Embed Size (px)
Citation preview

Vo1.165, No. 2,1989
December 15,1989
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Pages 590-594
NADPH-CYTOCHROME P-450 REDUCTASE IS NOT A COMPONENT OF
THE LIVER MICROSOMAL STEROID 5-ALPHA REDUCTASE
kuo-Chi Cheng*, Regina W. Wang and Anthony Y.H. Lu
~Dept. Pediatrics, Cornell University Medical College, NY, NY 10021
Dept. Animal and Exploratory Drug Metabolism, Merck, Sharp & Dohme Research Labs, Rahway, NJ 07065
Received August 30, 1989
Liver microsomal steroid 5-alpha-reduction is catalyzed by a NADPH-dependent enzyme system. The requirement of NADPH- cytochrome P-450 reductase to shuttle reduction equivalents from NADPH to steroid 5-alpha-reductase was investigated using an inhibitory antibody against NADPH-cytochrome P-450 reductase. This antibody preparation inhibited cytochrome c reduction in microsomes from female rat liver with an I50 of 0.75 mg anti- body/mg of microsomal protein. Benzphetamine N-demethylation and testosterone 6-beta-hydroxylation, two cytochrome P-450-mediated oxidative reactions, were inhibited by the antibody. On the other hand, testosterone 5-alpha-reductase was not affected by the antibody. These results suggest that NADPH-cytochrome P-450 reductase is not an obligatory component of the liver microsomal steroid 5-alpha-reduction. ©i989Acad~micPress, Zno.
Steroid 5-alpha-reductase (S5R) exists predominantly in
hepatic microsomes and in microsomal and nuclear fractions of
many steroid hormone-sensitive tissues (i). S5R activity in
hepatic microsomes of female animals is 3-10 times greater than
that of male animals. The difference is due to a higher level of
S5R in female liver microsomes (2). Several lines of evidence
indicate that S5R is an isozymic enzyme (3,4,5). Golf and
Graef (3) separated S5R into two fractions: one preferentially
reduced testosterone, the other preferentially reduced
progesterone and cortisone. In the same study, it was also
shown that NADPH-cytochrome P-450 reductase is an obligatory
component of S5R. On the other hand, Ichihara and Tanaka (6)
0006-291~89 $1.50 Copyr~ht© 1~9 ~ Aca~micPress,~c. Allrigh~ ~repro~ct ionina~rmrese~ed. 590

Vol. 165, No. 2, 1989 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
have recently demonstrated that progesterone 5-alpha-reduction
was independent of NADPH-cytochrome P-450 reductase. Therefore,
we decided to study the effect of antibody against NADPH--
cytochrome P-450 reductase on testosterone 5-alpha-reduction
by hepatic microsomes. This communication reports our findings.
Materials and Methods
Materials: 4-14C-Testosterone (58 uCi/mmol) and Aquasol-2 were obtained from New England Nuclear (Boston, MA). The TLC plates (PE sil G/UV) were purchased from Whatman (Kent, England). The antibody against NADPH-cytochrome P-450 reductase was a gift from Dr. B.S.S. Masters. Liver microsomes were prepared from female Sprague-Dawley rats (150-200gm) by a differential centri- fugation procedure (7).
Assays: Testosterone metabolism was assayed according to a previously established procedure (7). The assay was carried out in iml of volume containing 0.5mg of liver microsomes of female rats and various amounts of antibody solution. The mixture was preincubated at 37°C for 3 min. prior to adding 100ul of imM NADPH. After adding NADPH, the reaction was continued for another i0 mins. at 37°C, then stopped by adding 3ml of methylene chloride. The substrate plus metabolites were extracted with another 3ml of methylene chloride. The two extracts were combined, dried under nitrogen and applied to a TLC plate. When testosterone served as the substrate, a solvent system of chloroform/ethylacetate/ethanol (4/1/0.6) was used to separate the metabolites. After development, the substrate and metabo- lites were located either by UV light or by autoradiography. The radioactive spots were scraped from the TLC plate, dissolved in 5ml of Aquasol-2 and counted in a scintillation counter. Cytochrome c reduction by microsomal reductase was performed in a final volume of 1 ml containing 40 ug of microsomal protein and various amounts of antibody according to previously estab- lished method (8). Microsomal benzphetamine N-demethylase activity was determined according to Thomas, et al (9).
