Embed Size (px)
Citation preview

Cytochrome Systems Molecular Biology and Bioenergetics Edited by S. Papa U n i v e r s i t y o f B a r i
B a r i , Italy
B. Chance U n i v e r s i t y o f P e n n s y l v a n i a
P h i l a d e l p h i a , P e n n s y l v a n i a
and L. Ernster U n i v e r s i t y o f S t o c k h o l m
S t o c k h o l m , Sweden
Plenum Press • New York and London

Library of Congress Cataloging in Publication Data
U N E S C O International Workshop on Cytochrome Systems: Molecular Biology and Bioenergetics (1987: Bari , Italy) Cytochrome Systems.
"Proceedings of the U N E S C O International Workshop on Cytochrome Systems: Molecular Biology and Bioenergetics, which was IUB Symposium No. 159, held Apr i l 14-18, 1987, in Bari , I ta l y " -Verso t.p.
Includes bibliographical references and index. 1. Cytochrome-Congresses. I. Papa, S. II. Chance, Britton. III. Ernster, L . IV.
Title. QP671.C85U54 1987 574.19'218 87-25766 ISBN 0-306-42693-5
The Symposium was generously supported by the following organizations:
U N E S C O European Expert Committee on Biomaterials International Union of Biochemistry, Committee on Symposia International Union of Pure and Applied Biophysics Consiglio Nazionale delle Ricerche Universitä di Bari Regione Puglia Calabrese Veicoli Industriali S.p.A., Bari Sigma Chemical Company
Proceedings of the U N E S C O International Workshop on Cytochrome Systems: Molecular Biology and Bioenergetics, which was IUB Symposium No. 159, held Apr i l 14-18, 1987, in Bari , Italy
© 1987 Plenum Press, New York A Division of Plenum Publishing Corporation 233 Spring Street, New York, N .Y . 10013
A l l rights reserved
No part of this book may be reproduced, stored in a retrieval system, or transmitted in any form or by any means, electronic, mechanical, photocopying, microfilming, recording, or otherwise, without written permission from the Publisher
Printed in the United States of America

CONTENTS
INTRODUCTION
Cy t o c h r o m e S y s t e m s : f r o m D i s c o v e r y t o P r e s e n t D e v e l o p m e n t s E.C. S l a t e r 3
I . GENETICS
G e n e t i c s o f E . C o l i C y t o c h r o m e s w h i c h a r e Components o f t h e A e r o b i c R e s p i r a t o r y C h a i n
R.B. G e n n i s 15
M o l e c u l a r G e n e t i c A p p r o a c h e s t o S t u d y i n g t h e S t r u c t u r e and F u n c t i o n o f t h e C y t o c h r o m e c^ and t h e C y t o c h r o m e b c ^ Complex f r o m R h o d o b a c t e r C a p s u l a t u s
F. D a l d a l 23
S t r u c t u r a l H o m o l o g i e s i n t h e C y t o c h r o m e B/C^ Complex from R h o d o b a c t e r
N.A. G a b e l l i n i 35
Genes and Sequences f o r S u b u n i t s o f R e s p i r a t o r y Complexes i n P a r a c o c c u s d e n i t r i f i c a n s
B. L u d w i g , B. K u r o w s k i , G. P a n s k u s and P. Steinrücke.. 4 1
O r g a n i z a t i o n and E x p r e s s i o n o f N u c l e a r Genes f o r Y e a s t C y t o c h r o m e c_ O x i d a s e
R.M. W r i g h t , J.D. T r a w i c k , C.E. T r u e b l o o d , T.E. P a t t e r s o n and R.O. P o y t o n 49
The cDNA a S t r u c t u r a l Gene f o r Rat L i v e r C y t o c h r o m e c O x i d a s e S u b u n i t V I c
G. S u s k e , C. E n d e r s , T. Mengel and B. Kadenbach 57
G e n e t i c s o f Y e a s t Coenzyme QH^-Cytochrome c R e d u c t a s e M. C r i v e l l o n e , A. Gampel, I . M u r o f f , M. Wu and A. T z a g o l o f f 67
Sequence A n a l y s i s o f Mammalian Cyt o c h r o m e b M u t a n t s N. H o w e l l and K. G i l b e r t 79
P l a n t M i t o c h o n d r i a l G enes, Cy t o c h r o m e c O x i d a s e and C y t o p l a s m i c M a l e S t e r i l i t y
M.J. H a w k e s f o r d and C . J . L e a v e r 87
vii

A M u t a t i o n A f f e c t i n g L a r i a t F o r m a t i o n , b u t n o t S p l i c i n g o f a Y e a s t M i t o c h o n d r i a l Group I I I n t r o n
R. v a n d e r V e e n , J.H.J.M. Kwakman and L.A. G r i v e l l . . . . 97
The E v o l u t i o n o f M i t o c h o n d r i a l l y Coded C y t o c h r o m e Genes: A Q u a n t i t a t i v e E s t i m a t e
C. S a c c o n e , M. A t t i m o n e l l i , C. L a n a v e , R. G a l l e r a n i and G. P e s o l e 103
I n v o l v e m e n t o f N u c l e a r Genes i n S p l i c i n g o f t h e M i t o c h o n d r i a l COB T r a n s c r i p t i n S. C e r e v i s i a e
J . K r e i k e , G. Krummeck, T. Söllner, C. S c h m i d t and R . J . Schweyen I I I
I n t r o n s as Key E l e m e n t s i n t h e E v o l u t i o n o f M i t o c h o n d r i a l Genomes i n Lower E u k a r y o t e s
K. W o l f , A. Ahne, L. D e l G i u d i c e , G. O r a l e r , F. K anbay, A.M. M e r l o s - L a n g e , F. W e l s e r and M. Zimmer 119
GENETICS: S h o r t R e p o r t s
C o r e P r o t e i n D e f i c i e n c y i n Complex I I I o f t h e R e s p i r a t o r y C h a i n , i n a M i s s e n c e E x o n i c M i t o c h o n d r i a l Y e a s t M u t a n t o f C y t o c h r o m e b Gene
P. C h e v i l l o t t e - B r i v e t , G. S a l o u and D. M e u n i e r - L e m e s l e 13 1
L o c a l i z a t i o n and P a r t i a l S e q u e n c i n g A n a l y s i s o f M i t o c h o n d r i a l Gene f o r A p o c y t o c h r o m e b i n S u n f l o w e r
D. P a c o d a , A.S. T r e g l i a , L. S i c u l e l l a , C. P e r r o t t a and R. G a l l e r a n i 133
M o l e c u l a r B a s i s f o r R e s i s t a n c e t o I n h i b i t o r s o f t h e M i t o c h o n d r i a l U b i q u i n o l - C y t o c h r o m e c R e d u c t a s e i n S a c c h a r m o y c e s C e r e v i s i a e
A.M. C o l s o n , B. M e u n i e r and J . P . d i Rago 135
S t r u c t u r e o f t h e P r o t e i n s and Genes f o r t h e N u c l e a r - E n c o d e d S u b u n i t s o f N. C r a s s a C y tochrome O x i d a s e
M.D. S u a r e z , M. S a c h s , M. D a v i d and U.L. R a j B h a n d a r y . . 137
P a r t i a l Sequence o f Gene f o r C y t o c h r o m e Oxydase S u b u n i t I I o f S u n f l o w e r M i t o c h o n d r i a l DNA
L.R. C e c i , C. De B e n e d e t t o , C M . P e r r o t t a , L. S i c u l e l l a and R. G a l l e r a n i 139
U n u s u a l F e a t u r e s o f K i n e t o p l a s t DNA f r o m T r y p a n o s o m e s : N o v e l M e c h a n i s m o f Gene E x p r e s s i o n ( R N A - E d i t i n g ) , t h e A b s e n c e o f tRNA Genes and t h e V a r i a b l e P r e s e n c e o f a C y t o c h r o m e O x i d a s e S u b u n i t Gene
P. S l o o f , R. Benne, B. De V r i e s , T. H a k v o o r t and A. M u i j s e r s 14 1

I I . BIOSYNTHESIS
H e t e r o g e n e o u s E f f i c i e n c i e s o f mRNA T r a n s l a t i o n i n Human M i t o c h o n d r i a
A. Chomyn and G. A t t a r d i 145
R e g u l a t i o n o f t h e E x p r e s s i o n o f COI and C O I I I mRNAs i n Rat L i v e r M i t o c h o n d r i a
P. C a n t a t o r e , F. F r a c a s s o , A.M.S. L e z z a and M.N. G a d a l e t a 153
Two C o n s e n s u s Sequences f o r RNA P r o c e s s i n g i n N e u r o s p o r a C r a s s a M i t o c h o n d r i a
G. M a c i n o and M.A. N e l s o n 161
P r i m a r y S t r u c t u r e o f t h e Y e a s t N u c l e a r CBS 1 Gene P r o d u c t , N e c e s s a r y t o A c t i v a t e T r a n s l a t i o n o f M i t o c h o n d r i a l COB mRNA
V. F o r s b a c h and G. Rödel 169
A C a l m o d u l i n - L i k e P r o t e i n i n t h e Cy t o c h r o m e bc Complex R e q u i r e d f o r S y n t h e s i s o f B o t h C y t o c h r o m e bc and Cy t o c h r o m e c O x i d a s e Complexes i n Y e a s t M i t o c h o n d r i a
M.E . S c h m i t t and B.L. Trumpower 177
Im p o r t o f C y t o c h r o m e s b^ and c^ i n t o M i t o c h o n d r i a i s Dependent on B o t h Membrane P o t e n t i a l and N u c l e o s i d e T r i p h o s p h a t e s
F.U. H a r t l , J . O s t e r m a n n , N. P f a n n e r , M. T r o p s c h u g , B. G u i a r d and W. N e u p e r t 189
NADH: A Common R e q u i r e m e n t f o r t h e Import and M a t u r a t i o n o f C y t o c h r o m e s c and c ^
D.W. N i c h o l s o n , J . Ost e r m a n n and W. N e u p e r t 197
Imp o r t o f P r o t e i n s i n t o M i t o c h o n d r i a : S t r u c t u r a l and F u n c t i o n a l R o l e o f t h e P r e p e p t i d e
D. R o i s e 209
I n t e r a c t i o n o f t h e M i t o c h o n d r i a l P r e c u r s o r P r o t e i n A p o c y -t o c h r o m e c w i t h M o d e l Membranes and i t s I m p l i c a t i o n s f o r P r o t e i n T r a n s l o c a t i o n
B. de K r u i j f f , A. R i e t v e l d , W. J o r d i , T.A. B e r k h o u t , R.A. Demel, H. Görrissen and D. Mars h 2 15
I n f l u e n c e o f T h y r o i d Hormones on t h e E x p r e s s i o n o f N u c l e a r Genes E n c o d i n g S u b u n i t s o f t h e Cy t o c h r o m e bc Complex
V. J o s t e and B.D. N e l s o n 225
I I . BIOSYNTHESIS: S h o r t R e p o r t
I m p o r t o f C y t o c h r o m e c and Cy t o c h r o m e b^ i n t o M i t o c h o n d r i a J . O s t e r m a n n , F.U. H a r t l , M. T r o p s c h u n g , B. G u i a r d and W. N e u p e r t 235
ix

