Embed Size (px)
Citation preview

THE POPULATION BIOLOGY OF BACTERIAL PLASMIDS: A PRZORZ CONDITIONS FOR THE EXISTENCE OF
MOBILIZABLE NONCONJUGATIVE FACTORS
BRUCE R. LEVIN AND FRANK M. STEWART*
Department of Zoology, University of Massachusetts, Amherst, Massachusetts 01003 Department of Genetics, University College, London
Manuscript received February 12, 1979 Revised copy received August 20, 1979
ABSTRACT
A mathematical model for the population dynamics of nonconjugative plasmids that can be mobilized by conjugative factors is presented. In the analy- sis of the properties of this model, primary consideration is given to the condi- tions under which these nonself-transmissible extrachromosomal elements could become established and would be maintained in bacterial populations. The results of this analysis demonstrate the existence of conditions where, as a conse- quence of infectious transmission via mobilization, nonconjugative plasmids could become established and be maintained even when the bacteria carrying them have lower reproductive fitnesses than plasmid-free members of the popu- lation. However, these existence conditions are stringent and suggest, therefore, that it is highly unlikely that plasmids of this type would become established and maintained without some direct selection favoring their carriage. The general implications of these results and limitations of the model are discussed. Brief consideration is also given to the implications of these theoretical findings to the problems of the spread of multiple antibiotic resistance plasmids (R-factors) and the risk of contaminating natural populations of bacteria with chimeric plasmids produced by work with recombinant DNA.
number of the adaptations of bacteria to their environments are deter- A mined by plasmid-borne, rather than chromosomal, genes. Among these characters are resistance to antibiotics and heavy metals (WATANABE 1963; ANDERSON 1969; FALKOW 1975), the ability to ferment “atypical” carbon sources (REINWALD, CHAKRABARTY and GUNSALVES 1973) and to produce toxic and antibiotic sublstances ( SKERMAN, FORMAL and FALKOW 1972; KIRBY et al. 1975). In at least one case, there is evidence that plasmids are responsible for the production of tumors in the eukaryotic (plant) hosts of the bacteria carry- ing those factors, (CHILTON et al. 1977). In most cases, where there are no trans- forming systems, these autonomous hereditary units can be transmitted between cells by phage-mediated transduction (NOVICK and BOUANCHARD 1971). Many plasmids, particularly those in the Enterobacteriaceae, are also transmitted by conjugation (FALKOW 1975).
In an earlier investigation (STEWART and LEVIN 1977), we examined the
* Permanent address: Department of Mathematics, Brown University, Providence, Rhode Island 02912
Genetics 94: 425443 February, 1980.

426 B. R. LEVIN AND F. M. STEWART
population biology of a class of conjugationally transmitted plasmids where the genes coding for replication, conjugation and other plasmid characters are trans- mitted as a single inviolate element, i.e., ”conjugative plasmids” ( NOVICK et al. 1976). In this theoretical investigation, we demonstrated that, as a consequence of infectious transmission, there exists a broad set of (what seem to be) biologi- cally possible conditions where nonantibiotic (nonbacteriocin) -determining plasmids could become established and would be maintained at substantial fre- quencies in bacterial populations, even when the carriage of these plasmids results in a significant reduction in the reproductive fitness of their host cells.
There is a second major class of conjugationally transmitted plasmids to be considered: those for which the genes determining replication and other char- acters exist apart from those coding for conjugation. The conjugational trans- mission of these nonconjugative plasmids requires mobilization by conjugative factors that co-exist in their host cell (NOVICK et al. 1976). On the average, non- conjugative plasmids are smaller than conjugative factors, but, as a class, they seem to code fcr as broad a set of phenotypes as the self-transmissible elements (FALKOW 1975). In recent years, nonconjugative plasmids have come into par- ticular prominence as cloning vectors for work with recombinant DNA (NIH 1976).
In this investigation, we present a mathematical model for the population biology of nonconjugative plasmids that can be mobilized by conjugative factors. In the analysis of the properties of this model, we give primary consideration to the conditions under which cells carrying these nonself-transmissible plasmids can invade and become established in bacterial populations. We demonstrate that, as a consequence of their capacity for infectious transmission by mobiliza- tion, nonconjugative plasmids could become established and be maintained even when the bacteria carrying them have a lower reproductive fitness than plasmid- free cells. However, in spite of this possibility, we argue that it is highly unlikely that these nonself-transmissible hereditary units could exist without some selec- tion favoring the genes they are carrying.
A model for the mobilization transfer of nonconjugative plasmids The model presented here is an extension of one that we used in a study of
the population dynamics oE conjugative plasmids (STEWART and LEVIN 1977). We restrict the present consideration to the equable (chemostat) mode of re- source exploitation. A liquid containing a unique and limiting resource at a concentration c(pg/ml) enters a culture vessel (the “habitat”) at a constant rate, which is equal to the rate at which excess resource, wastes and organisms are removed. Let p be the rate of flow through this habitat as measured in turn- overs (fraction of the habitat exchanged) per hour and r ( p g / m l ) be the con- centration of the resource within the habitat. Four bacterial populations are considered: ( 1 ) plasmid-free recipients at a density of R/ml; (2) donors, which carry just the nonconjugative plasmid at a density of D/ml; (3) mobilizers, which carry- just the conjugative, mobilizing plasmids at a density of M/ml; and (4) mobilized donors, which carry both plasmids at a density of MD/ml.

CONDITIONS FOR THE EXISTENCE OF NONCONJUGATIVE PLASMIDS 427
These four bacterial populations reproduce at rates $ & ( r ) , $ D ( r ) , $M(r) and qYD(r) per hour, respectively. These rates of population growth are monotonic increasing functions of the resource concentration. I t is assumed that, in order to reproduce, each bacterium must consume e pg per cell of the limiting resource, which is taken up at a rate proportional to their growth rate. For example, individual cells of the recipient clone consume the limiting resource at a rate $R ( r ) e per cell per hour.
