Embed Size (px)
Citation preview

Pergamon
0305-'1978(95) 00109-3
BiochemicalSystematics and Ecology, VoL 24, N~ 3, pp. 237-242,1996 Copyright O 1996 Bsevier Science Ltd
Printsd in Great Britain. All rights reserved 0305-1978/96 $15.00+0.00
Limonoid and Flavonoid Composition in Varieties of Papeda and Papedocitrus
MARK A. BERHOW,* CHI H. FONGt and SHIN HASEGAWA~; Bioactive Constituents Research Unit, National Center forAgricultural Utilization Research,
Agricultural Research Service, U.S. Department of Agriculture, 1815 North University Street, Peoria, IL 61604-3902, U.S.A.
Key Word Index--Citrus; limonoids; flavonoids; papeda; Cabuyao; Citrus ladpe~,, C. hystrix;, C. macrophylla; C. webberii; C. junos; C. wilsonii; C. ichangen$is. Abst rac t - -The limonoids of the seeds and the flavonoids of the leaves of a number of species of the Citrus sub-genus Papeda were examined. The seeds contained only the normal citrus limonoids and did not contain ichangensin, the unique limonoid found in C. ichangensis. A flavonoid analysis showed that a number of papeda varieties including C. la~'pes and C. hys~ix may be a papeda hybrid species. C. junos had flavanone neoheperidosides, which are also found in cultivars related to the pummelo. All papedas examined had rutin present, which is characteristic of cultivars related to the citron. Citrus ichangensis had the most complex flavone/flavonol pattern of all the papedas examined. This suggests that Citrus ichangensis may truly be a unique papeda species unlike that of the other papedas. Copyright © 1996 Elsevier Science Ltd
Introduct ion The papedas represent a number of wild species of Citrus which are found growing in the forests of Eastern Asia. The subgenus Papeda has been classified by Tanaka (1977) into two sections: the section Papeda comprising the typical species and the section Papedocitrus which is intermediate in character between the subgenus Citrus and the section Papeda with flowers much like those in the subgenus Citrus and leaves like those of the section Papeda. The distinguishing characteristics of the papedas are the numerous, often acrid, oil droplets associated with the pulp vesicles and the large winged leaf petioles. The papedas can tolerate lower temperatures than do species of the Citrus subgenus and papeda hybrids may yet prove to be valuable as rootstock material. Because Citrus species hybridize easily, they have been difficult to classify and there have been several proposed classification schemes that are often at odds with one another (Swingle and Reece, 1967; Tanaka, 1969; Barret and Rhodes, 1976; Tanaka, 1977; Scora, 1988; Fang et al., 1991).
Limonoids are a group of chemically related triterpene derivatives that have received extensive study in plants of the Rutaceae and Meliaceae families. Among the 36 limonoids isolated from Citrus taxa and their hybrids, limonin is the predominant limonoid and is the primary cause of limonoid bitterness in citrus juices (Hasegawa and Herman, 1992). However, in Citrus ichangensis, a member of the sub- genus Papeda, ichangensin is the predominant limonoid and limonin is only accumulated at much lower concentrations (Bennett et aL, 1988; Herman et al., 1989).
*Author to whom correspondence should be addressed. tWork supported by the Citrus ProductsTechnical Committee. tPresent address: Western Regional Research Center, U.S. Department of Agriculture, Agricultural
Research Service, 800 Buchanan Street, Albany, Califomia 94710, U.S.A.
(Received 2 August 1995; accepted 1 November 1995)
237

238 M.A. BERHOW ETAL.
The f lavonoids are a major class of plant secondary metabol i tes wh ich have proved to have chemotaxonomic value. Citrus is a part icularly rich source of f lavonoids (Horowi tz and Gentili, 1977). A number of Citrus surveys ident i fy ing and quant i fy ing some of the major f lavonoids based on separat ions occur in the literature (Albach and Redman, 1969; Kefford and Chandler, 1970; Nishiura et aL, 1971; Park et al., 1983; Kanes et al., 1993), but l itt le wo rk has been done on the f lavonoid compos i t ion of the papedas.
