Embed Size (px)
Citation preview

Methods for measurements of chlorophyll fluorescence, luminiscence and photosynthesis in intact plants
by
Erik Sundbom
AKADEMISK AVHANDLING
som med tillstånd av rektorsämbetet vid Umeå universitet för erhållande av filosofie doktorsexamen, framlägges till offentlig granskning tisdagen den 9 juni 1981 kl. 10.00 vid Fysiologi- Botanik Hufo, seminarierum B.
Examinator: Professor Lennart Eliasson, UmeåFakultetsopponent: Amanuensis Thor Bernt Meltf, Trondheim

Titte:
Author:
Address:
Abstract:
Key words:
ISBN 91-717^
Methods for measurements of chlorophyll fluorescence, luminiscence and photosynthesis in intact plants.
Erik Sundbom
Department of Plant Physiology, University of Umeå, S-9OI 87 Umeå, Sweden
Methods were developed to study delayed light emission (luminiscence) and fluorescence changes in intact leaves of plants. Delayed light emission, detected from plants in darkness, was used to produce images of the plant leaves. The procedure was termed "phytoluminography". The use of the method is suggested for dia- nostic purposes at early stages of disturbances of the leaf tissues, not detectable with the naked eye. The delayed light emission is associated with the photochemistry of photosystem II and the light induced conversion and storage of energy in the thylakoid membrane system of chloroplasts.
Fluorescence yield changes were induced by lowering temperature between 20 C and -20 C. The temperature induced fluorescence changes in leaves parallel the temperature induced changes in isolated chloroplasts in reaction preparations mediating photosynthetic electron transport from endogenous water splitting to added NADP. At above freezing temperatures, lowering the temperature at a constant rate of 1 C per minute caused supressed electron transport and increased fluorescence yield which were linearely dependent on the temperature change in frost resistent plants.
Repeated freeze-thaw cycles between 20 °C and -20 °C induced variable fluorescence yield changes which were gradually depleated to F0 or Fm when the electron transport was injuried on the oxidizing or on the reduzing side of photosystem II, respectively.
The temperature induced fluorescence changes were used to characterize plants with different ability to withstand freezing temperatures. The method also discriminates between plants of different frost resistance, and the method was used in screening for frost tolerance.
Luminiscence, delayed light emission, phytoluminography, fluorescence, in vivo chlorophyll a fluorescence, photosynthesis, electron transport, frost tolerance, intact plants.
075-9
Umeå I98I. Distributed by the Department of Plant Physiology, University of Umeå, S-9OI 87 Umeå, Sweden.

Methods for measurements of chlorophyll fluorescence, luminiscence and photosynthesis in intact plants
byErik Sundbom
Department of Plant Physiology, University of Umeå, S-9OI 87 Umeå, Sweden
Doctoral thesis ISBN 91-717^-075-9


Titte:
Author:
Address:
Abstract:
Key words:
ISBN 91-717*»
Methods for measurements of chlorophyll fluorescence, luminiscence and photosynthesis in intact plants.
Erik Sundbom
Department of Plant Physiology, University of Umeå, S—901 87 Umeå, Sweden
Methods were developed to study delayed light emission (luminiscence) and fluorescence changes in intact leaves of plants. Delayed 1ight emission, detected from plants in darkness, was used to produce images of the plant leaves. The procedure was termed "phytoluminography". The use of the method is suggested for dia- nostic purposes at early stages of disturbances of the leaf tissues, not detectable with the naked eye. The delayed light emission is associated with the photochemistry of photosystem II and the light induced conversion and storage of energy in the thylakoid membrane system of chloroplasts.
Fluorescence yield changes were induced by lowering temperature between 20 C and -20 C. The temperature induced fluorescence changes in leaves parallel the temperature induced changes in isolated chloroplasts in reaction preparations mediating photosynthetic electron transport from endogenous water splitting to added NADP. At above freezing temperatures, lowering the temperature at a constant rate of 1 C per minute caused supressed electron transport and increased fluorescence yield which were linearely dependent on the temperature change in frost resistent plants.
Repeated freeze-thaw cycles between 20 °C and -20 °C induced variable fluorescence yield changes which were gradually depleated to F0 or Fm when the electron transport was injuried on the oxidizing or on the reduzing side of photosystem II, respectively.
The temperature induced fluorescence changes were used to characterize plants with different ability to withstand freezing temperatures. The method also discriminates between plants of different frost resistance, and the method was used in screening for frost tolerance.
Luminiscence, delayed light emission, phytoluminography, fluorescence, in vivo chlorophyll £ fluorescence, photosynthesis, electron transport, frost tolerance, intact plants.
075-9
Umeå I98I. Distributed by the Department of Plant Physiology, University of Umeå, S-901 87 Umeå, Sweden.


