Embed Size (px)
Citation preview

Le mouvement gCotropique des feuilles chez la Sensitive (Mimosapudica) soumise a l'influence d'un agent anesthCsique
FncirltP des Scietzces d e Poitiers, U.E.R. rles Scietlces Fot~rlrrtnet~trrles et AppliqiiPes, Laborntoire rlr Physiologic 13PgPtrrle (Ser18ic.e rlrr Cl~rotlophytotrot~),
Smriotl Biologiqre d e Benu-Site, 25, Frtitbolrrg S(1itlt Cyprietl, F-86000, Poitiers
R e ~ u le 6 mai 1976
GAILLOCHET, J., et P. FLEURAT-LESSARD. 1977. Le mouvement geotropique des feuilles chez la Sensitive (Mimosa prrdicrt) soumise B I'influence d'un agent anesthesique. Can. J . Bot. 55: 70-82.
Chez Mitnosrr prrrlicn L. , le mouvement gkotropique foliaire (mouvement lent) provoqud par I'inversion des plantes dans le champ gravifique, est inhibe de f a ~ o n reversible par I'ether kthylique utilisk aux mkmes degres d'ethirisation qui produisent chez cette plante I'inhibition reversible des mouvements seismonastiques foliaires (mouvements rapides) et chez d'autres organismes. I'inhibition de differentes fonctions cellulaires telles que photosynthtse. caryo- cintse et luminescence bactirienne.
Le degre d'inhibition du mouvement geotropique foliaire est lie a la concentration, B la duree d'action et au moment d'admission de I'Cther, ainsi qu'au type de l'organe moteur considere. Les risultats obtenus permettent de tirer les conclusions suivantes: une etherisation prealable des plantes en position verticale diminue la vitesse du mouvement geotropique; I'introduction de I'Cther au cours du developpement de la reaction peut bloquer le mouvement dans certaines conditions. Enfin, le mouvement geotropique des pulvinus primaires est plus inhibe que celui des pulvinus secondaires, I'inverse se produisant pour I'inhibition des mouvements ~Cismonastiques.
La contribution de ces resultats, a la connaissance du geotropisme foliaire, fait l'objet de la discussion.
GAILLOCHET. J., and P. FLEURAT-LESSARD. 1977. L e mouvement geotropique des feuilles chez la Sensitive (Mit~~osriprrrlicrr) soumise a I'influence d'un agent anesthesique. Can. J . Bot. 55: 70-82.
In Mitnosu prrdicrr L., the geotropic movement (a slow variety of leaf movement) induced by inverting plants in the gravific field is reversibly inhibited by diethyloxide used in a range of concentrations producing, reversibly, in the same plant, inhibition of the seismonastic leaf movement (a quick variety of leaf movement) and in other organisms, inhibition of various cellularfunctions such as photosynthesis, caryocinesis, and bacterial luminescence.
The degree of inhibition of geotropic leaf movement is dependent upon the concentration of ether, the duration of etherization, the time of anaesthetic admission, and the sequence of the motor organs. Results led to the following conclusions: preliminary etherization of plants. in vertical position. decreases rate of the geotropic movement; etherization occurring during the course of this movement can stop it in certain conditions; geotropic movement is more inhibited in primary than in secondary motor organs. whereas the opposite occurs in seismonastic move- ment.
The discussion deals with the contribution of these results to the knowledge of leafgeotropism.
Introduction On sait depuis Bonnet (1779) que les feuilles
dCplacCes de leur direction naturelle, par rapport au champ normal de la pesanteur, sont le sitge de mouvements lents de compensation posturale. Par la suite. des etudes ~recises ont CtC effectuees sur la gCorCactivitC des feuilles notamment chez Phaseolus vulgaris (Arslan 1949 ; Brauner et Rau 1966) et chez Mimosa pudiccl L. (Roblin 1968, 1975; Gavaudan et Roblin 1969). Les observa- tions publikes par ces auteurs ont mis en tvi- dence, chez les plantes inversees de 180 degres dans le champ gravifique, une regulation de
posture donnant aux feuilles une position iden- tique a celle qu'elles auraient en position nor- male, compte tenu de la symktrie du renverse- ment. Le developpement de cette regulation prCsente une allure diphasique: une phase nega- tive, au cours de laquelle les feuilles sont in- fluencees par la pesanteur, est suivie d'une phase positive au cours de laquelle les feuilles reagis- sent contre le champ gravifique. Chez Mimosa pudica L., dont la feuille prksente trois types d'articulations (fig. I) correspondant aux organes moteurs appeles pulvinus, primaires (P,), secon- daires (P,) et tertiaires (P, ou pulvinules), ce
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y SA
VA
NN
AH
RIV
NA
TL
AB
BF
on 1
1/13
/14
For
pers
onal
use
onl
y.

GAILLOCHET ET FLEURAT-LESSARD 7 1
mouvement rkversible, de rtgulation posturale geotropique, n'a CtC observC qu'au niveau des articulations primaire et secondaire, l'articula- tion tertiaire ne prCsentant aucune rCaction visible a l'ceil nu.
Dans le cadre de 1'Ctude des propriCtts sen- sorio-motrices chez les vCgCtaux, nous avons analysC le mouvement giotropique foliaire de Sensitives soumises a l'influence de 1'Cther Cthy- lique. Selon C. Bernard (1875), les propriCtCs anesthksiques de cet agent ont CtC fortuitement dkcouvertes sur l'homme en 1818, et sa premiere utilisation en chirurgie aurait CtC rCalisCe en 1842 par W. C. Long, mCdecin B Athtnes. D'apres le Docteur de la Bate (1869), l'usage de 1'Cther dans la pratique mCdicale remonterait 2 1846, date a laquelle le Docteur Morton (1847),
FIG. 1. Schema d'une feuille bifoliolee de Mimosa pridica L. comportant la terminologie des valeurs angu- laires mesurtes. & est la valeur de I'angle compris entre la tige et le petiole, I? zero du rapporteur etant dirigC vers la base de la plante. (1 represente la valeur de I'angle form6 A I'intersection du plan contenant le petiole (plan perpen- diculaire a la tige pour & = 90") et du plan ges folioles Ctalks. 7 est I'angle compris entre les rachis. 6 indique le degrk d'ouverture des foliolules. PI, P, et P, sont les organes moteurs de premier ordre (pulvinus primaires), de second ordre (pulvinus secondaires) et de troisikme ordre (pulvinus tertiaires ou pulvinules); ep, f, r, s t et t representent respectivement les Cpines, les foliolules, les rachis, les stipules et la tige. Rappelons que Mimosa pudica L. prisente un polymorphisme foliaire: la premiere feuille, composie pennee, est dotCe de trois paires de folioles: a partir de la seconde, les feuilles posskdent une paire de folioles dCcoupCes en foliolules (feuille bifo- liol6es); a I'approche de la floraison les feuilles possedent deux paires de folioles kgalement d6couptes en foliolules (feuilles quadrifoliol6es).
sur les conseils du chimiste Jackson. l'utilisa avec succts, en remplacement du protoxyde d'azote. Puis, ce fut en 1847 que Clemens observa ses effets sur les organes vCgCtaux douCs de mouve- ments spCctaculaires: Ctamines de Berberis vul- garis, feuilles de Dionaea muscipula, d'Hedy- sarum gyrans, d'Oxalis sensitiua et de Mimosa pudica. L'influence d'anesthesiques sur les mouvements foliaires ~Cismonastiques et nyc- thCmCraux, de cette dernitre plante a d'ailleurs fait l'objet de nombreuses observations: Marcet (1848), Leclerc (1 853), Bert (1 866), Pfeffer (1 874), Bernard (1 875), Correns (1 892), Herbert (1 925), Bose (1928), Wallace (1931a, 1931b), Toriyama (1953), mais seuls les travaux de Fondeville (1 964) et Gaillochet et al. (1 975) ont indiquC avec prCcision les conditions d'utilisation des agents anesthksiques sur ce matiriel.
Conditions expPrimentales Les experiences ont etC realistes dans une chambre
climatiste a 26 + 1 "C, sous humiditi relative de 70-75%. L'tclairement (LID = 14/10), Ctait assure quotidienne- ment de 7 a 21 h, par une rampe de tubes fluorescents blancs 'LF Phytor' (ACEC Charleroi, Belgique) dispen- sant une Cnergie de 5000 ergs cm-2 s-' (2500 Ix, mesures par une cellule photoelectrique a surface plane), au niveau de I'apex des plantes; la rtpartition de I'Cnergie spectrale emise par ces tubes a t te publiie par de Greef et Fredericq (1969).
Materiel biologique Provenant de cultures realisies dans les conditions
pricCdemment decrites, les plantes grouptes en lots homogknes ont CtC transferees dans la chambre expkri- mentale 8 jours avant I'expirience, et utilisees ensuite quand la deuxieme feuille quadrifoliolte Ctait de dimen- sion Cgale a ses stipules.
