Embed Size (px)
Citation preview

Insect Biochem. 1971, I, 19-26. [Scientechnica (Publishers) Ltd.] 19
M I C R O S O M A L N A D P H * - C Y T O C H R O M E c R E D U C T A S E
F R O M T H E H O U S E F L Y , M U S C A D O M E S T I C A
S O L U B I L I Z A T I O N A N D P U R I F I C A T I O N
THOMAS G. WILSON AND ERNEST HODGSON
Department of Entomology, North Carolina State University, Raleigh, North Carolina 276o7, U.S.A.
(Received 20 July, 197o)
ABSTRACT NADPH-cytochrome c reductase (E.C. 1.6.2.3) has been solubilized from
Musca domestica microsomes by treatment with isobutanol. The soluble enzyme has been partially purified by negative adsorption on zinc phosphate followed by chromatography on DEAE-cellulose and hydroxylapatite columns. The enzyme obtained by the above procedure is a flavoprotein with a molecular weight of approximately 57,ooo, as determined by Sephadex gel chromato- graphy.
NADPH-cYTOCHROME c reductase (E.C. 1.6.2.3) , an enzyme capable of reducing cyto- chrome c in the presence of NADPH, was first isolated from mammals by Horecker (I95O), who isolated it as a particulate enzyme from pig liver. Later, both Williams and Kamin (1962) and Phillips and Langdon (1962) solubilized the enzyme from mammalian microsomes and reported it to be a small molecular weight flavoprotein capable of reducing several dyes, ferricyanide, and inorganic iron. Nishibayashi, Omura, and Sato (1963) isolated a menadione-requiring NADPH oxidase from rabbit liver miCrosomes and recently postulated its identity with NADPH-cytochrome c reductase (Nishibayashi- Yamashita and Sato, 197o ). Iyanagi and Yamazaki (1969) isolated an NADPH-cyto- chrome c reductase from pig liver capable of reducing quinones. Ichikawa and Yamano (1969) isolated an NADPH-cytochrome c reductase from rabbit liver microsomesand reported trans-hydrogenase as well as weak microsomal cytochromes bs, P450, and P4ao reductase activity. Masters, Bilimoria, Kamin, and Gibson (1965) and Masters, Kamin, Gibson, and Williams (1965) elucidated the mechanism of electron transfer by pig liver NADPH-cytochrome c reductase.
The only previous reports on this enzyme in insects were by Lang (1959, 196i), who studied NADPH-cytochrome c reduction in mosquito mitochondrial supernatants and concluded that the activity was due to an NADPH reductase system and not trans- hydrogenation followed by NADH oxidation. NADPH-cytochrome c reductase activity was shown to be twice as high in mosquito larvae as in the adult insects.
NADPH-cytochrome c reductase is a component of the NADPH-cytochrome P45o mixed function oxidase system (Kamin and Masters, i968), which oxidizes, among other
* The following abbreviations a r e u s e d throughout: NADPH for reduced nicotinamide adenine dinucleotide phosphate; DEAE-cellulose for diethylaminoethyl-cellulose; FAD for flavin adenine dinucleotide, FMN for flavin mononucleotide.

20 WILSON AND HODGSON Insect Biochem.
xenobiotics, insecticides (O'Brien, 1967; Terriere, I968 ). An understanding of this enzyme in insects could be of interest not only to the comparative biochemist but also to the comparative toxicologist. In the present communication solubilization, purification, and some physical properties are described.
MATERIALS AND M E T H O D S
CHEMICALS
NADPH, trypsin, lipase, and DEAE-cellulose were obtained from Nutritional Biochemieals Company. Ovalbumin, myoglobin, FAD, FMN, and type I I I cytochrome c were obtained from Sigma Biochemicals Company. Bovine serum albumin, fraction V, was obtained from Armour Pharmaceutical Company. All other reagents were A.C.S. or analytical grade.
The zinc chloride solution used in the purification procedure was prepared by titrating a o" IO M ZnCl~ solution to pH 5"8 and removing the precipitated zinc hydroxide by centr ifugation. The supematant was used as the zinc chloride solution.