I I I . PROTEIN STRUCTURE
Cyt o c h r o m e c O x i d a s e : P a s t , P r e s e n t and F u t u r e M. Müller, N. L a b o n i a , B. Schläpfer and A. A z z i 23^
U n i v e r s a l F e a t u r e s i n C y t o c h r o m e O x i d a s e M. F i n e l , T. H a l t i a , L. Holm, T. J a l l i , T. M e t s o , A. P u u s t i n e n , M. R a i t i o , M. S a r a s t e and M. Wikström 24"
P o l y p e p t i d e S u b u n i t s Encoded by N u c l e a r Genes a r e E s s e n t i a l Components o f E u k a r y o t i c C y t o c h r o m e c_ O x i d a s e
T.E. P a t t e r s o n , C.E. T r u e b l o o d , R.M. W r i g h t and R.O. P o y t o n 25)
R e s p i r a t o r y Complex IV and C y t o c h r o m e a , a ^ G. B u s e , G.C.M. S t e f f e n s , R. B i e w a l d , B. B r u c h and S. H e n s e l 26
An Amino A c i d Sequence R e g i o n o f S u b u n i t I I o f C y t o c h r o m e O x i d a s e w h i c h may be R e s p o n s i b l e f o r E v o l u t i o n a r y Changes i n R e a c t i v i t y w i t h D i f f e r e n t C y t o c h r o m e s c
T.L. L u n t z and E. M a r g o l i a s h 27
E v i d e n c e f o r a F u n c t i o n a l R o l e o f N u c l e a r E n c o d e d S u b u n i t s i n M i t o c h o n d r i a l C y t o c h r o m e c O x i d a s e
R. B i s s o n and B. B a c c i 28!
S t r u c t u r a l A n a l y s i s o f t h e bc^ Complex f r o m B e e f H e a r t M i t o c h o n d r i a by t h e S i d e d H y d r o p a t h y P l o t and by C o m p a r i s o n w i t h O t h e r bc ^ C omplexes
T.A. L i n k , H. Schägger and G. v o n Jagow 289
C r y s t a l l i s a t i o n o f W a t e r - S o l u b l e P r e p a r a t i o n s o f t h e I r o n -S u l f u r - P r o t e i n and C y t o c h r o m e c^ o f U b i q u i n o l : C y t o c h r o m e R e d u c t a s e f r o m N e u r o s p o r a M i t o c h o n d r i a
J . Römisch, U. H a r n i s c h , U. S c h u l t e and H. W e i s s 303
I r o n C l u s t e r S i t e s o f C a r d i a c I p - S u b u n i t o f S u c c i n a t e D e h y d r o g e n a s e
T.E. K i n g , N.S. R e i m e r , M.T. Seaman, L.Q. Sun, Q.W. Wang, K.T. Y a s u n o b u and S.H. Ho 309
P o l y p e p t i d e s i n M i t o c h o n d r i a l E l e c t r o n - T r a n s f e r C o m p l e x e s T. Ozawa, M. N i s h i k i m i , H. S u z u k i , M. T a n a k a and Y. Shimomura 315
The F a t t y A c i d - A n c h o r e d F o u r Herne C y t o c h r o m e o f t h e P h o t o -s y n t h e t i c R e a c t i o n C e n t e r f r o m t h e P u r p l e B a c t e r i u m Rhodopseudomonas V i r i d i s
K.A. Weyer, F. L o t t s p e i c h , W. Schäfer and H. M i c h e l 325
x

I I I . PROTEIN STRUCTURE: S h o r t R e p o r t s
Membrane P r o t e i n i n P a r a c o c c u s D e n i t r i f i c a n s R e s e m b l e s S u b u n i t I I I o f B e e f H e a r t C y t o c h r o m e O x i d a s e
T. H a l t i a and M. F i n e l 335
The C i r c u l a r D i c h r o i s m P r o p e r t i e s o f t h e R i e s k e P r o t e i n and t h e b and c C y t o c h r o m e s o f t h e M i t o c h o n d r i a l b c | Complex
G. S o l a i n i , M. C r i m i , F. B a l l e s t e r , M. D e g l i E s p o s t i and G. L e n a z 337
M o n o m e r i z a t i o n o f C y t o c h r o m e O x i d a s e M i g h t Be E s s e n t i a l f o r S u b u n i t I I I Removal
M. F i n e l 339
F o u r i e r - T r a n s f o r m I n f r a - R e d S t u d i e s o f C y t o c h r o m e c O x i d a s e M.F. G r a h n , P . I . K a r i s , J.M. W r i g g l e s w o r t h and D. Chapman 34 1
I s o e n z y m e s o f Human C y t o c h r o m e c O x i d a s e T.B.M. H a k v o o r t , G.J.C. R u y t e r , K.M.C. S i n j o r g o and A.O. M u i j s e r s 343
F o l d i n g o f I n t e g r a l Membrane P r o t e i n s : R e n a t u r a t i o n E x p e r i ments w i t h B a c t e r i o r h o d o s p i n S u p p o r t a Two-Stage M e c h a n i s m
J . L . P o p o t and D.M. Engelman 345
I V . REACTION DOMAINS AND OXIDO-REDUCTION MECHANISMS
The N a t u r e o f F e r r y l , F e ( l V ) = 0 , Haem i n P e r o x i d a s e s A. J . Thomson, C. Greenwood, P.M.A. Gadsby and N. F o o t e 349
P o l a r i z a t i o n i n Herne P r o t e i n s and Cyto c h r o m e c O x i d a s e J.O. A l b e n 36 1
E f f e c t s o f F r e e z i n g on t h e C o o r d i n a t i o n S t a t e and L i g a n d O r i e n t a t i o n i n H e m o p r o t e i n s
H. A n n i and T. Y o n e t a n i 37 1
C a t a l y t i c M e c h a n i s m o f 0^ R e d u c t i o n by C y t o c h r o m e O x i d a s e M. Wikström 377
The E l e c t r o n - T r a n s f e r R e a c t i o n s i n and S u b s t r a t e B i n d i n g t o C y t o c h r o m e c_ O x i d a s e
B. F. v a n G e l d e r , A.C.F. G o r r e n , L. V l e g e l s and R. Wever 385
C o n t r o l o f C y t o c h r o m e O x i d a s e i n P r o t e o l i p o s o m e s : F l u x and S t o i c h i o m e t r y
P. N i c h o l l s , S. S h a u g h n e s s y and A.P. S i n g h 391
Cy t o c h r o m e c O x i d a s e A c t i v i t y i s R e g u l a t e d by N u c l e o t i d e s and A n i o n s
B. K a d e n b a c h , A. S t r o h , F . J . Hüther and J . B e r d e n . . . . 399
xi

Cytochrome O x i d a s e and N e u r o m u s c u l a r D i s e a s e s C.P. L e e , M.E. M a r t e n s , P.L. P e t e r s o n and J. S . H a t f i e l d 407
E l e c t r o n T r a n s f e r Between C a r d i a c C y t o c h r o m e c^ and c C.H. K i m , C. B a l n y and T.E. K i n g 415
Herne C l u s t e r S t r u c t u r e s and E l e c t r o n T r a n s f e r i n M u l t i h e m e C y t o c h r o m e s C^
R. H a s e r and J . Mosse 423
IV. REACTION DOMAINS AND OXIDO-REDUCTION MECHANISMS: S h o r t R e p o r t s
E l e c t r o n Exchange Between C y t o c h r o m e s a. P. S a r t i , F. M a l a t e s t a , G. A n t o n i n i , M.T. W i l s o n and M. B r u n o r i 433
P u l s e d , R e s t i n g and P e r o x y Forms o f C y t o c h r o m e O x i d a s e G. A n t o n i n i , F. M a l a t e s t a , P. S a r t i , M.T. W i l s o n and M. B r u n o r i 435
E f f e c t o f pH and T e m p e r a t u r e on E l e c t r o n T r a n s f e r i n M i x e d -V a l e n c e C y t o c h r o m e c O x i d a s e
P. B r e z e z i n s k i and Bo.G. Malmström 437
The pH-Dependence o f t h e R e d u c t i o n a l L e v e l s o f C y t o c h r o m e c and C y t o c h r o m e a Düring T u r n o v e r o f C y t o c h r o m e c O x i d a s e
P.E. Thörnstrom, B. M a i s o n - P e t e r i , P. B r z e z i n s k i , L. A r v i d s s o n , P.O. F r e d r i k s s o n and Bo.G. Malmström.... 439
V. Fe-S CENTERS
S t u d i e s o f B a c t e r i a l NADH-Ubiquinone ( o r M e n a q u i n o n e ) O x i d o r e d u c t a s e Systems
T. O h n i s h i , S.W. M e i n h a r d t and K. M a t s u s h i t a 443
L o c a l i z e d V a l e n c e S t a t e s i n I r o n - S u l f u r C l u s t e r s and T h e i r P o s s i b l e R e l a t i o n s h i p t o t h e A b i l i t y o f I r o n - S u l f u r C l u s t e r s t o C a t a l y z e R e a c t i o n s O t h e r Than E l e c t r o n T r a n s f e r
H. B e i n e r t 451
P r o b i n g t h e Fe/S Domain W i t h EPR: P a n d o r a ' s BOX A j a r W.R. Hagen 459
A s p e c t s o f S p i n C o u p l i n g Between Even and Odd E l e c t r o n S y s t e m s : A p p l i c a t i o n s t o S u c c i n a t e : Q R e d u c t a s e
J.C. S a l e r n o and Xu Yan 467
The I r o n - S u l f u r C l u s t e r s i n S u c c i n a t e D e h y d r o g e n a s e M.K. J o h n s o n , J . E . M o r n i n g s t a r , E.B. K e a r n e y , G. C e c c h i n i and B.A.C. A c k r e l l 473
Mechanisms o f E l e c t r o n T r a n s f e r i n S u c c i n a t e D e h y d r o g e n a s e and F u m a r a t e R e d u c t a s e : P o s s i b l e F u n c t i o n s f o r I r o n - S u l p h u r C e n t r e 2 and C y t o c h r o m e 1}
R. Cammack, J . M a g u i r e and B.A.C. A c k r e l l 485
xii