We assume that encounters between members of the different cell populations occur randomly at a frequency that is jointly proportional to their respective densities. A fraction of the encounters between cells carrying the conjugative, mobilizing plasmid (mobilizers or mobilized donors) result in the transfer of plasmid( s) to cells free of the conjugative factor (recipients and donors). A unique conjugational transfer rate constant, y ml per cell per hour, governs the rate of plasmid transfer for all “mating” pairs. The receipt of a plasmid results in the loss of that cell from its current clone and the recruitment into the appro- priate plasmid-bearing population. In the model, we assume that plasmid transfer is an instantaneous process and that cells carrying the mobilizing factor are unable to serve as recipients. In Table 1, we present the various “matings” pos- sible and the relative numbers 01 the different transconjugant types produced. Finally, we assume that: (1) cells carrying single plasmids ( D or M ) lose these factors with the same vegetative segregation rate, r per cell per hour, and that all these segregants become members of the R population, and ( 2 ) cells carrying both plasmids, MD, lose single factors at a rate 2~ per cell per hour and that half of the segregants become members of the D population, while the other half enter the M population. With these definitions and assumptions, the rates of change in the concentrations of the limiting resources and bacterial popula- tions are given by the following differential equations:
7 = p(c-r> - e [ R $R ( r ) + M $ ~ ( r ) + D $ D ( I ) + M D $ M D ( ~ ) 1, (1:l)
(1 :2) R = R$R(I) - y ~ ( ~ + M D ) + r ( ~ + D) - p ~ ,
M = M q M ( r ) + y R (M+aMMD) - M ( r + p ) + r M D , ~ = D $ D ( I ) ~ ~ [ R ~ D M D - D ( M + ~ M ’ M D ) ] - D ( r + p ) +TMD, M h = M D $YD ( r ) $. y [RaMD M D -k D ( M + ~ M ’ M D ) ] - M D (27 -k p ) ,
(1 :3)
(1:4)
(1 : 5 )
TABLE 1
Possible mntings and transconjugants produced
Mating Occurrence Transconjugants produced: cells/ml/hr
Number/ml/hr Mobilizers Donors Mobilized donors ~ ~ ~
Mobilizers x recipients 7M.R 7M.R - - Mobilizers x donors 7M.D - - 7M.D Mobilized donors x recepients 7MD.R yMD.RaM yMD.RaD yMD.Ra,, Mobilized donors x donors 7MD.D - - yMD.Da’,
where saY + aD + ayD = 1, and 0 5 5 1

428 B. R. LEVIN A N D F. M. STEWART
where a dot (.) denotes differentiation with respect to time. The parameters CYD, aM and aMD are, respectively, the proportions of transconjugants from matings between donors and recipient that acquire: (i) just the nonconjugative plasmid, (ii) just the mobilizing plasmid, and (iii) both plasmids; a D -I- CYM f a M D = 1. The parameter dM is the proportion of transconjugants from the mating between mobilized donors and donors that receive the mobilizing plasmid.
Conditions for invasion by the nonconjugative plasmid Our analysis of the properties of this model aims to determine the conditions
under which the nonconjugative plasmid could become established and would be maintained in a bacterial population that has been previously free of that factor. These “existence” conditions can be partitioned into two necessary sub- conditions: (1) those permitting the invasion by a plasmid of an equilibrium population of bacteria that is free of that factor, and (2) those allowing for its continued maintenance following invasion.
To facilitate our analysis, we shall consider invasion by the nonconjugative plasmid in two types of populations, namely: (i) equilibrium populations that are free of both conjugative and nonconjugative plasmids, and (ii) populations where there is a stable equilibrium with both plasmid-free bacteria and bacteria carrying the mobilizing, conjugative plasmid present. Both of these cases for invasion can be subsumed under the same general theory. The system of five differential equations, (1 : 1) through (1 : 5) , is of the form
xi = fi (21,22723724725) 7
where x1 = r, xz=R, x3=M, x4=D and x5=MD7
fl ( % Z Z 9 x 3 7 x b x 5 ) = p ( c x l ) - e .X xi$i (zl) (%I)
f i ( 2 1 7 2 2 , 2 3 7 2 4 7 2 5 ) =xZ$’Z (XI) - 7x2 ( 2 3 z 5 ) 7 (XS 2 4 ) - p T Z 7 (2:2) 2=2
f 3 ( 2 1 7 x 2 ? 2 3 , 2 4 , 2 5 ) = 2 3 $ 3 ( Z i ) + y [ x i ( x 3 + ‘3!Mz5) 1 - 5 3 [ 7 + p ] + 7 2 5 , (2:3)
f 4 (21,22,X3,X&5) = 2 ~ $ 4 ( 2 i ) + Y [ 2 ~ ~ ~ 2 5 - x 4 ( 2 3 f c y d l l x 5 ) ] - 2 4 ( T f p ) + 7 x 5 , (24)
and
f5 ( X 1 ~ 2 2 7 5 3 ~ 4 ~ 5 ) = z 5 $ 5 (si) + Y [ x z w D Z ~ + X ~ ( ~ 3 + ~ r ’ 2 5 ) ] - 2 5 ( 2 7 + p ) . (2~5)
(i) Invasion of a plasmid-free population: If the growth rate of the recipient population at the maximum concentration of the resource were sufficient to overcome its loss by flow, i.e.) if gE(c ) > p, then at least the population of recip- ients would maintain a stable equilibrium with the resource in that habitat. In the absense of plasmid-bearing cells, x3 = x4 = x5 = 0, and there would be a stable singular point (zl, zz) of the two-dimensional system of equations.
A A A
A A

CONDITIONS FOR THE EXISTENCE OF NONCONJUGATIVE PLASMIDS 429
A A
Thanks to the form of the differential equations, this singular point (xl, x,, 0: 0,O) is also a singular point of the full system of equations (2: 1) through (2 :5) : and we want to know if this point is stable or unstable in the larger space.
By inspection of the original set of equations, (2:l) through ( 2 : 5 ) , we see that if all of the partial derivitives are evaluated at the point in question, g2, 0,O. 0) , then, since 2, = x4 = x5 = 0,
A A A
3 f 3 - a f 3 - a f 3 - 3 f 4 - a f 4 - 3 f 4 - a f 5 - 3 f 5 - 3 f 5 - a f 5 -0 . - - _ _ _ _ _ _ - - - _ L _ _ - - - - ~ -
ax, ax, ax, ax, ax, ax3 ax, ax, ax, ax4
Thus, the Jacobian matrix of the system of equations takes the form: * * * * * * *
c I * * * 0 0 af3 0
0 0 0 af4 *
$ ax,
ax5
az3
0 0 0 0 -
where the asterisks indicate values that we do not need to know for the present consideration. W e are concerned only with the new eigenvalues, namely,
*
(2:6)
(2:7)
and - 3 f 5 - - $MD(r) + yff aMD - %' - p , 8 x 5
(2:8)
A A
where. as before. all partial derivatives are evaluated at (z,, z2, 0, 0,O). If any of these partial derivatives are positive, the corresponding population
once introduced would be able to invade the equilibrium population of plasmid- free cells.
The meanings of equations (2:6) through (2:s) are rather straightforward. Those cell types that are capable of transmitting their plasmids, M and MD, could invade if their rate of population growth through cell division and recruit- ment of plasmid-free cells into their population by conjugative transfer exceeds their rate of loss through segregation and flow. The population carrying just the nonconjugative plasmid, D, could invade only i f its rate of growth through cell division at the equilibrium concentration of resource is sufficiently high to over- come its rate of loss through segregation and flow.
