remarkable and totally unsuspectedaddition, the hydrogen bond between the carbamate

Cell70
similarity to the N-terminal ATP-binding fragment of the strand of the sheet. Conversely, in Hsp90 the C-terminalstrand from 205–220 extends from the body of the pro-bacterial type II topoisomerase, DNA gyrase B protein
(Wigley et al., 1991). This similarity was not initially rec- tein, making a dimeric interaction with a second mole-cule, whereas the equivalent sequence in gyrase B foldsognized by the authors of either the human or yeast
structures but was determined during the CASP2 struc- back to form the C-terminal strand of the sheet.The most significant difference between the struc-ture-prediction competition (Dunbrack et al., 1997), to
which the yeast Hsp90 N-domain was submitted. tures of the Hsp90 and gyrase B N-domains is the con-formation of the polypeptide sequence from residuesOptimal structural alignment of the N-terminal do-
mains of yeast Hsp90 N-domain and the gyrase B using 94–124 in Hsp90 and the corresponding sequence from95–119 in gyrase B. This segment in gyrase B is anthe SSAP algorithm (Orengo and Taylor, 1996) brings
six b strands and five helices in the Hsp90 structure extended loop of irregular conformation, folded downonto the ATP-binding site as a lid, making contact withinto equivalence with an rmsd between 79 common Ca
positions of approximately 4 A and almost superim- the base and phosphates of the bound ATP. In Hsp90,this segment consists of a short a helix, a loop, and aposes the bound nucleotides from the two structures
(Figure 4). This degree of structural homology, taken short 310 helix and is packed against the helix and loopformed by residues 10–27, away from the ADP/ATP-together with a clear functional similarity, would argue
strongly that both the Hsp90 and DNA gyrase adenine binding site. The difference in orientation of this other-wise topologically equivalent segment corresponds tonucleotide-binding domains are evolved from a com-
mon ancestor. However, even when aligned on the basis a hinge motion, pivoting at glycines 100 and 118 inHsp90 and at glycines 101 and 113 in gyrase B. Thisof this structural equivalence (Figure 5), the amino acid
sequences of the two proteins only have around 10% segment is conformationally flexible in gyrase B and isdisplaced from its closed conformation over the ATP-identity, suggesting that they diverged early in evolution.
While the central strands of the b sheet are very similar binding pocket in complexes with coumarin and cyclo-thialidine antibiotics and becomes disordered (Lewis etin both Hsp90 and gyrase B, the arrangement of the
terminal strands is different. In the gyrase B N-domain, al., 1996). While this segment is orientated away from theADP/ATP-binding site in both yeast and human Hsp90the amino-terminal sequence from 2–15 is detached
from the body of the protein and participates in a dimer structures, the local structure around the pivot residueGly-114 in human Hsp90 (Gly-100 in yeast) displays dif-interaction with a second molecule, whereas the equiv-
alent sequence in Hsp90 forms the amino-terminal ferent conformations in different crystal forms, sug-gesting that the mobility seen in gyrase might also bepresent in Hsp90. If this is the case, this segment inHsp90 might also be able to act as a lid closing overthe ADP/ATP-bindingsite and interacting with the nucle-otide. In the crystals of yeast Hsp90, residues in thissegment are involved in crystal contacts that would sig-nificantly stabilize the open conformation for the lid,despite the presence of nucleotides. From the presentdata, the possibility of a dynamic variation of the Hsp90lid conformation as a result of nucleotide binding isspeculative. Indeed, the extensive hydrophobic inter-face between the lid segment and the helix and loopfrom residues 10–27 in Hsp90 suggests that this openconformation is rather stable. The difference in confor-mation between the lids in gyrase B and Hsp90 mayactually be the result of selection of a different staticconformation in the evolution of Hsp90.