Rapid-flow hydroxylapatitc was prepared according to the procedure of Anacker and Stoy (~958).
PREPARATION OF MICROSOMES
Adult female houseflies of the CSMA strain, originally obtained from the Union Carbide Chemical Company, and maintained since x962 at North Carolina State University, were used in all experiments. Five- to seven-day-old flies were immobilized with carbon dioxide and cooled to o-5 ° C. The abdomens were removed and submerged in ice-cold distilled water; afterwards, they were drained, blotted, and weighed. The abdomens were then homogenized in a volume of o'I5 M phosphate buffer, pH 7"8, equal to 4-5 times their weight on a g. per ml. basis, in a Sorvall omnimixer at 75 per cent maximum speed for 3 minutes. The homogenate was filtered through x layer of No. IO cheese-cloth and centrifuged at lO,OOO g for io minutes in a SorvaU refriger- ated centrifuge to sediment debris, nuclei, and mitochondria. The supernatant was filtered through glass wool and centrifuged at ioo,ooo g for I hour in a Beckman L-2 ultracentrifuge. The microsomal pellet was resuspended in a homogenizer consisting of a cone-driven teflon pestle and a glass tube, in a volume of buffer equivalent to o '5-I times the original homogenization volume. Microsomes were resedimented at ioo,ooog and resuspended in 37 ml. of buffer. The micro- somal suspension was usually frozen overnight at this stage.
NADPH-CYTOCHROME c REDUCTASE PURIFICATION The microsomal suspension was diluted to xSo ml. with distilled water, and o'3 volumes of
isobutanol were slowly stirred in over a 3o-minute period. Stirring continued for an additional 2-3 hours. The suspension was then centrifuged at xo,ooo g for xo minutes, and the upper (iso- butanol) layer was removed by suction. The aqueous layer was filtered through glass wool and centrifuged at ioo,ooo g for I hour. A small, almost crystal-clear residue precipitated, which contained no reductase activity. The supernatant from this centrifugation was stirred rapidly while 1 I 'o ml. of zinc chloride solution was added over a i -minute period. The suspension was stirred for an additional 4 minutes and immediately centrifuged at io,ooog for io minutes to sediment the zinc phosphate precipitate, which was discarded. Concentration and time of stirring are critical; greater amounts of ZnC12 added or longer stirring time results in considerable enzyme adsorption on to the ZnHPO4 precipitate. The supernatant was then added directly to a I"7 × 2o crn. DEAR-cellulose column, previously equilibrated and washed with o'o4 M phosphate buffer, pH 7"8, and the column was then washed with equilibrating buffer. The enzyme was eluted with a linear gradient of xoo ml. each of o'o4-o'4 M phosphate buffer, pH 7"8, at a flow- rate of o'5 ml. per minute. Fraction volumes of 4'8 ml. were collected in a Gilson constant- volume fraction collector. Fractions containing enzyme were frozen at - - I5 ° C.
For preparation of rat or sheep liver enzyme, the procedure was the same except that 12-I6 g. of rat liver (Sprague-Dawley male white rats) or 5-6 g. of sheep liver (obtained at a local abattoir and frozen at --15 ° C. until used) were used instead of housefly abdomens.
For hydroxylapatite chromatography the enzymatically active fractions (above x-o unit per ml.) from the DEAR-cellulose column were diluted to 9 °ml . , a threefold dilution, with distilled

I 9 7 I , I HOUSEFLY NADPH-CYTOCHROME C REDUCTASE 2I
water and loaded on to a I '7 × x2 cm. hydroxylapatite column equilibrated and washed with 0"04 M phosphate buffer, p H 7"8. The column was eluted immediately with a linear gradient of Ioo ml. each of o.o4-o'3 M phosphate buffer, p H 7"8, at a flow-rate of o'4 ml. per minute. Fract ion volumes of 4"8 ml. were collected, and enzymatically active fractions were frozen at --15 o C.