Dispositif expPrimenta1 La realisation de I'etude a nCcessitC la construction d'un
appareillage (fig. 2) constituk par quatre enceintes en verre (cristallisoirs) presentant les caractkristiques suivantes: hauteur 29.5 cm, diametre 29 cm, volume 18.5 litres, monttes chacune sur un bbti en bois, permettant leur rotation complete autour d'un axe horizontal et leur fixation en position variable, dans le champ gravifique.
Protocole exp Primental Le jour pr6cCdent I'expCrience, les plantes, Ctayees et
arrostes, ont et6 solidement fix6es a un support de bois avant d'Ctre introduites, chacune, dans un cristallisoir place en position verticale.
Une heure avant I'experience, chaque cristallisoir a 6tC ferm6 par un couvercle de verre de 10 mm d16paisseur bloqui par des tcrous a serrage rapide, un joint de caoutchouc dur enduit de gel d'amidon assurant une 6tanchkitC parfaite.
L'inversion des plantes a 180 degrb dans le champ de la pesanteur s'effectue par un mouvement r6gulier des
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y SA
VA
NN
AH
RIV
NA
TL
AB
BF
on 1
1/13
/14
For
pers
onal
use
onl
y.

CAN. J. BOT. VOL. 55 , 1977
FIG. 2. Schema du dispositif utilise pour etudier ['action de substances volatiles sur la reaction gtotropique foliaire. (A) B%ti en bois supportant une enceinte en verre, cylindrique, (cristallisoir CR) pouvant tourner autour d'un axe horizontal de hauteur rtglable (systeme RH). Un volant (VM) soli- daire de cet axe, permet d'orienter I'enceinte, en position variable dans le champ gravifique et la vis de blocage (VB) permet de la fixer dans la position choisie. L'tclairage est assure par une rampe inferieure (REI) disposee sous le b%ti et une rampe suptrieure non representee ici. La plante est fixte sur un sup- port de bois (S). (B) La plante etant introduite, I'enceinte a ete fermee par un couvercle de verre (CV) solidement fixe par quatre vis a serrage rapide (VSR). L'inversion du dispositif, a I'aide du volant (VM), a ete realisee, selon I'exptrience, avant, pendant, ou apres admission de la substance volatile par le tube de caoutchouc (T) clampe par la pince (C).
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y SA
VA
NN
AH
RIV
NA
TL
AB
BF
on 1
1/13
/14
For
pers
onal
use
onl
y.
GAILLOCHET ET FLEURAT-LESSARD 73
enceintes autour de leur axe horizontal; une rarnpe lurnineuse disposee sous les bdtis apporte alors aux plantes inversees un tclairernent analogue a celui qu'elles recevaient en position verticale norrnale.
Trois sCries d'experiences ont Cte realisees. Dans la premiere, I'Cther a e t t introduit au moment de I'inversion des enceintes a 180 degres; dans la seconde, I'inversion des plantes a t te rtalisCe apres un temps variable d'ktheri- sation; dans la troisikrne, 1'Cther a etC introduit aprts I'inversion des plantes, a des moments variables de la reaction gkotropique.
Afin d'Cviter toute interference avec les variations angulaires induites par le rythrne nycthCrneral, toutes les experiences ont CtC effectukes entre 10 h 30 et 18 h 30.
Concentratiorts en Pther Ptl~ylique Un volume d'ether donne a etC introduit dans chaque
enceinte par un tube de plastique traversant le couvercle; le tube etant irnrnkdiaternent clampi, I'Cther presente alors dans chaque enceinte obturte une concentration donnee que nous avons exprirnee en terrne d'activitk therrno- dynarnique (syrnbolisee par 'a'). Cette notion, ClaborCe par le therrnodynarnicien Lewis (1907), introduite en toxicologic par Ferguson (1939) et Gavaudan et al. (1944) peut se definir, en phase gazeuse, cornrne une pression corrigee par rapport a la pression de vapeur saturante. Elle est reliCe a la variation d'energie libre de Gibbs AG par ]'equation
oh a = PIPo, P Ctant la pression de vapeur actuelle et Po la pression de vapeur saturante. Ferguson (1939) et Gavaudan (1947) adrnettent que l'ether, qui agit a des valeurs d'activitk therrnodynamique comprises entre quelques centiernes et I'unite, exerce une action de type physique, independant de la nature des molecules Ctran- gtres introduites; dans ce cas, seules les forces faibles de London sont rnises en cause, ce qui explique la reversi- bilite aisee de I'inhibition.
Les experiences n'ont CtC conduites que dans les con- ditions de reversibilite et de survie parfaites des feuilles choisies pour ]'experimentation c'est-a-dire pour les activitis therrnodynarniques de 0.05, 0.10, 0.15, 0.20 (et 0.25 pour les durees d'application inferieures a 240 rnin).
Expression des rPs~tltats Les angles foliaires (fig. 1) ont ete rnesures a l'aide d'un
rapporteur transparent rnuni d'un axe mobile pouvant se superposer au petiole ou au rachis des feuilles. L'etude a porte sur les deux dernieres feuilles bifoliolies et la pre- miere feuille quadrifoliolke (no d'ordre basifuge 6, 7 et 8) d'une rn&rne plante. Trois plantes ayant CtC utilistes dans chaque experience, chaque point constituant les courbes reprksente donc la rnoyenne des rnesures effectukes sur neuf feuilles.
En appelant Bo la valeur angulaire des organes rnoteurs prirnaires rnesuree sur la plante en position norrnale et 2, leur valeur angulaire rnesuree aux temps 'I' aprks I'inversion des plantes a 180 degris, dans le champ de la pesanteur, la reactivite gCotropique (gCorCactivitC) a Cte dkfinje pa! 2, y 2 , - 2,. La rnCrne definition exprirnke par D, = DO - D l a Ctt adoptee pour la gCorCactivitC des organes rnoteurs secondaires. L'introduction de cette
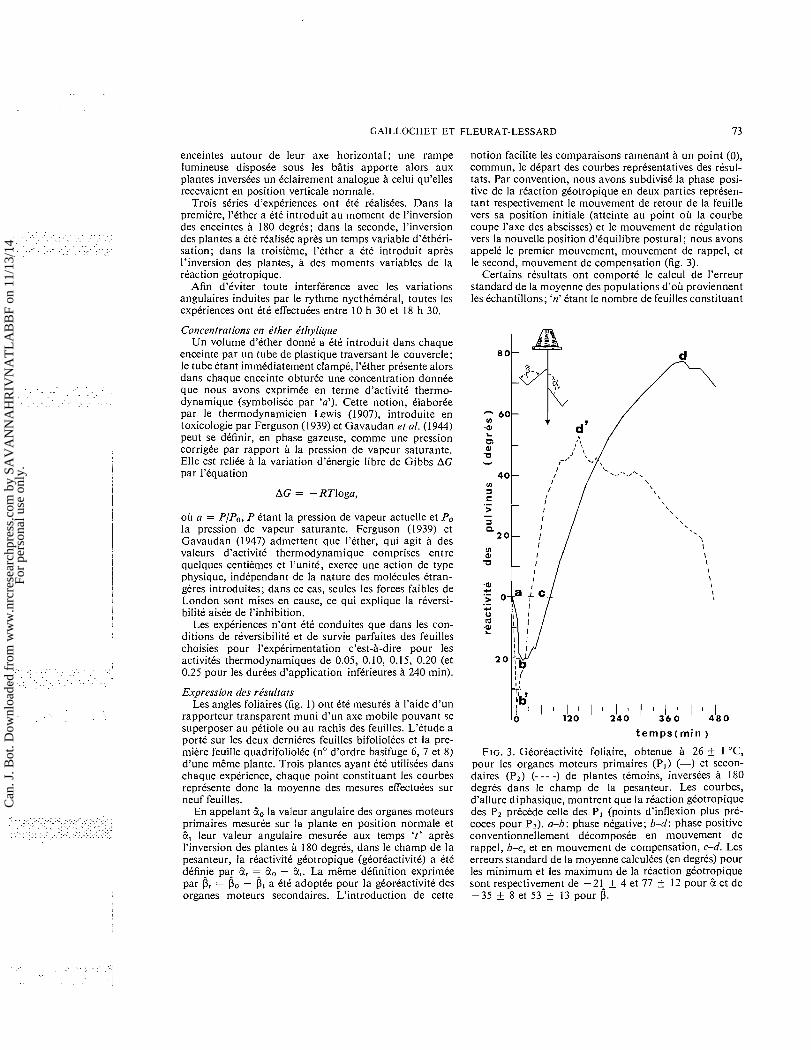
notion facilite les cornparaisons rarnenant a un point (0), cornrnun, le depart des courbes representatives des rtsul- tats. Par convention, nous avons subdivisk la phase posi- tive de la reaction gkotropique en deux parties represen- tant respectivernent le rnouvernent de retour de la feuille vers sa position initiale (atteinte au point oh la courbe coupe I'axe des abscisses) et le rnouvernent de regulation vers la nouvelle position d'kquilibre postural; nous avons appele le premier rnouvernent, rnouvernent de rappel, et le second, rnouvernent de compensation (fig. 3).