M a t e r i a l s a n d M e t h o d s Seeds and/or leaves of Citrus hystrix, C. hystrix var. 'Cabuyao', C. ichangensis, C. junos 'Yuzu', C. wilsonii 'lchang lemon', C. latipes, C. webberii var. 'Montana', and C. macrophylla were obtained from the Citrus Variety Collection of the Agricultural Experiment Station, University of California at Riverside. Seeds were dried at 60°C for 3 days prior to analysis. Collected leaves were oven-dried at 60°C in brown paper bags for at least 24 h.
Extraction and analysis of limonoids. Seeds (80 g) were ground into a meal and placed in a Soxhlet extractor. The meal was washed thoroughly with hexane to remove oils and then extracted with MeOH. After the MeOH was evaporated from the extract, the residue was extracted with a mixture of CH2CI 2 and H=O (1 : 1 ) at pH 3.0. The CH2CI2 fraction was used for analysis of the limonoid aglycones. The aqueous solution contained limonoid glucosides. The CH2CI2 extract was spotted onto TLC plates along with standards, and developed with three different solvent systems; (a) cyclohexane:EtOAc (2:3), (b) CH2CI2:MeOH (49:1) and (c) EtOAc:CH2CI2 (2:3). Silica gel HLF plates were purchased from Analtech (Newark, DE). Limonoid concentrations were determined quantitatively by HPLC using a C18 reversed-phase column and isocratic elution with 49% H20, 41% MeOH and 10% CH3CN at a flow rate of 1 ml/min. Limonoids were detected by UV absorption at 210 nm (Herman et al., 1992). Limonoid glucosides were analysed by HPLC with procedures described previously (Herman et al., 1992; Fong et al., 1993a). The analytical H PLC column used for limonoid analysis was a C-18 reverse phase, partisil ODS-3, 4.6 x 250 mm, particle size 5 Ilm was purchased from AIItech Associates (Deerfield, IL).
Extraction and analysis of flavonoids. Dried leaves were ground to a powder with a small coffee bean mill. For sample extraction, about 250 mg of the dried powder of each sample was extracted three times with 1 ml of a 1:1 DMSO: MeOH mixture at 50°C. Each extraction was allowed to soak for at least 1 h. The combined extraction mixtures were centrifuged for 1 min in a tabletop micro-centrifuge to pellet the cell debris. The resulting supernatant of each sample extract was filtered through a 0.45 micron syringe filter into an auto sampler vial for injection on to an automated HPLC system. The HPLC column used in the flavonoid analysis was a Licrosorb C18 reverse phase analytical column (25 x 0.4 cm, ODS 3, 5 micron particle size) which was used in conjunction with a UV diode-array detector. The gradient elution schedule consists of an initial 2 rain run of 80% 0.01 M H3PO4 and 20% MeOH followed by a linear gradient to 100% MeOH over 55 min at a flow rate of 1 ml per min (Kanes et al., 1993). Peak area percentages and extinction coefficients are calculated from chromatograms of standards detected at 285 nm. This wavelength was chosen for monitoring because all flavonoids examined in this study absorb at this wavelength and extinction coefficients were similar when calculated from several purified flavonoid standards. The relative concentrations were determined from peak areas using the extinction coefficients determined from standards.
