Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, May 1989, p. 1483-14900019-9567/89/051483-08$02.00/0Copyright C 1989, American Society for Microbiology
Immunomodulatory Activity of Monophosphoryl Lipid Ain C3H/HeJ and C3H/HeSnJ Mice
JACQUES R. HIERNAUX,' PHILIP W. STASHAK,2 JOHN L. CANTRELL,' JON A. RUDBACH,1AND PHILLIP J. BAKER2*
Ribi ImmunoChem Research, Inc., Hamilton, Montana 59840,1 and National Institute of Allergy and Infectious Diseases,Twinbrook-II Research Facility, 12441 Parklawn Drive, Rockville, Maryland 208522
Received 11 October 1988/Accepted 8 February 1989
Treatment with nontoxic monophosphoryl lipid A (MPL) derived from a polysaccharide-deficient, heptose-less Re mutant of either Salmonella typhimurium or Salmonella minnesota R595 enhanced the immunoglobulinM (IgM) anti-type III pneumococcal polysaccharide (SSS-III) antibody response of C3H/HeSnJ mice. Such anadjuvant effect was not observed in lipopolysaccharide-nonresponder C3H/HeJ mice. Nevertheless, C3H/HeJspleen cells produced a weak mitogenic response to both preparations of MPL in vitro, and C3H/HeJ miceshowed a significant increase in serum IgM levels without an increase in numbers of splenic IgM-secretingplaque-forming cells after in vivo treatment with MPL. A significant increase in serum IgG3 levels wasaccompanied by a transient decrease in serum IgGl levels in C3H/HeSnJ mice given MPL; such non-antigen-specific polyclonal effects were not observed in C3H/HeJ or in athymic nulnu mice. Since the enhancedantibody response to SSS-III has been attributed to the inactivation of suppressor T cells by MPL and sincesuppressor-T-cell activity is demonstrable in both C3H/HeSnJ and C3H/HeJ mice, these findings imply that (i)the suppressor T cells of C3H/HeJ mice are refractory to inactivation by MPL and (ii) some of the polyclonaland mitogenic effects produced in C3H/HeJ mice are due to the direct action of MPL on B lymphocytes.
The endotoxins or lipopolysaccharides (LPSs) isolatedfrom the cell walls of gram-negative bacteria are potentimmunomodulators, mainly because they possess mitogenicactivity for murine B lymphocytes in vitro, polyclonallyactivate murine B cells in vivo, stimulate the production ofinterleukin-1 (IL-1), promote granulopoiesis through therelease of endogenous colony-stimulating factor, and acti-vate macrophages (1, 2, 14, 25). LPS enhances the immuneresponse when injected with an antigen (20, 31) and sup-presses or enhances it when injected before an antigen (9,26). LPS also induces the production of tumor necrosisfactor, which has antitumor activity (10). All of these variousimmunomodulatory activities are elicited by the diphospho-ryl lipid A portion of the LPS molecule (for a review, seereference 24), which is extremely toxic and pyrogenic formost animal species, thereby precluding its use as a thera-peutic agent; however, the removal of a phosphate groupfrom the reducing end of diphosphoryl lipid A yields mono-phosphoryl lipid A MPL (N. Qureshi and K. Takayama, inB. H. Iglewski and V. L. Clark, ed., Molecular Basis ofBacterial Pathogenesis, in press). Since MPL retains all ofthe immunomodulatory properties of LPS and diphosphoryllipid A but is significantly less toxic and pyrogenic (30), itcan safely be used as an adjuvant in vivo.We have shown that MPL increases the antibody response
to type III pneumococcal polysaccharide (SSS-III) by abro-gating the activity of suppressor T (TJ) cells without influ-encing helper-T-cell (TH) and amplifier-T-cell function (5).MPL has been shown to be mitogenic for the spleen cells ofLPS-responsive (LPS') and LPS-defective (LPSd) mice (36)and to increase the antibody response of C3H/HeJ mice tosheep erythrocytes (37). The present study was designed toexamine the effects of treatment with MPL on the antibodyresponse to SSS-III in LPSr C3H/HeSnJ and LPSd C3H/HeJmice.
* Corresponding author.
MATERIALS AND METHODS
Mice. Female LPS' C3H/HeSnJ and LPSd C3H/HeJ mice,6 to 8 weeks of age, were obtained from Jackson Laboratory,Bar Harbor, Maine. Female athymic nude (nulnu) mice andthymus-bearing (nul+) littermate controls, 5 to 6 weeks ofage, were obtained from the Frederick Cancer ResearchCenter, Frederick, Md.
Antigen and immunization procedure. The immunologicalproperties of the preparation of SSS-III used and the methodby which it was prepared have been described (7). Forimmunization, mice were given a single intraperitoneal (i.p.)injection of an optimally immunogenic dose (500 ng) ofSSS-III. The magnitude of the antibody response producedwas determined 5 days after immunization.
Detection of antibody-secreting cells. The numbers ofsplenic antibody-producing plaque-forming cells (PFC) mak-ing antibody specific for SSS-III provided a measure of theantibody response elicited in individual mice 5 days afterimmunization with SSS-III, i.e., at the peak of the antibodyresponse. PFC making antibody of the immunoglobulin M(IgM) class (>90% of all PFC found [6]) were detected by aslide version of the localized hemolysis-in-gel technique,using indicator sheep erythrocytes coated with SSS-III bythe CrCl3 method (8). Corrections were made for numbers ofbackground sheep-erythrocyte-specific PFC found, so thatonly PFC making antibody specific for SSS-III were consid-ered. The values obtained (PFC per spleen), which are lognormally distributed (15), are expressed as the geometricmean (antilog) of the log1o number of PFC per spleen forgroups of similarly treated mice. This provides a reasonablygood measure of the magnitude of the total antibody re-sponse produced since SSS-III-specific PFC are detectedonly in the spleens of immunized mice (21).
Cells secreting non-antigen-specific immunoglobulin of theIgM class were detected by a slide version of the protein Aplaque assay (5, 16). The results are expressed as log1o
1483
Vol. 57, No. 5
on June 28, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
INFECT. IMMUN.
IgM-secreting PFC per spleen (mean + standard error of themean [SEM]) for groups of similarly treated mice.