(ii) Invasion of a bacterial population where the conjugative plasmid is main- tained: In this case, where g3 '= $2 # 0, there is a stable singular point, (z1,x2,z3)
A A A

430 B. R. LEVIN AND F. M. STEWART
for the three-dimensional system of equations
j i fi (Xi,2%%0,0)
Zz = fz (Xi,ZZJZa,O,O)
and 5 3 f3 ( & J z J ~ ~ O , O ) . However, this case is only slightly more complicated than the preceding two-
dimensional one; now - af5 - y & ? - is no longer zero. The new eigenvalues
introduced when we look at the differential equation system in the whole five- dimensional space as opposed to the subsystem in three-dimensional (r,R,M) subspace, are the eigenvalues of the matrix
a f 4
3f4 1
We now have at (&, A, z3, 070) :
.A 8f5 - $/ (* and -- HD r ) + y R CYMD - .p '- 2~ . ax,
If we think of matrix J as
its characteristic equation is
X z - (a+d) X + ad - bc = 0.
Since bc = yM (YRaD + T) >0, the roots of this equation are necessarily separated hythoseofhz- ( a f d ) i f a d = ( h - a ) ( h - d ) = O .
It follows that there will be a positive eigenvalue if and only if one or more of the following conditions are met:
!bD(;) - y h ? - T - P > o (2:9)
$MD(f-) + (YMD - 2 T-P > 0 (2:lO)
or [ $D (;) - Y M - T-p] [ $MO (>) f y h CYjrD - 2 T - P ] <'/& ( YfiCYDST). (2: 1 1 )
If any of these conditions is met, then the equilibrium with only recipients and

CONDITIONS FOR THE EXISTENCE OF NONCONJUGATIVE PLASMIDS 431
mobilizers present becomes unstable when cells carrying the nonconjugative plasmid are introduced and the latter will be able to invade the habitat.
The meanings of the first two of these invasion conditions for the nonconju- gative plasmid are relatively straightforward: (1 ) donors can invade if their rate of population growth through cell division is sufficiently high to make up for their loss through conversion into mobilized donors by conjugation with mobilizers and their loss by segregation and flow, and/or (2) mobilized donors can invade if the rate of their population growth through cell division and con- version of recipients is sufficiently high to make up for their loss through flow and segregation. A third condition says that even when neither (1) nor (2) obtains, it may be that the recruitment of mobilized donors by converting them first into donors and then into mobilized donors may be sufficient to tip the balance in favor of maintaining the nonconjugative plasmid. With realistic values for the parameters, this condition is delicate, and we expect that ( 2 : l l ) will rarely be fulfilled when (2:9) and (2: I O ) both fail.
Reparameterization to allow for a constant fitness relationship To facilitate our analysis of the conditions under which the invading non-
conjugative plasmid will be maintained in the new equilibrium bacterial popu- lation, we assume a constant relationship for the relative exponential growth rates of the different bacterial types. For this, we use the traditional “selection coefficient” parameter of population genetics (see, for example, CROW and KIMURA 1970). Letting $ ( r ) be the growth rate of the recipient clone at some resource concentration +, the growth rates of the mobilizer, donor and mobilized donor cell types are, respectively, (l-sIcI) $ ( r ) , (l-sD) + ( r ) and (I-sYD) $ ( r ) . The parameters, sM, SD and s X D are the selection coefficients. A cell type with a positive selection coefficient has a selective disadvantage relative to recipients, while a cell line with a negative selection coefficient has a selective advantage relative to recipients.
(i) Invasion of an equilibrium plasmid-free population: At equilibrium the growth rate of the recipients, $(^I), would be equal to the rate at which cells are removed by flow,
$ (4 = p *
Conditions (2:6) through (2:8) for the invasion of the different plasmid-bearing cell types can now be written,
I2
- sD > _I_ P SMD p + 27
and YffMD > R
for mobilizers, donors and mobilized donors, respectively.
(3:l)
(3:2)
(3:3)

432 B. R. LEVIN A N D F. M. STEWART
(ii) Invasion of an equilibrium population of mobilizers and recipients: At equilibrium in a population containing both mobilizers and recipients, the aver- age growth rate of both bacterial populations would be equal to the dilution rate,
$(r) R + $6) (l-sdl) M - - P 7
R + M but unless sy = 0, neither $(;) nor $(;) ( l-sy) will be equal to p. Consequently we can not make the simplifying $(;) = p assumption for this situation. Condi- tions (2:9) through (2:l l) can, nevertheless, be written in terms of the equi- librium growth rate of recipients and relative fitness values,
(3:4)
(3:5)
[$(;) ( i - s D ) - y&f- 7 - p ] [I/(;) (I-sXD) + Y R (YID - 27 - P ] <y&(y&+T) (3:6)
for (2:9), (2:lO) and (2:11), respectively.
Population dynamics To examine the behavior of these populations following the invasion by cells
carrying the nonconjugative plasmid and the fates and relative frequencies of the various cell types, we use a numerical solution to equations (1 :I ) through (1:5) (a computer simulation). In these numerical examples, we assume a
hyperbolic function for the growth rate, $ ( r ) =- pr (MONOD 1949), and use
values of P, Q and e similar to those estimated for E . coli growing in glucose- limited minimal medium (LEVIN, STEWART and CHAO 1977). For the plasmid transfer parameters, we use values in a range similar to those estimated for E . coli and some of its conjugative and nonconjugative plasmids in glucose minimal medium (LEVIN, STEWART and RICE 1979; LEVIN and RICE 1980).
(i) Nonconjugative plasmids in populations where a stable mobilizer-recipient equilibrium cannot be maintained: There are essentially two situations where the nonconjugative plasmid would have to become established in populations free of the mobilizing conjugative factor: (1 ) where mobilizers could maintain a stable equilibrium with recipients, but where, for historical reasons, such con- jugative factors did not exist, and (2) where the conditions were such that a stable mobilizer-recipient equilibrium could not exist because the conjugative plasmid could not become established. From the point of view of general theory, we consider the latter situation to be the more interesting one. In terms of the present notation, the population of potential recipients would not support a
Q+r

CONDITIONS FOR THE EXISTENCE O F NONCONJUGATIVE PLASMIDS 433
stable equilibrium population with mobilizers in the absence of the nonconju-
gative plasmid if condition (3:l) did not obtain, i.e., if y < s~~ + (see
STEWART and LEVIN 1977). The nonconjugative plasmid could then invade this population, consisting of just recipients, only if either condition (3:2) or 3:3) were
' l ID '+" . Since ayD I 1, for condition met, i.e., if ---so > - and/or yayD > (3:3) to be satisfied when condition (3:l) is not, srD would have to be less than sy. For a single host clone, this would occur only if there were some selection favoring the nonconjugative plasmid. This is also necessary for SD to be less than zero and, thus, for condition (3:2) to obtain.
If condition (3:2) obtains when conditions (3: l ) and (3:3) do not, the resulting population would consist of cells carrying the nonconjugative plasmid, and as long as there is a finite rate of vegetative segregation, i.e., T > 0, then cells free of the nonconjugative plasmid would also be maintained. In fact, there would be a stable equilibrium when the rate of recruitment of plasmid-free cells, R, by vegetative segregation from the donors, D, was equal to their rate of decline due to a lower growth rate (fitness). Furthermore, if the intensity of selection favoring the donors is substantial, with realistic rates of vegetative segregation, the vast majority of the population at equilibrium would consist of donors (Figure 1 ) .