R e s u l t s a n d Discussion Analysis o f the l imonoids in papeda taxa All citrus taxa and their hybrids wh ich have been examined have been shown to accumulate the bit ter-tast ing l imonoid l imonin through the l imonin b iosynthet ic pa thway (Hasegawa and Herman, 1992; Hasegawa and Ifuku, 1994). A second l imonoid biosynthet ic pa thway result ing in the format ion of the calamin group of l imonoids exists in the cultivars related to the genus Fortunella and their hybrids; the l imonin b iosynthet ic pa thway also exists in these cult ivars (Hasegawa and Herman, 1992; Hasegawa and Ifuku, 1994). A third l imonoid b iosynthet ic pa thway exists in the cult ivars related to C. ichangensis and its hybrids (Bennet t et aL, 1988; Hashinaga and Hasegawa, 1989; Herman et aL, 1989; Hashinaga et aL, 1990; Ohta et al., 1992; B e r h o w et al., 1994; Hasegawa and Ifuku, 1994). The end product of the ichangensin l imonoid pathway, ichangensin, is tasteless and its accumulat ion at the expense of l imonin wou ld el iminate l imonin bitterness in citrus juices. In this path- way, ichangensis is b iosynthesized f rom nomil in via deacety lnomi l in and deacety lnomi l inate.

L I M I N O I D A N D F L A V O N O I D IN CITRUS VARIETIES 239
In order to examine the extent that the genes for the ichangensin pathway are expressed in cultivars related to C. ichangensis, we have analysed the limonoids in seeds of some of the other papeda taxa. Some of the hybrids between C. ichangensis and other citrus taxa, such as C. junos '¥uzu; C. sudachi and C. sphaerocarpa "Mochiyu' and 'Yuko' have also been shown to possess ichangensin and its glucoside derivative, ichangensin 17-J3-D-glucopyranoside in their fruit tissues and seeds (Hashinaga and Hasegawa, 1989; Hashinaga et aL, 1990; Ozaki et aL, 1991; Ohta et aL, 1992; Berhow et aL, 1994). However, the seeds of C. hanaju and C. wilsonii 'lchang lemon'--two other C. ichangensis hybrids--do not contain ichangensin, but instead accumulate relatively high concentrations of deacetylnomilin, a precursor of ichangensin, and its corresponding glucoside (Ohta et al., 1992; Berhow et al., 1994).
As shown inTable 1, the presence of the limonin pathway in the papedas C. latipes and C. hystrix, the hybrids C. macrophylla, C. hystrix 'Cabayao' and C. webberii 'Montana' is clearly shown by the occurrence of limonin and to a lesser degree the limonin precursors nomilin, obacunone and ichangin in the seeds examined. However, none of these seeds contain any ichangensin, or the very high levels of deacetylnomilin, that indicates the presence of the initial stages of the ichangensin biosynthetic pathway. This shows that these other papeda species do not have this pathway and indeed the pathway is only found in cultivars directly related to C. ichangensis. The limonoid content of the non-ichangensis papedas resemble more closely that of the 'true' citrus species such as the mandarins, the citrons and the pummelos. Indeed, the low level of glucosides found in seeds from mature fruit seems to indicate a relationship with the pummelos, which have also been shown to have fairly low levels of limonoid glucosides (Fong et aL, 1993b).
