Embed Size (px)
Citation preview

Insect Biochem., Vol, 9. pp. 43 t048. 0020-1700/79/0201-0043 $02.00/0 t~) Pergamon Press Ltd. 1979. Printed in Great Britain
MICROSOMAL N A D P H - C Y T O C H R O M E c R E D U C T A S E FROM THE M I D G U T OF THE S O U T H E R N A R M Y W O R M
(SPODOPTERA ERIDANIA) D, L. CRANKSHAW, H. K. HETNARSKI and C. F. WILKINSON
Department of Entomology, Comstock Hall, Cornell University, Ithaca, NY 14853, U.S.A.
(Received 17 May 1978)
Abstract--Optimal conditions were established for the isolation and storage of microsomal NADPH- cytochrome c reductase from midgut tissues of southern armyworm (Spodoptera eridania) larvae. Compared with KCI (l.15%)-nicotinamide (0.2%), the use of an isolation medium consisting of sucros e (0.25 M), phenylmethane sulphonyl fluoride (0.5 mM), polyvinylpyrrolidone (1%) and EDTA (1 mM) increased by 1.7-fold the yield of reductase activity in the microsomal fraction. Washing microsomes isolated in the sucrose medium with sucrose (0.3 M)-pyrophosphate (0.1 M) removed 30% of the protein and caused a similar increase in the specific activity of the reductase. Characteristics of the m idgut microsomal N A DPH- cytochrome c reductase were similar to those of the rat liver microsomal enzyme except that the former was relatively insensitive to changes in ionic strength.
Key Word Index: Insect, southern armyworm, Spodoptera eridania, microsomes, NADPH-cytochrome c reductase, mixed-function oxidase, microsomal electron transport
INTRODUCTION
NADPH-CYTOCHROME c reductase is the major flavoprotein component of the microsomal electron transport chain where it serves to transfer reducing equivalents from NADPH to cytochrome P-450 (MANNERING, 1971; Lu et al., 1976). As such it plays an important physiological role in the cytochrome P-450- mediated metabolism of a large number of drugs, insecticides and other lipophilic foreign compounds by mammals, insects and other species (WILKINSON and BRATTSTEN, 1972). Although cytochrome P-450 is undoubtedly the functional electron acceptor for the flavoprotein, the enzyme was first recognized through its ability to reduce cytochrome c and subsequently this has proved a convenient electron acceptor for routine assay of enzyme activity (KAMIN et al., 1965; IYANAGI and MASON, 1975). Other acceptors to which the enzyme catalyzes electron transfer include various dyes (neotetrazolium and dichlorophenol indophenol) and quinones.
Since its initial isolation by HORECKER (1950) NADPH-cytochrome c reductase from several mammalian sources has been extensively studied and it has been solubilized, purified and well characterized from hepatic microsomes of a number of species. In contrast, information on the enzyme from insect tissue is more sparse. LANG (1959) first reported NADPH- cytochrome c reductase activity in mosquito mitochondrial supernatants and subsequently it has been measured in crude homogenates and microsomes isolated from whole insects or specific tissues of several species (WILKINSON and BRATTSTEN, 1972). Limited studies with solubilized, partially purified enzyme preparations from the house fly (Musca domestica, L.) have been carried out by WILSON and HODGSON (1971a,b) and by CAPDEVILLA et al. (1975) and a more detailed characterization of the purified lipase
43
solubilized enzyme from this species has recently been reported (MAYER and PROUGH, 1977).
This paper describes some of the characteristics of the NADPH-cytochrome c reductase in the microsomal fraction obtained from midgut homogenates of larvae of the southern armyworm. The study was undertaken to provide important background information necessary for further solubilization and purification studies on the flavoprotein.
MATERIALS AND METHODS
Insects
Larvae of the southern armyworm (Spodoptera eridania) were fed on bean plants under greenhouse conditions through the first five instars as previously described (KRIEGER and WILKINSON, 1969). The sixth instar larvae used in these experiments were kept in an environmental chamber maintained at a 16:8 photoperiod, 50-60~,, relative humidity, and at a temperature of 26 C.
