Embed Size (px)
Citation preview

Molecular and Cellular Biochemistry 51, 123-127 (1983). © 1983, Martinus Nijhoff Publishers, Boston. Printed in The Netherlands.
N A D P H / N A D P + ratio: regulatory implications in yeast glyoxylic acid cycle
Jorgina Satrustegui, Juan Bautista and Alberto Machado Departamento de Bioqu[mica y Biologia Molecular, Centro de Biologia Molecular, Universidad Aut6noma, Cantoblanco, Madrid-34, Spain
Summary
The utilization by yeast of two carbon sources is carried out through the operation of the glyoxylic acid cycle. Kinetic data from the isocitrate transforming enzymes suggest that the flow of isocitrate through the glyoxylic acid cycle depends upon the inhibition of the isocitrate decarboxylating enzymes. Both isocitrate dehydrogenases are inhibited by a mixture ofglyoxylate + oxaloacetate, but for the reasons described in the text we consider that this inhibition is of no physiological significance. On the other hand, we have found that N A D P H is a competitive inhibitor of NADP-isocitrate dehydrogenase with respect to NADP +, with a K I similar to its K M. It also produces an additive effect on the NADH-produced inhibition of NAD-isocitrate dehydrogenase. We propose N A D P H as the compound that channels the utilization of isocitrate into the glyoxylic acid cycle. This is supported by the finding of an increased N A D P H / N A D P + ratio in acetate grown yeast with respect to glucose grown cells.
Introduction
The glyoxylic acid cycle is an anaplerotic path- way that provides C 4 acids when two-carbon sub- strates such as acetate and ethanol are used as car- bon sources. The cycle involves several Krebs cycle steps as well as two additional specific enzymes, isocitrate lyase (E.C. 4.1.3.1) and malate synthase (E.C. 4.1.3.2). Isocitrate lyase (IL) catalyzes the cleavage of isocitrate to glyoxylate and succinate thus bypassing the two decarboxylating steps in the tricarboxylic acid cycle. The first of these steps is catalyzed by NAD- and NADP-isocitrate dehy- drogenases ( IDH). The channeling of isocitrate through I D H is strongly favoured over that flowing through the IL-glyoxylate pathway, in different or- ganisms ( Tetrahymena pyriformis (1), E. coli (2)) since I D H s have a higher activity and lower K M for isocitrate than IL (3). We have made a study of the isocitrate branchpoint in baker 's yeast in order to know whether regulation at the level of isocitrate
oxidation is also needed in order allow the opera- tion of the glyoxylate cycle in this organism.
Our results show that isocitrate dehydrogenases inhibition is indeed required in order to allow the function of glyoxylate cycle. We present evidence leading to the suggestion that N A D P H is the physi- ological inhibitor involved in that process.
Materials and methods
Chemicals
NADH, N A D P H , glucose-6-phosphate and malate dehydrogenases, were obtained from Boeh- ringer, and DL-isocitrate, oxoglutarate (monoso- dium salt), NAD +, N A D P +, lactate and glutamate dehydrogenases were obtained from Sigma. Other reagents were of the highest purity available from commercial sources.

124
Yeast growth and harvest conditions
A diploid strain of Saccharomyces cerevisiae was used. The cells were grown at 30 °C in minimal medium supplemented with Bacto Yeast Nitrogen base w / o amino acids (Difco) and 2% acetate o r2% glucose. Cultures were aerated by mechanical shak- ing. The cells were harvested at a round 1,2 mg dry weight /ml (20 and 48 hours for glucose and acetate- grown cells, respectively). The glucose concentra- t ion at harvest was 15-40 mM. Crude extracts were obtained by cell disruption in a MSK (B. Braun) cell homogenizer at 2 000 rpm for 1 min, and cen- trifuged at 5 000 X g for 10 min. Unless otherwise stated, supernatants were used for enzyme assays.
