Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, Sept. 2009, p. 4136–4149 Vol. 77, No. 90019-9567/09/$08.00�0 doi:10.1128/IAI.00453-09Copyright © 2009, American Society for Microbiology. All Rights Reserved.
Macrophage-Mediated Responses to Candida albicans inMice Expressing the Human Immunodeficiency
Virus Type 1 Transgene�‡Mathieu Goupil,1† Emilie Belanger Trudelle,1† Veronique Dugas,1 Catherine Racicot-Bergeron,1
Francine Aumont,1 Serge Senechal,1 Zaher Hanna,2,4,5
Paul Jolicoeur,1,4,5 and Louis de Repentigny1,3*Departments of Microbiology and Immunology1 and Medicine,2 Faculty of Medicine, University of Montreal, Sainte-Justine Hospital,3
Laboratory of Molecular Biology, Clinical Research Institute of Montreal,4 and Division of Experimental Medicine,McGill University,5 Montreal, Quebec, Canada
Received 22 April 2009/Returned for modification 27 May 2009/Accepted 20 June 2009
The critical impairments of innate and adaptive immunity that cause susceptibility to mucosal candidiasisin human immunodeficiency virus (HIV) infection have not been fully determined. We therefore conducted ananalysis of macrophage-mediated responses to Candida albicans in transgenic (Tg) mice expressing Nef, Env,and Rev of HIV type 1 (HIV-1) in CD4� T cells, dendritic cells, and macrophages and developing an AIDS-likedisease (CD4C/HIVMutA Tg mice). Macrophages were successfully recruited to the oral and gastric mucosae ofthese Tg mice in response to chronic carriage of C. albicans and displayed polarization toward an alternativelyactivated phenotype. Functionally, peritoneal macrophages from uninfected Tg mice exhibited increasedphagocytosis of C. albicans and enhanced production of interleukin 6 and monocyte chemoattractant protein1, demonstrating that the HIV-1 transgene independently activates selected macrophage functions. Productionof H2O2 by macrophages from Tg mice primed with gamma interferon and treated with phorbol 12-myristate13-acetate or C. albicans was moderately reduced, but expression of the HIV-1 transgene did not alterproduction of nitric oxide or reduce killing of C. albicans. A knockout of the inducible nitric oxide synthase(NOS2) gene in these Tg mice did not augment oral or gastrointestinal burdens during chronic carriage of C.albicans or cause systemic dissemination, likely due to a redundancy provided by partially preserved productionof H2O2 and oxygen-independent candidacidal mechanisms. Thus, the macrophage response to C. albicans islargely preserved in these Tg mice, and no functional macrophage defect appears to primarily determine thesusceptibility to mucosal candidiasis.
Oropharyngeal candidiasis (OPC) is the most frequent op-portunistic fungal infection among human immunodeficiencyvirus (HIV)-infected patients (64). Although the incidence ofOPC in HIV infection is sharply reduced by highly activeantiretroviral therapy (45), it remains a common coinfectionworldwide. The critical impairments of innate and adaptiveimmunity that are responsible for the onset and maintenanceof mucosal candidiasis in HIV infection have not been fullydetermined (15, 25). A correlation has been established in HIVinfection between symptomatic OPC and reduced CD4� cellcount (6, 46, 55), HIV viral load (6, 46), and the developmentof AIDS (55). Studies conducted with experimentally infectednormal, nude, and cytokine-specific gene knockout mice indi-cated that host defense against OPC requires intact Th1- andTh17-mediated immune responses to Candida albicans, includ-ing production of interleukin 12 (IL-12), CD4� T-cell augmen-tation of monocyte and polymorphonuclear leukocyte (PMN)
functions, and mucosal production of nitric oxide (NO) (1, 7,11, 17–21, 34, 61). Using a model of mucosal Candida infectionin transgenic (Tg) mice expressing HIV-1 Nef in CD4� T cells,dendritic cells, and macrophages which closely mimics the clin-ical and pathological features of candidal infection in humanHIV infection (14), we have previously shown that alteredCD4� T-cell phenotype and function determine the suscepti-bility to chronic carriage of C. albicans in these Tg mice (37).However, PMNs from the Tg mice were unimpaired in theircapacity to produce an oxidative burst and to phagocytose andkill C. albicans in vitro, and depletion of PMNs in these Tgmice did not alter the oral or gastrointestinal burdens of C.albicans or cause systemic dissemination (42). Accordingly, thedefective anti-Candida effector mechanisms that render theseTg mice susceptible to mucosal candidiasis have not yet beenidentified.
Oral colonization and infection of mice with C. albicanstrigger macrophage recruitment to the mucosa of the oralcavity (9), stomach (10, 71), and cecum (12), suggesting thatthese cells play a role in resistance to mucosal candidiasis(68). Activated macrophages have the capacity to kill C.albicans by their production of the reactive oxygen interme-diates (ROIs) O2
� and H2O2, by the formation of peroxyni-trite from O2
� and the reactive nitrogen intermediate NO,and by oxygen-independent candidacidal mechanisms (68–71,73). The participation of macrophages in host resistance
* Corresponding author. Mailing address: Department of Micro-biology and Immunology, Sainte-Justine Hospital and University ofMontreal, 3175 Cote Ste-Catherine, Montreal, QC, H3T 1C5, Can-ada. Phone: (514) 345-4643. Fax: (514) 345-4860. E-mail: [email protected].
† These authors contributed equally to this work.‡ Supplemental material for this article may be found at http://iai
.asm.org/.� Published ahead of print on 29 June 2009.
4136
on March 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

has been demonstrated by the enhanced susceptibility ofsevere combined immunodeficiency (SCID) mice to dissem-inated candidiasis of gastrointestinal origin after treatmentwith poly(I-C), an inhibitor of macrophage candidacidal ac-tivity (33).
Treatment of human monocyte-derived macrophages withHIV-1 Nef protein or infection of these cells with HIV-1 alterscellular signal transduction pathways and specifically activatesNF-�B, STAT1 and STAT3, mitogen-activated protein ki-nases, and genes for several inflammatory factors, includingmacrophage inflammatory protein 1�, macrophage inflamma-tory protein 1�, IL-1�, IL-6, and tumor necrosis factor alpha(TNF-�) (5, 24, 40, 59). Therefore, the anticandidal propertiesof macrophages could be altered either directly by the expres-sion of HIV-1 gene products within this cell population orindirectly by inadequate cytokine signaling from defectiveCD4� T cells. In several investigations producing conflictingresults, phagocytosis and killing of C. albicans by blood mono-cyte-derived macrophages from HIV-infected patients havebeen found to be either normal (56, 57) or reduced (13),possibly by HIV Nef (35, 62).
With the recognition that classically activated (M1) macro-phages (27, 44) primarily mediate the effector arm of a CD4�
T-cell-dependent protective Th1 adaptive immune response bytheir production of ROIs and reactive nitrogen intermediateswhich kill C. albicans (8, 43, 52, 54, 63), we asked whether adefective mucosal macrophage response to C. albicans contrib-utes to the phenotype of chronic oral candidiasis in these Tgmice expressing HIV-1. The likelihood of such a defect wasconsidered significant because CD4� T cells are quantitativelyand functionally defective in these Tg mice, and these alter-ations of CD4� T cells determine at least in part the suscep-tibility of these animals to chronic carriage of C. albicans (37).We found that macrophages from these Tg mice display apolarization toward an alternatively activated phenotype andare successfully recruited to the mucosa in response to C.albicans. Although the production of H2O2 was modestly re-duced, the production of NO and the killing of C. albicans bymacrophages were both unaltered by expression of the HIV-1transgene, and no further augmentation of oral burdens of C.albicans was found in NOS2�/� gene-deficient Tg mice. Thus,the macrophage response to C. albicans is largely preserved inthese Tg mice, and no functional macrophage defect appearsto primarily determine susceptibility to mucosal candidiasis.
MATERIALS AND METHODS
Generation of Tg mice expressing HIV-1. The Tg mice expressing Nef, Env,and Rev of HIV-1 in CD4� T cells, dendritic cells, and macrophages (CD4C/HIVMutA Tg mice) have been described elsewhere (30). CD4C/HIVMutA mutantDNA harbors mouse CD4 enhancer and human CD4 promoter elements to drivethe expression of HIV-1 genes in CD4� CD8� and CD4� thymocytes, in pe-ripheral CD4� T cells, and in macrophages and dendritic cells. Founder mouseF21388 was bred on the C3H background. Animals from this line express mod-erate levels of the transgene, with 50% survival at 3 months (14). Several HIV-1genes (gag, pol, vif, vpr, tat, and vpu) are mutated in the CD4C/HIVMutA DNA,whereas env, rev, and nef are intact. The generation of CD4C/HIVMutG micerevealed that selective expression of the nef gene is required and sufficient toelicit an AIDS-like disease in these Tg mice (30). This disease is characterized byfailure to thrive, wasting, severe atrophy and fibrosis of lymphoid organs, loss ofCD4� T cells, interstitial pneumonitis, and segmental glomerulosclerosis asso-ciated with tubulointerstitial nephritis (30).
Homozygous NOS2�/� mice were obtained from Foo Y. Liew and contained
a disruption of the inducible nitric oxide synthase (iNOS) 2 gene (74). They werebred for at least six generations on the C3H background and then with CD4C/HIVMutG Tg mice to generate homozygous NOS2�/� or heterozygous NOS2�/�
CD4C/HIVMutG Tg mice.Specific-pathogen-free male and female Tg mice and non-Tg littermates were
housed in sterilized individual cages equipped with filter hoods, supplied withsterile water, and fed with sterile mouse chow. All animal experiments wereapproved by the Animal Care Committee of the University of Montreal.