Chemicals
Pentamethylbenzene was purchased f rom Aldrich Chemical Co., Inc., Milwaukee, Wl, U.S.A. and p- dimethylaminobenzaldehyde f rom Eastman Kodak, Rochester, NY, U.S.A. Bovine serum albumin was from Pentex Biochemicals, Kankakee, IL, U.S.A. and sodium phenobarbital from Mallinkrodt, St. Louis, MO, U.S.A. Glucose-6-phosphate (G-6-P), G-6-P dehydrogenase, p- chloro-N-methylaniline, p-chloroaniline, and butylated hydroxytoluene (BHT) were purchased from Calbiochem, La Jolla, CA, U.S.A., and NADP, NADPH, NADH, polyvinyl- pyrrolidone (PVP-10), 2,6-dichlorophenolindophenol (DCIP), phenylmethane sulphonyl fluoride (PMSF) and cytochrome c, type VI, were.from Sigma Chemical Co., St. Louis, MO, U.S.A. All other solvents and chemicals employed were of analytical reagent grade.
44 D . L . CRANKSHAW, H. K. HI!TNARSKI AND C. F. WILKINSON
Enzyme preparations Armyworm midgut microsomes were prepared from the
midgut tissues of sixth-instar larvae, fed for the first 24 hr of the instar on kidney bean leaves and subsequently for about 48 hr on a semi-defined artificial diet containing 0.1'~i, pentamethylbenzene (BRATTSTEN and WILKINSON, 1973). Following excision of the guts from the larvae the gut contents were removed and the midgut tissues rinsed in ice- cold sucrose (0.25 M) containing EDTA (1 mM), polyvinylpyrrolidone (PVP, l°~,) and phenylmethane sulphonyl fluoride (PMSF, 2 mM). The gut tissue was homogenized (1:6, w/v) with a ground glass Potter-Elvehjem homogenizer and Teflon pestle in a similar medium adjusted to a PMSF concentration of 0.5 mM. The homogenate was centrifuged at 10,000g for 10 min (2-4°C) in an International Equipment Co. (IEC) B-20A centrifuge and the supernatant was filtered through glass wool. The pellet was rehomogenized in 809o of the original volume of fresh medium, recentrifuged and filtered as before. Microsomes were obtained by centrifuging the combined supernatants for 90 min at 105,000 g (0.4°C) in an IEC B-60 ultracentrifuge with an IEC A-211 fixed-angle rotor. The microsomal pellet was resuspended to approximately 25 mg/ml in 0.05 M Tris-HCI, pH 7.5 (0-4 C), containing 50", glycerol and 50"o glycerol and stored under nitrogen at - 1 5 C until required. In some of the initial studies microsomes were prepared in ice-cold 1.153,o KC1 containing 0.2°~o nicotinamide by a similar procedure.
Perfused livers of male Sprague-Dawley rats (Blue Spruce Farms, Altamont. NY, U.S.A.) were homogenized in ice- cold 1.15~o KCI (1:4, w/v) containing nicotinamide (0.2~o) using a Potter-Elvehjem homogenizer with a Teflon pestle. Phenobarbital induction was accomplished by feeding the rats water containing 0.17:o phenobarbital for ten days. Microsomes were sedimented from the postmitochondrial supernatant (10,000 g for 20 rain) by centrifugation at 105,000 g for 90 rain in an IEC B-60 preparative ultrancentrifuge equipped with an IEC A-321 fixed-angle rotor. The microsomal pellet was resuspended in 0.05 M Tris-HC1, pH 7.5 (0-4 C), containing 50"i, glycerol and butylated hydroxytoluene (0.01°o w/v) at a protein concentration of approximately 50 mg/ml and stored at
- 15"C under nitrogen until required. Protein concentration was determined by the method of
LowRY et al. (1951) using bovine serum albumin as a standard.
Enzyme assays Cytochrome c reductase activity was assayed directly at
25°C in 1 cm cuvettes in an Amino-Chance DW-2 spectrophotometer (PEDERSON et al., 1973). The 1 ml incubation mixture contained 25-50 /zg of microsomal protein, cytochrome c (75/iM) and 0.3 M phosphate buffer (pH 7.5 for rat liver and pH 7.8 for armyworm gut). The reactions were initiated by addition of either NADPH or NADH (0.1 mM for rat liver, 0.2 mM for armyworm gut) to the sample cuvette and the linear increase in absorbance at 550 nm was measured. A similar procedure employing 50 mM phosphate buffer was used to measure the enzymatic reduction of potassium ferricyanide (210 tiM) and 2,6- dichlorophenolindiphenol (DCIP) (95/IM) at 420 and 600 nm, respectively. In each case, reductase activity was calculated from the initial change in absorbance at the appropriate wavelength using the millimolar extinction coefficients of 21.1 for cytochrome c (MASSEY, 1959), 1.02 for ferricyanide (SCHELLENBERG and HELLERMAN, 1958) and 21.0 for DCIP (STEYN-PARVE and BEINERT, 1958). One unit of enzyme activity is that which reduces 1 /Jmole of electron acceptor/min.
