Embed Size (px)
Citation preview

ORIGINAL RESEARCH
Impaired Structural and Functional Development of CerebellumFollowing Gestational Exposure of Deltamethrin in Rats: Roleof Reelin
Kamendra Kumar • Nisha Patro • Ishan Patro
Received: 23 October 2012 / Accepted: 27 April 2013 / Published online: 17 May 2013
� Springer Science+Business Media New York 2013
Abstract Reelin is an extracellular matrix molecule that
is involved in the normal development of the cerebellar
lamination, Bergmann glial fibres alignment, Purkinje cell
monolayer arrangement and granule cell migration. In this
study, we have examined the effects of maternal exposure
of deltamethrin (DLT), a type II pyrethroid insecticide, on
the structural and functional development of rat cerebellum
during postnatal life. DLT (0.75 mg/kg body weight,
intraperitoneally dissolved in dimethylsulphoxide) was
administered in timed pregnant rats during two different
gestational time periods, i.e. gestational days of 7–10 and
11–14, respectively. In DLT exposed rats, a significant
overexpression of reelin was observed in the cells of the
external granule cell layer (EGL) and internal granule cell
layer along with an ectopic expression of reelin in the EGL
as well as in the migrating granule cells just below the
EGL, revealing an arrest of granule cell migration in this
zone. Mis-orientation and hypertrophy of the Bergmann
glial fibres further hampered the journey of the granule
cells to their final destination. Possibly reelin overexpres-
sion also caused misalignment of the Purkinje cells and
inhibited the neurite growth leading to a significant
decrease in the spine density, main dendritic length and
width of the dendritic arbour. Thus, it is proposed that the
DLT exerts its neurotoxic effects possibly via the intra-
cellular accumulation and low release of reelin leading to
an impaired granule cell and Purkinje cell migration,
inhibition of neurite outgrowth and reduced spine density.
Such impaired cerebellar development leads to motor
coordination deficits.
Keywords Cerebellum � Deltamethrin �Granule cell migration � Prenatal toxicity �Purkinje cells � Reelin
Introduction
The vulnerability of the developing nervous system to
environmental toxicants is related mainly to the period of
exposure and is accelerated, if the exposure occurs during
the development of that organ. Neural development com-
mences in the second week of gestation in rats and con-
tinues until postnatal day 21, thus drawing many
vulnerability windows for adverse exposure. Pyrethroids
are synthetic insecticides and are being increasingly used in
veterinary, agriculture and home pest control (Amweg
et al. 2005; Wolansky et al. 2009; Soderlund 2010). Human
exposure to pyrethroids is well documented to pregnant
women, infants and children (Berkowitz et al. 2003; Heu-
dorf et al. 2004; Shafer et al. 2005). Proposed mechanism
of action of pyrethroids is the prolonged opening of the
neuronal voltage sensitive sodium channels (Vijverberg
and van den Bercken 1990) leading to altered neuronal
excitability and firing rates (Narahashi 2000).
Deltamethrin (DLT), [IUPAC name: (S)-a-cyano-3-
phenoxyphenyl (1R)-cis-3-(2,2-dibromovinyl)-2,2-dime-
thyl-cyclopropanecarboxylate)] is a widely used a-cyano
group containing type-II synthetic pyrethroid insecticide
inducing a prototype type II neurological syndrome char-
acterized by choreoathetosis and salivation (‘CS’ syn-
drome; Verschoyle and Aldridge 1980; Soderlund et al.
K. Kumar � N. Patro � I. Patro (&)
School of Studies in Neuroscience, Jiwaji University,
Gwalior 474011, Madhya Pradesh, India
e-mail: [email protected]
I. Patro
School of Studies in Zoology, Jiwaji University,
Gwalior 474011, Madhya Pradesh, India
123
Cell Mol Neurobiol (2013) 33:731–746
DOI 10.1007/s10571-013-9942-7

2002; Ray and Fry 2006; Wolansky et al. 2009). Neuronal
development involves proliferation of neuronal stem cells
followed by differentiation and migration to their exact
location in the brain. Most of them form synapses with
neighbouring cells and become functional, while others
undergo apoptosis (Gohlke et al. 2008). The proliferation,
differentiation, migration and positioning of neuronal and
glial cells are essential steps during the development of the
CNS (D’Arca et al. 2010).
Developing cerebellum provides an ideal tissue system
to study the fundamental mechanism of these processes
because of its simple structure and clear cytoarchitecture
with only a few neuron types (Sergaki et al. 2010). The
principal neurons of the cerebellum are developed from
two regions. All glutamatergic neurons, i.e. granule cells
and some neurons of deep cerebellar nuclei originate from
the rhombic lip, while the Purkinje neurons and all cere-
bellar interneurons and GABAergic neurons of the deep
cerebellar nuclei develop from the ventricular zone
(Chizhikov et al. 2006; Hevner et al. 2006). During early
postnatal life, granule cell precursors proliferate exten-
sively in the external granule cell layer (EGL) and then
migrate radially via contact guided Bergmann glial cell
processes and form the internal granule cell layer (IGL).
While the Purkinje cells move radially from the germinal
zone of the fourth ventricle towards the cerebellar cortical
surface, and then gets arranged in a single layer and sub-
sequently develop extensively arbourized dendritic tree
(Altman and Bayer 1995). Thus, the cell migration and
correct positioning of specific cell populations are the most
critical aspect of the brain development. Reelin is one such
protein that plays an important role in the architectonic
development of the CNS.
Reelin, an extracellular matrix glycoprotein is exclu-
sively expressed by all glutamatergic granule cells in the
cerebellum at all postnatal ages in rodents (Ramos-Moreno
et al. 2006; Sinagra et al. 2008). During early embryonic
life, reelin regulates the migration and formation of proper
cytoarchitecture of laminated regions of the CNS, viz.
neocortex, hippocampus and cerebellum (D’Arcangelo
et al. 1995; Tissir and Goffinet 2003; Frotscher 2010). In
addition, the role of reelin in modulation of synaptic
plasticity (Weeber et al. 2002; Beffert et al. 2005) and
dendritic growth (Matsuki et al. 2008; Niu et al. 2008)
during postnatal and adult life is also well documented.
Sinagra et al. (2008) demonstrated in a cerebellar culture
that reelin is secreted and bound by a homogenous popu-
lation of glutamatergic granule cells and most of the
components of the reelin signalling pathway, i.e., reelin
and reelin receptors, very low-density lipoprotein receptors
(VLDLR) and Apolipoprotein E receptor-2 (ApoER2) are
also expressed by cerebellar granule cells, imposing auto-
crine and/or paracrine effects on these neurons. However,
the reelin receptors are exclusively expressed by Purkinje
cells in the developing cerebellum in vivo (Perez-Garcia
et al. 2004); while only the splice variant of ApoER2 was
found to be expressed on the cells of the internal granular
layer (Hibi et al. 2009). Thus, in the cerebellum, the reelin
signalling is important for the Bergmann glial guided
migration of Purkinje and granule cells, positioning and
maturation of Purkinje cells, axon guidance, dendritic
morphology and synaptic plasticity (Yoshiki and Kusakabe
1998; Hevner 2008). However, the precise role of reelin in
postnatal and adult cerebellum needs further investigation.
We have earlier reported that DLT exposure during
postnatal period significantly affect the cerebellar lamina-
tion (Patro et al. 1997), proliferation and migration of
postnatally generated granule cells due to the delayed
appearance, disorganization and hypertrophy of Bergmann
glial fibres and astrogliosis (Patro and Patro 2005) and
results in reduced Purkinje cell dendritic arbourization and
motor activity (Patro et al. 2009). The present study was
undertaken to study the effects of DLT when exposed
during prenatal life, the time when the neural tube forma-
tion and nervous system development is initiated, and to
investigate whether such changes are worsened or remain
the same. This study also focuses whether the migration
and subsequent lamination defects are associated with
reelin expression, being it is well established role in these
events and further, if reelin could be a potential target for
DLT neurotoxicity during early brain development.
Experimental Procedures
Test Material
Technical grade DLT purchased as pure compound (98 %
purity) from Sigma was used in the present study.
Test Species and Husbandry
All the animals used for the present study were raised in the
animal house of the School of Studies in Neuroscience
according to the lay down rules of the Jiwaji University
Ethical Committee on Animal handling. All the animals
were maintained in an animal room designed to maintain
temperature at 25 ± 2 �C, relative humidity at *50 % and
a 12-h light and 12-h dark photoperiod. All animals were
fed with standard rat pellet feed and water ad libitum
throughout the study.
Breeding and Treatment
Healthy albino Wistar rats were used for breeding with two
females and one male housed in standard plastic cages
732 Cell Mol Neurobiol (2013) 33:731–746
123

