Embed Size (px)
Citation preview

NAADP Receptors
Antony Galione
Department of Pharmacology, University of Oxford, Mansfield Road, Oxford OX1 3QT,United Kingdom
Correspondence: [email protected]
Of the established Ca2þmobilizing messengers, NAADP is arguably the most tantalizing. Itis the most potent, often efficacious at low nanomolar concentrations. Recent studies haveidentified a new class of calcium release channel, the two-pore channels (TPCs), as thelikely targets for NAADP. These channels are endolysosomal in localization where theymediate local Ca2þ release, and have highlighted a new role of acidic organelles as targetsfor messenger-evoked Ca2þmobilization. Three distinct roles of TPCs have been identified.The first is to effect local Ca2þ release that may play a role in endolysosomal function in-cluding vesicular fusion and trafficking. The second is to trigger global calcium releaseby recruiting Ca2þ-induced Ca2þ release (CICR) channels at lysosomal-ER junctions. Thethird is to regulate plasma membrane excitability by the targeting of Ca2þ release from ap-propriately positioned subplasma membrane stores to regulate plasma membrane Ca2þ-activated channels. In this review, I discuss the role of NAADP-mediated Ca2þ releasefrom endolysosomal stores as a widespread trigger for intracellular calcium signaling mech-anisms, and how studies of TPCs are beginning to enhance our understanding of the centralrole of lysosomes in Ca2þ signaling.
Calcium is the most evolutionarily ubiqui-tous of intracellular signals and controls
cellular mechanisms as diverse as cellular mo-tility, membrane fusion, ion channel function,enzyme activity, and gene expression (Berridgeet al. 2003). Free cytoplasmic calcium levelsare kept under tight control by pumps, ex-changers, and buffering mechanisms includ-ing storage by organelles (Pozzan et al. 1994).Ca2þ signals may be elicited when these mech-anisms are transiently overwhelmed by theopening of calcium permeable channels at theplasma membrane or in membranes of cal-cium-storing organelles. Chronic activation of
such channels may lead to cell death, for exam-ple, through the activation of apoptotic signal-ing cascades (Berridge et al. 1998). Many cellsurface receptors are linked to signaling path-ways that lead to the mobilization of calciumfrom intracellular storage organelles throughthe activation of specific Ca2þ release channels(Clapham 1995). Three major intracellularmessengers have been established to link cell stim-ulation with organellar Ca2þ release: inositol tri-sphosphate (IP3), cyclic adenosine diphosphateribose (cADPR), and nicotinic acid adeninenucleotide diphosphate (NAADP) (Bootmanet al. 2002).
Editors: Martin D. Bootman, Michael J. Berridge, James W. Putney, and H. Llewelyn Roderick
Additional Perspectives on Calcium Signaling available at www.cshperspectives.org
Copyright # 2011 Cold Spring Harbor Laboratory Press; all rights reserved; doi: 10.1101/cshperspect.a004036
Cite this article as Cold Spring Harb Perspect Biol 2011;3:a004036
1
on September 3, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

DISCOVERY OF NAADP AS ACa2þ-MOBILIZING MOLECULE
NAADP was discovered as a contaminant ofcommercial batches of b-NADPþ by Lee andcolleagues while they were investigating the ef-fects of various pyridine nucleotides on calci-um release from sea urchin egg homogenates(Clapper et al. 1987). The rationale for this wasthat at fertilization in sea urchin eggs, dramaticchanges in pyridine nucleotide levels occur at asimilar time to the generation of the calciumwave. Egg homogenates can be simply preparedfrom eggs and are remarkably stable, even afterfreezing, and sequester, and robustly release cal-cium when challenged with messengers anddrugs (Morgan and Galione 2008). Three dis-tinct calcium release mechanisms were shown.These were the early days of IP3, and IP3 wasfound to release calcium from microsomalstores. In addition, two metabolites of pyridinenucleotides, an enzyme-activated metaboliterelated to NADþ, subsequently identified ascADPR (Lee et al. 1989), and alkaline-treated
NADP, later shown to be NAADP (Lee and Aar-hus 1995), were found to release Ca2þ from dif-ferent subcellular nonmitochondrial fractionsfrom egg homogenate (Fig. 1). A key featureof each mechanism is their display of homolo-gous desensitization underscoring the inde-pendence of each of the three mechanisms.
NAADP AS A Ca2þMOBILIZING MESSENGER
NAADP is the most potent of Ca2þ mobilizingmessengers described, typically efficacious atpM or low nM concentrations. A growing num-ber of cellular stimuli and activation of cell sur-face receptors have been found to be coupled toincreases in NAADP levels, confirming its roleas an intracellular messenger (Churchill et al.2003; Masgrau et al. 2003; Rutter 2003; Yama-saki et al. 2005; Galione 2006; Gasser et al.2006; Kim et al. 2008). Mediation of calciumsignaling by NAADP has been implicated bytwo approaches: inhibition of agonist-evokedcalcium signals by prior self-inactivation of the
O
HO
HO
O
Lysosome
NAADP
Ca2+Ca2+
Cytosol
TPC2
OH
N+ O
HO
P P
N
N N
N
NH2
P
Figure 1. Structure and function of NAADP. NAADP differs from b -NADP in that the base nicotinic acid issubstituted for nicotinamide (upper panel). NAADP, unlike NADP, is a potent Ca2þmobilizing agent and inter-acts with two-pore channels in the membranes of lysosomes (lower panel).
A. Galione
2 Cite this article as Cold Spring Harb Perspect Biol 2011;3:a004036
on September 3, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

NAADP receptor or NAADP receptor phar-macological blocker (Naylor et al. 2009) andmeasurements of cellular NAADP levels. Mea-surements of NAADP have been performedusing either a radioreceptor assay, based on thehigh affinity NAADP binding protein of seaurchin eggs (Lewis et al. 2007), or by using acycling assay of coupled enzyme reactionsresulting in fluorescent resorufin production(Graeff and Lee 2002). Although some recep-tors appear to couple selectively to NAADP pro-duction, increasingly it is becoming apparentthat several receptors couple to multiple Ca2þ
mobilizing messengers (Aley et al. 2010), andthis may be the norm.
Ca2þ STORES TARGETED BY NAADP
Accumulating evidence suggests that the pri-mary Ca2þ stores targeted by NAADP are sepa-rate from the endoplasmic reticulum and aremembers of a group known as acidic organelles.The initial evidence for this came from thestudy of sea urchin eggs and was subsequentlyextended to mammalian cells.
Sea Urchin Eggs
The initial report of NAADP-evoked Ca2þ
release using alkaline-activated NADP showedthat the subcellular fraction reactive to NAADPin egg homogenates was largely separate fromthe microsomal/ER fraction sensitive to IP3
and cADPR (Clapper et al. 1987). Abrogationof Ca2þ storage by the ER SERCA inhibi-tor, thapsigargin, while inhibiting Ca2þ releaseby either IP3 or cADPR, only partially re-duced Ca2þ release evoked by NAADP in bothsea urchin egg homogenates (Genazzani andGalione 1996) and intact eggs (Churchill andGalione 2001a). Visualization of two separateCa2þ stores was observed in elegant sea urchinegg stratification studies (Lee and Aarhus 2000).Stratification by centrifugation of intact eggsresults in eggs forming elongated structureswith different organelles separating to different“poles.” Uniform photolysis of caged derivativesof Ca2þ mobilizing messengers resulted in IP3
and cADPR releasing Ca2þ from the nuclear
pole where ER accumulated, whereas NAADPreleased Ca2þ from the opposite end of the struc-ture. These experiments are consistent with theprimary Ca2þ store targeted by NAADPas beingdistinct from the ER.
In a series of important experiments usingpharmacological analyses and subcellular frac-tionation, lysosomal-related organelles wereimplicated as the primary target organelle forNAADP-evoked Ca2þ release in sea urchineggs (Churchill et al. 2002). Acidic stores, suchas lysosomes, have been shown to sequesterCa2þ by mechanisms dependent on their lowluminal pH (Patel and Docampo 2010). Inhib-ition of the vacuolar Hþ-ATPase by bafilomycindecreases proton uptake into acidic stores; iftheir membranes are sufficiently leaky to pro-tons, this leads to the alkalinization of their lu-men. Uptake of Ca2þ into these stores appearsto be dependent on the maintenance of the pro-ton gradient because bafilomycin and protono-phores inhibit Ca2þ storage by these organelles,although the detailed mechanisms are not wellunderstood. A dense membrane fraction fromsea urchin egg homogenates was isolated froma percoll gradient and consisted of “reserve gra-nules.” This fraction was enriched with lyso-somal markers and supported ATP-dependentCa2þ sequestration, which was inhibited bypreincubation with bafilomycin or the proto-nophore, nigericin, but not thapsigargin. Thisfraction was found to contain [32P]NAADPbinding sites, and displayed NAADP but notIP3/cADPR-evoked Ca2þ release. Reserve gran-ules from sea urchin eggs are lysosome-relatedorganelles. In intact sea urchin eggs, treatmentwith the lysosomotropic agent, glycyl-phenyl-alanine 2-naphthylamide (GPN), caused thereversible lysis of lysotracker-stained vesicles, re-sulting in a series of small-amplitude cytoplas-mic Ca2þ signals, consistent with their role asCa2þ stores. Importantly, GPN treatment in ei-ther intact eggs or egg homogenates selectivelyabolished NAADP-evoked Ca2þ release with lit-tle effect on Ca2þ release byeither IP3 or cADPR.From these data it was proposed that in seaurchin the primary target of NAADP are acidicstores rather than the endoplasmic reticulum.Consistent with this, experiments in sea urchin
NAADP Receptors
Cite this article as Cold Spring Harb Perspect Biol 2011;3:a004036 3
on September 3, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

