Embed Size (px)
Citation preview

1877Research Article
IntroductionThe olfactory system is the only region of the mammaliancentral nervous system (CNS) in which the olfactory receptorneurons are unique in retaining their ability to regeneratethroughout life, both in response to injury and as part of normalturnover (Graziadei and Monti Graziadei, 1980; Doucette etal., 1983). Olfactory ensheathing cells (OECs) are the glialcells that derive from the olfactory placode and envelopolfactory axons in the course of migration from the olfactoryepithelium to the bulb (Ramon-Cueto and Avila, 1998). Thecells are different from the typical glia in terms of existing inboth the peripheral nervous system and CNS and sharing thephenotypes of both astrocytes and Schwann cells (Ramon-Cueto and Avila, 1998; Gudino-Cabrera and Nieto-Sampedro,2000). OECs have been reported to pioneer the olfactory nervepathway and provide a conducive substrate for growingprimary olfactory axons, although the specific mechanismsremain undescribed (Tennent and Chuah, 1996).
The repair of CNS damage continues to be a majorchallenge, in particular that of spinal cord injury. Owing to theaxonal growth-promoting properties, OEC transplantation hasemerged as a very promising experimental therapy to treataxonal injuries (Franklin and Barnett, 2000; Raisman, 2001;Ramon-Cueto and Santos-Benito, 2001). Transplanted OECshave been shown to migrate with regenerating axons throughan unfavorable CNS environment (Li et al., 2004), and tomingle well with astrocytes in adult brain (Li et al., 1998;Lakatos et al., 2000; Richter et al., 2005). Therefore, themigrating ability of OECs in the CNS was thought to be
essential for neural regeneration and re-ensheathment afterspinal cord injury. However, whether transplanted OECs canmigrate over long distances in the CNS is still controversial(Smale et al., 1996; Gudino-Cabrera et al., 2000; Takami et al.,2002; Collazos-Castro et al., 2005; Deng et al., 2006).
The regeneration of CNS axons following injury isdrastically restricted by the presence of inhibitory moleculeswithin myelin (Schwab and Bartholdi, 1996; Ng et al., 1996).Three inhibitors that have been identified are Nogo, myelin-associated glycoprotein (MAG) and oligodendrocyte myelinglycoprotein (OMgp) (He and Koprivica, 2004). Nogo is amember of the reticulon family and occurs in three forms,Nogo-A, Nogo-B and Nogo-C, which are generated fromalternate splicing (GrandPre et al., 2000). All three isoforms ofNogo share a 66-amino-acid-residue luminal/extracellulardomain (Nogo-66), which inhibits axonal extension andfibroblast spreading (Brittis and Flanagan, 2001; Fournieret al., 2001). The molecular cloning of Nogo (Chen et al.,2000; GrandPre et al., 2000; Prinjha et al., 2000) led tothe identification of a neuronal surface glycosylphosphatidylinositol (GPI)-linked receptor that binds with highaffinity to Nogo-66, termed the Nogo-66 receptor (NgR)(Fournier et al., 2001). The low affinity neurotrophin receptor,p75, has been identified as a coreceptor for NgR, andtransduces a signal upon interaction with myelin ligands (Wanget al., 2002; Wong et al., 2002). The downstream signalingpathway involves the activation of small GTPases of the Rhofamily, which in turn regulate cytoskeletal protein assemblyand mediate inhibitory effects on neurite growth (Yamashita et
The migration of olfactory ensheathing cells (OECs) isessential for pioneering the olfactory nerve pathwayduring development and for promoting axonalregeneration when implanted into the injured centralnervous system (CNS). In the present study, recombinantNogo-66 enhanced the adhesion of OECs and inhibitedtheir migration. Using immunocytochemistry and westernblot, we showed that the Nogo-66 receptor (NgR) wasexpressed on OECs. When NgR was released from the cellsurface with phosphatidylinositol-specific phospholipase Cor neutralized by NgR antibody, the effect of Nogo-66 onOEC adhesion and migration was markedly attenuated.Nogo-66 was found to promote the formation of focaladhesion in OECs and inhibited their membrane
protrusion through the activation of RhoA. Furthermore,the co-culture migration assay demonstrated that OECmotility was significantly restricted by Nogo-A expressedon Cos7 cell membranes or oligodendrocytes. Moreover,treatment with anti-NgR antibody facilitated migration ofimplanted OECs in a spinal cord hemisection injury model.Taken together, we demonstrate, for the first time, thatNogo, a myelin-associated inhibitor of axon regeneration inthe CNS, enhances the adhesion and inhibits the migrationof OECs via NgR regulation of RhoA.
Key words: Nogo, Olfactory ensheathing cells, Migration, Adhesion,RhoA, Spinal cord injury
Summary
Nogo enhances the adhesion of olfactory ensheathingcells and inhibits their migrationZhida Su*, Li Cao*, Yanling Zhu, Xiujie Liu, Zhihui Huang, Aijun Huang and Cheng He‡
Department of Neurobiology, Second Military Medical University, Shanghai 200433, China*These authors contributed equally to this work‡Author for correspondence (e-mail: [email protected])
Accepted 28 March 2007Journal of Cell Science 120, 1877-1887 Published by The Company of Biologists 2007doi:10.1242/jcs.03448
Jour
nal o
f Cel
l Sci
ence
JCS ePress online publication date 8 May 2007
1878
al., 2002; Yiu and He, 2003). The mRNA for NgR has beenfound in abundance in OECs (Woodhall et al., 2003), and theNgR coreceptor p75 is well known as a marker protein forOECs. However, little is known about the function of thereceptors in OECs.
Although the neuronal growth inhibition activity of Nogohas been well documented, other important features of Nogoare only beginning to be understood. Nogo-A has beenreported to be an important determinant of the developmentof experimental autoimmune encephalomyelitis (Karnezis etal., 2004), as well as an essential player in modulating axon-glial junction architecture and possibly K+-channellocalization during development (Nie et al., 2003). Nogo-B,like other members of the reticulon family, is involved inmodulating BACE1 activity and amyloid-� peptide generation(He et al., 2004), and might function as a pro-apoptotic protein
(Tagami et al., 2000). Moreover, Nogo-B has beenreported as a regulator of vascular remodeling(Acevedo et al., 2004). In the present study, wedemonstrate that Nogo enhances the adhesion andinhibits the migration of OECs via RhoAactivation by NgR.
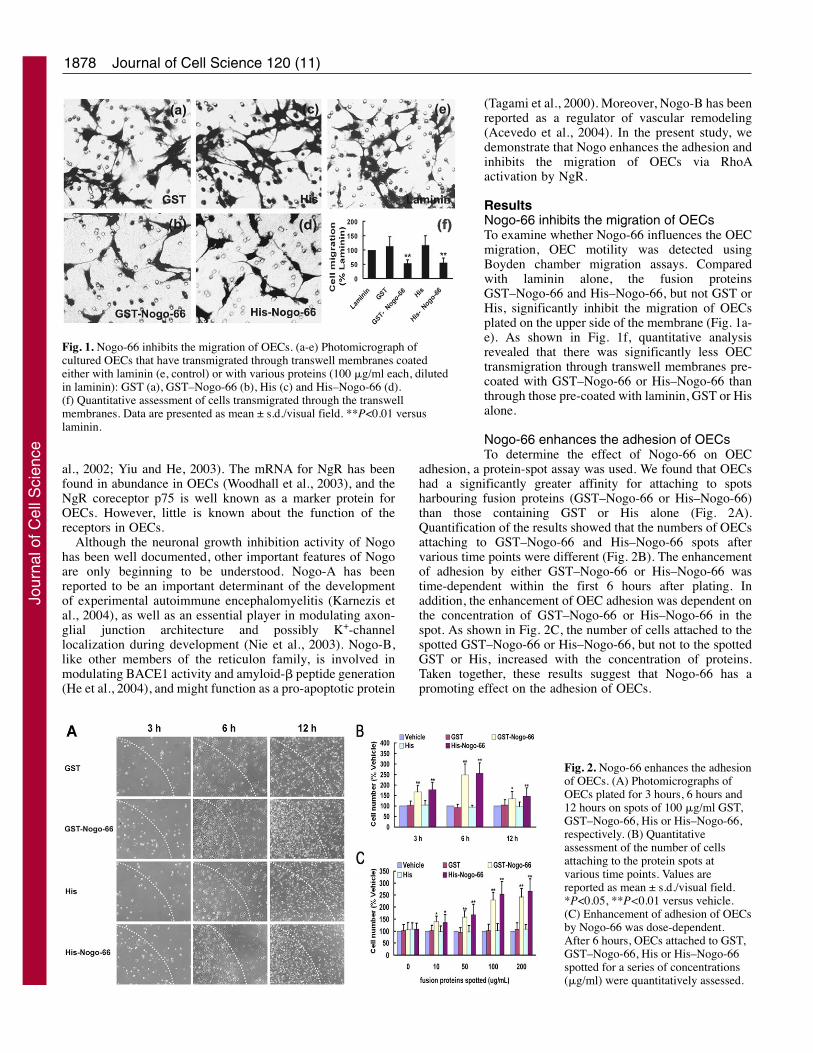
ResultsNogo-66 inhibits the migration of OECsTo examine whether Nogo-66 influences the OECmigration, OEC motility was detected usingBoyden chamber migration assays. Comparedwith laminin alone, the fusion proteinsGST–Nogo-66 and His–Nogo-66, but not GST orHis, significantly inhibit the migration of OECsplated on the upper side of the membrane (Fig. 1a-e). As shown in Fig. 1f, quantitative analysisrevealed that there was significantly less OECtransmigration through transwell membranes pre-coated with GST–Nogo-66 or His–Nogo-66 thanthrough those pre-coated with laminin, GST or Hisalone.
Nogo-66 enhances the adhesion of OECsTo determine the effect of Nogo-66 on OEC
adhesion, a protein-spot assay was used. We found that OECshad a significantly greater affinity for attaching to spotsharbouring fusion proteins (GST–Nogo-66 or His–Nogo-66)than those containing GST or His alone (Fig. 2A).Quantification of the results showed that the numbers of OECsattaching to GST–Nogo-66 and His–Nogo-66 spots aftervarious time points were different (Fig. 2B). The enhancementof adhesion by either GST–Nogo-66 or His–Nogo-66 wastime-dependent within the first 6 hours after plating. Inaddition, the enhancement of OEC adhesion was dependent onthe concentration of GST–Nogo-66 or His–Nogo-66 in thespot. As shown in Fig. 2C, the number of cells attached to thespotted GST–Nogo-66 or His–Nogo-66, but not to the spottedGST or His, increased with the concentration of proteins.Taken together, these results suggest that Nogo-66 has apromoting effect on the adhesion of OECs.