(52 9 28 9 22 cm3). The estrous cycle of the female rats
was checked each morning by collecting vaginal smear on
a glass slide using a Pasteur pipette containing phosphate
buffered saline (PBS) and examined under the microscope.
Mating was confirmed by the presence of spermatozoa in
the vaginal smear. The day on which the vaginal plug was
observed was considered as the gestational day 0 (GD 0).
DLT was dissolved in dimethyl sulfoxide (DMSO, Sigma)
to provide a rapid and complete absorption, and then
injected at a dosage of 0.75 mg/kg body wt/day, i.p, to the
pregnant females during GD 7–10 (n = 6) and 11–14
(n = 6). The day the pups were born was designated as
postnatal day 0 (PND0). The animals were observed for
any clinical sign of toxicity and change in body weight as a
reaction to treatment during the entire period of treatment
till parturition.
The pups born to mothers exposed during GD 7–10 were
considered as DLT-I group, while the pups born to mothers
exposed during GD 11–14 were considered as DLT-II
group. Similar quantity of DMSO was injected to control
females (n = 6) and the pups born to these females were
used as age matched controls (Cont). All the experimental
protocols were pre-approved by the Institutional Ethical
Committee.
The pups born to both treated and control females were
anesthetized on PND’s 0, 3, 7, 12, 15, 21, 30, 60 and 90
(n = 3). The animals were perfusion-fixed (transcardially)
with 2 % paraformaldehyde prepared in 0.01 M phosphate
buffer (pH 7.4) after flushing with phosphate buffer saline
(PBS). The brains were dissected out and the cerebella
were separated and transected at the tectal level and post-
fixed in the same fixative for 24 h at 4 �C. The tissues were
then washed with the phosphate buffer and cryoprotected
in sucrose gradients (10, 20 and 30 % in phosphate buffer).
The sagittal sections (15 lm) were cut through the vermis
region of the cerebellum with a Leica cryotome (CM1900)
and collected on serially numbered gelatin-coated slides.
These slides were stored at -20 �C until immunohisto-
chemical procedures were performed.
Immunohistochemistry
Cryocut sections through the vermis region from the various
groups were selected and processed for the immunocyto-
chemical labelling with anti-GFAP and anti-reelin anti-
bodies. The tissues were air dried and rinsed in PBS to
remove cryomount. The tissues were incubated with 1 %
triton X-100 in PBS for 30 min for membrane permeabili-
zation. This was followed by three washings in PBS and
incubation with 1 % H2O2 in PBS for 20 min to block the
endogenous peroxidases. After three washings with PBS, the
sections were incubated with 1 % normal goat serum in PBS
for 60 min at room temperature in a humid chamber for
non-specific protein blocking. The sections were subse-
quently incubated overnight at 4 �C with rabbit polyclonal
antibodies, i.e. anti-GFAP (glial fibrillary acidic protein;
Dako) and Reelin (Sigma) at a titre of 1:1000 diluted with
1 % BSA in PBS. Next day, the sections were brought to
room temperature and then rinsed with three changes of PBS
for 5 min each and further incubated with secondary anti-
body, i.e. biotinylated goat anti-rabbit (diluted with 1 %
BSA in PBS, 1:100, Sigma) for 90 min. The sections were
again rinsed in PBS three times for 5 min each and incubated
with streptavidin biotin–horseradish peroxidase complex
(diluted with 1 % BSA in PBS, 1:100, Amersham) for
90 min. The sections after washing with PBS were incu-
bated with 0.025 % 3,30-diaminobenzidine tetrahydrochlo-
ride (DAB, Sigma) and 0.03 % hydrogen peroxide (BDH,
England) in PBS as substrate chromogen system for 20 min.
The sections after thorough washing in distilled water were
counterstained with 0.1 % cresyl violet acetate (Sigma
certified stain, C-5042), prepared in acetate buffer (pH 3.5)
air dried, dehydrated quickly in n-butyl alcohol, cleared in
xylene and mounted in DPX.
Image Analyses and Quantification
High quality colour images were captured using Leica
DM6000 microscope equipped with Leica DFC 420 RC
digital camera and Leica Application Suite (LAS) software.
All images were acquired at the same exposure and digital
gain settings to remove occurrence of false-immunoposi-
tivity across sections. The area fraction of reelin expression
was measured with NIH ImageJ software (http://www.rsb.
info.nih.gov/ij).
Cerebellar Layer Thickness Measurements
A separate set of pups from all the three groups were per-
fused with 10 % buffered formalin and processed for par-
affin sectioning. The cerebella were removed by transecting
at the tectal level, post-fixed overnight and then washed to
clear the formalin from the tissues. Tissue blocks were
dehydrated in an ascending series of alcohols, viz. 30, 50, 70
and 90 % by immersing the tissue blocks for 45 min in each.
The tissues were finally dehydrated by immersing in two
changes of absolute alcohol for 30 min each. The tissues
were then cleared with two changes of toluene 45 min each,
infiltrated with Paraplast (Sigma, m. p. 56–58 �C) for 4 h in
a pre-heated incubator to 58 �C. Paraffin blocks were pre-
pared and serial sections were cut at a thickness of 6 lm
using Leica RM 2135 microtome. The sections were stained
with 0.1 % cresyl violet acetate (Sigma certified stain,
C-5042), prepared in acetate buffer (pH 3.5). Cresyl violet
stained sections (through vermis region) were used for
morphometric studies, i.e. the thickness measurement of
Cell Mol Neurobiol (2013) 33:731–746 733
123

various cerebellar layers using an ocular micrometer scale
fitted in the eye piece of a microscope. In total, 20 readings
were taken for each layer measurement, 2 from each folium
as per the sites shown in Fig. 1a, b.
Rapid Golgi Impregnation
Rapid Golgi impregnation method (modified from Stensaas
1967 by Patro et al. 2009) was used to study the dendritic
arbourization and dendritic spine counts in normal and DLT
treated developing rat cerebella. The cerebella from both the
treated and control pups (3 pups each parameter) were dis-
sected out after ether anaesthesia and directly immersed in
Golgi fixative (potassium dichromate—5 g, chloral
hydrate—5 g, glutaraldehyde—8 ml, formaldehyde—6 ml
and dimethyl sulphoxide—10 drops; total volume was made
to 100 ml with distilled water) in dark amber bottles for
4 days. The tissues blocks were rinsed briefly with 0.75 %
aqueous solution of silver nitrate (Qualigens) to remove any
precipitates and then transferred to the fresh solution of
AgNO3 and left for 5 days in dark. Following this the tissues
were properly washed in 70 % alcohol to remove any pre-
cipitates and 100 lm thick sections were cut using a Leica
automatic vibratome (VT1000S). The sections were picked
one by one and collected in 70 % alcohol, dehydrated in
absolute alcohol, cleared in xylene and mounted in DPX. All
the sections through the vermis regions were scanned and
visualized with Leica DM6000 microscope fitted with a
digital camera.
Motor Coordination
Motor coordination was assessed using a motor driven
treadmill, Rotamex-5 from Columbus instruments. The test
was consistently performed between 9.00 am to 4.00 pm in
rat pups from the age of 21 days onwards when true
walking develops. The Rotarod consists of a semi-enclosed
chamber which contains a series of 32 infrared beams
(diameter 3 cm and width 5 cm) and a rotating rod
suspended at a height of 35 cm above the floor. The ani-
mals were acclimatized for three consecutive days before
the day of final recording at start speed of 2 rpm and a
maximum speed of 8 rpm for a total duration of 100 s. The
final recording was performed at a start speed of 2 rpm and
maximum speed of 40 rpm for a total duration of 420 s,
24 h after the last acclimatization for all animal groups.
The latency to fall was recorded automatically by the
photocells as the total length of time spent by the animal on
the rotating rod with the help of software (Rotamex 5).
Minimum of four final recordings were taken for the same
animal group and the results were averaged to obtain a
single mean value for each animal.
Statistical Analysis
All the data was expressed as mean ± standard error of
means (SEM) and were analysed by one-way analysis of
variance (ANOVA) followed by Tukey’s post hoc test for
multiple comparisons using SigmaStat version 3.5 for
Windows. Values of p B 0.05 were considered significant.
Results
No signs of toxicity were observed during the period of
treatment. There was no mortality of the treated pregnant
females or the pups born to them. No significant difference
in body weight of the DLT-treated and vehicle treated
mothers was found. However, the body weight of both the
treated group pups (DLT-I and DLT-II) remained signifi-
cantly low as compared with their age matched controls
(Table 1; Fig. 2).
Cerebellar Layer Thickness
Cerebellar cortical lamination in rodents is a completely
postnatal event and simultaneously the expansion of the
cerebellar cortex also takes place. Such processes require
I
II
III
IV
VIaV
VIb-c
VII
VIII
IX
X
a
ML
EGL
PCL
IGL
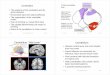
bFig. 1 Schematic
representation of sagittal section
from the vermis region of rat
cerebellum. a Rectangularboxes shown with red colour(folia VII) indicate the sites
selected from each of the X
folia. b Four different layers:
EGL external granular layer, MLmolecular layer, PCL Purkinje
cell layer, IGL internal granular
layer as delineated by arrows.
The magnification scale is
50 lm (Color figure online)
734 Cell Mol Neurobiol (2013) 33:731–746
123

