Embed Size (px)
Citation preview

Brain Research 868 (2000) 268–274www.elsevier.com/ locate /bres
Research report
Opposite effects of glutamate antagonists and antiparkinsonian drugson the activities of DOPA decarboxylase and 5-HTP decarboxylase in
the rat brain*Andrew Fisher, Michael S. Starr
Department of Pharmacology, The School of Pharmacy, 29 –39 Brunswick Square, London WC1N 1AX, UK
Accepted 28 March 2000
Abstract
This study measured the activities of L-DOPA and 5-HTP decarboxylase (DDC and 5-HTPDC) in the substantia nigra and corpusstriatum of reserpine-treated rats. Acute injection of the NMDA receptor antagonists CGP 40116 (5 mg/kg) and HA 966 (5 mg/kg), andto a lesser extent eliprodil (10 mg/kg), greatly elevated DDC in both structures, whilst having no effect on (nigra) or inhibiting (striatum)5-HTPDC. L-DOPA (25 mg/kg) on its own inhibited both enzymes in either brain region. The weak NMDA receptor-channel blockers(and antiparkinsonian drugs) budipine (10 mg/kg), memantine (40 mg/kg) and amantadine (40 mg/kg) strongly increased DDC, whilstnot affecting or decreasing 5-HTPDC activity in nigra and striatum. The L-DOPA-induced suppression of DDC was mostly reversed by allthree antiparkinsonian drugs, whilst L-DOPA-induced inhibition of 5-HTPDC was only reversed by CGP 40116 (striatum only). It isconcluded that glutamate exerts a differential physiological influence on the biosynthesis of dopamine and 5-HT in the brain, by tonicallysuppressing DDC and tonically stimulating 5-HTPDC. The L-DOPA-induced reduction in DDC may help to explain the eventual loss ofefficacy of L-DOPA therapy in parkinsonian patients. It is suggested, however, that it may be possible to extend the lifetime of L-DOPAtherapy with drugs which potentiate the activity of DDC, such as budipine and the 1-aminoadamantanes. 2000 Elsevier Science B.V.All rights reserved.
Keywords: Aromatic L-amino-acid decarboxylase; L-3,4-Dihydroxyphenylalanine; L-5-Hydroxytryptophan; Reserpine; Budipine; Amantadine; Memantine;NMDA antagonism
1. Introduction they are also differentially distributed in brain regions andin subcellular tissue fractions [5,31,33]. To accommodate
The first step in the parallel biosyntheses of dopamine these seemingly discordant molecular biological and bio-and 5-hydroxytryptamine (5-HT) is catalysed by a sub- chemical data, it has been proposed that AADC may be astrate-specific hydroxylase, but the ensuing decarboxyl- single enzyme with separate recognition and regulatoryation reaction appears to share the same enzyme, namely sites for L-DOPA and 5-HTP [5,6].aromatic L-amino-acid decarboxylase (AADC; E.C. Until recently DOPA decarboxylase (hereafter referred4.1.1.28). Although it has been claimed that a single gene to as DDC) was regarded as an unsaturated, unregulatedcodes for AADC [1], potentially different isoforms of and hence uninteresting enzyme. However, this opinionAADC may exist [29,40]. However, as yet there has been has radically altered with the discovery that the activity ofno satisfactory explanation for the many biochemical DDC is subject to both pre- and post-translational regula-properties which clearly differentiate between AADC tion [6,14,20,41]. The ability to increase the activity ofwhen L-3,4-dihydroxyphenylalanine (L-DOPA) and L-5-hy- DDC could be clinically important in the treatment ofdroxytryptophan (5-HTP) are used as substrates [6]. Thus Parkinson’s disease, where the disappearance of DDCnot only do the two enzymes exhibit different optima with caused by the degeneration of nigrostriatal dopamineregard to pH, temperature and substrate concentration, but neurones could contribute to the eventual loss of effective-
ness of dopamine replacement therapy with L-DOPA[6,30]. It may therefore be possible to prevent DDC*Corresponding author.
E-mail address: [email protected] (M.S. Starr) becoming rate-limiting in parkinsonism by coadministering
0006-8993/00/$ – see front matter 2000 Elsevier Science B.V. All rights reserved.PI I : S0006-8993( 00 )02339-8

A. Fisher, M.S. Starr / Brain Research 868 (2000) 268 –274 269
drugs which maintain or even increase the activity of were first injected intraperitoneally (i.p.) with reserpine (5residual DDC in the basal ganglia. Hadjiconstantinou et al. mg/kg). The following day (18–20 h later) they received[20] demonstrated that dopamine receptor antagonists drugs as indicated in the text, either alone or together withmodestly increased DDC activity in mice, but the use of L-DOPA (25 mg/kg, after giving benserazide, 50 mg/kg,such drugs in dopamine-deficient parkinsonian patients 30 min earlier), or with saline 1 ml /kg, controls). All drugwould exacerbate their condition. On the other hand, we doses were based on previous studies. One hour later theand others have found that various N-methyl-D-aspartate animals were sacrificed by guillotining, their brains re-(NMDA) receptor antagonists dramatically stimulated moved quickly into ice-cold saline and the substantia nigraDDC in the nigrostriatal dopamine pathway of rats, and corpus striatum dissected out.suggesting the adjunctive use of these compounds mayimprove pharmacotherapy with L-DOPA in Parkinson’s 2.3. Assay of DOPA and 5-HTP decarboxylasedisease [7,15,16,19].