Journal of Cell Science 120 (11)
Fig. 1. Nogo-66 inhibits the migration of OECs. (a-e) Photomicrograph ofcultured OECs that have transmigrated through transwell membranes coatedeither with laminin (e, control) or with various proteins (100 �g/ml each, dilutedin laminin): GST (a), GST–Nogo-66 (b), His (c) and His–Nogo-66 (d).(f) Quantitative assessment of cells transmigrated through the transwellmembranes. Data are presented as mean ± s.d./visual field. **P<0.01 versuslaminin.
Fig. 2. Nogo-66 enhances the adhesionof OECs. (A) Photomicrographs ofOECs plated for 3 hours, 6 hours and12 hours on spots of 100 �g/ml GST,GST–Nogo-66, His or His–Nogo-66,respectively. (B) Quantitativeassessment of the number of cellsattaching to the protein spots atvarious time points. Values arereported as mean ± s.d./visual field.*P<0.05, **P<0.01 versus vehicle.(C) Enhancement of adhesion of OECsby Nogo-66 was dose-dependent.After 6 hours, OECs attached to GST,GST–Nogo-66, His or His–Nogo-66spotted for a series of concentrations(�g/ml) were quantitatively assessed.
Jour
nal o
f Cel
l Sci
ence
1879Nogo regulates OEC migration
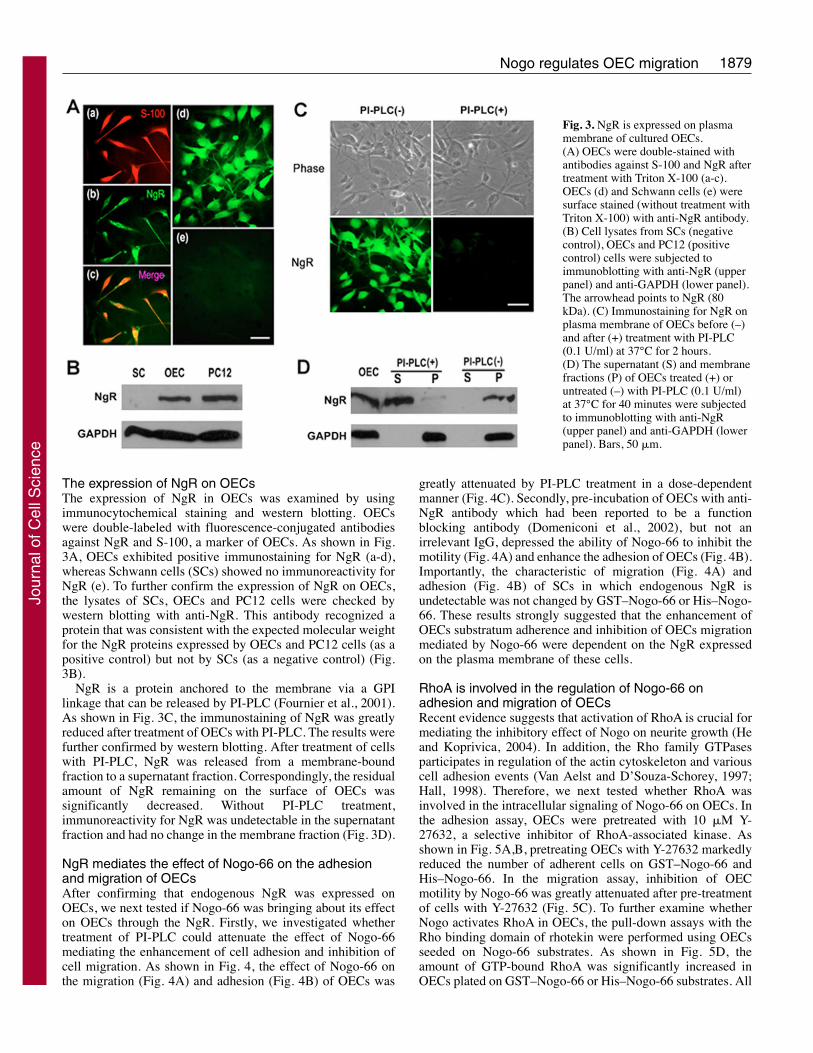
The expression of NgR on OECsThe expression of NgR in OECs was examined by usingimmunocytochemical staining and western blotting. OECswere double-labeled with fluorescence-conjugated antibodiesagainst NgR and S-100, a marker of OECs. As shown in Fig.3A, OECs exhibited positive immunostaining for NgR (a-d),whereas Schwann cells (SCs) showed no immunoreactivity forNgR (e). To further confirm the expression of NgR on OECs,the lysates of SCs, OECs and PC12 cells were checked bywestern blotting with anti-NgR. This antibody recognized aprotein that was consistent with the expected molecular weightfor the NgR proteins expressed by OECs and PC12 cells (as apositive control) but not by SCs (as a negative control) (Fig.3B).
NgR is a protein anchored to the membrane via a GPIlinkage that can be released by PI-PLC (Fournier et al., 2001).As shown in Fig. 3C, the immunostaining of NgR was greatlyreduced after treatment of OECs with PI-PLC. The results werefurther confirmed by western blotting. After treatment of cellswith PI-PLC, NgR was released from a membrane-boundfraction to a supernatant fraction. Correspondingly, the residualamount of NgR remaining on the surface of OECs wassignificantly decreased. Without PI-PLC treatment,immunoreactivity for NgR was undetectable in the supernatantfraction and had no change in the membrane fraction (Fig. 3D).
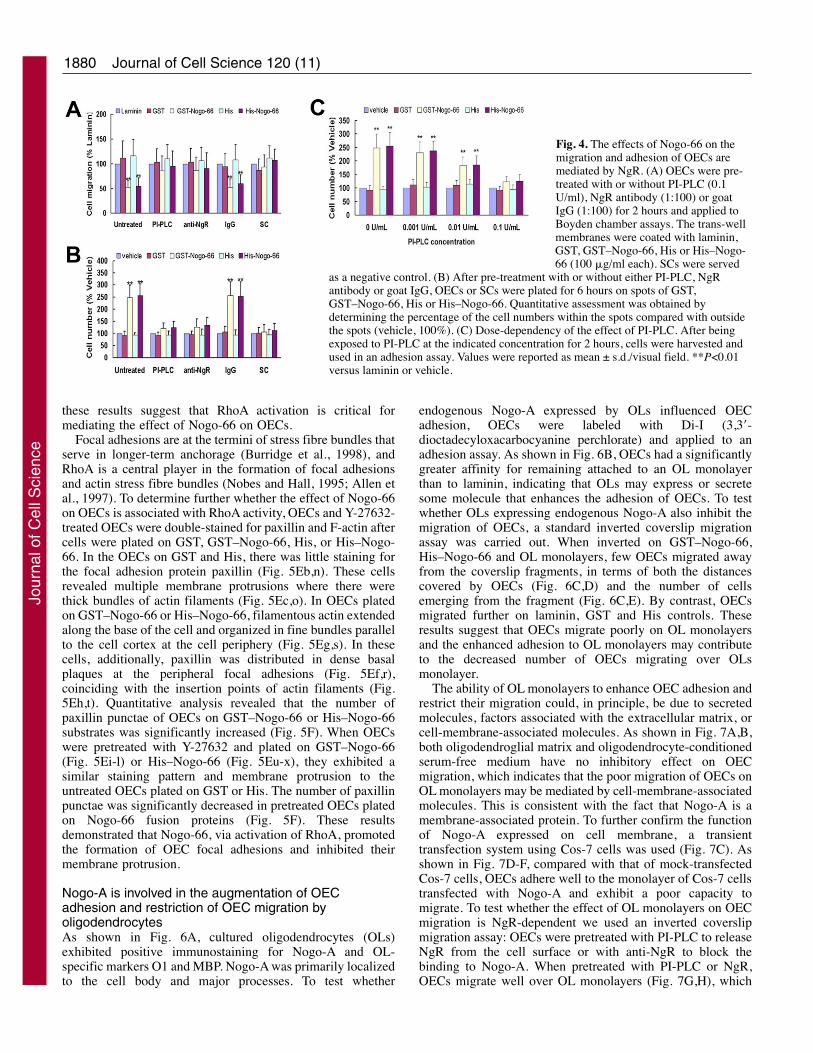
NgR mediates the effect of Nogo-66 on the adhesionand migration of OECsAfter confirming that endogenous NgR was expressed onOECs, we next tested if Nogo-66 was bringing about its effecton OECs through the NgR. Firstly, we investigated whethertreatment of PI-PLC could attenuate the effect of Nogo-66mediating the enhancement of cell adhesion and inhibition ofcell migration. As shown in Fig. 4, the effect of Nogo-66 onthe migration (Fig. 4A) and adhesion (Fig. 4B) of OECs was
greatly attenuated by PI-PLC treatment in a dose-dependentmanner (Fig. 4C). Secondly, pre-incubation of OECs with anti-NgR antibody which had been reported to be a functionblocking antibody (Domeniconi et al., 2002), but not anirrelevant IgG, depressed the ability of Nogo-66 to inhibit themotility (Fig. 4A) and enhance the adhesion of OECs (Fig. 4B).Importantly, the characteristic of migration (Fig. 4A) andadhesion (Fig. 4B) of SCs in which endogenous NgR isundetectable was not changed by GST–Nogo-66 or His–Nogo-66. These results strongly suggested that the enhancement ofOECs substratum adherence and inhibition of OECs migrationmediated by Nogo-66 were dependent on the NgR expressedon the plasma membrane of these cells.
RhoA is involved in the regulation of Nogo-66 onadhesion and migration of OECsRecent evidence suggests that activation of RhoA is crucial formediating the inhibitory effect of Nogo on neurite growth (Heand Koprivica, 2004). In addition, the Rho family GTPasesparticipates in regulation of the actin cytoskeleton and variouscell adhesion events (Van Aelst and D’Souza-Schorey, 1997;Hall, 1998). Therefore, we next tested whether RhoA wasinvolved in the intracellular signaling of Nogo-66 on OECs. Inthe adhesion assay, OECs were pretreated with 10 �M Y-27632, a selective inhibitor of RhoA-associated kinase. Asshown in Fig. 5A,B, pretreating OECs with Y-27632 markedlyreduced the number of adherent cells on GST–Nogo-66 andHis–Nogo-66. In the migration assay, inhibition of OECmotility by Nogo-66 was greatly attenuated after pre-treatmentof cells with Y-27632 (Fig. 5C). To further examine whetherNogo activates RhoA in OECs, the pull-down assays with theRho binding domain of rhotekin were performed using OECsseeded on Nogo-66 substrates. As shown in Fig. 5D, theamount of GTP-bound RhoA was significantly increased inOECs plated on GST–Nogo-66 or His–Nogo-66 substrates. All
Fig. 3. NgR is expressed on plasmamembrane of cultured OECs.(A) OECs were double-stained withantibodies against S-100 and NgR aftertreatment with Triton X-100 (a-c).OECs (d) and Schwann cells (e) weresurface stained (without treatment withTriton X-100) with anti-NgR antibody.(B) Cell lysates from SCs (negativecontrol), OECs and PC12 (positivecontrol) cells were subjected toimmunoblotting with anti-NgR (upperpanel) and anti-GAPDH (lower panel).The arrowhead points to NgR (80kDa). (C) Immunostaining for NgR onplasma membrane of OECs before (–)and after (+) treatment with PI-PLC(0.1 U/ml) at 37°C for 2 hours.(D) The supernatant (S) and membranefractions (P) of OECs treated (+) oruntreated (–) with PI-PLC (0.1 U/ml)at 37°C for 40 minutes were subjectedto immunoblotting with anti-NgR(upper panel) and anti-GAPDH (lowerpanel). Bars, 50 �m.