the migration of the prenatally born neurons to form spe-
cific layers, i.e. Purkinje cell layer and molecular layer and
also the genesis and migration of neurons forming internal
granular layer. Thus, the appearance and organization of
cortical lamination have been investigated following
maternal exposure of DLT in cresyl violet stained cere-
bellar sections through vermis region at various postnatal
ages, i.e. 0, 3, 7, 12, 15, 21, 30, 60 and 90 days (Fig. 3b, c,
e, f, h, i, k, l).
Extensive proliferation and migration of the progenitors
in the EGL during early postnatal period leads to the for-
mation of IGL. Histo-morphometric analysis depicted that
there was a gradual increase in the EGL thickness from 0 to
7 days control cerebella after which there was a gradual
decline, leading to a complete depletion of EGL in 21-day-
old pups (Fig. 3a, d, g, j). However, in both the DLT
treated groups studied, i.e. DLT-I and DLT-II, there was a
reduced and delayed growth of the EGL reaching to a peak
at P12, and then declined gradually (Table 2; Fig. 4a).
Thus, on day 7, there was a significant reduction in the
EGL thickness in both DLT-I and II groups (F2,39 = 3.8;
p B 0.05). However, a significantly thicker EGL was
recorded following DLT treatment in the DLT-I group on
days 12 and 15 (F2,39 = 4.3, 3.9, respectively; p B 0.05)
and in DLT-II group on day 12 (F2,39 = 4.2; p B 0.05). A
comparatively thicker EGL was recorded on day 21 fol-
lowing DLT exposure than 21 day controls, but was
insignificant.
Mean molecular layer (ML) thickness was observed to
be significantly reduced following DLT exposure in both
DLT-I and II groups at P12 (F2,39 = 3.5, 3.6, respectively;
p B 0.05) and DLT-I at P15 (F2,39 = 3.8; p B 0.05).
However, till P60, the ML thickness remained low in
treated group cerebella and was regained to normal values
by P90 only (Table 2; Fig. 4b).
During normal cerebellar development, Purkinje cell
monolayer arrangement is achieved by P7 and after that
there is an expansion of the layer because of the growth of
the Purkinje cell till 21 days. The Purkinje cell layer (PCL)
remained significantly thicker following DLT exposure and
in both the treated pups, i.e. DLT-I and DLT-II, there was a
significant increase in PCL thickness on P12 (F2,39 = 10.3,
4.4, respectively; p B 0.01, 0.05) and P15 (F2,39 = 9.6,
4.1, respectively; p B 0.01, 0.05). However, the PCL
remained thicker throughout the entire period of study till
P90, although, statistically insignificant at p = 0.05
(Table 2; Fig. 4c).
The mean thickness of the IGL was also found to be
reduced in the DLT-treated groups as compared with their
respective controls. The difference was found to be sig-
nificant on days 12 and 15 (F2,39 = 4.5, 4.2, respectively;
p B 0.01, 0.05) in DLT-I group and on day 12
(F2,39 = 4.8; p B 0.05) in DLT-II group. However, the
IGL thickness remained low throughout the period of
study, and did not reach to the normal control values until
maturity, i.e. P90 (Table 2; Fig. 4d).
Reelin Immunohistochemistry
Reelin is a large extracellular glycoprotein of 420–
450 kDa. In developing cerebellum, it is secreted by
Table 1 The body weight of the developing rat pups (g) following maternal exposure to deltamethrin during gestation
Groups 0 day 3 days 7 days 12 days 15 days 21 days 30 days 90 days
Body weight during postnatal life
Controla 9.7 ± 0.5 13.0 ± 0.4 19.3 ± 0.7 27.5 ± 0.4 32.7 ± 0.7 41.2 ± 1.1 48.9 ± 1.7 202 ± 1.2
DLT-Ib 7.5 ± 0.5 8.2 ± 0.5* 12.7 ± 0.5* 17.2 ± 0.9* 21.2 ± 0.6* 24.4 ± 0.8* 33.5 ± 1.1* 166 ± 3**
DLT-IIc 7.1 ± 0.4 9.9 ± 0.6* 14.7 ± 0.6* 19.5 ± 1.1* 23.3 ± 0.8* 25.4 ± 1.0* 34.8 ± 1.0* 173 ± 2.9**
Values are expressed as mean ± SEM (n = 6)
* p \ 0.05 for DLT-I and DLT-II versus control; ** p \ 0.01 for DLT-I and DLT-II versus control (one-way ANOVA followed by post hoc
Tukey’s test for multiple comparison among groups)a Vehicle treated (equal volume of DMSO only) groupb DLT dissolved in DMSO at a dose of 0.75 mg/kg body weight at GD 7–10c DLT dissolved in DMSO at a dose of 0.75 mg/kg body weight at GD 11–14
0
50
100
150
200
250
0d 3d 7d 12d 15d 21d 30d 90d
Bo
dy
wei
gh
t (g
)
Postnatal age (days)
Cont
DLT-I
DLT-II
* * * * ** ** * *
****
**
Fig. 2 Graph represents the body weight presented as the mean ±
SEM of the 6 pups/parameter/time point in control (Cont), DLT-I and
DLT-II groups. *DLT-I and DLT-II versus cont; p \ 0.05, **DLT-I
and DLT-II versus cont; p \ 0.01
Cell Mol Neurobiol (2013) 33:731–746 735
123

glutamatergic granule cells and plays a key role in
instructing neurons to achieve normal differentiation when
they reach their final destination at the end of the radial
migration. Reelin immunohistochemistry revealed its spe-
cific expression by the cells of EGL and the granule cells of
the IGL (Fig. 5a, d, g, j, n, q). Quantification of the reelin
expression in the control cerebella depicted an intense
expression on 3rd postnatal day, a subsequent down regu-
lation till 12th day, and then a steep decline with a
significant down regulation of expression by postnatal day
21 (Table 3; Fig. 6). Following DLT exposure, a signifi-
cantly enhanced expression of reelin was recorded in both
the cells of the EGL and IGL. In the DLT-I group, the
reelin expression was significantly high at all the age-
points studied, i.e. 0, 3, 7, 12 and 15 (Fig. 5b, e, h, k, l, o, r),
while in DLT-II group as well the reelin was over
expressed till postnatal day 21 (Fig. 5c, f, i, m, p, s), but
the significant difference was found only on 0, 7 and
Cont DLT-IIDLT-I
12d
EGL
ML
PCL
IGL
WM
a b c
15d
d e f
21d
g h i
90d
j k l
Fig. 3 Photomicrographs showing cresyl violet stained sagittal
sections from the cerebellar vermal region of control (a, d, g, j),DLT-I (b, e, h, k) and DLT-II (c, f, i, l) group pups at various age
points. Arrows (black) represents the granule cells in the EGL,
arrowheads (red) shows arrested granule cell migration and thick fatarrows (red) represents distorted Purkinje cell monolayer arrange-
ment and clustering. WM white matter. The scale bar is 50 lm (Color
figure online)
736 Cell Mol Neurobiol (2013) 33:731–746
123