Situations where condition (3:3) obtains when neither (3 : l ) nor (3:2) do are presented in Figure 2. When the transfer rate constant, is below the threshold value specified in (3: 3), the nonconjugative plasmid is unable to become estab- lished (Figure 2a) . Above this critical transfer rate, cells carrying the noncon- jugative plasmid increase in frequency, and a stable equilibrium with all four cell populations present results (Figures 2b, c and d) . A similar result obtains when conditions (3: 2) and (3: 3) both occur simultaneously.
(ii) Nonconjugative plasmids in populations where a stable mobilim- recipient equilibrium can be maintained: When condition (3: 1) is met, a stable equilibrium with cells carrying the mobilizing, conjugative plasmid and cells free of that factor would exist (see Figure 3a). There are three conditions under which the nonconjugative plasmid could invade the resulting equilibrium popu- lation of mobilizers and recipients. In the first of these, condition (3:4), the growth rate (reproductive fitness) of the donor population is sufficiently high to overcome their loss through segregation, flow and conversion into mobilized donors by conjugation. In Figure 3b, we illustrate the invasion of the noncon- jugative plasmid by this mechanism. In this example, we considered the worst possible case for invasion by this mechanism: where mobilized donors are unable to increase by cell division and are produced only by conjugation. In this case, when the concentration of donors is sufficiently high, the concentration of both mobilized donors and mobilizers declines (Figure 3c). That is, as a consequence of the fact that the transconjugants (mobilized donors) produced by matings between mobilizers o r mobilized donors and donors are of very low fitness, the
R
7
P R
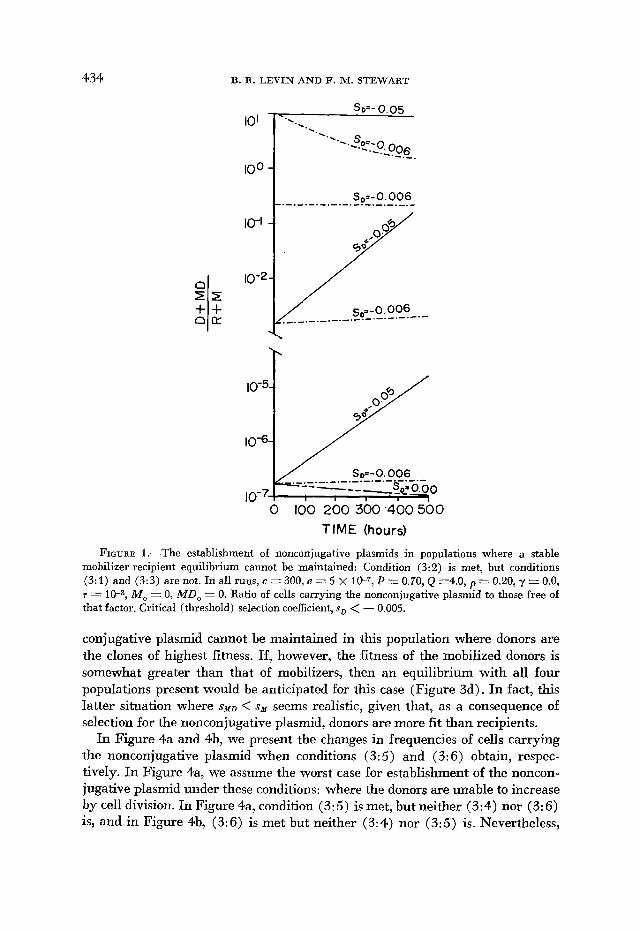
434 B. R. LEVIN AND F. NI. STEWART
a 2 + a
z + [r
10-5-
10-6-
10-7 0 100 200 300,400 500
1°"1 / So=-0. 006
0 100 200 300,400 500 TIME (hours)
FIGURE 1 .-The establishment of nonconjugative plasmids in populations where a stable mobilizer-recipient equilibrium cannot be maintained: Condition (3:2) is met, but conditions (3: 1) and (3:3) are not. In all runs, c = 300, e = 5 X P = 0.70, Q =4.0, p = 0.20, y = 0.0, T = 10-3, MO = 0, M D , = 0. Ratio of cells carrying the nonconjugative plasmid to those free of that factor. Critical (threshold) selection coefficient, sD < - 0.005.
conjugative plasmid cannot be maintained in this population where donors are the clones of highest fitness. If, however, the fitness of the mobilized donors is somewhat greater than that of mobilizers, then an equilibrium with all four populations present would be anticipated for this case (Figure 3d). In fact, this latter situation where s N D < sN seems realistic, given that, as a consequence of selection for the nonconjugative plasmid, donors are more fit than recipients.
In Figure 4a and 4b, we present the changes in frequencies of cells carrying the nonconjugative plasmid when conditions (3: 5 ) and (3: 6) obtain, respec- tively. In Figure 4a, we assume the worst case for establishment of the noncon- jugative plasmid under these conditions: where the donors are unable to increase by cell division. In Figure 4a, condition (3:5) is met, but neither (3:4) nor (3:6) is, and in Figure 4b, (3:6) is met but neither (3:4) nor (3:5) is. Nevertheless,
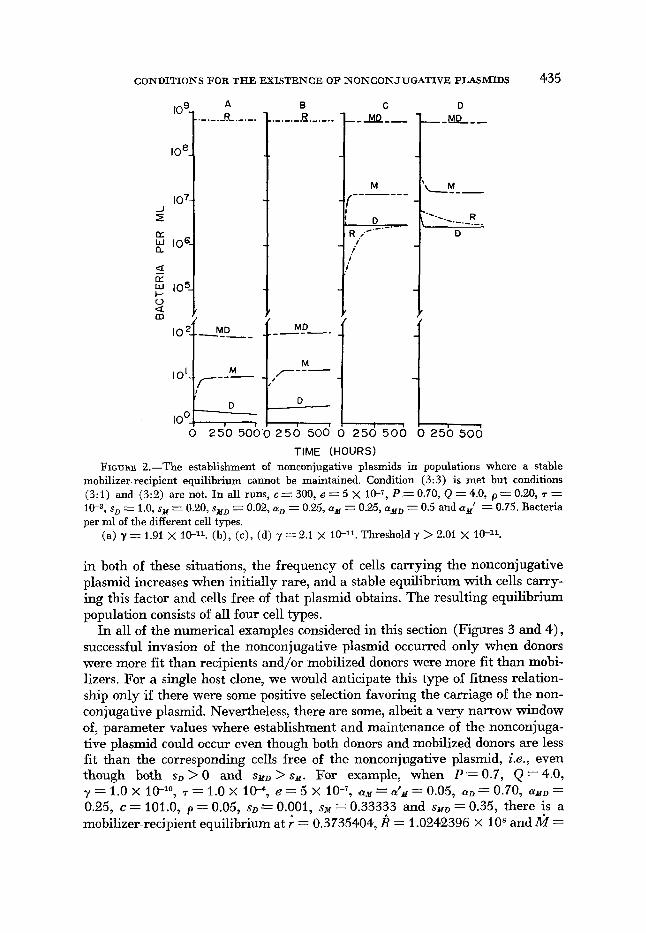
CONDITIONS FOR THE EXISTENCE O F NONCONJUGATIVE PLASMIDS 435
c D
I_ 0 250 500
TIME (HOURS) FIGURE %.-The establishment of nonconjugative plasmids in populations where a stable
mobilizer-recipient equilibrium cannot be maintained. Condition (3:3) is met but conditions (3 : l ) and (3:2) are not. In all runs, c = 300, e = 5 X IO-“, P = 0.70, Q = 4.0, p = 0.20, T = I p , s D - - 1.0, sM = 0.20, sxD = 0.02, LY D - - 0.25, = 0.25, ayD = 0.5 and cyy’ = 0.75. Bacteria per ml of the different cell types.