Consistent with the structural homology betweenHsp90 and DNA gyrase N-terminal domains, the confor-mation of ADP/ATP in the binding sites of Hsp90 andDNA gyrase is remarkably similar, and many of the pro-tein residues interacting with the nucleotides are con-served between the two proteins, constituting distinctsequence motifs characteristic of type II topoisomer-ases and Hsp90s (Bergerat et al., 1997). In particular,Asp-79, Gly-83, and Thr-171, which provide the directand solvent-linked interactions with the N1 and N6
Figure 4. Comparison of the ATP-Binding Domains of DNA Gyrase atoms of adenine in Hsp90, correspond to Asp-73,B and Hsp90 Gly-75, and Thr-165, which perform the same function(a and b) Side-by-side comparison of the backbone folds of Hsp90 in gyrase B. A magnesium ion is also present in the(right) and DNA gyrase B (left) N-terminal domains. The amino-termi-
Hsp90-ADP/ATP-binding site in essentially the samenal strand in gyrase B and the C-terminal strand in Hsp90, whichposition in gyrase B. This ion contacts the a and b phos-participate in dimer formation, are highlighted in cyan and red, re-phates in the ADP and ATP complexes and the side-spectively. The lidsegment is highlightedin magenta. Bound nucleo-
tides are shown as green CPK models. chain amide oxygen of an asparagine (37 in Hsp90, 46

Structure of ADP/ATP–Hsp90 N-Domain Complex71
Figure 5. Structural Alignment of the AminoAcid Sequences of Hsp90 and DNA Gyrase B
Alignment of the amino acid sequences of E.coli DNA gyrase B (SwissProt: GYRB_ECOLI)and S. cerevisiae Hsp90 (SwissProt:HS82_YEAST) N-domains, based on thealignment of their secondary structures bythe SSAP algorithm (Orengo and Taylor,1996). Helices are shown as cylinders, with a
helices colored red, and helices with a pri-marily 310 conformation colored magenta. b
strands are shown as green arrows. Se-quence identities are indicated by verticalbars between the sequences, and the threemotifs identified as common to type II topo-isomerases andHsp90s (Bergerat et al., 1997)are highlighted in red.
in gyrase B) conserved in both protein families (Bergerat Hsp90 is indeed variable, then a closed conformationsimilar to that in DNAgyrase might be favored by interac-et al., 1997). There are some small differences between
the phosphate interactions made by Hsp90 and gyrase tion with the g phosphate of a bound ATP. This wouldunmask a potential protein-binding surface formed byB. For example, the b phosphate forms an ion-pair–
hydrogen-bonding interaction with the side chain of the residues buried by the lid in its open conformation.Interestingly, temperature-sensitive mutants of yeastLys-98, which is conserved in Hsp90s but is Ala-100 in
gyrase. The major difference between the well-ordered Hsp90 (Nathan and Lindquist, 1995) that cannot be sim-ply attributed to destabilization of the protein core org phosphate in gyrase B and the disordered g phosphate
in Hsp90 may result from the open-lid conformation in disruption of the nucleotide-binding site map in this loop(Thr-22→Ile) or at the hinge of the lid (Thr-101→Ile).Hsp90, compared with the closed-lid conformation
found in gyrase B. However, interaction with the g phos- In both of the two well-characterized ATP-dependantchaperone systems, Hsp70/Hsc70/DnaK and Hsp60/phate comingfrom residues in the following domain may
also contribute to the ordering of this group in gyrase B. GroEL, binding and hydrolysis of ATP, and release ofADP, are used to drive conformational changes thatcycle the chaperones through high affinity and low affin-A Role for ADP/ATP Binding in the
Mechanism of Hsp90 ity states for incompletely folded protein substrates(Roseman et al., 1996; Zhu et al., 1996). Given that weThe results we present here unequivocally demonstrate
the existence of a specific ADP/ATP-binding site con- have unequivocally demonstrated the existence of anATP-binding site in Hsp90, it is reasonable to speculateserved throughout eukaryotic and bacterial Hsp90s,
necessitating a substantial reappraisal of much of the whether Hsp90 operates by a similar mechanism. TheATP affinity of the Hsp90 N-domain (≈132 mM) is suffi-current data relating to the biochemistry of Hsp90. The
ADP/ATP-binding site we have identified coincides ciently high as to guarantee that the ATP-binding sitewill be fully occupied at cellular ATP concentrations.structurally with the binding site for geldanamycin iden-
tified by Stebbins et al. (1996) and therefore clearly cor- Conversely, the affinity for ADP (29 mM), although higherthan for ATP, is probably insufficient to saturate theresponds to the ADP/ATP-binding site postulated by