15 days postnatally (Table 3; Fig. 6). On postnatal day 15
in DLT-I group cerebella, an ectopic expression of reelin
was observed in the EGL as well as in the migrating
granule cells just below the EGL, revealing an arrest of
granule cell migration in this zone (Fig. 5h, k, l). However,
by day 21, when most of the granule cell migration is over,
reelin expression still remain statistically different in both
DLT-I and DLT-II group cerebella as compared with their
age-matched controls, but values were significant only in
DLT-II group (Table 3; Fig. 6).
GFAP Immunohistochemistry
Bergmann glia, the unipolar cerebellar astrocytes are
involved in the migration of granule cells in the developing
cerebellum during early postnatal life. Similar to astro-
cytes, the GFAP antibody also acts as a specific marker for
Bergmann glia as well. Anti-GFAP immunolabeling and
co-staining with cresyl violet enabled us to study the
migration of granule cells from the EGL, the site of
their origin to the IGL where they are finally going to
reside in the adult cerebellum. During normal cerebellar
development, the granule cell proliferation and migration
occurs during PND0-21. The Bergmann glial fibres appears
immediately after birth and run straight between the ven-
tricular and the pial surface with their cell bodies lying at
the base of the Purkinje cells and the processes anchored at
the pial surface, leading to the normal migration of the
granule cells. The migration was observed to be at peak
from postnatal days of 7–15. However, the migration still
continued till PND21, when the EGL was completely
seized (Fig. 3h, i). In the DLT treated cerebella, the GFAP
immunolabelled Bergmann glial fibres were disorganised
and hypertrophied (Fig. 7b, c, e, f, h, i, k, l) as compared
with their thin and straight morphology in the controls
(Fig. 7a, d, g, j). At the same time, these fibres were more
strongly labelled with GFAP. Such disorganised morphol-
ogy and mal-orientation of Bergmann glial fibres were
more prominent in DLT-I group (Fig. 7b, e, h, k) as com-
pared with the DLT-II group preparations (Fig. 7c, f, i, l).
Even there was a clear cut indication of delayed migration as
evident from peak migration on 12th and 15th postnatal days
(Fig. 7d–i) and a persistence of EGL till PND21 (Figs. 3h, i,
and 7j, k, l). A comparatively more number of astrocytes with
Table 2 Thickness of various cerebellar cortical layers during postnatal development following maternal exposure to deltamethrin during
gestation
Groups 0 day 3 days 7 days 12 days 15 days 21 days 30 days
EGL thickness (lm)
Conta 17 ± 2.5 30 ± 0.9 38 ± 1.2 27 ± 1.4 15 ± 1.0 1.8 ± 0.8 1.4 ± 0.7
DLT-Ib 13 ± 2.4 25 ± 1.3 31 ± 1.9* 36 ± 1.9* 24 ± 1.5* 2.3 ± 0.4 1.5 ± 0.4
DLT-IIc 15 ± 1.5 27 ± 1.1 30 ± 1.8* 35 ± 1.3* 21 ± 1.7 2.5 ± 0.5 1.5 ± 0.4
Groups 12 days 15 days 21 days 30 days 60 days 90 days
ML thickness (lm)
Conta 86 ± 3.2 152 ± 7.3 170 ± 3 187 ± 8.1 198 ± 7.2 181 ± 14
DLT-Ib 41 ± 2.9* 105 ± 6.2* 146 ± 7.7 180 ± 14.8 174 ± 9.4 182 ± 18.5
DLT-IIc 53 ± 3.1* 122 ± 7.3 150 ± 3 173 ± 12.6 184 ± 13.4 181 ± 15.9
PCL thickness (lm)
Conta 20 ± 0.6 21 ± 1.2 23 ± 0.7 23 ± 1.3 22 ± 0.5 21 ± 0.7
DLT-Ib 26 ± 0.7** 27 ± 0.8** 27 ± 0.4 25 ± 0.5 25 ± 0.6 23 ± 0.5
DLT-IIc 24 ± 1.1* 25 ± 0.7* 24 ± 1.1 24 ± 1.2 24 ± 0.4 23 ± 0.6
IGL thickness (lm)
Conta 97 ± 11.5 122 ± 11.5 132 ± 9.2 135 ± 10.6 140 ± 13 150 ± 21.3
DLT-Ib 67 ± 11.7* 83 ± 9.1* 120 ± 10.4 125 ± 9.2 130 ± 11.5 145 ± 17.3
DLT-IIc 70 ± 12.2* 102 ± 8.2 117 ± 11.3 130 ± 11.4 135 ± 15 146 ± 19.5
Values are expressed as mean ± SEM (30 readings from 3 pups/parameter/time point)
EGL external granular layer, ML molecular layer, PCL Purkinje cell layer, IGL internal granular layer
* p \ 0.05 for DLT-I and DLT-II versus control; ** p \ 0.01 for DLT-I and DLT-II versus control (one-way ANOVA followed by post hoc
Tukey’s test for multiple comparison among groups)a Vehicle treated (equal volume of DMSO only) groupb DLT dissolved in DMSO at a dose of 0.75 mg/kg body weight at GD 7–10c DLT dissolved in DMSO at a dose of 0.75 mg/kg body weight at GD 11–14
Cell Mol Neurobiol (2013) 33:731–746 737
123

activated morphology were also observed in the granule cell
layer of both the treated groups as compared with their age
matched controls (Fig. 7b, c, e, f, h, i, k, l). Upregulation of
GFAP both in the Bergmann glial fibres and the astrocytes
were indicating the sign of astrogliosis following DLT-
exposure.
Phenotypic Maturation and Dendritogenesis of Purkinje
Neurons
Purkinje cell morphogenesis and dendritogenesis in the
developing cerebella of rats following maternal exposure
of DLT was studied by using rapid Golgi technique. Pre-
natally formed Purkinje neurons undergo a series of mor-
phogenetic changes during early postnatal life to become
mature. Such changes include their dispersion and align-
ment in a monolayer, appearance of a transient apical
growth cone and subsequent appearance of apical dendrites
invading the developing molecular layer. The primary
dendrite arbourizes and forms secondary and tertiary
branches with subsequent formation of spines that prolif-
erate in great numbers. Such changes were clearly seen in
the developing cerebella of control rat pups during post-
natal life, PND7 to PND21 days. The Purkinje cells were
aligned in a monolayer by the end of the 1st week and
started extending their primary dendrites in the ML. By
postnatal day 12, a well developed dendritic arbour with
primary, secondary and tertiary branches with spines were
observed (Fig. 8a). This dendritic arbour was further
elaborated to take up a rectangular area within the
molecular layer (Fig. 8d, g, j).
However, the Purkinje cells in the cerebella of rat pups
born to mothers exposed to DLT during two different
gestational periods possessed marked somal and dendritic
abnormalities. In both the DLT-treated groups, an incon-
sistent and non-uniform pattern of abnormal Purkinje cell
arrangement was observed in either or both ML and IGL.
At sites, the cells were present in clusters with their cell
soma closely adhering each other. Such clustering was
more often in 12- and 15-day-old treated preparations
(Fig. 8b) and were not at all seen once the rat pups became
mature, i.e. by PND30. Purkinje cells possessing more than
one somal process (extrasomal processes) with secondary
and tertiary dendrite like processes was a common occur-
rence (Fig. 8b, c, f). In addition, the Purkinje cell somas
with their dendrites lying parallel to the pial surface or
drooping towards the IGL (Fig. 8b, c, e, f, h) was also a
frequent feature following DLT exposure in contrast to the
straight Purkinje cell arbour, perpendicular to the pial
surface in age matched controls (Fig. 8a, d, g, j).
A very prominent impact of DLT exposure was
observed in the Purkinje cells dendritic arbourization. In
0
9
18
27
36
45
0d 3d 7d 12d 15d 21d 30d
EG
L T
hic
knes
s (µ
m)
Postnatal age (days)
Cont
DLT-I
DLT-II
*
**
**
a
0
50
100
150
200
250
12d 15d 21d 30d 60d 90d
ML
Th
ickn
ess
(µm
)
Postnatal age (days)
Cont
DLT-I
DLT-II**
*
b
0
5
10
15
20
25
30
12d 15d 21d 30d 60d 90d
PC
L T
hic
knes
s (µ
m)
Postnatal age (days)
Cont
DLT-I
DLT-II
** **
**
c
0
30
60
90
120
150
180
12d 15d 21d 30d 60d 90dIG
L T
hic
knes
s (µ
m)
Postnatal age (days)
Cont
DLT-I
DLT-II
**
*
d
Fig. 4 Graph represents the thickness (lm) of various cerebellar
cortical layers. Data is presented as the mean ± SEM of the 30
readings from 3 pups/parameter/time point in control, DLT-I and
DLT-II groups. *DLT-I and DLT-II versus cont, p \ 0.05; **DLT-I
and DLT-II versus cont, p \ 0.01
738 Cell Mol Neurobiol (2013) 33:731–746
123