Antibody Purification: In preliminary studies, antibody against pig NADPH-cytochrome P-450 reductase provided by Dr. B.S.S. Masters was used. In subsequent studies, antibody against rat NADPH-cytochrome P-450 was employed. The antisera was dialyzed against i0 mM potassium phosphate buffer, pH 7.0, containing 150 mM NaCl. The dialyzed antisera were then applied to a protein A-Agarose column (ix5 cm). Following washing with the phosphate buffer, the bound antibody was eluted with a i00 mM glycine-HCl buffer, pH 2.5. The eluted antibody was then extensively dialyzed against the phosphate buffer prior to use in the inhibition assay.
Results and Discussion
Golf and Graef (3) showed that NADPH-cytochrome P-450
reductase was needed to reconstitute steroid 5-alpha-reductase
591
Vol. 165, No. 2, 1989 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
activity. An alternative approach for studying the role of
NADPH-cytochrome P-450 reductase in steroid 5-alpha-reduction is
to employ an inhibitory antibody against P-450 reductase. If
S5R, like cytochrome P-450, accepts reducing equivalents from
NADPH-cytochrome P-450 reductase, then an inhibitory antibody
against P-450 reductase should be capable of affecting S5R
reductase activity.
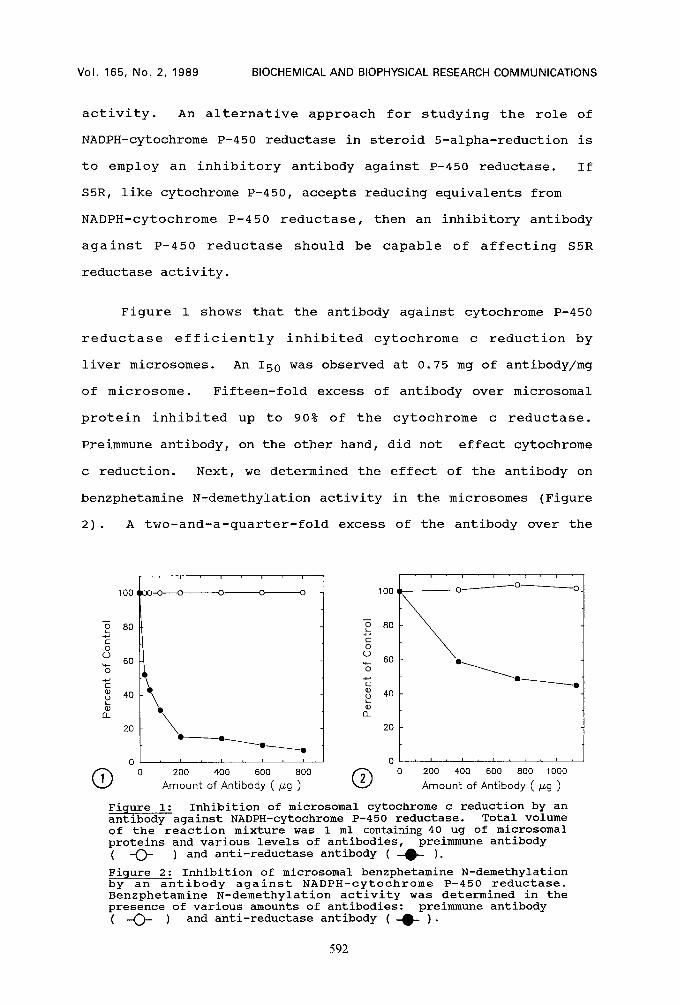
Figure 1 shows that the antibody against cytochrome P-450
reductase efficiently inhibited cytochrome c reduction by
liver microsomes. An I50 was observed at 0.75 mg of antibody/mg
of microsome. Fifteen-fold excess of antibody over microsomal
protein inhibited up to 90% of the cytochrome c reductase.