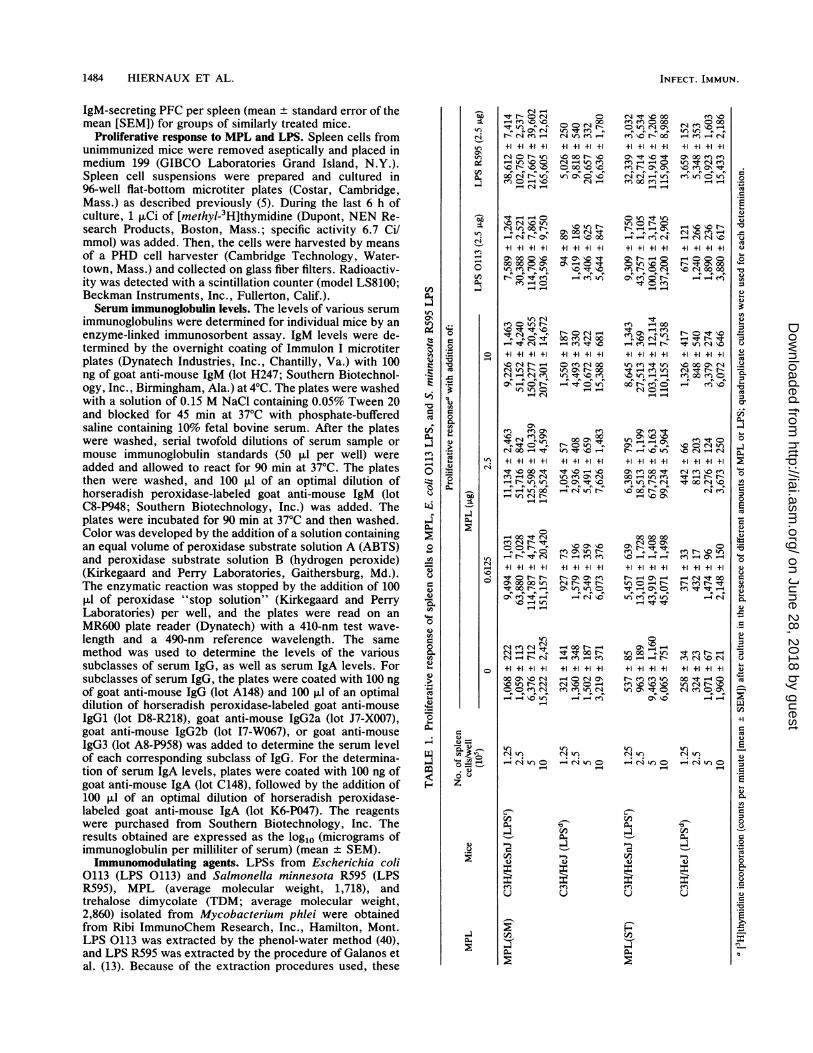
Proliferative response to MPL and LPS. Spleen cells fromunimmunized mice were removed aseptically and placed inmedium 199 (GIBCO Laboratories Grand Island, N.Y.).Spleen cell suspensions were prepared and cultured in96-well flat-bottom microtiter plates (Costar, Cambridge,Mass.) as described previously (5). During the last 6 h ofculture, 1 ,uCi of [methyl-3H]thymidine (Dupont, NEN Re-search Products, Boston, Mass.; specific activity 6.7 Ci/mmol) was added. Then, the cells were harvested by meansof a PHD cell harvester (Cambridge Technology, Water-town, Mass.) and collected on glass fiber filters. Radioactiv-ity was detected with a scintillation counter (model LS8100;Beckman Instruments, Inc., Fullerton, Calif.).Serum immunoglobulin levels. The levels of various serum
immunoglobulins were determined for individual mice by anenzyme-linked immunosorbent assay. IgM levels were de-termined by the overnight coating of Immulon I microtiterplates (Dynatech Industries, Inc., Chantilly, Va.) with 100ng of goat anti-mouse IgM (lot H247; Southern Biotechnol-ogy, Inc., Birmingham, Ala.) at 4°C. The plates were washedwith a solution of 0.15 M NaCl containing 0.05% Tween 20and blocked for 45 min at 37°C with phosphate-bufferedsaline containing 10% fetal bovine serum. After the plateswere washed, serial twofold dilutions of serum sample ormouse immunoglobulin standards (50 pul per well) wereadded and allowed to react for 90 min at 37°C. The platesthen were washed, and 100 ,ul of an optimal dilution ofhorseradish peroxidase-labeled goat anti-mouse IgM (lotC8-P948; Southern Biotechnology, Inc.) was added. Theplates were incubated for 90 min at 37°C and then washed.Color was developed by the addition of a solution containingan equal volume of peroxidase substrate solution A (ABTS)and peroxidase substrate solution B (hydrogen peroxide)(Kirkegaard and Perry Laboratories, Gaithersburg, Md.).The enzymatic reaction was stopped by the addition of 100,ul of peroxidase "stop solution" (Kirkegaard and PerryLaboratories) per well, and the plates were read on anMR600 plate reader (Dynatech) with a 410-nm test wave-length and a 490-nm reference wavelength. The samemethod was used to determine the levels of the varioussubclasses of serum IgG, as well as serum IgA levels. Forsubclasses of serum IgG, the plates were coated with 100 ngof goat anti-mouse IgG (lot A148) and 100 ,u1 of an optimaldilution of horseradish peroxidase-labeled goat anti-mouseIgGl (lot D8-R218), goat anti-mouse IgG2a (lot J7-X007),goat anti-mouse IgG2b (lot I7-W067), or goat anti-mouseIgG3 (lot A8-P958) was added to determine the serum levelof each corresponding subclass of IgG. For the determina-tion of serum IgA levels, plates were coated with 100 ng ofgoat anti-mouse IgA (lot C148), followed by the addition of100 ,ul of an optimal dilution of horseradish peroxidase-labeled goat anti-mouse IgA (lot K6-P047). The reagentswere purchased from Southern Biotechnology, Inc. Theresults obtained are expressed as the log1o (micrograms ofimmunoglobulin per milliliter of serum) (mean + SEM).Immunomodulating agents. LPSs from Escherichia coli
0113 (LPS 0113) and Salmonella minnesota R595 (LPSR595), MPL (average molecular weight, 1,718), andtrehalose dimycolate (TDM; average molecular weight,2,860) isolated from Mycobacterium phlei were obtainedfrom Ribi ImmunoChem Research, Inc., Hamilton, Mont.LPS 0113 was extracted by the phenol-water method (40),and LPS R595 was extracted by the procedure of Galanos etal. (13). Because of the extraction procedures used, these
Cl)
C')
-3
C)
c)
-.2CU
-3
..
0
0co
0
0
r_
0
C)
2
C)
C:co40-506
toy
CA.
0ta-
=12
O
c;C)o =
C)
z
Cl -R N 0 Cl
+l +1 +1 +1
- m O0N r tn
e0 0- t0- C-
C r~00 N-- ClN Oy+l +1 +1 +1
CN 00 ~
00 00 'rNW)^ b
F0 eC) k o
Cl -
+1 +1+1 +1
I", \C 00
- C
n
Cl4
+1 +1 +1 +1
ON 0 00 l)
CCN - ON C
0 r-
o'-N 'r'i
aNenI'lo-
\-4 r- 00T-44ICl N
tn
enCl N
Noo _C
+l +1 +1 +1
^ 0 N N
O~0000l
_ 00 N -
_~ _
0 -4
N Clt00o~ Cl
C F
o Ct 0
to_
0
+l +1 +1 +1'I 00 NmcCl '--4 CrC en000 110 IC
oC '1N000 Clq00 '-4 'IC 00+1 +1 +1 +1
oo cn " 00r- c t £o
+l +l +l +l-en " 0
00 CON) Cl 00
Hto-0C'~
r- 0 tn+1 +1 +1 +1:t 'IC r--
CC0CN 00'r ~.
+l +l +l +l
- -
IRt oo r-
+l +1 +1 +1
r4C') 0q Cl,
Nn c)
N 00 e
Cq'l \0 00)n e 000
en u0 N 00+l +1 +1 +1
cn) N ON
c') 00 en) '-C
0 C) t Cl)N -4 c0+i +1 +1 +1ON 0-V-C0 t 'ICC') rN Co
0tC) en
en') 00"t 't c
+l +1 +1 +1
tncnt-
n
- )-en
Cl - - o'- \OC
0 e 00 It00 l-) C)
10
0000 00N o o
4t_t :t
\C---f
+l +1 +1 +1
en \10 '\4Cl) al NR
l \0 90
C')~0'IC Cl-
+l +1 +1 +10 00 cC)
C WI
+l +l +l +l- t 00N 0000
- -Ct C)Mt r-Nle
+1 +1 +1 +1\C 000N lCl
oo Nf Ncq It r- r
0£o0 eiCl
+l +1 +1 +1
I~ 00 cl \10
al r bC')oN S0Cl
+1 +1 +1 +1r--4 le~00N en N t
_') N
+l +1 +1 +100 " 0oo
CNo
C C' 0
l) t l)
Cl4cl) W) (= ClCl ClCD n
00-
U)
--
00-
C)U
c0
AO
0.
