Embed Size (px)
Citation preview

Journal of Experimental Botany, Vol. 34, No. 142, pp. 493-506, May 1983
Positive and Negative Messages from Roots Induce
Foliar Desiccation and Stomatal Closure in Flooded
Pea Plants
MICHAEL B. JACKSON AND ALINA K. B. KOWALEWSKA
Agricultural Research Council Letcombe Laboratory, Letcombe Regis, Wantage,Oxfordshire, 0X12 9JT, UK.
Received 9 December 1982
ABSTRACTFlooding the soil for 5-7 d caused partial desiccation in leaves of pea plants {Pisum sativum. L. cv.'Sprite*)- The injury was associated with anaerobiosis in the soil, a large increase in the permeability ofleaf tissue to electrolytes and other substances, a low leaf water content and an increased watersaturation deficit {WSD). Desiccating leaves also lacked the capacity to rehydrate in humidatmospheres, a disability expressed as a water resaturation deficit (WRSD). This irreversible injurywas preceded during the first 4-5 d of flooding by closure of stomata within 24 h, decreasedtranspiration, an unusually large leaf water content and small WSD. Leaf water potentials were higherthan those in well-drained controls. Also, there was no appreciable WRSD. Leaflets detached fromflooded plants during this early phase retained their water more effectively than those from controlswhen left exposed to the atmosphere for 5 min.
Stomatal closure and the associated increase in leaf hydration could be simulated by excising leavesand incubating them with their petioles in open vials of water. Thus, such changes in flooded plantspossibly represented a response to a deficiency in the supply of substances that would usually betransported from roots to leaves in healthy plants (negative message). Ion leakage and the associatedloss of leaf hydration that occurs when flooding is extended for more than 5 d could not be simulated byisolating the leaves from the roots. Appearance of this symptom depended on leaves remainingattached to flooded root systems, implying that the damage is caused by injurious substances passingupwards (positive message). Both ethylene and ethanol have been eliminated as likely causes, butflooding increased phosphorus in the leaves to concentrations that may be toxic.
Keywords: Pisum sativum; Flooding; Foliar desiccation; Stomata; Ethylene.
I N T R O D U C T I O NWater is the major molecular constituent of herbaceous plants and must be continuouslyavailable in the soil for survival. An excess of soil water can, however, be just as damaging asdrought for the many species not adapted to a wetland environment. In these conditions,water slows gaseous exchange preventing both the ready diffusive penetration of soil byatmospheric oxygen and escape of gaseous products of metabolism such as ethylene andcarbon dioxide. The resulting chemical and biochemical changes within the rooting zonedamage not only roots but also indirectly, the aerial shoot system (Grable, 1966; Drew andLynch, 1980; Cannell and Jackson, 1981; Bradford, 1982). In peas, leaves rapidly senesce
at University of W
innipeg on August 25, 2014
http://jxb.oxfordjournals.org/D
ownloaded from

494 Jackson and Kowalewska—Foliar Desiccation in Flooded Pea Plants
and desiccate (Cannell, Gales, Snaydon, and Suhail, 1979; Jackson, 1979a; Bedford, Cannell,Thomson, and Dennis, 1980).
Unlike more flood-tolerant species such as sunflower (Kawase, 1974), maize (Jackson,Drew, and Giffard, 1981), barley (Trought and Drew, 1980) and tomato (Jackson andCampbell, 1975) peas fail to produce an adventitious root system at or near the surface offlooded soil. The shoots of flooded pea plants must therefore suffer from an absence ofsubstances normally provided by functioning roots (negative messages). They may also beaffected by anaerobically generated metabolites passing from anoxic roots or soil into theshoot (positive messages). The upward movement of one such substance explains theethylene-mediated epinastic leaf curvature of flooded tomato plants (Jackson and Campbell,1976; Bradford and Yang, 1981) and re-orientation of shoot extension in the 'diageotropic'mutant (Jackson, \919b). A third influence that may conceivably bear on shoot behaviour isan accumulation of hormones and other substances that would usually be exported to healthy,aerobic roots (Cannell and Jackson, 1981, p. 153). In this paper we assess the possiblecontribution of negative and positive messages causing foliar desiccation and stomatal closurein flooded pea plants.