LIST OF PAPERS
This thesis is based on the following papers, which will be referred
to in the text by the given Roman numerals.
I Sundbom, E. and Björn, L.O. 1977. Phytoluminography: Imaging plants
by delayed light emission. - Physiol. Plant. ^0: 39_i*1.
II Sundbom, E. and ö’quist, G. 1981. Temperature induced variable
f1uorescence-yield changes of intact leaves.
III Sundbom, E. and bquist, G. 1981. Temperature induced changes of
fluorescence yield and photosynthetic electron transport of iso
lated chloroplasts.
IV Hällgren, J.-E., Sundbom, E. and Strand, M. 1.981. Photosynthetic
responses to low temperature in Betula pubescens and Betula
tortuosa. - Physiol. Plant., submitted for publication, 1981.
V Sundbom, E. 1981. Temperature induced fluorescence changes, a
screening method for frost tolerance of potato (Solanum tuberosum
L.).


CONTENTS
Page1. INTRODUCTION 1
2. LUMINISCENCE (Delayed light emission) 42.1. The slow decay component of delayed 4
1ight emission2.2. Phytoluminography: Imaging plants by 5
delayed light emission
3. FLUORESCENCE 63.1. Fluorescence and the photochemistry 6
of photosystem 113.2. Methods to induce fluorescence yield changes 7
3.2.1. Temperature induced fluorescence 7 changes
3.2.2. Freeze-thaw induced fluorescence 8 changes
4. APPLICATION OF TEMPERATURE INDUCEDFLUORESCENCE CHANGES 94.1. Photosynthesis and temperature induced 9
fluorescence changes in birches adaptedto different temperatures
4.2. Temperature induced fluorescence changes: 9 A method in screening for frost tolerance
5. ACKNOWLEDGEMENTS 12
6. REFERENCES 13


- 1 -
I INTRODUCTION
The absorption of light by the photosynthetically active pigments of
plants results in creation of singlet excitation energy. This energy
conversion is the first step in photosynthesis, a complex reaction
scheme of photophysical, photochemical and biochemical processes. The
different processes take place at varoius time scales. The first step,
light absorption, is finished within 10 ^ seconds while, carbohydrate
formation, the last step may take several seconds.The processes have been
divided into different eras by Kamen (1963) and are summarized by
Govindjee (1975) as follows:
A. Era of radiation physics, 10 ^ to 10 ^ seconds:
Chi + hv — > Chi” Excitation (1)9 s£Chi — > Chl-j. Intercrossing system (2)
Chi s Chi — > Chi + Chi" Excitation energy transfer (3)
Chi — > Chi + hv' Fluorescence (*»)s gJU
Chi s + T — > T + Ch 1 g Trapping of energy (5)
y'cChlg, chlorophyll in ground state; Chi chlorophyll in singlet
excitated state; Chl ., chlorophyll triplet; T, energy trap.
- 1 0 -3B. Era of photochemistry, 10 to 10 seconds:
D+TA" (6)
DT"A ^ --- — > DT+A~ -------- > D+TA" (7)
^ D+T _A > D+TA~ (8)
D, electron donator; A, electron acceptor.

