Embed Size (px)
Citation preview

975
Alberto M. STCHIGEL, Lidia FIGUERA, Jose! CANO and Josep GUARRO
Unitat de Microbiologia, Facultat de Medicina i Cie[ ncies de la Salut, Universitat Rovira i Virgili, C}Sant Llorencn 21,
43201 Reus, Spain.
E-mail : umb!astor.urv.es
Received 5 December 2001; accepted 18 June 2002.
Two new thermotolerant species of Thielavia were isolated from Easter Island and India, after treating soil samples
with 65% (v}v) ethyl alcohol : T. intermedia sp. nov. characterized by large, obovate, dark brown ascospores with a
terminal germ pore; and T. arxii sp. nov. distinguished by the large, ellipsoidal to fusiform, brown ascospores with a
prominent, lateral to medial germ pore. A phylogenetic study of the ITS-5.8S rDNA sequences of 18 species of
Thielavia and Melanocarpus thermophilus was performed. This analysis revealed that T. intermedia could be the
ancestor of the remaining species and that all the species tested were genetically closely related.
INTRODUCTION
Thielavia (Chaetomiaceae, Sordariales) is characterized
by spherical, non-ostiolate ascomata, with a thin
peridium, and one-celled, darkly pigmented ascospores.
Revisions of this genus have been proposed by Booth
(1961), Mouchacca (1973), Malloch & Cain (1973), von
Arx (1975), and von Arx, Figueras & Guarro (1988).
Von Arx (1973) excluded T. sepedonium and similar
species with ascospores with two germ pores and
Myceliophthora anamorphs, and erected the genus
Corynascus to accommodate these species. The last,
most comprehensive, revision by von Arx et al. (1988)
accepted 14 species. Subsequently, two more species
have been described (Chen & Chen 1996, Ito, Okane &
Nakagiri 1998). Here we describe two new species
recently isolated from Easter Island and Indian soils
respectively, and compare them molecularly with other
species of the genus.
MATERIALS AND METHODS
Collection and specimen isolation
Indian soil samples were collected close to Jaipur,
Rajasthan State, a tropical semiarid region dominated
by a hot climate ; the average temperature is 23 °C in
winter and 35 ° in summer, and the total annual
precipitation about 900 mm. Vegetation is mainly
grasses and shrubs. Easter Island is a triangular,
volcanic, semiarid island of 162 km# located in the
middle of the Pacific Ocean. Soil samples were collected
near Hanga-Roa city. The vegetation is mainly grasses,
with reduced forest masses composed of a few imported
trees, such as Eucalyptus spp. and Melia azederach. The
area is dominated by a subtropical maritime climate ;
the average temperature is 17±8 ° in winter and 23±7 ° in
summer. The total annual precipitation is about
1140 mm, with no dry season.
Collections were taken mainly from the superficial
layer of soil with sterilized polyethylene bags. These
were closed with rubber bands and labelled. On their
arrival at the laboratory the materials were stored at
4–7 ° until used. Approx. 1 g of soil was treated with
65% (v}v) ethyl alcohol according to Warcup & Baker
(1963). The suspensions were cultured on tap water
agar (TWA; agar, 20 g; tap water, 1000 ml) and
incubated at 42 ° under 12 h darkness alternating with
12 h cool white fluorescent light.
Cultural and morphological characterization
The morphological study was performed by growing
the isolates on oatmeal agar (OA; Difco), potato carrot
agar (PCA; potatoes, 20 g; carrot, 20 g; agar, 20 g; tap
water, 1000 ml), potato dextrose agar (PDA; Difco)
and malt extract agar (MEA; Difco) at 15 °, room
temperature (22–25 °), 37 °, and 45 °, under 12 h
darkness alternating with 12 h cool white fluorescent
light. Colour notations in parentheses are from Kor-
nerup & Wanscher (1984). Measurements were per-
formed in water and lactophenol. Photomicrographs
were obtained with a Leitz Dialux 20 EB microscope.
Scanning electron microscopy techniques were as
described by Figueras & Guarro (1988).
Mycol. Res. 106 (8) : 975–983 (August 2002). # The British Mycological Society
DOI: 10.1017}S0953756202006299 Printed in the United Kingdom.