egg homogenates employing luminal pH in-dicators such as acridine orange or lysosensoralso have shown that NAADP uniquely amongCa2þ mobilizing messengers also causes thealkalinization of store lumena, representing an-other possible signaling mechanism for thismolecule (Morgan and Galione 2007b).
Mammalian Cells
Following these studies in sea urchin eggs, itwas shown that NAADP also targeted acidicstores in a wide range of mammalian cells andin response to a variety of cellular stimuli(Mitchell et al. 2003; Kinnear et al. 2004; Yama-saki et al. 2004; Galione 2006; Gerasimenkoet al. 2006; Menteyne et al. 2006; Zhang et al.2006; Macgregor et al. 2007; Gambara et al.2008; Jardin et al. 2008; Kim et al. 2008;Lloyd-Evans et al. 2008; Brailoiu et al. 2009b;Pandey et al. 2009; Thai et al. 2009; Dickinsonet al. 2010; Zhang et al. 2010).
DESENSITIZATION OF NAADP-EVOKEDCa2þ RELEASE
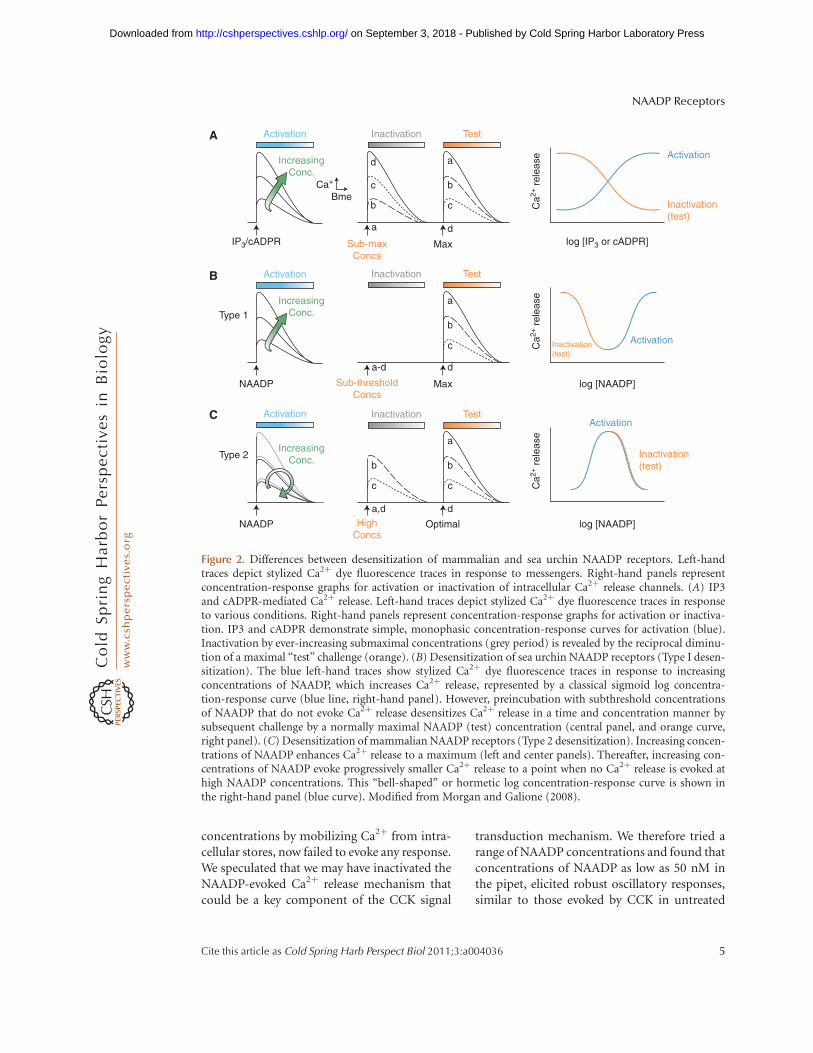
The Ca2þ release mechanism activated byNAADP shows unusual and profound inactiva-tion properties. One major area of confusion inthis field is that the inactivation properties ofCa2þ release varies markedly between sea urchinegg and mammalian systems, which we havetermed type I and type II, respectively (Morganand Galione 2008) (Fig. 2).
Sea urchin eggs: The initial report demon-strating the efficacy of NAADP as a Ca2þmobi-lizing molecule showed that NAADP releasedCa2þ by a mechanism independent of IP3 orryanodine receptors (RyRs), based on each ofthese mechanisms showing homologous desen-sitization (Lee and Aarhus 1995). After NAADPstimulated Ca2þ release in egg homogenates,they became refractory to subsequent challengewith NAADP, but still responded to either IP3
or cADPR. This was the first piece of evidencethat NAADP activated a novel Ca2þ releasechannel, distinct from the principal Ca2þ re-lease channels on the endoplasmic reticulum.
Further analysis of the phenomenon of self-inactivation of NAADP-evoked Ca2þ release in
sea urchin eggs and homogenates revealed sev-eral profound and unusual features. A sur-prising finding was that pM concentrations ofNAADP, although subthreshold for triggeringCa2þ release in egg homogenates, were able toinactivate completely the NAADP Ca2þ releasemechanism to a subsequent challenge by nMconcentration of NAADP that would normallyevoke a maximal Ca2þ release (Aarhus et al.1996; Genazzani et al. 1996). The extent of inac-tivation was dependent on both the concentra-tion and duration of incubation (Genazzaniet al. 1996; Genazzani et al. 1997b). Mecha-nisms of inactivation of the NAADP receptorare not understood, but may be related to theapparent irreversible binding of [32P]NAADP.The radioligand appears to become occludedon binding in a time-dependent manner (Aar-hus et al. 1996). Studies with the selectiveNAADP receptor antagonist, Ned-19, (Nayloret al. 2009) and its analogs (Rosen et al. 2009)have led to the proposal that there are twodistinct binding sites for NAADP. The first ishigh affinity, whose occupancy leads to slowinactivation of the receptor, and a second loweraffinity site that leads to rapid channel opening.Ned-20 blocks inactivation, but not activationof Ca2þ release by NAADP (Rosen et al. 2009).
Mammalian cells: There are key differencesbetween desensitization of the NAADP receptorbetween sea urchin eggs, in which subthresholdconcentrations of NAADP can fully inactivatethe NAADP-sensitive Ca2þ release mechanism;whereas in a mammalian cell, high concentra-tions of NAADP are needed for full inactivation,which can occur in the apparent absence ofreceptor activation. The first report of NAADPaction as a Ca2þ-mobilizing agent in a mam-malian cell was in the pancreatic acinar cell(Cancela et al. 1999), which was also the systemin which IP3 was first shown to mobilize Ca2þ
from nonmitochondrial stores (Streb et al.1983). Using whole cell patch and measuringCa2þ-activated currents, we found that a pi-pet concentration of 10 mM NAADP failed toelicit any responses. However, we noticed thatafter intracellular application of this concen-tration of NAADP, cholecystokinin (CCK),which usually increases cytosolic Ca2þ at pM
A. Galione
4 Cite this article as Cold Spring Harb Perspect Biol 2011;3:a004036
on September 3, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from
concentrations by mobilizing Ca2þ from intra-cellular stores, now failed to evoke any response.We speculated that we may have inactivated theNAADP-evoked Ca2þ release mechanism thatcould be a key component of the CCK signal
transduction mechanism. We therefore tried arange of NAADP concentrations and found thatconcentrations of NAADP as low as 50 nM inthe pipet, elicited robust oscillatory responses,similar to those evoked by CCK in untreated
Ca2+
rel
eased
cCa+
Bme
IncreasingConc.
cb
b
a
a
Sub-maxConcs
MaxIP3/cADPR
A
B
C
log [IP3 or cADPR]
Activation
Inactivation(test)
Test
Ca2+
rel
ease
log [NAADP]
ActivationInactivation(test)
Ca2+
rel
ease
log [NAADP]
Activation
Inactivation(test)
d
InactivationActivation
c
b
a-d
a
Sub-thresholdConcs
Max
Test
d
Inactivation
cc
bb
a,d
a
HighConcs
Optimal
Test
d
Inactivation
IncreasingConc.
NAADP
Type 1
Activation
IncreasingConc.
NAADP
Type 2
Activation
Figure 2. Differences between desensitization of mammalian and sea urchin NAADP receptors. Left-handtraces depict stylized Ca2þ dye fluorescence traces in response to messengers. Right-hand panels representconcentration-response graphs for activation or inactivation of intracellular Ca2þ release channels. (A) IP3and cADPR-mediated Ca2þ release. Left-hand traces depict stylized Ca2þ dye fluorescence traces in responseto various conditions. Right-hand panels represent concentration-response graphs for activation or inactiva-tion. IP3 and cADPR demonstrate simple, monophasic concentration-response curves for activation (blue).Inactivation by ever-increasing submaximal concentrations (grey period) is revealed by the reciprocal diminu-tion of a maximal “test” challenge (orange). (B) Desensitization of sea urchin NAADP receptors (Type I desen-sitization). The blue left-hand traces show stylized Ca2þ dye fluorescence traces in response to increasingconcentrations of NAADP, which increases Ca2þ release, represented by a classical sigmoid log concentra-tion-response curve (blue line, right-hand panel). However, preincubation with subthreshold concentrationsof NAADP that do not evoke Ca2þ release desensitizes Ca2þ release in a time and concentration manner bysubsequent challenge by a normally maximal NAADP (test) concentration (central panel, and orange curve,right panel). (C) Desensitization of mammalian NAADP receptors (Type 2 desensitization). Increasing concen-trations of NAADP enhances Ca2þ release to a maximum (left and center panels). Thereafter, increasing con-centrations of NAADP evoke progressively smaller Ca2þ release to a point when no Ca2þ release is evoked athigh NAADP concentrations. This “bell-shaped” or hormetic log concentration-response curve is shown inthe right-hand panel (blue curve). Modified from Morgan and Galione (2008).
NAADP Receptors
Cite this article as Cold Spring Harb Perspect Biol 2011;3:a004036 5
on September 3, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