- 2 -
-k -2C. Era of biochemistry, 10 to 10 seconds:
D+TA" + i NADP+ + H+ — > D+TA + i NADPH + i H+ ,reduction of NADP (9)
D+TA + i H20 — > DTA \ 0^ + H+ , oxygen evolution
D+TA + ADP + P. — > DTA + ATP, photophosphorylation
C02 + 2 NADPH + 3 ATP — > (CH20) + 2 NADP++ 3 ADP + 3 P.,
(1 0)(1 1 )
carbon fixation (12)
At a constant absorption of light quanta by the photosynthetic pigments
the excitation is influenced by the following deexcitation processes:
A. Light induced electron transport over photosystem II, involving
water splitting and secondary electron transfer redox pro
cesses.
B. Radi at ion less thermal deactivation
C. Radiant emission of Chi £ fluorescence
D. Distribution of excess excitation from Chi £ of photosystem II to
Chi £ of photosystem I, i.e. spillover.
The Chi £ excitation reemitted as fluorescence varies in response to the
rate of photosynthesis. It was shown by Lavorel (1959) and Clayton (19&9)
that the fluorescence emission consists of a constant and a variable
part. The variable part of Chi £ fluorescence is sensitive to the rate
of electron transport over photosystem II and also to changes in the
thylakoid membrane ultrastructure related to photophosphorylation.

- 3 -
Luminiscence (delayed light emission) is caused by a back reaction of
photosystem II created by the same singlet excitation that is respon
sible for the emission of fluorescence (promt fluorescence). The two
light phenomena thus have identical spectral distribution. They are
also subject to the same radiative or nonradiative decays, migration
and trapping by reaction center chlorophyll (Lavorel 1968). Thus lumi
niscence and variable fluorescence are both associated with the functio
ning of photosystem II (Goedheer 1962, 1963 and Bertch 1962).
The general purpose of this thesis was to study photosynthetic reactions
related to the photochemistry of intact plants, using methods non-
-destructive to the plant as a whole.
Delayed light emission, detected in darkness, was used to produce images
of the plants. The procedure was termed "phytoluminography". The use of
this method is suggested for diagnostic purposes at early stages of
disturbances in the leaf tissues, not detectable with the naked eye
(paper I).
An apparatus and a method to induce fluorescence yield changes in intact
leaves through temperature changes between 20 °C and -20 °C are described
(paper II). The temperature induced fluorescence changes at above free
zing temperatures are similar in intact leaves and in isolated chloro-
plasts and the fluorescence yield changes are, in this temperature
range, 1inearly dependent on the rate of photosynthetic electron tran
sport (paper III).
As a means to detect differences in frost tolerance, fluorescence yield
changes were also studied during repeated freeze-thaw cycles in intact
leaves (paper IV and V).

- k -
2. LUMINISCENCE (Delayed light emission)
2.1 The slow decay component of delayed light emission
Light production by previously illuminated green plants, which lasts
several minutes after the exciting light has been turned off, was dis
covered by Streler and Arnold (1951). The light phenomenon is known by
various names: delayed light, afterglow, luminiscence and delayed fluore
scence.
In general, the delayed light emission is caused by a reversal of the
photosynthetic conversion of light to chemical energy. Thus its intensity
and decay kinetics depend on the properties of the photosynthetic appara
tus. It is accepted that the delayed light emission originates from the
actively functioning photosystem II, and that the slow component, lasting
up to 10 minutes, correlates with the rate of the conversion
and S2 — > S.| (Jol iot e£ £l_. 1970 and Barbieri et aj_. 1970). The process
can be summarized as a back reaction between the reduced primary electron
acceptor Q and the oxidized primary electron donator that are produced
by photosystem II during excitation:
DP680A + hv'— > D+P680A"
D+p680A" — > DP680A + hv"
hv', exciting light; hv", delayed light emission.
Of several components with different decay rates of delayed light emission
(Björn 1971, Lavorel 1975), the slow component is suggested as the most
sensitive indicator of deviations from normal cell function. The slow
component is dependent on the properties of the thylakoids and on inter
actions between the chloroplasts and the rest of the cell (Björn 1971).