New species of Thielavia, with a molecular study of
representative species of the genus

New species of Thielavia 976
Table 1. List of strains, sources and sequences used in the molecular study.
Taxon Origin Strain no. EMBL no.
Gelasinospora bonaerensis soil ; Argentina FMR 5962 AJ002029
Melanocarpus thermophilus soil ; Irak FMR 6190 AJ271586
Thielavia appendiculata rabbit dung, UK CBS 731.68 AJ271584
T. arenaria (T) desert soil ; Egypt CBS 507.74 AJ271576
T. arxii sp. nov. soil ; Easter Island, Chile FMR 5875 AJ271580
T. australiensis (T) nesting material of incubator bird; Australia CBS 493.74 AJ271590
T. basicola on Arctostaphylos uva-ursi ; Switzerland CBS 229.82 AJ271591
T. coactilis salt marsh; Kuwait IMI 182707 AJ271585
T. fragilis (T) rhizosphere of Pennisetum thyphoideum ; India IMI 169641 AJ271578
T. hyalocarpa desert soil ; Egypt CBS 646.74 AJ271583
T. hyrcaniae Niger CBS 773.85 AJ271581
T. intermedia sp. nov. soil ; India FMR 5594 AJ271588
T. microspora saline desert soil ; Kuwait CBS 808.73 AJ271577
T. minuta cotton jacket ; Switzerland CBS 541.76 AJ271587
T. ovispora (T) root of Avena sativa ; Ukraine CBS 165.75 AJ271593
T. subthermophila on Vignia radiata ; India IMI 319643 AJ271575
T. terrestris ground soil ; IFO 9732 AJ271589
T. terricola on Polygala senega ; IFO 30068 AJ271579
T. minor on leaf of Elaeis guineensis ; CBS 611.74 AJ271582
T. tortuosa soil ; India FMR 5780 AJ271592
(T), ex-types ; CBS, Centraalbureau voor Schimmelcultures ; FMR, Facultad de Medicina de Reus; IFO, Institute for Fermentation, Osaka;
IMI, CABI Bioscience UK Centre; EMBL, European Molecular Biology Laboratory.
DNA extraction, amplification and sequencing
The strains sequenced are shown in the Table 1.
Gelasinospora bonaerensis (Stchigel, Cano & Guarro
1998) was used as outgroup. Fungal DNA was isolated
as described by Guillamo! n et al. (1996). Strains were
grown at 28 ° in Sabouraud broth (Difco) in Ehrlen-
meyer flasks shaken at 20 rpm. The mycelium was
collected by filtration through nytal mesh (42 µm pore
size), washed with distilled water, blotted with paper
towels, frozen with liquid nitrogen and ground to a fine
powder with a mortar and pestle. The powder was
incubed for 1 h at 65 ° in 2 ml of extraction buffer
(80±2 m TrisHCI pH 8±0, 0±25 m NaCI, 25 m
EDTA, 0±5% SDS). The lysate was extracted with
phenol–chloroform–isoamyl alcohol solution (25:24:1,
by vol.) and DNA was recovered by isopropanol
precipitation. The pellet was washed with 70% v}v
ethanol, dried under vacuum and resuspended in TE
buffer (10 m TrisHCI pH 8±0, EDTA 1 m). The
rDNA ITS region containing ITS 1 and ITS 2 and the
intervening 5.8S rRNA gene were amplified as described
by Gene! et al. (1996), by using a Perkin Elmer 2400
thermal cycler (Perking Elmer Cetus, Emervyville, CA).
The primers ITS5 (5«-GGAAGTAAAAGTCGTAAC-
AAGG-3«) and ITS4 (5«-TCCTCCGCTTATTGATA-
TGC-3«) (White et al. 1990) were used. The amplifi-
cation program consisted of pre-denaturalisation at
94–6 ° for 5 min, 30 cycles at 95 ° for 30 s, 50 ° for 1 min
and 72 ° for 1 min, and final incubation at 72 ° for
7 min to complete the last extension. The final products
were resolved by electrophoresis in a 2% agarose MP
gel (Boehringer Mannheim), and cleaned following the
GENECLEAN II protocol (BIO 101). The molecular
weights of amplified DNA were estimated by com-
parison with 100 bp DNA leader (Gibco BRL) standard
lane. The protocol Taq DyeDeoxy Terminator Cycle
Sequencing Kit (Applied Biosystems, Gouda) was used
for sequencing. Reactions were performed using the
primers ITS5 and ITS4 (White et al. 1990) and were run
on a 310 DNA sequencer (Applied Biosystems).