Jour
nal o
f Cel
l Sci
ence
1880
these results suggest that RhoA activation is critical formediating the effect of Nogo-66 on OECs.
Focal adhesions are at the termini of stress fibre bundles thatserve in longer-term anchorage (Burridge et al., 1998), andRhoA is a central player in the formation of focal adhesionsand actin stress fibre bundles (Nobes and Hall, 1995; Allen etal., 1997). To determine further whether the effect of Nogo-66on OECs is associated with RhoA activity, OECs and Y-27632-treated OECs were double-stained for paxillin and F-actin aftercells were plated on GST, GST–Nogo-66, His, or His–Nogo-66. In the OECs on GST and His, there was little staining forthe focal adhesion protein paxillin (Fig. 5Eb,n). These cellsrevealed multiple membrane protrusions where there werethick bundles of actin filaments (Fig. 5Ec,o). In OECs platedon GST–Nogo-66 or His–Nogo-66, filamentous actin extendedalong the base of the cell and organized in fine bundles parallelto the cell cortex at the cell periphery (Fig. 5Eg,s). In thesecells, additionally, paxillin was distributed in dense basalplaques at the peripheral focal adhesions (Fig. 5Ef,r),coinciding with the insertion points of actin filaments (Fig.5Eh,t). Quantitative analysis revealed that the number ofpaxillin punctae of OECs on GST–Nogo-66 or His–Nogo-66substrates was significantly increased (Fig. 5F). When OECswere pretreated with Y-27632 and plated on GST–Nogo-66(Fig. 5Ei-l) or His–Nogo-66 (Fig. 5Eu-x), they exhibited asimilar staining pattern and membrane protrusion to theuntreated OECs plated on GST or His. The number of paxillinpunctae was significantly decreased in pretreated OECs platedon Nogo-66 fusion proteins (Fig. 5F). These resultsdemonstrated that Nogo-66, via activation of RhoA, promotedthe formation of OEC focal adhesions and inhibited theirmembrane protrusion.
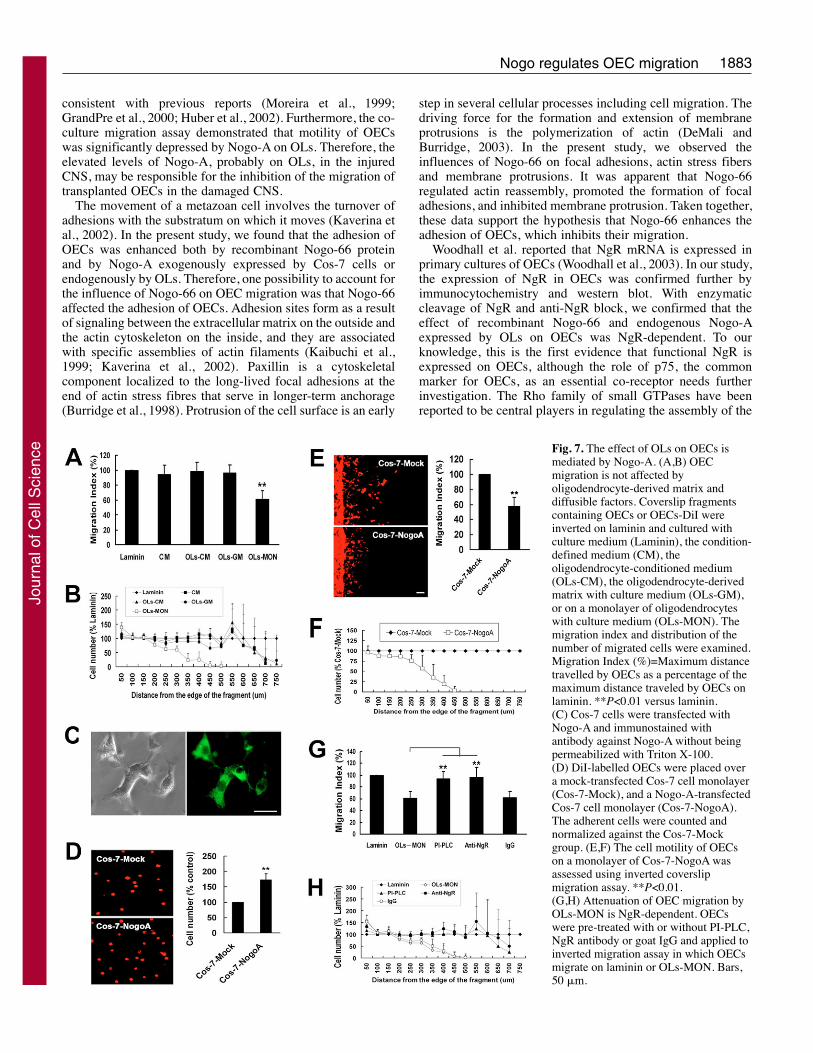
Nogo-A is involved in the augmentation of OECadhesion and restriction of OEC migration byoligodendrocytesAs shown in Fig. 6A, cultured oligodendrocytes (OLs)exhibited positive immunostaining for Nogo-A and OL-specific markers O1 and MBP. Nogo-A was primarily localizedto the cell body and major processes. To test whether
endogenous Nogo-A expressed by OLs influenced OECadhesion, OECs were labeled with Di-I (3,3�-dioctadecyloxacarbocyanine perchlorate) and applied to anadhesion assay. As shown in Fig. 6B, OECs had a significantlygreater affinity for remaining attached to an OL monolayerthan to laminin, indicating that OLs may express or secretesome molecule that enhances the adhesion of OECs. To testwhether OLs expressing endogenous Nogo-A also inhibit themigration of OECs, a standard inverted coverslip migrationassay was carried out. When inverted on GST–Nogo-66,His–Nogo-66 and OL monolayers, few OECs migrated awayfrom the coverslip fragments, in terms of both the distancescovered by OECs (Fig. 6C,D) and the number of cellsemerging from the fragment (Fig. 6C,E). By contrast, OECsmigrated further on laminin, GST and His controls. Theseresults suggest that OECs migrate poorly on OL monolayersand the enhanced adhesion to OL monolayers may contributeto the decreased number of OECs migrating over OLsmonolayer.
The ability of OL monolayers to enhance OEC adhesion andrestrict their migration could, in principle, be due to secretedmolecules, factors associated with the extracellular matrix, orcell-membrane-associated molecules. As shown in Fig. 7A,B,both oligodendroglial matrix and oligodendrocyte-conditionedserum-free medium have no inhibitory effect on OECmigration, which indicates that the poor migration of OECs onOL monolayers may be mediated by cell-membrane-associatedmolecules. This is consistent with the fact that Nogo-A is amembrane-associated protein. To further confirm the functionof Nogo-A expressed on cell membrane, a transienttransfection system using Cos-7 cells was used (Fig. 7C). Asshown in Fig. 7D-F, compared with that of mock-transfectedCos-7 cells, OECs adhere well to the monolayer of Cos-7 cellstransfected with Nogo-A and exhibit a poor capacity tomigrate. To test whether the effect of OL monolayers on OECmigration is NgR-dependent we used an inverted coverslipmigration assay: OECs were pretreated with PI-PLC to releaseNgR from the cell surface or with anti-NgR to block thebinding to Nogo-A. When pretreated with PI-PLC or NgR,OECs migrate well over OL monolayers (Fig. 7G,H), which
Journal of Cell Science 120 (11)
Fig. 4. The effects of Nogo-66 on themigration and adhesion of OECs aremediated by NgR. (A) OECs were pre-treated with or without PI-PLC (0.1U/ml), NgR antibody (1:100) or goatIgG (1:100) for 2 hours and applied toBoyden chamber assays. The trans-wellmembranes were coated with laminin,GST, GST–Nogo-66, His or His–Nogo-66 (100 �g/ml each). SCs were served
as a negative control. (B) After pre-treatment with or without either PI-PLC, NgRantibody or goat IgG, OECs or SCs were plated for 6 hours on spots of GST,GST–Nogo-66, His or His–Nogo-66. Quantitative assessment was obtained bydetermining the percentage of the cell numbers within the spots compared with outsidethe spots (vehicle, 100%). (C) Dose-dependency of the effect of PI-PLC. After beingexposed to PI-PLC at the indicated concentration for 2 hours, cells were harvested andused in an adhesion assay. Values were reported as mean ± s.d./visual field. **P<0.01versus laminin or vehicle.
Jour
nal o
f Cel
l Sci
ence

1881Nogo regulates OEC migration
indicates that the inhibitory effect of OL monolayers on themigration of OECs is NgR-dependent.