cells. The concentration-response relationshipfor NAADP was found to be “bell-shaped,” withmaximal responses occurring at around 100 nMNAADP, whereas with concentrations in excessof 1 mM no effects were seen. Using cagedNAADP, we showed that photolysis of this com-pound also evokes a series of spikes in Ca2þ-activated currents, which were suppressed in thepresence of supramicromolar concentrationsof free NAADP in the patch pipet. Bell-shapedconcentration-response curves seem to be amajor hallmark of mammalian NAADP-inducedCa2þ release. A subsequent study in a JurkatT-cell line showed maximal Ca2þ release uponmicroinjection of around 100 nM NAADP.However, concentrations of .1 mM failed toelicit any response per se whilst inhibitingT-cell receptor activation (Berg et al. 2000). Anumber of further studies in different cell typesused this phenomenon to implicate NAADP inthe Ca2þ signal transduction pathways activatedby various stimuli in the absence of selectiveNAADP antagonists at that time. These includeglucose-evoked Ca2þ spiking in MIN6 cells(Masgrau et al. 2003), ET1-evoked Ca2þ releasein pulmonary vascular smooth myocytes (Kin-near et al. 2004), and b1 adrenoreceptor en-hancement of Ca2þ signaling and contractilityin ventricular cardiac myocytes (Macgregoret al. 2007).
PHARMACOLOGICAL PROPERTIESOF NAADP RECEPTORS
The pharmacology of NAADP-evoked Ca2þ
release, initially investigated in sea urchin eggsystems, showed major differences with theknown Ca2þ release mechanisms in the ER. Inegg homogenates, NAADP-evoked Ca2þ releasewas unaffected by the competitive IP3R in-hibitor, heparin, or by ryanodine or eight-substituted cADPR analogs that antagonizeRyR-mediated Ca2þ release. An initial reportthat thio-NADP was a selective antagonist ofNAADP (Chini et al. 1995) was subsequentlyexplained by inactivation of the NAADP-sensitive Ca2þ release mechanism by traces ofcontaminating NAADP (Dickey et al. 1998).
A number of channel blockers were foundto inhibit NAADP-evoked Ca2þ release selective-ly in sea urchin egg homogenates with littleeffect on either IP3 or cADPR-mediated Ca2þ
release (Genazzani et al. 1997a). These includedvoltage-gated Ca2þ channel (VGCC) blockerssuch as diltiazem, nifedipine, and D600 (al-though greater concentrations were required toblock NAADP-evoked Ca2þ release than VGCCs),and purinoceptor antagonists such as aspyri-doxal-phosphate-6-azophenyl-2’,4’-disulfonate(PPADS) also display a degree of NAADP an-tagonism (Billington and Genazzani 2007).Because the NAADP receptor effectively dis-criminates between NAADP and NADP, whichdiffers only by the substitution of a nicotinicacid moiety instead of nicotinamide, nicotinicacid analogs were developed that antagonizeNAADP-induced Ca2þ release. These includeCMA008 (Dowden et al. 2006) and BZ194(Dammermann et al. 2009), which also havethe advantage of being membrane permeant.Recently, a series of novel compounds have beenidentified by in silico screening strategies basedon the three-dimensional shape and electro-static properties of NAADP that are the mostpotent of NAADP antagonists developed so far(Naylor et al. 2009; Rosen et al. 2009). Ned-19,the founding member of these analogs, is be-coming the most widely used antagonist onaccount of its potency, membrane permeability,and selectivity (Naylor et al. 2009; Rosen et al.2009; Thai et al. 2009; Aley et al. 2010).
Interestingly, Ned-19 analogs have been usedto dissect the activation and inactivation effectsof NAADP at the sea urchin egg NAADP recep-tor (Rosen et al. 2009). Ned-20, which differsonly from Ned-19 by the para rather than orthoposition of a fluorine, blocks the inactivation ofNAADP-sensitive Ca2þ release mechanism bysubthreshold NAADP concentrations for Ca2þ
release, without affecting NAADP-evoked Ca2þ
release by higher NAADP concentrations andinhibits high affinity [32P]NAADP binding toegg membranes (Rosen et al. 2009). These find-ings are consistent with multiple binding sitesfor the sea urchin egg NAADP receptor, withhigh affinity sites leading to inactivation andlower affinity sites leading to activation.
A. Galione
6 Cite this article as Cold Spring Harb Perspect Biol 2011;3:a004036
on September 3, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

TWO-PORE CHANNELS
A family of novel intracellular channels termedtwo-pore channels (TPCs) have emerged asthe leading candidates for NAADP-gated Ca2þ
release channels. The founding member of thisfamily, TPC1, was cloned in 2000 from a rat kid-ney cDNA library in a search for novel membersof voltage-gated cation channels (Ishibashi et al.2000). The putative channel had only a 20%homology with the transmembrane domains ofthe a subunit of voltage-gated Naþ and Ca2þ
channels, but the highest homology with adeposited sequence of a putative Ca2þ channelfrom the plant Arabidopsis thaliana. Subsequentanalysis of the plant clone, AtTPC1, implicateda role for this protein in Ca2þ transport and sig-naling when expressed in yeast and Arabidopsis(Furuichi et al. 2001), and a role in germinationand stomatal physiology as a component ofthe slow vacuolar channel (Peiter et al. 2005).The putative channel, rather than having fourrepeats of six transmembrane segments, as forvoltage-gated Naþ and Ca2þ channels, onlyhas two. Thus in effect, the protein is the equiv-alent of half a Naþ or Ca2þ channel, and may
represent an ancestral form that has been du-plicated later in evolution to give rise to thefour domain channels (Fig. 3).
IDENTIFICATION OF TWO-PORECHANNELS AS NAADP RECEPTORS
Two clues as to the candidature of TPCs asNAADP receptors emerged in the last few years.Michael Zhu, searching for novel TRP familymembranes in around 2000, had cloned a secondmember of the TPC family, termed TPC2, andfound that when heterologously expressed inHEK293 cells, it localized with the lysosomalmarker, LAMP1. The second was the furtheranalysis of AtTPC1 function by Sanders andcolleagues, showing that AtTPC1 localized toplant vacuoles, the major plant acidic organelleand the functional equivalent of lysosomes inplants (Peiter et al. 2005). The localization ofTPCs to acidic stores, and the partial phar-macological overlap of NAADP receptors withvoltage-gated Ca2þ channels and TRP proteins,which show homologies with TPCs, made theseproteins credible candidates as the elusive
Cav2.1Cav2.2Cav2.3
Cav1.2 (L)
Four-pore channels
Two-pore channels
One-pore channels
Cav3.1 (T)
TRPP2
TPC1TPC2
TRPM1TRPV1b
TRPC1TRPA1
TRPML1CatSper3CatSper4
CatSper1CatSper2
Figure 3. Phylogenetic tree for human two-pore channels and their relationship with voltage-gated Ca2þ chan-nels and TRP members. It is likely that voltage-gated Ca2þ channels have arisen from two rounds of tandemduplication in evolution. Thus, TPCs having 12 transmembrane domains (12TM) may be considered ancientintermediate proteins between TRP channels (6TM), such as CatSpers in sperm or mucolipins or polycystins,and voltage-gated Ca2þ channels (24TM).
NAADP Receptors
Cite this article as Cold Spring Harb Perspect Biol 2011;3:a004036 7
on September 3, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