Certains rtsultats ont comport6 le calcul de l'erreur standard de la rnoyenne des populations d'oh proviennent les Cchantillons; 'n' Ctant le nornbre de feuilles constituant
FIG. 3. Georeactivite foliaire, obtenue a 26 t 1 "C, pour les organes rnoteurs prirnaires (P,) (-) et secon- daires (PZ) (--- -) de plantes ternoins, inversees a 180 degres dans le champ de la pesanteur. Les courbes, d'allure diphasique, rnontrent que la reaction gkotropique des P, precede celle des P, (points d'inflexion plus pre- coces pour PI). a-b: phase negative; b-d: phase positive conventionnellernenl decornposee en rnouvernent de rappel, b-c, et en rnouvernent de compensation, c-d. Les erreurs standard de la rnoyenne calcultes (en degrts) pour les minimum et les maximum de la reaction giotropique sont respectivernent de -21- t 4 et 77 t 12 pour 2 et de -35 t 8 et 53 + 13 pour D.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y SA
VA
NN
AH
RIV
NA
TL
AB
BF
on 1
1/13
/14
For
pers
onal
use
onl
y.

74 CAN. J. BOT. VOL. 55, 1977
i'echantillon, 'tn' la moyenne des valeurs mesurtes, avec un Ccart-type '0,' I'erreur standard de I'Cchantillon, pour n - 1 degres de IibertC, est s = o/-.
Si 'p' est la moyenne estimCe des valeurs de la popula- tion parente et 'I' le coefficient donnt par les tables de Fisher et Yates (Mather 1960) pour un intervalle de con- fiance de 95%, la marge de variabiliti de cette population est p + st.
Aucune diffkrence significative n'a CtC dCcelCe entre les plantes temoins observtes dans les enceintes IaissCes ouvertes et les plantes tCmoins Ctudites dans les enceintes hermktiquement fermees, les enceintes Ctant maintenues retournees de 10 h 30 a 18 h 30.
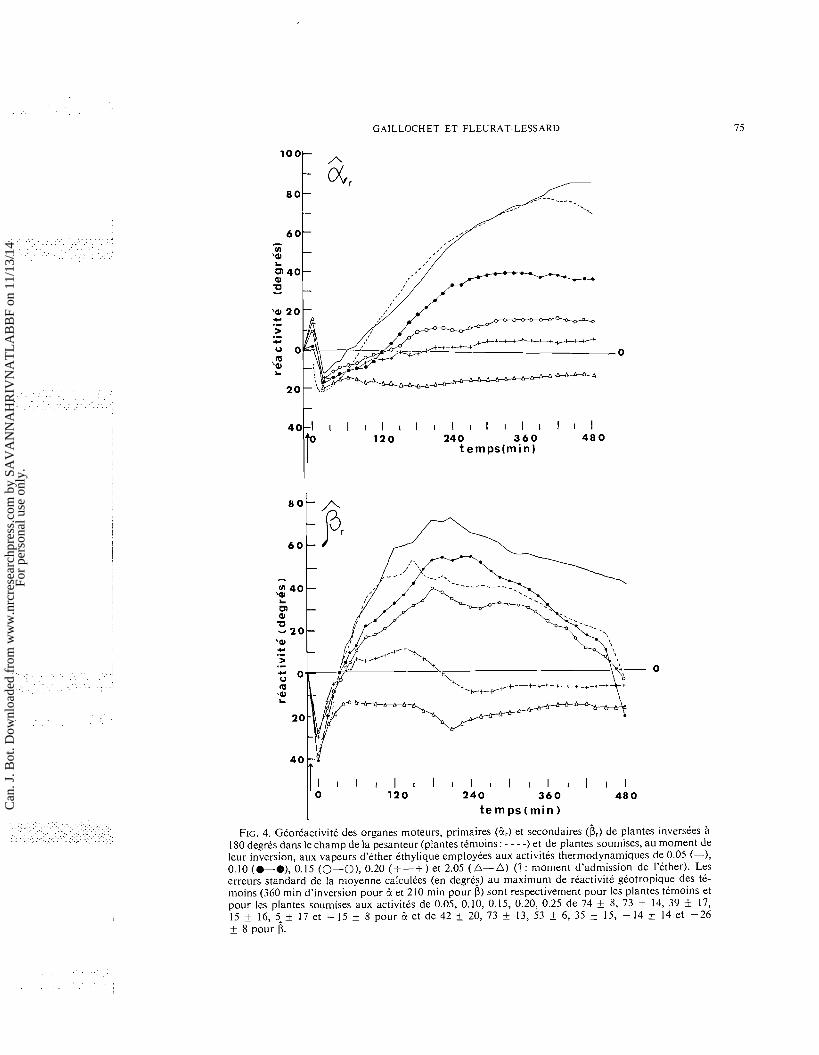
(1) L'inversion des plantes a t t t rtaliste im- mtdiatement apres l'introduction de l'tther dans les enceintes. Le mouvement gtotropique foliaire qui s'est dtroult dans les vapeurs d'tther pendant 480 min, a prtsentt comparativement a celui des ttmoins (fig. 31, les caracttres suivants (fig. 4): (a) Sous l'influence d'activitts thermodyna-
miques faibles (inftrieures ou tgales A 0.05), ]'amplitude du mouvement gtotropique est augmentte. Ce phtnomene d'excitation, an- ttrieurement observe dans l'ttude des mouve- ments stismonastiques (Gaillochet et al. 1975), et retrouvt ici pendant la rotation des enceintes qui dtclenche un mouvement stismonastique (mouvement rapide des pttioles vers la base des plantes), ne se produit pas chez les plantes ttmoins. Or, si l'admission d'tther est rtaliste immtdiatement aprts l'inversion des plantes, le mouvement stismonastique n'est pas pro- voqut, ce qui souligne l'influence de l'anes- thtsique sur la sensibilitt foliaire pendant les 30 s ntcessaires A la rotation des enceintes. En tout t ta t de cause, qu'il y ait ou non cette stimulation mtcanique initiale, le dtcours posi- tif de la rtaction gtotropique ne s'en trouve pas modifit de manitre significative.
(b) La phase ntgative de la rtaction gtotropique n'est pas modifite de maniere significative par l'anesthtsique; par contre, le degrt d'inhibi- tion de la phase positive varie, pour un mCme temps d'action de l'tther, avec l'intensitt de l'activitt thermodynamique. Ainsi, la vitesse du mouvement de rappel, de mCme que la vitesse et l'amplitude du mouvement de com- pensation, sont diminutes en fonction directe de l'intensitt de l'activitt thermodynamique (fig. 4).
(c) Les ttapes principales constituant la reaction gtotropique sont accentutes par influence de l'tther (fig. 4). Pour l'organe moteur primaire
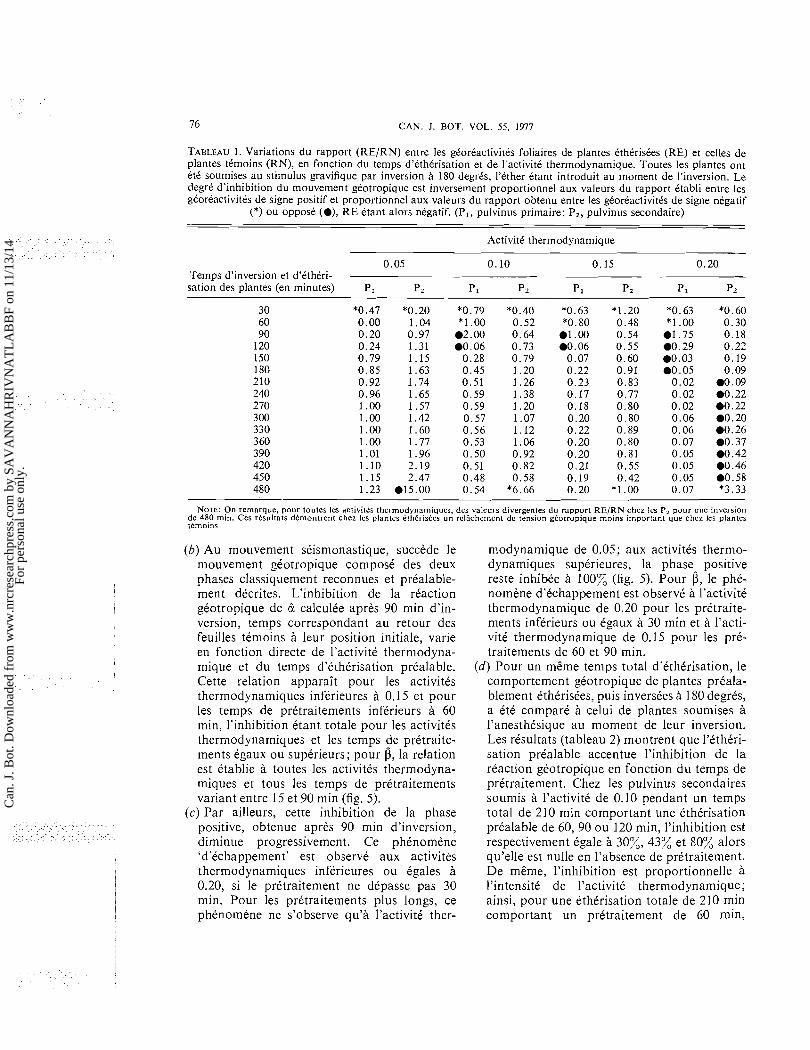
(B), la premiere ttape, effectute en 10 min, cor- respond au rapprochement feuille-tige (phase negative); la deuxitme, rtaliste pendant les 50 min suivantes indique une acctltration du mouvement gtotropique (mouvement de rap- pel); la troisieme effectute en 240 min montre un ralentissement de vitesse du mouvement aboutissant un optimum de remontte (mouvement de compensation) et la quatritme correspond a une phase de stabilkation du mouvement gtotropique.