Animal models of candidiasis. Oral inoculation with C. albicans LAM-1,assessments for signs of morbidity, quantification of C. albicans in the oralcavities of individual mice, and determination of burdens of C. albicans in thegastrointestinal tract and internal organs were conducted as described previously(14). Animals reaching predetermined morbidity end points were designatedpremortem and euthanized by CO2 inhalation.
In separate experiments, mice were infected intraperitoneally with 2 � 106
blastoconidia of C. albicans LAM-1 in 0.5 ml of 0.01 M phosphate-bufferedsaline, pH 7.4 (PBS). Sham-infected mice received PBS alone, and control micewere uninfected and did not receive PBS. Three days later, peritoneal cells werecollected by washing the peritoneal cavity twice with 8 ml of PBS. None of themice attained the morbidity end points, and all animals were euthanized by CO2
inhalation at the end of the experiment.Immunohistochemical analysis of mucosal immune cell populations. Tg mice
and control non-Tg littermates, infected or not infected with C. albicans, wereanesthetized with 0.3 ml of a solution containing 1.5 ml of ketamine hydrochlo-ride (Bimeda-MTC, Cambridge, Ontario, Canada), 0.1 ml of xylazine (Bimeda-MTC), and 8.4 ml of sterile PBS. From each animal, the spleen, cheeks, tongue,and stomach, including the cardial-atrium fold, were dissected and placed in aCryomold (Sakura Finetek, Torrance, CA) filled with OCT (10.24% [wt/wt]polyvinyl alcohol, 4.26% [wt/wt] polyethylene glycol, and 85.50% [wt/wt] nonre-active ingredients; Sakura Finetek). The base molds with tissue were depositedon a metal flat work surface placed on dry ice and were stored at �80°C untilsectioning. Sections (5 �m thick) were cut and deposited onto Superfrost Plusslides (Fisher Scientific, Ottawa, Ontario, Canada). The primary antibodies usedwere rat anti-mouse anti-F4/80 antigen, hamster anti-mouse anti-CD11c (Sero-tec, Raleigh, NC), and rat anti-mouse anti-CD4 or -CD8 (BD Biosciences,Mississauga, Ontario, Canada). Immunohistochemical staining was done by anindirect peroxidase-conjugated streptavidin procedure. All incubations weredone at room temperature. Sections were fixed for 15 s in 70% ethanol and 5 minin cold acetone and then washed three times in PBS. The sections were treatedfor 15 min with 0.03% H2O2 in PBS to inactivate endogenous peroxidase. Afterone further washing, the sections were incubated for 20 min with the followingserum dilutions before being labeled with primary antibodies for 1 h: 10%normal rabbit serum (Vector Laboratories, Burlington, Ontario) and 2.5% nor-mal mouse serum (Sigma-Aldrich, St. Louis, MO) for anti-F4/80, -CD4, and-CD8; 1.5% normal goat serum (Vector) for anti-CD11c. The sections werewashed three times, and biotinylated secondary antibodies (Vector) were addedfor 30 min. After further washing, the bound antibodies were detected using theVectastain Elite ABC kit (Vector). The sections were washed three times, andreaction sites were visualized with diaminobenzidine (Vector) and counter-stained with Meyer’s hematoxylin. Spleens from non-Tg mice were used aspositive controls, and negative controls were obtained by incubating sections withthe appropriate isotype controls.
Double fluorescence labeling of mucosal macrophages. For detecting classi-cally and alternatively activated macrophages (M1 and M2 macrophages, respec-tively), double staining of tissue sections was carried out by a sequential indirectimmunofluorescence technique using rat anti-mouse F4/80 (Serotec) and ratanti-mouse monocyte chemoattractant protein 1 (MCP-1) (Hycult Biotechnol-ogy, Uden, The Netherlands) or rat anti-mouse F4/80 and purified rat anti-mouse CD206 (BioLegend) as primary antibodies, respectively. All incubationswere done in the dark for 1 h at room temperature for F4/80 and CD206 and for18 h at 4°C for MCP-1. Sections were incubated for 30 min with 10% normalrabbit serum (Vector) and 2.5% normal mouse serum (Sigma-Aldrich) for 1 hprior to labeling with primary antibodies. The sections were washed three times,and biotinylated secondary antibodies (Vector) were added for 30 min. Afterfurther washing, sections were incubated with Texas Red streptavidin (Vector)for anti-F4/80 and with fluorescein streptavidin (Vector) for anti-CD206 andanti-MCP-1. Sections were viewed by confocal microscopy (AxioVert LSM 510Meta; Carl Zeiss, Jena, Germany).
As a positive control for detection of M1 and M2 macrophages, 1 � 106 restingor elicited peritoneal macrophages harvested from uninfected non-Tg mice wereincubated for 24 h at 37°C in 300 �l of supplemented RPMI medium containing100 ng/ml of lipopolysaccharide (LPS) (Sigma-Aldrich) or 2 � 102 U/ml of
VOL. 77, 2009 RESPONSE TO C. ALBICANS IN HIV GENE-EXPRESSING Tg MICE 4137
on March 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

mouse IL-4 (Cedarlane, Hornby, Ontario, Canada), respectively. Fluorescencelabeling was performed as described previously.
Flow cytometry analysis of mucosal macrophages. Single-cell suspensions ofsmall intestine and oral mucosal tissue were prepared using collagenase digestion aspreviously described (37). Cells were surface stained with different combinations ofanti-mouse anti-CD45, anti-CD11b, anti-I-A� (MHC class II alloantigen), anti-CD11c (BD Biosciences), and anti-F4/80 (Invitrogen, Carlsbad, CA) fluorescence-labeled monoclonal antibodies and their respective isotype controls (BD Biosciences
or Invitrogen). Small intestine cells were used as a control for comparison of flowcytometry profiles with the oral mucosal cell population. Cell surface marker analysiswas conducted on a FACSCalibur flow cytometer (BD Biosciences) equipped withCellQuest software. Data were acquired for 10,000 events by gating on CD45� cells,which composed 3% of mucosal cell suspensions. The percentages of oral mucosalmacrophages measured using two different combinations of antibodies (CD45�,I-A��, CD11b�, F4/80�; CD45�, CD11c�, CD11b�, F4/80�) (see Fig. S1 in thesupplemental material) did not differ significantly (P � 0.05).
FIG. 1. Immunohistochemical analysis of stomach tissue from CD4C/HIVMutA Tg and non-Tg mice. Tissues were obtained from mice both 70days after infection (or sham infection) with C. albicans (mean age, 124 days; range, 110 to 143 days) and premortem (mean age, 150 days; range,125 to 227 days). Tissues were stained with hematoxylin phloxine saffron (HPS) or Gomori-Grocott stain or with antibodies against F4/80, CD11c,CD4, and CD8 (see Materials and Methods). Squares represent areas enlarged in insets. Arrows indicate polymorphonuclear leukocytes (HPSpanel inserts, 70-day and premortem infected Tg mice). Images are representative of at least three experiments with consistent results.
4138 GOUPIL ET AL. INFECT. IMMUN.
on March 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
Phagocytosis and killing of C. albicans by peritoneal macrophages. Elicitedperitoneal macrophages were harvested 3 days after mice were given an intra-peritoneal injection of 1 ml of sterile 0.04% sodium thioglycolate (Sigma-Al-drich). Mice were anesthetized with ketamine/xylazine, and the peritoneal cellswere harvested by washing with PBS. The cells were washed with PBS, treatedwith lysing buffer to remove erythrocytes, washed again with PBS, and resus-pended in RPMI medium (Gibco, Grand Island, NY) supplemented with 20 mMHEPES buffer, 2 mM L-glutamine, 100 U/ml penicillin and streptomycin, and 50�g/ml of gentamicin. Elicited peritoneal cell suspensions (EPCs) from Tg andnon-Tg mice contained 80% macrophages, and cell viability was �95% by trypanblue exclusion.
To assay phagocytosis, C. albicans LAM-1 was grown in Sabouraud dextrosebroth (BD Diagnostics, Sparks, MD) at 30°C for 18 h with rotary agitation. Thecells were collected by centrifugation and washed twice with PBS. Then, 2.5 �
105 EPCs were incubated with fluorescein isothiocyanate-labeled, heat-inacti-vated (90°C, 45 min) C. albicans blastoconidia in 100 �l of supplemented RPMImedium at different EPC-to-blastoconidium ratios for 60 min at 37°C. Phagocy-tosis was stopped by cooling the samples to 4°C. As a negative control, phago-cytosis was conducted at 4°C (1:10 ratio) in the presence of cytochalasin B (5�g/ml; Sigma-Aldrich) (23). Ethidium bromide (10 �g/ml) was added, and thepercentage and mean fluorescence of macrophages with phagocytosed C. albi-cans were measured by flow cytometry (65).
To assay the killing of C. albicans, 2.5 � 105 EPCs were incubated withfluorescein isothiocyanate-labeled, live C. albicans blastoconidia in 100 �l ofsupplemented RPMI medium at a 1:1 effector/target (E/T) cell ratio at 37°C inindependent wells. A control assay was performed at 4°C. After EPCs were lysedwith 2.5% sodium deoxycholate, killed C. albicans cells were labeled withethidium bromide and quantitated by flow cytometry.
