Embed Size (px)
Citation preview

Perspectives in Diabetes ATP-Sensitive K + Channels in Pancreatic P-Cells Spare-Channel Hypothesis DANIEL L. COOK, LESLIE S. SATIN, MICHAEL L.J. ASHFORD, AND C. NICHOLAS HALES
Since their discovery in pancreatic p-cells, ATP- sensitive K+ channels in the cell membrane have been thought to mediate glucose-induced p-cell depolarization, which is required for triggering the voltage-dependent Ca2+ uptake subserving insulin release. The theory is that metabolism of glucose (and other fuel molecules) increases intracellular ATP or possibly other metabolites that diffuse to the membrane and inhibit the opening of ATP-sensitive K+ channels. This slows the efflux of positively charged K+ and depolarizes the cell. A recurrent source of contusion regarding this idea stems from the early observation that these channels are so exquisitely sensitive to intracellular ATP that channel opening is predicted to be -99% inhibited under physiological conditions. To account tor this apparent discrepancy, various mechanisms have been proposed that might render the channels less sensitive to intracellular ATP. We use a simple mathematical model to demonstrate that there is no major discrepancy and that, in tact, given the electrophysiological mechanisms existing in the p-cell, the extreme sensitivity of the channels to ATP is appropriate and even mandatory for their phvsioloaical function. Diabetes 37:495-98, 1988
nhibition of ATP-sensitive K+ channels by metabolites of glucose in pancreatic p-cells produces the glucose-de- pendent depolarization required for triggering glucose- induced electrical activity (1-3; Fig. 1). Thus, in the
absence of glucose, K + efflux through ATP-sensitive K + channels contributes a hyperpolarizing membrane current that maintains the hyperpolarized resting membrane poten-
From the Departments of Medlc~ne and of Physiology and Blophysics. Unl- verslty ot Wash~ngton, and the Division of Metabolism, SeattleVeterans Adm~n- istral~on Med~cal Center, Seattle. Wash~ngton; and the Departments of Clinical Blocherrlistry and of Pharmacology, University of Cambridge, Cambridge, United Klngdom Address correspondence and reprlnt requests to Dr Danlel L. Cook, Division of Metabolism (151), Seattle VA Medical Center, 1660 Columbian Way South, Seattle, \NA 98108 Rece~ved for publication 23 January 1988 and accepted 26 January 1988.
tial of the cell. As glucose stimulation is increased, ATP- sensitive K+ channels are closed, the hyperpolarizing cur- rent is reduced, and the cell depolarizes. The links between glucose metabolism and the closure of ATP-sensitive K + channels are yet to be fully described, but it seems that ATP tself is a key second messenger, whereas other glucose metabolites may contribute.
As in various other tissues that have ATP-sensitive K + channels [e.g., heart (4,5), skeletal muscle (6) , and neurons (7)], one of the most unusual unexplained properties of such channels is their great sensitivity to ATP in relation to the concentrations of ATP believed to exist in the cytoplasm of most tissues. The greatest apparent discrepancy between the sensitivity to ATP (K, -15 pM; 1,3) and the ambient intracellular ATP concentrations (2-5 mM) is that existing in p-cells. This discrepancy has troubled some investigators, and various explanations have been suggested to account for its existence, including cytosolic factors (e.g., ADP, which may modify the sensitivity of the channels to ATP in the intact cell; 1 ,%lo), compartmentalization of ATP ( I ) , and the pos- sibility that only a small fraction of channels is involved in the regulation of p-cell membrane potential (3). We propose a hypothesis, for which there is empirical support (1 I ) , that may resolve this discrepancy. Using a quantitative approach with a model incorporating the observed K, and levels of ATP, we can predict changes in the p-cell membrane po- tential very close to those that have been observed. We point out the analogy between this regulatory mechanism and sys- tems based on apparently "spare receptors" and thus intro- duce the concept of apparently "spare channels." We sug- gest that this concept may be applicable to membrane potential regulation in other systems. We propose that the metabolic advantages of systems operating on the basis of apparently spare channels include 1 ) maximal sensitivity to changes of channel activity by operating near to the fully closed state; 2) operation of the system at very low net cur- rent levels, thus minimizing ion fluxes and energy expended to maintain ion gradients; and 3) stability of the membrane potential by the summation of brief openings of many chan- nels.