- 5 -
In paper I the slow component of delayed light emission with a half-
-life of the order of 1 minute was studied in intact leaves.
2.2 Phytoluminography: Imaging plants by delayed light emission
One difficulty in studying the slow delayed light emission component is
its extremely low intensity. As demonstrated in paper I the use of an
image intensifier (Mullard XX 1063 three-stage focusing type) with a
gain of about 50.000 photons out per photon incident on the photocatode
could detect the slow component of delayed light emission in darkness.
The image produced on the image intensifier screen could then be re
corded on high-speed black and white photographic film.
It is also shown that thermal damage of the leaf tissues and also treat
ment with the photosynthetic inhibitor DCMU, 3~(3>^_dichlorophenyl)-
-1,1-dimethylurea, supress the delayed light emission at the affected
spots of the leaf. It was suggested that phytoluminography,imaging
plants by delayed light emiss ion, might be used for diagnostic purposes
to detect early stages of disturbances due to parasites, mineral defi
ciency, herbicides, frosts etc. in whole leaves. Later this possibility
has been extensively studied, with an improved apparatus, by Björn et
al. (1979).

- 6 -
3. FLUORESCENCE
3.1 Fluorescence and photochemistry of photosystem II
The chlorophyll content of higher plant leaves is very high (about
10 M) and at physiological temperatures chlorophyll £ of photosystem
11 is weakly fluorescent with a main emissionband at 685 nm and a sa
tellite band at 7**0 nm. Probably the most studied fluorescence para
meter is the relative fluorescence yield of Chi £ since this fluore
scence yield is an index of the amount of singlet Chi £ excitation of
photosystem 11. The amount of Chi £ excitation which is reemitted as
fluorescence varies in response to the rate of photosynthesis. Roughly
expressed, weak Chi £ fluorescence is typical in a situation where the
photosynthetic apparatus is working undisturbed at optimal (high) rates
of the photosynthetic electron transport, whereas high fluorescence is
typical when the photosynthesis is weak or inhibited. However all of
the Chi £ emission is not sensitive to the rate of photosynthetic
electron transport. Lavorel (1959) and Clayton (1969) first made the
distinction between fluorescence of variable (F ) and fluorescence ofvconstant (F ) yield. The variable part of the fluorescence emission is
sensitive to the rate of electron transport over photosystem II. Thus
the functioning of the electron donating and electron accepting side
of photosystem II would be expected to have great influence on the
variable fluorescence yield. Maximal fluorescence F is obtained when' mthe primary electron acceptor of photosystem II, Q, is fully reduced
(closed traps, P680Q ) and Fq is obtained when Q. is fully oxidized
(open traps, P680Q) (Butler 1978).

- 7 -
3.2 Methods to induce fluorescence yield changes
Changes in fluorescence yield that reflect changes in the energy con
version efficiency of photosystem II were first reported by Kautsky and
Hirsch (1931). Measurements of kinetics and yield of Fy have shown
that Fy can be quenched if plants are exposed to stress e.g. chilling
treatment (Smillie 1979) or prolonged water stress (Govindjee et al.
1981). Chlorophyll fluorescence has also been used as a probe to in
vestigate stress induced effects on the thylakoid membrane such as
1) heat induced separation of the light harvesting chlorophyll from
photosystem II (Schreiber and Armond 1978), 2) temperature induced
phase transitions in chilling sensitive plants (Murata et aj_. 1975)
and 3) temperature jump effects (Schreiber ejt <al_. 1976, Schreiber and
Berry 1977).
3.2.1. Temperature induced fluorescence changes
A fluorescence increase from Fq to F^ was induced through progressive
lowering of the temperature from 20 °C to -20 °C, at a constant cooling
rate of 1 °C per minute. The method is described and evaluated in paper
II and III. Lowering the temperature towards the freezing point induces
a linear increase in the fluorescence yield. At the same time the elec
tron transport between photosystem II and photosystem I is linearly in
hibited (Thorne and Boardman 1971, Nolan and Smillie 1977, Smillie and
Nott 1979). At the freezing point, the fluorescence shifts under influ
ence of exothermic heat, given off when ice is formed (Melcarek and
Brown 1979). Differences observed in the kinetics of the fluorescence
changes were related to the structure of the leaf, i.e. mesomorphic or
xeromorphic characters. Wheter applied to intact leaves or to isolated
chloroplasts, the method can reveal if the photosynthetic electron