(d) Le degrt d'inhibition du mouvement gtotro- pique varie tgalement, pour une activitt ther- modynamique et un temps d'action donnts de l'tther, avec le numtro d'ordre de l'articula- tion foliaire. Afin de comparer le mouvement des pulvinus primaires et secondaires, nous avons ttabli le rapport entre les valeurs de gtorCactivitt obtenues en atmosphere d'tther (RE) et les valeurs relevtes, au mCme instant, chez les plantes ttmoins (RN). Les valeurs du rapport RE/RN (tableau 1) indiquent que pendant la phase ntgative et la phase de rap- pel, l'inhibition des P, est gtntralement plus marqute que celle des P , . Par contre, pendant la phase de compensation, l'tther employt aux activitts thermodynamiques inftrieures ou tgales 0.15 exerce, sur les pulvinus primaires, une inhibition du mouvement gtotropique plus marqute que sur les pulvinus secondaires. (2) L'inversion des plantes a t t t rtaliste apres
un temps variable d'tthtrisation. Cette expt- rience a t t t entreprise dans le but de dtterminer les modalitts d'action d'une tthtrisation prta- lable des plantes sur le dtcours de la reaction gtotropique foliaire. Les plantes soumises, en position normale, aux vapeurs d'tther, pendant des durtes de 15 min, 30 min graduellement tchelonntes ensuite de 30 min jusqu'a 240 min, ont t t t inverstes par la suite dans l'atmosphere d'tther pendant 4 h. (a) Conformtment a nos premieres observations
rapporttes ci-dessus, la rotation des enceintes a provoqut, chez les plantes tthtristes, la stimulation mtcanique des pulvinus primaires. Cette stimulation a t t t abolie, dans nos con- ditions exptrimentales, apres 30 min d'action d'une activitt thermodynamique tgale ou suptrieure h 0.20; mais aucune relation n'a pu Ctre ttablie, de maniere significative, entre les conditions d'tthtrisation prtalables et l'ampli- tude de la rtponse mtcanique des pulvinus primaires.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y SA
VA
NN
AH
RIV
NA
TL
AB
BF
on 1
1/13
/14
For
pers
onal
use
onl
y.
GAlLLOCHET E T FLEURAT-LESSARD
FIG. 4. GCorCactivitC des organes moteurs, primaires (2,) et secondaires (b,) de plantes inverskes a 180 degrks dans le champ de la pesanteur (plantes tCmoins: - - - -) et de piantes soumises, au moment de leur inversion, aux vapeurs d'ether ethylique eniployCes aux activites thermodynamiques de 0.05 (-), 0.10 (@-a), 0.15 (0-O), 0.20 (+ -+ ) et 2.05 (A- A) (1 : moment d'admission de I'ether). Les erreurs standard de la nloyenne calculees (en degres) au maximum de reactivite geotropique des tC- noi ins (360 min d'inversion pour B et 210 min pour CJ) sont respectivement pour les plantes temoins et pour les plantes soumises aux activites de 0.05, 0.10, 0.15, 0.20, 0.25 de 74 + 8, 73 + 14, 39 17, 15 + 16, 5-+ 17 et -15 + 8 pour & e t d e 4 2 + 20, 73 + 13, 53 + 6, 35 + 15, -14 + 14 et -26 + 8 pour CJ.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y SA
VA
NN
AH
RIV
NA
TL
AB
BF
on 1
1/13
/14
For
pers
onal
use
onl
y.
76 CAN. J. BOT. VOL. 55. 1977
TABLEAU 1. Variations du rapport (RE/RN) entre les gCorCactivitts foliaires de plantes CthCrisCes (RE) et celles de plantes tCmoins (RN), en fonction du temps d'etherisation et de I'activite thermodynamique. Toutes les plantes ont ete soumises au stimulus gravifique par inversion a 180 degres, l'ether etant introduit au moment de I'inversion. Le degrt d'inhibition du mouvement gCotropique est inversement proportionnel aux valeurs du rapport Ctabli entre les gCorCactivites de signe positif et proportionnel aux valeurs du rapport obtenu entre les gCorCactivites de signe negatif
(*) ou oppose ( a ) , R E Ctant alors nCgatif. (PI, pulvinus primaire; P2, pulvinus secondaire)
ActivitC thermodynamique
0.05 0.10 0.15 0.20 Temps d'inversion et d3Cthtri- -
sation des plantes (en minutes) P, p2 PI p2 p1 Pz p 1 p z
NOTE: On remarque, pour toutes les activites thermodynamiques, des valeurs divergentes du rapport R E / R N chez les P, pour une inversion de 480 min. Ces resultats demontrent chez les plantes etherisees un relichement de tension georropique moins important que chez les plantes tkmoins
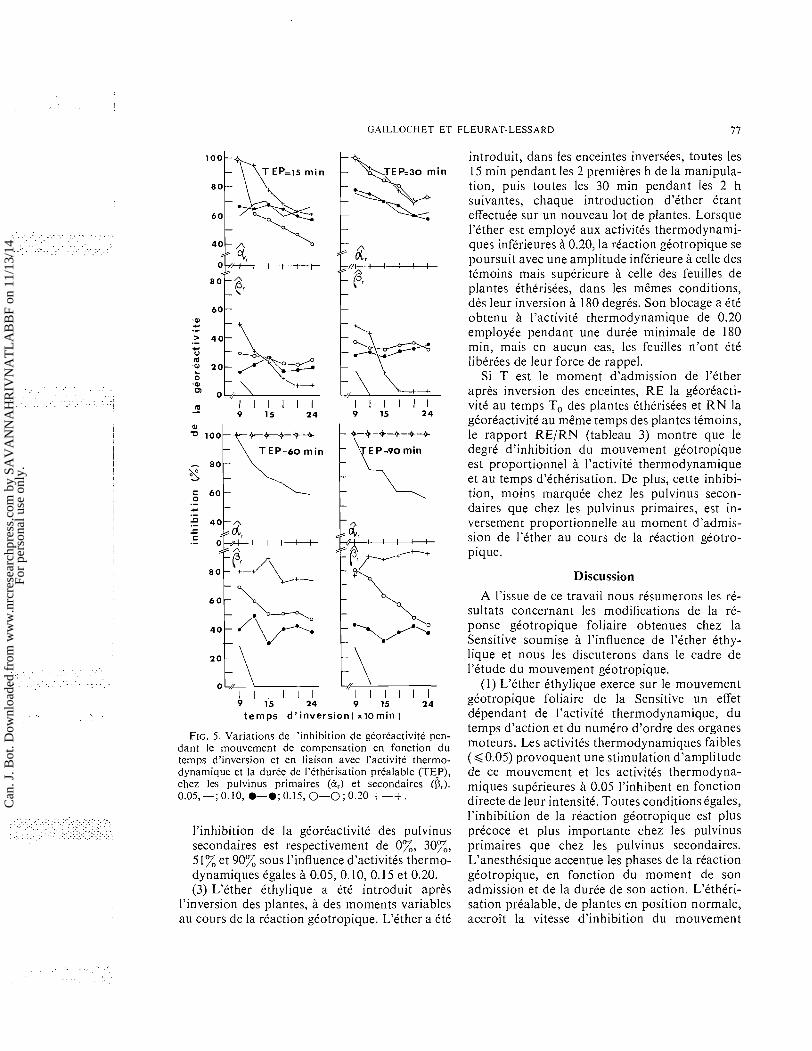
(6) Au mouvement seismonastique, succede le niouvement gtotropique compost des deux phases classiquement reconnues et prtalable- ment dtcrites. L'inhibition de la rtaction gtotropique de B calculte apres 90 rnin d'in- version, temps correspondant au retour des feuilles ttmoins a leur position initiale, varie en fonction directe de l'activitt thermodyna- mique et du temps d'tlhtrisation prtalable. Cette relation apparait pour les activitts thermodynamiques inftrieures a 0.15 et pour les temps de prttraitements inftrieurs a 60 min, l'inhibition t tant totale pour les activitts thermodynamiques et les temps de prttraite- ments tgaux ou suptrieurs; pour 0, la relation est ttablie a toutes les activitts thermodyna- miques et tous les temps de prttraitements variant entre 15 et 90 rnin (fig. 5).