V I . QUINONE BINDING SITES:STRUCTURE AND FUNCTION
S t r u c t u r a l and M e c h a n i s t i c A s p e c t s o f t h e Q u inone B i n d i n g S i t e s o f t h e bc Complexes
P.R. R i e h 495
QP-S-The E l e c t r o n A c c e p t o r o f S u c c i n a t e D e h y d r o g e n a s e T.E. K i n g and Y. Xu 503
C h a r a c t e r i s t i c s o f QP-C and R e c o n s t i t u t i o n o f t h e QH^-Cyt c r e d u c t a s e
T.Y. Wang, Z.P. Zhang and T.E. K i n g 509
The K i n e t i c s o f O x i d a t i o n o f C y t o c h r o m e b a r e i n Agreement w i t h t h e Q - C y c l e H y p o t h e s i s
S. De V r i e s , A.N. v a n Hoek, A. t e n Bookum and J.A. B e r d e n 517
The Q - R e a c t i o n Domains o f M i t o c h o n d r i a l QH^' C y t o c h r o m e c_ O x i d o r e d u c t a s e
J.A. B e r d e n , A.N. v a n Hoek, S. de V r i e s and P . J . S c h o p p i n k 523
EPR S t u d i e s o f t h e Q u i n o n e R e a c t i o n S i t e s i n B a c t e r i a S.W. M e i n h a r d t , X. Y a n g , B.L. Trumpower and T. O h n i s h i 533
EPR S i g n a l I I i n P h o t o s y s t e m I I : Redox and P a r a m a g n e t i c I n t e r a c t i o n s w i t h t h e 0^ E v o l v i n g Enzyme
A.W. R u t h e r f o r d and S. S t y r i n g 541
S t u d i e s on t h e U b i q u i n o n e P o o l i n M i t o c h o n d r i a l E l e c t r o n T r a n s f e r
G. L e n a z , R. F a t o and G. P a r e n t i C a s t e l l i 549
V I . QUINONE BINDING S I T E S : STRUCTURE AND FUNCTION: S h o r t R e p o r t s
E f f e c t s o f P r o t e o l y t i c D i g e s t i o n and C h e m i c a l M o d i f i c a t i o n o f b-c C o m p l e x on t h e Fe-S C e n t e r and t h e S t a b l e U b i s e m i -q u i n o n e (SQc)
T. C o c c o , S. M e i n h a r d t , D. G a t t i , M. L o r u s s o , T. O h n i s h i and S. Papa 559
K i n e t i c s o f U b i q u i n o l C y t o c h r o m e c R e d u c t a s e : L a c k o f a D i f f u s i o n - L i m i t e d S t e p w i t h S h o r t C h a i n U b i q u i n o l s as S u b s t r a t e s
R. F a t o , C. C a s t e l l u c c i o , G. L e n a z , M. B a t t i n o and G. P a r e n t i C a s t e l l i 561
V I I . S P E C I F I C REDOX SYSTEMS
B i o c h e m i s t r y and M o l e c u l a r B i o l o g y o f t h e E s c h e r i c h i a C o l i A e r o b i c R e s p i r a t o r y C h a i n
Y. A n r a k u 565
xiii

The I n t e r p l a y Between P h o t o s y n t h e s i s and R e s p i r a t i o n i n F a c u l t a t i v e A n o x y g e n i c P h o t o t r o p h i c B a c t e r i a
D. Z a n n o n i 575
The U b i q u i n o l - C y t o c h r o m e c O x i d o r e d u c t a s e o f P h o t o t r o p h i c B a c t e r i a
M. S n o z z i and J . Kälin 585
The Redox R e a c t i o n Between C y t o c h r o m e and t h e S e m i q u i -n o n e - Q u i n o l C o u p l e o f Q i s E l e c t r o g e n i c i n U b i q u i n o l -
c Cyt c^ O x i d o r e d u c t a s e
D.E. R o b e r t s o n and P.L. D u t t o n 593
C o n s t r u c t i o n and C h a r a c t e r i z a t i o n o f M o n o l a y e r F i l m s o f t h e R e a c t i o n C e n t e r Cytochrome-c_ P r o t e i n f r o m Rhodopseudomonas V i r i d i s
G. A l e g r i a and P.L. D u t t o n
S t r u c t u r e - F u n c t i o n R e l a t i o n s h i p s i n t h e Rhodopseudomonas V i r i d i s R e a c t i o n C e n t e r Complex: E l e c t r o g e n i c S t e p s C o n t r i -b u t i n g t o F o r m a t i o n
V.P. S k u l a c h e v , L.A. D r a c h e v , S.M. D r a c h e v a , A.A. K o n s t a n t i n o v , A.Yu. Semenov and V.A. S h u v a l o v . .
C a t a l y t i c S i t e s f o r R e d u c t i o n and O x i d a t i o n o f Q u i n o n e s A. C r o f t s , H. R o b i n s o n , K. An d r e w s , S. v a n Dören and Ed B e r r y 617
S t u d i e s o f t h e E l e c t r o g e n i c i t y o f t h e R e d u c t i o n o f C y t o chrome b t h r o u g h t h e A n t i m y c i n - S e n s i t i v e S i t e o f t h e U b i q u i n o 5 - ( i y t o c h r o m e c ^ O x i d o r e d u c t a s e Complex o f R h o d o b a c t e r S p h a e r o i d e s
E. G l a s e r and A.R. C r o f t s 625
M u t a n t s A f f e c t i n g t h e Q u i n o l O x i d a t i o n S i t e o f U b i q u i n o l -Cyt c ^ - O x i d o r e d u c t a s e
D.E. R o b e r t s o n , F. D a l d a l and P.L. D u t t o n 633
Cy t o c h r o m e o f B a c i l l u s S u b t i l i s H. F r i d e n and L. H e d e r s t e d t 641
M i t o c h o n d r i a l C y t o c h r o m e b C A . Yu and L. Yu 649
I s o l a t i o n o f t h e P h o t o s y s t e m Two R e a c t i o n C e n t r e and t h e l o c a t i o n and F u n c t i o n o f Cy t o c h r o m e D ^ . ^
J . B a r b e r , K. G o u n a r i s and D.J. Chapman 657
V I I . S P E C I F I C REDOX SYSTEMS: S h o r t R e p o r t s
E f f e c t o f A n t i m y c i n on t h e K i n e t i c s o f R e d u c t i o n o f b and c j C y t o c h r o m e s i n S i n g l e T u r n o v e r o f t h e M i t o c h o n d r i a l b - c ^ Complex
D. B o f f o l i , M. L o r u s s o and S. Papa 669
The b-Q C y c l e , a new model f o r t h e Pathways o f E l e c t r o n s and P r o t o n s t h r o u g h t h e b c ^ Complex! A c t i o n o f A n t i m y c i n A at C e n t e r 0
D. M e u n i e r - L e m e s l e and P. C h e v i l l o t t e - B r i v e t 673
xiv
601
. 609

R a p i d O x i d a t i o n K i n e t i c s o f Q u i n o l C y t o c h r o m e c O x i d o r e d u c t a s e : Q u i n o n e o r S e m i q u i n o n e C y c l e ?
A.N. v a n Hoek, J.A. B e r d e n and S. De V r i e s 675
E l e c t r o n C o n d u c t i o n Between C y t o c h r o m e s o f t h e M i t o c h o n d r i a l R e s p i r a t o r y C h a i n i n t h e P r e s e n c e o f A n t i m y c i n p l u s M y x o t h i a z o l
I.C. West 679
The R e l e a s e o f NAD* R a d i c a l i s t h e L i m i t i n g S t e p i n t h e Redox R e a c t i o n Between F e ( I I l ) C y t o c h r o m e c and NADH
A. F e r r i , P. C h i o z z i , M.E. C a t t o z z o and D. P a t t i 681
P r o p e r t i e s o f C y t o c h r o m e b i n Complex I I o f A s c a r i s 558 . .
M u s c l e M i t o c h o n d r i a and l t s F u n c t i o n i n A n a e r o b i c R e s p i r a t i o n
K. K i t a , S. T a k a m i y a , R. F u r u s h i m a , Y. Ma and H. Oya 683
V I I I . COOPERATIVITY AND ALLOSTERIC TRANSITIONS
P u l s e d and R e s t i n g C y t o c h r o m e O x i d a s e : an U p d a t e o f O p t i c a l and M o l e c u l a r P r o p e r t i e s
M. B r u n o r i , P. S a r t i , F. M a l a t e s t a , G. A n t o n i n i and M.T. W i l s o n 689
I s H O I n v o l v e d i n E l e c t r o n T r a n s p o r t Between C y t o c h r o m e s a_ and a_^ i n C y t o c h r o m e O x i d a s e ?
J.M. W r i g g l e s w o r t h , M.F. G r a h n , J . E l s d e n and H. Baum 697
C o n f o r m a t i o n a l T r a n s i t i o n s o f C y t o c h r o m e c_ O x i d a s e I n d u c e d by P a r t i a l R e d u c t i o n
C.P. S c h o l e s , C. F a n , J . Bank, R. D o r r and Bo.G. Malmström 705
R e d o x - L i n k e d C o n f o r m a t i o n a l Changes i n C y t o c h r o m e £ O x i d a s e T. A l l e y n e and M.T. W i l s o n 713
I X . PROTON-MOTIVE ACTIVITY
P r o t o n m o t i v e A c t i v i t y o f t h e C y t o c h r o m e C h a i n o f M i t o c h o n d r i a : M o d e l s and F e a t u r e s
S. Papa and N. C a p i t a n i o 723
The M e c h a n i s m o f E l e c t r o n G a t i n g i n C y t o c h r o m e c O x i d a s e Bo.G. Malmström 733
P r o t o n Pump A c t i v i t y i n B a c t e r i a l C y t o c h r o m e O x i d a s e s N. Sone 743
T o p o g r a p h y and P r o t o n m o t i v e M e c h a n i s m o f M i t o c h o n d r i a l C o u p l i n g S i t e 2
A.A. K o n s t a n t i n o v and E. Popova 751
xv

The Q-Gated P r o t o n Pump Model o f U b i q u i n o l C y t o c h r o m e c R e d u c t a s e : E l e c t r o n and P r o t o n T r a n s f e r P a t h ways and R o l e o f P o l y p e p t i d e s
M. L o r u s s o , D. B o f f o l i , T. C o c c o , D. G a t t i and S. Papa 767
P h o t o s y n t h e t i c C o n t r o l and A T P / E l e c t r o n R a t i o i n B a c t e r i a l P h o t o p h o s p h o r y l a t i o n
V. Adam, M. V i r g i l i , G. V e n t u r o l i , D. P i e t r o b o n and B.A. M e l a n d r i 777
I X . PROTON-MOTIVE ACTIVITY: S h o r t R e p o r t
P r o t o n m o t i v e A c t i v i t y o f M i t o c h o n d r i a l C y t o c h r o m e c O x i d a s e
N. C a p i t a n i o , E. De N i t t o and S. Papa 787
APPENDIX
P r o p o s a l f o r a N o v e l N o m e n c l a t u r e f o r t h e S u b u n i t s o f Cyto c h r o m e c O x i d a s e
G. B u s e , G.C.M. S t e f f e n s , R. B i e w a l d , B. B r u c h and S. H e n s e l 791
A u t h o r I n d e x 795
S u b j e c t I n d e x 799
xvi