- 8 -
transport is inhibited on the oxidizing or the reducing side of photo
system II (paper II, III). We suggest that the method provides a valu
able tool for investigation of effects of environmental stress imposed
on the function of photosynthesis.
3.2.2. Freeze-thaw induced fluorescence changes
In successively increased freeze-thaw stress, through repeated freeze-
-thaw cycles between 20 °C and -20 °C, principally two different
schemes of reaching a totally supressed variable fluorescence were ob
served (paper II). The variable fluorescence was eventually supressed
and invariable F or F was reached depending on the site of inhibition, o m r a
F or F was reached if inhibition of the electron transport occurred o m ron the oxidizing or reducing side of photosystem II, respectively. This
was shown in experiments with frost hardened and unhardened pine seed
lings. Furthermore, freeze-thaw experiments with different species
clearly discriminates between very frost sensitive (potato), moderately
frost resistent (pea,spi nach,summer active pine) and frost resistent
(hardened pine) plants.

- 9 -
4. APPLICATION OF TEMPERATURE INDUCED FLUORESCENCE CHANGES
4.1 Photosynthesis and temperature induced fluorescence changes in birches adapted to different temperatures
Adaptation and acclimation of plant photosynthetic reactions to tempe
rature has recently been described in detail by Berry and Björkman (1980)-
In Scandinavian mountains Betula tortuosa forms the tree limit. Whether
Jî. tortuosa d i ffers from B. pubescens in its ability to ma inta in a high
rate of photosynthesis at low temperatures was examined in paper IV. The
results show that jB. tortuosa has a higher rate of net CC^ exchange at
low temperatures than J3. pubescens. There was marked difference between
the birches in their ability to withstand freezing, as revealed from the
measurements of temperature induced fluorescence changes. It was conclu
ded that J5. tortuosa exhibits an adaptation to the prevailing climate in
the mountain region.
4.2 Temperature induced fluorescence changes:A method in screening for frost tolerance
Freezing injury in plants occurs over a broad spectrum of sub-zero
temperatures. The frost killing temperature depends on the actual species
and on the stage of acclimation. Although there is a great diversity in
the range of sub-zero temperatures that can be tolerated, there is how
ever one common feature involved in freezing tolerance. Membrane damages
are commonly inferred to be the primary effect of frost injuries (Lyons
jet a_K 1979). Further, the rapidity with which injury is manifested
suggests that the freezing injury is not due to metabolic disturbances,
as in chi 11ing injury.
In almost every country where potatoes are cultivated frosts during the
growing season are a well known problem. I northern Sweden frost occurs