(c) Par ailleurs, cette inhibition de la phase positive, obtenue a p r b 90 rnin d'inversion, diminue progressivement. Ce phtnomene 'd'tchappement' est observt aux activitts thermodynamiques inftrieures ou tgales a 0.20, si le prttraitement ne dtpasse pas 30 min. Pour les prdtraitements plus longs, ce phtnomene ne s'observe qu'a l'activitt ther-
modynamique de 0.05; aux activitts thermo- dynamiques suptrieures, la phase positive reste inhibte a 100% (fig. 5). Pour p, le pht- nomene d'tchappement est observt a l'activitt thermodynamique de 0.20 pour les prttraite- ments inftrieurs ou tgaux a 30 min et l'acti- vite thermodynamique de 0.15 pour les prt- traitements de 60 et 90 min.
(d) Pour un meme temps total d'tthtrisation, le comportement gtotropique de plantes prtala- blement ethtristes, puis inverstes a 180 degrts, a t t t compart a celui de plantes soumises a I'anesthtsique au moment de leur inversion. Les rtsultats (tableau 2) montrent que l'tthtri- sation prtalable accentue l'inhibition de la reaction gtotropique en fonction du temps de prttraitement. Chez les pulvinus secondaires soumis A l'activitt de 0.10 pendant un temps total de 210 rnin comportant une tthtrisation prtalable de 60, 90 ou 120 min, l'inhibition est respectivement tgale 30%, 43% et 80% alors qu'elle est nulle en I'absence de prttraitement. De meme, l'inhibition est proportionnelle a I'intensitt de l'activitt thermodynamique; ainsi, pour une tthtrisation totale de 210 rnin comportant un prttraitement de 60 min,
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y SA
VA
NN
AH
RIV
NA
TL
AB
BF
on 1
1/13
/14
For
pers
onal
use
onl
y.
GAILLOCHET ET FLEURAT-LESSARD 77
T EP=15 min
9-+-+-+-4-4
E P-90 rnin
t emps d l invers ion( x i o m i n )
FIG. 5. Variations de I'inhibition de gCoreactivite pen- dant le ~nouvement de compensation en fonction du temps d'inversion et en liaison avec I'activite thermo- dynamique et la duree de I'ethCrisation prealable (TE-P), chez les pulvinus primaires (2,) et secondaires (0,). 0.05,-; 0.10, 9-9;0.15, 0-0; 0.20 +-+.
l'inhibition de la gCorCactivitC des pulvinus secondaires est respectivement de 070, 30%, 51% et 90% sous l'influence d'activitis thermo- dynamiques Cgales A 0.05, 0.10, 0.15 et 0.20. (3) L'Cther Cthylique a CtC introduit apres
I'inversion des plantes, B des moments variables au cours de la rCaction gtotropique. L'Cther a t t t
introduit, dans les enceintes inversies, toutes les 15 min pendant les 2 premigres h de la manipula- tion, puis toutes les 30 min pendant les 2 h suivantes, chaque introduction d'tther Ctant effectuCe sur un nouveau lot de plantes. Lorsque l'tther est employ6 aux activitCs thermodynami- ques inferieures B 0.20, la rtaction giotropique se poursuit avec une amplitude infkrieure B celle des tCmoins mais superieure B celle des feuilles de plantes Ctherisees, dans les m&mes conditions, des leur inversion a 180 degrCs. Son blocage a CtC obtenu B llactivitC thermodynamique de 0.20 employte pendant une durte minimale de 180 min, mais en aucun cas, les feuilles n'ont Ctt libCrCes de leur force de rappel.
Si T est le moment d'admission de 1'Cther apres inversion des enceintes, RE la gCortacti- vitt au temps To des plantes CthCrisCes et R N la gCorCactivitC au m&me temps des plantes tCmoins, le rapport RE/RN (tableau 3) montre que le degrt d'inhibition du mouvement gtotropique est proportionnel B 1'activitC thermodynamique et au temps d1tthCrisation. De plus, cette inhibi- tion, moins ~narquCe chez les pulvinus secon- daires que chez les pulvinus primaires, est in- versement proportionnelle au moment d'admis- sion de I'Cther au cours de la reaction geotro- pique.
Discussion
A I'issue de ce travail nous rCsumerons les r t- sultats concernant les modifications de la rC- ponse gkotropique foliaire obtenues chez la Sensitive soumise B l'influence de l'tther Cthy- lique et nous les discuterons dans le cadre de 1'Ctude du mouvement gCotropique.
(1) L'Cther tthylique exerce sur le mouvement gkotropique foliaire de la Sensitive un effet dependant de I'activitC thermodynamique, du temps d'action et du numCro d'ordre des organes moteurs. Les activitCs thermodynamiques faibles (G0.05) provoquent une stimulation d'amplitude de ce mouvement et les activites thermodyna- miques supkrieures B 0.05 I'inhibent en fonction directe de leur intensitC. Toutes conditions Cgales, I'inhibition de la rCaction giotropique est plus prCcoce et plus importante chez les pulvinus primaires que chez les pulvinus secondaires. L'anesthtsique accentue les phases de la rtaction giotropique, en fonction du moment de son admission et de la durCe de son action. L'ttheri- sation prCalable, de plantes en position normale, accroit la vitesse d'inhibition du mouvement
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y SA
VA
NN
AH
RIV
NA
TL
AB
BF
on 1
1/13
/14
For
pers
onal
use
onl
y.

78 CAN. J. BOT. VOL. 55 , 1977
TABLEAU 2. Variations du degrC d'inhibition (pourcentage) de la phase positive de la reaction geotropique foliaire d e plantes prealablement etheristes avant d'ktre soumises au stimulus gravifique par inversion a 180 degrCs et d e plantes etherisies au moment de leur inversion. Le pourcentage d'inhibition a ete calcule chez les pulvinus primaires (P,) et secondaires (P,) en fonction de I'activitt thermodynamique, du temps dlCthCrisation totale (TTE), du temps d e pre-etherisation (TEP) et du temps d'action d e I'tther sur la reaction geotropique (To) (seules les valeurs de To superieures a 90 min ont e t t retenues, ce temps correspondant au moment oh les
pulvinus primaires des feuilles t h o i n s amorcent le mouvement de compensation)
Activite thermodynamique
P I pz T T E T E P To (min) (min) (min) 0.05 0.10 0.15 0.20 0.05 0.10 0.15 0.20
150 0 150 2 1 72 93 97 0 21 45 89 60 90 100 100 100 100 24 40 70 80
180 0 180 15 55 78 95 0 0 9 91 60 120 80 100 100 100 0 50 60 80 90 90 100 100 100 100 27 37 80 78
210 0 210 8 49 77 98 0 0 17 91 60 150 70 100 100 100 0 30 5 1 90 90 120 75 100 100 100 0 43 75 90
120 90 99 100 100 100 30 80 90 100
240 0 240 4 4 1 83 98 0 0 23 78 60 180 67 100 100 100 0 40 52 72 90 150 75 100 97 100 0 58 63 88
120 120 80 100 100 100 5 95 100 100 150 90 98 100 100 100 26 100 100 100
gkotropique; ce phknomtne s'attknue ('kchappe- ment') si l'activitk thermodynamique est- in- fkrieure a 0.15. L'kthkrisation produite au cours de la reaction geotropique provoque le blocage du mouvement Dar action de l'activite 0.20 uti- liske pendant le temps minimal de 180 min; un temps d'action infkrieur et les activitks thermo- dynamiques plus faibles utiliskes au minimum pendant 240 min, n'entrainent qu'une diminu- tion de vitesse du mouvement.
La confrontation entre ces rksultats et ceux aui ont kt6 obtenus a la suite de l'ktude du mouvement skismonastique foliaire provoque par agitation generale manuelle de la Sensitive soumise aux m@mes conditions expkrimentales (Gaillochet et a/. 1975) a permis de mettre en evidence les caracttres suivants: (a) Le mouvement foliaire geotropique (mouve-
ment lent provoquk) et le mouvement foliaire seismonastique (mouvement rapide provoquk) sont inhibks par 1'Cther kthylique utilisk dans une marge d7activitCs thermodynamiques pro- duisant, chez d'autres organismes, l'inhibition de certaines fonctions cellulaires: caryocintse (Gavaudan 1945 ; Deysson 1948), photosyn- these (Marchand 1965), bioluminescence bac- tkrienne (Hermelin 1966).