NAADP receptor. Over four years or so from2005, we worked extensively with Zhu and col-laborators, to test exhaustively the hypothesisthat TPCs represented a family of NAADP-gated intracellular channels from several keystandpoints. First, we examined the subcellularlocalization of the human TPC1 and TPC2 iso-forms in HEK293 cells. In addition, because thegenomes of many species, but not human orrodent, also express a third isoform, TPC3(Cai and Patel 2010; Zhu et al. 2010), we alsoexpressed the chicken TPC3 to examine its sub-cellular distribution in HEK293 cells (Calcraftet al. 2009). All three TPCs localize to the endo-lysosomal system with no apparent expressionin Golgi, mitochondria, or ER. Only TPC2 con-sistently colocalized with the lysosomal marker,LAMP2, but not early or late endosomes. Incontrast, TPC1 and TPC3 predominantly wereexpressed in endosomal and other unidentifiedcompartments, but with only sparse colocaliza-tion with lysosomal markers. In HEK293 cells,TPCs are endogenously expressed at low levels,and endogenous TPC2 was also immunolocal-ized to lysosomes. Overexpression of the humanHsTPC2 was associated with increased specific[32P]NAADP binding to HEK293 cell mem-branes and immunoprecipitated TPC2 pro-teins. Both high and low affinity binding siteswere manifest in membranes from TPC2-over-expressing cells, with Kd of 5 nM and 7 mM,remarkably similar to endogenous binding inmembranes from mouse liver, a tissue whichhas a particularly high expression of TPCs. Pho-tolysis of caged NAADP in patched wild-typeHEK293 cells elicited a small Ca2þ response,whereas in cells stably overexpressing TPC2, alarge biphasic Ca2þ response was evoked onNAADP uncaging or dialysis: an initial pace-maker-like ramp of Ca2þwas followed by a largerand faster transient Ca2þ release. Bafilomycintreatment abolished both phases of the Ca2þ
response, whereas the IP3R antagonist heparinblocked the second phase alone. This finding isconsistent with the “trigger” hypothesis for amode of NAADP action, whereby NAADPevokes a localized Ca2þ signal by mobilizing bafi-lomycin-sensitive acidic stores, which is thenglobalized by recruiting Ca2þ-induced Ca2þ
release (CICR) from nearby ER, in this case byactivating IP3Rs. The concentration-responserelationship between NAADP and Ca2þ releasewas of the characteristic bell-shape for NAADPin mammalian cells, with maximal Ca2þ releaseoccurring at between 10 nM and 1 mM, whereas1 mM was without effect. Importantly, shRNAagainst TPC2 completely abolished the res-ponse to NAADP. We also created Tpc2-/- mice,and found that NAADP-evoked activation ofoscillatory Ca2þ-dependent cation currents inpancreatic b cells seen in wild-type cells, wereabolished in cells from the knockout mice.
In contrast to TPC2, we found that HEKcells stably expressed with HsTPC1 evokedonly a localized Ca2þ release in response toNAADP, which failed to globalize, as was thecase for TPC2. One possibility is that the endo-somal localization of TPC1 means that there islittle close apposition with ER so that couplingwith CICR channels is weaker. Two subsequentpublications broadly confirmed our findings(Brailoiu et al. 2009a; Zong et al. 2009).
PROPERTIES OF ENDOGENOUS TPCsFROM SEA URCHIN EGGS
The properties of heterologously expressedmammalian TPCs made them strong candi-dates as NAADP receptors. However, most ofthe studies of NAADP-mediated Ca2þ releaseand [32P]NAADP binding sites have been per-formed in sea urchin egg preparations, wherethe Ca2þ mobilizing effects of NAADP werefirst discovered. It was important to ascertainwhether sea urchin eggs express TPCs andwhether they functioned as NAADP receptors.Screening of the genome of the sea urchinStrongylocentrotus purpuratus revealed threeTPC isoforms that were cloned from ovariesthat displayed around 30% sequence homologybetween the isoforms (Brailoiu et al. 2010; Ruaset al. 2010). Importantly, immunoprecipitationof TPCs from solubilized egg membranes withpolyclonal antibodies raised against each ofthe three TPC isoforms of TPCs producedimmunocomplexes that specifically bound[32P]NAADP with Kds of around 1 nM. Bindingof [32P]NAADP to the immunocomplexes
A. Galione
8 Cite this article as Cold Spring Harb Perspect Biol 2011;3:a004036
on September 3, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from
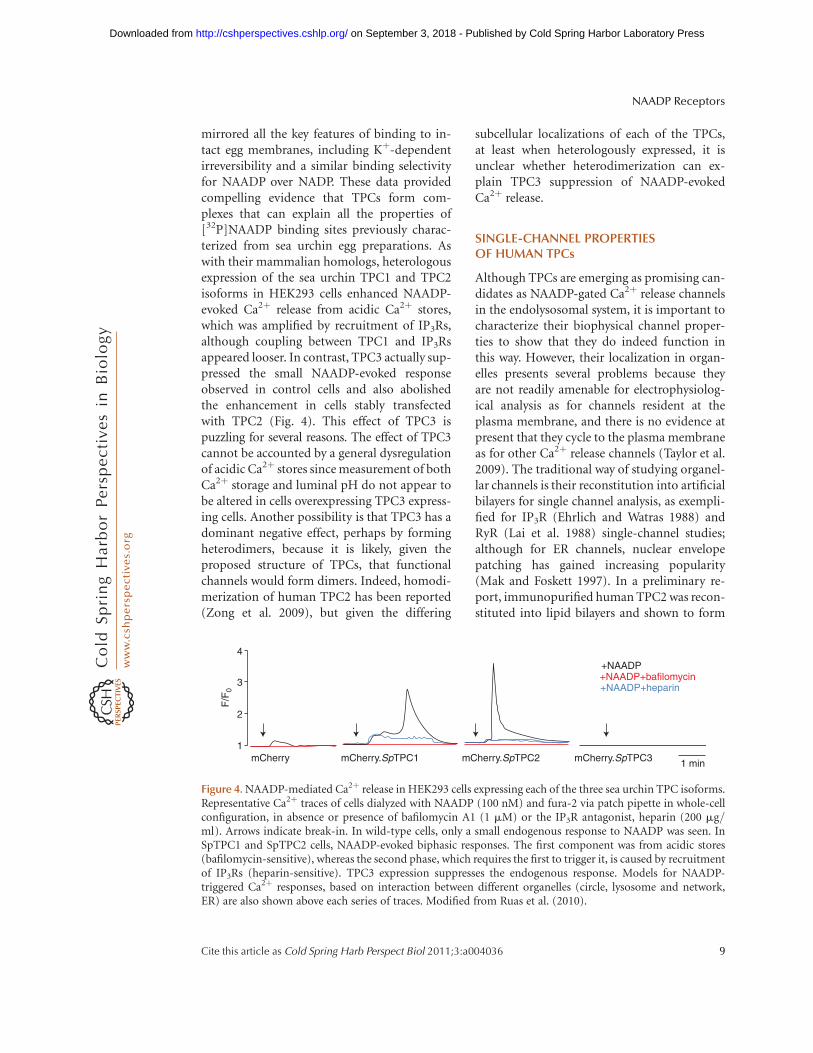
mirrored all the key features of binding to in-tact egg membranes, including Kþ-dependentirreversibility and a similar binding selectivityfor NAADP over NADP. These data providedcompelling evidence that TPCs form com-plexes that can explain all the properties of[32P]NAADP binding sites previously charac-terized from sea urchin egg preparations. Aswith their mammalian homologs, heterologousexpression of the sea urchin TPC1 and TPC2isoforms in HEK293 cells enhanced NAADP-evoked Ca2þ release from acidic Ca2þ stores,which was amplified by recruitment of IP3Rs,although coupling between TPC1 and IP3Rsappeared looser. In contrast, TPC3 actually sup-pressed the small NAADP-evoked responseobserved in control cells and also abolishedthe enhancement in cells stably transfectedwith TPC2 (Fig. 4). This effect of TPC3 ispuzzling for several reasons. The effect of TPC3cannot be accounted by a general dysregulationof acidic Ca2þ stores since measurement of bothCa2þ storage and luminal pH do not appear tobe altered in cells overexpressing TPC3 express-ing cells. Another possibility is that TPC3 has adominant negative effect, perhaps by formingheterodimers, because it is likely, given theproposed structure of TPCs, that functionalchannels would form dimers. Indeed, homodi-merization of human TPC2 has been reported(Zong et al. 2009), but given the differing
subcellular localizations of each of the TPCs,at least when heterologously expressed, it isunclear whether heterodimerization can ex-plain TPC3 suppression of NAADP-evokedCa2þ release.
SINGLE-CHANNEL PROPERTIESOF HUMAN TPCs
Although TPCs are emerging as promising can-didates as NAADP-gated Ca2þ release channelsin the endolysosomal system, it is important tocharacterize their biophysical channel proper-ties to show that they do indeed function inthis way. However, their localization in organ-elles presents several problems because theyare not readily amenable for electrophysiolog-ical analysis as for channels resident at theplasma membrane, and there is no evidence atpresent that they cycle to the plasma membraneas for other Ca2þ release channels (Taylor et al.2009). The traditional way of studying organel-lar channels is their reconstitution into artificialbilayers for single channel analysis, as exempli-fied for IP3R (Ehrlich and Watras 1988) andRyR (Lai et al. 1988) single-channel studies;although for ER channels, nuclear envelopepatching has gained increasing popularity(Mak and Foskett 1997). In a preliminary re-port, immunopurified human TPC2 was recon-stituted into lipid bilayers and shown to form
4
3
F/F
0
2
1mCherry mCherry.SpTPC1
+NAADP+NAADP+bafilomycin+NAADP+heparin
mCherry.SpTPC2 mCherry.SpTPC3 1 min
Figure 4. NAADP-mediated Ca2þ release in HEK293 cells expressing each of the three sea urchin TPC isoforms.Representative Ca2þ traces of cells dialyzed with NAADP (100 nM) and fura-2 via patch pipette in whole-cellconfiguration, in absence or presence of bafilomycin A1 (1 mM) or the IP3R antagonist, heparin (200 mg/ml). Arrows indicate break-in. In wild-type cells, only a small endogenous response to NAADP was seen. InSpTPC1 and SpTPC2 cells, NAADP-evoked biphasic responses. The first component was from acidic stores(bafilomycin-sensitive), whereas the second phase, which requires the first to trigger it, is caused by recruitmentof IP3Rs (heparin-sensitive). TPC3 expression suppresses the endogenous response. Models for NAADP-triggered Ca2þ responses, based on interaction between different organelles (circle, lysosome and network,ER) are also shown above each series of traces. Modified from Ruas et al. (2010).
NAADP Receptors
Cite this article as Cold Spring Harb Perspect Biol 2011;3:a004036 9
on September 3, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