(a) y = 1.91 x 1 W 1 . (b), (c), (d) y = 2.1 x Threshold y > 2.01 x
in both of these situations, the frequency of cells carrying the nonconjugative plasmid increases when initially rare, and a stable equilibrium with cells carry- ing this factor and cells free of that plasmid obtains. The resulting equilibrium population consists of all four cell types.
In all of the numerical examples considered in this section (Figures 3 and 4), successful invasion of the nonconjugative plasmid occurred only when donors were more fit than recipients and/or mobilized donors were more fit than mobi- lizers. For a single host clone, we would anticipate this type of fitness relation- ship only if there were some positive selection favoring the carriage of the non- conjugative plasmid. Nevertheless, there are some, albeit a very narrow window of, parameter values where establishment and maintenance of the nonconjuga- tive plasmid could occur even though both donors and mobilized donors are less fit than the corresponding cells free of the nonconjugative plasmid, i.e., even though both S D > 0 and s ~ D > s M . For example, when P = 0.7, Q =4.0,
0.25, c = 101.0, p = 0.05, sD = 0.001, sM = 0.33333 and sMD = 0.35, there is a mobilizer-recipient equilibrium at = 0.3735404, €? = 1.0242396 x lo8 and M =
y = I . O X IO-”, T = I . O X 10-4, e = 5 X aM=a’y=0.05, a D ~ 0 . 7 0 , C X M D =
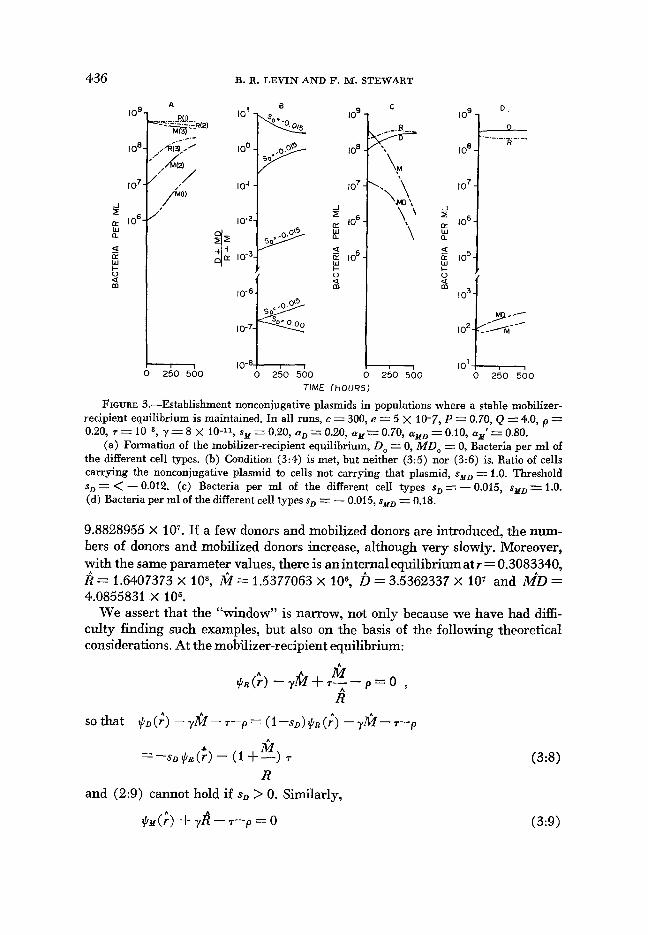
436
lo9
IO0
I o7
2 E IO6 a 5 U W I- U a m
- 0 250 500 0 250 500 0
B. R. LEVIN AND F. M. STEWART
I ,
250 500
I 0-7
lo9
108
lo7 J 2
a a
E IO6
E lo5
W
I- U a m
I06 W a
0 250 500
FIGURE 3.-Establishment nonconjugative plasmids in populations where a stable mobilizer- recipient equilibrium is maintained. In all runs, c = 300, e = 5 x 10-7, P = 0.70, Q = 4.0, p = 0.20, 7 =
(a) Formation of the mobilizer-recipient equilibrium, Do = 0, MD, = 0, Bacteria per ml of the different cell types. (b) Condition (3:4) is met, but neither (3:5) nor (3:6) is. Ratio of cells carrying the nonconjugative plasmid to cells not carrying that plasmid, sYD =Z 1.0. Threshold sD = < - 0.012. (c) Bacteria per ml of the different cell types sD = - 0.015, sMD = 1.0. (d) Bacteria per ml of the different cell types sD = - 0.015, sYD = 0.18.
y = 8 X 10-11, sY = 0.20, aD = 0.20, ay = 0.70, aMD = 0.10, mar'= 0.80.
9.8828955 X lo7. If a few donors and mobilized donors are introduced, the num- bers of donors and mobilized donors increase, although very slowly. Moreover, with the same parameter values, there is an internal equilibrium at r = 0.3083340, i$ = 1.6407373 x lo8, M = 1.5377063 x IO6, b = 3.5362337 X IO7 and M b = 4.0855831 x I O 6 .