Microsomal N-demethylation ofp-chloro N-rnethylaniline was assayed as described by BRATTSTEN and WILKINSON (1973).
.c
3
6 0 - -
40
2° l 0
24 48 72 84
Age in sixth instar, hr
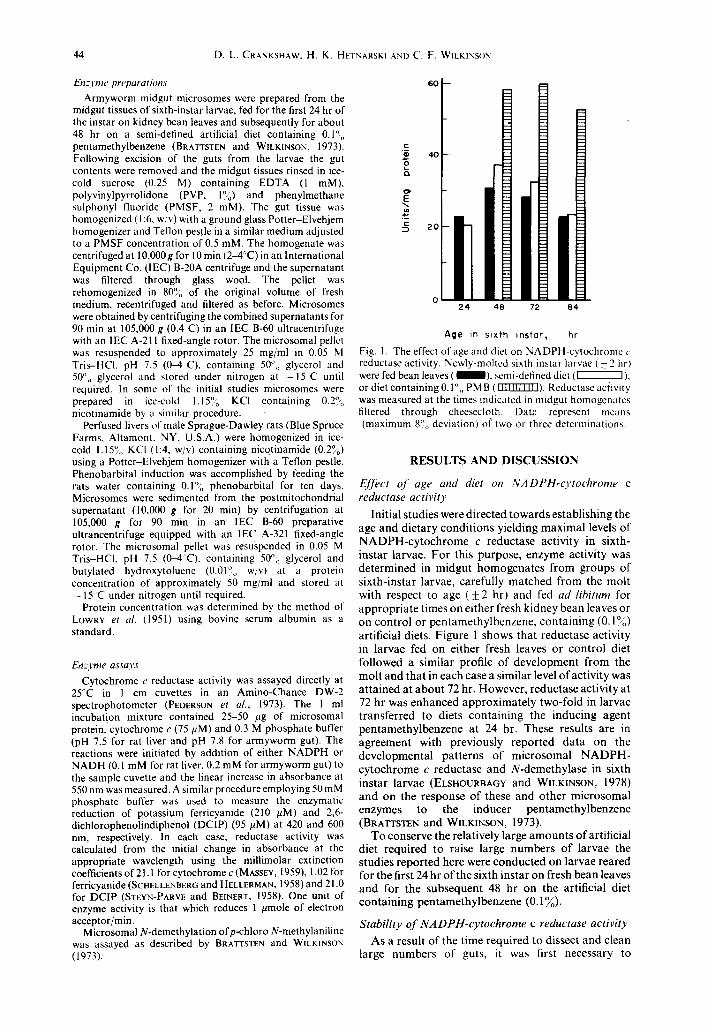
Fig. 1. The effect of age and diet on NADPH-cytochrome c reductase activity. Newly-molted sixth instar larvae ( + 2 hrl were fed bean leaves ( 1 ) , semi-defined diet ( ~ ) , or diet containing 0.1",, PM B (IIUIITZI~). Reductase aciivity was measured at the times indicated in midgut homogenates filtered through cheesecloth. Datt: represent means (maximum 8°0 deviation) o( two or three determinations.
R E S U L T S AND D I S C U S S I O N
EJfect o f age and diet on NADPH-cytochrome c reductase activity
Initial studies were directed towards establ ishing the age and dietary condi t ions yielding maximal levels of N A D P H - c y t o c h r o m e c reductase activity in sixth- instar larvae. Fo r this purpose, enzyme activity was de termined in midgut homogena te s f rom groups of s ixth-instar larvae, carefully matched from the mol t with respect to age ( + 2 hr) and fed ad libitum for appropr ia te times on either fresh kidney bean leaves or on control or pentamethylbenzene , con ta in ing (0.1°4~) artificial diets. Figure 1 shows tha t reductase activity in larvae fed on either fresh leaves or cont ro l diet followed a similar profile of deve lopment f rom the mol t and tha t in each case a similar level of activity was a t ta ined at a b o u t 72 hr. However, reductase activity at 72 hr was enhanced approximate ly two-fold in larvae t ransferred to diets con ta in ing the inducing agent pen tamethy lbenzene at 24 hr. These results are in agreement with previously reported da ta on the developmenta l pa t te rns of microsomal N A D P H - cy tochrome c reductase and N-demethylase in sixth instar larvae (ELSHOURBAGY and WILKINSON, 1978) and on the response of these and o ther microsomal enzymes to the inducer pen tamethy lbenzene (BRATTSTEN and WmKINSON, 1973).