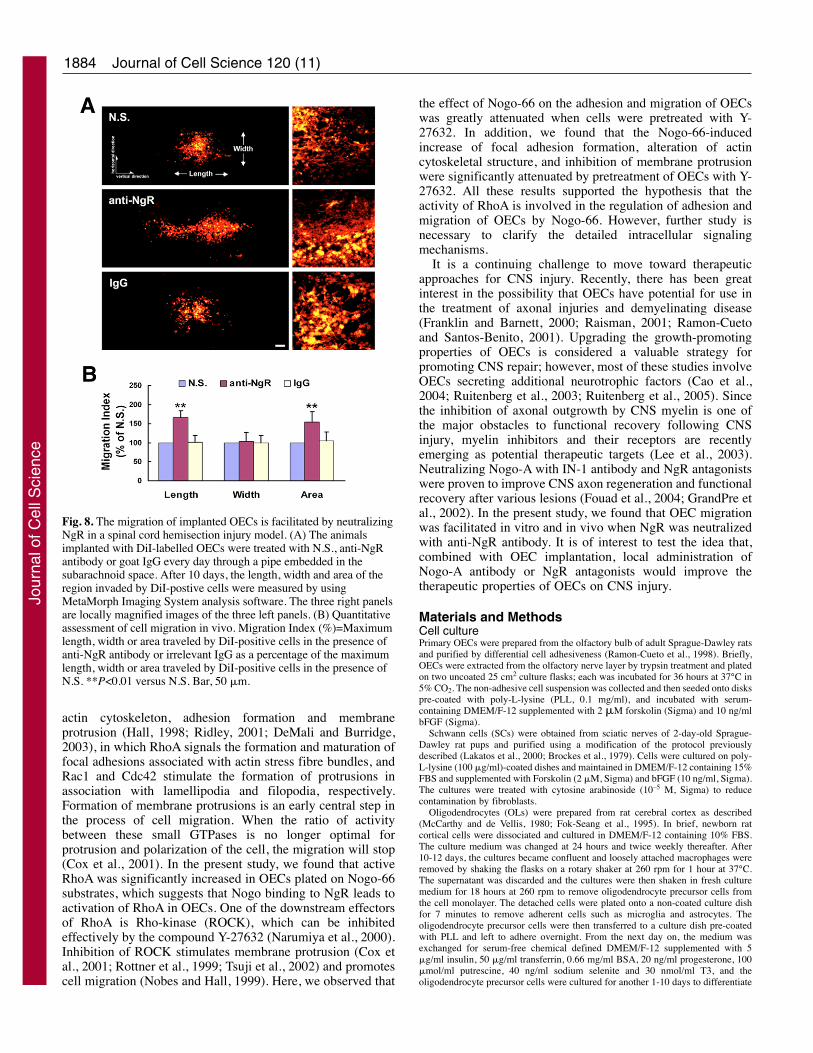
The migration of implanted OECs is facilitated byneutralizing NgR in vivoTo investigate whether the migration of implanted OECs wasaffected by Nogo, an in vivo migration assay was performedas previously reported (Cao et al., 2006). OECs prelabeled withDi-I were injected into the injured spinal cord. Ten days afterinjection, OECs were found to migrate longitudinally andlaterally from the injection sites. However, in the presence ofanti-NgR antibody, OECs migrated further in the verticaldirection (parallel with the long axis of spinal cord) whencompared with OEC migration using normal saline (N.S.) or
the irrelevant IgG, at the rostral (Fig. 8) as well as the caudalinjecton site. There was no difference in migration distancebetween OECs injected at rostral and caudal sites (data notshown). Quantitative analysis revealed that treatment of OECswith anti-NgR increased the maximum migration distancealong the vertical direction, and there was no significantdifference in the maximum migration distance along thehorizontal direction (Fig. 8B). These results indicated thatneutralizing NgR with anti-NgR antibody facilitated OECmigration through white matter tracts in vivo.
DiscussionCell migration is an essential process in embryonicdevelopment, growth, wound repair and inflammation, as well
Fig. 5. RhoA activation is critical formediating the effect of Nogo-66 onOECs. (A,B) Y-27632 treatmentsignificantly attenuates theenhancement of Nogo-66 on adhesionof OECs. OECs (a,c,e,g,i) and Y-27632-treated OECs (b,d,f,h,j) were plated ondishes coated with laminin (a,b), GST(c,d), GST–Nogo-66 (e,f), His (g,h) orHis–Nogo-66 (i,j). After incubation for30 minutes with gentle shaking,adherent cells were photographed andcounted. The values were normalizedwith respect to the values obtained forlaminin. (C) Y-27632 treatmentsignificantly attenuates the inhibition ofNogo-66 on migration of OECs. OECswere pre-treated with or without Y-27632 and applied to Boyden chamberassays. The trans-well membranes werecoated with laminin, GST, GST–Nogo-66, His or His–Nogo-66 (100 �g/mleach). Values were reported as mean ±s.d./visual field. *P<0.05, **P<0.01versus laminin or vehicle. (D) RhoAactivation in OECs was assessed withpull-down assays with the Rho-bindingdomain of rhotekin. The amount ofGTP-bound RhoA was normalized tothe amount of total RhoA. Results aremeans ± s.d. from three independentexperiments. *P<0.05 versus GST orHis. (E) Nogo-66 promotes theformation of focal adhesions andinhibits membrane protrusion of OECsthrough RhoA activation. OECs and Y-27632-treated OECs were plated for 60minutes on coverslips coated with GST(a-d), GST–Nogo-66 (e-l), His (m-p) orHis–Nogo-66 (q-x). Cultures were thenfixed, permeabilized, and stained withanti-paxillin to visualize focal adhesions(green) and rhodamine-conjugatedphalloidin to visualize filamentous actin(red). Arrows (in c,k,o,w) indicate thethick bundles of actin in the membraneprotrusions of OECs. Arrowheads (inh,t) indicate the stress fibers terminating at the adhesion plaques. Bar, 10 �m. (F) Quantification of the paxillin punctae. The number ofpunctae/cell were counted for 40 cells selected randomly in each group. The number of punctae was normalized by comparing with GST orHis, and the percentage of punctae in control and treated cells was compared. **P<0.01.
Jour
nal o
f Cel
l Sci
ence
1882
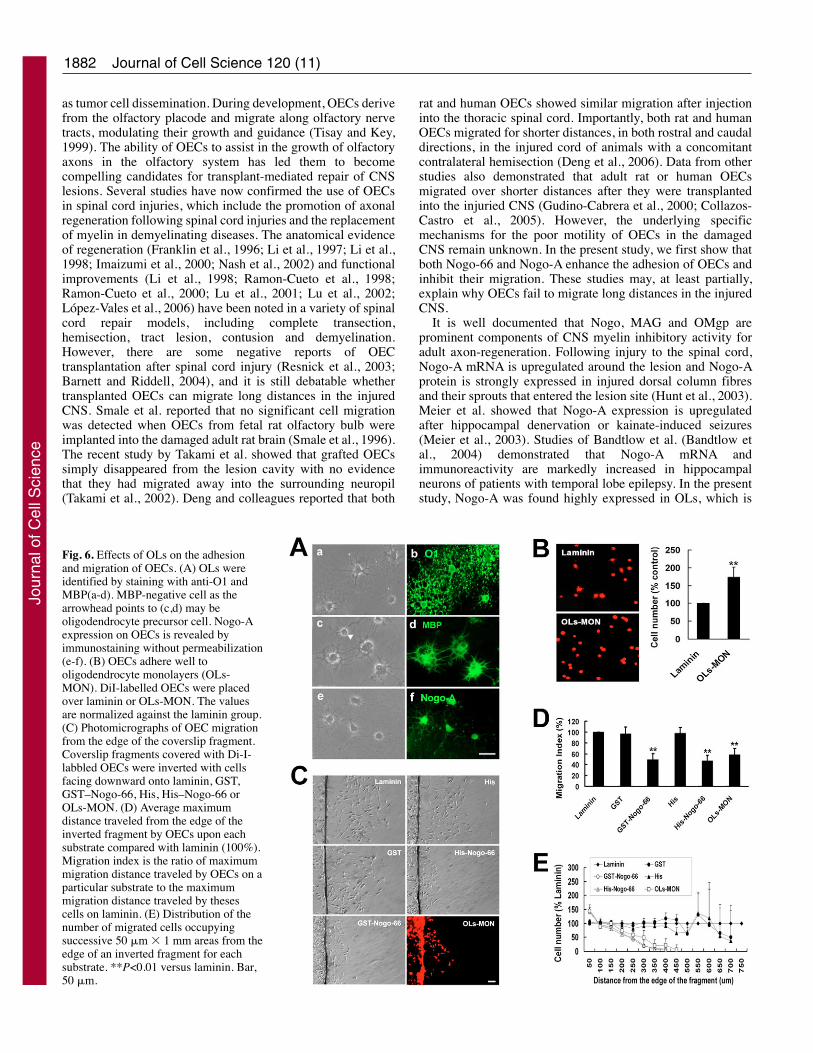
as tumor cell dissemination. During development, OECs derivefrom the olfactory placode and migrate along olfactory nervetracts, modulating their growth and guidance (Tisay and Key,1999). The ability of OECs to assist in the growth of olfactoryaxons in the olfactory system has led them to becomecompelling candidates for transplant-mediated repair of CNSlesions. Several studies have now confirmed the use of OECsin spinal cord injuries, which include the promotion of axonalregeneration following spinal cord injuries and the replacementof myelin in demyelinating diseases. The anatomical evidenceof regeneration (Franklin et al., 1996; Li et al., 1997; Li et al.,1998; Imaizumi et al., 2000; Nash et al., 2002) and functionalimprovements (Li et al., 1998; Ramon-Cueto et al., 1998;Ramon-Cueto et al., 2000; Lu et al., 2001; Lu et al., 2002;López-Vales et al., 2006) have been noted in a variety of spinalcord repair models, including complete transection,hemisection, tract lesion, contusion and demyelination.However, there are some negative reports of OECtransplantation after spinal cord injury (Resnick et al., 2003;Barnett and Riddell, 2004), and it is still debatable whethertransplanted OECs can migrate long distances in the injuredCNS. Smale et al. reported that no significant cell migrationwas detected when OECs from fetal rat olfactory bulb wereimplanted into the damaged adult rat brain (Smale et al., 1996).The recent study by Takami et al. showed that grafted OECssimply disappeared from the lesion cavity with no evidencethat they had migrated away into the surrounding neuropil(Takami et al., 2002). Deng and colleagues reported that both
rat and human OECs showed similar migration after injectioninto the thoracic spinal cord. Importantly, both rat and humanOECs migrated for shorter distances, in both rostral and caudaldirections, in the injured cord of animals with a concomitantcontralateral hemisection (Deng et al., 2006). Data from otherstudies also demonstrated that adult rat or human OECsmigrated over shorter distances after they were transplantedinto the injuried CNS (Gudino-Cabrera et al., 2000; Collazos-Castro et al., 2005). However, the underlying specificmechanisms for the poor motility of OECs in the damagedCNS remain unknown. In the present study, we first show thatboth Nogo-66 and Nogo-A enhance the adhesion of OECs andinhibit their migration. These studies may, at least partially,explain why OECs fail to migrate long distances in the injuredCNS.