(b) Dans les mouvements geotropique et skis- monastique, la loi d'inhibition s'exprime de faqon identique par des courbes d'allure sig- moi'de (fig. 6); ces courbes ont etk construites en prenant les valeurs d'inhibition obtenues apres 150 min d'ktherisation, temps corres- pondant a la gCoreactivitk maximale des pul- vinus secondaires des plantes temoins. D'autre part, le degre d'inhibition des reactions gko- tropique et skismonastique de ces organes moteurs secondaires n'a pu @tre calculk sur la meme valeur angulaire, n'ktant pas affecte de mani6re significative par la seismonastie, et 7 ne l'ktant pas par la gkorkactivitk (f ktant l'angle dklimitk par les rachis). Par ailleurs, l'inhibition partielle de gkorkactivite, obtenue pour les activites thermodynamiques <0.20, est suivie par un phenomtne 'd'kchappement' plus marque dans ce type de mouvement que dans le mouvement skismonastique (fig. 7).
(c) Le seuil d'inhibition totale du mouvement gkotropique suit, comme celui du mouvement seismonastique (Fondeville 1964 ; Gaillochet et a/. 1975), une courbe hyperbolique (fig. 8) rappelant celle qui a Ctk obtenue dans l'etude des inhibitions fonctionnelles du muscle dorsal de sangsue et de lombric (Guyot 1958) et du
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y SA
VA
NN
AH
RIV
NA
TL
AB
BF
on 1
1/13
/14
For
pers
onal
use
onl
y.
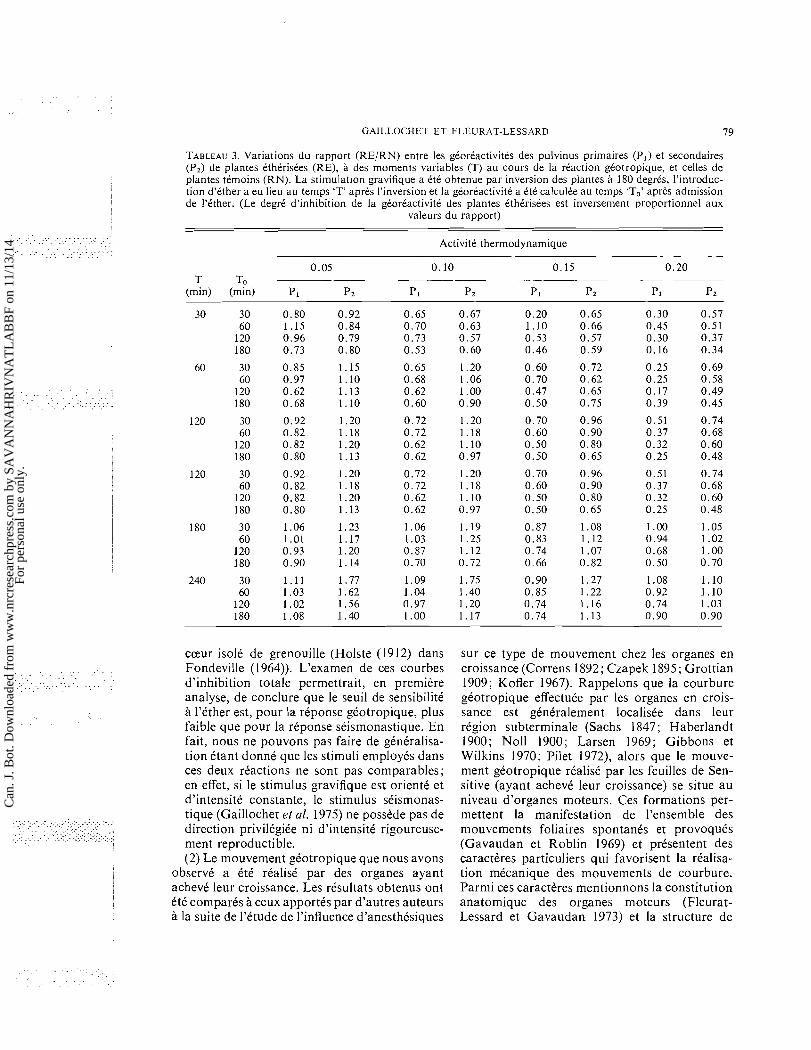
GAILLOCHET El' FLEURAT-LESSARD 7 9
TABLEAU 3. Variations du rapport (RE/RN) entre les gCorCactivites des pulvinus primaires (P,) et secondaires (P,) de plantes etherisees (RE), a des moments variables (T) au cours de la reaction geotropique, et celles de plantes temoins (RN). La stimulation gravifique a ete obtenue par inversion des plantes a 180 degres, l'introduc- tion d'ether a eu lieu au temps 'T' apres I'inversion et la georeactivite a ete calculee au temps 'To' aprts admission de I'ether. (Le degre d'inhibition de la georeactivite des plantes ttherisees est inversement proportionnel aux
valeurs du rapport)
ActivitC thermodynamique
0.05 0.10 0.15 0.20 T To -
(min) (min) P I Pz PI Pz PI Pz PI Pz
cceur isolt de grenouille (Holste (1912) dans Fondeville (1964)). L'examen de ces courbes d'inhibition totale permettrait, en premiere analyse, de conclure que le seuil de sensibilitt a l'tther est, pour la rtponse gtotropique, plus faible que pour la reponse stismonastique. En fait, nous ne pouvons pas faire de gentralisa- tion i tant donnt que les stimuli ernployts dans ces deux reactions ne sont pas cornparables; en effet, si le stimulus gravifique est orient6 et d'intensitt constante, le stimulus dismonas- tique (Gaillochet et a/. 1975) ne possede pas de direction priviltgite ni d'intensitt rigoureuse- ment reproductible. (2) Le mouvement gtotropique que nous avons
observt a t t t realist par des organes ayant achevt leur croissance. Les rtsultats obtenus ont ett comparts a ceux apportts par d'autres auteurs a la suite de I'ttude de l'influence d'anesthtsiques
sur ce type de mouvement chez les organes en croissance (Correns 1892; Czapek 1895 ; Grottian 1909; Kofler 1967). Rappelons que la courbure gtotropique effectute par les organes en crois- sance est gtntralement localiste dans leur region subterminale (Sachs 1847 ; Haberlandt 1900; No11 1900; Larsen 1969; Gibbons et Wilkins 1970; Pilet 1972), alors que le mouve- ment gtotropique rtalist par les feuilles de Sen- sitive (ayant achevt leur croissance) se situe au niveau d'organes moteurs. Ces formations per- mettent la manifestation de l'ensemble des mouvements foliaires spontants et provoquts (Gavaudan et Roblin 1969) et prtsentent des caracteres particuliers qui favorisent la rtalisa- tion mtcanique des mouvements de courbure. Parmi ces caracteres mentionnons la constitution anatomique des organes moteurs (Fleurat- Lessard et Gavaudan 1973) et la structure de
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y SA
VA
NN
AH
RIV
NA
TL
AB
BF
on 1
1/13
/14
For
pers
onal
use
onl
y.

CAN. J . BOT.
I I I I I 0.0 5 0.1 5 0.25
acti v i t e t herrnodynarnique
FIG. 6. DegrC d'inhibition de la gCorCactivitC foliaire apres 150 min d'etherisation chez les plantes soumises aux vapeurs d'ether au moment de leur inversion a 180 degres dans le champ gravifique et de la reaction skis- monastique de plantes identiques CthCrisCes en position normale. GeorCactivite des pulvinus primaires (G(PI), 0 - 0 ) . et secondaires (G(P2), 0 - - - 0 ) ; reactivite seis- monastlque des pulvinus primaires (S(P,), 8 -8 ) et secondaires (S(P2), 8-- -8) .
leurs parois parenchymateuses corticales (Wein- traub 1952) rappelant celle des parenchymes juvtniles observts dans les organes en croissance. ~ e ~ e n d a n t , dans les organ& en croissance et dans les organes moteurs, le siege de la percep- tion gravifique serait, selon la thCorie classique (Haberlandt 1900; Nemec 1900; Noll 1900), localis6 dans un tissu spCcial distinct de ceux oh s'effectue la courbure. Ce tissu contenant de volumineux plastes amyliferes, mobiles sous l'influence de variations d'orientation dans le champ de la pesanteur, est situC dans la colu- melle de la coiffe, donc au voisinage immCdiat de la rCgion de courbure racinaire. ~ou te fo i s . dans - les hypocotyles, tiges en croissance et organes moteurs foliaires, i l est localis6 dans la rCgion interne du parenchyme cortical, donc en associa-
VOL. 5 5 . 1977
l00C
"t I I I I I I I I I I I I I I 6 12 18 24 6 12 18 24
t e m p s d'et herisationcx lo min)
FIG. 7. Variations temporelles d'inhibition (pourcen- tage) de la georkactivitC foliaire de plantes inversees a 180 degrCs, etherisees au moment de leur inversion et de la reaction seismonastique de plantes etherisees, pendant les m&mes temps, en position normale. GCoreactivitC des pulvinus primaires (G(P,), 0 - 0 ) et secondaires (G(P2), 0- - - 0 ) ; reactiviti seismonastique des pulvinus primaires (S(P,), 8-8) et secondaires (S(P2), 8 - - -8) .
tion Ctroite avec les tissus corticaux pCriphC- riques oh s'effectue la courbure gtotropique.