Analysis of the flavonoids in papeda taxa This report is the first thorough examination of a large number of individual
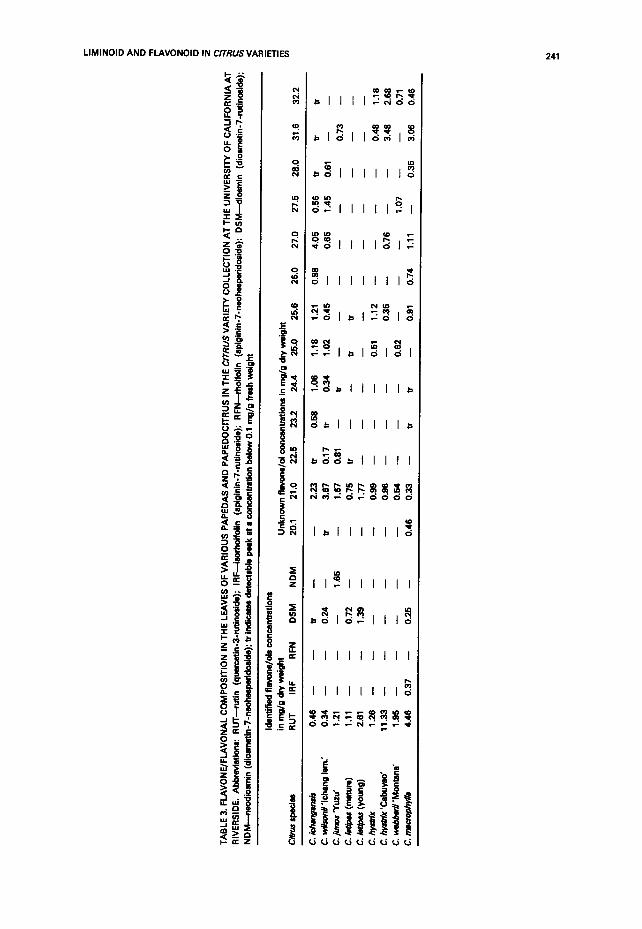
flavonoids in the leaves of cultivars in the Papeda subsection. While there have been some fairly substantial characterizations of the flavonoids in the major commercial species, at best only one or two of the papeda cultivars have been examined. Often these examinations have only been in regard to a few of the major flavonoids, especially naringin, hesperidin, narirutin and neohesperidin. We were able to look at the concentrations of 10 major flavanone glycosides (Table 2) five major identified flavone/ol glycosides and an additional 14 as yet unidentified flavone/ols (Table 3) in the papeda specimens present in the Riverside collection.
TABLE 1. L I M O N O I D COMPOSIT ION OF SEEDS OF VARIOUS PAPEDAS A N D PAPEDOCITRUS IN THE CITRUS
V A R I E T A L COLLECTION A T THE UNIVERSITY OF CAL IFORNIA A T RIVERSIDE. Abbreviat ions: L IM- - l imon in ; N O M - -
nomil in; O B A ~ o b a c u n o n e ; DAN--deecety lnomi l in ; I C N ~ c h a n g i n ; UNK---unident i f ied l imonoid aglycone; LG- - l imon in 17-
J3-D-glucopyranoside; N A G ~ o m o n i l i c acid 17-~-D-gfucopyranoeide; N G ~ n o m i l i n 17-~-o-g lucopyrenoside; D G - -
deecetylnornil in 17-[~-o-glucopyranoside; O G ~ b a c u n o n e 17-[~-o-glucopyranoside
L imonoid aglycones in seeds (ppm) Umono id glucosides in seeds (ppm)
Citrus species L I M N O M OBA DAN ICN UNK LG NAG NG DG OG
C/bus/aO~oes 1728 1512 756 1512 227 202 151 - - 403 - - - -
C#rus hystedx "Cabuyao "° 824 1098 261 - - - - - - 412 412 828 - - - -
300 800 90 60 - - - - 260 440 480 - - - -
C#rus hystedx 1987 858 909 - - - - - - 110 - - 346 - - 160
810 550 120 20 - - - - 56 - - 120 - - - -
C/bus webbadi "Montana" 5552 872 123 476 18 - - 1745 634 2380 - - - -
Citrus mecrophylla 4554 911 569 51 - - - - 256 - - 1025 - - - -
*The 'Cabuyao" cult ivars in the Rive~ ide Citrus Variety Collect ion have been labeled a C. hyst/ix varieties in the Col lect ion
records, but are referred to as a C. macroptera variety by Swingle.