Nucleotide sequence alignment and analyses
The sequences compared in this paper were aligned
using CLUSTAL W, version 1.5, of multiple sequence
alignment computer program (Thompson, Higgins &
Gibson 1994). Cladistic analyses using the neighbour-
joining method (Saitou & Nei 1987) with confidence
values for individual branches determined by bootstrap
analyses (500 pseudoreplicates) was performed with the
MEGA 1.0 program (Kumar, Tamura & Nei 1993).
The nucleotide composition, the observed frequencies
from pairwise comparisons, and the alignment gap
sequences were performed with the MEGA 1.0 pro-
gram. The EMBL accession numbers of the sequences
are in Table 1.
RESULTS
Taxonomy
Thielavia intermedia Stchigel & Guarro, sp. nov.(Figs 1–8)
Mycelium ex hyphis hyalinis vel brunneis, ramosis, anasto-
mosans, septatis, 1–5 µm diam composito. Coloniae 22–25 °Cin agaro cum decocto tuberorum et carotarum crescentes
supra 70 mm diam post 14 d, brunneae, expansae, planae,
granulosae; reversum brunneum; sine exsudatum. Ascomata
superficialia vel immersa, globosa, non-ostiolata, translucida,
atrobrunnea vel nigrae, glabrae, 130–300 µm diam, peridium
7–8 stratiorum compositum, 10–15 µm crassum, ex textura

A. M. Stchigel and others 977
1
2
3
Figs 1–3. Thielavia intermedia (FMR 5594). Fig. 1. Detail of the peridium. Fig. 2. Asci with inmature ascospores.
Fig. 3. Ascospores. Bar¯ 10 µm.
epidermoidea vel textura intrincata compositum; paraphisibus
nullis. Asci octospori, sub-globosi vel late ellipsoidei, brevi-
stipitati, 30–40¬22–30 µm, tenuitunicati, evanescentibus.
Ascosporae obovatis vel limoniformis, brunneae, laevis,
12–17¬9–12 µm, cum poro germinalibus apicalis unicus,
protrudente. Status conidialis nullis.
Typus : India : Rajasthan State : Jaipur, 26° N, 75° E, isol.ex
soil, 29 Oct. 1995, J. Guarro [isol. A. M. Stchigel ] (IMI
370868 – holotypus, FMR 5594 – cultura ex-typo).
Mycelium composed of hyaline to brown, branched,
anastomosing, septate, smooth hyphae, 1–5 µm broad.
Colonies on PCA attaining over 70 mm diam in 14 d at
room temperature, brown (M 6E4), expanding, flat,
granulose due to ascomata production; the reverse
present the same color than the surface; exudate and
soluble pigment absent. Ascomata superficial to im-
mersed, globose, non-ostiolate, translucent, dark brown
to black, glabrous, 130–300 µm diam; peridium 7–8-
layered, 10–15 µm thick, textura epidermoidea to textura
intrincata, composed of thickened, brown to dark
brown, irregular cells, 4–15¬3–13 µm; paraphyses
absent. Asci 8-spored, subglobose to broadly ellipsoidal,
short-stipitate, 30–40¬22–30 µm, thin-walled, evan-
escent. Ascospores obovate to limoniform, hyaline when
young and dark brown when mature, one-celled,
smooth walled, 12–17¬9–12 µm, with one distinctive,
apical, protruding, 1 µm diam, germ pore. Anamorph
not observed.
Colonies on OMA growing at room temperature
show the same characteristics as those on PCA. Colonies
on MEA growing at room temperature, attaining a
diam over 70 mm in 14 d, raw umber (M 5F8),
expanding, flat to slightly cottony, granulose by
ascomata production; exudate hyaline; reverse raw
umber (M 5F8); soluble pigment orange yellow.
Colonies on PDA growing at room temperature,
attaining a diam over 70 mm in 14 d, yellowish white to
greyish yellow (M 4A2 to 4B3), cottony, zonate ;
exudate hyaline; reverse raw umber to deep yellow
(M 5F8 to 4A8); soluble pigment orange yellow. Asco-
mata sterile produced. Thermotholerant. At 37 ° grow-
ing rapidly, producing white colonies, ascomata absent.