We assert that the "window" is narrow, not only because we have had diffi- culty finding such examples, but also on the basis of the following theoretical considerations. At the mobilizer-recipient equilibrium:
A M =-sLl$hC(I*) - (1 +--> T
R and (2:9) cannot hold if SD > 0. Similarly,
$M(;) + - T - p = 0
(3:8)
(3:9)
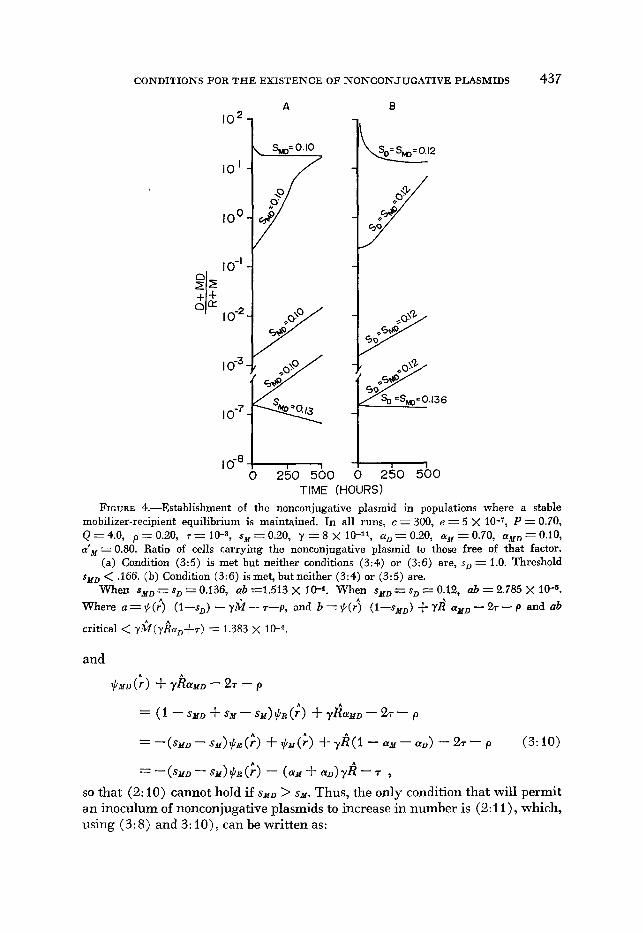
CONDITIONS FOR T H E EXISTENCE O F NONCONJUGATIVE PLASMIDS 437
A
lo* 1 IO'
too
to-'
I o - ~
IO-^
0 250 500 to-*
B
TIME (HOURS) FIGURE 4.-Establishment of the nonconjugative plasmid in populations where a stable
mobilizer-recipient equilibrium is maintained. In all runs, c = 300, e = 5 x IO-', P = 0.70, Q=4.0, p=0.20, r = l O - 3 , ~ ~ ~ 0 . 2 0 , y = 8 x 10-11, a,=0.20, cu,=0.70, cuM,=O.lO, a'= = 0.80. Ratio of cells carrying the nonconjugative plasmid to those free of that factor.
(a) Condition (3:5) is met but neither conditions (3:4) or (3:6) are, so = 1.0. Threshold sllD < .166. (b) Condition (3:6) is met, but neither (3:4) or (3:5) are.
When sMD = s, = 0.136, ab =1.513 x 10-4. When sllD = s, = 0.12, ab = 2.785 X 10-5. Where a = $(;) (I-s,) - y k - T-p, and b = $(d (l-sMD) + yR* aMD - 27 - p and ab
critical < y k ( y k o r , f r ) = 1.383 x IO-*.
and
#MI) <;) + YhCUMD - 27 - p
= (1 - SMJJ + SM - SM) $'E (E.) + y&MD - 27 - p
- ( S M D - sdi) #E (;) f #M (;) + y A ( 1 - @M - @D> 27 * p (3 : 10)
- - - ( sMD-sM)+R(F) - ( a M + a D ~ ) y f i - T 7
so that (2: 10) cannot hold if sMD > sM. Thus, the only condition that will permit an inoculum of nonconjugative plasmids to increase in number is (2:ll) , which, wing ( 3 : 8 ) and 3: l o ) , can be written as:

438 B. R. LEVIN AND F. M. STEWART
[sD+’R(hT) + (1 + ~ ) T ] ’ [ ( s M D - s M ) $ R ( ~ ) + ( a M + a D ) y a f T ] < y h ( y & a D f T ) - R
Clearly, this can occur only when &I and are of comparable size-already a rare situation-aM is small compared to aD, T is small, and S D and S J f D - S Y are very small.
(iii) Invasion of the nonconjugative plasmid in situations where it does not become established: In the cases presented, the conditions for the invasion of the population by the nonconjugative plasmid appear to be sufficient for the con- tinued maintenance of that plasmid in the population, because, in studying the invasion of a population of recipients alone, we tacitly assumed that no equi- librium between recipients and mobilizers was possible. If, however. a stable mobilizer-recipient equilibrium was possible, but did not exist at the time of invasion by the nonconjugative plasmid, the initial increase in the frequency of cells carrying the nonconjugative plasmid need not result in the continued persistence of that factor.
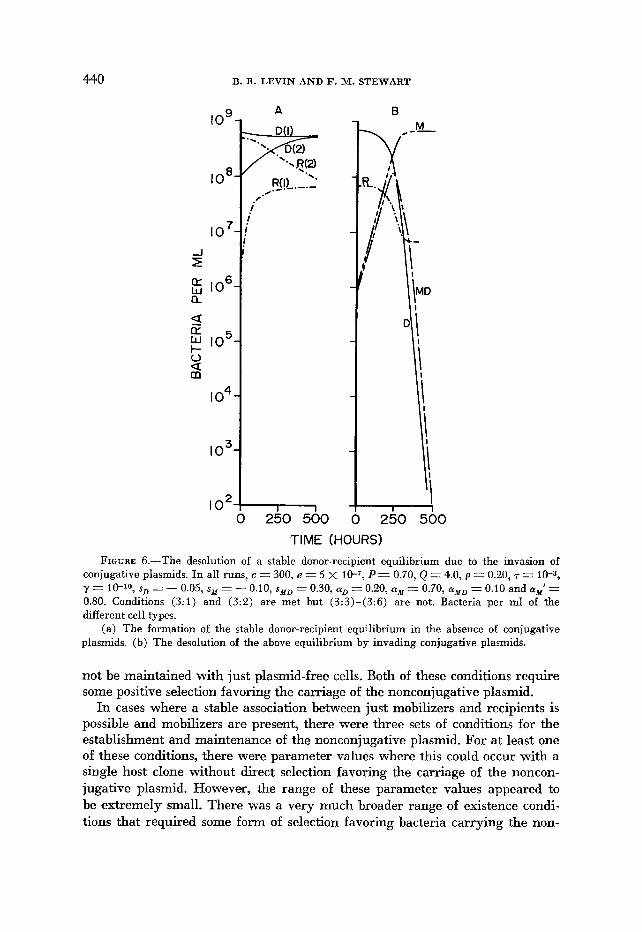
One situation of this type occurs when the donors could invade a population of recipients because of a higher intrinsic growth rate: condition (3:2) , -SO > r/p, but where condition (3: 1) is also met and (3:4) through (3:6) are not. If, in this situation, some mobilizers were present but were, for some reason, at a fre- quency below the equilibrium value at which they could be maintained, then the frequency of cells carrying the nonconjugative plasmid would initially increase. But when the mobilizing cells approach their equilibrium value the frequency of cells carrying this plasmid would decline (Figure 5a). A similar phenomenon occurs when conditions (3:l) and (3:3) are met, but (3:4) through (3:6) are not (Figure 5b). Finally, it is also possible that stable-equilibrium between recipients and donors can be upset by the invasion of conjugative plas- mids, and the nonconjugative plasmid will be eliminated. A situation of this type is presented in Figure 6. Here, conditions (3:l) and (3:2) are met, but conditions (3:3) through (3:6) are not.