M A T E R I A L S A N D M E T H O D SPlants of Pisum sativum L. (cv. 'Sprite*) were grown in pairs in 11-5 cm diameter pots containing'Levington' peat-based compost (Jackson, 1979a) either in a glasshouse (minimum temperature,15 °C) or in a controlled environment cabinet with a 16 h photoperiod (375—400 ̂ mol m~2 s~' PAR)at 24 °C and a night temperature of 14 °C. Intact plants were used when the sixth or seventh leaf wasexpanding, and were flooded to the soil surface for up to 7 d by plunging pots into 1200 ml vesselscontaining deionized water that was made up to volume daily. Non-flooded plants were watered byweight to 'pot capacity' each morning. In some experiments leaves were cut off singly from the base ofthe plant sequentially upwards during growth to maintain only two fully expanded leaves on each plant.When these leaves corresponded to positions 4 and 5 (the oldest leaf being in position 1) the plantswere de-topped and the subsequently emerging side-shoots removed. This allowed effects of flooding onleaves 4 and 5 to be studied without influence from large metabolic sinks elsewhere in the shoot. Thephysiology of these leaves was compared with that of similar leaves isolated from such de-leafed andde-topped plants and incubated for up to 7 d with their petioles in 10 ml vials of deionized water.
Transpiration rate for whole plants was estimated from the loss in weight (±0-1 g) occurring eachday between 2 h and 5 h after the start of the photoperiod, or over intervals of 24 h, after correcting forevaporation from the soil surface. Leaf water content (L WC) was calculated by difference between thefresh and oven-dry weights of leaflets of known area. Water saturation deficit (WSD) was measured inleaflets incubated for 5 h in 10 ml screw-top vials containing 1-0 ml of water at 23 °C. The resulting'saturated fresh weight' together with the initial fresh weight and dry weight were used to calculateWSD (Barrs, 1968). Water resaturation deficits (WRSD) were calculated from:
Mean saturated H2O content of Saturated H2O content ofleaves from non-flooded plants ~ leaves from flooded plants
WRSD = x 100Mean saturated H2O content of leaves from non-flooded plants
WRSD is a measure of the capacity of leaves from flooded plants to absorb and retain water in asaturated atmosphere compared with controls. Xylem water potentials in leaves were estimated within30 s of excision using a pressure chamber. It was necessary to slice off one side of the hollow petiole toensure a gas-tight seal. No corrections were made for the osmotic potential of the xylem sap. This wasmeasured in other experiments with a Roebling micro-osmometer in sap expressed from the roots ofde-topped plants by applying up to 0-15 MPa in a pressure chamber. Leaf conductance interpreted asstomatal conductance on the abaxial surface was assessed with a diffusion porometer (Parkinson andLegg, 1972) and expressed as conductance (cm s"1)- Rates of water loss from whole leaflets of knownarea were obtained from the change in weight over 5 min using a Cahn model 7550 electrobalancesensitive to ^0-01 mg. The rate of weight loss measured every 30 s was almost constant over this time.
Ethylene evolution from leaves was increased by spraying to 'run-off' at 10.00 h and 16.00 h each
at University of W
innipeg on August 25, 2014
http://jxb.oxfordjournals.org/D
ownloaded from

Jackson and Kowalewska—Foliar Desiccation in Flooded Pea Plants 495
day for 7 d with 1-aminocylopropane-l-carboxylic acid (ACC) dissolved in water containing 0-1%Tween 80.
Chlorophyll was extracted in 80% (v/v) ethanol in the dark at 4 °C for 7 d prior to optical densitymeasurements at 665 run. The outward leakage of ions after floating individual leaflets for 5 h ondouble distilled/deionized water containing 0-01% Tween 80 was estimated using a Philips PW9501conductivity meter sensitive to 0-1 //S. The intensity of fluorescence in these solutions was measuredwith a Baird-Atomic Fluoricord spectrofluorimeter. The combination of excitation and emissionwavelengths used (280 nm and 350 nm) gave the most intense fluorescence and was close to theoptimum for the aromatic amino acid tryptophan (Udenfriend, 1969). Leakage rate from the leafletswas constant throughout the 5 h incubation. Total nitrogen in leaf tissue was analysed with a CarloErba 1300 automatic nitrogen analyser; phosphorus was estimated in nitric and perchloric acid digestsof leaves by the molybdenum blue reaction followed by automated colorimetry. Atomic absorptionspectrophotometry was used for potassium, and gas chromatography for ethylene and oxygendeterminations in soil water or gas extracted under partial vacuum from roots and shoots (Jackson andCampbell, 1975; Hall, 1978).
R E S U L T S(1) Water relationshipsWaterlogging the soil caused extensive senescence and desiccation of the foliage. This hasbeen quantified previously together with other symptoms of injury (Jackson, 1979a; Jackson,Herman, and Goodenough, 1982). Typically, 4 d flooding partially or completely desiccatedmore than half the leaves.