NADH: A COMMON REQUIREMENT FOR THE IMPORT
AND MATÜRATION OF CYTOCHROMES c AND c,
D o n a l d W. N i c h o l s o n , J o a c h i m O s termann and W a l t e r N e u p e r t
I n s t i t u t für P h y s i o l o g i s c h e Chemie d e r Universität München G o e t h e s t r a s s e 33 D-8000 München 2 F e d e r a l R e p u b l i c o f Germany
SUMMARY
The c o v a l e n t a t t a c h m e n t o f heme t o a p o c y t o c h r o m e c, w h i c h i s c a t a l y z e d by t h e m i t o c h o n d r i a l enzyme c y t o c h r o m e c heme l y a s e , was d e p e n d e n t on NADH. I n a d d i t i o n , a c o f a c t o r p r e s e n t i n r e t i c u l o c y t e l y s a t e o r a N e u r o s p o r a c r a s s a c y t o s o l f r a c t i o n was r e q u i r e d f o r t h e NADH-dependent s t e p . I n t h e a b s e n c e o f NADH, ap o c y t o c h r o m e c was bound t o t h e m i t o c h o n d r i a l s u r f a c e and r e m a i n e d a c c e s s i b l e t o e x t e r n a l l y added p r o t e a s e s . I n t h e p r e s e n c e o f NADH, c o v a l e n t a t t a c h m e n t o f heme o c c u r r e d w i t h c o n c o m i t a n t t r a n s l o c a t i o n o f c y t o c h r o m e c a c r o s s t h e o u t e r m i t o c h o n d r i a l membrane t o a p r o t e a s e - r e s i s t a n t l o c a t i o n . B o t h heme a t t a c h m e n t and t r a n s l o c a t i o n were i n h i b i t e d by t h e heme a n a l o g u e d e u t e r o h e m i n .
The s e c o n d p r o t e o l y t i c - p r o c e s s i n g s t e p d u r i n g c y t o c h r o m e c. i m p o r t , f r o m t h e i n t e r m e d i a t e t o m a t u r e - s i z e p r o t e i n , was a l s o d e p e n d e n t on NADH and c o u l d a l s o be i n h i b i t e d by d e u t e r o h e m i n . We s u g g e s t t h a t t h i s o c c u r s as a r e s u l t o f c o n d i t i o n s w h i c h äffeet t h e h e m e - a t t a c h i n g r e a c t i o n f o r c y t o c h r o m e c^ and t h a t heme a t t a c h m e n t must p r e c e d e t h e s e c o n d p r o c e s s i n g s t e p . M o d e l s a r e p r e s e n t e d t o a e c o u n t f o r t h e s e o b s e r v a t i o n s .
INTRODUCTION
The m a j o r i t y o f m i t o c h o n d r i a l p r o t e i n s a r e encoded by n u c l e a r genes and a r e s y n t h e s i z e d as p r e c u r s o r s on f r e e r i b o s o m e s i n t h e c e l l c y t o s o l . They a r e t h e n i m p o r t e d i n t o m i t o c h o n d r i a a l o n g p a t h w a y s w h i c h c a n be s u b d i v i d e d i n t o a c o n s e c u t i v e s e r i e s o f d i s t i n e t s t e p s ( f o r r e v i e w see 1 - 4 ) . Most m i t o c h o n d r i a l p r o t e i n s , p a r t i c u l a r l y t h o s e w h i c h must be d i r e c t e d t o t h e i n n e r membrane o r m a t r i x , a r e i m p o r t e d by t h e f o l l o w i n g s e q u e n c e o f e v e n t s : i ) s y n t h e s i s i n t h e c y t o s o l a s a p r e c u r s o r c o n t a i n i n g an N - t e r m i n a l t a r g e t i n g s e q u e n c e ; i i ) b i n d i n g t o s p e c i f i c r e e e p t o r s on t h e m i t o c h o n d r i a l s u r f a c e ; i i i ) m e m b r a n e - p o t e n t i a l - d e p e n d e n t i n s e r t i o n i n t o t h e i n n e r membrane v i a t r a n s l o c a t i o n c o n t a c t s i t e s where t h e i n n e r and o u t e r membrane come c l o s e enough t o g e t h e r t o be spanned s i m u l t a n e o u s l y by t h e i m p o r t e d p r o t e i n ; i v ) r e m o v a l o f t h e N - t e r m i n a l p r e p i e c e by t h e c h e l a t o r - s e n s i t i v e p r o c e s s i n g p e p t i d a s e l o c a t e d i n t h e m a t r i x ; v ) s o r t i n g o f t h e p r o t e i n t o i t s f i n a l s u b m i t o c h o n d r i a l l o c a t i o n ; and v i ) a s s e m b l y i n t o f u n c t i o n a l c o m p l e x e s . I n a number o f c a s e s , t h e i m p o r t pathway i s a e c o m p a n i e d by c o v a l e n t m o d i f i c a t i o n
197

or the a c q u i s i t i o n of n o n - c o v a l e n t l y bound p r o s t h e t i c groups.
Energy i s r e q u i r e d f o r p r o t e i n import i n three known forms. F i r s t , i t has been w e l l e s t a b l i s h e d t h a t a l l p r o t e i n s which must be t r a n s l o c a t e d i n t o or across the in n e r membrane r e q u i r e the presence of a membrane p o t e n t i a l (5,6). Recently, i t has a l s o been demonstrated t h a t ATP i s r e q u i r e d f o r the maintenence of an import-competent conformation d u r i n g a l l steps which precede and i n c l u d e i n t e r a c t i o n of the precursor p r o t e i n w i t h the outer membrane (7, see accompanying a r t i c l e by H a r t l et a l ) . F i n a l l y , NAD(P)H i s r e q u i r e d f o r the maturation of cytochromes c (8,9) and c^ (10,11), f o r the Fe/S p r o t e i n of the bc^ complex ( H a r t l and Neupert, unp u b l i s h e d ) , and f o r cytochrome oxidase subunit I I ( D r i e v e r , Cook and Neupert, u n p u b l i s h e d ) .
The requirement f o r NADH by cytochromes c and c^ appears to be a common fea t u r e of the l a t e r stages of t h e i r r e s p e c t i v e import pathways. Despite a number of s i m i l a r i t i e s between the two p r o t e i n s (e.g. both mature p r o t e i n s are exposed to the m i t o c h o n d r i a l intermembrane space; they are the only two m i t o c h o n d r i a l p r o t e i n s to which heme i s covalently«bound; they both r e q u i r e NADH du r i n g import) , cytochromes c arid c^ f o l l o w markedly d i f f e r e n t import pathways. Cytochrome c i s s y n t h e s i z e d i n the c y t o s o l as apocytochrome c, which d i f f e r s from i t s mature counterpart i n conformation and by the absence of covalently-bound heme. I t does not c o n t a i n a c l e a v a b l e N-terminal prepiece and does not r e q u i r e a membrane p o t e n t i a l f o r import (12-18). Apocytochrome c i s t r a n s f e r r e d from a cytoplasmic pool to s p e c i f i c r e c e p t o r s a t the outer m i t o c h o n d r i a l membrane (18-21). Heme i s attached to the apopr o t e i n , by the enzyme cytochrome c heme l y a s e (8,9,20,22-24.), i n a step which i s coupled to the t r a n s l o c a t i o n of cytochrome c across the outer membrane to i t s f i n a l l o c a t i o n i n the intermembrane space. Cytochrome c^, on the other hand, f o l l o w s a somewhat more complicated sequence of events d u r i n g import. The precursor of cytochrome i s synthesized i n the c y t o s o l and contains an un u s u a l l y long N-terminal prepiece which i s processed i n two steps (10,11,25,26). The precursor binds to rec e p t o r s a t the outer membrane and the N-terminus i s then t r a n s l o c a t e d through the in n e r membrane v i a contact s i t e s (11) i n a step which i s dependent on a membrane p o t e n t i a l . The h y d r o p h i l i c f i r s t h a l f of the prepiece i s removed by the mat r i x peptidase f o l l o w e d by removal of the second p a r t by another p r o c e s s i n g a c t i v i t y which i s presumed to be l o c a t e d i n the intermembrane space. The l a t t e r step i s dependent on heme (25,26) and t h e r e f o r e might be coupled to the heme a t t a c h i n g r e a c t i o n or might have to be preceded by heme l i n k a g e . The mature p r o t e i n i s then assembled i n t o the bc^ complex, anchored to the in n e r membrane by a hydrophobic s t r e t c h of amino aci d s a t the C-terminus (27,28).
In t h i s r e p o r t we have examined the common requirement f o r NADH du r i n g the import of cytochromes c and c^ i n t o mitochondria. We demonstrate t h a t NADH i s r e q u i r e d f o r the co v a l e n t attachment of heme to apocytochrome c and t h a t the NADH-dependent step i s mediated by a c y t o s o l i c c o f a c t o r . Since the second pr o c e s s i n g step of cytochrome c. has n e a r l y i d e n t i c a l requirements, we suggest t h a t NADH i s needed f o r a s i m i l a r import event; namely, f o r the a c t i v i t y of cytochrome c 1 heme l y a s e .
MATERIALS AND METHODS
Ce l l Growth and Subcellular Fractionation
Neurospora c r a s s a w i l d type 74A was grown f o r U h at 25°C as p r e v i o u s l y d e s c r i b e d (29). M i t o c h o n d r i a were i s o l a t e d from f r e s h l y harvested hyphae by P e r c o l l - g r a d i e n t c e n t r i f u g a t i o n (Pharmacia) and suspended i n b u f f e r A (250 mM sucrose, 10 mM Mops/KOH (pH 7.2), 2 mM EDTA, 1 mM phenylmethylsulphonyl f l u o r i d e (PMSF)) at a p r o t e i n c o n c e n t r a t i o n of 5 mg/ml (8 ) . PMSF was not in c l u d e d i n b u f f e r A d u r i n g any of the f r a c t i o n a t i o n or import steps i f
198