DISCUSSION
In our study of the population biology of conjugative plasmids, we concluded that there were “biologically realistic” conditions where, as a consequence of their capacity for infectious transmission, conjugative plasmids could become established and would be maintained in bacterial populations, even in the absence of selection favoring the genes they are carrying (STEWART and LEVIN 1977). At first glance, one might assume that the conditions for the establish- ment and maintenance of mobilizable nonconjugative plasmids in the absence of selection for their genes would be only somewhat more restrictive than those for conjugative factors. For some nonconjugative plasmids, a major portion of the transconjugants of matings between mobilized donors and recipients receives the nonconjugative element. For example, for the nonconjugative plasmid pCRl
CONDITIONS FOR THE EXISTENCE O F NONCONJUGATIVE PLASMIDS 439
I o9
I o8
I o7
6 r’ E I O a 5 L T 3 E I O
3 I o2
I O ‘
IO0
0
250 500 0 250 500 TIME (HOURS)
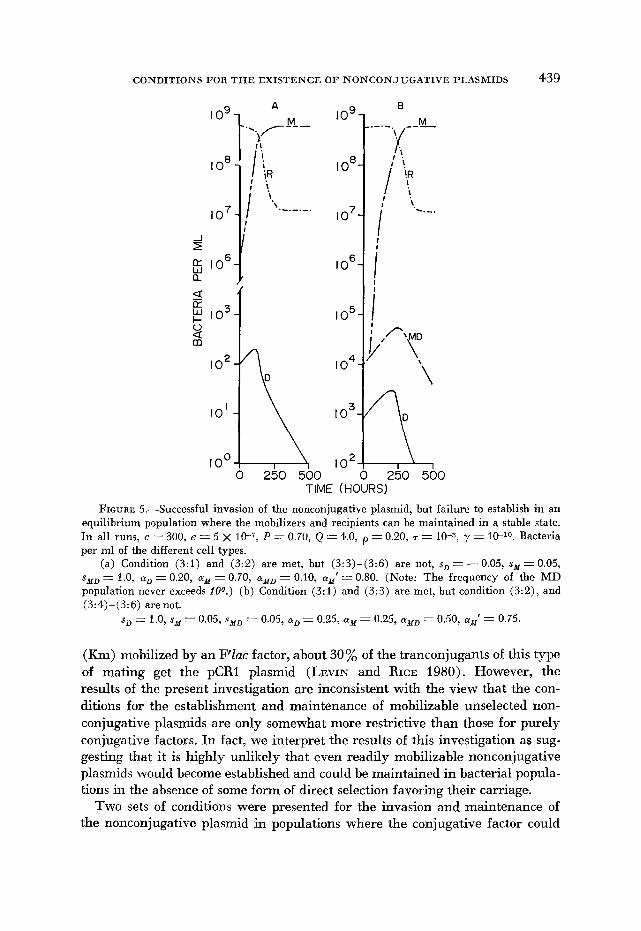
FIGURE 5.--Successful invasion of the nonconjugative plasmid, but failure to establish in an equilibrium population where the mobilizers and recipients can be maintained in a stable state. In all runs, c = 300, e = 5 x 10-7: P = 0.70, Q -- 4.0, p = 0.20, T = 10-3, y = 10-l0. Bacteria per ml of the different cell types.
(a) Condition (3:l) and (3:2) are met, but (3:3)-(3:6) are not, sD = - 0.05, sy = 0.05, syD = 1.0, aD = 0.20, ay = 0.70, cyzID = 0.10, ay’ = 0.80. (Note: The frequency of the MD population never exceeds 100.) (b) Condition (3:l) and (3:3) are met, but condition (3:2), and (3: 4)-( 3 : 6) are not.
sD 1.0, sM = 0.05, syD = 0.05, aD-= 0.25, aM = 0.25, ayD = 0.50, ay’ = 0.75.
(Km) mobilized by an PZac factor, about 30% of the tranconjugants of this type of mating get the pCRl plasmid (LEVIN and RICE 1980). However, the results of the present investigation are inconsistent with the view that the con- ditions for the establishment and maintenance of mobilizable unselected non- conjugative plasmids are only somewhat more restrictive than those for purely conjugative factors. In fact, we interpret the results of this investigation as sug- gesting that it is highly unlikely that even readily mobilizable nonconjugative plasmids would become established and could be maintained in bacterial popula- tions in the absence of some form of direct selection favoring their carriage.
Two sets of conditions were presented for the invasion and maintenance of the nonconjugative plasmid in populations where the conjugative factor could
440 B. R. LEVIN A N D F. M. STEWART
lo9
IO8
lo7
O6
o5
o4
o3
I o2 7 250 500
B
250 500 71ME (HOURS)
FIGURE 6.-The desolution of a stable donor-recipient equilibrium due to the invasion of conjugative plasmids. In all runs, c = 300, e = 5 x 10-7, P = 0.70, Q = 4.0, p = 0.20, r = 10-3, y = 10-~~,s~=-O.05,s,=-O.10,~,~=0.30,0i~=0.20,(~~=0.70,0l~~=0.10and~~~’= 0.80. Conditions (3: l ) and (3:2) are met but (3:3)-(3:6) are not. Bacteria per ml of the different cell types.
(a) The formation of the stable donor-recipient equilibrium in the absence of conjugative plasmids. (b) The desolution of the above equilibrium by invading conjugative plasmids.
not be maintained with just plasmid-free cells. Both of these conditions require some positive selection favoring the carriage of the nonconjugative plasmid.
In cases where a stable association between just mobilizers and recipients is possible and mobilizers are present, there were three sets of conditions for the establishment and maintenance of the nonconjugative plasmid. For at least one of these conditions, there were parameter values where this could OCCLW with a single host clone without direct selection favoring the carriage of the noncon- jugative plasmid. However, the range of these parameter values appeared to be extremely small. There was a very much broader range of existence condi- tions that required some f o r m of selection favoring bacteria carrying the non-

CONDITIONS FOR THE EXlSTENCE O F NONCONJUGATIVE PLASMIDS 4’4’1
conjugative plasmid. Nevertheless, as a consequence of infectious transmission via mobilization, there are conditions where nonconjugative plasmids could become established and would be maintained without the bacteria carrying those factors being the members of the population of highest reproductive fitness.