It is well documented that Nogo, MAG and OMgp areprominent components of CNS myelin inhibitory activity foradult axon-regeneration. Following injury to the spinal cord,Nogo-A mRNA is upregulated around the lesion and Nogo-Aprotein is strongly expressed in injured dorsal column fibresand their sprouts that entered the lesion site (Hunt et al., 2003).Meier et al. showed that Nogo-A expression is upregulatedafter hippocampal denervation or kainate-induced seizures(Meier et al., 2003). Studies of Bandtlow et al. (Bandtlow etal., 2004) demonstrated that Nogo-A mRNA andimmunoreactivity are markedly increased in hippocampalneurons of patients with temporal lobe epilepsy. In the presentstudy, Nogo-A was found highly expressed in OLs, which is
Journal of Cell Science 120 (11)
Fig. 6. Effects of OLs on the adhesionand migration of OECs. (A) OLs wereidentified by staining with anti-O1 andMBP(a-d). MBP-negative cell as thearrowhead points to (c,d) may beoligodendrocyte precursor cell. Nogo-Aexpression on OECs is revealed byimmunostaining without permeabilization(e-f). (B) OECs adhere well tooligodendrocyte monolayers (OLs-MON). DiI-labelled OECs were placedover laminin or OLs-MON. The valuesare normalized against the laminin group.(C) Photomicrographs of OEC migrationfrom the edge of the coverslip fragment.Coverslip fragments covered with Di-I-labbled OECs were inverted with cellsfacing downward onto laminin, GST,GST–Nogo-66, His, His–Nogo-66 orOLs-MON. (D) Average maximumdistance traveled from the edge of theinverted fragment by OECs upon eachsubstrate compared with laminin (100%).Migration index is the ratio of maximummigration distance traveled by OECs on aparticular substrate to the maximummigration distance traveled by thesescells on laminin. (E) Distribution of thenumber of migrated cells occupyingsuccessive 50 �m � 1 mm areas from theedge of an inverted fragment for eachsubstrate. **P<0.01 versus laminin. Bar,50 �m.
Jour
nal o
f Cel
l Sci
ence
1883Nogo regulates OEC migration
consistent with previous reports (Moreira et al., 1999;GrandPre et al., 2000; Huber et al., 2002). Furthermore, the co-culture migration assay demonstrated that motility of OECswas significantly depressed by Nogo-A on OLs. Therefore, theelevated levels of Nogo-A, probably on OLs, in the injuredCNS, may be responsible for the inhibition of the migration oftransplanted OECs in the damaged CNS.
The movement of a metazoan cell involves the turnover ofadhesions with the substratum on which it moves (Kaverina etal., 2002). In the present study, we found that the adhesion ofOECs was enhanced both by recombinant Nogo-66 proteinand by Nogo-A exogenously expressed by Cos-7 cells orendogenously by OLs. Therefore, one possibility to account forthe influence of Nogo-66 on OEC migration was that Nogo-66affected the adhesion of OECs. Adhesion sites form as a resultof signaling between the extracellular matrix on the outside andthe actin cytoskeleton on the inside, and they are associatedwith specific assemblies of actin filaments (Kaibuchi et al.,1999; Kaverina et al., 2002). Paxillin is a cytoskeletalcomponent localized to the long-lived focal adhesions at theend of actin stress fibres that serve in longer-term anchorage(Burridge et al., 1998). Protrusion of the cell surface is an early
step in several cellular processes including cell migration. Thedriving force for the formation and extension of membraneprotrusions is the polymerization of actin (DeMali andBurridge, 2003). In the present study, we observed theinfluences of Nogo-66 on focal adhesions, actin stress fibersand membrane protrusions. It was apparent that Nogo-66regulated actin reassembly, promoted the formation of focaladhesions, and inhibited membrane protrusion. Taken together,these data support the hypothesis that Nogo-66 enhances theadhesion of OECs, which inhibits their migration.
Woodhall et al. reported that NgR mRNA is expressed inprimary cultures of OECs (Woodhall et al., 2003). In our study,the expression of NgR in OECs was confirmed further byimmunocytochemistry and western blot. With enzymaticcleavage of NgR and anti-NgR block, we confirmed that theeffect of recombinant Nogo-66 and endogenous Nogo-Aexpressed by OLs on OECs was NgR-dependent. To ourknowledge, this is the first evidence that functional NgR isexpressed on OECs, although the role of p75, the commonmarker for OECs, as an essential co-receptor needs furtherinvestigation. The Rho family of small GTPases have beenreported to be central players in regulating the assembly of the
Fig. 7. The effect of OLs on OECs ismediated by Nogo-A. (A,B) OECmigration is not affected byoligodendrocyte-derived matrix anddiffusible factors. Coverslip fragmentscontaining OECs or OECs-DiI wereinverted on laminin and cultured withculture medium (Laminin), the condition-defined medium (CM), theoligodendrocyte-conditioned medium(OLs-CM), the oligodendrocyte-derivedmatrix with culture medium (OLs-GM),or on a monolayer of oligodendrocyteswith culture medium (OLs-MON). Themigration index and distribution of thenumber of migrated cells were examined.Migration Index (%)=Maximum distancetravelled by OECs as a percentage of themaximum distance traveled by OECs onlaminin. **P<0.01 versus laminin.(C) Cos-7 cells were transfected withNogo-A and immunostained withantibody against Nogo-A without beingpermeabilized with Triton X-100.(D) DiI-labelled OECs were placed overa mock-transfected Cos-7 cell monolayer(Cos-7-Mock), and a Nogo-A-transfectedCos-7 cell monolayer (Cos-7-NogoA).The adherent cells were counted andnormalized against the Cos-7-Mockgroup. (E,F) The cell motility of OECson a monolayer of Cos-7-NogoA wasassessed using inverted coverslipmigration assay. **P<0.01.(G,H) Attenuation of OEC migration byOLs-MON is NgR-dependent. OECswere pre-treated with or without PI-PLC,NgR antibody or goat IgG and applied toinverted migration assay in which OECsmigrate on laminin or OLs-MON. Bars,50 �m.
Jour
nal o
f Cel
l Sci
ence
1884
actin cytoskeleton, adhesion formation and membraneprotrusion (Hall, 1998; Ridley, 2001; DeMali and Burridge,2003), in which RhoA signals the formation and maturation offocal adhesions associated with actin stress fibre bundles, andRac1 and Cdc42 stimulate the formation of protrusions inassociation with lamellipodia and filopodia, respectively.Formation of membrane protrusions is an early central step inthe process of cell migration. When the ratio of activitybetween these small GTPases is no longer optimal forprotrusion and polarization of the cell, the migration will stop(Cox et al., 2001). In the present study, we found that activeRhoA was significantly increased in OECs plated on Nogo-66substrates, which suggests that Nogo binding to NgR leads toactivation of RhoA in OECs. One of the downstream effectorsof RhoA is Rho-kinase (ROCK), which can be inhibitedeffectively by the compound Y-27632 (Narumiya et al., 2000).Inhibition of ROCK stimulates membrane protrusion (Cox etal., 2001; Rottner et al., 1999; Tsuji et al., 2002) and promotescell migration (Nobes and Hall, 1999). Here, we observed that
the effect of Nogo-66 on the adhesion and migration of OECswas greatly attenuated when cells were pretreated with Y-27632. In addition, we found that the Nogo-66-inducedincrease of focal adhesion formation, alteration of actincytoskeletal structure, and inhibition of membrane protrusionwere significantly attenuated by pretreatment of OECs with Y-27632. All these results supported the hypothesis that theactivity of RhoA is involved in the regulation of adhesion andmigration of OECs by Nogo-66. However, further study isnecessary to clarify the detailed intracellular signalingmechanisms.
It is a continuing challenge to move toward therapeuticapproaches for CNS injury. Recently, there has been greatinterest in the possibility that OECs have potential for use inthe treatment of axonal injuries and demyelinating disease(Franklin and Barnett, 2000; Raisman, 2001; Ramon-Cuetoand Santos-Benito, 2001). Upgrading the growth-promotingproperties of OECs is considered a valuable strategy forpromoting CNS repair; however, most of these studies involveOECs secreting additional neurotrophic factors (Cao et al.,2004; Ruitenberg et al., 2003; Ruitenberg et al., 2005). Sincethe inhibition of axonal outgrowth by CNS myelin is one ofthe major obstacles to functional recovery following CNSinjury, myelin inhibitors and their receptors are recentlyemerging as potential therapeutic targets (Lee et al., 2003).Neutralizing Nogo-A with IN-1 antibody and NgR antagonistswere proven to improve CNS axon regeneration and functionalrecovery after various lesions (Fouad et al., 2004; GrandPre etal., 2002). In the present study, we found that OEC migrationwas facilitated in vitro and in vivo when NgR was neutralizedwith anti-NgR antibody. It is of interest to test the idea that,combined with OEC implantation, local administration ofNogo-A antibody or NgR antagonists would improve thetherapeutic properties of OECs on CNS injury.
Materials and MethodsCell culturePrimary OECs were prepared from the olfactory bulb of adult Sprague-Dawley ratsand purified by differential cell adhesiveness (Ramon-Cueto et al., 1998). Briefly,OECs were extracted from the olfactory nerve layer by trypsin treatment and platedon two uncoated 25 cm2 culture flasks; each was incubated for 36 hours at 37°C in5% CO2. The non-adhesive cell suspension was collected and then seeded onto diskspre-coated with poly-L-lysine (PLL, 0.1 mg/ml), and incubated with serum-containing DMEM/F-12 supplemented with 2 �M forskolin (Sigma) and 10 ng/mlbFGF (Sigma).
Schwann cells (SCs) were obtained from sciatic nerves of 2-day-old Sprague-Dawley rat pups and purified using a modification of the protocol previouslydescribed (Lakatos et al., 2000; Brockes et al., 1979). Cells were cultured on poly-L-lysine (100 �g/ml)-coated dishes and maintained in DMEM/F-12 containing 15%FBS and supplemented with Forskolin (2 �M, Sigma) and bFGF (10 ng/ml, Sigma).The cultures were treated with cytosine arabinoside (10–5 M, Sigma) to reducecontamination by fibroblasts.
Oligodendrocytes (OLs) were prepared from rat cerebral cortex as described(McCarthy and de Vellis, 1980; Fok-Seang et al., 1995). In brief, newborn ratcortical cells were dissociated and cultured in DMEM/F-12 containing 10% FBS.The culture medium was changed at 24 hours and twice weekly thereafter. After10-12 days, the cultures became confluent and loosely attached macrophages wereremoved by shaking the flasks on a rotary shaker at 260 rpm for 1 hour at 37°C.The supernatant was discarded and the cultures were then shaken in fresh culturemedium for 18 hours at 260 rpm to remove oligodendrocyte precursor cells fromthe cell monolayer. The detached cells were plated onto a non-coated culture dishfor 7 minutes to remove adherent cells such as microglia and astrocytes. Theoligodendrocyte precursor cells were then transferred to a culture dish pre-coatedwith PLL and left to adhere overnight. From the next day on, the medium wasexchanged for serum-free chemical defined DMEM/F-12 supplemented with 5�g/ml insulin, 50 �g/ml transferrin, 0.66 mg/ml BSA, 20 ng/ml progesterone, 100�mol/ml putrescine, 40 ng/ml sodium selenite and 30 nmol/ml T3, and theoligodendrocyte precursor cells were cultured for another 1-10 days to differentiate
Journal of Cell Science 120 (11)
Fig. 8. The migration of implanted OECs is facilitated by neutralizingNgR in a spinal cord hemisection injury model. (A) The animalsimplanted with DiI-labelled OECs were treated with N.S., anti-NgRantibody or goat IgG every day through a pipe embedded in thesubarachnoid space. After 10 days, the length, width and area of theregion invaded by DiI-postive cells were measured by usingMetaMorph Imaging System analysis software. The three right panelsare locally magnified images of the three left panels. (B) Quantitativeassessment of cell migration in vivo. Migration Index (%)=Maximumlength, width or area traveled by DiI-positive cells in the presence ofanti-NgR antibody or irrelevant IgG as a percentage of the maximumlength, width or area traveled by DiI-positive cells in the presence ofN.S. **P<0.01 versus N.S. Bar, 50 �m.