To conserve the relatively large amoun t s of artificial diet required to raise large number s of larvae the studies repor ted here were conducted on larvae reared for the first 24 hr of the sixth instar on fresh bean leaves and for the subsequent 48 hr on the artificial diet conta in ing pen tamethy lbenzene (0.1~o).
Stability o f NADPH-cytochrome c reductase activity
As a result of the t ime required to dissect and clean large numbers of guts, it was first necessary to
NADPH-Cytochrome c reductase 45
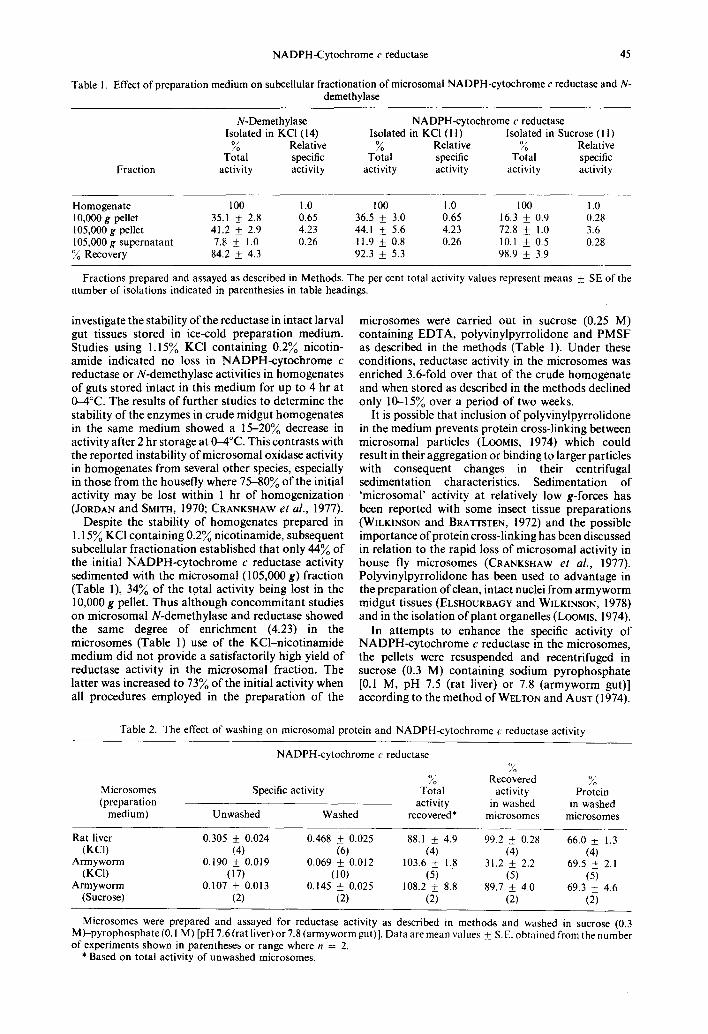
Table 1. Effect of preparation medium on subcellular fractionation of microsomal NADPH-cytochrome c reductase and N- demethylase
Fraction
N-Demethylase NADPH-cytochrome c reductase Isolated in KCI (14) Isolated in KCI (1 I) Isolated in Sucrose (I I)
~o Relative ~o Relative ~o Relative Total specific Total specific Total specific
activity activity activity activity activity activity
Homogenate 100 1.0 100 1.0 100 1.0 10,000g pellet 35.1 + 2.8 0.65 36.5 _ 3.0 0.65 16.3 _ 0.9 0.28 105,000g pellet 41.2 + 2.9 4.23 44.1 + 5.6 4.23 72.8 + 1.0 3.6 105,000g supernatant 7.8 + 1.0 0.26 11.9 + 0.8 0.26 10.1 + 0.5 0.28 O//o Recovery 84.2 _+ 4.3 92.3 _+ 5.3 98.9 _+ 3.9
Fractions prepared and assayed as described in Methods. The per cent total activity values represent means _+_ SE of the number of isolations indicated in parenthesies in table headings.
investigate the stability of the reductase in intact larval gut tissues stored in ice-cold preparation medium. Studies using 1.159/o KCI containing 0.29'0 nicotin- amide indicated no loss in NADPH-cytochrome c reductase or N-demethylase activities in homogenates of guts stored intact in this medium for up to 4 hr at 0-4°C. The results of further studies to determine the stability of the enzymes in crude midgut homogenates in the same medium showed a 15-20% decrease in activity after 2 hr storage at 0-4°C. This contrasts with the reported instability of microsomal oxidase activity in homogenates from several other species, especially in those from the housefly where 75-809/0 of the initial activity may be lost within 1 hr of homogenization (JORDAN and SMITH, 1970; CRANKSHAW et al., 1977).