The validity and generality of the preceding conclusions are, of course, con- tingent upon how realistic and how general this model of the population biology of mobilizable nonconjugative plasmids is. We consider this to be a “mechanis- tic” model in the sense that there is an empirical basis to the form of the relation- ship between resource concentrations and population growth and the form of the habitat is precisely defined (STEWART and LEVIN 1977). The assumptions that in this type of mass-culture habitat plasmid transfer (1) is initiated by the ran- dom collision of the donating and receiving cells, and (2) occurs at a rate that is proportional to the concentrations of cells carrying conjugative factors and those free of those plasmids, are also realistic. On the other hand, the assumptions that plasmid transfer is an instantaneous process governed by a unique and constant rate parameter are inconsistent with known observations. Plasmid transfer is a rather complex process. Measurable amounts of time elapse between the formation of specific pairs and the establishment of the plasmid DNA in the recipient cell (MEYNELL 1973; FALKOW 1975). Furthermore, the rate at which each of the steps of the plasmid transfer process occurs is very much contingent upon the physiological states of the host bacteria (CURTISS et al. 1969). Never- theless, for bacteria dividing at a constant rate in either exponential or chemo- stat equilibrium culture, mass-action models similar to those presented here offer a reasonably good analogue of conjugative plasmid transfer (LEVIN, STEWART and RICE 1979) and mobilization transfer (LEVIN and RICE 1980). The latter study of pCRl (Km) mobilized by F-lac pro in E. coti K12 hosts also offers some empirical justification for the present assumptions that: (1) the relative proportions at which the nonconjugative plasmid, the conjugative plasmid and both plasmids are transmitted in matings between mobilized donors and recipi- ents would be approximately constant, and (2) that the carriage of the noncom jugative plasmid could have little, if any, effect on the donor or recipient ability of a host bacterium.
In spite of the preceding positive statements, we are well aware of the fact that the present model represents a highly reduced and simplistic analogue of the biology of plasmids in real populations. It does not account for the physical het- erogeneity of real environments or for the interactions of bacteria maintained on the surfaces of these natural habitats. Furthermore, the present model does not account for the clonal heterogeneity and changes that would be occurring in natural populations. It is our intuitive feeling that the inclusion of these com- plexities of real environments and plasmid transmission will not alter the quali- tative conclusions drawn here. However, we reserve a strong statement of extra- polation until a more formal consideration of these complexities are considered.
In the DISCUSSION of STEWART and LEVIN (1977), we said (in a way that, in retrospect, appears overly dramatic) that the conclusion that conjugative

I42 B. R. LEVIN A N D F. M. STEWART
plasmids could become established and maintained, even in the face of negative selection, “frightened us.” This conclusion suggested that the more prudent use of antibiotics need not result in a decrease in the frequency of bacteria carrying R factors, and that it would be possible for the (likely to be unselected) chimeric plasmids produced in work with recombinant DNA to become established in natural populations of bacteria. We find the conclusion made in the present study of the existence conditions of nonconjugative plasmids to be more optimistic. These theoretical results most definitely support the position that the more prud- ent use of antibiotics will lead to reduction in the frequency of bacteria carrying R factors. (For an extensive review, see FEINMAN and MATHESON 1978). These results are also consistent with the view that the establishment and maintenance of chimeric plasmids in natural populations would require not only the trans- mission to wild hosts (LEVIN and STEWART 1977), but also some form of selec- tion favoring the carnage of the chimeric plasmid.
We wish to express our gratitude to JON D. GOGUEN, RICHARD K. KASAMA and FRANKLIN L. LERNER for useful commentary on this work. We also wish to thank VIRGINIA A. RICE, NANCY J. SERETTA and DAVID STILWELL for their various contributions to the preparation of the figures and to DEBBIE WILSON for typing and making corrections of this “more mathematical than most” manuscript. We owe a particular debt of gratitude to the reviewers for their perceptive and useful commentary. This investigation was supported by Public Health Service Research Grant GM19848 and Research Career Development Award KO4 GM00112 to B. R. LEVIN.
LITERATURE CITED
CHILTON, M. D., M. H. DRUMMOND, D. J. MERLO, D. SCIAKY, A. L. MONTOYA, M. P. GORDON Stable incorporation of plasmid DNA into higher plant cells.
Early stages of conjuga-
An Introduction io ihe Theory of Population Genetics.
and E. W. NESTER, 1977 The molecular basis of Crown Gall tumorigenesis. Cell 11: 263-271.
tion inE. coli. J. Bacteriol. 100: 1091-1104. CURTIS, R., 111, L. G. CARO, D. P. ALLISON and D. R. STALLIONS, 1969
CROW, J. F. and M. KIMURA, 1970 Harper Row, New York.
FALKOW, S., 1975
FEINMAN, S. E. and J. C. MATHESON 111, 1978
Infeciious Multiple Drug Resistance. Pion, London.
Draft Environmental Inpact Statement: Sub- therapeutic antibacterial agents in animal feeds. Prepared by the Bureau of Veterinary Medi- cine Food and Drug Administration.
LEVIN, B. R. and V. A. RICE, 1980 The kinetics of transfer of nonconjugative plasmids by mobilizing conjugative factors. Genet. Res. (in press).
LEVIN, B. R. and F. M. STEWART, 1977 Probability of establishing chimeric plasmids in natural populations of bacteria. Science 1%: 218-22G.
LEVIN, B. R., F. M. STEWART and L. CHAO, 1977 Resource limited growth, competition and pre- dation. A model and some experimental studies with bacteria and bacteriophage. Amer. Natur. 111: 3-24.
The kinetics of conjugative plasmid transmis-
Bacierial Plasmids. Massachusetts Institute of Technology Press, Cam-
LEVIN, B. R., F. M. STEWART and V. A. RICE, 1979 sion: Fit of a simple mass action model. Plasmid 2: 247-260.
MEYNELL, G. G. 1973 bridge, Mass.
MONOD, J., 1949 The growth of bacterial cultures. Ann. Review Microb. 3: 371-394.

CONDITIONS FOR T H E EXISTENCE OF NONCONJUGATIVE PLASMIDS 443 NOVICK, R. P. and D. BOUANCHAND, 1971 Extrachromosomal nature of drug resistance in
NOVICK, R. P., R. C. CLOWES, S. N. COHN, R. C. CURTISS 111, N. DATTA and S. FALKOW, 1976
NIH, 1976 REINWALD, J. G., A. M. CHAKRABARTY and I. C. GUNSALVES, 1973 A transmissible plasmid con-
trolling camphor oxidation in Pseudomonas putida. Proc. Natl. Acad. Sci. U.S. 70: 885-889. SKERMAN, F. J., S. B. FORMAL and S. FALKOW, 1972 Plasmid associated enterotoxin production
on a strain of Escherichia coli isolated from humans. Infect. and Immunity 5 : 622-624. STEWART, F. M. and B. R. LEVIN, 1977 The population biology of bacterial plasmids: a priori
conditions for the existence of conjugationally transmitted factors. Genetics 87 : 209-228. WATANABLE, T., 1963 Infective heredity of multiple drug resistance in bacteria. Bacter. Rev.
27: 87-115. Corresponding editor: H. ECHOLS
Staphylococcus aureaus. Ann. N.Y. Acad. Sci. 182: 279-294.
Uniform nomenclature for bacterial plasmids: A proposal. Bacter. Rev. 40: 168-1 89. Guidelines for Recombinant DNA Research. June 23,1978.





