C)C')
en
u
1484 HIERNAUX ET AL.
.3.0CU00'
0s
0-o
0t
3
C)
C.)0
C)
.0
.3
C)
CL
+,
CU
00
C)
C:0_0.
0
C)
on June 28, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

MPL ACTIVITY IN C3H MICE 1485
+
,a -:~ f
I-,
00 -
0nt-
cl I't (
00-
4z4z4z4z
o33 I3
C> 00 Ch_s00 .> .
K
1+ 1+ I+ I+ I+ 1+ 1+ 1+
-.J
C
'0'.0 P'3
LA 0%w-_ p. (-A'IO i 1- LA -.4
O CD CD 0 0 O O
o0 C'i s00
00 '0 0l C> 00
0o000wsS00000> 0 i0 00W
(20% W I.ALA No4.~I+ + I+ I+ I+ I+ I00
Is,)s)-4~
00 00 LA 00 "s ~- '.0I)O -- 'IO Oi ON O1+ 1+1+ 1+1+ 1+1+ 1+
C) C O CD Q Q-(=C
LI) LI)
W Ch4C6ONww
-i U.)_1 o U.)
:-4-
ON LA -4
A A 00 LA
1+ 1+ 1+1+1+ 1+1+ 1+
)00 'I0-. J .A LA-.
'000 -A 00 -
'00 %A '.0'0LA " --j LA
0 )0 0% -4Cw
1+ 1+1+ 1+1+ 1+1+ 1+
Q C) O C) ) C) C) Cb . .: . ;. .- .~ ;-00000000
0010 w1-' U.)* -P~
0% 000 <.i -A
U' I) LA Is1) --4 P. U)~I)0 00 LA 00'--'
1+ 1+1+ 1+
________
CD) 0) z z.b * * * .
I+ I+0 I+) LAI + +I
0-
U)
I."
Qaw0-4
(Q
Cw
C.
(DcIB
preparations of LPS contained minimal amounts of protein.One preparation of MPL [MPL(ST)] was isolated from apolysaccharide-deficient, heptoseless Re mutant of Salmo-nella typhimurium G30/C21, whereas the other MPL prepa-ration [MPL(SM)] was isolated from polysaccharide-defi-cient S. minnesota R595 as described previously (27, 28;Qureshi and Takayama, in press). MPL(ST) is a mixture ofpentaacyl and hexaacyl MPL, whereas MPL(SM) is a mix-ture of pentaacyl, hexaacyl, and heptaacyl MPL (27, 28;Qureshi and Takayama, in press). Lyophilized MPL wasreconstituted to 1 mg/ml in distilled water containing 0.2%triethylamine. It was mixed thoroughly and sonicated brieflyto obtain an opalescent stock solution which was stored at4°C until use. Data on the toxicity and pyrogenicity of MPL,as well as on the parent LPS from which they were derived,have been published (29).
Lyophilized TDM was solubilized in a mixture of chloro-form and methanol (9:1) to a concentration of 10 mg/ml anddried under vacuum. Then, TDM was emulsified in 400 ,ul ofsqualene oil (Sigma Chemical Co., St. Louis, Mo.) andadjusted to a concentration of 500 ,ug/ml in saline containing0.2% Tween 80. This stock solution was stored at 4°C untiluse.
Statistics. Student's t test was used to evaluate the signif-icance of the differences observed. Differences were consid-ered to be significant when probability (P) values <0.05 wereobtained.
r-nii
CD
<D
_.
0
0
2.CDU)
0
0.
U)U_Pzr-u)P.0
CD0.
0.0CD
RESULTS
Mitogenic activities of MPL(ST) and MPL(SM). Previousstudies (36, 38) have shown that both MPL(SM) and a lipidA precursor molecule derived from S. typhimurium aremitogenic for C3H/HeJ spleen cells in vitro. Consequently,we wished to examine the' proliferative response to C3H/HeJspleen cells to MPL(ST). The data in Table 1 show thatMPL(ST) was mitogenic for C3H/HeJ spleen cells under theassay conditions used. They also confirm that MPL(SM) ismitogenic for C3H/HeJ spleen cells; however, both induceda stronger proliferative response with C3H/HeSnJ spleencells [stimulation index of 23.6 and 18.2 at the peak of theresponse for MPL(SM) and MPL(ST), respectively] thanwith C3H/HeJ spleen cells [stimulation index of 7.1 and 3.1at the peak of the response for MPL(SM) and MPL(ST),respectively]. In both experiments, the proliferative' re-sponses of spleen cells to LPS 0113, which is nonmitogenicfor LPSd spleen cells (17), and to LPS R595, which ismitogenic for LPSd spleen cells (36), also were examined andused as controls.
Effect of MPL(ST) on serum immunoglobulin levels. MPL,which is a potent mitogen for B lymphocytes in vitro, is alsoa polyclonal activator of B cells in vivo (5). Since polyclonalB-cell activation causes an increase in the number of IgM-and IgG-secreting B lymphocytes (11), as well as an increasein serum immunoglobulin levels, we examined the serumimmunoglobulin levels of mice treated with MPL. The serumIgM levels of both nulnu and nul+ mice increased signifi-cantly after treatment with MPL(ST) (Table 2; P < 0.05 inboth cases), thereby indicating polyclonal activation inducedby MPL. nulnu mice have very low levels of all subclasses ofIgG in serum; these were not altered (P > 0.05) in responseto MPL(ST), although there was a significant (P < 0.01)increase in the level of serum IgG3 in nul+ mice treated withMPL(ST). Serum IgG2a and IgG2b levels remained un-changed (P > 0.05), whereas serum IgGl levels decreasedsignificantly (P < 0.05) in nul+ mice treated with MPL(ST).
VOL. 57, 1989
P:Z Pz °
LA
0 0
CA)
00
CL
0
ron
Or,
rA0-
0"'C)>3"'C_)_
00o: .
*()0
c,
CD
on June 28, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

1486 HIERNAUX ET AL.
TABLE 3. Serum immunoglobulin levels of C3H/HeSnJ mice treated with MPL(ST)
Treat- Level' in serummenta IgM IgGl IgG2a IgG2b IgG3 IgANaCl 2.608 ± 0.067 (405) 3.202 ± 0.045 (1,592) 3.006 ± 0.068 (1,014) 2.602 ± 0.061 (400) 2.314 ± 0.057 (206) 1.669 ± 0.049 (47)MPL 2.711 ± 0.113 (514) 3.363 ± 0.041 (2,306) 3.131 ± 0.043 (1,352) 2.836 ± 0.065 (686) 2.808 ± 0.076 (643) 1.736 ± 0.085 (54)
a Three-month-old female mice were given (i.p.) either 200 p.l of 0.15 M NaCI or 50 p.g of MPL(ST) 12 days before serum samples were collected for analysis.b Log1o serum immunoglobulin level (mean ± SEM) for five mice; geometric means (antilogs) are in parentheses (micrograms per milliliter).