Toft and his colleagues (Johnson and Toft, 1994, 1995), binding site with ADP at cellular concentrations. At leastin terms of nucleotide affinity, therefore, Hsp90 is setwhich appears to regulate the binding of the maturation
factor p23 to Hsp90–client–immunophilin complexes. up to go through an ATP-binding–ATP-hydrolysis–ADP-release cycle comparable to those of Hsp60/GroEL orBinding of p23 to Hsp90 is promoted by ATP or ATPgS
but inhibited by ADP (Sullivan et al., 1997) and thus Hsp70/DnaK.The binding of ATP by Hsp90 has been clearly demon-appears to be dependant on the presence of the g
phosphate. In the Hsp90–ATP complex, the disordered strated here, but the existence of an inherent ATPaseactivity remains unresolved. The ATP-binding site ofg phosphate will be exposed at the surface of the
N-domain and could either interact with p23 directly, Hsp90 is very similar to that of the proven ATPase DNAgyrase B, and the catalytic glutamate (Glu-42) responsi-forming part of a p23-binding surface on the N-domain,
or could interact with other parts of the Hsp90 molecule ble for the ATPase activity of gyrase B (Jackson andMaxwell, 1993) is conserved in the N-domain of Hsp90to stabilize a conformation that favors p23 binding else-
where. The nucleotide pocket lies near the middle of an (Glu-33), suggesting that Hsp90 is at least equipped foran ATPase activity with essentially the same catalyticelongated channel that traverses the helical face of the
N-domain, which might provide part of an extended mechanism as DNA gyrase B.In isolation, the 24 kDa N-domain of gyrase B willbinding site for p23. If the conformation of the lid in

Cell72
neither bind nor hydrolyze ATP (Gilbert and Maxwell, (Nieland et al., 1996). The size of the binding site offeredby the Hsp90 N-domain clamp conforms to the predomi-1994), and productive binding of ATP by DNA gyrase B
is dependent on interactions with the g phosphate from nant length of antigenic peptides presented by MHC-I,and the loading process appears to be ATP dependentresidues in the following domain, which may provide a
means for coupling ATP hydrolysis to changes in the (Li and Srivastava, 1993). The high degree of conserva-tion of the N-domain clamp between Hsp90s of therelative juxtaposition of these domains (Wigley et al.,
1991). Although the isolated N-domain of Hsp90 does cytoplasm and endoplasmic reticulum, suggest that thecytoplasmic Hsp90 might have an analogous role inbind ATP weakly, the g phosphate is disordered, and
an ATPase activity in the intact Hsp90 may also depend transporting antigenic peptides from the proteosome tothe cytoplasmic face of the TAP transporter.on interactions tostabilize the conformation of the disor-
dered g phosphate. As in DNA gyrase B, these interac- The data we present here finally resolves the contro-versy of ATP involvement in the function of Hsp90 andtions might be provided by residues in other domains,
giving a mechanism for ATPase-coupled conformational correctly defines the action of the antitumor agent gel-danamycin as an Hsp90-specific ATP mimetic. The pos-changes, or might be provided by a separate protein,
such as p23. sibility of ATPase activity inherent in Hsp90 is still notConformational changes of Hsp90 on addition of ATP
have been reported (Csermely et al., 1993), but the na-ture of these is unknown. The molecular-clamp structureformed by the yeast Hsp90 N-domain dimer (Prodromouet al., 1997) displays a conformational flexibility betweena closed form in which the loops at residues 160–168from each monomer are in contact and an open formin which they are separated by z8 A (Figure 6). Thestructure of the Hsp90 N-domain dimer is such that theC-terminal strands forming the dimer interface swapover topologically. This gives rise to the possibility thatregions of the structure beyond the C-terminus of theN-domain from one monomer could interact with theATP-binding site of the N-domain of the other monomerin the intact Hsp90 dimer. Such interactions either director mediated by p23 would be very likely to influence therelative juxtaposition of the N-domain with respect toother parts of Hsp90 and consequently affect the con-formation of the clamp (Figure 6c). Whether ATP bindingwould serve to open the clamp or close it can only bespeculated upon at this stage. The situation is furthercomplicated by the presence of two equivalent ATP-binding sites in the dimer, allowing for ATP2-, ADP2-, orADP1ATP-loaded states, with the possibility of positiveor negative cooperativity between them.