240 M . A . BERHOW ETAL.
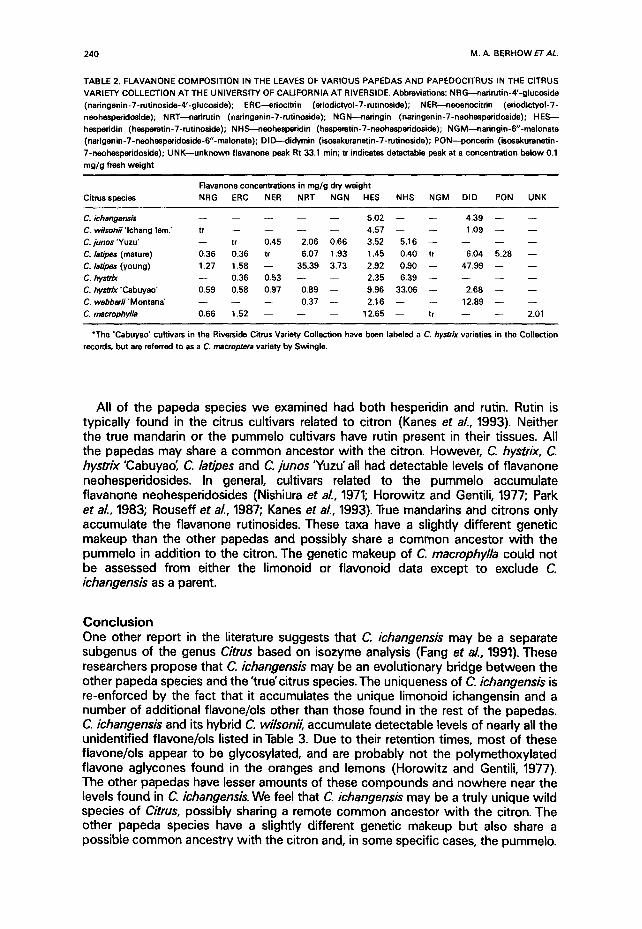
TABLE 2. FLAVANONE COMPOSIT ION IN THE LEAVES OF VARIOUS PAPEDAS A N D PAPEDOCITRUS IN THE CITRUS
VARIETY COLLECTION AT THE UNIVERSITY OF CAL IFORNIA AT RIVERSIDE, Abbreviat ions: NRG~nar i ru t in -4 ' -g lucos ide
(nar ingenin-7-rut inoside-4'-g lucoside); ERC--edoci t r in (edodictyol-7-rut inoside); NER---neoeriocitr in (er iodictyol -7-
neohespe¢idoside); NRT---narirutin (naringenin-7-rut inoside); NGN---naringin (naringenin-7-neohesperidoside); H E S - -
hesperidin (hesperet in-7-rut inoside); NHS---~eohesperidin (hesperetin-7-neohesperidoside); NGM--~a r ing in -6 " -ma lona te
(nar igenin-7-neohesper idoside-6"-malonate); DID--< l idymin ( isosakurenetin-7-rut inoside); PON---poncerin (isosakuranetin-
7*neohesperidoside); U N K - - u n k n o w n f lavanone peak Rt 33,1 min; tr indicates detectable peak at a concentrat ion be low 0.1
mg/g fresh weight
Flavanone concentrations in mg/g dw we igh t
Citrus species NRG ERC NER NRT NGN HES NHS N G M DID PON UNK
C. i c h a n g e n s i s . . . . . 5.02 - - - - 4 .39 - - - -
C. wilsonii ' lchang lem.' tr . . . . 4.57 - - - - 1.09 - - - -
C. junos 'Yuzu" - - tr 0.45 2.06 0.66 3.52 5.16 . . . . .
C. laO"pes (mature) 0.36 0.36 tr 6,07 1,93 1.45 0.40 tr 6.04 5.28 - -
C. latipe~ (young) 1.27 1.58 - - 35.39 3.73 2.92 0.90 - - 47.99 - - - -
C. hystrix - - 0.36 0.53 - - - - 2.35 6.39 . . . .
C. hystrix "Cabuyao' 0.59 0.58 0.97 0.89 - - 9.96 33.06 - - 2 .68 - - - -
C. webberii'Montana' - - - - - - 0.37 - - 2.16 - - - - 12.89 - - - -
C, macrophylla 0.66 1,52 - - - - - - 12.65 - - tr - - - - 2.01
*The "Cabuyao" cultivars in the Riverside Citrus Variety Collection have been labeled a C. hystrix varieties in the Collect ion
records, but are referred to as a C. macroptera variety by Swingle.