ASSAYS
All enzymatic assays were performed in a Beckman DB spectrophotometer at room temperature (25-27 ° C.). Absorbance was read directly from the spectrophotometer, usually for the first I '5 minutes of the reaction. The rate of cytochrome c reduction was linear during the initial 3-4 minutes of the reaction, and a plot of cytochrome c reduced versus amount of enzyme intersected the origin and remained linear beyond amounts of enzyme used in the reaction. No reduction of cytochrome c occurred in the absence of N A D P H or enzyme. All assays were initiated by addition of NADPH. Potassium phosphate with an ionic strength of o . io at pH 7"8 was used as the assay buffer.
NADPH-cytochrome c reductase was assayed by measuring the increase in ahsorhance at 55o nm. The cuvette contained 3"o × IO -7 moles of NADPH. The molar extinction coefficients used for reduced and oxidized cytochrome c at 55o nm. were 27"7 x io 3 M -2 cm.-1 and 9.0 × io 3 M -2 c m . - 2 respectively (Margoliash and Frohwirt, i959).
Protein was determined by the method of Miller (I959), using bovine serum albumin as a standard.
The molecular weight of the enzyme was determined by Sephadex gel filtration (Andrews, 2964). The flow-rate was I2 mL per hour through a 3 × 35 cm. column. Marker proteins used were cytochrome c, molecular weight 12,4oo; ovalbumin, molecular weight 43,500; and myo- globin, molecular weight 17,8oo (cited by Andrews, i964). NADPH-cytochrome c reductase from a peak tube from DEAE-ceUulose chromatography was concentrated with dried polyacrylamide gel particles to o'5 ml. and this, plus 2-4 rag. ovalbumin, was applied to the column. Cytochrome c or myoglobin was identified in the fractions by colour and absorbance at 28o nm., ovalbumin by absorbance at 28o nm., and NADPH-cytochrome c reductase by enzymatic activity.
Polyacrylamide disk electrophoresis was performed as described by Davis (1964). Electro- phoresis was carried out at 3 mA. per tube in o'o6 M Tris-glycine buffer, p H 7"4, for about 9o minutes (until the tracker dye, bromthymol blue, reached the bottom of the tubes). The gels were divided into two groups; half of the gels were stained for 3o minutes in i per cent naphthol blue- black in 7 per cent acetic acid and destained electrophoretically in 7 per cent acetic acid. Sections were sliced from the unstained half at positions corresponding to protein bands in the stained gels. These sections were homogenized for I minute in 5"o ml. of enzyme assay buffer in a Sorvall omnimixer with micro attachment, extracted overnight in buffer, and the supernatant was assayed for enzyme activity.
The prosthetic group was dissociated from the apoenzyme of NADPH-cytochrome c reductase by acid ammonium sulphate as described by Warburg and Christian (1938) and modified as follows: before ammonium sulphate addition, approximately I mg. per ml. of bovine serum albumin was dissolved in the enzyme solution to act as a carrier protein; after ammonium sulphate addition the enzyme solution was titrated to p H 4"1. Centrifugation then yielded the sedimented apo- enzyme.
R E S U L T S A N D D I S C U S S I O N
NADPH-CYTOCHROME c REDUCTASE PURIFICATION
Table I shows the pur i f i ca t ion of housef ly N A D P H - c y t o c h r o m e c r educ ta se o b t a i n e d wi th t he p r o c e d u r e ( t h r o u g h the D E A E - c e l l u l o s e c h r o m a t o g r a p h y s tep) ou t l i ned in Ma te r i a l s and M e t h o d s .
A buf fe r concen t r a t i on of o . i 5 M p h o s p h a t e buf fe r or a p r o t e i n concen t r a t i on grea te r t h a n 2 - 3 rag. p e r ml . in the mic ro soma l suspens ion r e su l t ed in i nc omple t e e n z y m e ex t rac t ion wi th i sobu t ano l ; hence , t he mic ro soma l su spens ion is d i l u t ed be fore iso- b u t a n o l ex t rac t ion . I s o b u t a n o l acts as a de t e rgen t to so lubi l ize the enzyme ( M o r t o n ,
x955).