Assay methods
A yeast NAD-isoci t ra te dehydrogenase prepara- tion essentially free f rom NADP-isoc i t ra te dehy- drogenase was obtained f rom an 0.1 M H CO3Na extract of e thanol-grown yeast by a m m o n i u m sul- fate precipitation. 28% (NH4) 2 SO 4 was added to the centrifuged extract (30 000 g, 30 min); and the pellet was resuspended in 0.1 M phosphate buffer pH 7.0 and again precipitated in 28% (NH4) 2 SO4. The resulting pellet was redissolved in the same buffer, and dialyzed over night. N A D - and N A D P - isocitrate dehydrogenase activities were determined by the method of Machado et al. (4). The reaction mixtures contained 1.0 ml 0.1 M phosphate buffer (KH2PO4-K2HPO4) pH 7, 0.1 M MgC12, 0.2 ml 50 mM DL-isocitrate, and the specific nucleotides of each assay in a final volume of 2.8 ml. The N A D + assay conta ined 0.3 ml of 0.1 M N A D + and, when added, 0.3 ml of 25 mM A M P . The NADP- l i nked activity was determined by adding0.3 ml of 2.5 m M N A D P +. Isocitrate lyase was assayed following Dixon et al. (5),
Determination o f metabolites
Samples of 100 or 200 ml of yeast cultures were rapidly gathered in precooled flasks and imme- diately immersed in liquid nitrogen. The frozen cultures were lyophilized, weighed, resuspended in acid or alkaline solutions (6), homogeneized in a MSK (B. Braun) cell homogenizer at 2 000 rpm for 2 min, and used as alkaline or acid extracts for the determinat ion of metabolites following the meth-
ods of Saez and Lagunas (6) and Bergmeyer (7). Measurements were made with a Unicam SPI700 Ultraviolet Spec t rophotometer and a Gilson oxy- graph K-IC. The dry weights of the corresponding cultures were obtained by filtration of 25 ml sam- ples and metaboli te concentrat ions expressed as #moles g dry weight -1 . In order to obtain the actual cellular concentrations, the intracellular volume was estimated through the distribution of ~4C-polie- thylenglycol and 3H20 (The Radiochemical Centre, Amersham) essentially as in (8).
Results and discussion
A ctivities and affinities for isocitrate o f NA D- and NADP-isocitrate dehydrogenases and isocitrate lyase in relation with isocitrate levels.
Table 1 shows the activities of different enzymes f rom yeast cells grown on glucose and acetate as sole carbon sources, In agreement with previous reports (9), IL was strongly induced in acetate cul- tures. This resulted in an increase in the I L / N A D P - I D H ratio in these cultures. The affinity for isoci- trate (Table 1) is strongly favourable to both IDHs . This difference in affinity has been reported in al- most every organism with both enzyme activities. The K M values for isocitrate of N A D P - I D H lie between 8 X 10 -6 M in Chlamydomonas (10)and 2 X 10 5 M in Blastocladiella (11); those of IL range f rom 3.3 X 10 3 M in Neurospora crassa (12) to 2.3 X 10 4 M in Chlorellapyrenoidosa (13). According
Table t. NAD , NADP isocitrate dehydrogenase and isocitrate lyase activities and affinities for isocitrate from glucose and acetate grown yeast.
2% glucose 2% ace ta te K M isocitrate
NAD-lsocitrate dehydrogenase AMP
+2.5 mM AMP NADP-isocitrate
dehydrogenase
lsocitrate lyase
22 38 2.5 X 10 -4 M 1.5 X 10 5 M
108 167 4.0 X 10 -5 M
36 124 1.3 X 10 -3 M
Enzyme activities are expressed in nmoles X rain -l X mg proF l . The results are the means of 3 experiments. K M for DL-isocitrate were obtained after Lineweaver-Burk.