NAADP-gated cation conductances (Pitt et al.2010). Channels were generally silent untilapplication of NAADP to the cis or cytoplasmicface of the bilayer, and the channels showed aselectivity for cations with conductances ofaround 300 pS and 15 pS for Kþ and Ca2þ
ions as the conducting species. Interestingly,NAADP sensitivity may be regulated by storefilling with Ca2þ, because NAADP sensitivitywas markedly dependent on trans or luminalCa2þ, with the EC50 for NAADP-evoked en-hancement of open probability decreasing from500 nM to 5 nM as luminal Ca2þ increased to200 mM. This is in the range of reported lu-minal free Ca2þ levels in lysosomes (Christen-sen et al. 2002; Lloyd-Evans et al. 2008). Thus,fluctuations in luminal Ca2þ because of cyclesof release and uptake of Ca2þ could be impor-tant determinants of the effects of NAADPon Ca2þ release, and offers one explanationfor how constant NAADP levels may elicit trainsof Ca2þ spikes, as widely observed in various celltypes (Cancela et al. 1999). Another variable isluminal pH of acidic stores, since NAADP hasalso been found to alkalinize acidic stores insea urchin eggs and homogenates (Morganand Galione 2007a; Morgan and Galione2007b), and it is possible that luminal pH hassignificant effects on TPC2 channel properties.Importantly, the NAADP antagonist was alsofound to block single channel TPC2 currents(Pitt et al. 2010). However, it should be stressedhere that although the immunopurified TPCcomplexes both form NAADP-gated Ca2þ chan-nels (Pitt et al. 2010) and bind [32P]NAADP(Calcraft et al. 2009; Ruas et al. 2010), the possi-bility remains that NAADP could interact withan accessory protein associated with TPCsinstead of a direct interaction with TPC proteinsthemselves (Galione et al. 2009).
A single-channel analysis of NAADP-gatedchannels has also been performed from lyso-somal enriched fractions derived from liver(Zhang and Li 2007) and bovine coronary vas-cular smooth muscle (Zhang et al. 2009). Thesechannels conducted Csþ and were sensitive toNAADP, with open probabilities displaying abell-shaped concentration dependence, andwith maximum Po occurring at 1 mM NAADP
in both preparations. The pharmacology wasconsistent with previous studies of NAADP-evoked Ca2þ release, with block by VGCC antag-onists, PPADS, and also amiloride. Interestingly,Po was increased at acidic pH. In contrast tothe situation in most mammalian cells exam-ined so far, pretreatment with concentrations ofNAADP as low as 0.5 nM blocked subsequentchannel openings by higher NAADP concentra-tions, as seen for sea urchin egg receptors andin liver (Mandi et al. 2006). The identity ofthese channels were ascribed to mucolipin-1(TRPML-1), a lysosomal TRP channel linkedto the lysosomal storage disease, mucolipidosisIV, on the basis of a blocking effect of an anti-TPRML1 antibody and reduction of channelactivity from cells treated with an siRNATPRML1 construct. However, the identity ofTRPML1 as an NAADP receptor candidateremains controversial (Pryor et al. 2006). Inaddition, a recent report suggests that NAADPmay increase levels of a short variant of aTRPML2 transcript in lymphoid cells (Samieet al. 2009), underscoring the likely complexinteractions between lysosomal channels.
INTERACTIONS OF NAADP AND OTHERCa2þ SIGNALING PATHWAYS
NAADP-evoked Ca2þ release from lysosomesappears to be small and highly localized. Giventhe dynamic properties of these organelles, theyare ideally suited to be targeted to the vicinityof Ca2þ-regulated effectors. Three modes ofNAADP-mediated Ca2þ signaling mechanismshave been highlighted (Fig. 5).
NAADP and Lysosomal-ER Interactions
Organelle interactions in Ca2þ-signaling isnot a new concept. For example, Ca2þ-micro-domains may arise around sites of ER Ca2þ
release, and neighboring organelles may beprofoundly affected physiologically. IndeedER-mitochondrial interactions have been wellstudied in the context of IP3R and RyR-mediated Ca2þ-release (Rizzuto et al. 1998;Csordas et al. 2001), which impacts on mito-chondrial metabolism and apoptotic pathways.
A. Galione
10 Cite this article as Cold Spring Harb Perspect Biol 2011;3:a004036
on September 3, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

The protein mitofusin 2 has been proposed totether ER and mitochondrial membranes (deBrito and Scorrano 2008).
NAADP-evoked Ca2þ release and its effectson Ca2þ-release channels on the ER/SR was firstnoted in pancreatic acinar cells (Cancela et al.1999). This phenomenon, whereby a localizedmicrodomain of Ca2þ release from acidic storestriggers a larger release from the ER, is widely ob-served in both the sea urchin egg and in manytypes of mammalian cell, and is one of thefundamental principles of NAADP-mediatedCa2þ signaling. The trigger hypothesis was for-mulated by the finding that NAADP-evokedresponses in pancreatic acinar cells could beblocked by either heparin or ryanodine, aswell as self-inactivation of the NAADP receptorwith NAADP itself (Cancela et al. 1999). Thiswas visualized in the larger sea urchin egg bydetailed imaging studies (Churchill andGalione 2000; Churchill and Galione 2001b;Churchill and Galione 2001a). NAADP was
found to act as a local messenger to formCa2þ gradients across the cell based on NAADPdiffusion. These gradients could be amplifiedand globalized by CICR through the recruit-ment of IP3R and RyR-dependent mechanismson the ER. Because of the distinct self-inactivation properties of NAADP receptors,subsequent NAADP-evoked Ca2þ-signaling pat-terns only occur in regions of the cell whereNAADP had not previously evoked a response(Churchill and Galione 2001b). This effect lastsfor many minutes, representing a basic type ofspatiotemporal memory in terms of the genera-tion of Ca2þ-signal patterning. As well as spatialcomplexities, NAADP could produce temporalpatterns in Ca2þ signals by the uptake of Ca2þ
released from NAADP-sensitive stores into theER to produce a series of Ca2þ spikes dependenton IP3R and RyRs (Churchill and Galione2001a).
In pulmonary vascular smooth musclecells, NAADP and the vasoactive hormone,
Trigger
ER
Acidic store
Resting
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
IP3R
SERCA
TPC
? NAADP
? ?
CICR amplification andglobalization
Acidic store
Plasmamembrane
Resting
CHX
TPC
?
Local Cross Talk
NAADP?
X+ or Y–
ΔΨ
A
B C
H+-ATPase
H+-ATPase
CHX
3. Fusion
NAADP
1. Docking
Lysosome
Endosome
2. Trigger
H+
H+
Figure 5. Three modes of NAADP-mediated Ca2þ signaling. (A) NAADP is a local trigger mechanism for det-onating global CICR responses from the ER. (B) Local Ca2þ release by NAADP from acidic stores positionedunder the plasma membrane may regulate membrane excitability (excitable cells) or ion fluxes (nonexcitablecells) by modulating Ca2þ-activated plasma membrane channels. (C) NAADP regulates local cytoplasmicCa2þ/pH and luminal Ca2þ/pH in endolysosomal compartments that may regulate vesicular fusion of lateendosomes/lysosomes. Modified from Galione et al. (2009) and Ruas et al. (2010).
NAADP Receptors
Cite this article as Cold Spring Harb Perspect Biol 2011;3:a004036 11
on September 3, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

endothelin-1, evoke a localized Ca2þ releasefrom lysosomes at lysosomal-SR junctions,which is then amplified and globalized by amechanism dependent on recruitment ofRyRs on the SR (Kinnear et al. 2004; Kinnearet al. 2008). Similar results have been reportedin coronary smooth muscle myocytes (Zhanget al. 2006), and also implicated for early Fas sig-naling processes, which eventually lead to apop-tosis (Zhang et al. 2010).
In Jurkat T cells, NAADP triggers Ca2þ
release, which can be amplified by RyRs andIP3Rs, but it has been proposed in this systemthat RyR1 may be the primary target of NAADPon the ER (Dammermann and Guse 2005;Dammermann et al. 2009). A role for RyR asthe direct target for NAADP has also beenproposed in pancreatic acinar cell ER/nuclearmembranes, although other evidence pointsto direct activation of acidic stores (Yamasakiet al. 2004; Menteyne et al. 2006) followed byamplification by CICR. Such discrepancies arenot surprising given the small release of Ca2þ
released by lysosomes that TPC studies haverevealed (Calcraft et al. 2009; Ruas et al. 2010),with amplification by ER mechanisms pro-viding much larger Ca2þ signals. Thus in smallcells, dissection of contributory Ca2þ releasemechanisms can prove difficult (Galione andPetersen 2005), but employment of emergingmolecular insights and tools may prove insight-ful (Galione et al. 2009).
Modulation of Plasma Membrane Excitability
As well as their involvement in organelle com-munication, NAADP and TPCs appear to playan important role in regulating ion fluxesacross the plasma membrane and hence alsoexcitability of excitable cells. NAADP has beenshown to stimulate Ca2þ influx across theplasma membrane of several cell types includingstarfish oocytes (Moccia et al. 2003; Moccia et al.2006), sea urchin eggs (Churchill et al. 2003),where it uniquely among Ca2þmobilizing mes-sengers mediates the polyspermic blocking“cortical flash,” and Jurkat T cells (Langhorstet al. 2004). What is not clear is whether NAADPdirectly activates plasma membrane channels or
whether NAADP first releases Ca2þ from intra-cellular stores, which then leads to activationof plasma membrane conductances. Indeed, atpresent, there is no evidence for TPC localiza-tion at the plasma membrane.
However, local NAADP-evoked Ca2þ releasefrom acidic stores in the vicinity of the plasmamembrane has been shown in several cell typesto open Ca2þ-activated ion channels. This wasfirst shown in non-excitable pancreatic acinarcells, where activation of such channels is likelyto contribute to fluid secretion (Cancela et al.1999). However, this may be a major mechanismin excitable cells. In pancreatic b cells, NAADPalso evokes Ca2þ-dependent currents, whichmay contribute to glucose-mediated depolariza-tion of the cells during stimulus-secretion cou-pling (Naylor et al. 2009), and which are absentin cells derived from Tpc2-/- mice (Calcraftet al. 2009). In neurons from the rat medullaoblongata (Brailoiu et al. 2009b), NAADP alsodepolarizes cells through a mechanism depend-ent on Ca2þ release from acidic stores.
NAADP and its Receptors in EndolysosomalPhysiology
NAADP may be unique among Ca2þ mobiliz-ing messengers in that in contrast to IP3 orcADPR, it may in most cases directly evokeCa2þ release from the endolysosomal system.NAADP-regulated TPCs are a new group ofchannels that are targeted to the endolysosomalsystem, along with mucolipins (Dong et al.2010), P2X4 receptors (Qureshi et al. 2007),and TRPM2 (Lange et al. 2009), all of whichare likely to influence the ionic environmentin acidic organelles. Interestingly, TRPM2 chan-nels have also been proposed to be NAADPreceptors (Beck et al. 2006); however, theyhave much lower affinities for NAADP, in thehighmM range. TRPM2 channels could providelocal Ca2þ signals that may directly impingeon the pleiotropic roles of the endolysosomalsystem including lysosomal biogenesis, vesicu-lar trafficking and transport, and autophagy.Both local and luminal Ca2þ is important formany of these processes including homotypicfusion processes of endosomes and heterotypic
A. Galione
12 Cite this article as Cold Spring Harb Perspect Biol 2011;3:a004036
on September 3, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