15d
j k l m
N O P
21d
q r s
h
3d
a b c
d e f
g i
EGL ML
PCL
IGL
n o p
Cont DLT-IIDLT-I
Fig. 5 Photomicrographs showing reelin immunolabelling in the
cerebellum (vermis) of control, DLT-I and DLT-II group pups
(n = 3). Arrows indicates the reelin positive cells in the EGL (black
arrows) and granule cells in ML and IGL (red arrows). The scale baris 50 lm (Color figure online)
Cell Mol Neurobiol (2013) 33:731–746 739
123

both the treated groups, there was a clear cut reduction in
the total rectangular area occupied by the dendritic arbour
in the molecular layer as compared with their age-matched
controls. This was clearly evident from the morphometric
studies for the measurement of length and width of the
dendritic arbour. A significant (p \ 0.01; 0.05) reduction in
both the length and width was found at all time points
studied in both the treated groups as compared with their
age-matched control (Table 4; Fig. 9b, c). In controls, the
Purkinje cells presented normal morphology and extensive
dendritic arbourization with primary, secondary and ter-
tiary branches with spiny branchlets extending into the
molecular layer up to the pial surface (Fig. 8a, d, g, j).
However, in both the treated groups the primary and sec-
ondary dendrites were quite prevalent, but tertiary branches
and spiny branches were remarkably reduced or absent.
Many Purkinje cells in all the parameters of treated
groups had long main dendrites without branching, in many
cases acquiring ‘S’ form rather than straight perpendicular
to the pial surface (Fig. 8h). Even the primary and
secondary dendrites were thick and stumpy and at times not
extending up to the pial surface (Fig. 8b, c, k) rather
swaying away from the pial surface (Fig. 8b, c, h). Such
changes were consistently seen throughout the period of
study, i.e. from PND12 to PND90 (Fig. 8b, c, e, f, h, i, k, l)
when the animals attain reproductive maturity.
Dendritic spine density was another feature that was
significantly affected in pups born to DLT exposed moth-
ers. In controls there was a gradual and linear increase in
the spine density with increasing age, i.e. in PND12 to
PND90 from 16.7 ± 0.5 to 22.3 ± 0.55. However, in both
the treated groups the increase in the number of spines was
much less and the spine density was significantly reduced
at all postnatal days studied. The data has been shown in
Table 4 and Fig. 9a.
Motor Coordination
Impact of maternal DLT exposure on the acquisition of
motor coordination was assessed as the total length of time
spent by the animals on the rotating rod of the rotarod. In
the accelerating rotarod test from 2 rpm to 40 rpm for
420 s, the latency to fall was lying between 27 ± 2.3 and
37.3 ± 2.5 s in PND21 to PND90 control rat pups,
respectively. However, the animals from both the treated
groups fell faster from the rotating rotarod leading to sig-
nificantly lower latency to fall at all the time points
(p \ 0.01; 0.05) and is expressed in Table 5 and Fig. 10.
Discussion
Developing cerebellum in neonates is a useful system to
study the basic mechanisms that regulates proliferation,
migration and differentiation of neuronal precursors
required for the histogenesis and the formation of proper
cytoarchitecture. The cerebellar development is complete
Table 3 Area fraction of reelin immunoreactivity in the cerebellar cortex during early postnatal life following maternal exposure to deltamethrin
during gestation
Group 0 day 3 days 7 days 12 days 15 days 21 days
Area fraction (%)
Conta 27 ± 11.02 29 ± 0.5 27 ± 1.04 25 ± 2.5 14 ± 0.65 3.9 ± 0.6
DLT-Ib 33 ± 0.5* 39 ± 0.7**, ## 31.8 ± 0.3* 36 ± 2.6**, # 27 ± 2.5** 5.9 ± 0.67
DLT-IIc 31 ± 0.34* 32 ± 0.6 37 ± 0.5**, # 29 ± 2.5 28 ± 2.9** 7.7 ± 0.5*
Values are expressed as mean ± SEM (30 readings from 3 pups/parameter/time point)
* p \ 0.05 for DLT-I and DLT-II versus control; ** p \ 0.01 for DLT-I and DLT-II versus control; # p \ 0.05 for DLT-I versus DLT-II;## p \ 0.05 for DLT-I versus DLT-II (one-way ANOVA followed by post hoc Tukey’s test for multiple comparison among groups)a Vehicle treated (equal volume of DMSO only) groupb DLT dissolved in DMSO at a dose of 0.75 mg/kg body weight at GD 7–10c DLT dissolved in DMSO at a dose of 0.75 mg/kg body weight at GD 11–14
0
5
10
15
20
25
30
35
40
45
0d 3d 7d 12d 15d 21d
Are
a fr
acti
on
(%
)
Postnatal age (days)
ContDLT-IDLT-II
**
** ** **
****
## #
*
#
*
Fig. 6 Area fraction of reelin immunoreactivity presented as the
mean ± SEM control, DLT-I and DLT-II groups. Control, DLT-I and
DLT-II groups (n = 3) at various postnatal ages. *DLT-I and DLT-II
versus cont, p \ 0.05, **DLT-I and DLT-II versus cont, P \ 0.01,#DLT-I versus DLT-II, p \ 0.05, ##DLT-I versus DLT-II, p \ 0.01
740 Cell Mol Neurobiol (2013) 33:731–746
123

by the third postnatal week in rodents. Granule cells, the
major excitatory neuron population of cerebellum are
generated postnatally in the EGL and migrate through
contact-guided migration along the Bergmann glial pro-
cesses (Rakic 1977; Hatten et al. 1997; Patro and Patro
2005). The Purkinje neurons generated prenatally from the
germinal zone of the fourth ventricle migrate towards the
cerebellar surface and settle into a monolayer. Reelin, a
large secreted protein is crucial for such migration and
laminar arrangement of neurons in the neocortex, hippo-
campus, cerebellum and spinal cord (D’Arcangelo et al.
1995; Ogawa et al. 1995; Tissir and Goffinet 2003; Sinagra
et al. 2008; Forster et al. 2010). The functions of reelin are
mediated through its receptors which trigger a complex
signalling cascade (Herz and Chen 2006).
In the present study, a significant overexpression of re-
elin was observed following prenatal exposure to DLT both
in the cells of EGL and IGL at all time points studied. In
addition, an ectopic expression of reelin was also observed
in the EGL as well as in the migrating granule cells just
below the EGL, indicating an arrest of granule cell
migration in this zone. Such reelin overexpression fol-
lowing DLT exposure suggests its intracellular accumula-
tion, either due to the failure of its release or blockade of its
Cont DLT-IIDLT-I
7d
12d
15d
21d
a b c
d e f
g h i
j k l
MLPCL
IGL
Fig. 7 Photomicrographs showing GFAP immunohistochemistry on
sagittal sections from the cerebellar vermal region of control, DLT-I
and DLT-II group (n = 3). Arrows (black) represents the cresyl violet
stained granule cells in EGL, arrowheads (red) represents GFAP
positive Bergmann glial fibres and arrows (red) represents GFAP
positive astrocytes in IGL. The scale bar is 50 lm (Color figure online)
Cell Mol Neurobiol (2013) 33:731–746 741
123

secretion from the granule cells. Such intracellular accu-
mulation accounts for an increase in its density and strong
immunopositivity near the EGL. Reelin expressing cere-
bellar granule cells both synthesize and secrete reelin,
blockade of its secretion or release interferes with the reelin
signalling and is thus directly responsible for the abnormal
and mis-oriented migration of granule cells. If such chan-
ges are dependent or secondary, the neurotoxic effects
imposed by DLT through voltage sensitive sodium chan-
nels need to be studied. In the absence of reelin signalling,
the migrating granule cells aggregate below the EGL due to
the arrest of their migration. Such cellular traffic jam was
also reported in Dab 1 mutants (Olson et al. 2006) and
reeler mouse (Pinto-Lord et al. 1982). Olson et al. (2006)
believed that the cells superficial to migrating neurons
obstruct further migration and thus result in the formation
12d
Cont DLT-IIDLT-I
21d
30d
90d
ba c
d fe
g ih
j lk
Fig. 8 Photomicrographs showing rapid Golgi stained sagittal sections from the cerebellar vermal region of control, DLT-I and DLT-II group
(n = 3). Arrows indicates the Golgi stained Purkinje neurons with distorted morphology. The scale bar is 50 lm
742 Cell Mol Neurobiol (2013) 33:731–746
123