P r o t e i n a s e K d i g e s t i o n was i n c l u d e d i n the experimental procedure. A c y t o s o l f r a c t i o n was prepared by g r i n d i n g hyphae w i t h quartz sand i n b u f f e r A and then p r e p a r i n g a post-ribosomal supernatant ( 8 ) .
Cell-Free Protein Synthesis
P r e c u r s o r p r o t e i n s of N. crassa cytochromes c and c^ were synthesized i n r a b b i t r e t i c u l o c y t e l y s a t e s ( 3 0 ) i n the presence of L-( S ) c y s t e i n e as p r e v i o u s l y d e s c r i b e d ( 8 ) . Synthesis of apocytochrome c was d i r e c t e d by N. crassa p o l y ( A ) - c o n t a i n i n g RNA ( 3 1 ) . Synthesis of the precu r s o r to cytochrome
was d i r e c t e d by a f u l l - l e n g t h cDNA clone of pre-cytochrome c^ i n pGEM-4 th a t was t r a n s c r i b e d w i t h SP6 RNA-Polymerase (Ostermann and Neupert, unpubl i s h e d ) .
Import of Cytochrome c
1. D i r e c t import: M i t o c h o n d r i a (75 |ig p r o t e i n ) were incubated f o r 10 min at 25 C i n a t o t a l volume of 2 0 0 ^1 b u f f e r A i n the presence of 50 i i l ( S ) c y s t e i n e - l a b e l l e d r e t i c u l o c y t e l y s a t e plus other additions' as i n d i c a t e d . Holocytochrome c formation was determined by r e - i s o l a t i n g the mitochondria by c e n t r i f u g a t i o n , immunoprecipitating t o t a l cytochrome c (holo plus apo), d i s s o c i a t i o n of the immunocomplexes w i t h urea, d i g e s t i o n w i t h t r y p s i n , and a n a l y s i s of the r e s u l t i n g peptides by reverse-phase HPLC ( 8 ) . The holocytochrome c c y s t e i n e - c o n t a i n i n g t r y p t i c peptide was c o l l e c t e d and i t s r a d i o a c t i v i t y determined as a measure of holocytochrome c formation.
2 . Chase of pre-bound apocytochrome c to holocytochrome c: Apocytochrome c was bound to mitochondria i n the absence of conversion to holocytochrome c (by w i t h h o l d i n g NADH) by i n c u b a t i n g mitochondria ( 7 5 iig p r o t e i n ) w i t h 50 \il ( S ) c y s t e i n e - l a b e l l e d r e t i c u l o c y t e l y s a t e i n a t o t a l volume of 2 0 0 i i l b u f f e r A f o r 10 min at 25 C. The mitochondria were r e -i s o l a t e d by c e n t r i f u g a t i o n at 17 ,4-00 xg f o r 12 min and resuspended i n f r e s h b u f f e r A. The bound apocytochrome c was then chased to holocytochrome c during a second i n c u b a t i o n f o r 10 min at 25 C i n a mixture c o n t a i n i n g 75 ng m i t o c h o n d r i a l p r o t e i n , plus a d d i t i o n s as i n d i c a t e d , i n 2 0 0 \il b u f f e r A. Holocytochrome c formation was then determined as de s c r i b e d above.
Import of Cytochrome
1. D i r e c t import: M i t o c h o n d r i a ( 5 0 |ig) were incubated f o r 30 min at 25 C i n a mixture ( t o t a l volume 100 nl) c o n t a i n i n g 3% (w/v) bovine serum albumin, 70,-mM K C l , 2 2 0 mM sucrose, 10 mM Mops/KOH (pH 7 . 2 ) i n the presence of 20 j i l ( S ) c y s t e i n e - l a b e l l e d r e t i c u l o c y t e plus other a d d i t i o n s as i n d i c a t e d . The sample's were cooled to 0 C and P r o t e i n a s e K was added to a f i n a l c o n c e n t r a t i o n of 20 ug/ml. F o l l o w i n g i n c u b a t i o n f o r 30 min at 0 C, the mitochondria were r e - i s o l a t e d as above and d i s s o c i a t e d i n SDS sample b u f f e r . The samples were then r e s o l v e d by SDS Polyacrylamide g e l e l e c t r o p h o r e s i s and v i s u a l i z e d by fluorography of the d r i e d g e l .
2 . Chase of intermediate to mature-size cytochrome c^: The precursor was imported i n t o mitochondria as above but i n the presence of 2 . 5 mM MgCl^ and absence of NADH to accumulate the i n t e r m e d i a t e - s i z e p r o t e i n . The mixtures were then t r e a t e d w i t h P r o t e i n a s e K ( F i g . 4-D) or not ( F i g 4.C) as des c r i b e d above. The mitochondria were r e - i s o l a t e d and suspended i n b u f f e r A then incubated f o r 30 min at 25 C i n the presence of a d d i t i o n s , as i n d i c a t e d , i n a t o t a l volume of 1 0 0 j i l . The samples were d i l u t e d w i t h 1 ml b u f f e r A and the mitochondria were sedimented by c e n t r i f u g a t i o n and then d i s s o c i a t e d i n SDS sample b u f f e r as above.
Miscellaneous Methods
Cytochrome c was p u r i f i e d from N. crassa and s p e c i f i c a n t i b o d i e s were r a i s e d i n r a b b i t s as p r e v i o u s l y d e s c r i b e d ( 2 9 ) . R a d i o a c t i v i t y determinations
199
were performed by s c i n t i l l a t i o n counting i n 0.1 ml of 10% (w/v) SDS, 1 M T r i s / H C l (pH 8.0), plus 10 ml ACS I I (Amersham). SDS Po l y a c r y l a m i d e g e l e l e c t r o p h o r e s i s was performed u s i n g Standard techniques (32). Gels were prepared f o r flu o r o g r a p h y by soaking them f o r 30 min i n A m g l i f y (Amersham) f o l l o w e d by d r y i n g and then exposure to X-ray f i l m s a t -80 C. The bands were q u a n t i f i e d by l a s e r densitometry (LKB-GSXL). P r o t e i n was determined as d e s c r i b e d p r e v i o u s l y (33).
RESULTS
Covalent Attachment of Heme to Apocytochrome c Requires NADH and a Cofactor from Reticulocyte Lysate or C e l l Cytosol
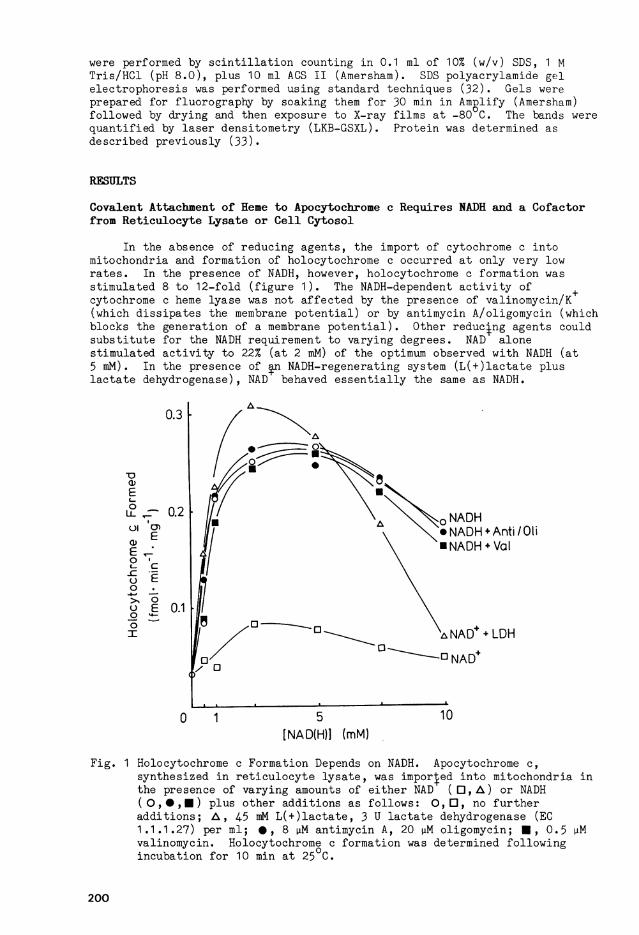
In the absence of redu c i n g agents, the import of cytochrome c i n t o m itochondria and formation of holocytochrome c occurred a t only very low r a t e s . In the presence of NADH, however, holocytochrome c f o r m a t i o n was st i m u l a t e d 8 to 1 2 - f o l d ( f i g u r e 1 ) . The NADH-dependent a c t i v i t y of cytochrome c heme l y a s e was not a f f e c t e d by the presence of valinomycin/K (which d i s s i p a t e s the membrane p o t e n t i a l ) or by ant i m y c i n A/oligomycin (which b l o c k s the generation of a membrane p o t e n t i a l ) . Other reduc|ng agents could S u b s t i t u t e f o r the NADH requirement to v a r y i n g degrees. NAD alone s t i m u l a t e d a c t i v i t y to 22% (at 2 mM) of the Optimum observed w i t h NADH (at 5 mM). In the presence of an NADH-regenerating System ( L ( + ) l a c t a t e p l u s l a c t a t e dehydrogenase), NAD behaved e s s e n t i a l l y the same as NADH.
0 1 5 10 [NAD(H)] (mM)
F i g . 1 Holocytochrome c Formation Depends on NADH. Apocytochrome c, syn t h e s i z e d i n r e t i c u l o c y t e l y s a t e , was imported i n t o m i t o c h o n d r i a i n the presence of v a r y i n g amounts of e i t h e r NAD ( D , A ) or NADH ( O , • , • ) p l u s other a d d i t i o n s as f o l l o w s : O , • , no f u r t h e r a d d i t i o n s ; A , 45 mM L ( + ) l a c t a t e , 3 U l a c t a t e dehydrogenase (EC 1.1.1.27) per ml; • , 8 \M a n t i m y c i n A, 20 jiM o l i g o m y c i n ; • , 0.5 uM val i n o m y c i n . Holocytochrome c formation was determined f o l l o w i n g i n c u b a t i o n f o r 10 min at 25 C.
200
NADPH (at 1 mM), which has been reported to be r e q u i r e d f o r holocytochrome c formation i n yeast ( 9 ) , stimulated a c t i v i t y to 45% of the NADH Optimum ( 8 ) , as d i d g l u t a t h i o n e (43% at 25 mM)(8), Na d i t h i o n i t e (100% a t 1 mg/ml, unpublished), and 2-mercaptoethanol (7% at 50 mM, unpubli s h e d ) .
We have p r e v i o u s l y i n v e s t i g a t e d the r o l e of NADH i n the cov a l e n t attachment of heme to apocytochrome c (8 ) . NADH was not r e q u i r e d f o r the bin d i n g of apocytochrome c to mitochondria and, u n l i k e the m a j o r i t y of imported m i t o c h o n d r i a l p r o t e i n s , a membrane p o t e n t i a l was not necessary f o r any of the stages of cytochrome c import. NADH was not i n v o l v e d i n the re d u c t i o n of the c y s t e i n e t h i o l s to which heme i s c o v a l e n t l y a t t a c h e d . P r e l i m i n a r y experiments i n d i c a t e t h a t r e d u c t i o n of the heme i r o n from F e ( I I I ) t o F e ( I I ) must occur before the enzymatic l i n k a g e of heme to apocytochrome c, and that t h i s r e d u c t i o n step r e q u i r e s NADH (Nicholson and Neupert, i n p r e p a r a t i o n ) .
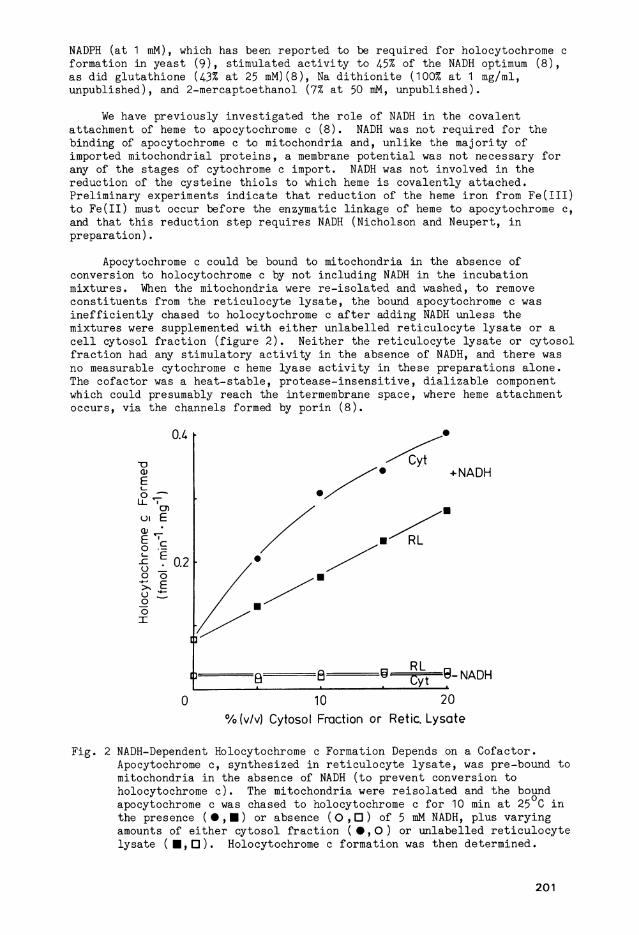
Apocytochrome c could be bound to mitochondria i n the absence of conversion to holocytochrome c by not i n c l u d i n g NADH i n the i n c u b a t i o n mixtures. When the mitochondria were r e - i s o l a t e d and washed, to remove c o n s t i t u e n t s from the r e t i c u l o c y t e l y s a t e , the bound apocytochrome c was i n e f f i c i e n t l y chased to holocytochrome c a f t e r adding NADH unless the mixtures were supplemented w i t h e i t h e r u n l a b e l l e d r e t i c u l o c y t e l y s a t e or a c e l l c y t o s o l f r a c t i o n ( f i g u r e 2 ) . N e i t h e r the r e t i c u l o c y t e l y s a t e or c y t o s o l f r a c t i o n had any s t i m u l a t o r y a c t i v i t y i n the absence of NADH, and there was no measurable cytochrome c heme ly a s e a c t i v i t y i n these p r e p a r a t i o n s a l o n e . The c o f a c t o r was a h e a t - s t a b l e , p r o t e a s e - i n s e n s i t i v e , d i a l i z a b l e component which could presumably reach the intermembrane space, where heme attachment occurs, v i a the Channels formed by p o r i n ( 8 ) .
n q B= = S Ö^T50" NADH 0 10 20
%(v/v) Cytosol Fraction or Retic. Lysate
F i g . 2 NADH-Dependent Holocytochrome c Formation Depends on a C o f a c t o r . Apocytochrome c, sy n t h e s i z e d i n r e t i c u l o c y t e l y s a t e , was pre-bound to mitochondria i n the absence of NADH (to prevent conversion to holocytochrome c ) . The mitochondria were r e i s o l a t e d and the bound apocytochrome c was chased to holocytochrome c f o r 10 min a t 25 C i n the presence (•,•) or absence ( 0 , D ) of 5 mM NADH, plus v a r y i n g amounts of e i t h e r c y t o s o l f r a c t i o n ( • , O ) or u n l a b e l l e d r e t i c u l o c y t e l y s a t e ( • , • ) . Holocytochrome c format i o n was then determined.
201