to their affinities and maximal activities, and the actual isocitrate concentrations, the contribution of each reaction to net isocitrate metabolism can be estimated. Intracellular citrate concentrations were found to be 1.4 and 8.3 nmoles /mg dry weight (see Materials and methods). The intracellular volume under different culture conditions was estimated to be around 1.55 #1/mg dry weight. Accordingly, the actual citrate concentrations in glucose and acetate grown yeast are 0.90 and 5.35 mM respectively. Assuming that aconitase is at equilibrium (7% isoci- trate, 93% citrate), isocitrate concentrations would be 0.06 X 10 -3 and 0.40 X 10 3 M in glucose and acetate grown yeast, respectively, although the con- centrations in specific compartments could be slightly different. According to their kinetic con- stants NAD- and NADP-isocitrate dehydrogenases would be almost fully saturated (depending on AMP levels) whereas isocitrate lyase would be well below saturation both in glucose or acetate cul- tures. In addition, NADP-isocitrate dehydrogenase has a higher activity than isocitrate lyase. More- over, a different cellular compartmentat ion of the various enzymes involved in isocitrate utilization has not been found in this organism; NADP-isoci- trate dehydrogenase is located in cytosol and also in glyoxysomes of Tetrahymena (14) and yeast (15). Thus isocitrate flux would be largely channelled via dehydrogenation instead of glyoxylate formation under both conditions. This supports the need of a strong inhibition of N A D P - I D H to allow the func- tion of the glyoxylate cycle.
Concerted inhibition of isocitrate dehydrogenase
The inhibition of isocitrate dehydrogenase by the concerted action of oxalacetate and glyoxylate has been assumed to act as the main regulating mecha- nism allowing glyoxylate cycle function.
Table 2 shows the inhibition of isocitrate dehy- drogenase by glyoxylate and oxaloacetate. The concentrations of these products were fixed accord- ing to those employed by other authors (16). The mxxture of both components each at a concentra- tion of 0.06 mM produced a 31% inhibition of N A D P - I D H . NAD-isocitrate dehydrogenase is strongly inb.ibited by oxaloacetate at the concentra- tions used in the concerted inhibition assay.
The concerted inhibition by glyoxylate and ox- aloacetate affects a large number of isocitrate de-
125
Table 2. Effect in glyoxylate and oxaloacetate on yeast NADP- isocitrate dehydrogenase.
Compounds added M X 10 3 % Activity
None 100 Oxaloacetate 3.3 45 Glyoxylate 3.3 50 Oxaloacetate 0.12 100 Glyoxylate " 0.12 95 Oxaloacetate + glyoxylate 0.12 + 0.12 74 Oxaloacetate + glyoxylate* 0.12 + 0.12 50 Oxaloacetate 0.06 100 Glyoxylate 0.06 95 Oxaloacetate + glyoxylate 0.06 + 0.06 95 Oxaloacetate ÷ glyoxylate* 0.06 + 0.06 69
* This combination of ketoacids was mixed and kept at room temperature from 20 rain before addition to the reaction mix- ture.
hydrogenases (11, 13, 16), and is thought to be brought about by the condensation product of ox- aloacetate and glyoxylate, namely oxalomalate (17). In agreement with this, the actual inhibition observed has been shown to depend on a number of experimental conditions affecting the assay, a) The inhibition of the pig heart enzyme by glyoxylate and oxaloacetate is greatly enhanced when the en- zyme is preincubated with the mixture of the two compounds (18 and Table 2 this paper), b) Inhibi- tion is observed when the reaction is started with the addition of isocitrate, but a concentration one order of magnitude higher of the mixture of inhibi- tors is required to produce the same effect when the reaction is started with N A D P (19). In addition, the inhibition has been shown to be non-revertible by lactate or malate dehydrogenases that consume glyoxylate and oxaloacetate respectively (18). This indicates that, at least in vitro, the concerted inhibi- tion is irreversible, with oxalomalate behaving as a structural analogue of isocitrate. Besides, the pre- sumptive physiological significance of concerted inhibition by glyoxylate and oxaloacetate faces the following problems, a) How is glyoxylate produced when isocitrate lyase is still repressed? b) How is inhibition released with the continuous production of glyoxylate? c) How is the concerted inhibition of isocitrate dehydrogenase accomplished if this en- zyme is localized in glyoxysomes (15, 20) and the enzymatic source of oxaloacetate, malate dehy- drogenase, has not been found in this organelle (21)?