of the ectopic zone. Similar results has also been recorded
with the transgenic mouse model where the reelin over-
expression in adult brain was found to alter the pattern of
neuronal migration of newly generated neurons in the SVZ
leading to the formation of an ectopic zone during adult
neurogenesis. A reduced number of granule cells in the
granule cell layer were also observed as a result of the
arrest of migration of the newly formed neurons (Pujadas
et al. 2010). In an experiment on cultured hippocampal
cells, blockage of reelin secretory pathway with brefeldin
A (BFA) perturbs the homeostasis of NMDA receptors
(NMDARs, Campo et al. 2009) as a result of the lack of
continuous secretion. Such defects are believed to play a
major role in the development of neuropsychiatric disor-
ders such as schizophrenia (Mohn et al. 1999; Guidotti
et al. 2000).
Our results further find support from a more recent study
of Duveau et al. (2011) where they reported in a model of
temporal lobe epilepsy induced by intra-hippocampal kai-
nic acid injection that the strong reelin immunopositivity of
the Cajal-Retzius cells was due to intracellular accumula-
tion resembling the morphology of these cells in reelin
Orleans (relnorl/orl) mice, which express the secretion-
deficient 310-kDa reelin fragment. Such intracellular
accumulation of reelin was explained due to the absence of
signal peptide for secretion located at the C-terminus (Rice
and Curran 2001).
Bergmann glial fibres act as specific guide for the cer-
ebellar granule cell migration were also found to be mis-
aligned, hypertrophied and strongly GFAP labelled,
indicating astrogliosis following DLT exposure during
prenatal life. Such defects in Bergmann glial processes
were also reported to affect granule cell migration conse-
quent upon DLT exposure during early postnatal life (Patro
and Patro 2005). Radial glial cells have also been reported
to be specifically vulnerable to a variety of prenatal insults,
viz. alcohol (Miller and Robertson 1993), methylmercury
(Choi et al. 1996), ionising radiation (Roper et al. 1997)
and methylazoxymethanol (MAM, Spalice et al. 2009).
Reelin secretion defects observed in this investigation
following DLT exposure could also be correlated with
Bergmann glia mis-orientation and hypertrophy as reelin
has been reported to promote extension and maintenance of
radial glial scaffold in the hippocampus of reeler mutant
mouse when exposed to reelin exogenously (Frotscher
et al. 2003). Lack of reelin has been found to result in
aberrant migration of cortical neuron and misalignment
radial glial fibres (Hunter-Schaedle 1997; Hartfuss et al.
2003). Any disruption in radial glial scaffold causes
migrational disorders leading to mental retardation, epi-
lepsy, schizophrenia, autism, etc. (Pang et al. 2008; Spalice
et al. 2009; Verrotti et al. 2009).
An ectopic Purkinje cell arrangement with marked so-
mal and dendritic abnormalities in the DLT treated cere-
bella of both the treated groups were also a prominent
observation of this study. Purkinje cell possessing extra-
somal processes, drooping dendritic tree, a significantly
reduced and stunted dendritic arbour and spine density as
seen through Golgi studies, was a common occurrence.
Reelin being a major component mediating the migration
Table 4 Purkinje cell spine density, and width and length of the dendritic arbour (lm) in the cerebellum of developing rats following maternal
exposure to deltamethrin during gestation
Groups 12 days 15 days 21 days 30 days 60 days 90 days
Spine density/10 lm of dendrite
Conta 16.7 ± 0.5 18.7 ± 0.9 20.7 ± 0.4 20.5 ± 0.6 21.7 ± 0.6 22.3 ± 0.5
DLT-Ib 11.9 ± 1.2* 14.8 ± 0.6* 17.5 ± 1.2* 17.7 ± 1.1* 18.2 ± 0.5* 20.1 ± 0.6*
DLT-IIc 14.1 ± 0.5* 15 ± 0.6* 19 ± 0.9* 17 ± 1.3* 19.6 ± 0.9 20.2 ± 0.7*
Width of the dendritic arbour (lm)
Conta 85.5 ± 0.5 106 ± 1.5 135.2 ± 3.3 131.8 ± 3.5 134. ± 2.3 146.5 ± 2.3
DLT-Ib 52.7 ± 3.4** 70 ± 4.5** 99.2 ± 3.6** 97.7 ± 4.8* 98.3 ± 4.3** 97.5 ± 3.6**
DLT-IIc 61 ± 3.6** 79.5 ± 4.4** 104 ± 3.6** 102.7 ± 5.7* 91.2 ± 4.8** 97.5 ± 2.9**
Length of the dendritic arbour (lm)
Conta 101.5 ± 1.6 128 ± 2.2 163 ± 2.9 171.2 ± 2.8 173.1. ± 3.5 170 ± 2.6
DLT-Ib 70 ± 3.5** 91.5 ± 3.5** 116.2 ± 4.8** 141.4 ± 2.9** 151 ± 2.9* 141.9 ± 4.5**
DLT-IIc 75.7 ± 4** 102 ± 5.1** 125.7 ± 4.6** 131.8 ± 4.3** 147.6 ± 4.8* 130.6 ± 5.3**
Values are expressed as mean ± SEM (from 3 pups/parameter/time point)
* p \ 0.05 for DLT-I and DLT-II versus control; ** p \ 0.01 for DLT-I and DLT-II versus control (one-way ANOVA followed by post hoc
Tukey’s test for multiple comparison among groups)a Vehicle treated (equal volume of DMSO only) groupb DLT dissolved in DMSO at a dose of 0.75 mg/kg body weight at GD 7–10c DLT dissolved in DMSO at a dose of 0.75 mg/kg body weight at GD 11–14
Cell Mol Neurobiol (2013) 33:731–746 743
123

and positioning of the Purkinje cell, defective reelin sig-
nalling is possibly responsible for their ectopic arrange-
ment and much affected dendritogenesis and spine density.
Reelin is known to exert its function by binding to and
activating its very low-density lipoprotein receptors
(VLDLR) and ApoER2 (Trommsdorff et al. 1999; Herz
and Chen 2006) on the surface of the Purkinje cells. The
abnormalities associated with the dendritic arbourization
and spine formation following DLT exposure could be a
secondary defect resulting from the misalignment and
ectopic arrangement of the Purkinje cells. Dendritic
arbours are also substantially stunted and disorganised in
reeler mice (Tabata and Nakajima 2002; Niu et al. 2004),
scrambler mutant mice (Rice and Curran 1999), Cend1
knock-out mice (Sergaki et al. 2010) and Dab-1 suppressed
cortical neurons (Olson et al. 2006), many of them directly
or indirectly influencing the reelin pathway.
Impaired cerebellar lamination and morphological
defects in Purkinje neurons are directly associated with the
functional impairment related to motor coordination. Our
findings with rotarod test suggests that the latency to fall of
the rotating rod was significantly less in the DLT treated
animals even at adulthood clearly, suggesting that the
structural abnormalities due to defective reelin expression
possibly directly affects the cerebellar functional output.
However, the role of reelin in developmental neurotoxicity
0
20
40
60
80
100
120
140
160
12d 15d 21d 30d 60d 90d
Wid
th o
f th
e d
end
riti
c ar
bo
r (µ
m)
Postnatal age (days)
Cont
DLT-I
DLT-II
****
********
**
** ** ***
*
b
0
40
80
120
160
200
12d 15d 21d 30d 60d 90dLen
gth
of
den
dri
tic
arb
or
(µm
)
Postnatal age (days)
Cont
DLT-I
DLT-II
*
*
**
** **
****
****
**
**
**
c
0
5
10
15
20
25
12d 15d 21d 30d 60d 90dSp
ine
den
sity
/10µ
m o
f d
end
rite
Postnatal age (days)
Cont
DLT-I
DLT-II
** * **
***
*
**
a
Fig. 9 Graph represents the measurement (mean ± SEM) of the
Spine density (a), width of the dendritic arbour (b) and length of the
dendritic arbour (c) of Purkinje neurons in control, DLT-I and DLT-II
group animals (n = 3). *DLT-I and DLT-II versus cont, p \ 0.05,
**DLT-I and DLT-II versus cont, p \ 0.01
Table 5 Motor coordination impairment in the developing rat pups
following maternal exposure to deltamethrin during gestation
Groups 21 days 30 days 60 days 90 days
Latency to fall (s) from the accelerating rotarod
Conta 27.2 ± 2.3 34.8 ± 1.9 37.0 ± 2.1 37.3 ± 2.5
DLT-Ib 12.1 ± 1.9** 23.0 ± 2.3** 16.1 ± 1.3** 21.0 ± 1.2**
DLT-IIc
16.4 ± 1.4* 24.9 ± 1.9 20.8 ± 1.6* 17.1 ± 1.3**
Values are expressed as mean ± SEM (n = 6)
* p \ 0.05 for DLT-I and DLT-II versus control; ** p \ 0.01 for DLT-Iand DLT-II versus control (one-way ANOVA followed by post hocTukey’s test for multiple comparison among groups)a Vehicle treated (equal volume of DMSO only) groupb DLT dissolved in DMSO at a dose of 0.75 mg/kg body weight at GD7–10c DLT dissolved in DMSO at a dose of 0.75 mg/kg body weight at GD11–14
0
9
18
27
36
45
21d 30d 60d 90d
Lat
ency
to
dro
p (
sec)
Postnatal age (days)
Cont
DLT-I
DLT-II
*
**
**** **
***
Fig. 10 Rotarod performance of the control, DLT-I and DLT-II
group animals (n = 6), presented as the mean ± SEM of the time
spent on the rotating rod. *DLT-I and DLT-II versus cont, p \ 0.05,
**DLT-I and DLT-II versus cont, p \ 0.01
744 Cell Mol Neurobiol (2013) 33:731–746
123