Similar results were obtained with nulnu and nul+ micetreated with MPL(SM) (data not shown). All serum samplesused in these experiments were collected 3 days aftertreatment with MPL.Treatment with MPL(ST) significantly increased the se-
rum IgM (P < 0.001) and IgG3 (P < 0.01) levels of C3H/HeSnJ mice (Table 2), whereas serum IgG2a, IgG2b, andIgA levels were not changed significantly (P > 0.05); serum
IgGl levels were decreased significantly (P < 0.01) inC3H/HeSnJ mice treated with MPL(ST). Although LPSdC3H/HeJ mice showed a significant increase in serum IgMlevels after treatment with MPL(ST), the levels of otherimmunoglobulin isotypes were not significantly affected.Thus, the increased levels of serum IgM and IgG3 inC3H/HeSnJ mice are indicative of polyclonal B-cell activa-tion induced by MPL(ST). In contrast, there was an increasein the serum IgM levels, but not the serum IgG3 levels, ofC3H/HeJ mice. These differences might have been due to thefact that MPL(ST) was not as mitogenic for C3H/HeJ spleencells as for C3H/HeSnJ spleen cells (Table 1). Since aprevious study (19) showed that in mice treated with LPSR595, there is a rapid increase in serum IgM levels and adelayed increase in serum IgG levels, we also examinedserum immunoglobulin levels of C3H mice 12 days after thetreatment with MPL(ST). We found a significant increase (P< 0.05) in the serum IgGl, IgG2b, and IgG3 levels and nosignificant increase (P > 0.05) in the serum IgM, IgG2a, andIgA levels of C3H/HeSnJ mice treated with MPL (Table 3).Thus, although we detected an initial decrease in the level ofserum IgGl in C3H/HeSnJ mice 3 days after treatment withMPL (Table 2), an increase was noted 12 days after theadministration of MPL. The increased levels of serum IgG2bobserved at day 12 were not noted 3 days after the treatmentwith MPL. C3H/HeJ mice treated with MPL(ST) showed nosignificant differences (P > 0.05) in serum immunoglobulinlevels 12 days after the treatment (data not shown).Adjuvant activity ofMPL in nude mice. MPL(ST) does not
increase the magnitude of the SSS-Ill-specific antibody(PFC) response of nude mice, even though it induces signif-icant polyclonal activation of IgM synthesis (5). Conse-quently, we wished to examine the effects of MPL(SM) on
both the antigen-specific and polyclonal IgM responses ofnude mice. MPL(SM) and MPL(ST) both induced a signifi-cant (P < 0.001) polyclonal IgM response in nulnu and nul+mice, as evidenced by the increased numbers of IgM-secreting PFC detected (Table 4). MPL(ST) had no effect (P> 0.05) on the magnitude of the SSS-III-specific PFC re-
sponse of nulnu mice, as reported previously (5); however,MPL(SM) significantly (P < 0.05) increased the SSS-III-specific PFC response of nulnu mice. Although the basis forthis difference is not known, it might be related to thestronger polyclonal (Table 4) and mitogenic (unpublishedresults) responses elicited by MPL(SM) in nulnu mice.
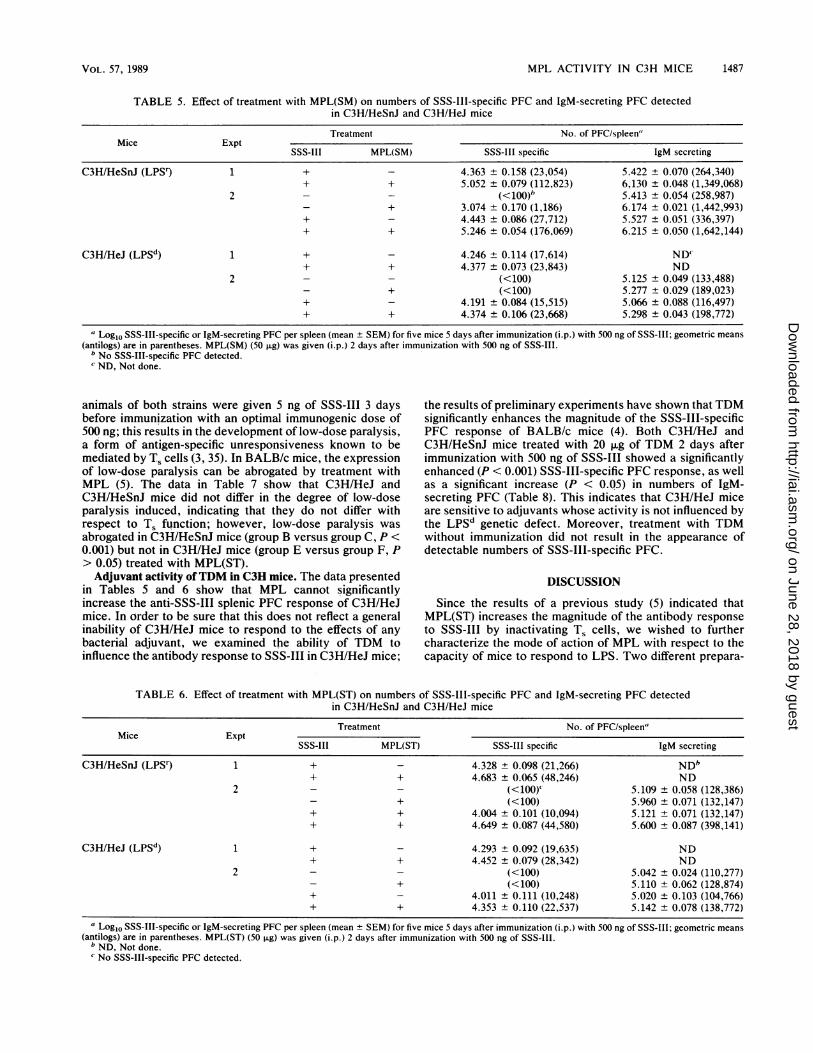
Adjuvant activity of MPL in C3H mice. C3H/HeSnJ andC3H/HeJ mice were immunized (i.p.) with 500 ng of SSS-III,and the numbers of SSS-III-specific, as well as IgM-se-
creting, PFC were determined 5 days later with or withouttreatment with MPL(SM) or MPL(ST). Although treatmentwith both preparations of MPL significantly increased (P <0.001) the magnitude of the polyclonal antibody response ofunimmunized C3H/HeSnJ mice, it did not result in thedetection of SSS-III-specific PFC in unimmunized mice.Both preparations of MPL significantly increased the mag-nitude of the SSS-III-specific and polyclonal (IgM) re-sponses in immunized C3H/HeSnJ mice (Tables 5 and 6; P <0.01). In C3H/HeJ mice, neither type of MPL had a signifi-cant effect (P > 0.05) on the numbers of SSS-III-specific andIgM-secreting PFC detected (Tables 5 and 6); this seems tobe at variance with the data in Table 2 showing an increasein the levels of serum IgM observed for C3H/HeJ micetreated with MPL(ST). This might be due to inherent differ-ences between the two methods used. Even though serumIgM levels are a reflection of the activity of MPL(ST) in vivoin various organs between the time of its injection and thetime of bleeding, the assay for IgM-secreting PFC deter-mines the number of IgM-secreting 13 lymphocytes presentin the spleen at the time of sacrifice. Thus, for C3H/HeJmice, both types of MPL were mitogenic for spleen cells invitro (Table 1) and polyclonally stimulated B lymphocytes invivo in one of the assays used (Table 2); however, they hadno detectable effect on the magnitude of the antibody re-sponse to SSS-III, thereby indicating a dissociation betweenthese phenomena.