- 10 -
frequently during the last month before the crop Solanum tuberosum
reaches maturity. Temperature induced fluorescence changes of different
and closely related clones of tuberosum were measured in one freeze-
thaw cycle between 20 °C and -10 °C (paper V). The short term frost
simulation by the freeze-thaw cycle and its influences on the fluore
scence changes was compared with long term frost treatment in a climate
chamber. The results of the two separately performed "blind-tests" were
identical with respect to their ranking of the different clones for
frost tolerance. The temperature induced fluorescence changes also moni
tored progressive damages to the chloroplast membranes when plants were
exposed to successively lower temperatures. It was found that the diffe
rences in freezing temperature of the leaves of the different clones
did not correlate with the estimated frost tolerance in the climate
chamber test or fluorescence test.
The fluorescence pattern observed indicates a primary effect of the
freezing injury to the oxidizing side of photosystem II, since variable
fluorescnece decreased to F .o
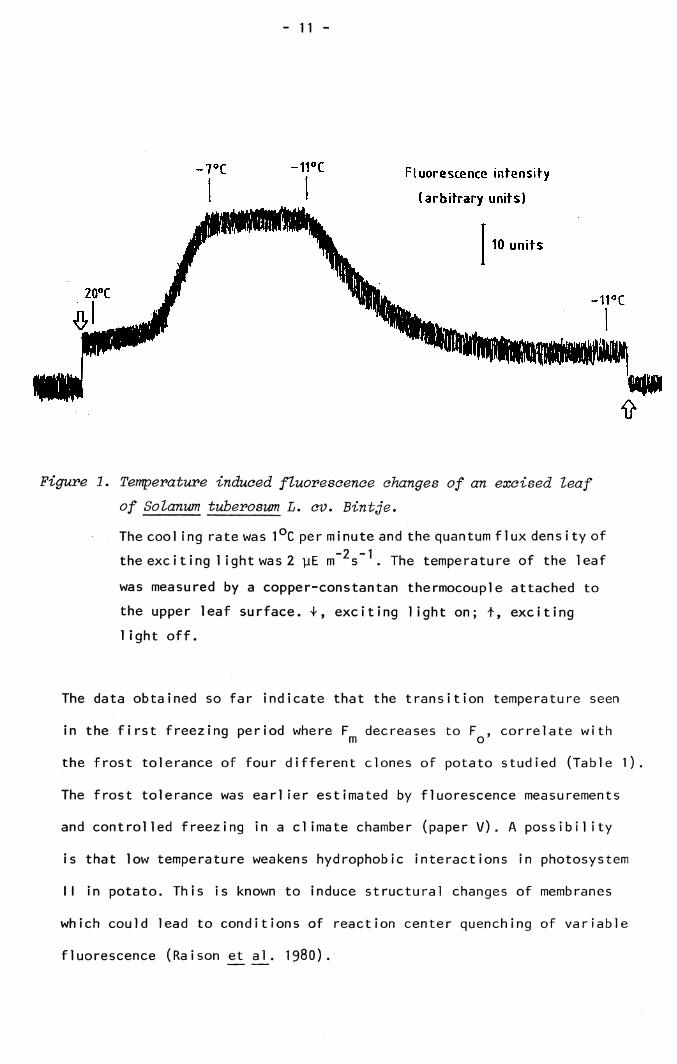
An interesting discovery was made on the low temperature induced Fm
fluorescence in potato. As shown in paper V, invariable Fq was induced
after one freeze-thaw cycle. This situation also occurs in the first
freezing period introduced, if the temperature falls below a critical
value (Figure 1). This temperature is found to be -11 °C for the potato
cultivar Bintje, irrespective of the time (5-30 minutes) of exposure to
sub-zero temperatures above -11 °C. Hence the low temperatures of the
first freezing period causes irreversible inhibition of the electron
transport of the oxidizing side of photosystem II.
- 11 -
Fluorescence intensity
(arb i t r ary units)
10 un i ts
Figure 1. Temperature induced fluorescence changes of an excised leaf of Solanum tuberosum L. cv. Bintje.
The cool ing rate was 1°C per minute and the quantum flux density of-2 -1the exciting 1 ight was 2 yE m s . The temperature of the leaf
was measured by a copper-constantan thermocouple attached to the upper leaf surface. T, exciting light on; t, exciting 1ight off.
The data obtained so far indicate that the transition temperature seen
in the first freezing period where F decreases to F , correlate with3 r m o ’the frost tolerance of four different clones of potato studied (Table 1).
The frost tolerance was earlier estimated by fluorescence measurements
and controlled freezing in a climate chamber (paper V). A possibility
is that low temperature weakens hydrophobic interactions in photosystem
II in potato. This is known to induce structural changes of membranes
which could lead to conditions of reaction center quenching of variable
fluorescence (Raison et al. 1980).