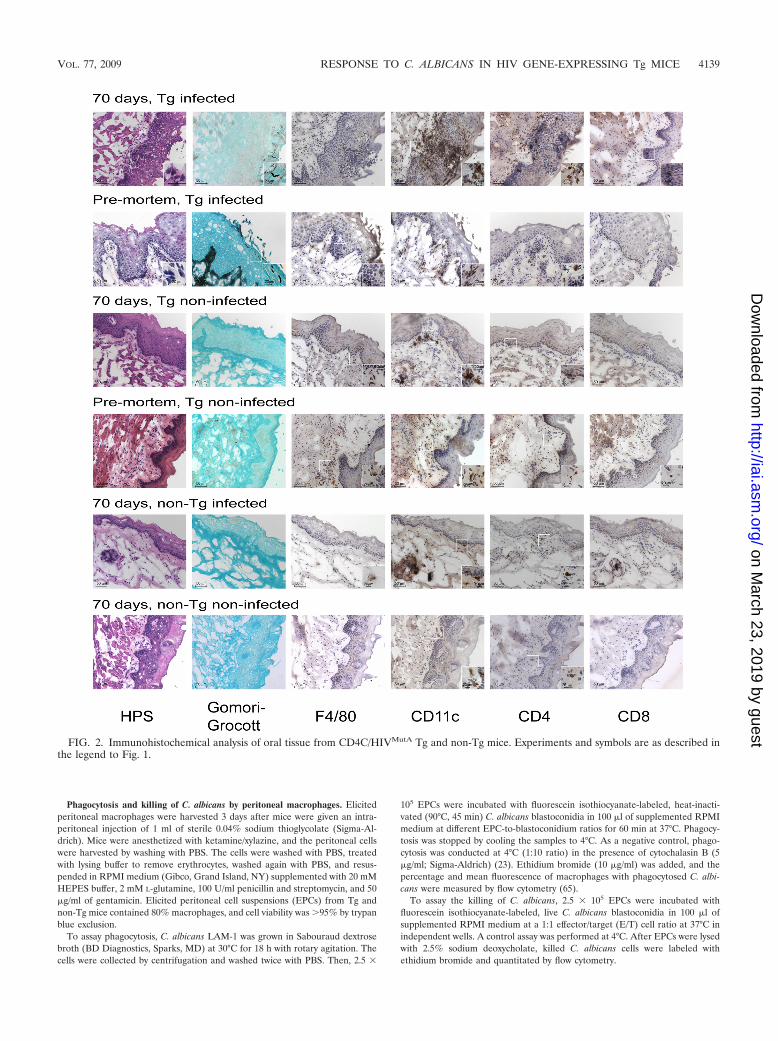
FIG. 2. Immunohistochemical analysis of oral tissue from CD4C/HIVMutA Tg and non-Tg mice. Experiments and symbols are as described inthe legend to Fig. 1.
VOL. 77, 2009 RESPONSE TO C. ALBICANS IN HIV GENE-EXPRESSING Tg MICE 4139
on March 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

Production of H2O2 and NO by peritoneal macrophages. To assay the pro-duction of H2O2, 2 � 106 resting peritoneal cells suspended in 100 �l of sup-plemented RPMI medium were deposited in 96-well cell culture plates (model3595; Corning Inc., Corning, NY) and incubated for 2 h at 37°C in 5% CO2 anda humidified atmosphere. The plates were washed with Hanks balanced salt solu-tion (Gibco) to remove nonadherent cells, and adherent cells were incubated for48 h in 100 �l of supplemented RPMI medium containing 500 U of mouse gammainterferon (IFN-) (Cedarlane). The plates were washed with PBS, and the adherentcells were treated with 4 � 104 live C. albicans blastoconidia, or 1 �M of phorbol12-myristate 13-acetate (PMA) as a positive control, in 100 �l of Krebs-Ringerphosphate glucose containing 50 �M Amplex Red reagent (Invitrogen). After 90min of incubation at 37°C, H2O2 in individual wells was quantitated according to themanufacturer’s protocol (Amplex Red hydrogen peroxide assay kit; Invitrogen).
To assay the production of NO, adherent EPCs in 100 �l of supplementedRPMI medium containing 10% of heat-inactivated fetal bovine serum (Gibco)per well were incubated for 48 h at 37°C with 2.5 � 105 heat-inactivated C.albicans blastoconidia or, as positive and negative controls, 500 U/well of mouseIFN- or 400 �M of NG-monomethyl L-arginine (Sigma-Aldrich), respectively.To quantitate nitrite (NO2
�) as an indicator of NO production, 50 �l of Greissreagent (28) was added to 50 �l of culture supernatant; the mixture was kept inthe dark for 15 min, and the A490 was measured and compared to a standardcurve. In selected experiments, 2.5 � 105 heat-inactivated C. albicans blasto-conidia were added to the EPCs at the start of the 2-h period of adherence,followed by stimulation of the adherent cells with IFN- or without stimulation.
Production of cytokines by peritoneal macrophages. To assay the productionof cytokines, resting peritoneal cells were harvested as described above, andadherent cells were obtained by incubating 2 � 106 cells/well in 96-well cellculture plates for 2 h at 37°C and 5% CO2. After being washed, adherent cellswere treated or not treated with 2.5 � 105 heat-inactivated C. albicans in 100 �lof supplemented RPMI medium and were incubated at 37°C and 5% CO2 for48 h to promote cytokine accumulation in the supernatant. Supernatants werecollected from the wells, and cytokine production was assayed using the BDcytometric bead array mouse inflammation kit (BD Biosciences) according to themanufacturer’s protocol on a FACSCalibur flow cytometer equipped with BDFCAP array software.
Expression of classical and alternative activation marker genes by peritonealmacrophages. Peritoneal cells were harvested from Tg and non-Tg mice infectedintraperitoneally or not infected with C. albicans. RNA was extracted frompelleted peritoneal cells using the RNeasy Mini Plus kit (Qiagen). Ten nano-grams of total RNA was used in a real-time quantitative reverse transcription-PCR (qRT-PCR) with the QuantiTect SYBRGreen RT-PCR kit (Qiagen) toassess expression of 18S, F4/80, TNF-�, NOS2, YM1 (encoding the macrophageprotein Ym1) and ARG1 (arginase 1). The probes used were obtained throughthe GeneGlobe Search Center (Qiagen). Cycle threshold values were normalizedusing the 18S housekeeping gene. In addition, the F4/80 gene was used tonormalize results for macrophage expression of the target genes, since the peri-toneal fluid contains a mixed cell population. Efficiencies of amplification wereevaluated for each gene by assessing dilutions of a reference RNA in triplicate.The expression of the target genes in the different groups of mice was calculatedrelative to that in young non-Tg control animals, not infected with C. albicansand not sham infected with PBS, using REST-MCS beta software version 2(http://www.gene-quantification.de).
Statistical analysis. Kaplan-Meier modeling and a log-rank test were used tocompare survival functions of NOS2�/� and NOS2�/� Tg and non-Tg mice.Fisher’s exact test was utilized to compare the proportions of surviving animalsat the conclusion of the observation period. Oral burdens of C. albicans werecompared as described previously (14, 37, 42). A pairwise fixed reallocationrandomization test (60) was used to analyze gene expression results in real-timeqRT-PCR. All other data were analyzed with SPSS version 14 software (SPSS,Chicago, IL) or PROC MIXED version 9.1 software (SAS Institute, Cary, NC),using analysis of variance, followed by Tukey-Kramer contrast analysis whennecessary. Differences were considered to be significant at a P value of 0.05.
RESULTS
Chronic orogastric candidiasis in Tg mice elicits a mixedinflammatory cell response, including macrophages, in themucosa. To characterize the mucosal inflammatory cell re-sponse which we previously observed in Tg mice infected withC. albicans (14), immunohistochemical analysis was performedon stomach and oral mucosae of Tg and non-Tg mice, infected
or not infected with C. albicans, both premortem and at 70days after infection. At 70 days after infection of Tg mice withC. albicans, hyphae destroyed the cardial-atrium fold of thestomach, penetrated the mucosa to the lamina propria, andwere surrounded by a dense infiltrate of PMNs (Fig. 1). The
FIG. 3. Double-fluorescence analysis of stomach and oral tissuesobtained premortem from CD4C/HIVMutA Tg mice infected with C.albicans. F4/80� macrophages almost uniformly coexpressed the man-nose receptor (CD206) but very rarely MCP-1, consistent with polar-ization toward an alternatively activated (M2) phenotype. Images arerepresentative of three experiments with consistent results.
4140 GOUPIL ET AL. INFECT. IMMUN.
on March 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

submucosa directly below the penetrating hyphae containeddense localized cellular infiltrates mainly composed of F4/80�
macrophages, CD11c� dendritic cells, and CD4� T cells, withfewer CD8� T cells, surrounded by these same cell populationsin the interstitium (Fig. 1). In premortem Tg mice, Candidahyphae penetrated far more deeply, reaching the submucosa,which contained a massive infiltrate of F4/80�, CD11c�,CD4�, and CD8� cells (Fig. 1). In contrast, the gastric muco-sae of uninfected Tg mice were devoid of Candida hyphae andcontained only scattered F4/80� and CD11c� cells in the lam-ina propria, very rare CD4� cells, and no CD8� T cells (Fig.1). Therefore, these Tg mice maintain the ability to mount achronic inflammatory cell response by successfully recruitingall of these cell populations in response to Candida, despite theprofound quantitative depletion and functional impairments ofboth dendritic cells and CD4� T cells previously demonstratedin these animals (14, 37). Cell populations in the gastric mu-cosae of infected or uninfected non-Tg control mice were com-parable to those in the uninfected Tg mice, with the exceptionof more frequent CD4� cells in the non-Tg animals and theiraugmentation by Candida infection as previously observed (14)(Fig. 1).