L-DOPA is not only decarboxylated by DDC in dopa- The assay has been described in detail elsewhere [16]minergic neurones, but also by 5-HTP decarboxylase and is based on that described by Hadjiconstantinou et al.(hereafter distinguished as 5-HTPDC) in 5-HTergic neuro- [20]. Tissue samples were homogenized in 0.25 M sucrosenes [3,39]. Since 5-HT neurones sprout to occupy synapses solution (1 ml for nigra, 3 ml for striatum) and thevacated by dopamine axons in the dopamine-lesioned brain homogenates centrifuged in Eppendorf tubes at 3000 rpm[35], it is distinctly possible that 5-HT systems in the for 10 min. A 100-ml volume of the supernatant wasparkinsonian brain are critically involved in ‘dopamine assayed for protein by the Bradford method [13], while 20replacement’ with L-DOPA. ml were assayed for DDC and 5-HTPDC as follows. An
During the course of studies into the effects of glutamate incubation mixture (total volume 400 ml) was formed ofantagonists on the decarboxylation of L-DOPA by the the following: sodium phosphate buffer (pH 7.2); eithernigrostriatal dopamine system in the reserpine-treated rat, L-DOPA (0.5 mM) or 5-HTP (0.5 mM); ascorbic acid (0.1we noticed that 5-HTPDC in this brain region responded mM); pyridoxal-59-phosphate (0.01 mM); 2-mercap-differently from DDC to various NMDA antagonists. This toethanol (1 mM); EDTA (0.1 mM); pargyline (0.1 mM);paper will show that glutamate blockade strongly poten- enzyme (20 ml). Blank values were determined by sub-tiates DDC but not 5-HTPDC, suggesting the two forms of stituting water for L-DOPA/5-HTP substrate. Incubationthe decarboxylase are differentially regulated by glutamate. was carried out for 10 min at 378C and was stopped by the
addition of ice-cold 80 ml perchloric acid (0.5 M). Themixture was then centrifuged at 3000 rpm for 10 min and
2. Materials and methods the resulting supernatant assayed by high-performanceliquid chromatography (HPLC). Full details of the HPLC
2.1. Materials assay have been given elsewhere [9]. Enzyme activity wasexpressed as nmol dopamine or 5-HT/mg protein /h. Sims
Methanol (Rathburn, Scotland) and other chromato- et al. [33] reported that the pH optimum for the 5-HTPDCgraphic materials (Fluka, UK) were of the highest grade. assay was 8.3, but in our hands the rate of decarboxylationReserpine, a-methyl-p-tyrosine hydrochloride, L-DOPA, 5- of 5-HTP was not significantly increased at this pHHTP, benserazide hydrochloride, amantadine hydrochlo- (unpublished observations).ride, pargyline hydrochloride and general chemicals usedfor the enzyme assays were obtained from Sigma (UK) 2.4. Data analysis(RS)-3-amino-1-hydroxypyrrolidin-2-one (HA 966) was
´obtained from Tocris (UK). Eliprodil (Synthelabo Re- Results were calculated as means6S.E.M. for at leastcherche, Bagneux), R-DL-(E)-2-amino-5-phosphonopen- six separate determinations. DDC and 5-HTPDC activitiestanoic acid (CGP 40116; Ciba Geigy, Basel, Switzerland), in drug- and saline-treated animals were compared bybudipine (Byk Gulden, Konstanz, Germany) and meman- Analysis of variance and post hoc by Dunnett’s t-test.tine (Merz, Frankfurt, Germany) were generously donated. Significance was assumed at P,0.05.
2.2. Animals and treatments3. Results
Male Wistar albino rats (A.R. Tuck) weighing 200–260g were used for these experiments. All experiments were 3.1. Effects of reserpine treatment on DDC and 5-conducted in accordance with the Animals (Scientific HTPDCProcedures) Act 1986 and with local Ethical Committeeguidelines. Animals were initially housed in groups of six One day after receiving reserpine, 5 mg/kg i.p., ratsat 22618C, under fluorescent lighting from 07.00 to 17.00 were highly sedated and completely akinetic. The basalh, and allowed free access to rat chow and water. Animals activities of DDC were 70.163.1 and 80.264.5 nmol
270 A. Fisher, M.S. Starr / Brain Research 868 (2000) 268 –274
dopamine /mg protein /h for nigra and striatum, respective-ly. Corresponding basal values for 5-HTPDC were30.565.0 and 28.361.6 nmol 5-HT/mg protein /h fornigra and striatum, respectively. The greater preponderanceof DDC over 5-HTPDC activity in these brain regionsaccords with earlier findings [31,33].