Previous studies suggest that MPL(ST) increases themagnitude of the SSS-III-specific antibody response byinactivating T1 cells (5). The failure to augment the antibodyresponse to SSS-III in C3H/HeJ mice, in conjunction withthe absence of appreciable B-cell polyclonal activation byMPL, suggests that both processes may be under commongenetic control and involve the absence of a cell receptorrequired for the binding of MPL. In order to evaluate thedegree of Ts function in C3H/HeJ and C3H/HeSnJ mice,
TABLE 4. Effect of treatment with MPL(SM) or MPL(ST)on numbers of SSS-III-specific PFC and IgM-secreting
cells detected in nulnu and nul+ mice
Treat- No. of PFC":Mlcement" SSS-III specific IgM secreting
nulnu 3.492 ± 0.104 (3,102) 5.116 + 0.041 (130,494)MPL(SM) 3,992 ± 0.076 (9,828) 6.021 ± 0.042 (1,049,385)
3.654 ± 0.071 (4,506) 5.006 ± 0.070 (101,327)MPL(ST) 3.682 ± 0.161 (4,811) 5.699 ± 0.039 (500,035)
nul+ 3.503 ± 0.187 (3,183) 5.020 ± 0.059 (104,339)MPL(SM) 4.514 ± 0.107 (32,685) 5.985 ± 0.081 (966,849)
3.790 ± 0.056 (6,159) 4,923 ± 0.068 (83,800)MPL(ST) 4.053 + 0.087 (11,304) 5.425 ± 0.072 (265,810)
" Mice were given 50 p.g of MPL (i.p.) 2 days after immunization (i.p.) with500 ng of SSS-III.
b Log1o PFC per spleen (mean t SEM) for five mice; geometric means arein parentheses.
INFECT. IMMUN.
on June 28, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
MPL ACTIVITY IN C3H MICE 1487
TABLE 5. Effect of treatment with MPL(SM) on numbers of SSS-III-specific PFC and IgM-secreting PFC detectedin C3H/HeSnJ and C3H/HeJ mice
Treatment No. of PFC/spleen"Mice Expt
SSS-III MPL(SM) SSS-III specific IgM secreting
C3H/HeSnJ (LPSr) 1 + _ 4.363 ± 0.158 (23,054) 5.422 ± 0.070 (264,340)+ + 5.052 ± 0.079 (112,823) 6,130 ± 0.048 (1,349,068)
2 - _ (<100) 5.413 ± 0.054 (258,987)+ 3.074 ± 0.170 (1,186) 6.174 ± 0.021 (1,442,993)
+ - 4.443 + 0.086 (27,712) 5.527 ± 0.051 (336,397)+ + 5.246 ± 0.054 (176,069) 6.215 ± 0.050 (1,642,144)
C3H/HeJ (LPSd) 1 + _ 4.246 ± 0.114 (17,614) ND'+ + 4.377 ± 0.073 (23,843) ND
2 - _ (<100) 5.125 ± 0.049 (133,488)+ (<100) 5.277 ± 0.029 (189,023)
+ - 4.191 ± 0.084 (15,515) 5.066 ± 0.088 (116,497)+ + 4.374 ± 0.106 (23,668) 5.298 ± 0.043 (198,772)
a Log1o SSS-III-specific or IgM-secreting PFC per spleen (mean + SEM) for five mice 5 days after immunization (i.p.) with 500 ng of SSS-I1I; geometric means(antilogs) are in parentheses. MPL(SM) (50 ,ug) was given (i.p.) 2 days after immunization with 500 ng of SSS-III.
b No SSS-III-specific PFC detected.c ND, Not done.
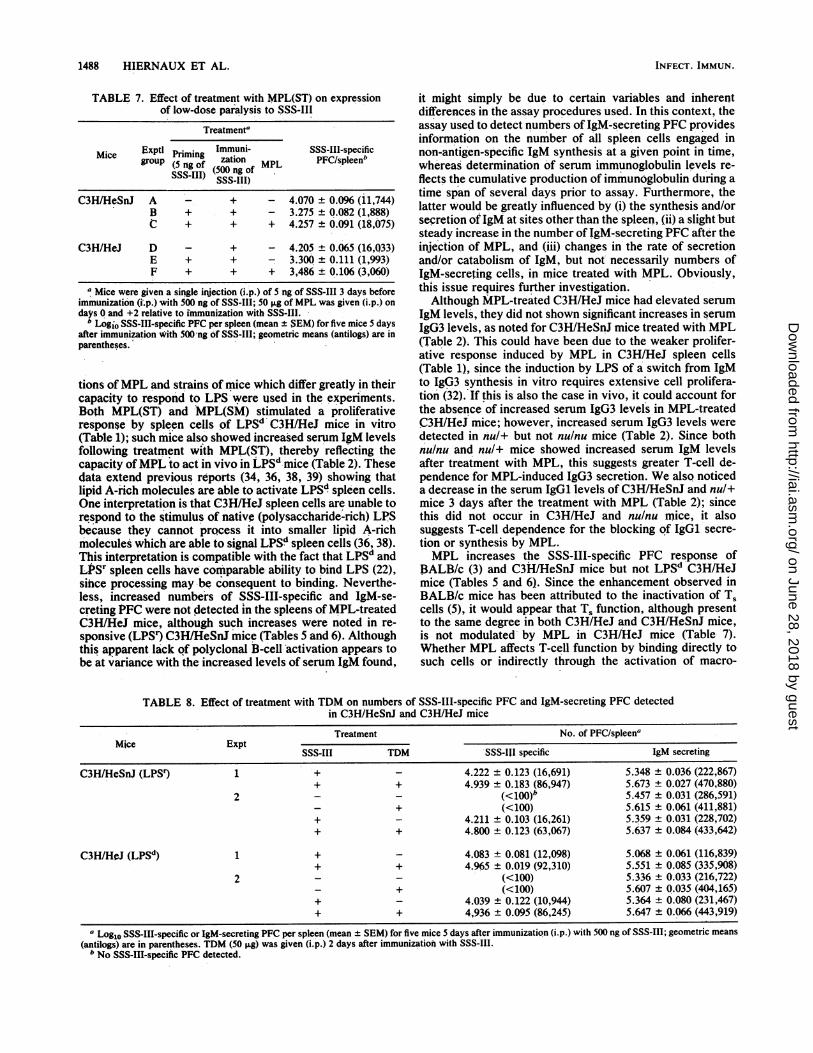
animals of both strains were given 5 ng of SSS-IlI 3 daysbefore immunization with an optimal immunogenic dose of500 ng; this results in the development of low-dose paralysis,a form of antigen-specific unresponsiveness known to bemediated by T5 cells (3, 35). In BALB/c mice, the expressionof low-dose paralysis can be abrogated by treatment withMPL (5). The data in Table 7 show that C3HIHeJ andC3H/HeSnJ mice did not differ in the degree of low-doseparalysis induced, indicating that they do not differ withrespect to T. function; however, low-dose paralysis was
abrogated in C3H/HeSnJ mice (group B versus group C, P <0.001) but not in C3H/HeJ mice (group E versus group F, P> 0.05) treated with MPL(ST).Adjuvant activity ofTDM in C3H mice. The data presented
in Tables 5 and 6 show that MPL cannot significantlyincrease the anti-SSS-III splenic PFC response of C3H/HeJmice. In order to be sure that this does not reflect a generalinability of C3H/HeJ mice to respond to the effects of anybacterial adjuvant, we examined the ability of TDM toinfluence the antibody response to SSS-Ill in C3H/HeJ mice;
the results of preliminary experiments have shown that TDMsignificantly enhances the magnitude of the SSS-III-specificPFC response of BALB/c mice (4). Both C3H/HeJ andC3H/HeSnJ mice treated with 20 ,ug of TDM 2 days afterimmunization with 500 ng of SSS-III showed a significantlyenhanced (P < 0.001) SSS-III-specific PFC response, as wellas a significant increase (P < 0.05) in numbers of 1gM-secreting PFC (Table 8). This indicates that C3H/HeJ miceare sensitive to adjuvants whose activity is not influenced bythe LPSd genetic defect. Moreover, treatment with TDMwithout immunization did not result in the appearance ofdetectable numbers of SSS-III-specific PFC.