In contrast to the gastric mucosa, penetration of the oralmucosa (cheeks, tongue) by Candida hyphae in the Tg micepremortem or at 70 days after infection was confined to thesuperficial layer of the epithelium and consistently failed toelicit a PMN response (Fig. 2). As with the stomach, however,Candida infection in premortem Tg mice induced the recruit-ment of F4/80�, CD4�, and CD11c� cells, and more rarelyCD8� cells, within discrete inflammatory foci in the laminapropria and the basal layer of the epithelium directly below theinvading hyphae (Fig. 2). Accordingly, although the inflamma-tory response in the oral mucosa was far more modest than inthe stomach, Candida infection of these Tg mice induced therecruitment of F4/80�, CD11c�, CD4�, and CD8� cells inboth tissues. In contrast to the oral mucosae of the infected Tgmice, the oral mucosae of uninfected Tg mice contained widelyscattered F4/80� and CD11c� cells, very rare CD4� cells, andextremely rare CD8� cell populations (Fig. 2), while unin-fected non-Tg animals showed only rare CD4� cells and a
moderate density of CD11c� cells in the basal layer of theepithelium and lamina propria (Fig. 2). The oral mucosae ofinfected non-Tg animals contained sparse focal infiltrates ofCD4� cells in the lamina propria and basal epithelial layer butno evidence of Candida hyphae (Fig. 2).
Recruited macrophages are polarized toward an alterna-tively activated phenotype. The F4/80� macrophages recruitedto the gastric submucosa and to the oral mucosa of premortemTg mice in response to C. albicans almost uniformly expressedthe mannose receptor (CD206) but only very infrequentlyMCP-1, consistent with an alternatively activated (M2) pheno-type (27) (Fig. 3). Identical results were obtained at 70 daysafter infection of Tg mice with C. albicans (data not shown).
qRT-PCR analysis of classical (TNF-�, NOS2) and alterna-tive (YM1, ARG1) macrophage activation marker gene ex-pression was conducted on peritoneal macrophages harvestedfrom mice infected intraperitoneally with C. albicans or shaminfected with PBS. In both young and old Tg and non-Tg mice,C. albicans infection significantly augmented expression of allmarker genes relative to expression of those genes in younguninfected non-Tg control animals (P 0.05) (Fig. 4). How-ever, C. albicans infection consistently increased expression ofYM1 and ARG1 to a greater degree than TNF-� and NOS2 inboth Tg and non-Tg mice, showing that these macrophageshave a mixed phenotype but are nevertheless polarized to-ward alternative activation in response to C. albicans, inde-pendently of the anatomic site of infection. Interestingly, amore modest but significant (P 0.05) enhancement ingene expression was found in old uninfected Tg controlmice, demonstrating that transgene expression also activatesthese peritoneal macrophages independently of Candida in-fection.
The percentage of macrophages in the oral mucosa is unal-tered by transgene expression but decreased by Candida infec-tion. To quantitatively assess the impact of the HIV-1 trans-gene on the entire oral mucosal macrophage population,multiparametric flow cytometry analysis was conducted onCD4C/HIVMutA Tg mice and non-Tg littermates infected ornot infected with C. albicans. Transgene expression did notsignificantly alter the percentage of oral macrophages in mice
FIG. 4. qRT-PCR analysis of classical (TNF-�, NOS2) and alternative (YM1, ARG1) activation marker gene expression by peritonealmacrophages from CD4C/HIVMutA Tg and non-Tg mice, 7 (young mice) or 70 (old mice) days after intraperitoneal infection with C. albicans (CA)or sham infection with PBS. Data represent the means � standard deviations of results of six independent observations. *, P 0.05 compared tountreated young non-Tg controls.
VOL. 77, 2009 RESPONSE TO C. ALBICANS IN HIV GENE-EXPRESSING Tg MICE 4141
on March 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

infected or not infected with C. albicans, either early or late inthe course of AIDS-like disease (P � 0.05) (Fig. 5). However,oral infection with C. albicans resulted in a decreased percent-age (P 0.05) of this cell population on day 70 after infectionin both Tg and non-Tg animals, compared with the percentagefor infected mice on day 7 or uninfected control animals on day70 (Fig. 5). In contrast to the percentage of macrophages in theoral cavity, however, the percentages of macrophages in thesmall intestine were not significantly different in mice infectedor not infected with C. albicans (P � 0.05).
Macrophages from Tg mice display enhanced phagocytosisbut unaltered killing of C. albicans. The percentage of macro-phages from Tg mice with phagocytosed C. albicans was mod-estly increased (P 0.05) over that of macrophages fromnon-Tg mice at all E/T cell ratios and in both young and old
mice (Fig. 6). Phagocytosis by macrophages from both Tg andnon-Tg mice was enhanced at low (1:5 and 1:10) compared tohigh (1:1) E/T cell ratios at all times of incubation and was alsoenhanced in old compared to young mice (P 0.05). Themean fluorescence of macrophages with phagocytosed C. albi-cans was also enhanced in old compared to young mice at allE/T cell ratios (P 0.001) but was decreased in old but notyoung Tg mice, in comparison to non-Tg mice, at a 1:10 E/Tcell ratio (P 0.001) (data not shown). The percentages of C.albicans killed by macrophages from Tg and non-Tg mice didnot differ (P � 0.05) in either young or old mice (Fig. 6).
Reduced production of H2O2 by macrophages from Tg micein response to C. albicans. Production of H2O2 by peritonealmacrophages, in comparison to that by untreated macro-phages, was significantly augmented (P 0.001) by incubationwith PMA or live C. albicans (Fig. 7A and B) and was signif-icantly greater (P � 0.02) in old than in young Tg or non-Tgmice. H2O2 production by untreated or treated (PMA or C.albicans) macrophages was significantly reduced (P � 0.04) inboth young and old Tg mice in comparison to non-Tg animals,demonstrating that macrophages from the Tg mice have asomewhat defective H2O2 response to these stimulants. Incontrast, levels of NO2
� production by macrophages fromyoung or old Tg and non-Tg mice were not significantly dif-ferent, either without treatment or after stimulation withIFN- and/or C. albicans (P � 0.05) (Fig. 7C and D).
Enhanced production of IL-6 and MCP-1 by macrophagesfrom Tg mice. Peritoneal macrophages from uninfected Tgmice produced significantly greater (P 0.001) concentrationsof IL-6 and MCP-1 than those from non-Tg controls whenincubated either alone or with heat-inactivated C. albicans(Fig. 8), demonstrating that macrophages from Tg mice displayan activated phenotype. In addition, production of TNF-� bymacrophages from both Tg and non-Tg mice was significantlyenhanced (P � 0.003) by incubation with C. albicans. However,transgene expression, stimulation with C. albicans, and age ofmice did not significantly alter production of IL-10, IFN-, orIL-12p70. Finally, production of IL-6, MCP-1, and IFN- wassignificantly decreased (P 0.001) in old compared to youngmice, independently of transgene expression or stimulationwith C. albicans, consistent with the reported lower productionof proinflammatory cytokines and chemokines in aged micethan in young mice (51).
Tg mice lacking iNOS are not more susceptible to oralcandidiasis or systemic dissemination. To determine if macro-phage production of NO contributes to mucosal containment ofC. albicans, Tg mice and non-Tg littermates containing a homozy-gous or heterozygous disruption of the macrophage iNOS gene(NOS2) were orally infected with C. albicans and longitudinallyassessed for mortality and oral burdens of C. albicans (Fig. 9).Over a 157-day observation period, mortality occurred only inNOS2�/� and NOS2�/� Tg mice, not in their respective non-Tgcontrols, which all survived (P 0.005) (Fig. 9A). However,survival of NOS2�/� and survival of NOS2�/� Tg mice werenot significantly different (P � 0.66), indicating that abrogationof NO production does not independently hasten mortality ofthese Tg mice. During both primary infection (days 1 to 15after infection) and chronic carriage of C. albicans (days 22 to92), oral burdens were significantly elevated in both CD4C/HIVMutG NOS2�/� and NOS2 �/� Tg mice over those in their
FIG. 5. Longitudinal assessment of oral and small intestine macro-phage cell populations in CD4C/HIVMutA Tg mice infected or notinfected with Candida albicans. Data are presented as the percentageof CD45� cells and are the means � standard deviations of resultsfrom four or five independent experiments. Significant differences: *,P 0.05 compared to infected mice on day 7; **, P 0.05 comparedto uninfected control mice on day 70.