Preimmune antibody, on the other hand, did not effect cytochrome
c reduction. Next, we determined the effect of the antibody on
benzphetamine N-demethylation activity in the microsomes (Figure
2). A two-and-a-quarter-fold excess of the antibody over the
I00~
-~ 8O
¢D 6O
4O
O_ 20
l ' , ' , , • , ' , '
~o-o--o O O o 100 O~°--------------o
O 6O ~d e ~ ~ ~o~, ~ 40
• @
n 20 0-- - - I ~ S _ _ _ ~ _ ~ _ . O
0 I I I I 0 I I I I I
2 0 0 4 0 0 6 0 0 ~ 0 0 Q 0 2 0 0 4 0 0 6 0 0 8 0 0 1 0 0 0
A m o u n t of Ant ibody ( ~ g ) A m o u n t of Ant ibody ( ~g )
Figure i: Inhibition of microsomal cytochrome c reduction by an antibody against NADPH-cytochrome P-450 reductase. Total volume of the reaction mixture was 1 ml containing 40 ug of microsomal proteins and various levels of antibodies, preimmune antibody ( -O- ) and anti-reductase antibody ( -~- ).
Figure 2: Inhibition of microsomal benzphetamine N-demethylation by an antibody against NADPH-cytochrome P-450 reductase. Benzphetamine N-demethylation activity was determined in the presence of various amounts of antibodies: preimmune antibody ( -O- ) and anti-reductase antibody ( -~ ).
592
V o l . 165 , N o . 2, 1 9 8 9 B IOCHEMICAL A N D BIOPHYSICAL RESEARCH C O M M U N I C A T I O N S
microsomes inhibited the enzyme activity by about 55%, whereas
preimmune antibody again did not affect the enzyme activity.
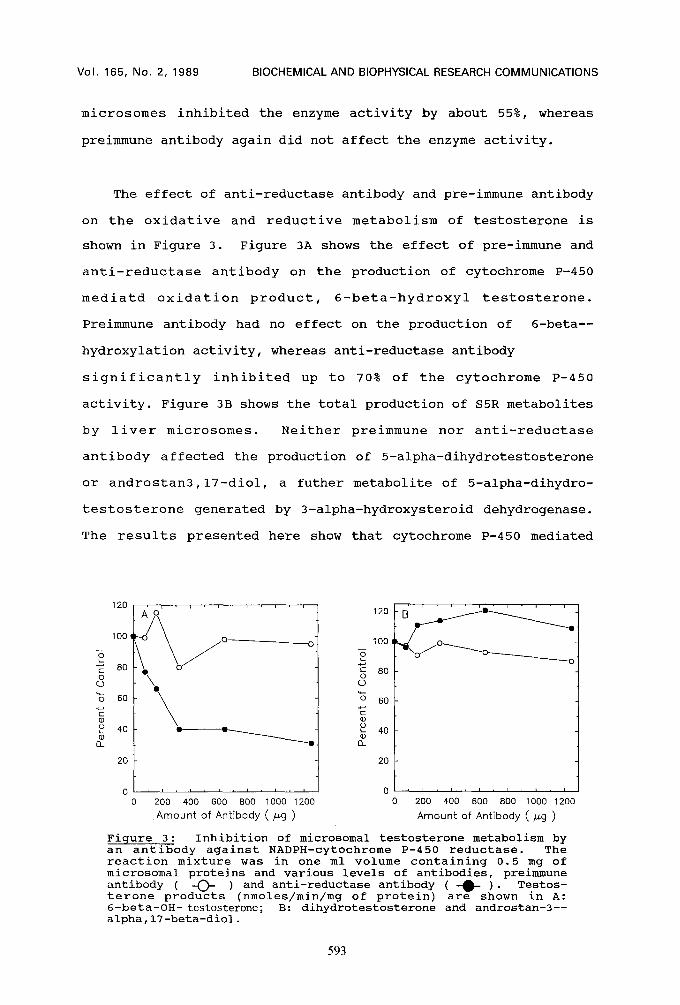
The effect of anti-reductase antibody and pre-immune antibody
on the oxidative and reductive metabolism of testosterone is
shown in Figure 3. Figure 3A shows the effect of pre-immune and
anti-reductase antibody on the production of cytochrome P-450
mediatd oxidation product, 6-beta-hydroxyl testosterone.
