Embed Size (px)
Citation preview

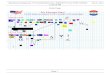
New material
• The following slides will be posted on Blackboard
• They cover in more detail and with some good examples material we’ve already introduced
• But they also cover some new material on cells, signaling and energetics

Intermediate filament — DYSTROPHIN
• In muscle, Dystrophin binds actin and dystrophin glycoprotein complex, a Golgi-secreted complex that lies outside cell
• Either failure to glycosylate dystroglycan OR mutations in dystrophin can lead to MUSCULAR DYSTROPHY, destroying muscle cells over time.
The extracellular matrix, composed of collagen and protein-carbohyrate complexes (proteoglycans) stabilizes or orients tissues by binding to cell ADHESINS like dystroglycan

Microtubulesa-tubulin and b-tubulin form dimers that then, in groups of 13, form a microtubule. Like actin filaments, they can polymerize at one end and depolymerize at the other to lead to movement.
Also, microtubules can be connected by flexible NEXIN crosslinks. When DYNEIN, a “motor protein” moves, the nexin crosslinks cause the 9+2 arrangement of microtubules to bend, found in CILIA and eukaryotic FLAGELLA.

Phospholipids orient themselves in H2O as AMPHIPATHIC molecules, partly dissolved in H2O and partly “dissolved” in lipid oil. Biological membranes construct their own “oil”, which is the interior of the phospholipid membrane. The red phospholipid polar heads are dissolved in water while the hydrocarbon (hydrophobic) tails form lo-energy hydrophobic interactions. Like an oil slick, the phospholipids slide around by each other laterally, very rapidly, leading to an oily “fluid” membrane
OUTSIDE of cell
INSIDE of cell

Integral proteins span the membrane. Regions of membrane, called RAFTS, are associated with particular proteins groups. Some proteins (caveolin) lead to pocket formation and beginning of endocytosis.
Caveolin and start of vesicle formation
Membrane raft Phospholipids spontaneously form vesicles in aqueous solutions, trapping H2O other molecules inside and forming the basis of all cells


SUMMARY: CAVEOLIN causes membrane bilayer to form a pit or to “cave in”. Once that begins to happen, pit may be coated with CLATHRIN, resulting in a “coated pit” that forms a vesicle. Clathrin is shed and vesicle may fuse with LYSOSOME for hydrolysis of contents or have some other fate.
Virus can enter cells through normal vesicles
Cholesterol is brought into cells via vesicles

AQUAPORINS are specific H2O channels that significantly increase the diffusion rate of water across membranes. Some membranes allow diffusion of water very poorly, and such membranes (in kidney, for example) rely exclusively on aquaporins for water movement. Aquaporins have very small channels that are lined with polar amino acid R-groups. Aquaporins can easily flex to close the water channel – very little movement is needed – which makes aquaporins susceptible to REGULATION by cofactors or hormones.
water
water

MEMBRANE RECEPTORSAdditional vocabulary
• Proteins, e.g. enzymes, often bind other molecules by virtue of their shape and aa functional groups.
• The molecule that remains primarily stable in space is called a RECEPTOR.
• The molecule that binds to the receptor is a LIGAND.
• The process is RECEPTOR-LIGAND BINDING
• Receptor-ligand binding is through the affinity of weak bonds (H-bonds, ionic bonds)
• Some ligands are substrates for chemical reactions (receptor is an enzyme catalyst)
• Other ligands participate in a catalytic reaction (enzyme cofactors)
• Other ligands change the shape (conformation) of the protein receptor and launch secondary reactions. These ligands are SIGNAL molecules.

cAMP (second messenger) commonly activates another enzyme, which catalyzes a reaction whose product activates a second enzyme, and so on and so on. Together, these enzymes constitute a CASCADE of reactions.
The final enzyme in the epinephrine cascade hydrolyzes glycogen to glucose
Enzymes in red are inactive, and enzymes in green are active
An enzyme cascade also results in amplification, since each step is a catalytic one, and multiple copies of the product are formed. About 10,000 molecules of glucose are released for each 1 molecule of epinephrine
G-protein signaling in detail

• G-proteins are active in virtually all living cells
• One is involved with transducing light to cellular changes in vision
• In this case light is the first signal (messenger) and cGMP is the second messenger.
• cGMP or cAMP — both are inactivated by hydrolysis by key enzyme, PHOSPHODIESTERASE, turning off the effect of original signal.

CAFFEINE and Adenosine have similar structures. Adenosine is also the “A” part of cAMP and ATP. Adenosine is a signal molecule in brain cells, reducing brain activity (increasing sleepiness) during excessive stress (hopefully). Caffeine is a competitive inhibitor of brain adenosine receptors and reduces ability of adenosine to induce sleepiness.Caffeine also gets into cells and can fit into active site of phosphodiesterase, reducing enzyme activity and keeping cAMP levels artificially high.

Intracellular Receptors• Lipid-soluble
molecules like steroids cross membranes easily
• Notice the steroid hormone signal (ligand) binds an intracellular receptor, not a membrane receptor
• Hormone-receptor complex travels to nucleus and causes genetic changes