- 12 -
Table 1. Transition temperature of decrease to Fq of four different clones of potato, S. tuberosum. X ± SE, n - 1-5.
-, * transitionClone no. . /O-x ntemperature ( C)
1 - 11,0 ± 0,5 52 -11,3 1k -12,7 (±0,7) 25 - 12,0 1
*The clone no. is identical with the clone no. used in paper V.
5. ACKNOWLEDGEMENTS
I wish to express my sincere thanks to all members of the Department of
Plant Physiology, University of Umeå. I am especially grateful to my
supervisor and friend Docent Gunnar öquist. This work could not have
been accomplished without his valuable ideas, encouragement and never
failing spirit. I am also grateful to Professor Lennart Eliasson for his
support and for introducing me to the field of agricultural plant physio
logy, and to Professor Lars Olov Björn for fruitful cooperation. The
close cooperation with Doctor Jan-Erik Hällgren is deeply acknowledged.
This work was supported by grants from the Swedish Natural Science
Research Council, Swedish Counsil for Forestry and Agricultural Research
and National Swedish Board for Technical Development.

- 13 -
REFERENCES
Barbierie, G., Delosme, R. and Joliot, P. (1970). Photochem. Photobiol. 12, 187.
Berry, J. and Björkman, 0. (1980). Ann. Rev. Plant Physiol. 31, 491. Bertch, W. (1962). Proc. Nat. Acad. Sei. U.S. 48, 2000.Björn, L.O. (1971). Photochemistry and Photobiology 13, 5.Björn, L.O. and Forsberg, A.S. (1979). Physiol. Plant. 47, 215.Butler, W.L. (1978). Ann. Rev. Plant Physiol. 29, 345- Clayton, R.K. (1969). Biophys. J. 9, 60.Goedheer, J.C. (1962). Biochim. Biophys. Acta 64, 29.Goedheer, J.C. (1963). Biochim. Biophys. Acta 66, 6.Govindjee, Downton, W.J.S., Fork, D.C. and Armond, P.A. (1981). Plant
Sei. Lett. 20, 191.Govindjee änd Govindjee, R. (1975). In: Bioenergetics of Photosynthesis
(Govindjee ed.) pp 1-50, Academic Press, New York.Joliot, P., Joliot, A., Bouges, B. and Barbieri, G. (1970). Photochem.
Photobiol. 12, 287.Kamen, M.D. (1963). Primary Processes in Photosynthesis. Academic Press,
New York.Kautsky, H. and Hirsch, A. (1931). Naturwissenschaften 19» 964.Lavorel, J. (1959). Plant Physiol. 34, 204.Lavorel, J. (1968). Biochim. Biophys. Acta 153, 727.Lavorel, J. (1975). In: Bioenergetics of Photosynthesis (Govindjee ed.)
pp 223-317, Academic Press, New York.Lyons, J.M., Raison, J.K. and Steponkus, P.L. (1979). In: Low Temperature
Stress in Crop Plants, The role of the membrane (James M. Lyons,Douglas Graham, John K. Raison Eds.) pp 1-24, Academic Press, NewYork.
Melcarek, P.K. and Brown, G.N. (1977). Plant and Cell Physiol. 18, 1099.Melcarek, P.K. and Brown, G.N. (1979). Cryobiology 16, 69.Murata, N., Troughton, J.H. and Fork, D.C. (1975). Plant Physiol. 56,
508.Nolan, W.G. and Smillie, R.M. (1977). Plant Physiol. 59, 1141.Raison, J.K., Berry, J.A., Armond, P.A. and Pike, C.S. (1980) . In:
Adaption of Plants to Water and High temperature Stress (Neil C. Turner and Paul J. Kramer Eds.) pp 261-273, John Wiley and Sons, New York.

- I lf -
Schreiber, U. and Armond, P. (1978). Biochim. Biophys. Acta 502, I38. Schreiber, U. and Berry, J.A. (1977). Planta I36, 233.Schreiber, ü., Colbow, K. and Vidaver, W. (1976). Biochim. Biophys. Acta
1|23, 249.Smillie, R.M. (1979). In: Low Temperature Stress in Crop Plants, The
Role of the Membrane (James M. Lyons, Douglas Graham, John K. Raison Eds.) pp 187“202, Academic Press, New York.
Smillie, R.M. and Nott, R. (1979). Plant Physiol. 63, 796.Strehler, B.L. and Arnold, W. (1951). J. Gen; Physiol. 3^, 809.Thorne, S.W. and Boardman, N.K. (1971). Biochim. Biophys. Acta 23A, 113-