126
Inhibition by NADPH of yeast NADP-isocitrate dehydrogenase
We have found that N A D P H is a s t rong compet i - tive inh ib i to r o f N A D P - I D H with respect to N A D P +, wi th a K I of 4.0 X 10 -5 M, s imilar to its K M for N A D P +, 2.6 )< 10 5 M. S imi la r results have been r epor t ed for the enzyme of o ther sources, Te- trahymena (22) and Blastocladiella (11). Glucose 6-phosphate and 6-phospho-g lucona te dehydrogen- ases are also inh ib i ted by N A D P H , and the cel lular N A D P H / N A D P + ra t io is cons idered the ma jo r regu la tory pa rame te r act ing on the pentose phos- pha te cycle. We p ropose tha t the inhib i t ion by N A D P H of N A D P - i s o c i t r a t e dehydrogenase also plays a regu la tory role on this enzyme, s imilar to tha t pos tu l a t ed for the pentose phospha te cycle dehydrogenases .
Inhibition by NADPH of NAD-isocitrate dehydrogenase
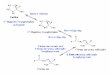
A n y p r o p o s e d r egu la to r of the g lyoxyl ic acid pa thway t h rough an inh ib i t ion of i soci t ra te ox ida - tive d e c a r b o x y l a t i o n should also p roduce the inhi- b i t ion of N A D - I D H , the th i rd enzyme ut i l iz ing i soci t ra te in eukaryo t i c cells. F igure 1 shows that N A D P H is also an inh ib i to r o f N A D - I D H . N A D H and N A D P H are a compet i t ive inh ib i to r of yeast N A D - i s o c i t r a t e dehydrogenase with respect to N A D +. In the presence of 0.18 m M A M P , its ap- pa ren t K I are 0.023 and 0.11 raM, respectively. The presence of 0.293 nM N A D P H great ly enhances the inhib i t ion p roduced by N A D H (apparen t K I = 0.010 mM, Fig. 1). S imi la r results were ob ta ined by P lau t and col leagues for the rat liver and hear t enzymes (23, 24).
Concentrations of oxidized and reduced pyridine nucleotides in glucose- and acetate-grown yeast
The var ia t ions in N A D P H and N A D P + c o n c e n -
300
200 >,-
,.r
i -
5 100
i I J i I L I
1 2 3 4 5 6 7
I/[NAD] (raM)
Fig. 1. Effect of reduced pyridine nucleotides on yeast NAD-- isocitrate dehydrogenase. An NAD-isocitrate dehydrogenase preparation essentially free from NADP-dependent activity (Materials and methods) was used. Lineweaver-Burk plots have been obtained by changing NAD concentrations with a constant amount of DL-isocitrate (0.066 mM) and AMP (0.18 raM) in the presence of 0.058 mM NADH (O), 0.29 mM NADPH (D), 0.058 mM NADH + 0.29 mM NADPH (A), and in absence (e) of reduced pyridine nucleotides.
t ra t ions , s imi lar to those previous ly found in E. coli (2), are consis tent with the p roposed role of N A D P H as a phys io logica l inhib i tor of isoci t ra te dehydrogenases (Table 3). The N A D P H / N A D P + ra t io increases f rom 7.4 in glucose to 12 in aceta te- g rown cells. As j udged by those ra t ios and the K M and K I for N A D P + and N A D P H of isoci t ra te de- hydrogenase , the N A D P - l i n k e d enzyme would be inhib i ted under bo th condi t ions . However , this in- h ib i t ion would be increased dur ing growth on ace- tate with respect to glucose and the rise in the N A D P H / N A D P + ra t io would enhance the inhibi-
Table 3. Effect of the carbon source on the concentrations of metabolites.
Carbon NADPH NADP source
NADPH NADH NAD NADH Citrate Oxoglutarate Citrate
NADP NAD (oxoglutarate)
2%glucose 0.408±0.061 0.055_+0.007 7.4 0.472--+0.05 2.33_+0.300.20 1.40--+0.10 1.10___0.15 1.27 2%acetate 0.628-+0.07 0.052±0.006 1 2 . 0 0.717-+0.09 2.64+0.320.27 8.37_+0.91 0.58-+0.06 14.4
Concentrations are expressed in #moles >( g dry weighr I . The values are the means (of 3 experiments) -+ S.D.