Jour
nal o
f Cel
l Sci
ence

1885Nogo regulates OEC migration
into OLs. OLs were identified by indirect immunofluorescence labeling usingmonoclonal anti-oligodendrocyte marker O1 and anti-myelin basic protein (MBP).
Unless indicated otherwise, cells were pretreated with or without PI-PLC (0-0.1U/ml), anti-NgR (1:100), IgG (1:100) or Y-27632 (15 �M) for 2 hours before contactwith Nogo-66. For PI-PLC treatment, cells were washed once with 0.1 M PBS (PH7.4) and then incubated with phosphatidylinositol-specific phospholipase C (PI-PLC,Sigma; 0.001, 0.01, 0.1 U/ml) in DMEM/F-12 at 37°C for the times indicated. Thecells were washed three times and then processed for the migration and adhesionassay or for western blot analysis. To confirm cell viability, cells were stained with0.4% Trypan Blue. Trypan-Blue-incorporating cells were <1% in each experiment.
Fluorescent stainingFor immunocytochemical analysis, immunocytochemistry of cells cultured oncoverslips were performed as previously described (Yan et al., 2003). Briefly,coverslips were fixed with 4% paraformaldehyde in PBS for 20 minutes, thenpermeabilized with or without 0.3% Triton X-100 in 0.1 M PBS for 15 minutes andincubated overnight at 4°C with polyclonal antibodies to NgR (Santa Cruz), S-100(Boster) or Nogo-A (Santa Cruz), or monoclonal antibodies to paxillin (BDTransduction Laboratories), O1 (R&D Systems) or MBP (myelin basic protein,Chemicon international) diluted in PBS containing 10% normal goat serum. Afterwashing three times with 0.1 M PBS (pH 7.4), cells were incubated withfluorescence-conjugated secondary antibody (Sigma) for 90 minutes at roomtemperature. For visualization of F-actin, cells were incubated with 0.1 �g/mlTRITC-phalloidin (Sigma) overnight at 4°C. Cells were then examined byfluorescence microscopy.
Boyden chamber migration assayTo measure the ability of OECs to migrate, OEC migration studies were performedusing a 24-well Boyden chamber (Costar) containing polycarbonate membranes (8�m pore size), with slight modification of a previously described protocol (Klemkeet al., 1997; Yan and Rivkees, 2002; Yamauchi et al., 2003). In brief, the undersidesof polyethylene terephthalate filter membranes were coated with laminin or fusionproteins (100 ng/ml diluted in laminin) overnight at 4°C. Cells were detached bytrypsin/EDTA and then seeded onto the upper chamber at a density of 4�105 cellsin 250 �l of culture medium containing 1% serum per well. The upper chamberswere inserted into the tissue-culture wells and 750 �l culture medium containing1% serum was added to the lower chambers. After incubation for 8 hours at 37°C,non-migratory cells on the upper membrane surface were removed with a cottonswab, and migratory cells migrating through the membrane pores and invading tothe underside surface of the membrane were fixed with 4% paraformaldehyde andstained with Coomassie Brilliant Blue. For quantitative assessment, the number ofstained, migrating cells was then counted under a microscope at five fields per filterin three independent experiments.
Inverted coverslip migration assayThe inverted migration assay was performed according to a previous protocol (Fok-Seang et al., 1995). To prelabel SCs or OECs, cells were incubated with 25 �g/mlDi-I (Molecular Probes, a sulfonated carbocyanine fluorescent tracer: 3,3�-dioctadecyloxacarbocyanine perchlorate) for 5 min at 37°C. Coverslips (8 mm) werebroken to produce an approximately 1�2 mm fragment. Di-I-labeled or nonlabeledcells were plated onto the coverslip fragments precoated with PLL. After the cellshad been allowed to attach for 16-18 hours, the coverslips were washed with culturemedium three times to remove any loose cells and then inverted so that cells faceddownward onto tissue culture surfaces coated with laminin, GST, GST–Nogo-66,His or His–Nogo-66, or onto cell monolayers or extracellular matrix. These cultureswere then incubated for a further 3 days to allow cell migration from the edge of theinverted fragment and fixed with 4% paraformaldehyde. The maximum migrationdistance from the edge of each of the coverslip fragments was measured and thenumber of cells present within rows (50 �m � 1 mm area) progressing outward fromthe edge of the coverslip fragment was counted. Experiments were carried out in thepresence of 12 �g/ml aphidicolin (Sigma), a mitotic inhibitor, in order to be certainthat movement away from the coverslip fragment was due to migration alone anddid not contribute to proliferation.
Preparation of culture conditioned medium and matrixOligodendrocyte-conditioned serum-free medium was obtained by growingconfluent OLs for 2 days in condition-defined medium that consisted of DMEM/F-12 supplemented with 1% N2 (vol/vol), 10.1 ng/ml T3, 400 ng/ml T4, 0.035%bovine serum albumin (BSA) and 20 �M leupeptin. Oligodendrocyte-conditionedserum-free medium prepared in this way was filtered (0.22 �m filter) and usedwithout further dilution. Extracellular matrix from OLs was obtained fromoligodendrocyte monolayers by incubating these cells in distilled water for 2 hours.Cellular debris was removed by several washes with culture medium prior to assay.
Cells adhesion assayThe adherent ability of OECs when contacted with Nogo-66 was detected by twodifferent approaches. OECs or SCs were plated on recombinant GST or His and therespective GST or His fusion proteins (10-100 �g/ml solution in 1 �l-sized spots)
that were dried down onto dishes coated with poly-L-lysine (PLL). Cells wereincubated at 37°C before fixing with 4% paraformaldehyde after various time points(Dorries et al., 1996). The number of cells attached to GST or His fusion proteinspots were then counted under a microscope. To analyze the adhesion of OECs tosubstrates or cell monolayers, a total of 20,000 Di-I-labeled OECs or SCs wereplaced onto 24-well plates coated with laminin GST, GST–Nogo-66, His orHis–Nogo-66, or a complete monolayer of OLs or Cos-7 cells transfected withNogo-A. The cultures were incubated for 30 minutes at 37°C on a rotary shaker at25 rpm. After several washes with culture medium, cultures were fixed in 4%paraformaldehyde for 20 minutes at room temperature and the number of adheringcells was determined by counting the number of stained cells in a counting gridunder a 20� objective and repeating the counts in adjacent grids to cover onecomplete diameter of each well.
Western blot analysisTo detect the expression of NgR, OECs, SCs (as negative control) and PC12 cells(as positive control) were harvested after being rinsed briefly with ice-cold PBS andlysed in SDS gel sample buffer. To confirm the ability of PI-PLC to release NgRfrom the membrane, the supernatant and membrane fractions of OECs werecollected after cells were treated with 0.1 U/ml PI-PLC. The samples weredenatured by boiling for 10 minutes with SDS gel sample buffer, then centrifugedfor 10 minutes at 12,000 g at 4°C. Proteins in the supernatants were separated by12% SDS-polyacrylamide gel and then transferred onto nitrocellulose membranes.Membranes were then blocked with 10% low-fat milk in 1� TBST and incubatedwith specific primary antibody against NgR (Chemicon). To control for differencesin protein loading, membranes were also incubated with anti-GAPDH antibody(Sigma). After incubating with horseradish peroxidase (HRP)-conjugated secondaryantibodies (Sigma; 1:10,000), immunoreactive bands were visualized bychemiluminescence reagents (ECL, Amersham).
RhoA activity assayActive RhoA was determined with the GST-Rhotekin-binding domain as describedpreviously (Ren et al., 1999). OECs were plated onto the dishes coated with Nogo-66 fusion proteins (GST–Nogo-66 or His–Nogo-66) or control proteins (GST orHis) for 6 hours. Cells were washed with ice-cold PBS and lysed in RIPA buffer(50 mM Tris, pH 7.2, 1% Triton X-100, 0.5% sodium deoxycholate, 0.1% SDS,500 mM NaCl, 10 mM MgCl2, 10 �g/ml each of leupeptin and aprotinin, 1 mMphenylmethylsulfonyl fluoride). Cell lysates were clarified by centrifugation at12,000 rpm at 4°C for 5 minutes, and equal volumes of lysates were incubated withGST-Rhotekin-binding domain (20 �g) bound to beads at 4°C for 60 minutes. Thebeads were then washed four times in buffer B (Tris buffer containing 1% TritonX-100, 150 mM NaCl, 10 mM MgCl2, 10 �g/ml each of leupeptin and aprotinin,and 0.1 mM phenylmethylsulfonyl fluoride) at 4°C. Bound RhoA proteins weredetected by western blotting using a monoclonal antibody against RhoA (SantaCruz, 1:500). The amount of GTP-bound RhoA was normalized to the total amountof RhoA in cell lysates as previously described (Ren et al., 1999), and statisticalanalysis was performed for the results of three independent experiments.
In vivo migration assayTo investigate whether Nogo affected the migration of implanted OECs, an in vivomigration assay was performed as previously described (Cao et al., 2006). Animalcare and use followed recommended NIH guidelines. Female adult Sprague-Dawleyrats (200-250 g) were anaesthetized with 2% pentobarbital sodium. Laminectomywas performed to expose the dorsal surface of the T7-9 segment, followed by ahemisection at T8 using scalpels. A 1 mm segment was removed on the left side ofthe T8 spinal cord. Animals were divided into three experimental groups: (1) Normalsaline (N.S.) group (n=6), (2) anti-NgR group (n=8) and (3) IgG group (n=7). Di-I-labeled OECs were deposited into two injection sites at the rostral as well as thecaudal stump, � mm from the lesion cavity, using a sterile glass needle. A volumeof 0.5 �l containing 5�105 cells in DMEM was grafted into each site. Rats in group1 were injected with untreated OECs and received N.S. (10 �l/day) through a pipeembedded in the subarachnoid space. In group 2, rats were injected with OECspretreated with anti-NgR antibody (Santa Cruz, 1:100) for 30 minutes and receivedthe antibody (10 �l/day) instead of N.S. In group 3, rats were injected with OECspretreated with an irrelevant goat IgG (1:100) for 30 minutes and received the IgG(10 �l/day).