fusions of late endosomes with lysosomes, aswell as condensation of luminal contents (Piperand Luzio 2004; Luzio et al. 2007); release ofCa2þ from endolysosomal stores is thought tobe a crucial regulatory mechanism. Overexpres-sion of TPCs in HEK293 causes profoundchanges in trafficking, lysosomal size, and dis-tribution as observed in certain lysosomal stor-age diseases (Ruas et al. 2010). These effects canbe ameliorated by treatment with the NAADPantagonist, Ned-19. These data are suggestiveof a major role for NAADP and TPC proteinsin the regulation of luminal Ca2þ, Ca2þ release,and local Ca2þ signaling in endolysosomalphysiology, and are thus likely to be key regula-tors of trafficking, autophagy, and other func-tions of these organelles.
Conclusions: Why Have Multiple Messengersfor Ca2þ Release?
Over the last decade or so, NAADP has joined IP3
and cADPR as a major Ca2þmobilizing messen-ger. A major question in Ca2þ signaling researchis how ubiquitous Ca2þ signals can encode spe-cificity. A general view is that the complex spatialand temporal patterns of Ca2þ signals widelyobserved in cells are key to understanding thisproblem. The coordination of Ca2þ signals bymultiple messengers acting at differentially dis-tributed target Ca2þ release channels with dif-ferent properties offers one possible solution.For example, NAADP-evoked Ca2þ release leadsto neuronal cell differentiation (Brailoiu et al.2006), whereas cADPR-mediated Ca2þ releaseleads to cell proliferation, but delays differen-tiation (Yue et al. 2009). On the other hand, acti-vation of certain cell surface receptors mayproduce different combinations of messengersthat are required to mimic the specific Ca2þ sig-naling patterns evoked by the particular recep-tor agonist (Cancela et al. 2002; Yamasaki et al.2005), thus increasing the repertoire of cellularresponses mediated by Ca2þ.
The emerging view that NAADP directly tar-gets acidic stores rather than the ER is an impor-tant new principle in Ca2þ signaling and cellularhomeostasis, and allows NAADP to evoke distinctCa2þ signals from those directly mobilizing Ca2þ
from the ER. This was initially proposed on thebasis of pharmacological studies, but the identifi-cation of endolysosomal TPC proteins as majortargets for NAADP has begun to cement thishypothesis in molecular terms. Three major con-sequences of NAADP-evoked Ca2þ release havebeen identified. The unifying principle is thatNAADP, by mobilizing acidic stores, leads to lo-calized Ca2þ signals that may trigger key cellularresponses. Depending on the subcellular locali-zation of these stores, there are fundamentally dif-ferent consequences of NAADP-mediated Ca2þ
release. For stores proximal to the plasma mem-brane, Ca2þ-activated plasma channels may beactivated. Such ion fluxes produced in nonexcit-able cells may, for example, be important in fluidsecretion. In excitable cells, depolarization andchanges in membrane excitability may result.For stores apposed to the ER, NAADP-evokedCa2þ release from acidic stores may triggerglobalized Ca2þ responses by activating IP3Rs orRyRs by CICR. The third major aspect is the reg-ulation of luminal Ca2þ and pH, as well as localCa2þ signals in the endolysosomal system thatmay have a major impact on the many roles ofthese organelles in key cellular processes thatthey control, including vesicular trafficking,autophagy, apoptosis, and autolysis, as well astheir role in fighting infection. Cellular stimulimay be selectively coupled to NAADP signalingpathways, or as is commonly observed, to mul-tiple messenger pathways, either providing dis-tinct patterns of Ca2þ signals leading to specificresponses.
The establishment of a role of the endolyso-somal system in Ca2þ signaling, the identifica-tion of specific Ca2þ release channels of acidicorganelles as the targets for NAADP, open upnew possibilities for a better understanding ofthe mechanisms of cellular Ca2þ signaling andhow this goes awry in disease, and its controland pharmacological manipulation.
ACKNOWLEDGMENTS
This work in AG’s laboratory is funded by theWellcome Trust, and AG is a Principal Investiga-tor of the British Heart Foundation Centre ofResearch Excellence at the University of Oxford.
NAADP Receptors
Cite this article as Cold Spring Harb Perspect Biol 2011;3:a004036 13
on September 3, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

I would like to thank Dr. Anthony Morgan forhis helpful discussion and help with preparingthe figures.
REFERENCES
Aarhus R, Dickey DM, Graeff RM, Gee KR, Walseth TF, LeeHC. 1996. Activation and inactivation of Ca2þ release byNAADPþ. J Biol Chem 271: 8513–8516.
Aley PK, Noh HJ, Gao X, Tica AA, Brailoiu E, Churchill GC.2010. A functional role for nicotinic acid adeninedinucleotide phosphate (NAADP) in oxytocin-mediatedcontraction of uterine smooth muscle from rat. J Phar-macol Exp Ther 333: 726–735.
Beck A, Kolisek M, Bagley LA, Fleig A, Penner R. 2006.Nicotinic acid adenine dinucleotide phosphate and cyclicADP-ribose regulate TRPM2 channels in T lymphocytes.FASEB J 20: 962–964.
Berg I, Potter BV, Mayr GW, Guse AH. 2000. Nicotinic acidadenine dinucleotide phosphate (NAADPþ) is an essen-tial regulator of T-lymphocyte Ca2þ-signaling. J Cell Biol150: 581–588.
Berridge MJ, Bootman MD, Lipp P. 1998. Calcium–a lifeand death signal. Nature 395: 645–648.
Berridge MJ, Bootman MD, Roderick HL. 2003. Calcium:Calcium signalling: dynamics, homeostasis and remodel-ling. Nat Rev Mol Cell Biol 4: 517–529.
Billington RA, Genazzani AA. 2007. PPADS is a reversiblecompetitive antagonist of the NAADP receptor. Cell Cal-cium 41: 505–511.
Bootman MD, Berridge MJ, Roderick HL. 2002. Calciumsignalling: More messengers, more channels, more com-plexity. Curr Biol 12: R563–565.
Brailoiu GC, Brailoiu E, Parkesh R, Galione A, ChurchillGC, Patel S, Dun NJ. 2009b. NAADP-mediated channel‘chatter’ in neurons of the rat medulla oblongata. Bio-chem J 419: 91–97.
Brailoiu E, Churamani D, Cai X, Schrlau MG, Brailoiu GC,Gao X, Hooper R, Boulware MJ, Dun NJ, Marchant JS,et al. 2009a. Essential requirement for two-pore channel1 in NAADP-mediated calcium signaling. J Cell Biol 186:201–209.
Brailoiu E, Churamani D, Pandey V, Brailoiu GC, Tuluc F,Patel S, Dun NJ. 2006. Messenger-specific role for nico-tinic acid adenine dinucleotide phosphate in neuronaldifferentiation. J Biol Chem 281: 15923–15928.
Brailoiu E, Hooper R, Cai X, Brailoiu GC, Keebler MV,Dun NJ, Marchant JS, Patel S. 2010. An ancestral deu-terostome family of two-pore channels mediates nico-tinic acid adenine dinucleotide phosphate-dependentcalcium release from acidic organelles. J Biol Chem285: 2897–2901.
Cai X, Patel S. 2010. Degeneration of an intracellular ionchannel in the primate lineage by relaxation of selectiveconstraints. Mol Biol Evol (in press).
Calcraft PJ, Ruas M, Pan Z, Cheng X, Arredouani A, Hao X,Tang J, Rietdorf K, Teboul L, Chuang KT, et al. 2009.NAADP mobilizes calcium from acidic organellesthrough two-pore channels. Nature 459: 596–600.
Cancela JM, Churchill GC, Galione A. 1999. Coordinationof agonist-induced Ca2þ-signalling patterns by NAADPin pancreatic acinar cells. Nature 398: 74–76.
Cancela JM, Van Coppenolle F, Galione A, Tepikin AV,Petersen OH. 2002. Transformation of local Ca2þ spikesto global Ca2þ transients: The combinatorial roles ofmultiple Ca2þ releasing messengers. EMBO J 21: 909–919.
Chini EN, Beers KW, Dousa TP. 1995. Nicotinate adeninedinucleotide phosphate (NAADP) triggers a specific cal-cium release system in sea urchin eggs. J Biol Chem 270:3216–3223.
Christensen KA, Myers JT, Swanson JA. 2002. pH-depend-ent regulation of lysosomal calcium in macrophages. JCell Sci 115: 599–607.
Churchill GC, Galione A. 2000. Spatial control of Ca2þ sig-naling by nicotinic acid adenine dinucleotide phosphatediffusion and gradients. J Biol Chem 275: 38687–38692.
Churchill GC, Galione A. 2001a. NAADP induces Ca2þ
oscillations via a two-pool mechanism by primingIP3- and cADPR-sensitive Ca2þ stores. EMBO J 20:2666–2671.
Churchill GC, Galione A. 2001b. Prolonged inactivation ofnicotinic acid adenine dinucleotide phosphate-inducedCa2þ release mediates a spatiotemporal Ca2þ memory. JBiol Chem 276: 11223–11225.
Churchill GC, Okada Y, Thomas JM, Genazzani AA, Patel S,Galione A. 2002. NAADP mobilizes Ca2þ from reservegranules, lysosome-related organelles, in sea urchineggs. Cell 111: 703–708.
Churchill GC, O’Neill JS, Masgrau R, Patel S, Thomas JM,Genazzani AA, Galione A. 2003. Sperm deliver a new sec-ond messenger: NAADP. Curr Biol 13: 125–128.
Clapham DE. 1995. Calcium signaling. Cell 80: 259–268.
Clapper DL, Walseth TF, Dargie PJ, Lee HC. 1987. Pyridinenucleotide metabolites stimulate calcium release from seaurchin egg microsomes desensitized to inositol trisphos-phate. J Biol Chem 262: 9561–9568.
Csordas G, Thomas AP, Hajnoczky G. 2001. Calcium signaltransmission between ryanodine receptors and mito-chondria in cardiac muscle. Trends Cardiovasc Med 11:269–275.
Dammermann W, Guse AH. 2005. Functional ryanodinereceptor expression is required for NAADP-mediatedlocal Ca2þ signaling in T-lymphocytes. J Biol Chem280: 21394–21399.
Dammermann W, Zhang B, Nebel M, Cordiglieri C,Odoardi F, Kirchberger T, Kawakami N, Dowden J,Schmid F, Dornmair K, et al. 2009. NAADP-mediatedCa2þ signaling via type 1 ryanodine receptor in T cellsrevealed by a synthetic NAADP antagonist. Proc NatlAcad Sci 106: 10678–10683.
de Brito OM, Scorrano L. 2008. Mitofusin 2 tethersendoplasmic reticulum to mitochondria. Nature 456:605–610.
Dickey DM, Aarhus R, Walseth TF, Lee HC. 1998. Thio-NADP is not an antagonist of NAADP. Cell Biochem Bio-phys 28: 63–73.
Dickinson GD, Churchill GC, Brailoiu E, Patel S. 2010.Deviant NAADP-mediated Ca2þ-signalling upon lyso-some proliferation. J Biol Chem 285: 13321–13325.
A. Galione
14 Cite this article as Cold Spring Harb Perspect Biol 2011;3:a004036
on September 3, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