At present, we do not know what function this clampserves in the molecular mechanism of Hsp90. The chan-nel defined by the clamp in the closed conformation(Prodromou et al., 1997) is of the right shape and sizeto accommodate a peptide chain in an extended confor-mation, making contact with 8–10 amino acids. Theclamp might provide the common binding site sug-gested for the various accessory factors, such as p60/Hop/Sti1, p50/CDC37, and immunophilins, which needto be exchanged at various stages of Hsp90-dependantprotein folding and activation. Alternatively, the clampmight provide a site for binding parts of client proteinsduring folding, i.e., binding and releasing segments of Figure 6. A Model for the ATP-Dependent Conformational Changespolypeptide chain in response toATP binding and hydro- in the Molecular Clamp of the Hsp90 N-Terminal Domainlysis, in an analogous manner to the Hsp70 chaperone. (a and b) Conformational flexibility of the N-domain dimer, in the (a)
The clamp might also be involved in an emerging role closed and (b) open conformations (Prodromou et al., 1997). Thebound nucleotides observed in the closed form of the dimer isfor the endoplasmic reticulum Hsp90, GRP94/GP96, inmodeled into the corresponding position on the open conformation.loading antigenic peptides, imported into the ER lumen(c) Cartoon of a possible model for ATP-switched opening and clos-by the TAP transporter, onto MHC Class I moleculesing of the N-domain molecular clamp. Binding of ATP to the
(Li and Srivastava, 1993). Thus, immunodominant viral N-domains recruits p23, which interacts with the bound ATP and apeptides eluted from MHC-I on the surface of cells in- site in the C-terminal region of Hsp90, promoting a conformationalfected with vesicular stomatitis virus, are also elutable change that allows the release of a bound protein or peptide. Hydro-
lysis of the ATP releases bound p23, and the clamp closes.from the GRP94/GP96 from the ER of the same cells

Structure of ADP/ATP–Hsp90 N-Domain Complex73
the limits of detection of ITC and consequently have large apparentproven but appears highly likely from the structural data,errors in measurement.although an ADP/ATP mechanism that functions by nu-
cleotide exchange but not hydrolysis cannot be ruledGraphical Representationsout. Finally, the conformationaly flexible molecularFigures 1a, 1b, 2c, 3, 4, 6a, and 6b were produced using Molscript
clamp previously identified in the structure of the (Kraulis, 1991) and Raster3D (Merrit and Murphy, 1994). Figure 1cyeast Hsp90 N-domain would appear to offer an ATP- was produced using Raster3D and Robert Esnouf’s adaptation ofswitchable binding site for accessory proteins or for Molscript (Bobscript), 2a by GRASP (Nicholls et al., 1993), and 2b
by Ligplot (Wallace et al., 1995).client proteins and may also play a role in the intracellulartrafficking of antigenic peptides.
Acknowledgments
Experimental ProceduresWe are very grateful to Dale Wigley for making the undepositedcoordinates of DNA gyrase B available to us and to Dietlind GerloffCrystal Growth and Data Collectionand Fred Cohen for drawing our attention to the results of CASP2.Tetragonal crystals of Hsp90 N-domain complexes with ATP,We thank Christine Orengo for assistance with structure alignment,ATPgS, and ADP were grown by vapor diffusion in hanging dropsRoman Laskowski for much assistance with surface analysis andunder very similar conditions to native crystals (Prodromou et al.,generation of figures, and Tony Maxwell for very useful discussion.1996) but with the addition of nucleotides (5 mM) and MgCl2 (5 mM)We are grateful to the Daresbury Laboratory, Warrington, U.K., for
to the mother liquor. Crystalswere stabilized for freezing ina solutionaccess to the Synchrotron Radiation Source and to the Ludwig
containing 30% glycerol, 90 mM ammonium sulphate, 45 mM so-Institute for Cancer Research for provision of X-ray diffraction facili-
dium succinate (pH 5) and 13.5% polyethylene glycol methyl esterties. J. E. L. is a Wellcome Trust Career Development Fellow. This
550. Diffraction data for Hsp90 cocrystals with ATP and ADP werework was supported by a Wellcome Trust project grant to P. W. P.
collected from crystals frozen at 110 K on Station 9.5 (l 5 0.92 A)and L. H. P.