4142 GOUPIL ET AL. INFECT. IMMUN.
on March 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

non-Tg controls (P 0.001), which in contrast rapidly clearedC. albicans from oral cavities immediately after primary infec-tion (Fig. 9B). The level of sustained enhancement of oralburdens during chronic carriage in these CD4C/HIVMutG
NOS2�/� and NOS2 �/� mice compared to non-Tg mice wassimilar to what we previously observed for CD4C/HIVMutG
NOS2�/� animals (42). However, Tg or non-Tg homozygousNOS2�/� mice did not have elevated burdens of C. albicanscompared to their respective heterozygous NOS2�/� controls
(P � 0.05), demonstrating that a lack of NOS2 does not en-hance oral proliferation of C. albicans in either Tg or non-Tgmice. Furthermore, a lack of NO production did not en-hance the burdens of C. albicans in the stomachs or smalland large intestines of CD4C/HIVMutG Tg mice or causesystemic dissemination of C. albicans (Table 1). The levelsof nitrite in culture supernatants of peritoneal macrophagesfrom non-Tg mice stimulated with IFN- for 48 h were 22�M from NOS2�/� mice, 12 �M from NOS2�/� macro-
FIG. 6. Phagocytosis and killing of C. albicans blastoconidia by peritoneal macrophages from CD4C/HIVMutA Tg and non-Tg mice. Analysiswas performed at the indicated macrophage/blastoconidium ratios (1:1, 1:5, and 1:10). Data represent the means � standard deviations of resultsfrom 5 to 11 independent observations. *, P 0.05 compared to non-Tg mice.
VOL. 77, 2009 RESPONSE TO C. ALBICANS IN HIV GENE-EXPRESSING Tg MICE 4143
on March 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
phages, and 5 �M for macrophages from NOS2�/� mice(the RPMI medium control was 2 �M), demonstratingthat the NOS2�/� mice have approximately 50% of thenormal level of iNOS activity.
DISCUSSION
In this study, we demonstrate that (i) Tg mice expressingHIV-1 successfully recruit macrophages to the oral and gastricmucosae in response to chronic carriage of C. albicans, despiteexpression of the HIV-1 transgene; (ii) macrophages from theTg mice display an activated phenotype; (iii) macrophagesfrom these Tg mice have a modestly reduced production ofH2O2 in response to C. albicans in vitro but unimpaired pro-duction of NO and killing of C. albicans; and (iv) production ofNO by macrophages is not required to limit mucosal prolifer-ation of C. albicans or to prevent systemic dissemination inthese Tg mice. Taken together, these results indicate that themacrophage response to C. albicans is largely preserved andthat no quantitative or functional macrophage defect directlycauses the susceptibility to mucosal candidiasis in these Tgmice.
The Tg mice were able to recruit macrophages to the mu-cosa in response to C. albicans, as previously reported for bothintact (9, 10) and immunodeficient SCID (71) and RAG-2�/�
(12) mice. This intact recruitment of macrophages is consistentwith unaltered numbers of circulating monocytes (42) and en-hanced expression of the CC chemokine RANTES (CCL5) inthe lamina propria of the gastrointestinal tract of these Tgmice (29). An increased expression of RANTES has beenfound in the oral mucosae of HIV-infected patients, with afurther enhancement in those with OPC (38).
The observed decrease in the percentage of macrophages inthe oral mucosae of mice infected with C. albicans, measuredrelative to the number of CD45� cells by flow cytometry, mayhave resulted from the more abundant recruitment, relative tomacrophages, of other CD45� cell populations, including den-dritic cells and CD4� and CD8� cells found within the muco-sal inflammatory foci. This interpretation is supported by theunaltered percentage of macrophages in the small intestines ofthese infected Tg mice, which in contrast to the oral cavity andstomach resist Candida infection, as evidenced by an absenceof Candida hyphae and inflammatory cell infiltrates (14).
The predominant expression of the mannose receptor but
FIG. 7. Production of H2O2 (A and B) and nitrite (NO2�) (C and D) by peritoneal macrophages from young (A and C) or old (B and D)
CD4C/HIVMutA Tg and non-Tg mice. Adherent macrophages were stimulated with IFN-, PMA, or C. albicans blastoconidia. In selectedexperiments for nitrite production (C and D), C. albicans was added to elicited peritoneal cells at the start of the 2-h period of adherence (C.albicans/2h), followed by stimulation of the adherent cells with IFN- or no stimulation. Data represent the means � standard deviations of resultsfrom four or five (H2O2) and five (NO2
�) independent observations. *, P � 0.04 compared to non-Tg mice.
4144 GOUPIL ET AL. INFECT. IMMUN.
on March 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

not MCP-1 by macrophages recruited to the oral mucosae andgastric submucosae of Tg mice premortem and at 70 days afterinfection with C. albicans is consistent with polarization towardan alternatively activated (M2) phenotype mediated by Th2-type cytokines IL-4 and IL-13 (26, 27, 47). We have previouslyshown that CD4� T cells harvested from Tg mice 70 days afterinfection with C. albicans display a polarization toward a Th2response, demonstrated by a sharp decline in the percentage ofcervical lymph node CD4� T cells producing the Th1 cytokineIFN- but a marked rise in the percentage of cells producingIL-4 (37). A direct effect of transgene expression on polariza-tion toward an M2 phenotype was shown by the preferentiallyenhanced expression of YM1 and ARG1 genes by peritonealmacrophages from uninfected old Tg mice. However, Candidainfection also contributed to this phenotype, since these geneswere also preferentially expressed by peritoneal macrophagesfrom non-Tg mice infected intraperitoneally with C. albicans.Although the M2 phenotype was predominant, the M1 TNF-�and NOS2 marker genes were nevertheless expressed at lowerbut similar levels in Tg and non-Tg mice infected with C.albicans. Accordingly, macrophages from these infected micedisplay an intermediate activation phenotype (47) that is pre-dominantly M2. Although evidence indicates that alternativelyactivated macrophages are more susceptible to some intracel-lular pathogens, including Cryptococcus neoformans (49), IL-13was found to attenuate gastrointestinal candidiasis in normaland immunodeficient RAG-2�/� mice, and this protection wascorrelated with recruitment of macrophages overexpressingthe mannose receptor to the intestinal mucosa (12).
Functionally, peritoneal macrophages from uninfected Tgmice displayed features of an activated phenotype, as shown byincreased phagocytosis of C. albicans and enhanced productionof IL-6 and MCP-1 after stimulation with C. albicans in vitro orwithout stimulation. These results demonstrate that the HIV-1transgene independently activates selected macrophage func-tions in the absence of Candida infection. HIV-1 and its Nefprotein have been shown to cause the release of several mac-rophage proinflammatory cytokines, including IL-6 by activa-tion of the NF-�B transcription factor (3, 59), and HIV-1induces a primed, proinflammatory state, which increases mac-rophage production of IL-6 in response to the primarily TLR4agonist LPS (5).
Peritoneal macrophages from these uninfected Tg mice pro-duced an unimpaired TNF-� response to C. albicans in vitro (2,36), consistent with the reported intact TLR4 and dectin-1/TLR2 receptor complex-dependent signaling pathways inHIV-1-infected macrophages (4, 5, 50, 53). In addition, TNF-�gene expression by peritoneal macrophages from Tg mice andthat by peritoneal macrophages from non-Tg mice were aug-mented to the same degree in vivo in response to C. albicansinfection. Because an intact TNF-� response is required forsuccessful host defense against oral and systemic candidiasis(21, 41), no defect in the capacity of macrophages to produceTNF-� appears to contribute to the susceptibility of these Tgmice to Candida infection.
Production of H2O2 by macrophages primed with IFN- andtreated with PMA or C. albicans was moderately reduced inthe Tg mice, while production of NO in response to IFN-
FIG. 8. Production of cytokines by peritoneal macrophages from young or old CD4C/HIVMutA Tg and non-Tg mice, incubated with heat-inactivated C. albicans blastoconidia or not incubated. Data represent the means � standard deviations of results from five to seven independentobservations. *, P 0.001 compared to non-Tg mice; **, P � 0.003 compared to macrophages incubated without C. albicans.
VOL. 77, 2009 RESPONSE TO C. ALBICANS IN HIV GENE-EXPRESSING Tg MICE 4145
on March 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

and/or C. albicans was unaffected by expression of the HIV-1transgene. This defective H2O2 production was insufficient todiminish killing of C. albicans by macrophages from the Tgmice, likely due to a redundancy provided by intact NO andoxygen-independent candidacidal mechanisms. The preservedinduction of macrophage NO production by C. albicans andIFN- is consistent with intact signaling via TLR4 (5, 66),STAT1, and NF-�B (67, 72) and concurs with a previous studywhich showed that thioglycolate-elicited peritoneal macro-phages from Nef and Vpr-expressing Tg mice, stimulated withLPS and IFN-, had deficient NO production while productionwas unaltered in Nef-expressing Tg mice (16). The CD4C/HIVMutA Tg mice used in the present study express Nef, Env,and Rev but not Vpr (30). Deficient production of H2O2 bymacrophages from the Tg mice in response to PMA suggests adefect at the level of protein kinase C (39) or faulty signalingthrough the JNK, ERK1/2, p38, and JAK-STAT pathways (32).In rat astrocytes, PMA activates these pathways, whereasIFN- activates the JAK-STAT and ERK1/2 pathways only(32). Therefore, the HIV transgene may interfere with selectedsignaling pathways involved in the response to PMA but notthose involved in the response to IFN-, potentially explainingwhy the macrophage H2O2 response to PMA is reduced whilethe NO response to IFN- is intact in these Tg mice. Thishypothesis is supported by the results of a study examining thegenomewide innate immune response to LPS of monocyte-derived macrophages infected in vitro with replicating CCR5-tropic HIV-1, which demonstrated a selective inhibitory effecton the classical NF-�B activation pathway while other signal
transduction pathways, such as the mitogen-activated proteinkinase pathway involving p38 phosphorylation, were not af-fected (58). In accordance with our present findings, theseinnate immune responses in HIV-infected macrophages werebroadly preserved, with limited exceptions (58). Production ofROIs by PMA-stimulated human monocyte-derived macro-phages infected with HIV-1 in vitro has been found to be eithernormal (57) or reduced (31), while production by monocyte-derived macrophages from HIV-1-infected patients was defec-tive in comparison to that of uninfected controls (48).