22 WILSON AND HODGSON Insect Biochem.
Usually, all of the enzyme assayed in the microsomal suspension was recovered by extraction with isobutanol. Frequently, up to 12o per cent of the original enzyme activity was recovered in the soluble fraction. This may be due to ' l a ten t ' enzyme located in the microsomal membranes in such a position as to be unavailable to the sub- strate during assay, but freed by isobutanol. However, the possibility of isobutanol activation was not eliminated. Release o f ' latent ' enzyme was observed by Phillips and Langdon (i962) when they solubilized the enzyme from rat liver with trypsin. No NADPH-cytochrome c reductase has been assayed in resuspended mitochondria, even after extraction with isobutanol.
NADPH-cytochrome c reductase from houseflies could be solubilized with crude pancreatic lipase (o. 7 rag. per ml. microsomal suspension) by stirring at 5 ° C. for 20 hours, and 95 per cent of the enzyme was recovered with a release of 25 per cent of the microsomal protein. Solubilization by 2 × crystallized trypsin (o. 5 rag. per ml. micro- somal suspension) by stirring at 5 ° C. for 2. 5 hours yielded 4 ° per cent of the enzyme
Table/.--PURIFICATION OF HOUSEFLY NADPH-cYTOCHROME c REDUCTASE
FRACTION
Microsomes Isobutanol supernatant ZnHPO~ supernatant DEAE eluant
SPECIFIC ACTIVITY (units per rag. protein)
I'4 5"6 8"4
6I'O
TOTAL UNITS
i6o I98 195 IIO
3:3 g. of housefly abdomens were used in this purification.
PERCENTAGE RECOVERY
I00
x24 122
69
PURIFICATION
I'OX
4.o× 6"0 ×
44.o x
and completely digested the microsomes. Lipase has been used to solubilize pig liver reductase (Williams and Kamin, i962), and trypsin solubilizes rat liver reductase (Phillips and Langdon, i962; Omura and Takesue, I97o ). However, attempts to solubilize pig liver reductase with n-butanol yielded poor results (Williams and Kamin, 1962).
NADPH-cytochrome c reductase from houseflies can be precipitated from the iso- butanol aqueous layer by stirring in o'9 volumes of acetone ( - I 5 ° C.) for 5 minutes, sedimenting by centrifugation, and redissolving in buffer with a recovery of 6o-9o per cent of the enzyme activity. Attempts to purify the reductase from the isobutanol aqueous layer by acetone fractionation yielded erratic results and were abandoned. Precipitation of the enzyme by acetone is a function of temperature and the amount of isobutanol dissolved in the aqueous layer; since these varied from preparation to preparation, a volume of acetone which gave consistent results could not be established.
The elution pattern of NADPH-cytochrome c reductase from a DEAE-cellulose column is shown in F/g. i. The enzyme was eluted from the column at approximately o.o 7 M phosphate buffer.
NADPH-cytochrome c reductase can be further purified by hydroxylapatite chromato- graphy. An elution pattern is shown in Fig. 2; the reductase was eluted from the column at a lower molarity than from DEAE-cellulose. The coincidence of the enzyme peak with a 28o nm. absorption peak is a good indication of the predominance of enzyme among the proteins in that peak.

197 I, I HOUSEFLY NADPH-CYTOCHROME c REDUCTASE 2 3
The electrophoretic pattern of the enzyme after DEAE-ceUulose and hydroxylapatite purification is shown in Fig. 3. The enzyme band was considerably enhanced by hydroxylapatite chromatography, but contaminating protein was still present. By visual examination of the post-hydroxylapatite gel, NADPH-cytochrome c reductase was estimated to be 7o-8o per cent of the total protein present.
Although whole flies could be used for preparation of NADPH-cytochrome c reduc- tase, both total and specific activity of the enzyme in the microsomal suspension were
>,. i - o3 Z hi E~
J (D
0.6
0.4
I . - ~0.2 =1. E o 06 N
o-OPTICAL DENSITY
I I ...... I I0 20 30
FRACTION NUMBER
6.0
r-- n- o
4.0w < 13.
ttl
2.0 ~=~ - - > -
g... 0 ta
FIG. z .--DEAE-cel lulose chromatography of the NADPH-cytochrome c reductase from Musca microsomes following negative adsorption on zinc phosphate, z9o units applied to the column, zz8 recovered. Ninefold purification.