At 45 ° and at 15 ° growing scarcely, producing white
colonies ; ascomata absent. No growth below 12 °.The main features of the T. intermedia ITS 1-2 and
5.8S rDNA region sequences are : 498 bp; 117 A;
152 C; 124 G and 105 T. The location of the ITS 1
region is from nucleotides 7 to 154, the gene 5.8S rRNA
from nucleotides 155 to 328, and the ITS 2 region from
nucleotides 329 to 465.
T. intermedia shows certain similarities with T. fragilis
and T. ovispora. All have obovate ascospores, but these

New species of Thielavia 978
4 5
6
87
Figs 4–8. Thielavia intermedia (FMR 5594). Fig. 4. Detail of the peridium. Fig. 5. Asci. Figs 6–7. Mature ascospores.
Fig. 8. Ascospores (SEM). Bar Figs 4–7¯ 15 µm, Fig. 8¯ 12±5 µm.
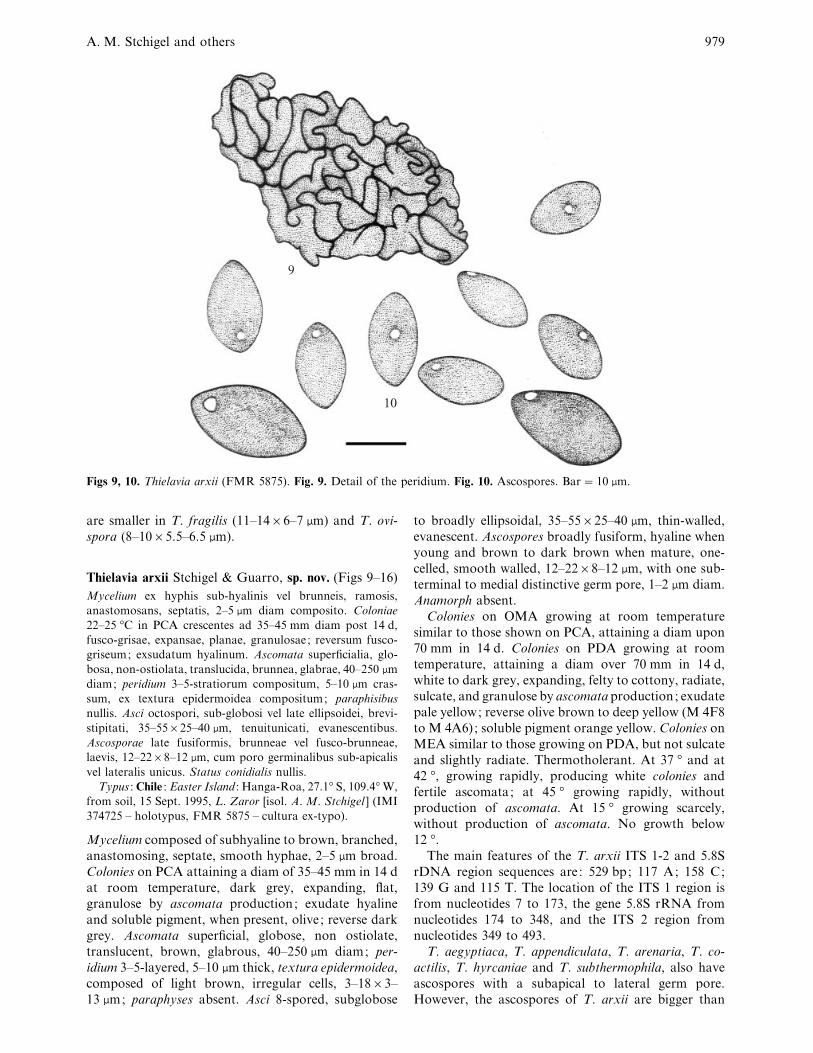
A. M. Stchigel and others 979
9
10
Figs 9, 10. Thielavia arxii (FMR 5875). Fig. 9. Detail of the peridium. Fig. 10. Ascospores. Bar¯ 10 µm.
are smaller in T. fragilis (11–14¬6–7 µm) and T. ovi-
spora (8–10¬5±5–6±5 µm).