Covalent attachment of heme to apocytochrome c, by the enzyme cytochrome c heme l y a s e , r e q u i r e s NADH i n a step which i s a l s o dependent on a f a c t o r p r e sent i n r e t i c u l o c y t e l y s a t e or N. crassa c e l l c y t o s o l . T his step appears to be the r e d u c t i o n of the heme i r o n and the c o f a c t o r probably mediates t h i s r e d u c t i o n . The conformation of the heme group and/or i t s a b i l i t y to form c r i t i c a l alignments w i t h apocytochrome c may be e s s e n t i a l f o r the cytochrome c heme lyas e r e a c t i o n .
The Translocation of Cytochrome c to a Protease-Resistant Location Depends on NADH and i s Inhibited by Deuterohemin
We have p r e v i o u s l y demonstrated t h a t the apocytochrome c a s s o c i a t e d w i t h mitochondria i n the presence of deuterohemin (an analogue of heme which does not c o n t a i n the v i n y l groups at p o s i t i o n s 3 and 8 of the p o r p h y r i n r i n g t h a t are necessary f o r formation of the t h i o e t h e r l i n k a g e s w i t h apocytochrome c) coul d be d i s p l a c e d by excess added apocytochrome c; e.g. import was blocked at the stage of h i g h - a f f i n i t y r e c e p t o r b i n d i n g ( 2 0 ) . We t h e r e f o r e proposed t h a t the t r a n s l o c a t i o n of cytochrome c across the outer m i t o c h o n d r i a l membrane du r i n g import i s i n t r i n s i c a l l y coupled t o the cov a l e n t attachment of heme (8,20).
T r a n s l o c a t i o n of cytochrome c across the outer m i t o c h o n d r i a l membrane was determined by the a c c e s s i b i l i t y of the p r o t e i n to e x t e r n a l l y added proteases which d i d not penetrate the outer membrane under the c o n d i t i o n s used here ( f i g u r e 3 ) . In the presence of NADH, fo r m a t i o n of holocytochrome c occ u r r e d a t 23-times the r a t e compared to samples which d i d not c o n t a i n NADH (mixtures I I and I , r e s p e c t i v e l y ) . T r a n s l o c a t i o n of the cytochrome c to a p r o t e a s e - r e s i s t a n t l o c a t i o n a l s o depended on NADH ( c f . lane 1 vs 3 ) . As a c o n t r o l , the imported cytochrome c was s e n s i t i v e to protease treatment when the mitochondria were l y s e d w i t h detergent f o l l o w i n g import (lanes 2 and U)•
In the presence of NADH, 80% of the conve r s i o n of apocytochrome c t o holocytochrome c was i n h i b i t e d by deuterohemin ( c f . mixture I I I vs I V ) . In an i d e n t i c a l manner, the t r a n s l o c a t i o n of cytochrome c to a pr o t e a s e -r e s i s t a n t l o c a t i o n was i n h i b i t e d by approximately 80% i n the presence of deuterohemin ( c f . lane 5 vs 7 ) . Therefore, whenever holocytochrome c fo r m a t i o n was s t i m u l a t e d by NADH, or i n h i b i t e d by deuterohemin, there were p a r a l l e l changes i n the formation of a p r o t e a s e - r e s i s t a n t cytochrome c s p e c i e s .
The c l o s e c o r r e l a t i o n between the formation of holocytochrome c and the t r a n s p o r t of the p r o t e i n to a l o c a t i o n i n mitochondria where i t was not a c c e s s i b l e to e x t e r n a l l y added proteases confirms our hypothesis t h a t the two events are t i g h t l y coupled.
Formation of Mature-Size Cytochrome c^ Depends on NADH and i s Inhibited by Deuterohemin
The import of cytochrome c^ could be a r r e s t e d a t d i f f e r e n t stages under v a r i o u s c o n d i t i o n s ( f i g u r e 4A). When import was performed i n the presence of EDTA/o-phenanthroline, which i n h i b i t s the mat r i x p e p t i d a s e , the p r e c u r s o r of cytochrome c^ (p-C^) was accumulated i n a p r o t e a s e - r e s i s t a n t l o c a t i o n (lane 1 ) . In the presence of MgCl^, the imported cytochrome c^ was processed to the i n t e r m e d i a t e - s i z e p r o t e i n ( i - C ^ ; lane 2 ) . P r o c e s s i n g to the mature-size cytochrome c^ (m-C^) depended on the presence of NADH i n a d d i t i o n to MgCl^ (la n e 3 ) . The second p r o c e s s i n g step, or events t i g h t l y coupled to the second pr o c e s s i n g step (e.g. co v a l e n t attachment of heme), t h e r e f o r e appear to r e q u i r e NADH.
To determine whether the second p r o c e s s i n g step of cytochrome c^ maturation i s coupled to heme l i n k a g e , cytochrome c^ was imported i n the presence or absence of deuterohemin ( f i g u r e 4-B). In the absence of NADH,
202