All of the papeda species we examined had both hesperidin and rutin. Rutin is typically found in the citrus cultivars related to citron (Kanes et al., 1993). Neither the true mandarin or the pummelo cultivars have rutin present in their tissues. All the papedas may share a common ancestor with the citron. However, C. hystrix, C. hystrix "Cabuyao; C. latipes and C. junos 'Yuzu' all had detectable levels of flavanone neohesperidosides. In general, cultivars related to the pummelo accumulate flavanone neohesperidosides (Nishiura et al., 1971; Horowitz and Gentili, 1977; Park et al., 1983; Rouseff et aL, 1987; Kanes et aL, 1993). True mandarins and citrons only accumulate the flavanone rutinosides. These taxa have a slightly different genetic makeup than the other papedas and possibly share a common ancestor with the pummelo in addition to the citron. The genetic makeup of C. rnacrophylla could not be assessed from either the limonoid or flavonoid data except to exclude C. ichangensis as a parent.
Conclusion One other report in the literature suggests that C. ichangensis may be a separate subgenus of the genus Citrus based on isozyme analysis (Fang et aL, 1991), These researchers propose that C. ichangensis may be an evolutionary bridge between the other papeda species and the 'true" citrus species. The uniqueness of C. ichangensis is re-enforced by the fact that it accumulates the unique limonoid ichangensin and a number of additional flavone/ols other than those found in the rest of the papedas. C. ichangensis and its hybrid C. wilsonii, accumulate detectable levels of nearly all the unidentified flavone/ols listed in Table 3. Due to their retention times, most of these flavone/ols appear to be glycosylated, and are probably not the polymethoxylated flavone aglycones found in the oranges and lemons (Horowitz and Gentili, 1977). The other papedas have lesser amounts of these compounds and nowhere near the levels found in C. ichangensis. We feel that C. ichangensis may be a truly unique wild species of Citrus, possibly sharing a remote common ancestor with the citron. The other papeda species have a slightly different genetic makeup but also share a possible common ancestry with the citron and, in some specific cases, the pummelo.
r-
o z z _o
c~
Z,
TA
BL
E 3
. F
LA
VO
NE
/FL
AV
ON
AL
C
OM
PO
SIT
ION
IN
TH
E L
EA
VE
S O
F V
AR
IOU
S
PA
PE
DA
S A
ND
P
AP
ED
OC
ITR
US
IN
TH
E C
ITR
US
VA
RIE
TY
CO
LL
EC
TIO
N
AT
TH
E U
NIV
ER
SIT
Y
OF
CA
LIF
OR
NIA
A
T
RIV
ER
SID
E.
Ab
lxe
vle
tio
ns:
R
UT
~ru
tin
(q
uw
ceti
n-3
-ru
tin
osi
de
);
IRF
~is
orh
oif
oli
n
(al~
gin
in-7
-~n
~k
le);
R
FN
---c
I~Ii
n
(ap
igin
in-7
-n~
d~
de
)"
DS
k~
--d
i~in
(d
i~ti
n-7
-~in
ce
k:l
e);
~
ND
M--
-ne
od
iosm
in
(dio
#m
leti
n-7
-ne
oh
esp
ed
do
sid
e);
tr
ind
ica
tes
de
tect
ab
le I
xmk
at
a co
nce
ntr
ati
on
b
elo
w 0
.1
mg
/g
fi'u
h
we
igh
t X
:o
C/~
u,
spec
ies
Ide
nti
fie
d f
lavo
ne
lob
co
nce
ntr
ati
on
s
in m
g/g
d
ry w
eig
ht
RU
T
IRF
R
FN
D
SM
N
DM
Un
kno
wn
fl
evo
ne
/ol
con
cen
tra
tio
ns
in m
g/g
d
ry w
eig
ht
20
.1
21
.0
22
.5
23
.2
24
.4
25
.0
25
.6
26
,0
27
.0
27
.5
28
.0
31
.6
32
.2
c. k
:hJn
gwl~
0.