Despite the stability of homogenates prepared in 1.15~ KC1 containing 0.29/o nicotinamide, subsequent subcellular fractionation established that only 449/0 of the initial NADPH-cytochrome c reductase activity sedimented with the microsomal (105,000 g) fraction (Table 1), 34~ of the totat activity being lost in the 10,000 g pellet. Thus although concommitant studies on microsomal N-demethylase and reductase showed the same degree of enrichment (4.23) in the microsomes (Table 1) use of the KCl-nicotinamide medium did not provide a satisfactorily high yield of reductase activity in the microsomal fraction. The latter was increased to 739/0 of the initial activity when all procedures employed in the preparation of the
microsomes were carried out in sucrose (0.25 M) containing EDTA, polyvinylpyrrolidone and PMSF as described in the methods (Table 1). Under these conditions, reductase activity in the microsomes was enriched 3.6-fold over that of the crude homogenate and when stored as described in the methods declined only 10-159/o over a period of two weeks.
It is possible that inclusion of polyvinylpyrrolidone in the medium prevents protein cross-linking between microsomal particles (LOOMIS, 1974) which could result in their aggregation or binding to larger particles with consequent changes in their centrifugal sedimentation characteristics. Sedimentation of 'microsomal' activity at relatively low g-forces has been reported with some insect tissue preparations (WILKINSON and BRATTSTEN, 1972) and the possible importance of protein cross-linking has been discussed in relation to the rapid loss of microsomal activity in house fly microsomes (CRANKSHAW e l al., 1977). Polyvinylpyrrolidone has been used to advantage in the preparation of clean, intact nuclei from armyworm midgut tissues (ELsHOURBAGY and WILKINSON, 1978) and in the isolation of plant organelles (LOOMIS, 1974).
In attempts to enhance the specific activity of NADPH-cytochrome c reductase in the microsomes, the pellets were resuspended and recentrifuged in sucrose (0.3 M) containing sodium pyrophosphate [0.1 M, pH 7.5 (rat liver) or 7.8 (armyworm gut)] according to the method of WELTON and AusT (1974).
Table 2. The effect of washing on microsomal protein and NADPH-cytochrome c reductase activity
NADPH-cytochrome c reductase %
% Recovered % Microsomes Specific activity Total activity Protein (preparation activity in washed in washed
medium) Unwashed Washed recovered* microsomes microsomes
Rat liver 0.305 ___ 0.024 0.468 __+ 0.025 88.1 _+ 4.9 99.2 + 0.28 66.0 + 1.3 (KCI) (4) (6) (4) (4) (4)
Armyworm 0.190 _ 0.019 0.069 + 0.012 103.6 _+ 1.8 31.2 _ 2.2 69.5 + 2.1 (KC1) (17) (10) (5) (5) (5)
Armyworm 0.107 _+ 0.013 0.145 + 0.025 108.2 _+ 8.8 89.7 _+ 4.0 69.3 _ 4.6 (Sucrose) (2) (2) (2) (2) (2)
Microsomes were prepared and assayed for reductase activity as described in method~ and washed in sucrose (0.3 M)-pyrophosphate (0. I M) [pH 7.6 (rat liver) or 7.8 (armyworm gut)]. Data are mean values _+_ S.E. obtained from the number of experiments shown in parentheses or range where n = 2.
* Based on total activity of unwashed microsomes.