of environmental agents has not been explored till date,
while its importance in neurodegeneration and b-amyloid
induced synaptic depression was reported in Alzheimer’s
disease (Beffert et al. 2006; Botella-Lopez et al. 2006;
Durakoglugil et al. 2009; Knuesel et al. 2009), suggesting
its role in the pathogenesis of neurological diseases. Thus,
it is proposed that DLT at a dose as low as 1/200th of the
LD50 which could be close to the environmental exposure
to the pregnant women due to their occupational or resi-
dential proximity to such insecticide treated farmlands
causes the defects in neuronal migration and subsequent
lamina formation through reelin by its overexpression and/
or blockade of its release and signalling.
Acknowledgments Financial support from the Indian Council of
Medical Research, New Delhi is thankfully acknowledged. Kamendra
Kumar is a Department of Biotechnology (DBT) Senior Research
Fellow. Facilities developed through the DBT-Human Resource
Development and Bioinformatics Infrastructural facility through
Department of Biotechnology Grants used in this study are thankfully
acknowledged.
Conflict of interest Authors have no conflict of interest.
References
Altman J, Bayer S (1995) Atlas of prenatal brain development. CRC
Press, Boca Raton
Amweg EL, Weston DP, Ureda NM (2005) Use and toxicity of
pyrethroid pesticides in the Central Valley, California, USA.
Environ Toxicol Chem 24(4):966–972
Beffert U, Weeber EJ, Durudas A, Qiu S, Masiulis I, Sweatt JD, Li
WP, Adelmann G, Frotscher M, Hammer RE, Herz J (2005)
Modulation of synaptic plasticity and memory by reelin involves
differential splicing of the lipoprotein receptor Apoer2. Neuron
47(4):567–579
Beffert U, Durudas A, Weeber EJ, Stolt PC, Giehl KM, Sweatt JD,
Hammer RE, Herz J (2006) Functional dissection of reelin
signaling by site-directed disruption of disabled-1 adaptor binding
to apolipoprotein E receptor 2: distinct roles in development and
synaptic plasticity. J Neurosci 26(7):2041–2052
Berkowitz GS, Obel J, Deych E, Lapinski R, Godbold J, Liu Z,
Landrigan PJ, Wolff MS (2003) Exposure to indoor pesticides
during pregnancy in a multiethnic, urban cohort. Environ Health
Perspect 111(1):79–84
Botella-Lopez A, Burgaya F, Gavin R, Garcia-Ayllon MS, Gomez-
Tortosa E, Pena-Casanova J, Urena JM, Del Rio JA, Blesa R,
Soriano E, Saez-Valero J (2006) Reelin expression and glyco-
sylation patterns are altered in Alzheimer’s disease. Proc Natl
Acad Sci USA 103(14):5573–5578
Campo CG, Sinagra M, Verrier D, Manzoni OJ, Chavis P (2009)
Reelin secreted by GABAergic neurons regulates glutamate
receptor homeostasis. PLoS ONE 4(5):e5505
Chizhikov VV, Lindgren AG, Currle DS, Rose MF, Monuki ES,
Millen KJ (2006) The roof plate regulates cerebellar cell-type
specification and proliferation. Development 133(15):2793–2804
Choi BH, Yee S, Robles M (1996) The effects of glutathione
glycoside in methyl mercury poisoning. Toxicol Appl Pharmacol
141(2):357–364
D’Arca D, Zhao X, Xu W, Ramirez-Martinez NC, Iavarone A,
Lasorella A (2010) Huwe1 ubiquitin ligase is essential to
synchronize neuronal and glial differentiation in the developing
cerebellum. Proc Natl Acad Sci USA 107(13):5875–5880
D’Arcangelo G, Miao GG, Chen SC, Soares HD, Morgan JI, Curran T
(1995) A protein related to extracellular matrix proteins deleted
in the mouse mutant reeler. Nature 374(6524):719–723
Durakoglugil MS, Chen Y, White CL, Kavalali ET, Herz J (2009)
Reelin signaling antagonizes beta-amyloid at the synapse. Proc
Natl Acad Sci USA 106(37):15938–15943
Duveau V, Madhusudan A, Caleo M, Knuesel I, Fritschy JM (2011)
Impaired reelin processing and secretion by Cajal–Retzius cells
contributes to granule cell dispersion in a mouse model of
temporal lobe epilepsy. Hippocampus 21(9):935–944
Forster E, Bock HH, Herz J, Chai X, Frotscher M, Zhao S (2010)
Emerging topics in reelin function. Eur J Neurosci
31(9):1511–1518
Frotscher M (2010) Role for reelin in stabilizing cortical architecture.
Trends Neurosci 33(9):407–414
Frotscher M, Haas CA, Forster E (2003) Reelin controls granule cell
migration in the dentate gyrus by acting on the radial glial
scaffold. Cereb Cortex 13(6):634–640
Gohlke JM, Armant O, Parham FM, Smith MV, Zimmer C, Castro
DS, Nguyen L, Parker JS, Gradwohl G, Portier CJ, Guillemot F
(2008) Characterization of the proneural gene regulatory
network during mouse telencephalon development. BMC Biol
6:15
Guidotti A, Auta J, Davis JM, Di-Giorgi-Gerevini V, Dwivedi Y,
Grayson DR, Impagnatiello F, Pandey G, Pesold C, Sharma R,
Uzunov D, Costa E (2000) Decrease in reelin and glutamic acid
decarboxylase67 (GAD67) expression in schizophrenia and
bipolar disorder: a postmortem brain study. Arch Gen Psychiatr
57(11):1061–1069
Hartfuss E, Forster E, Bock HH, Hack MA, Leprince P, Luque JM,
Herz J, Frotscher M, Gotz M (2003) Reelin signaling directly
affects radial glia morphology and biochemical maturation.
Development 130(19):4597–4609
Hatten ME, Alder J, Zimmerman K, Heintz N (1997) Genes involved
in cerebellar cell specification and differentiation. Curr Opin
Neurobiol 7(1):40–47
Herz J, Chen Y (2006) Reelin, lipoprotein receptors and synaptic
plasticity. Nat Rev Neurosci 7(11):850–859
Heudorf U, Angerer J, Drexler H (2004) Current internal exposure to
pesticides in children and adolescents in Germany: urinary levels
of metabolites of pyrethroid and organophosphorus insecticides.
Int Arch Occup Environ Health 77(1):67–72
Hevner RF (2008) Reelin and the cerebellum. In: Fatemi SH (ed)
Reelin glycoprotein. Springer, Heidelberg, pp 141–158
Hevner RF, Hodge RD, Daza RA, Englund C (2006) Transcription
factors in glutamatergic neurogenesis: conserved programs in
neocortex, cerebellum, and adult hippocampus. Neurosci Res
55(3):223–233
Hibi T, Mizutani M, Baba A, Hattori M (2009) Splicing variations in
the ligand-binding domain of ApoER2 results in functional
differences in the binding properties to reelin. Neurosci Res
63(4):251–258
Hunter-Schaedle KE (1997) Radial glial cell development and
transformation are disturbed in reeler forebrain. J Neurobiol
33(4):459–472
Knuesel I, Nyffeler M, Mormede C, Muhia M, Meyer U, Pietropaolo
S, Yee BK, Pryce CR, LaFerla FM, Marighetto A, Feldon J
(2009) Age-related accumulation of reelin in amyloid-like
deposits. Neurobiol Aging 30(5):697–716
Matsuki T, Pramatarova A, Howell BW (2008) Reduction of Crk and
CrkL expression blocks reelin-induced dendritogenesis. J Cell
Sci 121(Pt 11):1869–1875
Cell Mol Neurobiol (2013) 33:731–746 745
123