DIABETES VOL 37, MAY 1988 495

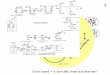
FIG. 1. Dlagram showing essential elements of system. Glucose metabolism produces ATP in mlllimolar levels sufficient to bind to and inhlblt opnlng of all but few ATP-msI tM K+ channels In p-cell membrane. At low levels of lntracellular ATP, many ATP-sensitive K+ channels are open,
I producing large hyperpolarlzlng K+ current that completely dominates small depolarlzlng leak current so that membrane potential Is held near K+ equilibrium potentlal (ca. -90 mV). As ATP level increases, progressively fewer ATP-sensitive K+ channels remain open, balance
ATP of currents begins to tip in favor of leak current, end membrane potential beglns to depolarize. Only when ATP level is high enough to inhiblt
GLUCOSE METABOLISM channel opening by >99% does cell finally depolarize into physlologlc range in whlch electrical activity Is triggered.
QUANTITATIVE APPROACH PREDICTS OBSERVED BEHAVIOR Two equations were derived for a single cell that model the effect of ATP-sensitive K + channels and an unspecified leak conductance on the depolarization of pancreatic p-cells while extracellular glucose is increased from zero to the threshold level (typically 25 mM), where regenerative elec- trical activity is triggered. The first equation relates intra- cellular ATP concentration to the K + conductance through ATP-sensitive K + channels (GK). We have assumed a single site of binding with constant affinity and a Hill coefficient of -1 (1,8), yielding
where Go is the total ATP-suppressible K + conductance that has been measured directly (3; unpublished observations), and where a value of 10,000 pS was used. The half-maximal inhibitory concentration of ATP (K,) was taken as 15 kM, as directly measured in excised patch-clamp studies in p-cells (1,8,10).
The second equation is a standard parallel conductance equation used to calculate the cell's resting membrane po- tential (Em) based on two competing conductances: an ATP- independent leak conductance (GL) and the ATP-dependent conductance through ATP-sensitive K + channels as deter- mined by Eq. 1
This standard equation calculates a resting membrane po- tential as a balance between the equilibrium potentials (EL and EK) for two parallel pathways weighted by their respec- tive conductances (G, and GK). In this case, the K + equilib- rium potential (E,) was assumed to be -90 mV, whereas the equilibrium potential of the leak conductance was as- sumed to be zero, i.e., EL = 0 mV. For Fig. 2, the total leak conductance was taken as 15 pS for the following reasons. Rorsman and Trube (3) found -50 pS of leak conductance with 3 mM ATP in their whole-cell patch-clamp pipette. How- ever, this leak was probably largely due to residual con- ductance through ATP-sensitive K + channels. Because Eq.
1 predicts, with 3 mM ATP in the pipette (and cell), that of the 10,000 pS at ATP-inhibitable conductance, -0.5% (or 50 pS) would still be active and account for virtually all of the measured leak conductance. Because a nonzero value of ATP-independent leak conductance must be used in Eq. 2, and acknowledging the problems of directly measuring such small conduc-tances, we chose 15 pS as a low value between zero and the measured value of 50 pS. Although its physiological nature is unknown, such a leak conductance must exist, because p-cells depolarize when their membrane K + conductance is decreased. Thus, a depolarizing leak current must counterbalance the hyperpolarizing ATP-sen- sitive resting K + current.
In Fig. 2, the normalized membrane conductance (GK/Go, Eq. 1) and the membrane potential (Em, Eq. 2) are plotted versus the logarithm of ATP concentration. The striking result here is that the membrane potential curve lies -2.5 log units (i.e., -300-fold) to the right of the GK/Go curve. Significant changes in membrane potential are not seen until the ATP- sensitive K + conductance is reduced to a small fraction of the total ATP-suppressible K + conductance by ATP con- centrations 100-fold higher than the K, for ATP. Clearly, the membrane potential will depolarize from - 75 to -60 mV (the threshold potential at which p-cell electrical activity is triggered) with a change of intracellular ATP concentration from 2 to 5 mM (Fig. 2, box).