46 D.L. CRANKSHAW. H. K. HETNARSKI AND C. F. WILKINSON
i S - -
x 12
6
0 0 . 2 0 . 4 0 . 6 0 8
Protein, mg/m[
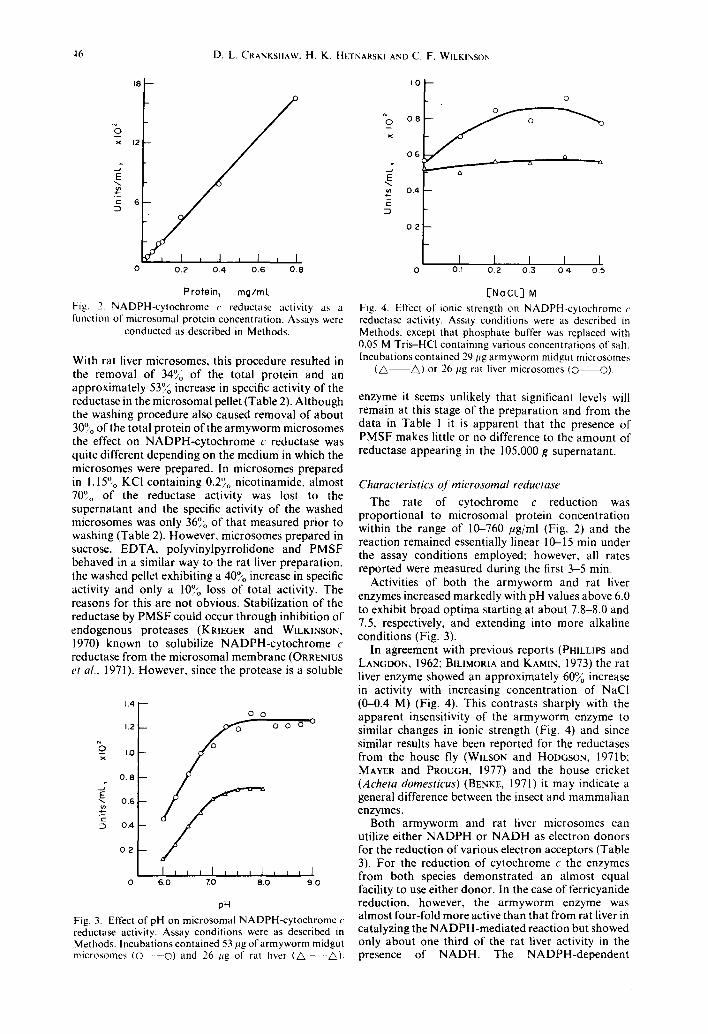
Fig. 2. NADPH-cytochrome r reductase activity as a runction of microsomal protein concentration. Assays were
conducted as described in Methods.
With rat liver microsomes, this procedure resulted in the removal of 34}0 of the total protein and an approximately 53~o increase in specific activity of the reductase in the microsomal pellet (Table 2). Although the washing procedure also caused removal of about 30°,0 of the total protein of the armyworm microsomes the effect on NADPH-cytochrome c reductase was quite different depending on the medium in which the microsomes were prepared. In microsomes prepared in 1.157o KCI containing 0.2'~o nicotinamide, almost 70°0 of the reductase activity was lost to the supernatant and the specific activity of the washed microsomes was only 369o of that measured prior to washing (Table 2). However, microsomes prepared in sucrose, EDTA, polyvinylpyrrolidone and PMSF behaved in a similar way to the rat liver preparation, the washed pellet exhibiting a 40}0 increase in specific activity and only a 10°~, loss of total activity. The reasons for this are not obvious. Stabilization of the reductase by PMSF could occur through inhibition of endogenous proteases (KRIEGER and WILKINSON, 1970) known to solubilize NADPH-cytochrome c reductase from the microsomal membrane (ORRENIUS et al., 1971). However, since the protease is a soluble
x
._J
~n
1.4 i
1 . 2 ' - -
1,0 --
0 . 8
0 . 6
0 .4
0 . 2
0 0
0 0 0 0 0
I , J ~ I , , , l i i t I 6.0 7 0 8 0 9 0
pH Fig. 3. Effect of pH on microsomal NADPH-cytochrome c reductase activity. Assay conditions were as described in Methods. Incubations contained 53 pg of armyworm midgut microsomes ( 0 - - 0 ) and 26 l~g of rat liver (A ~).
1.0
~ O 0 . 8
0 . 6 p
E N tn 0 . 4
0 . 2
m
o
1 1 I I I 0 O.i 02 03 0.4 0.5
[Ne CL] M Fig. 4. Effect of ionic strength on NADPH-cytochrome r reductase activity. Assay conditions were as described in Methods, except that phosphate buffer was replaced with 0.05 M Tris-HCl containing various concentrations of salt. Incubations contained 29 ltg armyworm midgut microsomes
(A A) or 26 l~g rat liver microsomes (O O).
enzyme it seems unlikely that significant levels will remain at this stage of the preparation and from the data in Table 1 it is apparent that the presence of PMSF makes little or no difference to the amount of reductase appearing in the 105,000 g supernatant.
Characteristics oJ microsomal reductase
The rate of cytochrome c reduction was proportional to microsomal protein concentration within the range of 10-760 #g/ml (Fig. 2) and the reaction remained essentially linear 10-15 min under the assay conditions employed; however, all rates reported were measured during the first 3-5 min.
Activities of both the armyworm and rat liver enzymes increased markedly with pH values above 6.0 to exhibit broad optima starting at about 7.8-8.0 and 7.5, respectively, and extending into more alkaline conditions (Fig. 3).