Preimmune antibody had no effect on the production of 6-beta--
hydroxylation activity, whereas anti-reductase antibody
significantly inhibited up to 70% of the cytochrome P-450
activity. Figure 3B shows the total production of S5R metabolites
by liver microsomes. Neither preimmune nor anti-reductase
antibody affected the production of 5-alpha-dihydrotestosterone
or androstan3,17-diol, a futher metabolite of 5-alpha-dihydro-
testosterone generated by 3-alpha-hydroxysteroid dehydrogenase.
The results presented here show that cytochrome P-450 mediated
120 ' ' ' ' . . . . " ' ' ' 120
1 0 0 f ! / ~ o / ° o 100
80 ~ 80 o \ o
~d 60 ~ ~ 60
40 • • ~ 40 el_ • [3_
20
0 I I I I i I I 0
0 200 400 600 800 1000 1200 0 A m o u n t of Ant ibody ( ~g )
20
B ' ' ' -
O ~ " " ~ O
"i/~/~/°~o ~ o ~ o
, i i i i I ] ~ I
200 400 600 800 1000 1200
A m o u n t of Ant ibody ( ~g )
Figure 3: Inhibition of microsomal testosterone metabolism by an antibody against NADPH-cytochrome P-450 reductase. The reaction mixture was in one ml volume containing 0.5 mg of microsomal proteins and various levels of antibodies, preimmune antibody ( -O- ) and anti-reductase antibody ( -0- )" Testos- terone products (nmoles/min/mg of protein) are shown in A: 6-beta-OH- testosterone; B: dihydrotestosterone and androstan-3-- alpha,17-beta-diol.
593

Vol. 165, No. 2, 1989 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
reactions, which depend upon NADPH-cytochrome P-450 reductase,
are affected by the anti-reductase antibody. The fact that S5R
activity is not affected by the anti-reductase antibody, there-
fore, suggests that S5R is independent of NADPH-cytochrome P-450
reductase.
During the course of our study, Ichihara and Tamaka reported
that progesterone 5-alpha-reduction by liver microsomes was not
affected by antibody against NADPH-cytochrome P-450 reductase
(6). Their results are consistent with our findings that only
cytochrome P-450 products, and not those of S5R or 3-alpha--
hydroxyldehydrogenase, are affected by anti-reductase anti-
body.
Acknowledqments
We are grateful to Dr. B.S.S. Masters for providing the antibody for our preliminary studies. K.-C. Cheng is partially supported by a JFRA from the American Cancer Society and a Cornell Scholar Award.
References
i. Marrini, L. (1982) Endocrine Rev. 3:1-25. 2. Liang, T., Cheung, A. H., Renolds, G. D. and Rasmusson, G. H.
(1985) J. Biol. Chem. 260: 4890-4895. 3. Golf, S. W. and Graef, V. (1978) J. Steroid Biochem. 9:
1087-1092. 4. Nozu, K. and Tamaoki, B. (1974) Acta Endocrinol. (Copenhagen)
76: 608-624. 5. Frederikson, D. W. and Wilson, J. D. (1971) J. Biol. Chem.
246: 2584-2593. 6. Ichihara, K. and Tanaka, C. (1987) Biochem. Internatl. 15:
1005-1011. 7. Cheng, K-C. and Schenkman, J. B. (1983) J. Biol. Chem. 258:
11738-11744. 8. Yasukochi, Y. and Masters, B. S. S. (1976) J. Biol. Chem. 251:
5337-5344. 9. Thomas, P.E., Lu, A.Y.H., Ryan, D., West, S.B., Kawalek, J.
and Levin, W. (1976) J. Biol. Chem. 251: 1385-1391.
594





