Ten days after injury, the experimental animals were perfused and the fixed spinalcords (T7-T9) were post-fixed for 8 hours in 4% PFA. A series of consecutive sagittalsections (8 �m) were cut and collected in PBS. To quantify the motility of theimplanted OECs in the spinal cord, we selected five representative midsagittalsections in each animal and measured the size of the region invaded by Di-I-labeledcells from the graft site under a fluorescence microscope (IX70, Olympus). Todescribe the motorial characteristic of grafted OECs in detail, the area covered bygrafted OECs (area) and the maximum migration distance along the vertical (length)or horizontal (width) direction were included in the parameters measured.
Statistical analysisAll data present represent the results of at least three independent experiments, using
Jour
nal o
f Cel
l Sci
ence

1886
cells prepared at different times. Statistical analysis was performed using unpairedStudent’s t-test. All data are presented as mean ± s.d.
We thank Dana Dodd in the Schwab lab for the gift of plasmidencoding Nogo-A. This work was supported by the National KeyBasic Research Program (2005CB724302, 2006CB500702), theNational Natural Science Foundation (30400128, 30325022,30530240), the Program for Changjiang Scholars and InnovativeResearch Team in University (IRT0528), the Shanghai Young Scienceand Technology Phosphor Projects (04QMX1437), and the ShanghaiMetropolitan Fund for Research and Development (04DZ14005,04XD14004).
ReferencesAcevedo, L., Yu, J., Erdjument-Bromage, H., Miao, R. Q., Kim, J. E., Fulton, D.,
Tempst, P., Strittmatter, S. M. and Sessa, W. C. (2004). A new role for Nogo as aregulator of vascular remodeling. Nat. Med. 10, 382-388.
Allen, W. E., Jones, G. E., Pollard, J. W. and Ridley, A. J. (1997). Rho, Rac and Cdc42regulate actin organization and cell adhesion in macrophages. J. Cell Sci. 110, 707-720.
Bandtlow, C. E., Dlaska, M., Pirker, S., Czech, T., Baumgartner, C. and Sperk, G.(2004). Increased expression of Nogo-A in hippocampal neurons of patients withtemporal lobe epilepsy. Eur. J. Neurosci. 20, 195-206.
Barnett, S. C. and Riddell, J. S. (2004). Olfactory ensheathing cells (OECs) and thetreatment of CNS injury; advantages and possible caveats. J. Anat. 24, 57-67.
Brittis, P. A. and Flanagan, J. G. (2001). Nogo domains and a Nogo receptor:implications for axon regeneration. Neuron 30, 11-14.
Brockes, J. P., Field, K. L. and Raff, M. C. (1979). Studies on cultured rat Schwanncells. I. Establishment of purified populations from cultures of peripheral nerve. BrainRes. 165, 105-118.
Burridge, K., Fath, K., Kelly, T., Nuckolls, G. and Turner, C. (1988). Focal adhesions:transmembrane junctions between the extracellular matrix and the cytoskeleton. Annu.Rev. Cell Biol. 4, 487-525.
Cao, L., Liu, L., Chen, Z. Y., Wang, L. M., Ye, J. L., Qiu, H. Y., Lu, C. L. and He,C. (2004). Olfactory ensheathing cells genetically modified to secrete GDNF topromote spinal cord repair. Brain 127, 535-549.
Cao, L., Su, Z. D., Zhou, Q., Lv, B. L., Liu, X. J., Jiao, L., Li, Z. H., Zhu, Y. L.,Huang, Z. H., Huang, A. J. and He, C. (2006). Glial cell line-derived neurotrophicfactor promotes olfactory ensheathing cells migration. Glia 54, 536-544.
Chen, M. S., Huber, A. B., van der Haar, M. E., Frank, M., Schnell, L., Spillmann,A. A., Christ, F. and Schwab, M. E. (2000). Nogo-A is a myelin-associated neuriteoutgrowth inhibitor and an antigen for monoclonal antibody IN-1. Nature 403, 434-439.
Collazos-Castro, J. E., Muneton-Gomez, V. C. and Nieto-Sampedro, M. (2005).Olfactory glia transplantation into cervical spinal cord contusion injuries. J. Neurosurg.Spine 3, 308-317.
Cox, E. A., Sastry, S. K. and Huttenlocher, A. (2001). Integrin-mediated adhesionregulates cell polarity and membrane protrusion through the Rho family of GTPases.Mol. Biol. Cell 12, 265-277.
DeMali, K. A. and Burridge, K. (2003). Coupling membrane protrusion and celladhesion. J. Cell Sci. 116, 2389-2397.
Deng, C., Gorrie, C., Hayward, I., Elston, B., Venn, M., Mackay-Sim, A. and Waite,P. (2006). Survival and migration of human and rat olfactory ensheathing cells in intactand injured spinal cord. J. Neurosci. Res. 83, 1201-1212.
Domeniconi, M., Cao, Z., Spencer, T., Sivasankaran, R., Wang, K., Nikulina, E.,Kimura, N., Cai, H., Deng, K. et al. (2002). Myelin-associated glycoprotein interactswith the Nogo66 receptor to inhibit neurite outgrowth. Neuron 35, 283-290.
Dorries, U., Taylor, J., Xiao, Z., Lochter, A., Montag, D. and Schachner, M. (1996).Distinct effects of recombinant tenascin-C domains on neuronal cell adhesion, growthcone guidance, and neuronal polarity. J. Neurosci. Res. 43, 420-438.
Doucette, J. R., Kiernan, J. A. and Flumerfelt, B. A. (1983). The re-innervation ofolfactory glomeruli following transection of primary olfactory axons in the central orperipheral nervous system. J. Anat. 137, 1-19.
Fok-Seang, J., Mathews, G. A., ffrench-Constant, C., Trotter, J. and Fawcett, J. W.(1995). Migration of oligodendrocyte precursors on astrocytes and meningeal cells.Dev. Biol. 171, 1-15.
Fouad, K., Klusman, I. and Schwab, M. E. (2004). Regenerating corticospinal fibersin the Marmoset (Callitrix jacchus) after spinal cord lesion and treatment with the anti-Nogo-A antibody IN-1. Eur. J. Neurosci. 20, 2479-2482.
Fournier, A. E., GrandPre, T. and Strittmatter, S. M. (2001). Identification ofa receptor mediating Nogo-66 inhibition of axonal regeneration. Nature 409, 341-346.
Franklin, R. J. M. and Barnett, S. C. (2000). Olfactory ensheathing cells and CNSregeneration: the sweet smell of success? Neuron 28, 15-18.
Franklin, R. J. M., Gilson, J. M., Franceschini, I. A. and Barnett, S. C. (1996).Schwann cell-like myelination following transplantation of an olfactorybulbensheathing cell line into areas of demyelination in the adult CNS. Glia 17, 217-224.
GrandPre, T., Nakamura, F., Vartanian, T. and Strittmatter, S. M. (2000).
Identification of the Nogo inhibitor of axon regeneration as a Reticulon protein. Nature403, 439-444.
GrandPre, T., Li, S. and Strittmatter, S. M. (2002). Nogo-66 receptor antagonistpeptide promotes axonal regeneration. Nature 417, 547-551.
Graziadei, P. P. and Monti Graziadei, G. A. (1980). Neurogenesis and neuronregeneration in the olfactory system of mammals. III. Deafferentation andreinnervation of the olfactory bulb following section of the fila olfactoria in rat. J.Neurocytol. 9, 145-162.
Gudino-Cabrera, G. and Nieto-Sampedro, M. (2000). Schwann-like macroglia in adultrat brain. Glia 30, 49-63.
Gudino-Cabrera, G., Pastor, A. M., de la Cruz, R. R., Delgado-Garcia, J. M. andNieto-Sampedro, M. (2000). Limits to the capacity of transplants of olfactory glia topromote axonal regrowth in the CNS. Neuroreport 11, 467-471.
Hall, A. (1998). Rho GTPases and the actin cytoskeleton. Science 279, 509-514.He, W., Lu, Y., Qahwash, I., Hu, X. Y., Chang, A. and Yan, R. (2004). Reticulon family
members modulate BACE1 activity and amyloid-beta peptide generation. Nat. Med.10, 959-965.
He, Z. and Koprivica, V. (2004). The Nogo singnaling pawthway for regeneration block.Annu. Rev. Neurosci. 27, 341-368.
Huber, A. B., Weinmann, O., Brosamle, C., Oertle, T. and Schwab, M. E. (2002).Patterns of nogo mRNA and protein expression in the developing and adult rat andafter CNS lesions. J. Neurosci. 22, 3553-3567.
Hunt, D., Coffin, R. S., Prinjha, R. K., Campbell, G. and Anderson, P. N. (2003).Nogo-A expression in the intact and injured nervous system. Mol. Cell. Neurosci. 24,1083-1102.
Imaizumi, T., Lankford, K. L., Burton, W. V., Fodor, W. L. and Kocsis, J. D. (2000).Xenotransplantation of transgenic pig olfactory ensheathing cells promotes axonalregeneration in rat spinal cord. Nat. Biotechnol. 18, 949-953.
Kaibuchi, K., Kuroda, S. and Amano, M. (1999). Rgulation of the cytoskeleton andcell adhesion by the Rho family GTPases in mammalian cells. Annu. Rev. Biochem.68, 459-486.
Karnezis, T., Mandemakers, W., McQualter, J. L., Zheng, B., Ho, P. P., Jordan, K.A., Murray, B. M., Barres, B., Tessier-Lavigne, M. and Bernard, C. C. (2004). Theneurite outgrowth inhibitor Nogo-A is involved inautoimmune-mediateddemyelination. Nat. Neurosci. 7, 736-744.
Kaverina, I., Krylyshkina, O. and Small, J. V. (2002). Regulation of substrate adhesiondynamics during cell motility. Int. J. Biochem. Cell Biol. 34, 746-761.
Klemke, R. L., Cai, S., Giannini, A. L., Gallagher, P. J., de Lanerolle, P. and Cheresh,D. A. (1997). Regulation of cell motility by mitogen-activated protein kinase. J. CellBiol. 137, 481-492.
Lakatos, A., Franklin, R. J. M. and Barnett, S. C. (2000). Olfactory ensheathing cellsand Schwann cells differ in their in vitro interactions with astrocytes. Glia 32, 214-225.
Lee, D. H., Strittmatter, S. M. and Sah, D. W. (2003). Targeting the Nogo receptor totreat central nervous system injuries. Nat. Rev. Drug Discov. 2, 872-878.