In agreement with previous reports (PHILLIPS and LANGDON, 1962; BILIMORIA and KAMIN, 1973) the rat liver enzyme showed an approximately 60~o increase in activity with increasing concentration of NaCl (0-0.4 M) (Fig. 4). This contrasts sharply with the apparent insensitivity of the armyworm enzyme to similar changes in ionic strength (Fig. 4) and since similar results have been reported for the reductases from the house fly (WILSON and HODGSON, 1971b: MAYER and PROUGH, 1977) and the house cricket (Acheta domesticus) (BENKE, 1971) it may indicate a general difference between the insect and mammalian enzymes.
Both armyworm and rat liver microsomes can utilize either NADPH or NADH as electron donors for the reduction of various electron acceptors (Table 3). For the reduction of cytochrome c the enzymes from both species demonstrated an almost equal facility to use either donor. In the case of ferricyanide reduction, however, the armyworm enzyme was almost four-fold more active than that from rat liver in catalyzing the NADPH-mediated reaction but showed only about one third of the rat liver activity in the presence of NADH. The NADPH-dependent

NADPH-Cytochrome e reductase 47
Table 3. Microsomal NADPH-cytochrome e reductase activities
Specific activity (units, mg protein)
Electron Electron Armyworm donor acceptor midgut Rat liver
NADPH Cytochrome c 0.17 0.14 (0.15--0.19) (0.138-0.140)
NADH Cytochrome c 0.19 0.28 (0.18-0.19) (0.23-0.32)
NADPH Ferricyanide 0.41 0.12 (0.40-0.41) (0.10-0.14)
NADH Ferricyanide 0.79 2.06 (0.77-0.82) (1.97-2.15)
NADPH DCIP 0.11 0.05 (0.10-0.12) (0.04-0.06)
Microsomes from armyworm midgut and rat liver prepared and assayed as described in methods. Results are the mean values of assays with ranges conducted with two different microsomal preparations.
reduct ion of D C I P by a r m y w o r m midgut microsomes was two-fold greater than tha t catalyzed by rat liver.
L ineweaver -Burk double reciprocal plots were l inear with bo th N A D P H and cy tochrome c over concen t ra t ion ranges of 13--103 and 12-71 /~M, respectively. Appa ren t Km values were 30.0 /~M for N A D P H and 35 /aM for cy tochrome c which are similar to those repor ted for house fly microsomal enzyme (WILSON and HODGSON, 1971b; MAYER and PROUGH, 1977) and for the rat liver microsomal enzyme (ERNSTER and ORRENIUS, 1965).
Studies to solubilize, purify and fur ther character ize the N A D P H - c y t o c h r o m e c reductase in a rmyworm midgut microsomes are current ly in progress.
Acknowledgement--The work was supported in part by grants (ES 00400 and ES 00098) from the U.S. Public Health Service.
R E F E R E N C E S
BENKE G. M. (1971) Microsomal oxidases in selected orthopteran insects, with special reference to the house cricket, Acheta domestieus and the Madagascar roach, GromphadorhhTa portentosa. M. S. thesis, Cornell University, Ithaca, New York.
BILMORIA M. H. and KAMIN H. (1973) The effect of high salt concentrations upon cytochrome c. cytochrome b s. and iron-EDTA reductase activities of liver microsomal NADPH-cytochrome e reductase. Ann. N.Y. Acad. ScL 212, 428-448.
BRATTSTEN L. B. and WILKINSON C. F. (1973) Induction of microsomal enzymes in the southern armyworm (Prodenia eridania). Pestic biochem. Physiol. 3, 393-407.
CAPDEVILLA J., AHMAD N. and AGOSIN M. (1975) Soluble cytochrome P-450 from housefly microsomes: partial purification and characterization of two hemoprotein forms. J. biol. Chem. 250, 1048--1060.
CRANKSHAW D. L., ZABIK M. and AUST S. D. (1977) The role of tyrosinase in the inactivation of house fly microsomal mixed-function oxidases. Pestic. Biochem. Physiol. 7, 564-572.
ELSHOURBAGY N. and WILKINSON C. F. (1978) The role of DNA-dependent RNA polymerases in microsomal enzyme induction in southern armyworm (Spodoptera eridania) larvae. Insect Biochem. 8, 425-433.
ERNSTER L. and ORRENICS S. (1965) Substrate-induced
synthesis of the hydroxylating enzyme system of liver microsomes. Fedn. Proe. Fedn. Am. Sotw exp. Biol. 24, 1190-1199.
HORECKER B. U (1950) Triphosphopyridine nucleotide- cytochrome c reductase in liver. J. biol. Chem. 183, 593-605.