DISCUSSION
Since the results of a previous study (5) indicated thatMPL(ST) increases the magnitude of the antibody responseto SSS-Ill by inactivating T, cells, we wished to furthercharacterize the mode of action of MPL with respect to thecapacity of mice to respond to LPS. Two different prepara-
TABLE 6. Effect of treatment with MPL(ST) on numbers of SSS-III-specific PFC and IgM-secreting PFC detectedin C3H/HeSnJ and C3H/HeJ mice
Treatment No. of PFC/spleen"Mice Expt
SSS-III MPL(ST) SSS-I11 specific IgM secreting
C3H/HeSnJ (LPSr) 1 + _ 4.328 ± 0.098 (21,266) NDb+ + 4.683 ± 0.065 (48,246) ND
2 - _ (<100)' 5.109 ± 0.058 (128,386)+ (<100) 5.960 ± 0.071 (132,147)
+ + 4.004 ± 0.101 (10,094) 5.121 ± 0.071 (132,147)+ + 4.649 ± 0.087 (44,580) 5.600 ± 0.087 (398,141)
C3H/HeJ (LPSd) 1 + _ 4.293 ± 0.092 (19,635) ND+ + 4.452 ± 0.079 (28,342) ND
2 - _ (<100) 5.042 ± 0.024 (110,277)+ (<100) 5.110 ± 0.062 (128,874)
+ - 4.011 ± 0.111 (10,248) 5.020 ± 0.103 (104,766)+ + 4.353 ± 0.110 (22,537) 5.142 ± 0.078 (138,772)
a Log1o SSS-III-specific or IgM-secreting PFC per spleen (mean ± SEM) for five mice 5 days after immunization (i.p.) with 500 ng of SSS-III; geometric means(antilogs) are in parentheses. MPL(ST) (50 ,ug) was given (i.p.) 2 days after immunization with 500 ng of SSS-III.
b ND, Not done.No SSS-III-specific PFC detected.
VOL. 57, 1989
on June 28, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
1488 HIERNAUX ET AL.
TABLE 7. Effect of treatment with MPL(ST) on expressionof low-dose paralysis to SSS-III
Treatmenta
Mice Exptl Priming Immuni- SSS-III-specificgrouPr(5ingo zation ML PFC/spleenb
SSS-gIII) (500 ng of MPLSSS-III)
C3H/HeSnJ A - + - 4.070 ± 0.096 (11,744)B + + - 3.275 ± 0.082 (1,888)C + + + 4.257 ± 0.091 (18,075)
C3H/HeJ D - + - 4.205 ± 0.065 (16,033)E + + - 3.300 ± 0.111 (1,993)F + + + 3,486 ± 0.106 (3,060)
a Mice were given a single injection (i.p.) of 5 ng of SSS-III 3 days beforeimmunization (i.p.) with 500 ng of SSS-III; 50 Fg of MPL was given (i.p.) ondays 0 and +2 relative to immunization with SSS-III.
b Logio SSS-III-specific PFC per spleen (mean ± SEM) for five mice 5 daysafter immunization with 500Ong of SSS-III; geometric means (antilogs) are inparentheses.
tions ofMPL and strains of mice which differ greatly in theircapacity to respond to LPS were used in the experiments.Both MPL(ST) and MPL(SM) stimulated a proliferativeresponse by spleen cells of LPSd C3H/HeJ mice in vitro(Table 1); such mice also showed increased serum IgM levelsfollowing treatment with MPL(ST), thereby reflecting thecapacity ofMPL to act in vivo in LPSd mice (Table 2). Thesedata extend previous reports (34, 36, 38, 39) showing thatlipid A-rich molecules are able to activate LPSd spleen cells.One interpretation is that C3H/HeJ spleen cells are unable torespond to the stimulus of native (polysaccharide-rich) LPSbecause they cannot process it into smaller lipid A-richmolecules which are able to signal LPSd spleen cells (36, 38).This interpretation is compatible with the fact that LPSd andLPSr spleen cells have comparable ability to bind LPS (22),since processing mnay be consequent to binding. Neverthe-less, increased numbers of SSS-III-specific and IgM-se-creting PFC were not detected in the spleens of MPL-treatedC3H/HeJ mice, although such increases were noted in re-sponsive (LPSr) C3H/HeSnJ mice (Tables 5 and 6). Althoughthis apparent lack of polyclonal B-cell activation appears tobe at variance with the increased levels of serum IgM found,
it might simply be due to certain variables and inherentdifferences in the assay procedures used. In this context, theassay used to detect numbers of IgM-secreting PFC providesinformation on the number of all spleen cells engaged innon-antigen-specific IgM synthesis at a given point in time,whereas determination of serum immunoglobulin levels re-
flects the cumulative production of immunoglobulin during a
time span of several days prior to assay. Furthermore, thelatter would be greatly influenced by (i) the synthesis and/orsecretion of IgM at sites other than the spleen, (ii) a slight butsteady increase in the number of IgM-secreting PFC after theinjection of MPL, and (iii) changes in the rate of secretionand/or catabolism of IgM, but not necessarily numbers ofIgM-secreting cells, in mice treated with MPL. Obviously,this issue requires further investigation.Although MPL-treated C3H/HeJ mice had elevated serum
IgM levels, they did not shown significant increases in serum
IgG3 levels, as noted for C3H/HeSnJ mice treated with MPL(Table 2). This could have been due to the weaker prolifer-ative response induced by MPL in C3H/HeJ spleen cells(Table 1), since the induction by LPS of a switch from IgMto IgG3 synthesis in vitro requires extensive cell prolifera-tion (32). If this is also the case in vivo, it could account forthe absence of increased serum IgG3 levels in MPL-treatedC3H/HeJ mice; however, increased serum IgG3 levels were
detected in nul+ but not nulnu mice (Table 2). Since bothnulnu and nul+ mice showed increased serum IgM levelsafter treatment with MPL, this suggests greater T-cell de-pendence for MPL-induced IgG3 secretion. We also noticeda decrease in the serum IgGl levels ofC3H/HeSnJ and nul+mice 3 days after the treatment with MPL (Table 2); sincethis did not occur in C3H/HeJ and nulnu mice, it alsosuggests T-cell dependence for the blocking of IgGl secre-tion or synthesis by MPL.MPL increases the SSS-III-specific PFC response of
BALB/c (3) and C3H/HeSnJ mice but not LPSd C3H/HeJmice (Tables 5 and 6). Since the enhancement observed in
BALB/c mice has been attributed to the inactivation of Tscells (5), it would appear that Ts function, although presentto the same degree in both C3H/HeJ and C3H/HeSnJ mice,is not modulated by MPL in C3H/HeJ mice (Table 7).Whether MPL affects T-cell function by binding directly tosuch cells or indirectly through the activation of macro-
TABLE 8. Effect of treatment with TDM on numbers of SSS-IIl-specific PFC and IgM-secreting PFC detectedin C3H/HeSnJ and C3H/HeJ mice
Treatment No. of PFC/spleenaMice Expt
SSS-III TDM -SSS-IJI specific IgM secreting
C3H/HeSnJ (LPS9 1 + _ 4.222 ± 0.123 (16,691) 5.348 ± 0.036 (222,867)+ + 4.939 ± 0.183 (86,947) 5.673 ± 0.027 (470,880)
2 - _ (<10O)b 5.457 ± 0.031 (286,591)+ (<100) 5.615 ± 0.061 (411,881)
+ - 4.211 ± 0.103 (16,261) 5.359 ± 0.031 (228,702)+ + 4.800 ± 0.123 (63,067) 5.637 + 0.084 (433,642)
C3H/HeJ (LPSd) 1 + _ 4.083 ± 0.081 (12,098) 5.068 ± 0.061 (116,839)+ + 4.965 ± 0.019 (92,310) 5.551 ± 0.085 (335,908)
2 - - (<100) 5.336 ± 0.033 (216,722)+ (<100) 5.607 ± 0.035 (404,165)
+ - 4.039 ± 0.122 (10,944) 5.364 ± 0.080 (231,467)+ + 4,936 ± 0.095 (86,245) 5.647 ± 0.066 (443,919)
a Log1o SSS-III-specific or IgM-secreting PFC per spleen (mean ± SEM) for five mice 5 days after immunization (i.p.) with 500 ng of SSS-III; geometric means
(antilogs) are in parentheses. TDM (50 Ixg) was given (i.p.) 2 days after immunization with SSS-III.b No SSS-III-specific PFC detected.