Miller MW, Robertson S (1993) Prenatal exposure to ethanol alters
the postnatal development and transformation of radial glia to
astrocytes in the cortex. J Comp Neurol 337(2):253–266
Mohn AR, Gainetdinov RR, Caron MG, Koller BH (1999) Mice with
reduced NMDA receptor expression display behaviors related to
schizophrenia. Cell 98(4):427–436
Narahashi T (2000) Neuroreceptors and ion channels as the basis for
drug action: past, present, and future. J Pharmacol Exp Ther
294(1):1–26
Niu S, Renfro A, Quattrocchi CC, Sheldon M, D’Arcangelo G (2004)
Reelin promotes hippocampal dendrite development through the
VLDLR/ApoER2-Dab1 pathway. Neuron 41(1):71–84
Niu S, Yabut O, D’Arcangelo G (2008) The reelin signaling pathway
promotes dendritic spine development in hippocampal neurons.
J Neurosci 28(41):10339–10348
Ogawa M, Miyata T, Nakajima K, Yagyu K, Seike M, Ikenaka K,
Yamamoto H, Mikoshiba K (1995) The reeler gene-associated
antigen on Cajal–Retzius neurons is a crucial molecule for
laminar organization of cortical neurons. Neuron 14(5):899–912
Olson EC, Kim S, Walsh CA (2006) Impaired neuronal positioning
and dendritogenesis in the neocortex after cell-autonomous Dab1
suppression. J Neurosci 26(6):1767–1775
Pang T, Atefy R, Sheen V (2008) Malformations of cortical
development. Neurologist 14(3):181–191
Patro N, Patro IK (2005) Effects of deltamethrin on granule cell
migration during postnatal development of rat cerebellum.
Indian J Exp Biol 43(2):158–162
Patro N, Mishra S, Chattopadhyay M, Patro I (1997) Neurotoxico-
logical effects of deltamethrin on the postnatal development of
cerebellum of rat. J Biosci 22(2):117–130
Patro N, Shrivastava M, Tripathi S, Patro IK (2009) S100beta
upregulation: a possible mechanism of deltamethrin toxicity and
motor coordination deficits. Neurotoxicol Teratol 31(3):169–176
Perez-Garcia CG, Tissir F, Goffinet AM, Meyer G (2004) Reelin
receptors in developing laminated brain structures of mouse and
human. Eur J Neurosci 20(10):2827–2832
Pinto-Lord MC, Evrard P, Caviness VS Jr (1982) Obstructed neuronal
migration along radial glial fibers in the neocortex of the reeler
mouse: a Golgi–EM analysis. Brain Res 256(4):379–393
Pujadas L, Gruart A, Bosch C, Delgado L, Teixeira CM, Rossi D, de
Lecea L, Martinez A, Delgado-Garcia JM, Soriano E (2010)
Reelin regulates postnatal neurogenesis and enhances spine
hypertrophy and long-term potentiation. J Neurosci 30(13):
4636–4649
Rakic P (1977) Genesis of the dorsal lateral geniculate nucleus in the
rhesus monkey: site and time of origin, kinetics of proliferation,
routes of migration and pattern of distribution of neurons.
J Comp Neurol 176(1):23–52
Ramos-Moreno T, Galazo MJ, Porrero C, Martinez-Cerdeno V,
Clasca F (2006) Extracellular matrix molecules and synaptic
plasticity: immunomapping of intracellular and secreted reelin in
the adult rat brain. Eur J Neurosci 23(2):401–422
Ray DE, Fry JR (2006) A reassessment of the neurotoxicity of
pyrethroid insecticides. Pharmacol Ther 111(1):174–193
Rice DS, Curran T (1999) Mutant mice with scrambled brains:
understanding the signaling pathways that control cell position-
ing in the CNS. Genes Dev 13(21):2758–2773
Rice DS, Curran T (2001) Role of the reelin signaling pathway in
central nervous system development. Annu Rev Neurosci
24:1005–1039
Roper SN, Abraham LA, Streit WJ (1997) Exposure to in utero
irradiation produces disruption of radial glia in rats. Dev
Neurosci 19(6):521–528
Sergaki MC, Guillemot F, Matsas R (2010) Impaired cerebellar
development and deficits in motor coordination in mice lacking
the neuronal protein BM88/Cend1. Mol Cell Neurosci
44(1):15–29
Shafer TJ, Meyer DA, Crofton KM (2005) Developmental neurotox-
icity of pyrethroid insecticides: critical review and future
research needs. Environ Health Perspect 113(2):123–136
Sinagra M, Gonzalez Campo C, Verrier D, Moustie O, Manzoni OJ,
Chavis P (2008) Glutamatergic cerebellar granule neurons
synthesize and secrete reelin in vitro. Neuron Glia Biol
4(3):189–196
Soderlund DM (2010) State-dependent modification of voltage-gated
sodium channels by pyrethroids. Pestic Biochem Physiol
97(2):78–86
Soderlund DM, Clark JM, Sheets LP, Mullin LS, Piccirillo VJ,
Sargent D, Stevens JT, Weiner ML (2002) Mechanisms of
pyrethroid neurotoxicity: implications for cumulative risk
assessment. Toxicology 171(1):3–59
Spalice A, Parisi P, Nicita F, Pizzardi G, Del Balzo F, Iannetti P
(2009) Neuronal migration disorders: clinical, neuroradiologic
and genetics aspects. Acta Paediatr 98(3):421–433
Stensaas LJ (1967) The development of hippocampal and dorsolateral
pallial region of the cerebral hemisphere in fetal rabbits. V. Sixty
millimeter stage, glial cell morphology. J Comp Neurol
131(1967):423–436
Tabata H, Nakajima K (2002) Neurons tend to stop migration and
differentiate along the cortical internal plexiform zones in the
reelin signal-deficient mice. J Neurosci Res 69(6):723–730
Tissir F, Goffinet AM (2003) Reelin and brain development. Nat Rev
Neurosci 4(6):496–505
Trommsdorff M, Gotthardt M, Hiesberger T, Shelton J, Stockinger W,
Nimpf J, Hammer RE, Richardson JA, Herz J (1999) Reeler/
disabled-like disruption of neuronal migration in knockout mice
lacking the VLDL receptor and ApoE receptor 2. Cell
97(6):689–701
Verrotti A, Agostinelli S, Mohn A, Manco R, Coppola G, Franzoni E,
Cerminara C, Parisi P, Iannetti P, Spalice A (2009) Clinical
features of psychogenic non-epileptic seizures in prepubertal and
pubertal patients with idiopathic epilepsy. Neurol Sci 30(4):
319–323
Verschoyle RD, Aldridge WN (1980) Structure-activity relationships
of some pyrethroids in rats. Arch Toxicol 45(4):325–329
Vijverberg HP, van den Bercken J (1990) Neurotoxicological effects
and the mode of action of pyrethroid insecticides. Crit Rev
Toxicol 21(2):105–126
Weeber EJ, Beffert U, Jones C, Christian JM, Forster E, Sweatt JD,
Herz J (2002) Reelin and ApoE receptors cooperate to enhance
hippocampal synaptic plasticity and learning. J Biol Chem
277(42):39944–39952
Wolansky MJ, Gennings C, DeVito MJ, Crofton KM (2009) Evidence
for dose-additive effects of pyrethroids on motor activity in rats.
Environ Health Perspect 117(10):1563–1570
Yoshiki A, Kusakabe M (1998) Cerebellar histogenesis as seen in
identified cells of normal-reeler mouse chimeras. Int J Dev Biol
42(5):695–700
746 Cell Mol Neurobiol (2013) 33:731–746
123