>- 1-- O3 z 0,06 Ld C3 /
• ¢ 0.04 ¢o I.-
o 0.02
E
_ . 0 - 2 8 0 m/~ O D
/ ~ D-ENZYME ACTIVITY
3.0 _
2.0,,, Po
o3 I.=J
Z > - ~ N
Z 0 "' o 0 .... ~ I, I
oo 0 I0 20 30 oJ
FRACTION NUMBER FXG, z . - -Hydroxylapat i te chromatography of the NADPH-cytochrome c reductase
from Musca microsomes following DEAE-ceUulose chromatography. 97 units applied to column, 44 units recovered. PT-fold purification.
lower, and frequently large losses of enzyme activity occurred at one or more of the purification steps. This may be due to endogenous inhibitors occurring in the head and/ or thorax (Matthews and Hodgson, x966 ). Enzyme activity is low (less than 15 per cent) in the head and thorax as compared with that in the abdomen. Thus, abdominal excision is a purification step.
This purification method has been attempted for NADPH-cytochrome c reductase from sheep and rat liver microsomes. Although isobutanol did solubilize the enzyme, recovery from that step and remaining purification steps was low. Only trace amounts of enzyme could be recovered from the DEAE-cellulose column.

24 WILSON AND HODGSON Insect Biochem.
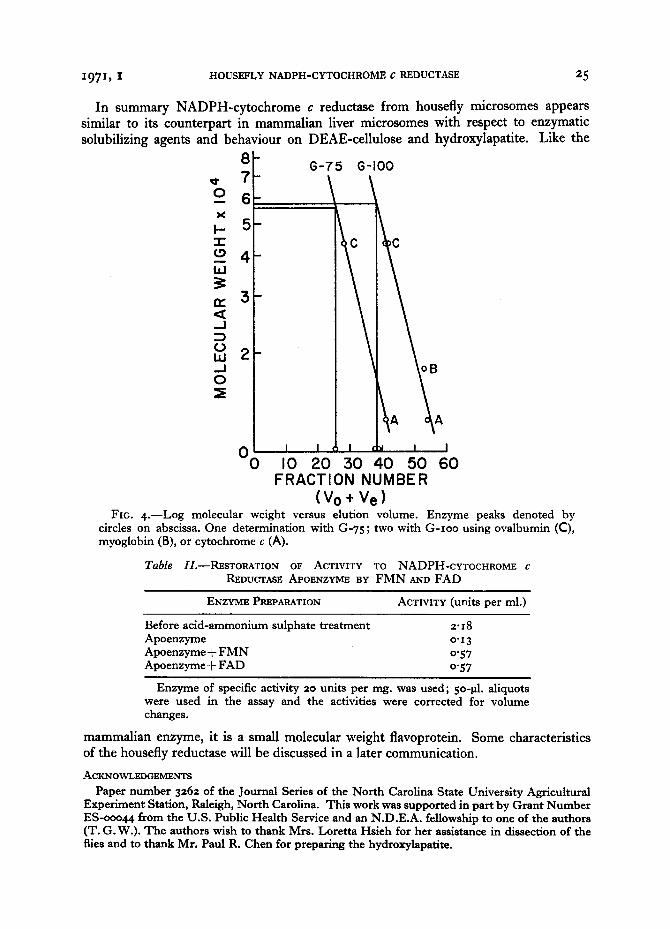
MOLECULAR WEIGHT
The molecular weight of NADPH-cytochrome c reductase was determined by Sephadex gel filtration as outlined in Materials and Methods. Using a plot of log molecular weight versus elution volume (Andrews, 1964) for the enzyme and marker proteins (Fig. 4), the molecular weight of NADPH-cytochrome c reductase was deter- mined to be 57,ooo, an average of 3 separate determinations. This molecular weight is similar to those reported for the same enzyme from mammals, ranging from 5%o00 (Phillips and Langdon, 1962 ) to 89,ooo (Nishibayashi-Yamashita and Sato, 197o ).