Thielavia arxii Stchigel & Guarro, sp. nov. (Figs 9–16)
Mycelium ex hyphis sub-hyalinis vel brunneis, ramosis,
anastomosans, septatis, 2–5 µm diam composito. Coloniae
22–25 °C in PCA crescentes ad 35–45 mm diam post 14 d,
fusco-grisae, expansae, planae, granulosae; reversum fusco-
griseum; exsudatum hyalinum. Ascomata superficialia, glo-
bosa, non-ostiolata, translucida, brunnea, glabrae, 40–250 µm
diam; peridium 3–5-stratiorum compositum, 5–10 µm cras-
sum, ex textura epidermoidea compositum; paraphisibus
nullis. Asci octospori, sub-globosi vel late ellipsoidei, brevi-
stipitati, 35–55¬25–40 µm, tenuitunicati, evanescentibus.
Ascosporae late fusiformis, brunneae vel fusco-brunneae,
laevis, 12–22¬8–12 µm, cum poro germinalibus sub-apicalis
vel lateralis unicus. Status conidialis nullis.
Typus : Chile : Easter Island : Hanga-Roa, 27.1° S, 109.4° W,
from soil, 15 Sept. 1995, L. Zaror [isol. A. M. Stchigel ] (IMI
374725 – holotypus, FMR 5875 – cultura ex-typo).
Mycelium composed of subhyaline to brown, branched,
anastomosing, septate, smooth hyphae, 2–5 µm broad.
Colonies on PCA attaining a diam of 35–45 mm in 14 d
at room temperature, dark grey, expanding, flat,
granulose by ascomata production; exudate hyaline
and soluble pigment, when present, olive; reverse dark
grey. Ascomata superficial, globose, non ostiolate,
translucent, brown, glabrous, 40–250 µm diam; per-
idium 3–5-layered, 5–10 µm thick, textura epidermoidea,
composed of light brown, irregular cells, 3–18¬3–
13 µm; paraphyses absent. Asci 8-spored, subglobose
to broadly ellipsoidal, 35–55¬25–40 µm, thin-walled,
evanescent. Ascospores broadly fusiform, hyaline when
young and brown to dark brown when mature, one-
celled, smooth walled, 12–22¬8–12 µm, with one sub-
terminal to medial distinctive germ pore, 1–2 µm diam.
Anamorph absent.
Colonies on OMA growing at room temperature
similar to those shown on PCA, attaining a diam upon
70 mm in 14 d. Colonies on PDA growing at room
temperature, attaining a diam over 70 mm in 14 d,
white to dark grey, expanding, felty to cottony, radiate,
sulcate, and granulose by ascomata production; exudate
pale yellow; reverse olive brown to deep yellow (M 4F8
to M 4A6); soluble pigment orange yellow. Colonies on
MEA similar to those growing on PDA, but not sulcate
and slightly radiate. Thermotholerant. At 37 ° and at
42 °, growing rapidly, producing white colonies and
fertile ascomata; at 45 ° growing rapidly, without
production of ascomata. At 15 ° growing scarcely,
without production of ascomata. No growth below
12 °.The main features of the T. arxii ITS 1-2 and 5.8S
rDNA region sequences are : 529 bp; 117 A; 158 C;
139 G and 115 T. The location of the ITS 1 region is
from nucleotides 7 to 173, the gene 5.8S rRNA from
nucleotides 174 to 348, and the ITS 2 region from
nucleotides 349 to 493.
T. aegyptiaca, T. appendiculata, T. arenaria, T. co-
actilis, T. hyrcaniae and T. subthermophila, also have
ascospores with a subapical to lateral germ pore.