Macrophages from Tg mice had an unimpaired ability toupregulate NOS2 expression in vivo and NO production invitro in response to C. albicans. However, disruption of theNOS2 gene in these Tg mice did not further augment oralburdens during chronic carriage of C. albicans or cause sys-temic dissemination to deep organs. These results concur witha lack of enhancement of oral C. albicans burdens duringprimary infection of immunocompetent iNOS�/� mice (22)and indicate that macrophage NO production is also dispens-able during chronic oral carriage in these Tg mice with defec-tive adaptive immunity to C. albicans (37), likely due at least inpart to the redundancy provided by partially preserved produc-tion of H2O2 and oxygen-independent candidacidal mecha-nisms.
We have previously shown that PMNs from these Tg miceare quantitatively and functionally intact and that depletion ofPMNs does not alter oral or gastrointestinal burdens of C.albicans or cause systemic dissemination of endogenous (gas-trointestinal) origin (42). However, interference with both
FIG. 9. (A) Percent survival and (B) oral burdens of Candida albicans in CD4C/HIVMutG Tg mice and non-Tg littermates containing ahomozygous (NOS2�/�) or heterozygous (NOS2�/�) mutation of the macrophage iNOS gene. Mice were inoculated intraorally with 108 CFU ofC. albicans, and burdens were assessed longitudinally by sampling the oral cavity. Data represent the means � standard deviations (SD) of resultsfor 9 mice in the “non-Tg iNOS�/�” group and for 10 mice in each of the three other groups.
4146 GOUPIL ET AL. INFECT. IMMUN.
on March 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

macrophage and PMN functions was necessary to enhance thesusceptibility of T- and B-cell-deficient SCID mice to orogas-tric candidiasis (33). Therefore, macrophages and PMNs couldboth contribute to mucosal containment of C. albicans in theseTg mice but without observable enhancement in susceptibilitybecause of their functional redundancy.
In conclusion, we found that the mucosal macrophage re-sponse to C. albicans is largely preserved in these Tg mice. Thisresult was unexpected because dendritic cells and CD4� T cellsare both profoundly defective in the Tg mice and, conse-quently, one would have predicted a defect of macrophages askey effector cells of a classical M1-mediated Th1 response. Thefact that this was not the case appears to result at least in partfrom macrophage activation by the HIV-1 transgene. Furtherstudies will be required to identify the defective anti-Candidamucosal effector mechanisms which cause susceptibility to can-didiasis in these Tg mice and to determine if and how alteredCD4� T cells cause these effector mechanism defects.
ACKNOWLEDGMENTS
This work was supported by the Canadian Institutes of Health Re-search HIV/AIDS Research Program (grant H0P-41544). VeroniqueDugas is the recipient of a studentship award from the University ofMontreal.
We are grateful to Foo Y. Liew for providing NOS2-deficient mice.We thank Marie-Andree Laniel and Stephanie Lemay for support inmaintaining the Tg and NOS2-deficient mouse colony, Louis Gabouryfor interpretation of the histopathology, Christian Charbonneau forassistance with photomicrography, Miguel Chagnon and Yves Lepagefor statistical analysis, and Claire St-Onge for manuscript preparation.
REFERENCES
1. Balish, E., H. Filutowicz, and T. D. Oberley. 1990. Correlates of cell-medi-ated immunity in Candida albicans-colonized gnotobiotic mice. Infect. Im-mun. 58:107–113.
2. Barker, K. S., T. Liu, and P. D. Rogers. 2005. Coculture of THP-1 humanmononuclear cells with Candida albicans results in pronounced changes inhost gene expression. J. Infect. Dis. 192:901–912.
3. Bergamini, A., F. Bolacchi, E. Faggioli, R. Placido, S. Vendetti, L. Cappan-noli, L. Ventura, G. Cerasari, I. Uccella, M. Andreoni, and G. Rocchi. 1998.HIV-1 does not alter in vitro and in vivo IL-10 production by human mono-cytes and macrophages. Clin. Exp. Immunol. 112:105–111.
4. Brown, G. D., J. Herre, D. L. Williams, J. A. Willment, A. S. Marshall, andS. Gordon. 2003. Dectin-1 mediates the biological effects of beta-glucans. J.Exp. Med. 197:1119–1124.
5. Brown, J. N., J. J. Kohler, C. R. Coberley, J. W. Sleasman, and M. M.Goodenow. 2008. HIV-1 activates macrophages independent of Toll-likereceptors. PLoS ONE 3:e3664.
6. Campo, J., R. J. Del, J. Castilla, S. Garcia, C. Rodriguez, and A. Bascones.2002. Oral candidiasis as a clinical marker related to viral load, CD4 lym-phocyte count and CD4 lymphocyte percentage in HIV-infected patients.J. Oral Pathol. Med. 31:5–10.
7. Cantorna, M. T., and E. Balish. 1991. Role of CD4� lymphocytes in resis-tance to mucosal candidiasis. Infect. Immun. 59:2447–2455.
8. Cenci, E., L. Romani, A. Mencacci, R. Spaccapelo, E. Schiaffella, P. Puccetti,and F. Bistoni. 1993. Interleukin-4 and interleukin-10 inhibit nitric oxide-dependent macrophage killing of Candida albicans. Eur. J. Immunol. 23:1034–1038.
9. Chakir, J., L. Cote, C. Coulombe, and N. Deslauriers. 1994. Differentialpattern of infection and immune response during experimental oral candi-diasis in BALB/c and DBA/2 (H-2d) mice. Oral Microbiol. Immunol. 9:88–94.
10. Cole, G. T., A. A. Halawa, and E. J. Anaissie. 1996. The role of the gastro-intestinal tract in hematogenous candidiasis: from the laboratory to thebedside. Clin. Infect. Dis. 22(Suppl. 2):S73–S88.
11. Conti, H. R., F. Shen, N. Nayyar, E. Stocum, J. N. Sun, M. J. Lindemann,A. W. Ho, J. H. Hai, J. J. Yu, J. W. Jung, S. G. Filler, P. Masso-Welch, M.Edgerton, and S. L. Gaffen. 2009. Th17 cells and IL-17 receptor signaling areessential for mucosal host defense against oral candidiasis. J. Exp. Med.206:299–311.
12. Coste, A., C. Lagane, C. Filipe, H. Authier, A. Gales, J. Bernad, V. Douin-Echinard, J. C. Lepert, P. Balard, M. D. Linas, J. F. Arnal, J. Auwerx, and
TA
BL
E1.
Viable
CF
Uin
organsof
CD
4C/H
IVM
utG
iNO
ST
gm
iceinoculated
intraorallyw
ithC
.albicans a
Mouse
type
Kidneys
Stomach
Smallintestine
Large
intestine
Mean
C.albicans
count,CF
U/g
(range)
No.of
mice
culturepositive
forC
.albicans/totalno.of
mice
Mean
C.albicans
count,C
FU
/g(range)
No.of
mice
culturepositive
forC
.albicans/totalno.of
mice
Mean
C.albicans
count,C
FU
/g(range)
No.of
mice
culturepositive
forC
.albicans/totalno.of
mice
Mean
C.albicans
count,C
FU
/g(range)
No.of
mice
culturepositive
forC
.albicans/totalno.of
mice
TgiN
OS
�/�
b8.0
�10
1(N
Af)
1/108.1
�10
4(2.0
�10
2–4.9�
105)
8/101.9
�10
3(1.4
�10
2–9.9�
103)
6/105.1
�10
3(3.9
�10
2–1.2�
104)
8/10iN
OS
�/�
cN
A(N
A)
0/105.6
�10
4(1.8
�10
2–3.3�
105)
8/102.1
�10
4(5.3
�10
1–1.4�
105)
7/104.2
�10
3(1.1
�10
2–1.8�
104)
9/10
Controlnon-T
giN
OS
�/�
dN
A(N
A)
0/103.3
�10
3(N
A)
1/102.8
�10
2(N
A)
1/107.7
�10
2(N
A)
1/10iN
OS
�/�
eN
A(N
A)
0/9N
A(N
A)
0/9N
A(N
A)
0/9N
A(N
A)
0/9
aM
icestudied
includedT
gand
controlnon-Tg
offspringderived
fromfounder
mouse
F27367.N
oculture-positive
resultsw
ereobtained
with
brains,lungs,andlivers
fromany
mouse
type.b
The
mean
ageat
assessment
was
155days
(range,106to
205days).A
ssessment
was
doneon
theday
ofeuthanization,because
ofsevere
illness;survivorsw
ereeuthanized
157days
afterinoculation
with
C.albicans.
cThe
mean
ageat
assessment
was
145days
(range,112to
205days).A
ssessment
was
doneon
theday
ofeuthanization,because
ofsevere
illness;survivorsw
ereeuthanized
157days
afterinoculation
with
C.albicans.
dT
hem
eanage
atassessm
entw
as209
days(range,203
to222
days).All10
mice
survivedand
were
euthanized155
daysafter
inoculationw
ithC
.albicans.eT
hem
eanage
atassessm
entw
as210
days(range,204
to222
days).All9
mice
survivedand
were
euthanized156
daysafter
inoculationw
ithC
.albicans.fN
A,not
applicable.