3.2. Effects of NMDA receptor antagonists on DDC and5-HTPDC
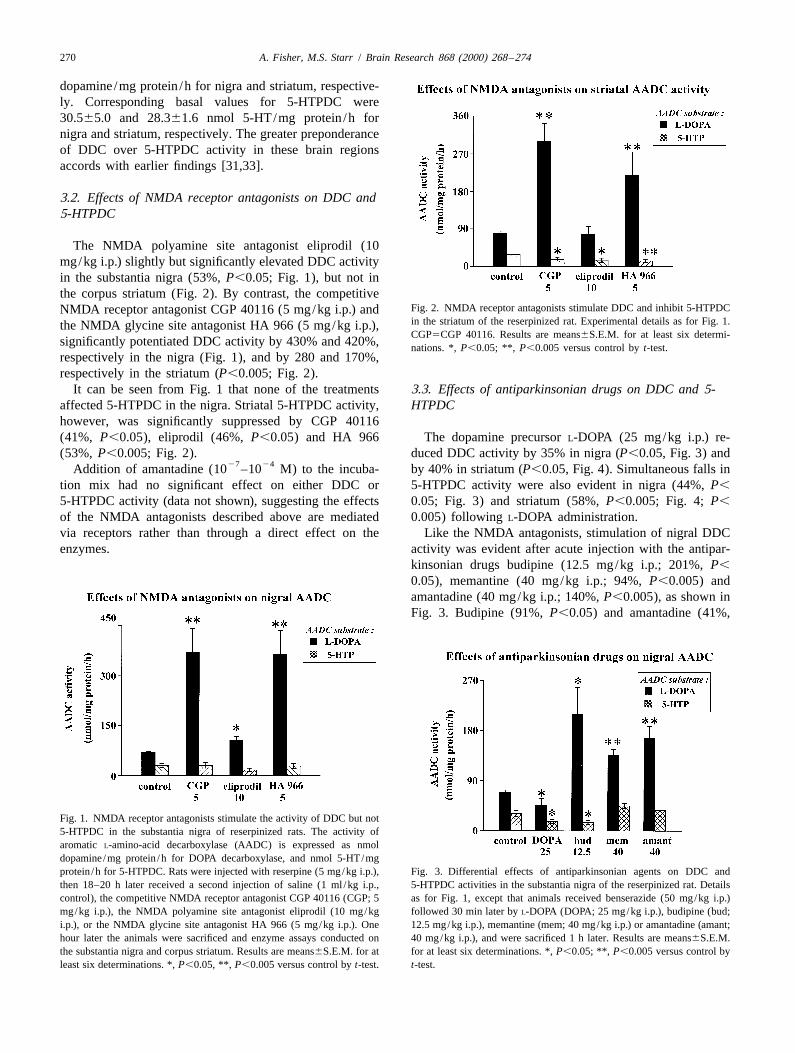
The NMDA polyamine site antagonist eliprodil (10mg/kg i.p.) slightly but significantly elevated DDC activityin the substantia nigra (53%, P,0.05; Fig. 1), but not inthe corpus striatum (Fig. 2). By contrast, the competitive
Fig. 2. NMDA receptor antagonists stimulate DDC and inhibit 5-HTPDCNMDA receptor antagonist CGP 40116 (5 mg/kg i.p.) andin the striatum of the reserpinized rat. Experimental details as for Fig. 1.the NMDA glycine site antagonist HA 966 (5 mg/kg i.p.),CGP5CGP 40116. Results are means6S.E.M. for at least six determi-
significantly potentiated DDC activity by 430% and 420%, nations. *, P,0.05; **, P,0.005 versus control by t-test.respectively in the nigra (Fig. 1), and by 280 and 170%,respectively in the striatum (P,0.005; Fig. 2).