46
--
--
C.
w//
son
//"l
cha
ng
le
m.'
0
.34
--
--
C. ]
un
os
"Yu
zu"
1.21
m
--
C.
/at/
peS
(m
atu
re)
1.11
--
--
C./
ag
pe
s (y
ou
ng
) 2.
61
--
--
C.
hy~
/r/x
1.
26
--
--
C,
~ 'C
ab
uya
o'
11
.33
--
--
C.
we
bb
ed
/"M
on
tan
a"
1.9
5
--
--
C.
ma
cro
ph
y//a
4
.46
0
.37
--
tr
--
--
2.2
3
tr
0.5
8
1.0
6
1.1
8
1.21
0
.98
4
.06
0
.56
tr
tr
tr
0.2
4
--
tr
3.6
7
0.1
7
tr
0.3
4
1.0
2
0.4
5
--
0.6
5
1.4
5
0.5
1
--
--
--
1.6
5
--
1.6
7
0.8
1
--
tr
..
..
..
0
.73
--
0.7
2
--
--
0.7
5
tr
--
--
tr
tr
..
..
..
1.3
9
--
--
1 .~
.
..
..
..
..
..
I I
I O
'~
I I
I O
e5
1
1.1
2
..
..
O
.~
1.1
8
--
I I
0"~
.
..
.
0'3
5
--
0"7
6
I I
3"4
8
2"6
8
--
--
--
0"5
4
--
--
--
0"6
2
--
--
--
1 "0
7
--
--
0"71
0"2
5
--
0'4
6
0"3
3
--
tr
1T
--
0.9
1
0.7
4
1.11
I
0.3
5
3.0
6
0,4
6

242 M.A. BERHOW ETAL.
Acknowledgements---Research conducted at the USDA, ARS, Fruit and Vegetable Chemistry Laboratory, Pasadena, California, closed by the USDA in 1994. Names are necessary to report factually on available data; however, the USDA neither guarantees nor warrants the standard of the product, and the use of the name by the USDA implies no approval of the product to the exclusion of others that may also be suitable.
References Albach, R. F. and Redman, G. H. (1969) Composition and inheritance of flavanones in citrus fruit.
Phytochemistry 8, 127-143. Barret, H. C. and Rhodes, A. M. (1976) A numerical taxonomic study of affinity relationships in cultivated
Citrus and its close relatives. Syst. Bot. 1, 105-136. Bennett, R. D., Herman, Z. and Hasegawa, S. (1988) Ichangensin: a new citrus limonoid. Phytochemistry
27, 1543-1 545. Berhow, M. A., Omura, M., Ohta, H., Ozaki, Y. and Hasegawa, S. (1994) Limonoids in seeds of three
Citrus hybrids related to Citrus ichangensis. Phytochemis~/ 36, 923-925. Fang, D., Xiao, S., Zhang, W. and Wan, S. (1991 ) Studies on the taxonomy and evolution of Citrus species
by isozyme analysis. Proc. Int. Ci~us Syrup. Guangzhou, China, November, 1990 (Bangyan, H. and Qian, Y., eds), pp. 237-246. International Academic Publishers, Beijing, China.
Fong, C. H., Hasegawa, S., Miyake, M., Ozaki, Y., Coggins, C. W. and Atkin, D. R. (1993a) Limonoids and their glucosides in Valencia orange seeds during fruit growth and development. J. Agric. Food Chem. 112-115.
Fong, C. H., Suhayda, C., Berhow, M. and Hesegawa, S. (1993b) Limonoids in Pummelo. Citrus Research Conference Abstracts of Papers, p. 10, Pasadena, California.