VOL. 77, 2009 RESPONSE TO C. ALBICANS IN HIV GENE-EXPRESSING Tg MICE 4147
on March 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

B. Pipy. 2008. IL-13 attenuates gastrointestinal candidiasis in normal andimmunodeficient RAG-2(-/-) mice via peroxisome proliferator-activated re-ceptor-gamma activation. J. Immunol. 180:4939–4947.
13. Crowe, S. M., N. J. Vardaxis, S. J. Kent, A. L. Maerz, M. J. Hewish, M. S.McGrath, and J. Mills. 1994. HIV infection of monocyte-derived macro-phages in vitro reduces phagocytosis of Candida albicans. J. Leukoc. Biol.56:318–327.
14. de Repentigny, L., F. Aumont, J. S. Ripeau, M. Fiorillo, D. G. Kay, Z. Hanna,and P. Jolicoeur. 2002. Mucosal candidiasis in transgenic mice expressinghuman immunodeficiency virus type 1. J. Infect. Dis. 185:1103–1114.
15. de Repentigny, L., D. Lewandowski, and P. Jolicoeur. 2004. immunopatho-genesis of oropharyngeal candidiasis in human immunodeficiency virus in-fection. Clin. Microbiol. Rev. 17:729–759.
16. Dickie, P., A. Roberts, and R. Lee. 2001. A defect in HIV-1 transgenicmurine macrophages results in deficient nitric oxide production. J. Leukoc.Biol. 70:592–600.
17. Elahi, S., G. Pang, R. B. Ashman, and R. Clancy. 2001. Nitric oxide-en-hanced resistance to oral candidiasis. Immunology 104:447–454.
18. Elahi, S., G. Pang, R. Clancy, and R. B. Ashman. 2000. Cellular and cytokinecorrelates of mucosal protection in murine model of oral candidiasis. Infect.Immun. 68:5771–5777.
19. Farah, C. S., S. Elahi, K. Drysdale, G. Pang, T. Gotjamanos, G. J. Seymour,R. L. Clancy, and R. B. Ashman. 2002. Primary role for CD4� T lymphocytesin recovery from oropharyngeal candidiasis. Infect. Immun. 70:724–731.
20. Farah, C. S., S. Elahi, G. Pang, T. Gotjamanos, G. J. Seymour, R. L. Clancy,and R. B. Ashman. 2001. T cells augment monocyte and neutrophil functionin host resistance against oropharyngeal candidiasis. Infect. Immun. 69:6110–6118.
21. Farah, C. S., Y. Hu, S. Riminton, and R. B. Ashman. 2006. Distinct roles forinterleukin-12p40 and tumour necrosis factor in resistance to oral candidiasisdefined by gene-targeting. Oral Microbiol. Immunol. 21:252–255.
22. Farah, C. S., J. M. Saunus, Y. Hu, A. Kazoullis, and R. B. Ashman. 2009.Gene targeting demonstrates that inducible nitric oxide synthase is notessential for resistance to oral candidiasis in mice, or for killing of Candidaalbicans by macrophages in vitro. Oral Microbiol. Immunol. 24:83–88.
23. Fattorossi, A., R. Nisini, J. G. Pizzolo, and R. D’Amelio. 1989. New, simpleflow cytometry technique to discriminate between internalized and mem-brane-bound particles in phagocytosis. Cytometry 10:320–325.
24. Federico, M., Z. Percario, E. Olivetta, G. Fiorucci, C. Muratori, A. Micheli,G. Romeo, and E. Affabris. 2001. HIV-1 Nef activates STAT1 in humanmonocytes/macrophages through the release of soluble factors. Blood 98:2752–2761.
25. Fidel, P. L., Jr. 2006. Candida-host interactions in HIV disease: relationshipsin oropharyngeal candidiasis. Adv. Dent. Res. 19:80–84.
26. Gordon, S. 2007. The macrophage: past, present and future. Eur. J. Immu-nol. 37(Suppl. 1):S9–S17.
27. Gordon, S. 2003. Alternative activation of macrophages. Nat. Rev. Immunol.3:23–35.
28. Green, L. C., D. A. Wagner, J. Glogowski, P. L. Skipper, J. S. Wishnok, andS. R. Tannenbaum. 1982. Analysis of nitrate, nitrite, and [15N]nitrate inbiological fluids. Anal. Biochem. 126:131–138.
29. Hanna, Z., D. G. Kay, M. Cool, S. Jothy, N. Rebai, and P. Jolicoeur. 1998.Transgenic mice expressing human immunodeficiency virus type 1 in im-mune cells develop a severe AIDS-like disease. J. Virol. 72:121–132.
30. Hanna, Z., D. G. Kay, N. Rebai, A. Guimond, S. Jothy, and P. Jolicoeur.1998. Nef harbors a major determinant of pathogenicity for an AIDS-likedisease induced by HIV-1 in transgenic mice. Cell 95:163–175.
31. Howell, A. L., D. S. Groveman, P. K. Wallace, and M. W. Fanger. 1997.HIV-1-infected monocytes and monocyte-derived macrophages are impairedin their ability to produce superoxide radicals. Int. J. Clin. Lab Res. 27:111–117.
32. Hwang, M. N., K. S. Kim, Y. W. Choi, I. Jou, and S. Yoon. 2007. PMAactivates Stat3 in the Jak/Stat pathway and induces SOCS5 in rat brainastrocytes. Mol. Cells 23:94–99.
33. Jensen, J., T. Warner, and E. Balish. 1994. The role of phagocytic cells inresistance to disseminated candidiasis in granulocytopenic mice. J. Infect.Dis. 170:900–905.
34. Jones-Carson, J., A. Vazquez-Torres, H. C. van der Heyde, T. Warner, R. D.Wagner, and E. Balish. 1995. Gamma delta T cell-induced nitric oxideproduction enhances resistance to mucosal candidiasis. Nat. Med. 1:552–557.
35. Kedzierska, K., R. Azzam, P. Ellery, J. Mak, A. Jaworowski, and S. M.Crowe. 2003. Defective phagocytosis by human monocyte/macrophages fol-lowing HIV-1 infection: underlying mechanisms and modulation by adjunc-tive cytokine therapy. J. Clin. Virol. 26:247–263.
36. Kim, H. S., E. H. Choi, J. Khan, E. Roilides, A. Francesconi, M. Kasai, T.Sein, R. L. Schaufele, K. Sakurai, C. G. Son, B. T. Greer, S. Chanock, C. A.Lyman, and T. J. Walsh. 2005. Expression of genes encoding innate hostdefense molecules in normal human monocytes in response to Candidaalbicans. Infect. Immun. 73:3714–3724.
37. Lewandowski, D., M. Marquis, F. Aumont, A. C. Lussier-Morin, M. Ray-mond, S. Senechal, Z. Hanna, P. Jolicoeur, and L. de Repentigny. 2006.Altered CD4� T cell phenotype and function determine the susceptibility to
mucosal candidiasis in transgenic mice expressing HIV-1. J. Immunol. 177:479–491.
38. Lilly, E. A., D. J. Hart, J. E. Leigh, S. Hager, K. M. McNulty, D. E. Mercante,and P. L. Fidel, Jr. 2004. Tissue-associated cytokine expression in HIV-positive persons with oropharyngeal candidiasis. J. Infect. Dis. 190:605–612.
39. Liu, W. S., and C. A. Heckman. 1998. The sevenfold way of PKC regulation.Cell. Signal. 10:529–542.
40. Mangino, G., Z. A. Percario, G. Fiorucci, G. Vaccari, S. Manrique, G.Romeo, M. Federico, M. Geyer, and E. Affabris. 2007. In vitro treatment ofhuman monocytes/macrophages with myristoylated recombinant Nef of hu-man immunodeficiency virus type 1 leads to the activation of mitogen-activated protein kinases, I�B kinases, and interferon regulatory factor 3 andto the release of beta interferon. J. Virol. 81:2777–2791.
41. Marino, M. W., A. Dunn, D. Grail, M. Inglese, Y. Noguchi, E. Richards, A.Jungbluth, H. Wada, M. Moore, B. Williamson, S. Basu, and L. J. Old. 1997.Characterization of tumor necrosis factor-deficient mice. Proc. Natl. Acad.Sci. USA 94:8093–8098.
42. Marquis, M., D. Lewandowski, V. Dugas, F. Aumont, S. Senechal, P. Joli-coeur, Z. Hanna, and L. de Repentigny. 2006. CD8� T cells but not poly-morphonuclear leukocytes are required to limit chronic oral carriage ofCandida albicans in transgenic mice expressing human immunodeficiencyvirus type 1. Infect. Immun. 74:2382–2391.
43. Marr, K. A., S. A. Balajee, T. R. Hawn, A. Ozinsky, U. Pham, S. Akira, A.Aderem, and W. C. Liles. 2003. Differential role of MyD88 in macrophage-mediated responses to opportunistic fungal pathogens. Infect. Immun. 71:5280–5286.
44. Martinez, F. O., A. Sica, A. Mantovani, and M. Locati. 2008. Macrophageactivation and polarization. Front. Biosci. 13:453–461.
45. Martins, M. D., M. Lozano-Chiu, and J. H. Rex. 1998. Declining rates oforopharyngeal candidiasis and carriage of Candida albicans associated withtrends toward reduced rates of carriage of fluconazole-resistant C. albicansin human immunodeficiency virus-infected patients. Clin. Infect. Dis. 27:1291–1294.