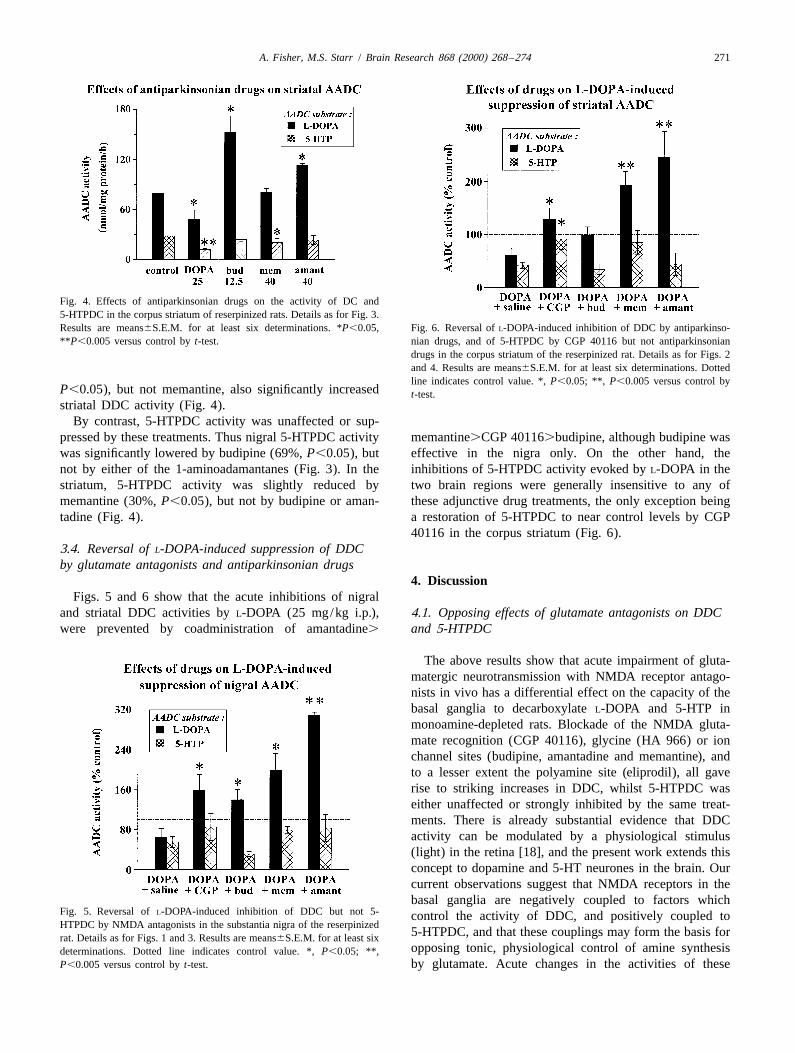
It can be seen from Fig. 1 that none of the treatments 3.3. Effects of antiparkinsonian drugs on DDC and 5-affected 5-HTPDC in the nigra. Striatal 5-HTPDC activity, HTPDChowever, was significantly suppressed by CGP 40116(41%, P,0.05), eliprodil (46%, P,0.05) and HA 966 The dopamine precursor L-DOPA (25 mg/kg i.p.) re-(53%, P,0.005; Fig. 2). duced DDC activity by 35% in nigra (P,0.05, Fig. 3) and
27 24Addition of amantadine (10 –10 M) to the incuba- by 40% in striatum (P,0.05, Fig. 4). Simultaneous falls intion mix had no significant effect on either DDC or 5-HTPDC activity were also evident in nigra (44%, P,
5-HTPDC activity (data not shown), suggesting the effects 0.05; Fig. 3) and striatum (58%, P,0.005; Fig. 4; P,
of the NMDA antagonists described above are mediated 0.005) following L-DOPA administration.via receptors rather than through a direct effect on the Like the NMDA antagonists, stimulation of nigral DDCenzymes. activity was evident after acute injection with the antipar-
kinsonian drugs budipine (12.5 mg/kg i.p.; 201%, P,
0.05), memantine (40 mg/kg i.p.; 94%, P,0.005) andamantadine (40 mg/kg i.p.; 140%, P,0.005), as shown inFig. 3. Budipine (91%, P,0.05) and amantadine (41%,
Fig. 1. NMDA receptor antagonists stimulate the activity of DDC but not5-HTPDC in the substantia nigra of reserpinized rats. The activity ofaromatic L-amino-acid decarboxylase (AADC) is expressed as nmoldopamine /mg protein /h for DOPA decarboxylase, and nmol 5-HT/mgprotein /h for 5-HTPDC. Rats were injected with reserpine (5 mg/kg i.p.), Fig. 3. Differential effects of antiparkinsonian agents on DDC andthen 18–20 h later received a second injection of saline (1 ml /kg i.p., 5-HTPDC activities in the substantia nigra of the reserpinized rat. Detailscontrol), the competitive NMDA receptor antagonist CGP 40116 (CGP; 5 as for Fig. 1, except that animals received benserazide (50 mg/kg i.p.)mg/kg i.p.), the NMDA polyamine site antagonist eliprodil (10 mg/kg followed 30 min later by L-DOPA (DOPA; 25 mg/kg i.p.), budipine (bud;i.p.), or the NMDA glycine site antagonist HA 966 (5 mg/kg i.p.). One 12.5 mg/kg i.p.), memantine (mem; 40 mg/kg i.p.) or amantadine (amant;hour later the animals were sacrificed and enzyme assays conducted on 40 mg/kg i.p.), and were sacrificed 1 h later. Results are means6S.E.M.the substantia nigra and corpus striatum. Results are means6S.E.M. for at for at least six determinations. *, P,0.05; **, P,0.005 versus control byleast six determinations. *, P,0.05, **, P,0.005 versus control by t-test. t-test.
A. Fisher, M.S. Starr / Brain Research 868 (2000) 268 –274 271
Fig. 4. Effects of antiparkinsonian drugs on the activity of DC and5-HTPDC in the corpus striatum of reserpinized rats. Details as for Fig. 3.Results are means6S.E.M. for at least six determinations. *P,0.05, Fig. 6. Reversal of L-DOPA-induced inhibition of DDC by antiparkinso-**P,0.005 versus control by t-test. nian drugs, and of 5-HTPDC by CGP 40116 but not antiparkinsonian
drugs in the corpus striatum of the reserpinized rat. Details as for Figs. 2and 4. Results are means6S.E.M. for at least six determinations. Dottedline indicates control value. *, P,0.05; **, P,0.005 versus control by
P,0.05), but not memantine, also significantly increased t-test.striatal DDC activity (Fig. 4).