L'ttude du mouvement geotropique d'organes en croissance, soumis i I'influence d'anesthksi- ques, a montrC que les faibles concentrations d'anesthksiques stimulent la courbure et la croissance, alors que les fortes concentrations d'anesthksiques inhibent la courbure sans sus- pendre totalement la croissance (Grottian 1909). De mEme, chez la Sensitive, la rCaction gCotro- pique de feuilles ayant achevt leur croissance a CtC stimulte par les faibles concentrations d'Cther Cthylique et inhibee par les fortes concentrations. En outre, pour ces dernieres concentrations d'kther Cthylique, les altCrations morphologiques produites chez les feuilles apicales soulignent l'effet inhibiteur exercC par l'anesthesique sur les processus de croissance. Par ailleurs, Kofler (1967) a montrC que l'hydrate de chloral pro- voque chez les filaments issus des spores de Funaria hygrometrica un renforcement de polari-
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y SA
VA
NN
AH
RIV
NA
TL
AB
BF
on 1
1/13
/14
For
pers
onal
use
onl
y.
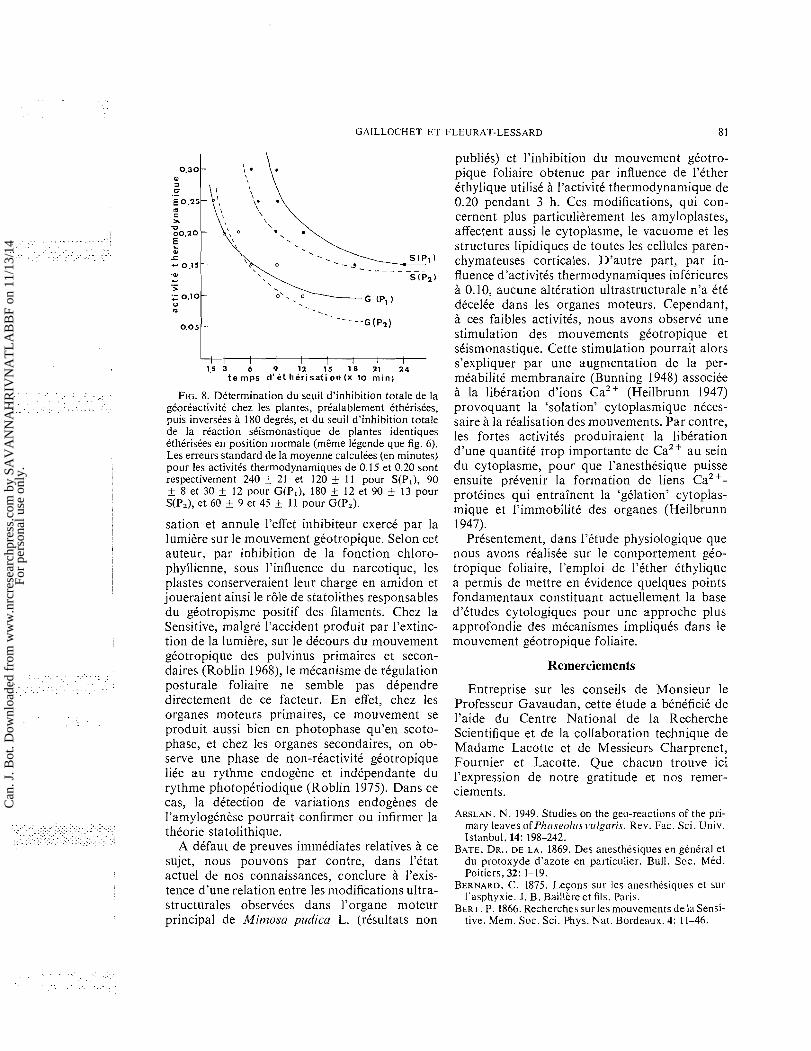
GAILLOCHET ET FLEURAT-LESSARD 81
1 . 5 3 6 9 12 15 1 8 21 2 4 t e m p s d ' e t her isat i on ( X 10 min)
. . . . . . . . . . . . . . . . . . . . . . FIG. 8. Determination du seuil d'inhibition totale de la . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . gCortactivitC chez les plantes, preaiablernent CthCrisCes,
puis inverstes 180 degres, et du seuil d'inhibition totale de la reaction stisrnonastique de plantes identiques CthCrisees en position norrnale (rnerne legende que fig. 6). Les erreurs standard de la rnoyenne calculees (en minutes) pour les activitCs therrnodynarniques de 0.15 et 0.20 sont respectivernent 240 + 21 et 120 + 11 pour S(P,), 90 i 8 et 30 ? 12 pour G(P,), 180 1 12 et 90 + 13 pour S(P,), et 60 + 9 et 45 + 11 pour G(P,).
i sation et annule l'effet inhibiteur exerct par la lumitre sur le mouvement gtotropique. elo on cet auteur, par inhibition de la fonction chloro- phyllienne, sous ]'influence du narcotique, les plastes conserveraient leur charge en amidon et joueraient ainsi le r6le de statolithes responsables du gtotropisme positif des filaments. Chez la Sensitive, malgrt l'accident produit par I'extinc- tion de la lumiere, sur le dtcours du mouvement gtotropique des pulvinus primaires et secon-
. . . . . . . . . . . . . . . . . . . daires (Roblin 1968), le mtcanisme de rtgulation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . posturale foliaire ne semble pas dtpendre
directement de ce facteur. En effet, chez les . .
. . organes moteurs primaires, ce mouvement se . - . produit aussi bien en photophase qu'en scoto-
phase, et chez les organes secondaires, on ob- serve une phase de non-rtactivitt gtotropique lite au rythme endogene et indtpendante du rythme photoptriodique (Roblin 1975). Dans ce cas, la detection de variations endogtnes de l'amylogtntse pourrait confirmer ou infirmer la thtorie statolithique.
A dtfaut de preuves immtdiates relatives ce sujet, nous pouvons par contre, dans l'ttat actuel de nos connaissances. conclure a I'exis- tence d'une relation entre les modifications ultra- structurales observtes dans l'organe moteur principal de Mimosa pudica L. (rtsultats non
publits) et l'inhibition du mouvement gtotro- pique foliaire obtenue par influence de l'tther tthylique utilist A l'activitt thermodynamique de 0.20 pendant 3 h. Ces modifications, qui con- cernent plus particulitrement les amyloplastes, affectent aussi le cytoplasme, le vacuome et les structures lipidiques de toutes les cellules paren- chymateuses corticales. D'autre part, par in- fluence d'activitts thermodynamiques inftrieures B 0.10, aucune alttration ultrastructurale n'a t t t dCcelte dans les organes moteurs. Cependant, a ces faibles activitts, nous avons observt une stimulation des mouvements gtotropique et stismonastiaue. Cette stimulation ~ o u r r a i t alors s'expliquer par une augmentation de la per- mtabilitt membranaire (Bunning 1948) associte a la libtration d'ions Ca2+ (Heilbrunn 1947) provoquant la 'solation' cytoplasmique ntces- saire a la rtalisation des mouvements. Par contre, les fortes activitts ~roduiraient la libtration d'une quantite trop importante de Ca2+ au sein du cytoplasme, pour que I'anesthtsique puisse ensuite prevenir la formation de liens Ca2+- prottines qui entrainent la 'gtlation' cytoplas- mique et l'immobilitt des organes (Heilbrunn 1947).
Prtsentement, dans l'ttude physiologique que nous avons rtaliste sur le comportenlent gto- tropique foliaire, I'emploi de l'tther tthylique a permis de mettre en tvidence quelques points fondamentaux constituant actuellement la base d'ttudes cytologiques pour une approche plus approfondie des mtcanismes impliquts dans le mouvement gtotropique foliaire.
Remerciements
Entreprise sur les conseils de Monsieur le Professeur Gavaudan, cette t tude a btntficit de l'aide du Centre National de la Recherche Scientifique et de la collaboration technique de Madame Lacotte et de Messieurs Charprenet, Fournier et Lacotte. Que chacun trouve ici I'expression de notre gratitude et nos remer- ciements.
ARSLAN. N. 1949. Studies on the geo-reactions of the pri- mary leaves of Pilcr~seol~rs \>~~lgrrr-is. Rev. Fac. Sci. Univ. Istanbul. 14: 198-242.