An alternative approach to determining whether this theory models the behavior of p-cells is to consider whether the probability of channel opening is predicted for normal or physiological concentrations of glucose as measured on cell-attached patches. In the presence of 5 mM glucose, Misler et al. (10) measured average channel activity [i.e., the product of the number of channels in the patch (n) times the probability that each channel will be open (P)] and found nP = ,017. With n = 6 channels per patch, an estimate ( I I; unpublished observations) between the estimates of Misler et al. (8 per 1 kM2; 10) and Ashcroft et al. (1 per 3.5 kM2; 2), the open-channel probability can be calculated as ,0171 6 channels = ,003. Because the maximum open-channel probability for ATP-sensitive K+ channels in the absence of ATP is only -.2 (unpublished observations), the G,/Go for a single channel is .003/.2 = ,015. This value for GK/Go is equivalent to a concentration between -2.0 and 2.5 mM ATP (Fig. 2).
DIABETES, VOL. 37, MAY 1988

ASSUMED PARAMETERS HAVE REASONABLE VALUES In pointing out that the observed behavior of the p-cell mem- brane potential and the cell's ATP content are not in gross discrepancy, we do not intend to imply that the only factor capable of regulating the ATP-sensitive K t channel is ATP. Furthermore, several assumptions that we have used in our calculations may be in error, and it is worthwhile to consider what effect they may have on this theoretical relationship. The complete molecular basis of ATP-mediated inhibition is unknown. We have used a simple model involving only the net open-channel probability. In keeping with the behavior of other channels and detailed kinetic analysis, the existence of several open and closed states will probably be indicated. Although the consideration of such states might refine our model closer to reality, the overall conclusions should not be affected.
The total cellular conductance due to maximally open ATP- sensitive K+ channels is not certain. The activity of this chan- nel "runs down" after patch excision (13), and the same is true when cells are dialyzed in the whole-cell recording con- figuration on which this measurement relies ( I ; unpublished observations). Thus, 10,000 pS may be taken as a minimum. The effect of increasing total ATP-suppressible K+ con- ductance is to shift the curve relating membrane potential tc log ATP to the right. This would exaggerate the dose shift relative to the GJG, curve and improve the force of our argument.
The leak conductance was taken as 15 pS. Doubling the leak conductance to 30 pS shifts the membrane potential curve to the left by -0.5 log unit. This illustrates the impor- tance of obtaining a precise measurement of this parameter and of understanding its physiological basis. We do not feel that we have grossly underestimated leak conductance enough to shift the membrane potential curve out of the range of physiologic ATP concentration. Note that if leak conductance were itself subject to reciprocal control by adenine nucleotides (as the Ca2+-activated nonselective cat- ion channel may be; 14), the slope of the response of mem- brane potential to changes in ATPIAMP would be consid- erably enhanced. Changing the equilibrium potential would not shift the membrane potential versus log-ATP curve but
FIG. 2. Results of mathematical formalization of concepts in Fig. 1. Normalized conductance (GK/G,, Eq. 1; broken line, left axis) and membrane potential (Em, Eq. 2; solid line, right axis) are plotted as functions of logarithm of ATP concentration in micromoles. Equations predict >2.5 log unit (-300-fold) right shlR of membrane potential curve relative to conductance- suppressing effect of ATP. Box encloses physiologic range of intracellular ATP concentration between 2 and 5 mM and physiologic membrane potential range of -75 to - 60 mV.
would only change its slope. For example, if EL = +50 mV, as for a pure Na+ permeability, the slope would increase.
Whether the actual changes in ATP content of cells ex- posed to metabolic stimulants of insulin secretion are suffi- cient to account for the observed changes in membrane potential is unknown. In other words, is the dose-response curve to ATP in the right region, and is the slope of the response steep enough? In the absence of knowledge of possible local changes in ATP concentration it is impossible to answer these questions rigorously (15,16). However, be- cause changes in total ATP content on the order of 30-50% can be detected when glucose concentration is raised to the stimulatory threshold (17,18), it seems entirely feasible that local changes in ATP concentration near the membrane would be even larger, if anything.