However, the ascospores of T. arxii are bigger than
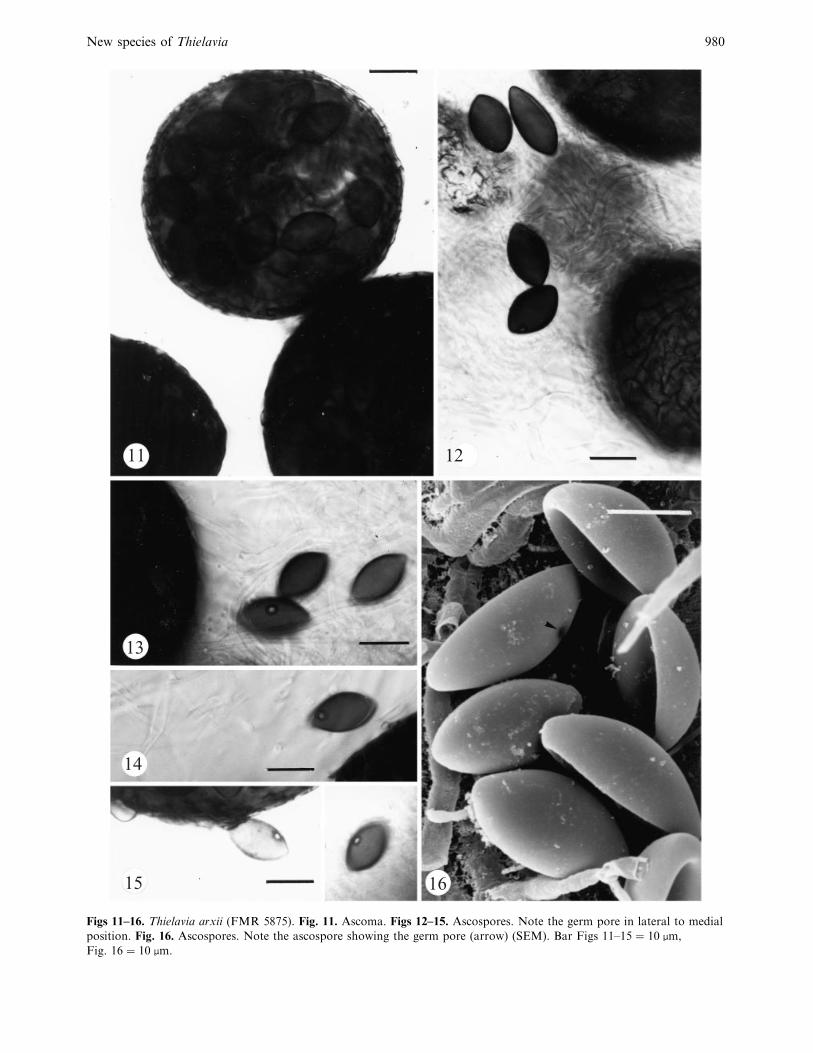
New species of Thielavia 980
11 12
13
14
15 16
Figs 11–16. Thielavia arxii (FMR 5875). Fig. 11. Ascoma. Figs 12–15. Ascospores. Note the germ pore in lateral to medial
position. Fig. 16. Ascospores. Note the ascospore showing the germ pore (arrow) (SEM). Bar Figs 11–15¯ 10 µm,
Fig. 16¯ 10 µm.

A. M. Stchigel and others 981
94
99
T. arxii
T. terricola
T. hyrcaniae
T. minor
T. hyalocarpa
T. fragilis
T. coactilis
T. appendiculata
T. terrestris
T. basicola
T. australiensis
T. tortuosa
Melanocarpus thermophilus
T. minuta
T. microspora
T. subthermophila
T. arenaria
T. intermedia
Gelasinosporabonaerensis
0.01
86
100
94
67
92
57
92
62
Fig. 17. Neighbour-joining tree based on the nucleotide sequences of ITS – 5.8S rRNA regions. Branch lengths are
proportional to genetic distance. The scale bar represents 0±1% sequence divergence. Bootstrap replication frequences
greatest than 50% are indicated above the internodes.
those of T. aegyptiaca (13–14¬8–10 µm), T. arenaria
(9–12±5¬6–7±5 µm), T. coactilis (9–12±5¬(5–)6–7 µm)
and T. hyrcaniae (12–15¬5–6 µm). T. appendiculata
and T. subthermophila have ascospores of similar size to
those of the new species, but T. appendiculata has setose
ascomata and T. subthermophila has ascomata associ-
ated with numerous aleurioconidia which are absent in
T. arxii. Moreover, T. arxii is the only species of the
genus that has a germ pore in a median position in the
ascospores.
Molecular results
The phylogram obtained from the analysis of the ITS-
5.8S rDNA sequences using the neighbour-joining
method (Fig. 17) in general has clades with low
bootstrap values. Some of the terminal clades received
consistent statistical support which demonstrated some
species of the genus are genetically very close. The new
species Thielavia intermedia constituted a basal clade.