cytochrome c^ accumulated as the i n t e r m e d i a t e - s i z e p r o t e i n w i t h n e g l i g i b l e formation of m-C (lane 1 ) . In the presence of NADH, more than 65% of the imported cytochrome c^ appeared as m-C (lane 2 ) . When deuterohemin was included i n import mixtures t h a t were otherwise i d e n t i c a l to t h a t i n lane 2 , formation of m-C was i n h i b i t e d (lane 3 ) • Formation of m-C. i n the presence of deuterohemin occurred at 39% of the r a t e compared to th a t i n the absence of deuterohemin ( c f . lane 2 vs 3 ) » Although the t o t a l amount of imported cytochrome c^ was lower i n the presence of deuterohemin, the r a t i o of m-C t o i - C j i n the presence of deuterohemin ( 0 . 6 6 ) was c o n s i d e r a b l y lower than the r a t i o i n the absence of deuterohemin ( 2 . 1 ) i n d i c a t i n g t h a t the second processing step was s p e c i f i c a l l y blocked.
S i m i l a r r e s u l t s were observed when cytochrome c^ was imported from r e t i c u l o c y t e l y s a t e i n t o mitochondria i n the absence of NADH, then the mitochondria were r e i s o l a t e d and the bound cytochrome c^ was chased under vari o u s c o n d i t i o n s ( f i g u r e 4 0 ) . Low amounts of the bound cytochrome c. were chased to m-C i n the absence of NADH (lane 1 ) . In the presence of NADH, however, m-C formation was s t i m u l a t e d 3 . 2 - f o l d (lane 2 ) . This NADH-dependent chase to m-G^ was blocked when deuterohemin was added to the inc u b a t i o n mixture (lane 3 ) . When u n l a b e l l e d r e t i c u l o c y t e l y s a t e was added during the chase i n c u b a t i o n , the NADH-dependent m-C formation was s t i m u l a t e d a f u r t h e r 39% ( c f . lane 4 vs 2 ) . P r o c e s s i n g to m-C1 under these c o n d i t i o n s
A. B. + NADH
1 11 1 in nT~
-NADH +NADH - DH + DH Holoc Formed: Q01Ö (U08 0.558 0.109
+ 8
F i g . 3 A. T r a n s l o c a t i o n of Cytochrome c to a P r o t e a s e - R e s i s t a n t L o c a t i o n Depends on NADH. Apocytochrome c, syn t h e s i z e d i n r e t i c u l o c y t e l y s a t e , was imported i n t o mitochondria i n the absence (pool I) or presence (pool I I ) of 5 mM NADH. A l i q u o t s of the two samples were removed f o r determinatjj.on of holocytochrome c formation (expressed as fmol x min" x mg~ ). The samples were then d i v i d e d i n t o two a l i q u o t s , one of which r e c e i v e d T r i t o n X - 1 0 0 (1% (w/v) f i n a l c o n c e n t r a t i o n ) ( l a n e s 2 & 4 ) . A l l four samples were t r e a t e d w i t h 4 0 \ig P r o t e i n a s e K/ml f o r 2$ min at 25 C a t a m i t o c h o n d r i a l p r o t e i n c o n c e n t r a t i o n of 1 mg/ml. (Using t h i s procedure, l e s s than 5% of adenylate kinase a c t i v i t y , a marker f o r the intermembrane space, was l o s t i n the absence of T r i t o n X - 1 0 0 w h i l e more than 90% was l o s t i n the presence of detergent.) T o t a l cytochrome c was immunoprecipitated from the samples, r e s o l v e d by SDS-PAGE, and v i s u a l i z e d by flu o r o g r a p h y . B. T r a n s l o c a t i o n of Cytochrome c to a P r o t e a s e - R e s i s t a n t L o c a t i o n i s I n h i b i t e d by Deuterohemin. Apocytochrome c was imported i n t o mitochondria i n mixtures c o n t a i n i n g 5 mM NADH and i n e i t h e r the absence ( p o o l I I I ) or presence (pool IV) of 100 JIM deuterohemin. Holocytochrome c formation and t r a n s l o c a t i o n to a p r o t e a s e - r e s i s t a n t l o c a t i o n were then determined as de s c r i b e d f o r panel A.
203

EDTA/o-phe + - - NADH - + + MgCl2 + + Deuterohem- - + NÄDH - - + 1 2 3
1 2 3
NADH - + + + + + -Deuterohemin - - + - + - -Retic. Lysate _ - - + + - -Cytosol _ _ _ _ - + -Na Dithionite _ _ - - - - +
1 2 3 ^ 5 6 7
1 2 3 4 5 6 7 F i g . 4 A. Import of Cytochrome c, i n t o M i t o c h o n d r i a . The precursor of
cytochrome c 1 was iraportea i n t o mitochondria i n the presence of e i t h e r 5 mM EDTA plus 0.2 mM o-phenanthroline, 2.5 mM M g C l 2 > or 5 mM NADH as i n d i c a t e d . F o l l o w i n g i n c u b a t i o n f o r 30 min at 25 C, the samples were d i g e s t e d w i t h P r o t e i n a s e K f o r 30 min at 0 C and then the mitochondria were r e i s o l a t e d , d i s s o c i a t e d i n SDS sample b u f f e r and processed as des c r i b e d i n Methods. p, pr e c u r s o r ; i , i n t e r m e d i a t e ; m, mature. B . The NADH-Dependent Second P r o c e s s i n g Step of Cytochrome i s I n h i b i t e d by Deuterohemin. The precursor of cytochrome c^ was imported i n t o mitochondria, as d e s c r i b e d i n panel A, i n the presence of 2.5 mM MgCl ?, plus 5 mM NADH or 0.1 mM deuterohemin as i n d i c a t e d . C. Chase of Mitochondrial-Bound Cytochrome c^. The precursor of cytochrome c^ was imported i n t o mitochondria i n the presence of 2.5 mM MgCl^ f o r 30 min at 25 C. The mitochondria were r e - i s o l a t e d by c e n t r i f u g a t i o n and suspended i n b u f f e r A. The bound cytochrome c^ was chased f o r 30 min at 25 C i n b u f f e r A alone or i n the presence of 5 mM NADH, 0.1 mM deuterohemin, 25% (v/v) u n l a b e l l e d r e t i c u l o c y t e l y s a t e , 20% (v/v) c y t o s o l f r a c t i o n , or 1 mg Na d i t h i o n i t e / m l as i n d i c a t e d . M i t o c h o n d r i a were r e i s o l a t e d and analyzed as above. D. Chase of Pr o t e a s e - R e s i s t a n t I n t e r m e d i a t e - S i z e Cytochrome c 1 . The experiment was performed as d e s c r i b e d f o r panel C except t h a t f o l l o w i n g the f i r s t i n c u b a t i o n p e r i o d the samples were t r e a t e d w i t h P r o t e i n a s e K as des c r i b e d i n panel A.
204

was a l s o i n h i b i t e d by deuterohemin (lane 5 ) . The a d d i t i o n a l Stimulation of m-C formation observed i n the presence of r e t i c u l o c y t e l y s a t e (lane 4 ) occurred only m a r g i n a l l y (about 5%) when a c e l l c y t o s o l f r a c t i o n was used i n s t e a d (lane 6 ) . L i k e the NADH-requiring step of cytochrome c import, Na d i t h i o n i t e could be used as a reducing agent f o r m-Cj formation w i t h comparable e f f i c i e n c y ( c f . lane 7 vs 2 ) .
A s i m i l a r experiment as de s c r i b e d i n f i g u r e 4 0 was attempted t o examine only the chase from i-C^ to m-C^. The i n t e r m e d i a t e - s i z e cytochrome c was accumulated by impor t i n g p-C^ i n t o mitochondria i n the absence of NADH f o l l o w e d by treatment w i t h P r o t e i n a s e K to remove p-C^ t h a t might be bound to the m i t o c h o n d r i a l s u r f a c e . Under these c o n d i t i o n s the mitochondria contained mostly i-C^ and very l i t t l e protease r e s i s t a n t p-C^ ( f i g u r e 4 A , lane 2 ) . The mitochondria were then r e - i s o l a t e d and i-C^ was chased to m-C under v a r i o u s c o n d i t i o n s ( f i g u r e 4 D ) . Although the e f f i c i e n c y of the chase was low f o l l o w i n g t h i s treatment, the same b a s i c f e a t u r e s d e s c r i b e d i n f i g u r e 4 0 were observed, namely: m-C f o r m a t i o n was s t i m u l a t e d by NADH ( 2 . 2 - f o l d ; lane 2) and somewhat f u r t h e r i n the presence of u n l a b e l l e d r e t i c u l o c y t e l y s a t e (40% over NADH alone; lane 4 ) or c y t o s o l f r a c t i o n (26% over NADH alone; lane 6 ) ; t h i s S t i m u l a t i o n over NADH-independent m-C formation was i n h i b i t e d by deuterohemin (lanes 3 and 5); formation of m-C i n the presence of Na d i t h i o n i t e (lane 7) was 85% of the amount observed w i t h NADH (lane 2 ) .
I t t h e r e f o r e appears tha t the second p r o c e s s i n g step of cytochrome c^ i s a f f e c t e d by c o n d i t i o n s which a f f e c t heme attachment to cytochrome c and thu s , by analogy, might a f f e c t heme attachment to cytochrome c^. I t i s p o s s i b l e then t h a t heme attachment to cytochrome c^ enhances the r a t e of the second p r o c e s s i n g step, though i t might not be o b l i g a t o r y .
DISCUSSION
Cytochromes c and c^ are imported i n t o mitochondria by d i s t i n c t l y d i f f e r e n t mechanisms. They do, however, share the common requirement f o r c o v a l e n t heme attachment du r i n g the f i n a l stages of t h e i r r e s p e c t i v e import pathways. During the import of cytochrome c, t h i s step occurs i n the m i t o c h o n d r i a l intermembrane space (8) and i s r e s p o n s i b l e f o r the completion of t r a n s l o c a t i o n of the p r o t e i n across the outer membrane, probably owing t o a c o n f o r m a t i o n a l change r e s u l t i n g from the c o v a l e n t l y attached heme group. To account f o r previous o b s e r v a t i o n s and those presented here we propose the f o l l o w i n g model. Apocytochrome c spontaneously i n s e r t s p a r t way i n t o the l i p i d b i l a y e r of the outer membrane w i t h low a f f i n i t y (34-39). The p a r t i a l l y i n s e r t e d p r o t e i n i s then bound w i t h high a f f i n i t y by a b i n d i n g p r o t e i n which i s l o c a l i z e d i n the intermembrane Space (Köhler, S t u a r t and Neupert, i n p r e p a r a t i o n ) . Heme i s c o v a l e n t l y attached to the c y s t e i n e t h i o l s a t p o s i t i o n s 14 and 17 ( u n i v e r s a l numbering) i n a r e a c t i o n which i s c a t a l y z e d by the enzyme cytochrome c heme l y a s e and i s dependent on NADH plus a c y t o s o l i c c o f a c t o r . The NADH appears to be r e q u i r e d f o r r e d u c t i o n of the heme i r o n t o F e ( I I ) before t h i s step, though i t i s unclear where and how t h i s might occur ( N i c h o l s o n and Neupert, i n p r e p a r a t i o n ) . The c o v a l e n t l y attached heme group i s presumably then the nucleus around which the r e s t of the Po l y p e p t i d e c h a i n f o l d s , drawing the remainder of the molecule across the outer membrane as i t does so to i t s f i n a l l o c a t i o n i n the intermembrane space.
Since cytochrome c^ maturation shares so many common f e a t u r e s w i t h the heme-attaching step i n cytochrome c import, i t might be expected t h a t the same enzyme c a t a l y z e s the r e a c t i o n f o r both. Cytochrome c 1 , l i k e cytochrome c, i s a l s o exposed to the intermembrane space where heme attachment occurs i n the l a t t e r case. Furthermore, there i s sequence homology between the two p r o t e i n s i n the heme-binding r e g i o n , e.g. i n N. cr a s s a (40-42):
205