By contrast, 5-HTPDC activity was unaffected or sup-pressed by these treatments. Thus nigral 5-HTPDC activity memantine.CGP 40116.budipine, although budipine waswas significantly lowered by budipine (69%, P,0.05), but effective in the nigra only. On the other hand, thenot by either of the 1-aminoadamantanes (Fig. 3). In the inhibitions of 5-HTPDC activity evoked by L-DOPA in thestriatum, 5-HTPDC activity was slightly reduced by two brain regions were generally insensitive to any ofmemantine (30%, P,0.05), but not by budipine or aman- these adjunctive drug treatments, the only exception beingtadine (Fig. 4). a restoration of 5-HTPDC to near control levels by CGP
40116 in the corpus striatum (Fig. 6).3.4. Reversal of L-DOPA-induced suppression of DDCby glutamate antagonists and antiparkinsonian drugs
4. DiscussionFigs. 5 and 6 show that the acute inhibitions of nigral
and striatal DDC activities by L-DOPA (25 mg/kg i.p.), 4.1. Opposing effects of glutamate antagonists on DDCwere prevented by coadministration of amantadine. and 5-HTPDC
The above results show that acute impairment of gluta-matergic neurotransmission with NMDA receptor antago-nists in vivo has a differential effect on the capacity of thebasal ganglia to decarboxylate L-DOPA and 5-HTP inmonoamine-depleted rats. Blockade of the NMDA gluta-mate recognition (CGP 40116), glycine (HA 966) or ionchannel sites (budipine, amantadine and memantine), andto a lesser extent the polyamine site (eliprodil), all gaverise to striking increases in DDC, whilst 5-HTPDC waseither unaffected or strongly inhibited by the same treat-ments. There is already substantial evidence that DDCactivity can be modulated by a physiological stimulus(light) in the retina [18], and the present work extends thisconcept to dopamine and 5-HT neurones in the brain. Ourcurrent observations suggest that NMDA receptors in thebasal ganglia are negatively coupled to factors which
Fig. 5. Reversal of L-DOPA-induced inhibition of DDC but not 5- control the activity of DDC, and positively coupled toHTPDC by NMDA antagonists in the substantia nigra of the reserpinized
5-HTPDC, and that these couplings may form the basis forrat. Details as for Figs. 1 and 3. Results are means6S.E.M. for at least sixopposing tonic, physiological control of amine synthesisdeterminations. Dotted line indicates control value. *, P,0.05; **,
P,0.005 versus control by t-test. by glutamate. Acute changes in the activities of these

272 A. Fisher, M.S. Starr / Brain Research 868 (2000) 268 –274
enzymes could involve an alteration in their phosphoryla- explain the ability of CGP 40116 and HA 966 to facilitatetion status [28]. DDC in reserpine-treated and not intact rats, and why these
Not only can we conclude from these experiments that compounds greatly potentiated the release of dopamineDDC and 5-HTPDC are differentially regulated by gluta- newly formed from L-DOPA in dialysis experiments inmate, but our findings also suggest that DDC and 5- reserpine-treated rats [8,10]. However, it does not explainHTPDC probably function as separate entities. This is why the reverse was true for amantadine and memantine,because the homogenisation of the brain destroys any which are also NMDA antagonists [24], yet potentiatednatural compartmentalisation of the tissue and produces a DDC to a lesser extent in reserpine-treated as compared tomixture in which the two enzymes are equally exposed to normal rats [16]. We do not yet have sufficient informationadded substrate, whether this be L-DOPA or 5-HTP. Thus, on 5-HTPDC in normal animals to say whether theif DDC were capable of utilising both L-DOPA and 5-HTP reserpine status of the animal similarly alters the responseas substrates, then any drug-induced increases in DDC of this enzyme to glutamate antagonists.activity should not only hasten the biotransformation ofL-DOPA, but also that of 5-HTP as well. Clearly this didnot happen, which leads us to conclude that 5-HTP was not 4.3. Reversal of L-DOPA-induced suppression of DDCa substrate for DDC and conversely that L-DOPA was not a — implications for Parkinson’s diseasesubstrate for 5-HTPDC, at least under the in vitro incuba-tion conditions employed here. Using immunohistochemis- We have shown here that systemically-administered L-
¨try, Hokfelt et al. [21] reached a different conclusion, DOPA not only inhibited its own decarboxylation, but thatwhich is that DDC and 5-HTPDC are immunologically of 5-HTP as well. Acutely, L-DOPA is believed to interfereindistinguishable and can utilise each others’ substrates. with the action of DDC by combining with the coenzymeWhatever the reason for the discrepancy in these ex- pyridoxal-59-phosphate to form an inhibitory tetrahydro-perimental data, there is nevertheless compelling evidence isoquinoline complex [32,38]. If this is the case, then it isthat L-DOPA loading in vivo can increase dopamine not surprising that L-DOPA also impaired 5-HTPDCfluorescence in 5-HT neurones [3,39], and that 5-HTP activity, since this utilizes the same coenzyme. Chronicadministered in large doses can similarly be taken up and treatment with L-DOPA has also been reported to suppressconverted to 5-HT in catecholaminergic neurones [4]. AADC [37], although continued exposure to the aminoUnder exceptional conditions, therefore, the sources of acid could involve an action at the pre-translational stagethese biogenic amines in the brain become more diffuse to suppress enzyme induction (e.g. [20,41]).and extend beyond their natural neuronal boundaries. In either event, a diminution in the capacity of the basal