DISCUSSION
The main clade is formed by the species studied, apart
from one newly described (Thielavia intermedia), and
was divided into two sister clusters supported by a
bootstrap index of 57%. One grouped T. microspora,
T. subthermophila and T. arenaria, with a bootstrap
value of 86%; these species have ellipsoidal ascospores
and aleurioconidia. The terminal group constituted by
T. subthermophila and T. arenaria received a bootstrap
value of 100%; both species are thermotolerant and
have ascospores with a subapical to lateral germ pore,
and are differentiated only in their sizes. The second

New species of Thielavia 982
cluster was of the remaining species and comprised two
clades that received a low statistical support (48%).
The first clade is composed of T. tortuosa, Melanocarpus
thermophilus and T. minuta, grouped together with a
bootstrap index of 67%. Moreover, M. thermophilus
and T. minuta clustered with a very high bootstrap
value (94%); these species have striking morphological
similarities, short, septate and rugose setae, a peridium
of textura epidermoidea, and 8-spored, broadly ovoid
asci, differing only in temperature requirements and
morphology of the ascospores (Guarro et al. 1996).
Melanocarpus and Thielavia are distinguished by per-
idium type (textura angularis in Melanocarpus and
textura epidermoidea in Thielavia), the morphology of
the ascospores (oblate, thick-walled and opaque in
Melanocarpus and fusiform, ellipsoidal or ovate, thin-
walled and brownish in Thielavia), and in the ana-
morphs when present (Chrysonilia-like arthroconidia in
Melanocarpus, and Chrysosporium-like aleurioconidia
in Thielavia) (von Arx et al. 1988). However, it is
probable that future molecular studies will demonstrate
that these genera are not too genetically distinct. These
species are morphologically distant from T. tortuosa,
which has a peculiar combination of cylindrical, sinuose
asci and large ascospores with a subapical to lateral
germ pore.
The other clade was successively ramified forming
several branches also with a low confidence value.
However, three groups received strong statistical
support. One of them is composed of T. australiensis,
T. basicola and T. terrestris, with a bootstrap value of
92%. T. australiensis and T. terrestris have a thermo-
philic or thermotolerant ecology, and their ascospores
are morphologically similar. However, T. australiensis
has a Chrysosporium-like anamorph, and T. terrestris
produces enteroblastic conidia in chains. T. basicola is
not close morphologically to those, because it has
inequilateral ovate-fusiform ascospores and lacks an
anamorph. Two morphologically related species,
T. appendiculata and T. coactilis, were placed in a clade
supported by a bootstrap value of 62%. They show
ellipsoid to slightly fusiform ascospores with a subapical
germ pore. However, T. appendiculata can be differen-
tiated from T. coactilis by the presence of setose
ascomata, larger ascospores, and the absence of
chlamydospores. Finally, in another clade the new
species T. arxii was placed together with T. terricola,
T. hyrcaniae and T. minor, they formed a well defined
clade, supported by a bootstrap value of 99%. On the
basis of the morphology of T. terricola and T. minor
(ellipsoid ascospores with a terminal germ pore), we
expected them to cluster together, but not with the
other two species of the clade which show clearly
distinct features. T. arxii is easily distinguishable by its
fusiform ascospores with a lateral to median germ pore,
and is thermotolerant, while T. terricola and T. minor
have ellipsoidal ascospores with a terminal germ pore,
and are mesophilic.
This study confirmed that the ITS region is highly
conserved in Sordariales, as indicated previously (Stchi-
gel et al. 1998, 1999), and that it is not very useful to
establish phylogenetical relationships at species level.
We are currently evaluating the usefulness of the LSU
rDNA gene for this purpose.
ACKNOWLEDGEMENTS
We are indebted to Lori M. Carris (Washington State University) and
David H. Griffin (State University of New York) for useful editorial
suggestions on this manuscript, to Luı!s Zaror (Universidad de
Valdivia) for providing the Chilian soil samples, and to the curators
of CBS, IFO and IMI for kindly providing fungal cultures.
REFERENCES
Booth, C. (1961) Studies of pyrenomycetes : VI. Thielavia, with notes
on some allied genera. Mycological Papers 83 : 1–15.