at the SRS, CLRC Daresbury Laboratory, Warrington, UK. The ATPset consisted of 74,267 reflections collected to 2.0 A (21,438 unique
Received May 21, 1997; revised June 9, 1997.reflections, 99.9% complete, Rmerge 5 11.6% with I/sI 5 3.0 in thefinal shell). The ADP crystals diffracted further, giving 95,477 reflec-
Referencestions to 1.84 A (27,155 unique, 99.0% complete, Rmerge 5 9.3 withI/sI 5 3.2 in the final shell). The refined cell of a 5 73.91 A, c 5
Aligue, R., Akhavannik, A., and Russell, P.A. (1994). A role for Hsp90110.95 A with spacegroup P4322 was the same for both data setsin cell-cycle control—Wee1 tyrosine kinase activity requires interac-and close to that observed for tetragonal crystals of the proteintion with Hsp90. EMBO J. 13, 6099–6106alone. A third dataset from a cocrystal with ATPgS was collected
locally on a Rigaku/MAR system with CuKa radiation to 2.5 A (T 5 Bergerat, A., de Massy, B., Gadelle, D., Varoutas, P.-C., Nicolas, A.,110 K, 11,273 unique, 99.9% complete, Rmerge 5 6.3 with I/sI 5 2.6 and Forterre, P. (1997). An atypical topoisomerase II from archaeain the final shell). with implications for meotic recombination. Nature 386, 414–417.
Borkovich, K.A., Farrelly, F.W., Finkelstein, D.B., Taulien, J., andStructure Refinement Lindquist, S. (1989). Hsp82 is an essential protein that is requiredThe ATP and ADP data sets were subject to the same refinement in higher concentrations for growth of cells at higher temperatures.protocol, consisting of X-PLOR (Brunger, 1992) rigid-body refine- Mol. Cell. Biol. 9, 3919–3930.ment of the tetragonalcrystal form of the apo yeast Hsp90 N-domain Brunger, A. (1992). X-PLOR Version 3.1. A System for X-Ray Crystal-structure (Prodromou et al., 1997; Brookhaven Protein Databank lography and NMR, (New Haven, Connecticut: Yale Universityaccession no. 1AH6) in the new cell, followed by simulated anneal- Press).ing-refinement from 40008C with 258C step cooling. The ATP or ADP
CCP4 (1994). Collaborative computational project No. 4. Acta Cryst.was placed in the clear difference density, and the complex wasD50, 760–763.subjected to a further 100 cycles of positional refinement. WatersChen, C.F., Chen, Y.M., Dai, K., Chen, P.L., Riley, D.J., and Lee, W.H.were added automatically using 2 3 10 cycles of REFMAC (CCP4,(1996). A new member of the Hsp90 family of molecular chaperones1994) and ARP (Lamzin and Wilson, 1993) (230 for ATP, 360 forinteracts with the retinoblastoma protein during mitosis and afterADP). Final R-factors from REFMAC were: ATP, Rw 5 18.3%, Rfree 5heat-shock. Mol. Cell. Biol. 16, 4691–4699.24.3%; ADP, Rw 5 16.4%, Rfree 5 23.1%. The ATPgS data was refined
using X-PLOR only. All electron density map interpretation and Csermely, P., and Kahn, C.R. (1991). The 90 kDa heat-shock proteinmodel building was performed with O (Jones et al., 1991). (Hsp-90) possesses an ATP binding-site and autophosphorylation
activity. J. Biol. Chem. 266, 4943–4950.Isothermal Titration Calorimetry Csermely, P., Kajtar, J., Hollosi, M., Jalsovszky, G., Holly, S., Kahn,Isothermal titration calorimetry (ITC) gives a complete thermody- C.R., Gergely, P., Jr., Soti, C., Mihaly, K., and Somogy, J. (1993).namic characterization of an interaction based on the equation ATP induces a conformational change in the 90-kDa heat shock2RT.ln KB 5 DG8 5 DH8 2 TDS8, where R is the gas constant, T is protein (hsp90). J. Biol. Chem. 268, 1901–1907.the absolute temperature, and DG8, DH8, and DS8 are the standard Cutforth, T., and Rubin, G. (1994). Mutations in Hsp83 and CDC37free-energy, enthalpy, and entropy changes on going from unbound impair signaling by the Sevenless receptor tyrosine kinase in Dro-to bound states, respectively. sophila. Cell 77, 1027–1036.