ganglia to decarboxylate L-DOPA could have far-reaching4.2. Effects of reserpine on DDC and 5-HTPDC implications for dopamine replacement therapy with L-
DOPA in Parkinson’s disease, which is dependent on DDCReserpine treatment itself elicited a modest increase in for dopamine synthesis. This is because the levels of DDC
DDC activity in the brains of mice [20] and rats [7], most in the human brain are low to begin with [12] and areprobably as the result of depleting brain dopamine and further reduced by degeneration of the dopamine neuronesremoving feedback inhibition of the enzyme via dopamine [17,26,27]. Consequently any additional burden that isautoreceptors [14,20,41]. By contrast, reserpine had a more placed on DDC activity by exogenously-administered L-profound effect on the ability of glutamate antagonists to DOPA, could mean that a critical point is reached wherestimulate DDC. In a previous study with intact rats, we the activity of the enzyme and not the availability of thefound that eliprodil and CGP 40116 did not affect DDC in substrate now determines the rate of dopamine synthesis. Itthe substantia nigra or corpus striatum, and that HA 966 is distinctly possible, therefore, that the motor complica-was only marginally effective at stimulating DDC, and tions which beset L-DOPA in advanced stages of parkin-then only in the striatum [16]. Here, however, CGP 40116 sonism are self-inflicted, and are due to an insufficiency ofand HA 966 greatly enhanced DDC activity in both of dopamine synthesis by DDC brought about by the treat-these brain regions, while a smaller increase in DDC (this ment itself.time in nigra only) was evident with eliprodil. Following We have shown above, however, that it may be possibledegeneration of the nigrostriatal dopamine pathway, or to circumvent this problematic situation by supplementingafter depletion of dopamine stores by reserpine, there are L-DOPA with an agent that enhances DDC activity, such assecondary increases in glutamate release in the striatum a glutamate antagonist. In the reserpine model, the ability[2,25,36,40] and in striatal efferent pathways [9,36]. This of these compounds to potentiate the enzyme was evident-increase in excitatory neurotransmission could therefore ly much greater than the capacity of L-DOPA to inhibit it,exercise a greater glutamate-mediated suppression of DDC so that when the two were combined the result was a netin the nigra and striatum, such that disinhibition by certain gain in dopamine synthesis. The weak NMDA receptor-ionNMDA antagonists is now able to evoke a rebound channel antagonists budipine, memantine and amantadineincrease in DDC activity. Such a mechanism would were particularly effective in this respect, which might

A. Fisher, M.S. Starr / Brain Research 868 (2000) 268 –274 273
D receptors via the indirect striatal output pathway?, Brain Res.account for their antiparkinsonian efficacy when adminis- 2
753 (1997) 163–175.tered in conjunction with L-DOPA [22,23,34].[10] C.S. Biggs, M.S. Starr, Microdialysis study of the effects of the
The exact whereabouts of the site(s) of L-DOPA decarb- antiparkinsonian drug budipine on L-DOPA-induced release ofoxylation in the brains of parkinsonian patients remains dopamine and 5-hydroxytryptamine by rat substantia nigra andsomething of a mystery. The dopaminergic neurones corpus striatum, Synapse 33 (1999) 1–12.
¨[11] B. Bjelke, I. Stromberg, W.T. O’Connor, B. Andbjer, L.F. Agnati, K.themselves obviously make a major contribution, but thisFuxe, Evidence for volume transmission in the dopamine denervatedis expected to decrease as the neurones degenerate. Animalneostriatum of the rat after a unilateral nigral 6-OHDA microinjec-studies suggest that a fraction of exogenously-administeredtion. Studies with systemic D-amphetamine treatment, Brain Res.
L-DOPA is also taken up and converted to dopamine by 662 (1994) 11–24.5-HTPDC in 5-HTergic neurones [3,39]. Upon release this [12] R.R. Bowsher, D.P. Henry, Aromatic L-amino-acid decarboxylase
biochemistry and functional significance, in: A.A. Boulton, G.B.dopamine is then assumed to be capable of diffusing largeBaker, P.H. Yu (Eds.), Neuromethods, Vol. Series I: Neurochemistry,distances through the extracellular milieu in order to reachNeurotransmitter Enzymes, Humana Press, New Jersey, 1986, pp.and stimulate its target receptors [11]. The promiscuous33–77.
synthesis and release of dopamine by 5-HTergic elements [13] M.M. Bradford, A rapid and sensitive method for the quantitation ofin the parkinsonian brain is therefore an attractive hypoth- microgram quantities of protein utilizing the principle of protein–
dye binding, Anal. Biochem. 72 (1976) 248–254.esis, although its contribution may be lessened in the[14] S. Cho, N.H. Neff, M. Hadjiconstantinou, Regulation of tyrosinepresence of an NMDA antagonist that promotes the
hydroxylase and aromatic L-amino-acid decarboxylase by dopa-activity of DDC and not 5-HTPDC.minergic drugs, Eur. J. Pharmacol. 323 (1997) 149–157.