Dong XP, Wang X, Xu H. 2010. TRP channels of intracellularmembranes. J Neurochem 113: 313–328.
Dowden J, Berridge G, Moreau C, Yamasaki M, ChurchillGC, Potter BV, Galione A. 2006. Cell-permeant small-molecule modulators of NAADP-mediated Ca2þ release.Chem Biol 13: 659–665.
Ehrlich BE, Watras J. 1988. Inositol 1,4,5-trisphosphate acti-vates a channel from smooth muscle sarcoplasmic retic-ulum. Nature 336: 583–586.
Furuichi T, Cunningham KW, Muto S. 2001. A putative twopore channel AtTPC1 mediates Ca2þ flux in Arabidopsisleaf cells. Plant Cell Physiol 42: 900–905.
Galione A. 2006. NAADP, a new intracellular messenger thatmobilizes Ca2þ from acidic stores. Biochem Soc Trans 34:922–926.
Galione A. 2008. Investigating cADPR and NAADP in intactand broken cell preparations. Methods 46: 194–203.
Galione A, Petersen OH. 2005. The NAADP receptor: Newreceptors or new regulation? Mol Interv 5: 73–79.
Galione A, Evans AM, Ma J, Parrington J, Arredouani A,Cheng X, Zhu MX. 2009. The acid test: The discoveryof two-pore channels (TPCs) as NAADP-gated end-olysosomal Ca2þ release channels. Pflugers Arch 458:869–876.
Gambara G, Billington RA, Debidda M, D’Alessio A, Pal-ombi F, Ziparo E, Genazzani AA, Filippini A. 2008.NAADP-induced Ca2þ signaling in response to endothe-lin is via the receptor subtype B and requires the integrityof lipid rafts/caveolae. J Cell Physiol 216: 396–404.
Gasser A, Bruhn S, Guse AH. 2006. Second messenger func-tion of nicotinic acid adenine dinucleotide phosphaterevealed by an improved enzymatic cycling assay. J BiolChem 281: 16906–16913.
Genazzani AA, Galione A. 1996. Nicotinic acid-adeninedinucleotide phosphate mobilizes Ca2þ from a thapsigar-gin-insensitive pool. Biochem J 315: 721–725.
Genazzani AA, Empson RM, Galione A. 1996. Unique inac-tivation properties of NAADP-sensitive Ca2þ release. JBiol Chem 271: 11599–11602.
Genazzani AA, Mezna M, Dickey DM, Michelangeli F, Wal-seth TF, Galione A. 1997a. Pharmacological properties ofthe Ca2þ-release mechanism sensitive to NAADP in thesea urchin egg. Br J Pharmacol 121: 1489–1495.
Genazzani AA, Mezna M, Summerhill RJ, Galione A,Michelangeli F. 1997b. Kinetic properties of nicotinicacid adenine dinucleotide phosphate-induced Ca2þ
release. J Biol Chem 272: 7669–7675.
Gerasimenko JV, Flowerdew SE, Voronina SG, SukhomlinTK, Tepikin AV, Petersen OH, Gerasimenko OV. 2006.Bile acids induce Ca2þ release from both the endoplasmicreticulum and acidic intracellular calcium stores throughactivation of inositol trisphosphate receptors and ryano-dine receptors. J Biol Chem 281: 40154–40163.
Graeff R, Lee HC. 2002. A novel cycling assay for nicotinicacid-adenine dinucleotide phosphate with nanomolarsensitivity. Biochem J 367: 163–168.
Ishibashi K, Suzuki M, Imai M. 2000. Molecular cloning of anovel form (two-repeat) protein related to voltage-gatedsodium and calcium channels. Biochem Biophys ResCommun 270: 370–376.
Jardin I, Lopez JJ, Pariente JA, Salido GM, Rosado JA. 2008.Intracellular calcium release from human platelets: Dif-ferent messengers for multiple stores. Trends CardiovascMed 18: 57–61.
Kim BJ, Park KH, Yim CY, Takasawa S, Okamoto H, Im MJ,Kim UH. 2008. Generation of nicotinic acid adeninedinucleotide phosphate and cyclic ADP-ribose byglucagon-like peptide-1 evokes Ca2þ signal that is essen-tial for insulin secretion in mouse pancreatic islets. Dia-betes 57: 868–878.
Kinnear NP, Boittin FX, Thomas JM, Galione A, Evans AM.2004. Lysosome-sarcoplasmic reticulum junctions. Atrigger zone for calcium signaling by nicotinic acidadenine dinucleotide phosphate and endothelin-1. JBiol Chem 279: 54319–54326.
Kinnear NP, Wyatt CN, Clark JH, Calcraft PJ, Fleischer S,Jeyakumar LH, Nixon GF, Evans AM. 2008. Lysosomesco-localize with ryanodine receptor subtype 3 to form atrigger zone for calcium signalling by NAADP in ratpulmonary arterial smooth muscle. Cell Calcium 44:190–201.
Lai FA, Erickson HP, Rousseau E, Liu QY, Meissner G. 1988.Purification and reconstitution of the calcium releasechannel from skeletal muscle. Nature 331: 315–319.
Lange I, Yamamoto S, Partida-Sanchez S, Mori Y, FleigA, Penner R. 2009. TRPM2 functions as a lysosomalCa2þ-release channel in beta cells. Sci Signal 2: pra23.
Langhorst MF, Schwarzmann N, Guse AH. 2004. Ca2þrelease via ryanodine receptors and Ca2þ entry: Majormechanisms in NAADP-mediated Ca2þ signaling in T-lymphocytes. Cell Signal 16: 1283–1289.
Lee HC, Aarhus R. 1995. A derivative of NADP mobilizescalcium stores insensitive to inositol trisphosphate andcyclic ADP-ribose. J Biol Chem 270: 2152–2157.
Lee HC, Aarhus R. 2000. Functional visualization of the sep-arate but interacting calcium stores sensitive to NAADPand cyclic ADP-ribose. J Cell Sci 113: 4413–4420.
Lee HC, Walseth TF, Bratt GT, Hayes RN, Clapper DL. 1989.Structural determination of a cyclic metabolite of NADwith intracellular calcium-mobilizing activity. J BiolChem 264: 1608–1615.
Lewis AM, Masgrau R, Vasudevan SR, Yamasaki M, O’NeillJS, Garnham C, James K, Macdonald A, Ziegler M,Galione A, et al. 2007. Refinement of a radioreceptorbinding assay for nicotinic acid adenine dinucleotidephosphate. Anal Biochem 371: 26–36.
Lloyd-Evans E, Morgan AJ, He X, Smith DA, Elliot-Smith E,Sillence DJ, Churchill GC, Schuchman EH, Galione A,Platt FM. 2008. Niemann-Pick disease type C1 is a sphin-gosine storage disease that causes deregulation of lysoso-mal calcium. Nat Med 14: 1247–1255.
Luzio JP, Bright NA, Pryor PR. 2007. The role of calcium andother ions in sorting and delivery in the late endocyticpathway. Biochem Soc Trans 35: 1088–1091.
Macgregor A, Yamasaki M, Rakovic S, Sanders L, Parkesh R,Churchill GC, Galione A, Terrar DA. 2007. NAADP con-trols cross-talk between distinct Ca2þ stores in the heart. JBiol Chem 282: 15302–15311.
Mak DO, Foskett JK. 1997. Single-channel kinetics, inacti-vation, and spatial distribution of inositol trisphosphate(IP3) receptors in Xenopus oocyte nucleus. J Gen Physiol109: 571–587.
NAADP Receptors
Cite this article as Cold Spring Harb Perspect Biol 2011;3:a004036 15
on September 3, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