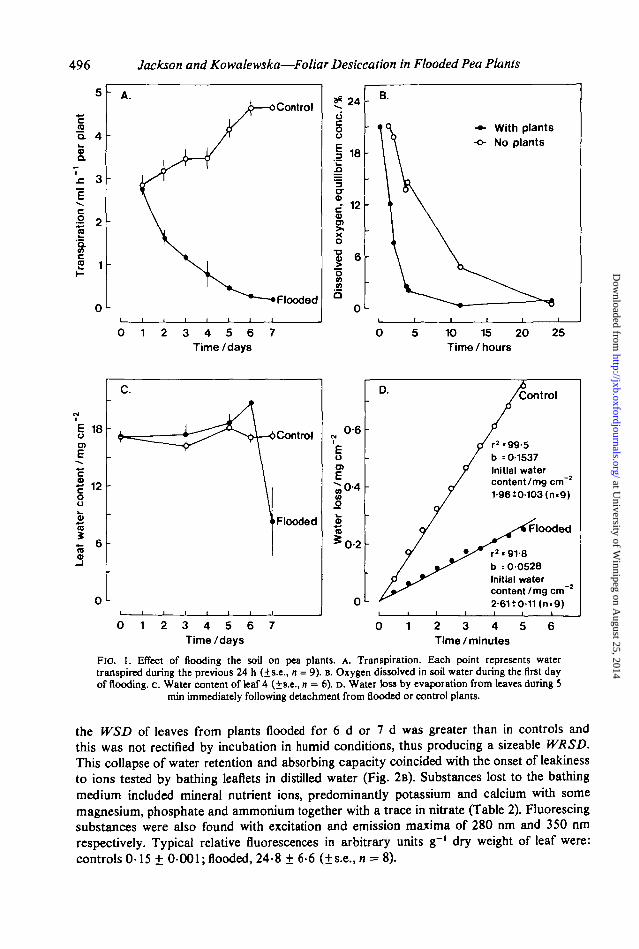
Desiccation was not preceded by fast rates of water loss from flooded plants. On thecontrary, transpiration slowed during the first 24 h and this trend continued for a further 6 d(Fig. 1A) in association with de-oxygenation of the soil that was partly brought about by thepresence of the plants (Fig. 1B). Unexpectedly, desiccation was not preceded by an increasedleaf water deficit Instead, leaves of flooded plants initially showed slightly larger rather thansmaller L WCs (Fig. lc). Only after about 5 d of flooding did the position reverse when leaveslost their water rather suddenly (Fig. lc). Rates of water loss from individual leaflets weremeasured after detachment from the plant and weighing every 30 s for 5 min. Leaves fromplants flooded for 4 d showed a higher initial water content than those from controls andretained their water more effectively during the subsequent 5 min on the balance pan (Fig.ID). Comparable results were obtained with leaves from plants flooded for 2, 3 or 7 d exceptthat by the seventh day their initial water content was less than controls, because desiccationhad begun.
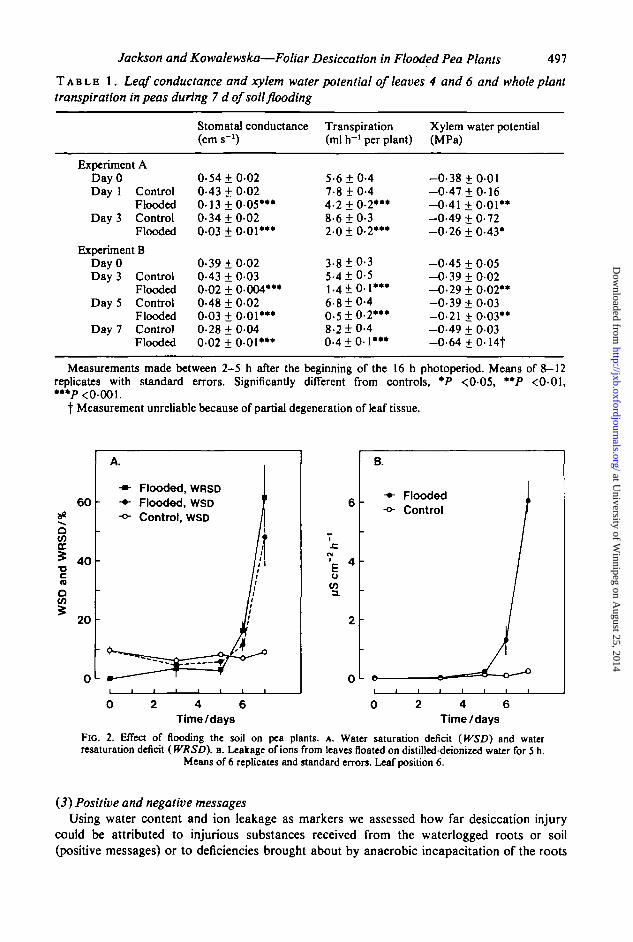
Water in the form of vapour is lost by transpiration predominantly through stomata.Microscopical examination of nitrocellulose ('clear' nail varnish) impressions of the adaxialleaf surface showed partial stomatal closure in flooded plants. Diffusion porometry confirmedthis quantitatively. Stomatal conductance was extremely small after 24 h of flooding (Table1A) and remained so throughout the 7 d of the experiment. This accounted both for the slowertranspiration and the xylem leaf water potentials that were higher (less negative) than controlsduring the first 5 d of the experiment (Table IB). By the seventh day partial desiccationprevented further reliable estimates of the xylem leaf water potential of flooded plants.