tion produced by N A D H on NAD-isocitrate dehy- drogenase. Moreover, this increased inhibition has been confirmed by other means. The intracellular ci trate/oxoglutarate ratio has been used to calcu- late the N A D P H / N A D P + ratio of the free nucleo- tides through the equilibrium constant of the NADP-isoci trate dehydrogenase reaction (25), since the reactants of this dehydrogenase system are in equilibrium with the free N A D P H / N A D P + ra- tio. On the basis of the data in Table 3 and the equilibrium constant of the aconitase reaction (26), the free N A D P H / N A D P ÷ ratio of glucose grown cells would be one order of magnitude lower than that of the acetate grown cells (96 in glucose and 1090 in acetate for CO2 concentration of 1.16 raM, (25)). This difference in free ratios amplifies that found with the total ratios and supports the pro- posed role o f N A D P H / N A D P as a regulator of the glyoxylic acid cycle function.
Acknowledgements
This work was supported by grants from the Fondo de Investigaciones Sanitarias and f rom the Comisi6n Asesora de Investigaci6n Cientifica y T6cnica. We thank Miss Maria Victoria Mora Gil for her technical assistance, and Dr. Gerald Frank for his help in correcting the manuscript.
References
1. Machado, A. and Satrflstegui, J., 1981. Biochimie 63: 247 249.
2. Bautista, J., Satrtlstegui, J. and Machado, A., 1979. FEBS Letters 105:333 336.
127
3. Cooper, T. G. and Beevers, H., 1969. J. Biol. Chem. 244: 3507-3513.
4. Machado, A., Nflfiez de Castro, 1, and Mayor, F., 1975. Mol. Cell. Biochem. 6: 93-100.
5. Dixon, G. and Kornberg, H. L., 1959. Biochem. J. 72:3 p. 6. Saez, H. J. and Lagunas, R., 1976. Mol. and Cell. Biochem.
13:73 78. 7. Bergmeyer, H. U., 1974. Methods in enzymatic analysis. Vol.
III y IV. 2nd ed. Academic Press, New York. 8. Rottenberg, H., 1979. Methods in Enzymology vol. LV, 549. 9. Kornberg, H. L., 1966. Biochem. J. 99:1 11.
10. Foo, S. S. K. and Badour, S.S. , 1977. Can. J. Bot. 55: 2178 2185.
11. Ingebretsen, O. C., 1975. J. Bacteriol. 124: 65-72. 12. Johanson, R. M., Hill, J. M. and McFadden, B. A., 1974.
Biochim, Biophys. Acta 364: 327-340. 13. John, P. C. L. and Syrett, P . J . , 1967. Biochem. J. 105:
409-416. 14. Muller, M., Hogg, J. F. and de Duve, C., 1968. J. Biol.
Chem. 243:5385 5395. 15. Fukui, S. and Tanaka, A., 1979. Trends in Biochem. Sci, 4:
246-249. 16. Satoh, Y., 1972. Plant Cell Physiol. 13:493 503. 17. Ramaley, F. R. and Hudock, M.O. , 1973. Biochem. Bio-
phys. Acta 315:22 36. 18. Ruffo, A., Moratti, R., Montani, A. and Melzi D'Eril, G. L.,
1974. Ital. J. Biochem. 23:357 370. 19. Levy, M. R., 1972. Arch. Biochem. Biophys. 152:463 471. 20. Tanaka, A., Nabeshima, S., Tokuda, M. and Fukui, S.,
1977. Agric. Biol. Chem. 41: 795-799. 21. Kawamoto, S., Tanaka, A., Jamamura, M., Teranishi, J.,
Fukui, S. and Osumi, M., 1979. Arch. Microbiol. 112: 1-8. 22. Vidal, P. and Machado, A., 1977. Mol. Cell. Biochem. 17:
151-156. 23. Chen, R .F . and Plaut, G. W. E., 1963. Biochemistry 2:
1023-1032. 24. Plaut, G. W. E. and Aogaichi, T,, 1968. J. Biol. Chem. 243:
5572 5583. 25. Veech, R. L., Egleston, R. V. and Krebs, H. A., 1969. Bio-
chem. J. 115:609 619. 26. Garland, P. B. and Randle, P .J . , 1964. Biochem. J. 93:
678-687.
Received 13 July 1982.