Mandi M, Toth B, Timar G, Bak J. 2006. Ca2þ release trig-gered by NAADP in hepatocyte microsomes. Biochem J395: 233–238.
Masgrau R, Churchill GC, Morgan AJ, Ashcroft SJ, Gali-one A. 2003. NAADP: A new second messenger forglucose-induced Ca2þ responses in clonal pancreaticbeta cells. Curr Biol 13: 247–251.
Menteyne A, Burdakov A, Charpentier G, Petersen OH,Cancela JM. 2006. Generation of specific Ca2þ signalsfrom Ca2þ stores and endocytosis by differential couplingto messengers. Curr Biol 16: 1931–1937.
Mitchell KJ, Lai FA, Rutter GA. 2003. Ryanodine receptortype I and nicotinic acid adenine dinucleotide phosphatereceptors mediate Ca2þ release from insulin-containingvesicles in living pancreatic beta-cells (MIN6). J BiolChem 278: 11057–11064.
Moccia F, Billington RA, Santella L. 2006. Pharmacologicalcharacterization of NAADP-induced Ca2þ signals in star-fish oocytes. Biochem Biophys Res Commun 348: 329–336.
Moccia F, Lim D, Nusco GA, Ercolano E, Santella L. 2003.NAADP activates a Ca2þ current that is dependent onF-actin cytoskeleton. FASEB J 17: 1907–1909.
Morgan AJ, Galione A. 2007a. Fertilization and nicotinicacid adenine dinucleotide phosphate induce pH changesin acidic Ca2þ stores in sea urchin eggs. J Biol Chem 282:37730–37737.
Morgan AJ, Galione A. 2007b. NAADP induces pH changesin the lumen of acidic Ca2þ stores. Biochem J 402: 301–310.
Naylor E, Arredouani A, Vasudevan SR, Lewis AM, ParkeshR, Mizote A, Rosen D, Thomas JM, Izumi M, Ganesan A,et al. 2009. Identification of a chemical probe for NAADPby virtual screening. Nat Chem Biol 5: 220–226.
Pandey V, Chuang CC, Lewis AM, Aley PK, Brailoiu E,Dun NJ, Churchill GC, Patel S. 2009. Recruitment ofNAADP-sensitive acidic Ca2þ stores by glutamate. Bio-chem J 422: 503–512.
Patel S, Docampo R. 2010. Acidic calcium stores open forbusiness: Expanding the potential for intracellular Ca2þ
signaling. Trends Cell Biol 20: 277–286.
Peiter E, Maathuis FJ, Mills LN, Knight H, Pelloux J, Hether-ington AM, Sanders D. 2005. The vacuolar Ca2þ-acti-vated channel TPC1 regulates germination andstomatal movement. Nature 434: 404–408.
Piper RC, Luzio JP. 2004. CUPpling calcium to lysosomalbiogenesis. Trends Cell Biol 14: 471–473.
Pitt SJ, Funnell T, Sitsapesan M, Venturi E, Rietdorf K, RuasM, Ganesan A, Gosain R, Churchill GC, Zhu MX, et al.2010. TPC2 is a novel NAADP-sensitive Ca2þ-releasechannel, operating as a dual sensor of luminal pHand Ca2þ. J Biol Chem M110.156927[pii]10.1074/jbc.M110.156927 (in press).
Pozzan T, Rizzuto R, Volpe P, Meldolesi J. 1994. Molecularand cellular physiology of intracellular calcium stores.Physiological Reviews 74: 595–636.
Pryor PR, Reimann F, Gribble FM, Luzio JP. 2006. Mucoli-pin-1 is a lysosomal membrane protein required for intra-cellular lactosylceramide traffic. Traffic 7: 1388–1398.
Qureshi OS, Paramasivam A, Yu JC, Murrell-Lagnado RD.2007. Regulation of P2X4 receptors by lysosomal
targeting, glycan protection and exocytosis. J Cell Sci120: 3838–3849.
Rizzuto R, Pinton P, Carrington W, Fay FS, Fogarty KE, Lif-shitz LM, Tuft RA, Pozzan T. 1998. Close contacts withthe endoplasmic reticulum as determinants of mito-chondrial Ca2þ responses. Science 280: 1763–1766.
Rosen D, Lewis AM, Mizote A, Thomas JM, Aley PK, Vasu-devan SR, Parkesh R, Galione A, Izumi M, Ganesan A,et al. 2009. Analogues of the nicotinic acid adenine dinu-cleotide phosphate (NAADP) antagonist Ned-19 indi-cate two binding sites on the NAADP receptor. J BiolChem 284: 34930–34934.
Ruas M, Rietdorf K, Arredouani A, Davis LC, Lloyd-EvansE, Koegel H, Funnell TM, Morgan AJ, Ward JA, WatanabeK, et al. 2010. Purified TPC isoforms form NAADPreceptors with distinct roles for Ca2þ signaling and endo-lysosomal trafficking. Curr Biol 20: 703–709.
Rutter GA. 2003. Calcium signalling: NAADP comes out ofthe shadows. Biochem J 373: e3–4.
Samie MA, Grimm C, Evans JA, Curcio-Morelli C, Heller S,Slaugenhaupt SA, Cuajungco MP. 2009. The tissue-spe-cific expression of TRPML2 (MCOLN-2) gene is influ-enced by the presence of TRPML1. Pflugers Arch 459:79–91.
Streb H, Irvine RF, Berridge MJ, Schulz I. 1983. Release ofCa2þ from a nonmitochondrial intracellular store in pan-creatic acinar cells by inositol-1,4,5-triphosphate. Nature306: 67–69.
Taylor CW, Prole DL, Rahman T. 2009. Ca2þ channels on themove. Biochemistry 48: 12062–12080.
Thai TL, Churchill GC, Arendshorst WJ. 2009. NAADPreceptors mediate calcium signaling stimulated byendothelin-1 and norepinephrine in renal afferent arte-rioles. Am J Physiol Renal Physiol 297: F510–516.
Yamasaki M, Masgrau R, Morgan AJ, Churchill GC, Patel S,Ashcroft SJ, Galione A. 2004. Organelle selection deter-mines agonist-specific Ca2þ signals in pancreatic acinarand beta cells. J Biol Chem 279: 7234–7240.
Yamasaki M, Thomas JM, Churchill GC, Garnham C, LewisAM, Cancela JM, Patel S, Galione A. 2005. Role ofNAADP and cADPR in the induction and maintenanceof agonist-evoked Ca2þ spiking in mouse pancreatic aci-nar cells. Curr Biol 15: 874–878.
Yue J, Wei W, Lam CM, Zhao YJ, Dong M, Zhang LR, ZhangLH, Lee HC. 2009. CD38/cADPR/Ca2þ pathway pro-motes cell proliferation and delays nerve growth factor-induced differentiation in PC12 cells. J Biol Chem 284:29335–29342.
Zhang F, Li PL. 2007. Reconstitution and characterizationof a nicotinic acid adenine dinucleotide phosphate(NAADP)-sensitive Ca2þ release channel from liver lyso-somes of rats. J Biol Chem 282: 25259–25269.
Zhang F, Xia M, Li PL. 2010. Lysosome-dependent Ca2þ
release respsone to fas activation in coronary arterialmyocytes through NAADP: evidence from cd38 geneknockouts. Am J Physiol Cell Physiol 298: C1209–C1216.
Zhang F, Jin S, Yi F, Li PL. 2009. TRP-ML1 functions as alysosomal NAADP-sensitive Ca2þ release channel incoronary arterial myocytes. J Cell Mol Med 13: 3174–3185.
A. Galione
16 Cite this article as Cold Spring Harb Perspect Biol 2011;3:a004036
on September 3, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

Zhang F, Zhang G, Zhang AY, Koeberl MJ, Wallander E, LiPL. 2006. Production of NAADP and its role in Ca2þ
mobilization associated with lysosomes in coronaryarterial myocytes. Am J Physiol Heart Circ Physiol 291:H274–282.
Zhu MX, Ma J, Parrington J, Galione A, Mark Evans A. 2010.TPCs: Endolysosomal channels for Ca2þ mobilization
from acidic organelles triggered by NAADP. FEBS Lett584: 1966–1974.
Zong X, Schieder M, Cuny H, Fenske S, Gruner C, Rotzer K,Griesbeck O, Harz H, Biel M, Wahl-Schott C. 2009. Thetwo-pore channel TPCN2 mediates NAADP-dependentCa2þ-release from lysosomal stores. Pflugers Arch 458:891–899.
Cite this article as Cold Spring Harb Perspect Biol 2011;3:a004036 17
NAADP Receptors
on September 3, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

November 3, 20102011; doi: 10.1101/cshperspect.a004036 originally published onlineCold Spring Harb Perspect Biol
Antony Galione NAADP Receptors
Subject Collection Calcium Signaling
Perspectives on Mechanism and FunctionStore-Operated Calcium Channels: New
Richard S. Lewis
Calcium Signaling in Neuronal DevelopmentSheila S. Rosenberg and Nicholas C. Spitzer
Calcium Signaling in Smooth Muscle
Thomas J. Heppner, et al.David C. Hill-Eubanks, Matthias E. Werner,
Calcium Signaling in Cardiac Myocytes
D. BootmanClaire J. Fearnley, H. Llewelyn Roderick and Martin
LipidProtein Kinase C: The ''Masters'' of Calcium and
Peter Lipp and Gregor Reither
Voltage-Gated Calcium ChannelsWilliam A. Catterall
CommunicationCalcium Signaling in Synapse-to-Nucleus
Anna M. Hagenston and Hilmar BadingAdenylyl Cyclases
-Signaling Pathways of2+Regulation by Ca
Michelle L. Halls and Dermot M.F. CooperNAADP Receptors
Antony Galione DiseaseEndoplasmic-Reticulum Calcium Depletion and
Djalila Mekahli, Geert Bultynck, Jan B. Parys, et al.
and Golgi Apparatus Pumps of the Endoplasmic Reticulum2+The Ca
Raeymaekers, et al.Ilse Vandecaetsbeek, Peter Vangheluwe, Luc
Requirements, Players, and Adaptations Signaling During Mammalian Fertilization:2+Ca
FissoreTakuya Wakai, Veerle Vanderheyden and Rafael A.
Neuronsand Calcium Signaling in Mammalian Central mGluR1/TRPC3-mediated Synaptic Transmission
KonnerthJana Hartmann, Horst A. Henning and Arthur
Calcium Oscillations
Bird, et al.Geneviève Dupont, Laurent Combettes, Gary S.
Organellar Calcium BuffersDaniel Prins and Marek Michalak Skeletal Muscle Formation in Vertebrates
Signaling During Embryonic2+Visualization of Ca
Sarah E. Webb and Andrew L. Miller
http://cshperspectives.cshlp.org/cgi/collection/ For additional articles in this collection, see
Copyright © 2011 Cold Spring Harbor Laboratory Press; all rights reserved
on September 3, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from






















![[Pharma] receptors](https://img.pdfslide.net/doc/110x75/55c466e6bb61eb94478b470c/pharma-receptors.jpg)