INFECT. IMMUN.
on June 28, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

MPL ACTIVITY IN C3H MICE 1489
phages and the subsequent release of various immunomod-ulators remains to be elucidated; however, it is most likelythat the LPS-binding proteins identified on the surface of Bcells, T cells, and macrophages play an important role in thetransmission of signals relevant to the immunomodulatoryproperties of LPS and MPL (22, 23). Since MPL has beenshown to influence Ts but not TH or amplifier-T-cell function(5), perhaps T, cells possess unique binding sites for eitherMPL or the immunomodulatory products generated upontreatment with MPL. This is now being investigated.The surprising decrease in the serum IgGl levels of
C3H/HeSnJ mice 3 days after the treatment with MPL isonly transient, since serum IgGl levels were significantlyenhanced in mice 12 days after treatment with MPL (Tables2 and 3). Since the production of IgGl can be stimulated byIL-4, which is produced by TH2 cells (18, 33), further studiesshould enable one to determine whether this is due to anegative influence of MPL on the production of IL-4 and/orthe interaction of IL-4 with B lymphocytes soon aftertreatment with MPL. For example, MPL could affect TH2cells by inducing the production of gamma interferon by THicells; gamma interferon has been shown to have a negativeeffect on the proliferation of TH2 cells (12). It should benoted that, although the genetic defect of C3H/HeJ miceprevents them from responding in the same manner asC3H/HeSnJ mice to the adjuvant effect of MPL, it does notalter their capacity to respond to other immunomodulatingagents. TDM, an adjuvant molecule derived from M. phlei,increased the magnitude of the SSS-III-specific antibodyresponse in C3H/HeJ mice, as well as in C3H/HeSnJ mice(Table 8). This suggests that the B cells of both strains ofmice are receptive to stimulation by adjuvants and that themechanisms involved in producing the adjuvant effects byMPL and TDM may differ.
ACKNOWLEDGMENTS
This work was supported through funds available from PublicHealth Service contract 263-80-D-0394 from the National Institute ofAllergy and Infectious Diseases.We are indebted to Brenda Marshall and Vernon Robinson for
expert editorial assistance.
LITERATURE CITED1. Alexander, P., and R. Evans. 1971. Endotoxins and double
stranded RNA render macrophages cytotoxic. Nature (London)New Biol. 232:76-78.
2. Apte, R. N., C. Galanos, and D. V. Pluznik. 1975. Lipid A, theactive part of bacterial endotoxins in inducing serum colonystimulating activity and macrophage progenitor cells. J. Cell.Physiol. 87:71-78.
3. Baker, P. J., D. F. Amsbaugh, P. W. Stashak, G. Caldes, and B.Prescott. 1982. Direct evidence for the involvement of T sup-pressor cells in the expression of low-dose paralysis to type IIIpneumococcal polysaccharide. J. Immunol. 128:1059-1062.
4. Baker, P. J., M. B. Fauntleroy, P. W. Stashak, J. R. Hiernaux,J. L. Cantrell, and J. A. Rudbach. 1989. Adjuvant effects oftrehalose dimycolate on the antibody response to type IIIpneumococcal polysaccharide. Infect. Immun. 57:912-917.
5. Baker, P. J., J. R. Hiernaux, M. B. Fauntleroy, B. Prescott, J. L.Cantrell, and J. A. Rudbach. 1988. Inactivation of suppressorT-cell activity by nontoxic monophosphoryl lipid A. Infect.Immun. 56:1076-1083.
6. Baker, P. J., and P. W. Stashak. 1969. Quantitative and quali-tative studies on the primary antibody response to pneumococ-cal polysaccharide at the cellular level. J. Immunol. 103:1342-1348.
7. Baker, P. J., P. W. Stashak, D. F. Amsbaugh, and B. Prescott.1971. Characterization of the antibody response to type III
pneumococcal polysaccharide at the cellular level. I. Dose-response studies and the effect of prior immunization on themagnitude of the antibody response. Immunology 20:469-481.
8. Baker, P. J., P. W. Stashak, and B. Prescott. 1969. The use oferythrocytes sensitized with purified pneumococcal polysaccha-rides for the assay of antibody and antibody-producing cells.Appl. Microbiol. 17:422-426.
9. Behling, U. H., and A. H. Nowotny. 1982. Bacterial endotoxinsas modulators of specific and nonspecific immunity, p. 127-140.In C. DeLisi and J. R. Hiemaux (ed.), Regulation of immuneresponse dynamics, vol. 2. CRC Press, Inc., Boca Raton, Fla.
10. Carswell, E. A., L. J. Old, R. L. Kassel, S. Green, N. Fiore, andB. Williamson. 1975. An endotoxin-induced serum factor thatcauses tumor necrosis. Proc. Natl. Acad. Sci. USA 72:3666-3670.
11. Coutinho, A., R. Benner, M. Bjorklund, L. Forni, D. Holmberg,F. Ivars, C. A. Martinez, and S. Pettersson. 1982. A "trans"perspective on the control of immunoglobulin C gene expres-sion. Immunol. Rev. 67:87-114.
12. Gajewski, T. F., and F. W. Fitch. 1988. Anti-proliferative effectof IFN--y in immune regulation. I. IFN--y inhibits the prolifera-tion of Th2 but not Thl murine helper T lymphocyte clones. J.Immunol. 140:2245-2252.
13. Galanos, C., 0. Luderitz, and 0. Westphal. 1969. A new methodfor the extraction of R lipopolysaccharides. Eur. J. Biochem.9:245-249.