Li, Y., Field, P. M. and Raisman, G. (1997). Repair of adult rat corticospinal tract bytransplants of olfactory ensheathing cells. Science 277, 2000-2002.
Li, Y., Field, P. M. and Raisman, G. (1998). Regeneration of adult rat corticospinalaxons induced by transplanted olfactory ensheathing cells. J. Neurosci. 18, 10514-10524.
Li, Y., Carlstedt, T., Berthold, C. H. and Raisman, G. (2004). Interaction oftransplanted olfactory-ensheathing cells and host astrocytic processes provides abridge for axons to regenerate across the dorsal root entry zone. Exp. Neurol. 188,300-308.
López-Vales, R., Forés, J., Verdú, E. and Navarro, X. (2006). Acute and delayedtransplantation of olfactory ensheathing cells promote partial recovery after completetransaction of the spinal cord. Neurobiol. Dis. 21, 57-68.
Lu, J., Feron, F., Ho, S. M., Mackay-Sim, A. and Waite, P. M. (2001). Transplantationof nasal olfactory tissue promotes partial recovery in paraplegic adult rats. Brain Res.889, 344-357.
Lu, J., Feron, F., Mackay-Sim, A. and Waite, P. M. (2002). Olfactory ensheathing cellspromote locomotor recovery after delayed transplantation into transected spinal cord.Brain 125, 14-21.
McCarthy, K. D. and de Vellis, J. (1980). Preparation of separate astroglial andoligodendroglial cell cultures from rat cerebral tissue. J. Cell Biol. 85, 890-902.
Meier, S., Brauer, A. U., Heimrich, B., Schwab, M. E., Nitsch, R. and Savaskan, N.E. (2003). Molecular analysis of Nogo expression in the hippocampus duringdevelopment and following lesion and seizure. FASEB J. 17, 1153-1155.
Moreira, E. F., Jaworski, C. J. and Rodriguez, I. R. (1999). Cloning of a novel memberof the reticulon gene family (RTN3): gene structure and chromosomal localization to11q13. Genomics 58, 73-81.
Narumiya, S., Ishizaki, T. and Uehata, M. (2000). Use and properties of ROCKspecificinhibitor Y-27632. Methods Enzymol. 325, 273-284.
Nash, H. H., Borke, R. C. and Anders, J. J. (2002). Ensheathing cells andmethylprednisolone promote axonal regeneration and functional recovery in thelesioned adult rat spinal cord. J. Neurosci. 22, 7111-7120.
Ng, W. P., Cartel, N., Roder, J., Roach, A. and Lozano, A. (1996). Human centralnervous system myelin inhibits neurite outgrowth. Brain Res. 720, 17-24.
Nie, D. Y., Zhou, Z. H., Ang, B. T., Teng, F. Y., Xu, G., Xiang, T., Wang, C. Y., Zeng,L., Takeda, Y., Xu, T. L. et al. (2003). Nogo-A at CNS paranodes is a ligand of Caspr:possible regulation of K(+) channel localization. EMBO J. 22, 5666-5678.
Nobes, C. D. and Hall, A. (1995). Rho, Rac, and Cdc42 GTPases regulate the assemblyof multimolecular focal complexes associated with actin stressfibers, lamellipodia andfilopodia. Cell 81, 53-62.
Journal of Cell Science 120 (11)
Jour
nal o
f Cel
l Sci
ence

1887Nogo regulates OEC migration
Nobes, C. D. and Hall, A. (1999). Rho GTPases control polarity, protrusion, and adhesionduring cell movement. J. Cell Biol. 144, 1235-1244.
Plant, G. W., Currier, P. F., Cuervo, E. P., Bates, M. L., Pressman, Y., Bunge, M. B.and Wood, P. M. (2002). Purified adult ensheathing glia fail to myelinate axons underculture conditions that enable schwann cells to form myelin. J. Neurosci. 22, 6083-6091.
Prinjha, R., Moore, S. E., Vinson, M., Blake, S., Morrow, R., Christie, G.,Michalovich, D., Simmons, D. L. and Walsh, F. S. (2000). Inhibitor of neuriteoutgrowth in humans. Nature 403, 383-384.
Raisman, G. (2001). Olfactory ensheathing cells – another miracle cure for spinal cordinjury? Nat. Rev. Neurosci. 2, 369-374.
Ramon-Cueto, A. and Avila, J. (1998). Olfactory ensheathing glia: properties andfunction. Brain Res. Bull. 46, 175-187.
Ramon-Cueto, A., Plant, G. W., Avila, J. and Bunge, M. B. (1998) Long-distanceaxonal regeneration in the transected adult rat spinal cord is promoted by olfactoryensheathing glia transplants. J. Neurosci. 18, 3803-3815.
Ramon-Cueto, A., Cordero, M. I., Santos-Benito, F. F. and Avila, J. (2000). Functionalrecovery of paraplegic rats and motor axon regeneration in their spinal cords byolfactory ensheathing glia. Neuron 25, 425-435.
Ramon-Cueto, A. and Santos-Benito, F. F. (2001). Cell therapy to repair injured spinalcords: olfactory ensheathing glia transplantation. Restor. Neurol. Neurosci. 19, 149-156.
Ren, X. D., Kiosses, W. B. and Schwartz, M. A. (1999). Regulation of the small GTP-binding protein Rho by cell adhesion and the cytoskeleton. EMBO J. 18, 578-585.
Resnick, D. K., Cechvala, C. F., Yan, Y., Witwer, B. P., Sun, D. and Zhang, S. (2003).Adult olfactory ensheathing cell transplantation for acute spinal cord injury. J.Neurotrauma 20, 279-285.
Richter, M. W., Fletcher, P. A., Liu, J., Tetzlaff, W. and Roskams, A. J. (2005). Laminapropria and olfactory bulb ensheathing cells exhibit differential integration andmigration and promote differential axon sprouting in the lesioned spinal cord. J.Neurosci. 25, 10700-10711.
Ridley, A. J. (2001). Rho proteins, PI 3-kinases and monocyte/macrophage motility.FEBS Lett. 498, 168-171.
Rottner, K., Hall, A. and Small, J. V. (1999). Interplay between Rac and Rho in thecontrol of substrate contact dynamics. Curr. Biol. 9, 640-648.
Ruitenberg, M. J., Plant, G. W., Hamers, F. P., Wortel, J., Blits, B., Dijkhuizen, P.A., Gispen, W. H., Boer, G. J. and Verhaagen, J. (2003). Ex vivo adenoviral vector-mediated neurotrophin gene transfer to olfactory ensheathing glia: effects onrubrospinal tract regeneration, lesion size, and functional recovery after implantationin the injured rat spinal cord. J. Neurosci. 23, 7045-7058.
Ruitenberg, M. J., Levison, D. B., Lee, S. V., Verhaagen, J., Harvey, A. R. and Plant,G. W. (2005). NT-3 expression from engineered olfactory ensheathing glia promotesspinal sparing and regeneration. Brain 128, 839-853.
Schwab, M. E. and Bartholdi, D. (1996). Degeneration and regeneration of axons in thelesioned spinal cord. Physiol. Rev. 76, 319-370.
Smale, K. A., Doucette, R. and Kawaja, M. D. (1996). Implantation of olfactoryensheathing cells in the adult rat brain following fimbria-fornix transection. Exp.Neurol. 137, 225-233.
Tagami, S., Eguchi, Y., Kinoshita, M., Takeda, M. and Tsujimoto, Y. (2000). A novelprotein, RTN-XS, interacts with both Bcl-xL and Bcl-2 on endoplasmic reticulum andreduces their antiapoptotic activity. Oncogene 19, 5736-5746.
Takami, T., Oudega, M., Bates, M., Wood, P., Kleitman, N. and Bunge, M. (2002).Schwann cell but not olfactory ensheathing glia transplants improve hindlimblocomotor performance in the moderately contused adult rat thoracic spinal cord. J.Neurosci. 22, 6670-6681.
Tennent, R. and Chuah, M. I. (1996). Ultrastructural study of ensheathing cells in earlydevelopment of olfactory axons. Dev. Brain Res. 95, 135-139.
Tisay, K. T. and Key, B. (1999). The extracellular matrix modulates olfactory neuriteoutgrowth on ensheathing cells. J. Neurosci. 19, 9890-9899.
Tsuji, T., Ishizaki, T., Okamoto, M., Higashida, C., Kimura, K., Furuyashiki, T.,Arakawa, Y., Birge, R. B., Nakamoto, T., Hirai, H. et al. (2002). ROCK and mDia1antagonize in Rho-dependent Rac activation in Swiss 3T3 fibroblasts. J. Cell Biol. 157,819-830.
Van Aelst, L. and D’Souza-Schorey, C. (1997). Rho GTPases and signaling networks.Genes Dev. 11, 2295-2322.
Wang, K. C., Kim, J. A., Sivasankaran, R., Segal, R. and He, Z. (2002). P75 interactswith the Nogo receptor as a co-receptor for Nogo, MAG and OMgp. Nature 420, 74-78.
Wong, S. T., Henley, J. R., Kanning, K. C., Huang, K. H., Bothwall, M. and Poo, M.M. (2002). A p75(NTR) and Nogo receptor complex mediates repulsive signaling bymyelin-associated glycoprotein. Nat. Neurosci. 5, 1302-1308.
Woodhall, E., West, A. K., Vickers, J. C. and Chuah, M. I. (2003). Olfactoryensheathing cell phenotype following implantation in the lesioned spinal cord. Cell.Mol. Life Sci. 60, 2241-2253.
Yamashita, T., Higuchi, H. and Tohyama, M. (2002). The p75 receptor transduces thesignal from myelin-associated glycoprotein to Rho. J. Cell Biol. 157, 565-570.
Yamauchi, J., Chan, J. R. and Shooter, E. M. (2003). Neurotrophin 3 activation of TrkCinduces Schwann cell migration through the c-Jun N-terminal kinase pathway. Proc.Natl. Acad. Sci. USA 100, 14421-14426.
Yan, H. and Rivkees, S. A. (2002). Hepatocyte growth factor stimulates the proliferationand migration of oligodendrocyte precursor cells. J. Neurosci. Res. 69, 597-606.
Yan, H., Lu, D. and Rivkees, S. A. (2003). Lysophosphatidic acid regulates theproliferation and migration of olfactory ensheathing cells in vitro. Glia 44, 26-36.
Yiu, G. and He, Z. (2003). Signaling mechanisms of the myelin inhibitors of axonregeneration. Curr. Opin. Neurobiol. 13, 545-551.
Jour
nal o
f Cel
l Sci
ence





