[15] A. Fisher, C.S. Biggs, M.S. Starr, Effects of glutamate antagonistson the activity of aromatic L-amino-acid decarboxylase, AminoAcids 14 (1998) 43–49.Acknowledgements
[16] A. Fisher, C.S. Biggs, M.S. Starr, Differential effects of NMDA andnon-NMDA antagonists on the activity of aromatic L-amino-acidWe are grateful for financial support from The Sir Julesdecarboxylase activity in the nigrostriatal dopamine pathway of the
Thorn Charitable Trust and the Medical Research Council, rat, Brain Res. 792 (1998) 126–132.and for gifts of drugs to Byk Gulden (budipine) and Merz ´[17] A. Gjedde, G.C. Leger, P. Cumming, Y. Yasuhara, A. Evans, M.(memantine). Guttman, H. Kuwabara, Striatal L-DOPA decarboxylase activity in
Parkinson’s disease in vivo: implications for the regulation ofdopamine synthesis, J. Neurochem. 61 (1993) 1538–1541.
[18] M. Hadjiconstantinou, Z.L. Rossetti, C. Silvia, D. Krajnc, N.H. Neff,References Aromatic L-amino-acid decarboxylase activity of the rat retina is
modulated in vivo by environmental light, J. Neurochem. 51 (1988)[1] V.R. Albert, J. Allen, T. Joh, A single gene codes for aromatic 1560–1564.
L-amino-acid decarboxylase in both neuronal and non-neuronal [19] M. Hadjiconstantinou, Z.L. Rossetti, T.A. Wemlinger, N.H. Neff,tissues, J. Biol. Chem. 262 (1987) 9404–9411. Dizocilpine enhances striatal tyrosine hydroxylase and aromatic
[2] R.L. Albin, A.B. Young, J.B. Penney, The functional anatomy of L-amino-acid decarboxylase activity, Eur. J. Pharmacol. [Mol.basal ganglia disorders, Trends Neurosci. 12 (1989) 366–375. Pharmacol. Sect.] 289 (1995) 97–101.
[3] R. Arai, N. Karasawa, M. Geffard, T. Nagatsu, I. Nagatsu, Immuno- [20] M. Hadjiconstantinou, T.A. Wemlinger, C.P. Silvia, J. Hubble, N.H.histochemical evidence that central serotonin neurons produce Neff, Aromatic L-amino-acid decarboxylase activity of mousedopamine from exogenous L-DOPA in the rat, with reference to the striatum is modulated via dopaminergic receptors, J. Neurochem. 60involvement of aromatic L-amino-acid decarboxylase, Brain Res. (1993) 2175–2180.
¨667 (1994) 295–299. [21] T. Hokfelt, K. Fuxe, M. Goldstein, Immunohistochemical localiza-[4] R. Arai, N. Karasawa, T. Nagatsu, I. Nagatsu, Exogenous L-5- tion of aromatic L-amino-acid decarboxylase (DOPA decarboxylase)
hydroxytryptophan is decarboxylated in neurons of the substantia in central dopamine and 5-hydroxytryptamine nerve cell bodies ofnigra pars compacta and locus coeruleus of the rat, Brain Res. 669 the rat, Brain Res. 53 (1973) 175–180.(1995) 145–149. [22] J. Iizuka, Controlled single-blind trial of budipine, L-dopa, and
[5] D.A. Bender, W.F. Coulson, Variations in aromatic amino acid amantadine in 102 patients with parkinson’s disease, in: F. Ger-decarboxylase activity towards DOPA and 5-hydroxytryptophan stenbrand, W. Poewe, G. Stern (Eds.), Clinical Experiences Withcaused by pH changes and denaturation, J. Neurochem. 19 (1972) Budipine in Parkinson Therapy, Springer-Verlag, New York, 1985,2801–2810. pp. 129–139.
[6] M.D. Berry, A.V. Juorio, X.M. Li, A.A. Boulton, Aromatic L-amino- [23] K. Jellinger, H. Bliesath, Adjuvant treatment of Parkinson’s diseaseacid decarboxylase: a neglected and misunderstood enzyme, Neuro- with budipine: a double-blind trial versus placebo, J. Neurol. 234chem. Res. 21 (1996) 1075–1087. (1987) 280–282.
¨[7] C.S. Biggs, A. Fisher, M.S. Starr, The antiparkinsonian drug [24] J. Kornhuber, J. Bormann, M. Hubers, K. Rusche, P. Riederer,budipine stimulates the activity of aromatic L-amino-acid decarbox- Effects of the 1-aminoadamantanes at the MK-801 binding site ofylase and enhances L-DOPA-induced dopamine release in rat the NMDA-receptor-gated ion channel: a human postmortem brainsubstantia nigra, Synapse 30 (1998) 309–317. study, Eur. J. Pharmacol. [Mol. Biol. Sect.] 206 (1991) 297–300.