PHYSIOLOGICAL ADVANTAGES OF SPARE-CHANNEL MECHANISM Inhibition of the ATP-sensitive K+ channel by ATP as a reg- ulatory system differs from classic agonist-receptor regula- tory systems because the channel is most active in the ab- sence of the regulator. Nevertheless, the basic properties of a regulatory system based on inhibition are probably similar to those based on activation, including the need for maxi- mum sensitivity and the need for minimum energy expen- diture. Only a few receptors need to be activated to give the full biologic effect of the agonist in many receptor-agonist systems. This property of response was originally consid- ered to be due to the existence of spare receptors (19). The apparent excess of receptors is not due to truly spare re- ceptors in the sense that they are unnecessary for the re- sponse to the agonist, because all receptors are capable of interacting with the agonist and exerting an effect. Only a few such units need be activated to exert a full biologic effect that is somehow lim~ted at a later stage of the signaling process. The net effect is to shift the dose-response curve to the left, to an agonist concentration considerably lower than the midpoint of the agonist-receptor binding curve. A major difference with this is that the spare-channel mecha- nism shifts the dose-response curve (Em in this case) to the right rather than to the left of the agonist-receptor binding
1 10 100 1 10 100 micromolar millimolar
lntracellular [ATP]
DIABETES, VOL. 37, MAY 1988

curve (coincident with the G,IG, curve). Another conse- include the effect of channel activity on membrane potential, quence is that of maximizing sensitivity because, in the ab- the key physiological endpoint. In doing this, we are able to sence of cooperativity, the bottom end of the dose-response predict that, given the number of ATP-sensitive K + channels curve provides the greatest change of receptor activity per in p-cells, it is indeed appropriate, even mandatory, that only change of agonist concentration. a few be open at any one time to control membrane potential
We suggest that it is valuable and valid to adapt these in the physiologic voltage range. concepts to the situation in which regulation occurs via in- hibition of an otherwise active system. Maximum sensitivity in this case results from the modulation of the net conduc- tance of many almost fully closed channels (Eq. 1). This effect is then amplified by the conductance-balancing mech- anism that controls the membrane potential (Eq. Z), which means that small changes in the open-state probability of many ATP-sensitive channels can dramatically alter the bal- ance of inward and outward currents and thus affect the membrane potential. At first, the production of many chan- nels by cells only to have them spend most of their time in the closed state would suggest that the channels are un- necessary or spare. However, we propose that the closed channels are an essential element of the system as is the situation with apparently spare receptors, conferr~ng on it advantageous characteristics.
It is possible to envisage a system that would be equally sparing of ion fluxes simply by the presence of far fewer ATP-sensitive channels with commensurately lower sensitiv- ity to inhibition by ATP. The main disadvantage of such a system would be the stochastic noise of the opening and closing of these few channels. ATP-sensitive channels, when mainly inhibited, open only briefly [e.g., in isolated patches (1,8) and cell-attached patches (2,3)]. In contrast, many of these channels, each contributing only brief pulses of current that are filtered by the cell's membrane capacitance, would produce a relative quiet and stable membrane potential. If, on the other hand, there were only 10 channels that were inhibited by 50% (thereby satisfying the energy requirements for a low level of ion movement), the statistical possibility of all the channels being open or closed at the same time would lead to a noisy and unstable membrane potential. Thus, we see several advantages of apparently spare channels in a system based on a regulator that inhibits an otherwise active channel; maximum sensitivity (i.e., the largest relative change of signal for the smallest relative change in the reg- ulator) is achieved by regulation close to the inactive state, the inactive state is the least energy expensive, and the sum of the activity of many channels operating at low levels of open probability confers greater stability than fewer chan- nels generating the same current with longer open times.