A
B c÷) FIG. 3.--Electrophoresis of NADPH-cytochrome c reductase from Musca microsomes
purified by DEAE-cellulose and hydroxylapatite chromatography. Right gel: protein from DEAE column. Left gel: protein from HA column. The middle band (A) contained NADPH-cytochrome c reductase activity. The large bottom band (B) is bromthymol blue (tracker dye). Impurities in hydroxylapatite-purified enzyme consist of a single band above the tracker dye band and a slow band just below the stacking gel. Protein applied to each gel was 15 gg. from a DEAE-cellulose column (specific activity 98 units per mg.) and 19 ~tg. from a hydroxylapatite column (specific activity 155 units per mg.).
PROSTHETIC GROUP The apoenzyme of NADPH-cytochrome c reductase was prepared as outlined in
Materials and Methods. The sedimented apoenzyme was dissolved in assay buffer and divided into 2 equal portions. To one portion lO -8 M FAD was added, and lO -3 M FMN was added to the other. After IO minutes' incubation at o ° C., the fractions were assayed for activity, and the results are shown in Table II. Both FAD and FMN have equal ability to restore activity to the apoenzyme. Unfortunately, the enzyme is not present in sufficient quantity in 3 ° g. of housefly abdomens to establish the identity of the prosthetic group clearly or to obtain an adsorption spectrum of the flavoprotein. The small quantity of enzyme present in 3 ° g. of housefly abdomens is reflected by the observation that no yellow colour, a flavoprotein characteristic, could be seen at any stage of the reductase purification. Reports on mammalian enzyme indicate, for example, that the solubilized enzyme solution is yellow in colour (Horecker, 195o; Phillips and Langdon, 1962; Williams and Kamin, 1962).
i971, I HOUSEFLY NADPH-CYTOCHROME C REDUCTASE 25
In summary NADPH-cytochrome c reductase from housefly mierosomes appears similar to its counterpart in mammalian liver microsomes with respect to enzymatic solubilizing agents and behaviour on DEAE-cellulose and hydroxylapatite. Like the
o_ X
I- " l -
la.I
_.1
t . ) ia.I ._1 0
8 7
6
5
4
3
2
O0 I0 20 30 40 50 60 FRACTION NUMBER
(Vo+ Ve) FIG. 4 . - -Log molecular weight versus elution volume. Enzyme peaks denoted by
circles on abscissa. One determination with G-75; two with G- Ioo using ovalbumin (C), myoglobin (B), or cytochrome c (A).
Table //.--RESTORATION OF ACTIVITY TO NADPH-cYTOCHROME c REDUCTASE APOENZYME BY F M N AND FAD
ENZYME PREPARATION ACTIVITY (units per rnl.)
Before acid-ammonium sulphate treatment 2.i 8 Apoenzyme o" 13 Apoenzyme + F M N 0"57 Apoenzyme + FAD 0.57
Enzyme of specific activity 20 units per nag. was used; 5o-gl. aliquots were used in the assay and the activities were corrected for volume changes.
mammalian enzyme, it is a small molecular weight flavoprotein. Some characteristics of the housefly reduetase will be discussed in a later communication.
ACKNOWLEDGEMENTS
Paper number 3262 of the Journal Series of the North Carolina State University Agricultural Experiment Station, Raleigh, North Carolina. This work was supported in part by Grant Number ES-ooo44 from the U.S. Public Health Service and an N.D.E.A. fellowship to one of the authors (T. G. W.). The authors wish to thank Mrs. Loretta Hsieh for her assistance in dissection of the flies and to thank Mr. Paul R. Chen for preparing the hydroxylapatite.

26 WILSON AND HODGSON
REFERENCES
ANACKER, W. F., and STOY, V. (1958), ' Protinchromatographie an calcium-phosphate '. Biochem. Z., 330, 141-I59.
ANDREWS, P. (I 964), ' Estimation of the molecular weights of proteins by Sephadex gel-filtration ', Biochem. ~., 9I, 222-233.
DAVIS, B. J. (1964), ' Disc electrophoresis. II . Method and application to human serum proteins ', Ann. N.Y. Acad. Sci., I l I , 404-427.
HORECKER, B. L. (195o), ' Triphosphopyridine nucleotide-cytochrome c reductase in liver ', J . biol. Chem., I83, 593-6oo.