BATE, DR.. DE LA. 1869. Des anesthesiques en general et du protoxyde d'azote en particuiier. Bull. Soc. Med. Poitiers, 32: 1-19.
BERNARD. C. 1875. Lecons sur les anesthtsiques et sur I'asphyxie. J. B. Baillereet fils. Paris.
BERT, P. 1866. Recherches sui- les mouvernents de la Sensi- tive. Mern. Soc. Sci. Phys. Nat. Bordeaux. 4: 11-46.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y SA
VA
NN
AH
RIV
NA
TL
AB
BF
on 1
1/13
/14
For
pers
onal
use
onl
y.

82 CAN. J. BOT. VOL. 55. 1977
BONNET. C. 1779. Recherches sur I'usage des feuilles dans les plantes et sur quelques autres sujets relatifs a I'his- toire de la vegetation. D N ~ I S Collection complete des cruvres de Charles Bonnet. Samuel Fauche, NeuchBtel. pp. 179-505.
BOSE. J . CH. 1928. The motor mechanism of plants. Longmans, Green and Co.. London.
BRAUNER, L . , et W. RAU. 1966. Versuche zur Bewe- gungsphysiologie der Pflanzen. Drrtls Pflanzenphysio- logische Praktika. Vol. 3. Springer-Verlag, Berlin. pp. 13-14.
BUNNING, E. 1948. Entwicklungs und Bewegungs- physiologiederPflanzen.Sp~inger-Verlag, Berlin. p. 286.
CLEMENS, M. 1847. Etherisation de plantes douees de mouvements spontanis visibles. Bull. Soc. Vaud. Sci. Nat. 2: 257-259.
CORRENS. C . 1892. ~ b e r die Abhingigkeit der Reizer- scheinungen hoherer Pflanzen von der Gegenwart freien Sauerstoffes. Flora (Jena), 75: 87-15 1.
CZAPEK, F. 1895. Untersuchungen iiber Geotropismus. Jahrd. Wiss. Bot. 27: 249-339.
DE GREEF. J.. et H. FREDERICQ. 1969. Photomorphogenic and chlorophyll studies in the Bryophyte M ~ r c l ~ r r t ~ t i n polytt~orphrr. 11. Photobiological responses to terminal irradiations with different redlfar-red ratios. Physiol. Plant. 22: 462-468.
DEYSSON, G. 1948. Contribution B I'itude du syndrome mitoclasique. These de Pharmacie, Paris.
FERGUSON. J . 1939. The use of chemical potentials as indices of toxicity. Proc. R. Soc. London, Ser. B. 127: 387-404.
FLEURAT-LESSARD. P., et P. G A V A U D A N . 1973. Orga- nisation st~ucturale et ~ ~ l t r a s t r ~ ~ c t u r > ~ l e de I'organe sensorio-moteur principalen relation avec la physiologie du mouvement chez Mittrostr p~rdic(r L. C.R. SCances Soc. Biol., Paris, 167: 332-336.
FONDEVILLE. J . C. 1964. Recherches sur la sensibilite, la rnotl-iciti et les rythmes endogenes chez Mitt~ostrprrdictr LinnC. These Doctorat d'Etat. Poitiers.
GAILLOCHET, J.. H. POUSSEL et P. GAVAUDAN. 1975. Re- cherche de conditions "standard" de narcose chez Mit?los(r prrrlicn L . en vue de I'application i divers pro- blemes du mouvement foliaire. C.R. Soc. SCances Biol., Paris. 169: 652-659.
C A V A U D A N P. 1945. Pharmacodynamie de I'inhibition de la ca~yocinese. These de Doctorat en Pharmacie, Paris.
1947. La narcose par les narcotiques indiffkrents. Anesth. Analg. 6: 193-214.
GAVAUDAN, P., M. DODE et H. POUSSEL. 1944. La toxicologic genkrale et la notion d'activite thermo- dynamique. MCm. Ind. Chim. de I'Etat. Station d'essais du Boucher, 31: 384-423.
G A V A U D A N , P., et G. ROBLIN. 1969. Enregistrement et analyse des reactions motrices des feuilles de MOrrosct prrtliccr L . soumises B diverses orientations dans le champ de gravite. C.R. SCances Soc. Biol., Paris, 163: 743-746.
GIBBONS, B. S . , et M. B. WILKINS. 1970. Growth inhibitor production by root caps in relation to geotropic re- sponse. Nature, 226: 558-559.
GROTTIAN, W. 1909. Beitlige zur Kenntnis des Geotro- pismus. Beih. Bot. Centralbl. 24: 255-285.
GUYOT, M. 1958. Analyse par les hautes Dressions de . . quelques fonctions physiologiques des vegitaux et des animaux. These Doctorat Etat Sci. Nat.. Poitiers.
HABERLANDT. G. 1900. Ueber die Perception des geotro- pischen Reizes. Ber. Dtsch. Bot. Ges. 18: 261-272.
HEILBRUNN, L. V. 1947. An outline of general physiology. Ed. W. Saunders and Co.. Philadelphia and London.
HERBERT, D. A. 1925. Movement of Mimosrr prrrlica as affected by anesthetics and other substances. Proc. R. Soc. Queensland, 36: 121-147.
HERMELIN, B. 1966. Contribution I'etude de la rkver- sibilite des inhibitions thermiques et chimiques. These de Doctorat de SpCcialite. Poitiers.
KOFLER, L. 1967. Polarisation et geotropisme des spores de Fiit~trrirr hygrometrico en presence d'hydlxte de chloral. Drrtls Colloque de morphologie experimentale, Grenoble, 28-29 mars 1966. Bull. Soc. Bot. Fr. 3eme partie. pp. 138-150.
LARSEN. P. 1969. The optimum angle ofgeotropic stimula- tion and its relation to the starch srarolith hypothesis. Physiol. Plant. 22: 469-488.
LECLERC. 1853. Recherches physiologiques e t anato- miques sur I'appareil nerveux des vCg6taux. L'action des substances anesthesiques sur la Sensitive et sur d'autres vegetaux. C.R. Acad. Sci. Paris. 37: 526528.
LEWIS, G. N. 1907. Outline of a new system of ther- modynamic chemistry. Proc. Am. Acad. Arts Sci. 43: 259.
MARCET. 1848. Note sur I'action du chloroforme sur la Sensitive (Mimoscr prrrlic.tr). Arch. Sci. Phys. Nat. 9: 204-207.
MARCHAND. C. 1965. Recherches sur I'action des pres- sions ClevCes et des inhibiteurs chimiques sur la lumi- nescence bactkrienne. These Doctorat de SpCcialitC, Poitiers.
MATHER. K . 1960. Statistical analysis in biology. Methurn and Co. Ltd., London. (First published in 1943.)
MORTON. W. T . G . 1847. Memoire sur la decouverte du nouvelemploi de I'ether sulfurique. P. Baut~uche . Paris.
NEMEC. B. 1900. Ueber die Wahrnehmung des Schwer- kraftreizes bei den Pflanzen Jahrb. Wiss. Bot. 36: 8&178.
NOLL. F . 1900. Ueber Geotrooismus. Jahrd. Wiss. Bot. 34: 458-506.
PFEFFER. W. 1874. i jber Fortpflanzung des Reizes bei Mittrostr prrdictr. Jahrb. Wiss. Bot. 9: 308-326.
PILET, P. E. 1972. Geoperception et gkorkaction raci- naires. Physiol. Veg. 10: 147-367.
ROBLIN, G. 1968. Recherches sur les phinomenes ryth- miques er sur I'influence de la gravire chez Mitt~osri pirrlicci L . These de Doctorat de Specialit&. Poitiers.
1975. Chronobiologie et PI-oprietes sensorio- motrices de la Sensitive. These d e Doctorat d'Etat. Poitiers.
SACHS, J. 1874. Trait6 d e botanique. F . Savy Libr.. Paris. pp. 993-1003.
T O R I Y A M A , H. 1953. Observational and experimental studies of sensitive plants. 1. The structure of paren- chymatous cells of pulvinus. Cytologia, 18(4): 283-292.
WALLACE, R . H. 1931tr. Studies on the sensitivity of Mittloso prrtlictr. 1. Effect of certain general anesthetics upon sleep movement. Am. J. Bot. 18: 102-1 11.
19310. Studieson the sensitivity of Mittro.snprrrlictr. 11. Effecl of animal anesthetics and certain other com- pounds upon seismonastic sensitivity. Am. J . Bot. 18: 215-235.
WEINTRAUB. M. 1952. Leaf movements in Mitnos(r prrdicrr L. New Phytol. 50: 357-382.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y SA
VA
NN
AH
RIV
NA
TL
AB
BF
on 1
1/13
/14
For
pers
onal
use
onl
y.





