The titration experiments were performed using the MSC systemCzar, M.J., Owens-Grillo, J.K., Dittmar, K.D., Hutchinson, K.A., Za-(MicroCal Inc., MA) as describedelsewhere (Ladbury and Chowdhry,charek, A.M., Leach, K.L., Deibel, M.R., Jr., and Pratt, W.B. (1994).1996). All experiments involved injecting 16 aliquots of 15 ml of 1Characterisation of the protein-protein interactions determining themM ATP or ADP into 1.3 ml of Hsp90 N-domain at 100 mM at 258C.heat shock protein (hsp90-hsp70-hsp56) heterocomplex. J. Biol.All experiments were carried out in 20 mM Tris (pH 7.4) in the pres-Chem. 269, 11155–11161.ence or absence of 5 mM MgCl2. The resulting data were fit asDai, K., Kobayashi, R., and Beach, D. (1996). Physical interaction ofdescribed elsewhere (Wiseman et al., 1989; Ladbury and Chowdhry,mammalian CDC37 with CDK4. J. Biol. Chem. 271, 22030–22034.1996) after subtracting the heats of dilution. Heats of dilution were
determined in separate experiments from addition of ATP or ADP Dunbrack, R.L., Jr., Gerloff, D.L., Bower, M., Chen, X., Lichtarge, O.,and Cohen, F.E. (1997). Meeting review: the second meeting on theinto buffer and buffer into protein. Titration data were fit using a
nonlinear least-squares curve-fitting algorithm with three floating critical assessment of techniques for protein structure prediction(CASP2), Asilomar, California, December 13–16, 1966. Folding andvariables: stoichiometry, binding constant (KB 5 1/KD), and change
of enthalpy of interaction (DH8). The data for ATP binding are close to Design 1, R27–R42.

Cell74
Flaherty, K.M., Wilbanks, S.M., De Luca-Flaherty, C., and McKay, possess ATPase activity and bind heat-shock transcription factorsD.B. (1994). Structural basisof the 70kDa heat shock cognate protein and peptidyl prolyly isomerases. J. Biol. Chem. 268, 1479–1487.ATP hydrolytic activity. II. Structure of the active site with ATP or Nathan, D.F., and Lindquist, S. (1995). Mutational analysis of Hsp90ADP boundto wild-type andmutant ATPase fragment. J. Biol. Chem. function: interactions with a steroid receptor and a protein kinase.269, 12899–12907. Mol. Cell. Biol. 15, 3917–3925.Flynn, G.C., Chapell, T.G., and Rothman, J.E. (1991). Peptide binding Nicholls, A., Bharadwaj, R., and Honig, B. (1993) GRASP: graphicaland release by proteins implicated as catalysts of protein assembly. representation and analysis of surface-properties. Biophys. J. 64,Science 245, 385–390. A166.Freeman, B.C., and Morimoto, R.I. (1996). The human cytosolic mo- Nieland, T.J.F., Tan, M.C.A.A., Monnee-van Muijen, M., Koning, F.,lecular chaperones hsp90, hsp70 (hsc70) and hdj-1 have distinct Kruisbeek, A.M., and van Bleek, G.M. (1996). Isolation of an immuno-roles in recognition of a non-native protein and protein refolding. dominant viral peptide that is endogenously bound to the stressEMBO J. 15, 2969–2979. protein GP96/GRP94. Proc. Natl. Acad. Sci. USA 93, 6135–6139.Gilbert, E.J., and Maxwell, A. (1994). The 24kDa N-terminal subdo- Opperman, H., Levinson, W., and Bishop, J.M. (1981). A cellularmain of the DNA gyrase B protein binds coumarin drugs. Mol. Micro- protein that associates with the transforming protein of Rous Sar-biol. 12, 365–373. coma Virus is also a heat-shock protein. Proc. Natl. Acad. Sci. USAHohfeld, J., Minami, Y., and Hartl, F.-U. (1995). Hip, a novel cochap- 78, 1067–1071.erone involved in the eukaryotic Hsc70/Hsp40 reaction cycle. Cell Orengo, C.A., and Taylor, W.R. (1996). SSAP—sequential structure83, 589–598. alignment program for protein-structure comparison. Meth. Enzy-Jackson, A.P., and Maxwell A. (1993). Identifying the catalytic resi- mol. 266, 617–635.due of the ATPase reaction of DNA gyrase. Proc. Natl. Acad. Sci. Owens-Grillo, J.K., Czar, M.J., Hutchinson, K.A., Hoffman, K., Per-USA 90, 11232–11236. dew, G.H., and Pratt, W.B. (1996). A model of protein targettingJakob, U., Scheibel, T., Bose, S., Reinstein, J., and Buchner, J. mediated by immunophilins and other proteins that bind to hsp90via(1996). Assessment of the ATP binding properties of Hsp90. J. Biol. tetratricopeptide repeat domains. J. Biol. Chem. 271, 13468–13475.Chem. 271, 10035–10041.