Cytochrome c Cytochrome c.
1
(n)...KTJ (n)...RE1
)EG...(c) ;RV...(C)
Despite these l i k e l y circumstances, however, a yeast mutant which l a c k s the heme lyas e enzyme f o r cytochrome c ( c y c 3 ~ ) contains normal l e v e l s of cytochrome c^ ( 4 3 , 4 4 ) • We have found t h i s to a l s o be the case f o r a mutant of N. crassa ( c y t - 2 - 1 ) t h a t i s d e f i c i e n t i n cytochrome c heme l y a s e a c t i v i t y , and t h e r e f o r e contains no cytochrome c, but which has normal l e v e l s of cytochrome c^ (Nargang, N i c h o l s o n and Neupert, i n p r e p a r a t i o n ) . I t t h e r e f o r e appears th a t two separate enzymes are i n v o l v e d , perhaps because of t o p o l o g i c a l requirements d u r i n g import and maturation.
How might the covalent attachment of heme to cytochrome c^ be r e l a t e d t o the second p r o t e o l y t i c p r o c e s s i n g event? I t i s apparent t h a t heme attachment must precede p r o c e s s i n g of i-C«. t o m-C.j. This i s i n d i c a t e d by the requirement of t h i s step f o r NÄDH and i t s i n h i b i t i o n by deuterohemin. S i m i l a r l y , a heme-deficient mutant of yeast accumulates i-C^ i n vi v o ( 2 5 - 2 6 ) . One p o s s i b i l i t y i s t h a t heme attachment a f f e c t s the conformation of i-C^ around the r e g i o n of the second p r o c e s s i n g s i t e . which enables r e c o g n i t i o n by the second p r o c e s s i n g protease. A more l i k e l y p o s s i b i l t y i s t h a t f o l l o w i n g heme attachment and the ensuing c o n f o r m a t i o n a l changes, the second p r o c e s s i n g s i t e becomes a c c e s s i b l e to the peptidase. For example, a p p l y i n g the p r i n c i p l e s of the 'stop t r a n s p o r t ' model ( 4 5 ) , the f o l l o w i n g sequence of events might occur. The prec u r s o r of cytochrome c^ binds to i t s receptor a t the outer m i t o c h o n d r i a l membrane and the N-terminus i s t r a n s l o c a t e d through the i n n e r membrane, v i a contact s i t e s , i n a step which i s dependent on a membrane p o t e n t i a l . The p o s i t i v e l y charged f i r s t p a r t of the prepiece i s reraoved by the c h e l a t o r - s e n s i t i v e m atrix peptidase l e a v i n g the more hydrophobic 'stop t r a n s p o r t ' domain (the second p a r t of the prepiece) i n s e r t e d through the i n n e r membrane. At some p o i n t f o l l o w i n g t r a n s l o c a t i o n of the remainder of the p r o t e i n across the outer membrane, the C-terminus (which anchors the mature p r o t e i n to the in n e r membrane) becomes embedded i n the inner membrane. At t h i s stage the second p r o c e s s i n g s i t e i s presumably not a c c e s s i b l e to the intermembrane-space-localized second p r o c e s s i n g protease. F o l l o w i n g covalent heme attachment, however, the r e s u l t i n g c o n f o r m a t i o n a l change might expose t h i s s i t e a l l o w i n g the removal of the remainder of the prepiece to occur.
Another p o s s i b i l i t y i s t h a t cytochrome c^ f o l l o w s a 'conservative s o r t i n g ' pathway s i m i l a r to t h a t of the Fe/S p r o t e i n of the bc^ complex ( 4 6 ) i n which p-C^ would be t r a n s p o r t e d completely i n t o the matrix where the f i r s t p art of the prepiece i s removed. The second part of the prepiece might then r e d i r e c t i-C^ back to the i n n e r membrane. Covalent heme attachment and the ensuing conformational change i s then r e s p o n s i b l e f o r drawing the p r o t e i n back across the in n e r membrane, exposing the second p r o c e s s i n g s i t e i n the meantime. As the C-terminus i s p u l l e d through i t anchors the p r o t e i n to the i n n e r membrane. I f t h i s pathway were to be the case, then the co u p l i n g of t r a n s l o c a t i o n to heme attachment f o r cytochrome c 1 might be analogous to the same event d u r i n g cytochrome c import, except t h a t i n s t e a d of being t r a n s l o c a t e d across the outer membrane i n t o the intermembrane space (as i s the case f o r cytochrome c ) , cytochrome c^ i s t r a n s l o c a t e d from the matrix across the inne r membrane to the intermembrane space. The intermembrane space i - C j should t h e r e f o r e have heme attached to i t wh i l e the matrix l o c a l i z e d i - C 1 species should not. Fu r t h e r work i s r e q u i r e d i n t h i s regard to determine ine exact route t h a t cytochrome c^ takes d u r i n g import. Where and how the heme a t t a c h i n g step occurs, and how i t compares w i t h the e q u i v a l e n t step of cytochrome c import w i l l a l s o be of co n s i d e r a b l e i n t e r e s t .
206

REFERENCES
1. Neupert, W. & Schatz, G. (1981) Trends Biochem. Sei. 6:1-4. 2. Hay, R., Böhni, P. & Gasser, S. (1984) Biochim. Biophys. Acta 779:65-87. 3. Harmey, M.A. & Neupert, W. (1985) i n The enzymes of b i o l o g i c a l membranes
(Martonosi, A., ed.) 4:431-464, Plenum Press, New York. 4. Nicholson, D.W. & Neupert, W. (1987) i n Protein transfer and organelle
biogenesis (Das, R.C. & Robbins, P.W., eds.) Academic Press, New York (in press).
5. Schleyer, M., Schmidt, B. & Neupert, W. (1982) Eur. J . Biochem. 125:109-116.
6. Pfanner, N. & Neupert, W. (1985) EMBO J. 4:2819-2825. 7. Pfanner, N. & Neupert, W. (1986) FEBS Lett. 209:152-156. 8. Nicholson, D.W., Köhler, H. & Neupert, W. (1987) Eur. J . Biochem.
164:147-157. 9. Basile, G., DiBello, C. & Taniüchi, H. (1980) J . B i o l . Chem.
255:7181-7191. 10. Teintze, M., Slaughter, M., Weiss, H. & Neupert, W. (1982) J. B i o l .
Chem. 257:10364-10371. 11. Schleyer, M. & Neupert, W. (1985) C e l l 43:339-35.0. 12. Zimmermann, R., Paluch, V. & Neupert, W. (1979) FEBS Lett. 108:141-146. 13. Stewart, J.W., Sherman, F., Shipman, N.A. & Jackson, M. (1971) J. B i o l .
Chem. 246:7429-7445. 1 4 . Zitomer, R.S. & H a l l , B.D. (1976) J. B i o l . Chem. 251:6320-6326. 15. Smith, M., Leung, D.W., Gillam, S., A s t e i l , CR., Montgomery, D.L. &
H a l l , B.D. (1979) C e l l 16:753-761. 16. Matsuura, S., Arpin, M., Hannum, C, Margoliash, E,, Sabatini, D.D. &
Morimoto, T. (1981) Proc. Natl. Acad. Sei. USA 78:4368-4372. 17. Scarpulla, R.C, Agne, K.M. & Wu, R. (1981) J . B i o l . Chem.
256:6480-6486. 18. Zimmermann, R., Hennig, B. & Neupert, W. (1981) Eur. J . Biochem.
116:455-460. 19. Korb, H. & Neupert, W. (1978) Eur. J. Biochem. 91:609-620. 20. Hennig, B. & Neupert, W. (1981) Eur. J. Biochem. 121:203-212. 21. Hennig, B., Köhler, H. & Neupert, W. (1983) Proc. Natl. Acad. Sei. USA
80:4963-4967. 22. Veloso, D., Basile, G., & Taniüchi, H. (1981) J. B i o l . Chem.
256:8646-8651. 23. Taniüchi, H., Basile, C , Taniüchi, M. & Veloso, D. (1983) J. B i o l .
Chem. 258:10963-10966. 24. Visco, C , Taniüchi, H. & Berlett, B.S. (1985) J. B i o l . Chem.
260:6133-6138. 25. Gasser, S.M., Ohashi, A., Daum, C , Böhni, P.C., Gibson, J . , Reid,
G.A., Yonetani, T. & Schatz, G. (1982) Proc. Natl. Acad. Sei. USA 79:267-271.
26. Ohashi, A., Gibson, J . , Gregor, I. & Schatz, G. (1982) J. B i o l . Chem. 257:13042-13047.
27. Wakabayashi, S., Matsubara, H., Kim, C.H., Kawai, K. & King, T.E. (1980) Biochem. Biophys. Res. Commun. 97:1548-1554«
28. L i , Y., Leonard, K. & Weiss, H. (1981) Eur. J . Biochem. 116:199-205. 29. Hennig, B. & Neupert, W. (1983) Methods Enzymol. 97:261-274. 30. Pelham, H.R.B. & Jackson, R.J. (1976) Eur. J . Biochem. 67:247-256. 3 1 . Zimmermann, R. & Neupert, W. ( 1 9 8 3 ) Methods Enzymol. 97:275-286. 32. Laemmli, U.K. (1970) Nature (Lond.) 227:680-685. 33. Bradford, M. (1976) Anal. Biochem. 72:248-254. 34. Rietveld, A., Sijens, P., V e r k l e i j , A.J. & de K r u i j f f , B. (1983) EMBO J.
6:907-913.
207

35. Dumont, M.E. & Richards, F.M. (1984) J. B i o l . Chem. 259:4147-4156. 36. Rietveld, A. & de K r u i j f f , B. (1984) J . B i o l . Chem. 259:6704-6707. 37. Rietveld, A., Ponjee, G.A.E., Schiff e r s , P,, J o r d i , W., van de Coolwijk,
P.J.F.M., Demel, R.A., Marsh, D. & de K r u i j f f , B. (1985) Biochim. Biophys. Acta 818:398-409.
38. Rietveld, A., J o r d i , W. & de K r u i j f f , B. (1986) J. B i o l . Chem. 261:3846-3856.
39. Berkhout, T.A., Rietveld, A. & de K r u i j f f , B. (1987) Biochim. Biophys. Acta 897:1-4.
40. Heller, J . & Smith, E.L. (1966) J . B i o l . Chem. 241:3165-3180. 4 1 . Lederer, F. & Simon, A.M. (1974) Biochem. Biophys. Res. Commun.
56:317-323. 42. Römisch, J . , Tropschug, M., Sebald, W. & Weiss, H. (1987) Eur. J .
Biochem. 164:111-115. 43. Rothstein, R.J. & Sherman, F. (1980) Genetics 94:871-889. 44. Dumont, M.E., Ernst, J.F., Hampsey, D.M. & Sherman, F. (1987) EMBO J .
6:235-241. 45. Hurt, E.C. & van Loon A.P.G.M. (1986) Trends Biochem. Sei. 11:204-207. 46. H a r t l , F.-U., Schmidt, B., Wächter, E., Weiss, H. & Neupert, W. (1986)
C e l l 47:939-951.
208