Regardless of the evolutionary history of the system, or the teleological rationale for its existence in p-cells, it is clear from our quantitative analysis that the initial intuitive concern about the degree of channel inhibition was well based; the channels are, as predicted, -99% inhibited at any one time. The importance of this study is to extend the analysis to
ACKNOWLEDGMENTS D.L.C. and L.S.S. were supported by the National Institutes of Health, Diabetes Research Council, Seattle, Washington, and the U.S. Veterans Administration. M.L.J.A. and C.N.H. were supported by the British Diabetes Association, Medical Research Council, and Wellcome Trust.
REFERENCES 1 Cook DL. Hales CN lntracellular ATP directly blocks K + channels in
pancreatic 0-cells Nature (Lond) 31 1 :271-73, 1984 2 Ashcroft FM, Harrison DE, Ashcroft SJH: Glucose induces closure of
single potassium channels in isolated rat pancreatlc beta cells Nature (Lond) 312 446-48, 1984
3 Rorsman P, Trube G Glucose-dependent K+ channels ~n pancreatic beta- cells are regulated by lntracellular ATP. Pfluegers Arch 405 305-309, 1985
4 Noma A ATP-regulated K+ channels ~n cardlac muscle. Nature (Lond) 305.1 47-48, 1983
5 Kake~ M, Noma A. Shibasaki T. Propert~es of adenos~ne-triphosphate- regulated potasslum channels in gulnea-p~g ventricular cells. J Physiol (Lond) 363.441-62, 1985
6. Spruce AE, Standen NB, Stanf~eld PR: Voltage-dependent, ATP-sensitive potassium channels of skeletal muscle membrane. Nature (Lond) 31 6 736-38, 1985
7 Ashford MLJ, Sturgess NC, Trout NJ, Gardener NJ, Hales CN ATP-sen- sitive channels In neonatal rat cultured neurones (Abstract) Br J Phar- macol91 301 P, 1987
8 Kake~ M, Kelly RP, Ashcroft SJH, Ashcroft FM: The ATP-sens~tivity of K + channels In 0-cells is modulated by ADP. FEES Lett 208 63-66, 1986
9 Dunne MJ, Petersen OH. lntracellular ADP activates K t channels that are inhib~ted by ATP in an ~nsulin-secreting cell Ine. FEES Lett 208:59- 62. 1986
10 Misler S, Falke LC, Gillis K, McDanlel ML. A metabolite-regulated potas- sium channel In rat pancreatlc 0 cells. Proc Natl Acad Sci USA 83 71 19- 23, 1986
11. Arkhammar P, N~lsson T, Rorsman P, Berggren P-0 lnhibit~on of ATP- reoulated K ' channels orecedes deoolarization-induced increase in cv- topasmic free Ca2+ concentration in pancreatic beta cells J Biol hem 262:5448-54, 1987
12. Ohno-Shosak~ T, Zuckler BJ, Trube G. Dual effects of ATP on K+-currents of mouse pancreatic beta cells. Pfluegers Arch 408 133-38, 1987
13. F~ndlay I, Dunne MJ, Petersen OH, ATP-sensitive inward rectifier and voltage and calcium-activated K+ channels in cultured pancreatic islet cells J Membr El01 88.165-72, 1985
14 Sturgess NC. Hales CN, Ashford MLJ Inhibition of a calcium activated, nonselective cation channel, in a rat insulinoma cell line, by aden~ne der~vatives FEES Lett 208,397-400, 1986
15 We~ss JN, Lamp ST: Glycolysis preferentially inhibits ATP-sensitive K+ channels In Isolated gu nea pig cardiac myocytes Science 238:67-69, 1987
16 M~ller DS Horow~tz SB lntracellular compartmental~zat~on of adenos~ne tr~phosphate J E ~ o l Chem 261 13911-15 1986
17 Mala~sse WJ Sener A Glucose-~nduced changes n cytosol~c ATP content In pancreallc Islets B~ochim E~ophys Acta 927 190-95 1987
18 Ashcroft SJH Chatra C Weers~nahe C Randle PJ lnterrelat~onsh~os of Islet metabol~sm, adenos~ne trlphosphate content and ~nsul~n relAase Blochem J 132 223-31 1973
19 Kahn CR Membrane receptors for hormones and neurotransmitters J Cell Eiol 70 261 -86 1976
DIABETES. VOL 37. MAY 1988