IYANAGI T. and MASON H. S. (1975) Hepatic NADPH- cytochrome e oxidoreductase. In Reactivity ofFlavins (Ed. by YAGI K.) pp. 145-156. University of Tokyo Press, Tokyo.
JOROAN T. W. and SMITH J. N. (1970) Oxidation inhibitors in homogenates of houseflies and blowflies. Int. J. Biochem. I, 139-149.
KAMIN H., MASTERS B. S., GIBSON Q. H. and WILLIAMS C. H., Jr. (1965) Microsomal TPNH-cytochrome e reductase. Fe~bL Proc. Fe~hl. Am. Sots e.vp. Bhd. 24, 1164-1171.
KRIEGER R. I. and WILKINSON C. F. (1969) Microsomal mixed-function oxidases in insects--I. Localization and properties of an enzyme system effecting aldrin epoxidation in larvae of the southern armyworm (Prodenia eridania). Biochem. Pharmaeol. 18, 1403-1415.
KRIEGER R. I. and WILKINSON C. F. (1970) An endogenous inhibitor of microsomal mixed-function oxidases in homogenates of the southern armyworm (Prodenia eridania). Biochem. J. 116, 781-789.
LANG C. A. { 1959) Cytochrome c reductase activities during dcvclopmcnt. EvpL Cell Res. 17, 516-518.
L(×)MIS W. D. (1974) Overcoming problems of phenolics and quinones in the isolation of plant enzymes and organelles. Methods Enzymol. 31,528-544.
LOWRY O. H., ROSEBROUGH N. J., FARR A. L. and RANDALL R. J. (1951) Protein measurement with the Folin phenol reagent. J. biol. Chem. 193, 265-275.
Lu A. Y. H., KUNTZMAN R. and CONNEY A. H. (1976) The liver microsomal hydroxylation enzyme system, k)'ont. gastrointest. Res. 2, 1-31.
MANNERINO G. J. (1971) Microsomal enzyme systems which catalyze drug metabolism. In Fundamentals o] Drug Metabolism and Drug Di.sposition (Ed. by LADU B. N., MANDEL H. G. and WAY E. L.) pp. 206-252. Williams & Wilkins, Baltimore.
MASSEY V. (1959) The microestimation of succinate and the extinction coefficient of cytochrome c. Biochim. bioph3's. Acta 34, 255-256.
MAYER R. T. and PROUGH R. A. (1977) Purification and characterization of NADPH-cytochrome c (P4s0) reductase from the house fly, Musea domestica. Comp. biochem. Physiol. 57B, 81-87.
ORRENIUS S,, BERGGREN M., MOLDEUS P. and KRIEGER R. I. (1971) Mechanism of inhibition of microsomal mixed- function oxidation by the gut-contents inhibitor of the southern armyworm (Prodenia eridania). Biochem J. 124, 427-430.
PEDERSON T. C., BUEGE J. A. and ACST S. D. (1973) Microsomal electron transport: the role of reduced nicotinamide adenine dinucleotide phosphate-cytochrome c reductase in liver microsomal lipid peroxidation. J. biol. Chem. 248, ? 134-7141.
PHILIPS A. H. and LANGDON R. G. (1962) Hepatic triphosphopyridine nucleotide-cytochrome c reductase: isolation, characterization, and kinetic studies. J. biol, Chem. 237, 2652-2660.
SCHELLENBERG K. A. and HELLERMAN L. (1958) Oxidation of reduced pyridine nucleotide. J. biol. Chem. 231, 54%556.
STEVN-PARvE E. P. and BEINERT H. (1958) On the mechanism of dehydrogenation of fatty acyl derivatives of coenzyme A-VI. Isolation and properties of stable enzyme-substrate complexes. J. biol. Chem. 233, 843-852.
WELTON A. F. and AUST S. D. (1972) Lipid peroxidation during enzymatic iodination of rat liver endoplasmic reticulum. Bioehem.. biophfs. Res. Commun. 49, 661-666.
WILKINSON C. F. and BRATTSTEN L. B. (1972) Microsomal drug metabolizing enzymes in insects. Drug Metab. Rev. 1, 153-228.

48 D .L . CRANKSHAW, H. K. HETNARSKI AND C. F. WILKINSON
WILSON T. G. and HODGSON E. (1971a) Microsomal NADPH-cytochrome c reductase from the housefly, Musca domestica: solubilization and purification, bisect Biochem. I, 19---26.
WILSON T. G. and HODGSON E. (1971b) Microsomal NA DPH-cytochrome c red uctase from the housefly. Musca domestica: properties of the purified enzyme. Insect Biochem. 1, 171-180.