[8] C.S. Biggs, L.J. Fowler, P.S. Whitton, M.S. Starr, NMDA receptor [25] N. Lindefors, U. Ungerstedt, Bilateral regulation of glutamate tissueantagonists increase the release of dopamine in the substantia of extracellular levels in caudate-putamen by midbrain dopaminereserpine-treated rats, Eur. J. Pharmacol. 299 (1996) 83–91. neurones, Neurosci. Lett. 115 (1990) 248–252.
[9] C.S. Biggs, L.J. Fowler, P.S. Whitton, M.S. Starr, Extracellular [26] K.G. Lloyd, O. Hornykiewicz, Parkinson’s disease: activity of L-levels of glutamate and aspartate in the entopeduncular nucleus of DOPA decarboxylase in discrete brain regions, Science 170 (1970)the rat determined by microdialysis: regulation by striatal dopamine 1212–1213.

274 A. Fisher, M.S. Starr / Brain Research 868 (2000) 268 –274
[27] E. Melamed, F. Hefti, D.J. Pettibone, J. Liebman, R.J. Wurtman, [35] M.L. Stachowiak, J.P. Bruno, A.M. Snyder, E.M. Stricker, M.J.Aromatic L-amino-acid decarboxylase in rat corpus striatum: impli- Zigmond, Apparent sprouting of striatal serotonergic terminals aftercations for action of L-DOPA in parkinsonism, Neurology [NY] 31 dopamine-depleting brain lesions in neonatal rats, Brain Res. 291(1981) 651–655. (1984) 164–167.
[28] N.H. Neff, M. Hadjiconstantinou, Aromatic L-amino-acid decarbox- [36] M.S. Starr, Glutamate /dopamine D /D balance in the basal ganglia1 2
ylase modulation and Parkinson’s disease, Progr. Brain Res. 106 and its relevance to Parkinson’s disease, Synapse 19 (1995) 264–(1995) 91–97. 293.
[29] K.L. O’Malley, S. Harmon, M. Moffat, A. Uhland-Smith, S. Wong, [37] H. Takashima, M. Tsujihata, M. Niwa, S. Nagataki, Effects ofThe human aromatic L-amino-acid decarboxylase gene can be chronic administration of L-DOPA and bromocriptine on the dopa-alternatively spliced to generate unique protein isoforms, J. Neuro- minergic metabolism in the rat striatum, Biogenic Amines 4 (1987)chem. 65 (1995) 2409–2416. 15–22.
[30] J. Opacka-Juffry, D.J. Brooks, L-Dihydroxyphenylalanine and its [38] S.S. Tate, R. Sweet, F.H. McDowell, A. Meister, Decrease of thedecarboxylase: new ideas on their neuroregulatory roles, Mov. 3,4-dihydroxyphenylalanine (DOPA) decarboxylase activities inDisord. 10 (1995) 241–249. human erythrocytes and mouse tissues after administration of
[31] M.H. Rahman, T. Nagatsu, T. Kato, Aromatic L-amino-acid de- DOPA, Proc. Natl. Acad. Sci. 68 (1971) 2121–2123.carboxylase activity in central and peripheral tissues and serum of [39] F. Tison, N. Mons, M. Geffard, P. Henry, The metabolism ofrats with L-DOPA and L-5-hydroxytryptophan as substrates, Bio- exogenous L-DOPA in the brain: an immunohistochemical study ofchem. Pharmacol. 30 (1981) 645–649. its conversion to dopamine in non-catecholaminergic cells in the rat
[32] H.F. Schott, W.G. Clark, DOPA decarboxylase inhibition through the brain, J. Neural Transm. 3 (1991) 27–39.interaction of coenzyme and substrate, J. Biol. Chem. 196 (1952) [40] M.Y. Zhu, A.V. Juorio, Aromatic L-amino-acid decarboxylase: bio-449–462. logical characterization and functional role, Gen. Pharmacol. 26
[33] K.L. Sims, G.A. Davis, F.E. Bloom, Activities of 3,4-dihydroxy-L- (1995) 681–696.phenylalanine and 5-hydroxy-L-tryptophan decarboxylases in rat [41] M.Y. Zhu, A.V. Juorio, I.A. Paterson, A.A. Boulton, Regulation ofbrain: assay characteristics and distribution, J. Neurochem. 20 aromatic L-amino-acid decarboxylase by dopamine receptors in the(1973) 449–464. rat brain, J. Neurochem. 58 (1992) 637–641.
[34] G. Skuza, Z. Rogoz, G. Quack, W. Danysz, Memantine, amantadine,and L-deprenyl potentiate the action of L-DOPA in monoamine-depleted rats, J. Neural Transm. [Gen. Sect.] 98 (1994) 57–67.





