14. Gery, I., J. Kruger, and S. Spiesel. 1972. Stimulation of Blymphocytes by endotoxin. Reaction of thymus-deprived miceand karyotypic analysis of dividing cells in mice bearing T6T6thymus grafts. J. Immunol. 108:1088-1091.
15. Gottlieb, C. F. 1974. Application of transformations to normal-ize the distribution of plaque-forming cells. J. Immunol. 113:51-57.
16. Gronowicz, E., A. Coutinho, and F. Melchers. 1976. A plaqueassay for all cells secreting Ig of a given type or class. Eur. J.Immunol. 6:588-590.
17. Hiernaux, J. R., J. M. Jones, J. A. Rudbach, F. Rollwagen, andP. J. Baker. 1983. Antibody response of immunodeficient (Xid)CBA/N mice to Escherichia coli 0113 lipopolysaccharide, athymus-independent antigen. J. Exp. Med. 157:1197-1207.
18. Isakson, P., E. Pure, E. S. Vitetta, and P. H. Krammer. 1982. Tcell-derived B cell differentiation factor(s). Effect on the isotypeswitch of murine B cells. J. Exp. Med. 155:734-748.
19. Izui, S., R. A. Eisenberg, and F. J. Dixon. 1981. Subclass-restricted IgG polyclonal antibody production in mice injectedwith lipid A-rich lipopolysaccharides. J. Exp. Med. 153:324-338.
20. Johnson, A. G., S. Gaines, and M. Landy. 1956. Studies on the0-antigen of Salmonella typhosa. V. Enhancement of antibodyresponse to protein antigens by the purified lipopolysaccharide.J. Exp. Med. 103:225-246.
21. Jones, J. M., D. F. Amsbaugh, P. W. Stashak, B. Prescott, P. J.Baker, and D. W. Alling. 1976. Kinetics of the antibody re-sponse to type III pneumococcal polysaccharide. III. Evidencethat suppressor cells function by inhibition of the recruitmentand proliferation of antibody-producing cells. J. Immunol. 116:647-656.
22. Lei, M.-G., and D. C. Morrison. 1988. Specific endotoxiclipopolysaccharide-binding proteins on murine splenocytes. I.Detection of lipopolysaccharide-binding sites on splenocytesand splenocyte subpopulations. J. Immunol. 141:996-1005.
23. Lei, M.-G., and D. C. Morrison. 1988. Specific endotoxiclipopolysaccharide-binding proteins on murine splenocytes. II.Membrane localization and binding characteristics. J. Immunol.141:1006-1011.
24. Luderitz, O., C. Galanos, and E. T. Rietschel. 1982. Endotoxinsof gram-negative bacteria. Pharmacol. Ther. 15:383-402.
25. Mizel, S. B., J. J. Oppenhein, and D. L. Rosenstreich. 1978.Characterization of lymphocyte-activating factor (LAF) pro-duced by a macrophage cell line, P388D. II. Biochemicalcharacterization of LAF induced by activated T cells and LPS.J. Immunol. 120:1504-1514.
26. Perrson, U. 1977. Lipopolysaccharide-induced suppression of
VOL. 57, 1989
on June 28, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

1490 HIERNAUX ET AL.
the primary immune response to a thymus-dependent antigen. J.Immunol. 118:789-796.
27. Qureshi, N., P. Mascogni, E. Ribi, and K. Takayama. 1985.Monophosphoryl lipid A obtained from lipopolysaccharides ofSalmonella minnesota R595. Purification of the dimethyl deriv-ative by high performance liquid chromatography and completestructural determination. J. Biol. Chem. 260:5271-5278.
28. Qureshi, N., K. Takayama, and E. Ribi. 1982. Purification andstructural determination of nontoxic lipid A from the roughmutant of Salmonella typhimurium. J. Biol. Chem. 257:11808-11815.
29. Ribi, E. 1984. Beneficial modification of the endotoxin mole-cule. J. Biol. Response Medif. 3:1-9.
30. Ribi, E., J. L. Cantrell, K. Takayama, H. 0. Ribi, K. R. Meyers,and N. Qureshi. 1986. Modulation of humoral and cell-mediatedimmune responses by a structurally established nontoxic lipidA, p. 407-420. In A. Szentivanyi and H. Friedman (ed.),Immunology and immunopharmacology of bacterial endotoxins.Plenum Publishing Corp., New York.
31. Seppala, I. J. T., and 0. Makela. 1984. Adjuvant effect ofbacterial LPS and/or alum precipitation in response to polysac-charide and protein antigens. Immunology 53:827-836.
32. Severinson, E., S. Bergstedt-Lindqvist, W. van der Loo, and C.Fernandez. 1982. Characterization of the IgG response inducedby polyclonal B cell activators. Immunol. Rev. 67:73-85.
33. Stevens, T. L., A. Bossie, V. M. Sanders, R. Fernandez-Botran,R. L. Coffman, T. R. Mosmann, and E. S. Vitetta. 1988.Regulation of antibody isotype secretion by subsets of antigen-
specific helper T cells. Nature (London) 334:255-258.34. Sultzer, B. M., and R. Castagna. 1988. Inhibition of activated
nonresponder C3H/HeJ lymphocytes by lipopolysaccharide en-dotoxin. Infect. Immun. 56:3040-3045.
35. Taylor, C. E., D. F. Amsbaugh, P. W. Stashak, G. Caldes, B.Prescott, and P. J. Baker. 1983. Cell surface antigens and othercharacteristics of T cells regulating the antibody response totype III pneumococcal polysaccharide. J. Immunol. 130:19-23.
36. Tomai, M. A., A. G. Johnson, and E. Ribi. 1988. Glycolipidinduced proliferation of lipopolysaccharide hyporesponsiveC3H/HeJ splenocytes. J. Leukocyte Biol. 43:11-17.
37. Tomai, M. A., L. F. Solem, A. G. Johnson, and E. Ribi. 1987.The adjuvant properties of a nontoxic monophosphoryl lipid Ain hyporesponsive and aging mice. J. Biol. Response Modif.6:99-107.
38. Vogel, S. N., G. S. Madonna, L. M. Wahl, and P. D. Rick. 1984.In vitro stimulation of C3H/HeJ spleen cells and macrophagesby a lipid A precursor molecule derived from Salmonellatyphimurium. J. Immunol. 132:347-353.
39. Vukajlovich, S. W., and D. C. Morrison. 1983. Conversion oflipopolysaccharides to molecular aggregates with reduced sub-unit heterogeneity: demonstration of KPS responsiveness in"endotoxin-unresponsive" C3H/HeJ splenocytes. J. Immunol.130:2804-2808.
40. Westphal, O., 0. Luderitz, and F. Bister. 1952. Uber dieExtraktian von Bakterien mit Phenol-Wasser. Z. Naturforsch.Teil B 7:148-155.
INFECT. IMMUN.
on June 28, 2018 by guesthttp://iai.asm
.org/D
ownloaded from










![4,800 122,000 135M · 2020. 5. 16. · other chronic diseases, such as rhinosinusitis and chronic obstructive pulmonary disease (COPD) [2]. ... mast cells, NKT cells, γδ-Tcells,](https://img.pdfslide.net/doc/110x75/60ffef71ac04547dec6006b4/4800-122000-135m-2020-5-16-other-chronic-diseases-such-as-rhinosinusitis.jpg)


