Prodromou, C., Piper, P.W., and Pearl, L.H. (1996). Expression andJoab, I., Radanyi, C., Renoir, M.,Buchou, T., Catelli, M.-G., Binart,N., crystallisation of the yeast Hsp82 chaperone, and preliminary X-rayand Mester, J. (1984). Common non-hormone binding component in diffraction studies of the amino-terminal domain. Prot. Struct. Funct.non-transformed chick oviduct receptors of four natural steroids. Genet. 25, 517–522.Nature 308, 850–853.
Prodromou, C., Roe, S.M., Piper, P.W., and Pearl, L.H. (1997). AJohnson, J.L., and Toft, D.O. (1994). A novel chaperone complex molecular clamp in the crystal structure of the N-terminal domainfor steroid-receptors involving heat-shock proteins, immunophilins, of the yeast Hsp90 chaperone. Nature Struct. Biol. 4, 477–482.and p23. J. Biol. Chem. 269, 24989–24993.
Roseman, A.M., Chen, S., White, H., Braig, K., and Saibil, H.R. (1996).Johnson, J.L., and Toft, D.O. (1995). Binding of p23 and Hsp90 The chaperonin ATPase cycle: mechanism of allosteric switchingduring assembly with the progesterone-receptor. Mol. Endocrinol. and movements of substrate-binding domains in GroEL. Cell 87,9, 670–678. 241–251.Jones, T.A., Zou, J.-Y., Cowan, S.W., and Kjeldgaard, M. (1991). Schneider, C., Sepp-Lorenzino, L., Nimmesgern, E., Ouerfelli, O.,Improved methods for building protein models in electron density Danishefsky, S., Rosen, N., and Hartl, F.U. (1996). Pharmacologicmaps and the location of errors in these models. Acta Cryst. A47, shifting of a balance between protein folding and degradation medi-110–119. ated by Hsp90. Proc. Natl. Acad. Sci. USA 93, 14536–14541.Kimura, Y., Yahara, I., and Lindquist, S. (1993). The role of the protein Sepehrnia, B., Paz, I.B., Dasgupta, G., and Momand, J. (1996). Heat-chaperone Ydj1 in establishing Hsp90 mediated signal transduction shock protein 84 forms a complex with mutant p53 protein predomi-pathways. Science 268, 1362–1365. nantly within a cytoplasmic compartment of the cell. J. Biol. Chem.Kraulis, P.J. (1991). MOLSCRIPT—a program to produce both de- 271, 15084–15090.tailed and schematic plots of protein structures. J. Appl. Cryst. 24,
Smith, D.F., and Toft, D.O. (1993). Steroid receptors andtheir associ-946–950.
ated proteins. Mol. Endocrinol. 7, 4–11.Ladbury, J.E., and Chowdhry, B.Z. (1996). Sensing the heat: the
Smith, D.F., Sullivan, W.P., Marion, T.N., Zaitsu, K., Madden, B.,application of isothermal titration calorimetry to thermodynamicMcCormick, D.J., and Toft, D.O. (1993). Identification of a 60kDastudies of biomolecular interactions. Chem. Biol. 3, 791–801.stress related protein, p60, which interacts with hsp90 and hsp70.
Lamzin, V.A., and Wilson, K.S. (1993). Automated refinement of pro- Mol. Cell. Biol. 13, 869–876.teins. Acta Cryst. D49, 129–147.
Stancato, L.F., Chow, Y.-H., Hutchinson, K.A., Perdew, G.H., Jove,Lewis, R.J., Singh, O.M.P., Smith, C.V., Skarzynski, T., Maxwell, A., R., and Pratt, W.B. (1993). Raf exists in a native heterocomplex withWonacott, A.J., and Wigley, D.B. (1996). The nature of inhibition of Hsp90 and p50 that can be reconstituted in a cell-free system. J.DNA gyrase by the coumarins and the cyclothialidines revealed by Biol. Chem. 268, 21711–21716.X-ray crystallography. EMBO J. 15, 1412–1420.