Hasegawa, S. and Herman, Z. (1992) Biosynthesis of limonoids in Citrus. In Secondary Metabolite Biosynthesis and Metabolism (Petroski, R. J. and McCormick, S. P., eds), pp. 305-317. Plenum Press, New York.
Hasegawa, S. and Ifuku, Y. (1994) Nippon Shokuhin Kogyo Gakkaishi41, 372. Hashinaga, F. and Hasegawa, H. (1989) Limonoids in seeds of Sudachi. J. Japan. Soc. Hort. Sci. 58, 227-
229. Hashinaga, F., Herman, Z. and Hasegawa, S. (1990) Limonoids in seeds of Yuzu. Nippon Shokuhin Kogyo
Gakkaishi 37, 380-382. Herman, Z., Hasegawa, S., Fong, C. H. and Ou, P. (1989) Limonoids in Citrus ichangensis. Agric. Food
Chem. 37, 850-851. Herman, Z., Fong, C. H. and Hasegawa, S. (1992) Analysis of limonids in citrus seeds. In Modern Methods
of Plant Analysis, New Series Vol. 14, Seed Analysis (Linskens, H. F. and Jackson, J. F., eds), pp. 361- 375. Springer-Verlag, Berlin, Germany.
Horowitz, R. M. and Gentili, B. (1977) Flavonoid constituents of Citrus. In Citrus Science and Technology Vo/. I (Nagy, S., Shaw, P. E. and Veldhuis, M. K., eds), pp. 397-426. AVI Publishing Company, Inc., Westport, CT.
Kanes, K., Tisserat, B., Berhow, M. and Vandercook, C. (1993) Phenolic composition of various tissues of Rutaceae species. Phytochemisb7 32, 967-974.
Kefford, J. F. and Chandler, B. V. (1970) The Chemical Constituents of Citrus Fruits. Academic Press, New York.
Nishiura, M., Kamiya, S. and Esaki, S. (1971 ) Flavonoids in Citrus and related genera III. Flavonoid pattern and Citrus taxonomy. Agric. Biol. Chem. 35, 1691-1706.
Ohta, H., Berhow, M., Bennett, R. D. and Hasegawa, S. (1992) Limonoids in seeds of Citrus hanaju. Phytochemis~y 31, 3905-3907.
Ozaki, Y., Miyake, M., Maeda, H., Ifuku, Y., Bennett, R., Herman, D. Z., Fong, C. H. and Hasegawa, S. (1991 ) Ichangensin glucoside in Citrus junos, Citrus sudachi, and C~us sphaerocarpa. Phytochemistry 30, 2659-2661.
Park, G. L., Avery, S. M., Byers, J. L. and Nelson, D. B. (1983) Identification of bioflavonoids from Citrus. Food Technol. 34, 98-105.
Rouseff, R. L., Martin, S. F. and Youtsey, C. O. (1987) Quantitative survey of narirutin, naringin, hesperidin, and neohesperidin in Citrus. J. Agric. Food Chem. 35, 1027-1030.
Scora, R. W. (1988) Biochemistry, taxonomy and evolution of modern cultivated citrus. In Proceedings of the Sixth International Citrus Congress (Goren, R. and Mendel, K., eds), pp. 277-289. Balaban Publishers, Tel Aviv, Israel.
Swingle0 W. R. and Reece, P. C. (1967) The botany of Citrus and its wi ld relatives. In The Citrus Industry Vol. 1 (Reuther, W., Webber, H. J. and Batchelor, L. D., eds), pp. 190-430. University of California, Berkeley, CA.
Tanaka, T. (1969) Misunderstandings with regards to Citrus classification and nomenclature. Bull. Univ. Osaka Pref. Ser. B Agric. 21, 139-145.
Tanaka, T. (1977) Fundamental discussion of Citrus classification. Studia Ci~ilogica 14, 1-6.





