Chen, K.-Y. & Chen, Z.-C. (1996) Thielavia pingtungia sp. nov., a
thermophilic ascomycete from Taiwan. Mycotaxon 60 : 241–247.
Figueras, M. J. & Guarro, J. (1988) A scanning electron microscopic
study of ascoma development in Chaetomium malaysiense. Mycolo-
gia 80 : 298–306.
Gene! , J., Guillamo! n, J. M., Guarro, J., Pujol, I. & Ulfig, K. (1996)
Molecular characterization, relatedness and antifungal suscep-
tibility of the basidiomycetous Hormographiella species and
Coprinus cinereus from clinical and environmental sources. Antonie
van Leewenhoek 70 : 49–57.
Guarro, J., Abdullah, S. K., AI-Bader, S. M., Figueras, M. J. &
Gene! , J. (1996) The genus Melanocarpus. Mycolological Research
100 : 75–78.
Guillamo! n, J. M., Cano, J., Ramo! n, D. &Guarro, J. (1996) Molecular
differentiation of Keratinomyces (Trichophyton) species. Antonie
van Leewenhoek 69 : 223–227.
Kornerup, A. & Wanscher, J. H. (1984) Methuen Handbook of
Colour. 3rd. edn. Eyre Methuen, London.
Kumar, S., Tamura, K. & Nei, M. (1993) MEGA: molecular
evolutionary genetics analysis. Version 1.0. Pennsylvania State
University, Philadelphia.
Ito, T., Okane, I. & Nakagiri, A. (1998) Thielavia aurantiaca, a new
species from Japanese soil. Mycoscience 39 : 93–96.
Malloch, D. & Cain, R. F. (1973) The genus Thielavia. Mycologia 65 :
1055–1077.
Mouchacca, J. (1973) Les Thielavia des sols arides : espe' cies nouvelles
et analyse ge!ne! rique. BulletıUn de la SocieU teU Mycologique de France
89 : 295–311.
Saitou, N. & Nei, M. (1987) The neighbour-joining method: a new
method for reconstructing phylogenetic trees. Molecular Biology
and Evolution 4 : 406–425.
Stchigel, A. M., Cano, J. & Guarro, J. (1998) A new species of
Gelasinospora from Argentina. Mycological Research 102 : 1405–
1408.
Stchigel, A. M., Sague! s, M., Cano, J. & Guarro, J. (1999) Three new
thermotolerant species of Corynascus (Sordariales, Chaetomiaceae)
from soil, with a key of the known species. Mycological Research
104 : 879–887.
Thompson, J. D., Higgins, D. G. & Gibson, T. J. (1994) CLUSTAL
W: Improving the sensitivity of progressive multiple sequence
alignment through sequence weighting, positions-specific gap
penalties and weight matrix choice. Nucleic Acids Research 22 :
4673–4680.
von Arx, J. A. (1973) Ostiolate and non-ostiolate pyrenomycetes.
Proceeding of the Koninklijke Nederlandse Akademie van Weten-
schappen 76 : 289–296.
von Arx, J. A. (1975) On Thielavia and some similar genera of
ascomycetes. Studies in Mycology 8 : 1–29.
von Arx, J. A., Figueras, M. J. & Guarro, J. (1988) Sordariaceous
ascomycetes without ascospore ejaculation. Nova Hedwigia 94 :
1–104.

A. M. Stchigel and others 983
Warcup, J. H. & Baker, K. F. (1963) Ocurrence of dormant
ascospores in soil. Nature 197 : 1317–1318.
White, T. J., Bruns, T., Lee, S. & Taylor, J. (1990) Amplification and
direct sequencing of fungi ribosomal RNA genes for phylogenetics.
In PCR Protocols : a guide to methods and applications (M. A.
Innis, D. H. Gelfand, J. J. Sninsky & T. J. White, eds) : 315–322.
Academic Press, San Diego.
Corresponding Editor: D. L. Hawksworth
![Tigers By :Tabby Griffith Organism Family, Genus, Species Organism Family: Felidae Genus: Panthera Species: Tigers (Sumatran Tiger, Amur [or Siberian]](https://img.pdfslide.net/doc/110x75/56649ef25503460f94c04af6/tigers-by-tabby-griffith-organism-family-genus-species-organism-family.jpg)




