46. Mercante, D. E., J. E. Leigh, E. A. Lilly, K. McNulty, and P. L. Fidel, Jr.2006. Assessment of the association between HIV viral load and CD4 cellcount on the occurrence of oropharyngeal candidiasis in HIV-infected pa-tients. J. Acquir. Immune Defic. Syndr. 42:578–583.
47. Mosser, D. M., and J. P. Edwards. 2008. Exploring the full spectrum ofmacrophage activation. Nat. Rev. Immunol. 8:958–969.
48. Muller, F., H. Rollag, and S. S. Froland. 1990. Reduced oxidative burstresponses in monocytes and monocyte-derived macrophages from HIV-in-fected subjects. Clin. Exp. Immunol. 82:10–15.
49. Muller, U., W. Stenzel, G. Kohler, C. Werner, T. Polte, G. Hansen, N.Schutze, R. K. Straubinger, M. Blessing, A. N. McKenzie, F. Brombacher,and G. Alber. 2007. IL-13 induces disease-promoting type 2 cytokines, alter-natively activated macrophages and allergic inflammation during pulmonaryinfection of mice with Cryptococcus neoformans. J. Immunol. 179:5367–5377.
50. Murciano, C., A. Yanez, M. L. Gil, and D. Gozalbo. 2007. Both viable andkilled Candida albicans cells induce in vitro production of TNF-alpha andIFN-gamma in murine cells through a TLR2-dependent signalling. Eur.Cytokine Netw. 18:38–43.
51. Murciano, C., A. Yanez, J. E. O’Connor, D. Gozalbo, and M. L. Gil. 2008.Influence of aging on murine neutrophil and macrophage function againstCandida albicans. FEMS Immunol. Med. Microbiol. 53:214–221.
52. Netea, M. G., K. Gijzen, N. Coolen, I. Verschueren, C. Figdor, J. W. Van derMeer, R. Torensma, and B. J. Kullberg. 2004. Human dendritic cells are lesspotent at killing Candida albicans than both monocytes and macrophages.Microbes Infect. 6:985–989.
53. Netea, M. G., N. A. Gow, C. A. Munro, S. Bates, C. Collins, G. Ferwerda,R. P. Hobson, G. Bertram, H. B. Hughes, T. Jansen, L. Jacobs, E. T.Buurman, K. Gijzen, D. L. Williams, R. Torensma, A. McKinnon, D. M.MacCallum, F. C. Odds, J. W. Van der Meer, A. J. Brown, and B. J.Kullberg. 2006. Immune sensing of Candida albicans requires cooperativerecognition of mannans and glucans by lectin and Toll-like receptors. J. Clin.Investig. 116:1642–1650.
54. Netea, M. G., J. W. Meer, I. Verschueren, and B. J. Kullberg. 2002. CD40/CD40 ligand interactions in the host defense against disseminated Candidaalbicans infection: the role of macrophage-derived nitric oxide. Eur. J. Im-munol. 32:1455–1463.
55. Nielsen, H., K. D. Bentsen, L. Hojtved, E. H. Willemoes, F. Scheutz, M.Schiodt, K. Stoltze, and J. J. Pindborg. 1994. Oral candidiasis and immunestatus of HIV-infected patients. J. Oral Pathol. Med. 23:140–143.
56. Nielsen, H., A. Kharazmi, and V. Faber. 1986. Blood monocyte and neutro-phil functions in the acquired immune deficiency syndrome. Scand. J. Im-munol. 24:291–296.
57. Nottet, H. S., L. de Graaf, N. M. de Vos, L. J. Bakker, J. A. van Strijp, M. R.Visser, and J. Verhoef. 1993. Phagocytic function of monocyte-derived mac-rophages is not affected by human immunodeficiency virus type 1 infection.J. Infect. Dis. 168:84–91.
58. Noursadeghi, M., J. Tsang, R. F. Miller, S. Straschewski, P. Kellam, B. M.Chain, and D. R. Katz. 2009. Genome-wide innate immune responses in
4148 GOUPIL ET AL. INFECT. IMMUN.
on March 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

HIV-1-infected macrophages are preserved despite attenuation of the NF-kappa B activation pathway. J. Immunol. 182:319–328.
59. Olivetta, E., Z. Percario, G. Fiorucci, G. Mattia, I. Schiavoni, C. Dennis, J.Jager, M. Harris, G. Romeo, E. Affabris, and M. Federico. 2003. HIV-1 Nefinduces the release of inflammatory factors from human monocyte/macro-phages: involvement of Nef endocytotic signals and NF-kappa B activation.J. Immunol. 170:1716–1727.
60. Pfaffl, M. W., G. W. Horgan, and L. Dempfle. 2002. Relative expressionsoftware tool (REST) for group-wise comparison and statistical analysis ofrelative expression results in real-time PCR. Nucleic Acids Res. 30:e36.
61. Pirofski, L. A., and A. Casadevall. 2009. Rethinking T cell immunity inoropharyngeal candidiasis. J. Exp. Med. 206:269–273.
62. Pugliese, A., D. Torre, F. M. Baccino, G. Di Perri, C. Cantamessa, L.Gerbaudo, A. Saini, and V. Vidotto. 2000. Candida albicans and HIV-1infection. Cell Biochem. Funct. 18:235–241.
63. Romani, L. 1999. Immunity to Candida albicans: Th1, Th2 cells and beyond.Curr. Opin. Microbiol. 2:363–367.
64. Samaranayake, L. P., and P. Holmstrup. 1989. Oral candidiasis and humanimmunodeficiency virus infection. J. Oral Pathol. Med. 18:554–564.
65. Saresella, M., K. Roda, L. Speciale, D. Taramelli, E. Mendozzi, F. Guerini,and P. Ferrante. 1997. A rapid evaluation of phagocytosis and killing ofCandida albicans by CD13� leukocytes. J. Immunol. Methods. 210:227–234.
66. Schilling, D., K. Thomas, K. Nixdorff, S. N. Vogel, and M. J. Fenton. 2002.Toll-like receptor 4 and Toll-IL-1 receptor domain-containing adapter pro-tein (TIRAP)/myeloid differentiation protein 88 adapter-like (Mal) contrib-ute to maximal IL-6 expression in macrophages. J. Immunol. 169:5874–5880.
67. Schroder, K., M. J. Sweet, and D. A. Hume. 2006. Signal integration betweenIFNgamma and TLR signalling pathways in macrophages. Immunobiology.211:511–524.
68. Vazquez-Torres, A., and E. Balish. 1997. Macrophages in resistance to can-didiasis. Microbiol. Mol. Biol. Rev. 61:170–192.
69. Vazquez-Torres, A., J. Jones-Carson, and E. Balish. 1994. Candidacidalactivity of macrophages from immunocompetent and congenitally immuno-deficient mice. J. Infect. Dis. 170:180–188.
70. Vazquez-Torres, A., J. Jones-Carson, and E. Balish. 1996. Peroxynitritecontributes to the candidacidal activity of nitric oxide-producing macro-phages. Infect. Immun. 64:3127–3133.
71. Vazquez-Torres, A., J. Jones-Carson, T. Warner, and E. Balish. 1995. Nitricoxide enhances resistance of SCID mice to mucosal candidiasis. J. Infect.Dis. 172:192–198.
72. Vila-del Sol, V., M. Díaz-Munoz, and M. Fresno. 2007. Requirement oftumor necrosis factor � and nuclear factor-�B in the induction by IFN- ofinducible nitric oxide synthase in macrophages. J. Leukoc. Biol. 81:272–283.
73. Vonk, A. G., C. W. Wieland, M. G. Netea, and B. J. Kullberg. 2002. Phago-cytosis and intracellular killing of Candida albicans blastoconidia by neutro-phils and macrophages: a comparison of different microbiological test sys-tems. J. Microbiol. Methods. 49:55–62.
74. Wei, X. Q., I. G. Charles, A. Smith, J. Ure, G. J. Feng, F. P. Huang, D. Xu,W. Muller, S. Moncada, and F. Y. Liew. 1995. Altered immune responses inmice lacking inducible nitric oxide synthase. Nature 375:408–411.
Editor: A. Casadevall
VOL. 77, 2009 RESPONSE TO C. ALBICANS IN HIV GENE-EXPRESSING Tg MICE 4149
on March 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

















![Prevalence and predictors of Haikael Martin oral thrush ... · Oral candidiasis (also known as “oral thrush”) is a fungal infection caused mainly by Candida albicans (c.alb) [1]](https://img.pdfslide.net/doc/110x75/5c96d57e09d3f26b0a8d0743/prevalence-and-predictors-of-haikael-martin-oral-thrush-oral-candidiasis.jpg)





![Cronicon · Candida albicans (C. albicans) ... is a well-known therapeutic compound which is successfully used to treat vaginal candidiasis (VC) [14]. However, its efficacy to treat](https://img.pdfslide.net/doc/110x75/5f7c3ae023a7c12cf86688df/cronicon-candida-albicans-c-albicans-is-a-well-known-therapeutic-compound.jpg)

![Comparison of the Virulence of Different Candida albicans ......The features mentioned above are present in 50% of patients with HIV infection and 80% in patients with AIDS [8]. Candidiasis](https://img.pdfslide.net/doc/110x75/5f4befd088913e44a653cc8b/comparison-of-the-virulence-of-different-candida-albicans-the-features-mentioned.jpg)