ICHIKAWA, Y., and YAMANO, T. (1969), ' Studies on the microsomal reduced nicotinamide adenine dinucleotide phosphate-cytochrome e reductase from rabbit liver' , ~. Bioehem., Tokyo, 66, 351-359.
IYANAGI, T., and YAMAZAKI, I. (I969), ' One-electron-transfer reactions in biochemical systems. I I I . One-electron reduction of quinones by microsomal flavin enzymes ', Biochim. biophys. Acta, 172 ~ 37o-381.
KAMIN, H., and MASTERS, B. S. S. (I968), ' Electron transport in microsomes ', in Enzymatic Oxidation of Toxicants (ed. HODGSON, E.), pp. 5-26. Raleigh: North Carolina State University.
LANO, C. A. (I959), ' Cytochrome c reductase activities during development ', ExpI Cell Res., 17, 516-518.
LANG, C. A. (I96I), ' TPNH-cytochrome c reductase in the mosquito ', Fedn Proc. Fedn Am. Socs exp. Biol., 2o, 47.
MARGOLIASH, E., And FROHWlRT, N. (I959), ' Spectrum of horse-heart cytochrome c ', Biochem.J., 7I, 57o-572.
MASTERS, B. S. S., BILIMORIA, M. H., KAMIN, H., and GIBSON, Q. H. (I965), ' The mechanism of i - and 2-electron transfers catalyzed by reduced triphosphopyridine nucleotide-cytochrome c reductase ', ~. biol. Chem., 24o, 4o81-4o88.
MASTERS, B. S. S., KAMIN, H., GIBSON, Q. H., and WILLIAMS, C. H. , jun . (I965), ' Studies on the mechanism of microsomal triphosphopyridine nucleotide-cytochrome c reductase ', ] . biol. Chem., 2 0 , 921-931.
MATTHEWS, H., and HODGSON, E. (I966), 'Na tu ra l ly occurring inhibitor(s) of microsomal oxidations from the housefly ', ~. econ. Ent., 59~ 1286-1288.
MILLER, G. L. (I959) , ' Protein determination for large numbers of samples ', Analyt. Chem., 3I, 964.
MORTON, R. K. (I955), ' Methods of extraction of enzyme from animal tissue ', in Methods of En¢ymology (ed. COLOWlCK, S. P., and KAPLAN, W. O.), vol. I, pp. 25-51. New York: Academic Press.
NISHIBAYASHI, H., OMURA, T., and SATO, R. (i963) , ' A flavoprotein oxidizing N A D P H isolated from liver microsomes ', Biochim. biophys. Acta, 67, 520-522.
NISHIBAYASHI-YAMASHITA, H., and SATO, R. (197o), ' Vitamin K3-dependent N A D P H oxidase of liver microsomes ', j~. Bioehem., Tokyo, 67, 199-21o.
O'BRIEN, R. D. (I967), Insecticides-Action and Metabolism. New York: Academic Press. OMURA, T., and TAKESUE, S. (I97O), ' A new method for simultaneous purification of cytochrome
b~ and NADPH-cytochrome c reductase from rat liver microsomes ', .7. Biochem., Tokyo, 67, 249-257.
PHILLIPS, A. H., and LANODON, R. G. (I962), 'Hepa t i c triphosphopyridine nucleotide-cyto- chrome c reductase: Isolation, characterization, and kinetic studies ', ~. biol. Chem., 237, 2652- 2660.
TERRIERE, L. C. (1968), ' The oxidation of pesticides: The comparative approach ', in Enzymatic Oxidation of Toxicants (ed. HODQSON, E.), pp. 175-196. Raleigh: North Carolina State University.
WARBURG, O., and CHRISTIAN, W. (1938), ' Isolierung der Prosthetischen gruppe der d-amino- saure oxydase ', Biochem. Z., 298, 15o-168.
WILLIAMS, C. H., jun., and KAMIN, H. (x962), ' Microsomal triphosphopyridine nucleotide- cytochrome e reductase: isolation, characterization, and kinetic studies ', J . biol. Chem., z37, 587-595.
Key Word Index: Microsomes, NADPH-cytochrome c reductase, Musca domestica.





