Stebbins, C.E., Russo, A.A., Schneider, C., Rosen, N., Hartl, F.U., andLi, Z., and Srivastava, P.K. (1993). Tumour rejection antigen GP96/Pavletich, N.P. (1997). Crystal structure of an Hsp90-geldanamycinGRP94 is an ATPase: implication for antigen presentation and pro-complex: targetting of a protein chaperone by an antitumor agent.tein folding. EMBO J. 12, 3143–3151.Cell 89, 239–250.
Melnick, J., Aviel, S., and Argon, Y. (1992). The endoplasmic-reticu-Stepanova, L., Leng, X.H., Parker, S.B., and Harper, J.W. (1996).lum stress protein-GRP94, in addition to BiP associates with unas-Mammalian p50 (CDC37) is a protein kinase targetting subunit ofsembled immunoglobulin-chains. J. Biol. Chem. 267, 21303–21306.Hsp90 that binds and stabilises CDK4. Genes Dev. 10, 1491–1502.
Merrit, E.A., and Murphy, M.E.P. (1994). Raster3D version 2.0—aSullivan, W., Stensgard, B., Caucutt, G., Bartha, B., McMahon, N.,program for photorealistic molecular graphics. Acta Cryst. 50,Alnemri, E.S., Litwack, G., and Toft, D.O. (1997). Nucleotides and869–873.two functional states of Hsp90. J. Biol. Chem. 272, 8007–8012.Moodie, S.L., and Thornton, J.M. (1993). A study into the effects ofWallace, A.C., Laskowski, R.A., and Thornton, J.M. (1995). LIPGLOT:protein binding on nucleotide conformation. Nucleic Acids Res. 21,a program to generate schematic diagrams of protein–ligand inter-1369–1380.actions. Prot. Eng. 8, 127–134.Nadeau, K., Sullivan, M.A., Bradley, M., Engman, D.M., and Walsh,Weich, H., Buchner, J., Zimmermann, R., and Jakob, U. (1992).C.T. (1992). 83-kilodalton heat-shock proteins of Trypanasomes areHSP90 chaperones protein folding in vitro. Nature 358, 169–170.potent peptide-stimulated ATPases. Prot. Sci. 1, 970–979.
Nadeau, K., Das, A., and Walsh, C.T. (1993). Hsp90 chaperonins Weich, H., Buchner, J., Zimmermann, M., Zimmermann, R., and

Structure of ADP/ATP–Hsp90 N-Domain Complex75
Jakob, U. (1993). Hsc70, immunoglobulin heavy-chain binding-pro-tein, and Hsp90 differ in their ability to stimulate transport of precur-sor proteins into mammalian microsomes. J. Biol. Chem. 268, 7414–7421.
Whitesell, L., and Cook, P. (1996). Stable and specific binding of heatshock protein 90 by geldanamycin disrupts glucocorticoid receptorfunction in intact cells. Mol. Endocrinol. 10, 705–712.
Wigley, D.B., Davies, G.J., Dodson, E.J., Maxwell, A., and Dodson,G. (1991). Crystal structure of an N-terminal fragment of the DNAgyrase B protein. Nature 351, 624–629.
Wilhelmson, A., Cuthill, S., Denis, M., Wikstrom, A.-C., Gustafsson,J.-A., and Poellinger, L. (1990). The specific DNA binding activity ofthe dioxin receptor is modulated by the 90 kDa heat-shock protein.EMBO J. 9, 69–76.
Wiseman, T., Williston, S., Brandtas, J.F., and Lin, L.N. (1989). Rapidmeasurement of binding constants and heats of binding using anew titration calorimeter. Anal. Biochem. 179, 131–137.
Zhu, X.T., Zhao, X, Burkholder, W.F., Gragerov, A., Ogata, C.M.,Gottesman, M.E., and Hendrickson, W.A. (1996). Structural analysisof substrate binding by the molecular chaperone DnaK. Science272, 1606–1614.
Brookhaven Protein Databank Accession Number
The Brookhaven Protein Databank accession numbers for coordi-nates of the ADP–Hsp90 and ATP-Hsp90 complexes reported hereare 1AMW and 1AM1, respectively.