(2) DesiccationThe ability of leaves to absorb and retain water when detached and incubated for 5 h in
sealed vials with petioles in 1 0 ml of water was measured. This enabled water saturation deficits(WSD) prior to the incubation and water resaturation deficits (WRSD) at the end of theincubation to be calculated. During the first 5 d of flooding, leaves from waterlogged plantshad slightly smaller WSDs than controls (Fig. 2A). This was expected from the water contentand xylem water potential measurements already given (Fig. l c and Table 1). HoweveT,
at University of W
innipeg on August 25, 2014
http://jxb.oxfordjournals.org/D
ownloaded from
496 Jackson and Kowalewska—Foliar Desiccation in Flooded Pea Plants
c
"a 4
I
a 2
Iro 1
Control
Flooded
0 1 2 3 4 5 6Time /days
ug
3
libr
3
1Ko
u
SO
h
CO
5
24
18
12
6
0
B.
' 1S
-
•*• With plants\ -o- No plants\\K\ \\\ V
~——• ~9
10 15 20Time/hours
25
en
I 12o
Control
"Flooded
0-6
0-4
0-2
D.
-
-
7/ ^
0" - *"i i i
/Control/
/</ rJ«99-5
/ b =0-15379 Initial water
/ content/mg cm"2
7 1-96i0-103(n.9)
^ ^ T F I o o d e d
• ^ r2.91-8^ b =00528
Initial watercontent /mg cm"2
2-6U0H(ni9)i i i i
1 2 3 4 5 6 7Time/days
2 3 4 5Time/minutes
FIG. 1. Effect of flooding the soil on pea plants, A. Transpiration. Each point represents watertranspired during the previous 24 h (+s.e., n = 9). B. Oxygen dissolved in soil water during the first dayof flooding, c. Water content of leaf 4 (±s.e., n = 6). D. Water loss by evaporation from leaves during 5
min immediately following detachment from flooded or control plants.
the WSD of leaves from plants flooded for 6 d or 7 d was greater than in controls andthis was not rectified by incubation in humid conditions, thus producing a sizeable WRSD.This collapse of water retention and absorbing capacity coincided with the onset of leakinessto ions tested by bathing leaflets in distilled water (Fig. 2B). Substances lost to the bathingmedium included mineral nutrient ions, predominantly potassium and calcium with somemagnesium, phosphate and ammonium together with a trace in nitrate (Table 2). Fluorescingsubstances were also found with excitation and emission maxima of 280 nm and 350 runrespectively. Typical relative fluorescences in arbitrary units g~' dry weight of leaf were:controls 0-15 ± 0-001; flooded, 24-8 ± 6 6 (±s.e., n = 8).
at University of W
innipeg on August 25, 2014
http://jxb.oxfordjournals.org/D
ownloaded from
Jackson and Kowalewska—Foliar Desiccation in Flooded Pea Plants 497
T A B L E 1. Leaf conductance and xylem water potential of leaves 4 and 6 and whole planttranspiration in peas during 7 d of soil flooding
Experiment ADayODay 1
Day 3
ControlFloodedControlFlooded
Experiment BDayODay 3
Day 5
Day 7
ControlFloodedControlFloodedControlFlooded
Stomatal conductance Transpiration(cm s~')
0-54 + 0-020-43 + 0020-13 + 0-05***0-34 + 002003 ± 001***
0-39 + 0020-43 + 0030-02 + 0-004***0-48 + 002003 + 0-01***0-28 + 0040-02 + 001***
(ml h"1 per plant)
5-6 ±0-47-8 + 0-44-2 + 0-2***8-6 + 0-32-0±0-2**»
3-8 ±0-35-4 ±0-5l-4± 0-1***6-8 ±0-4O-5 + O-2***8-2 ±0-40-4±0-l**»
Xylem water potential(MPa)
-0-38 + 001-0-47 + 0-16-0-41 ±0-01**-0-49 ±0-72-0-26 ±0-43*
-0-45 + 005-0-39 ± 0 0 2-0-29 + 002**-0-39 + 003-0-21 ±0-03**-0-49 + 003-O-64±0-14f
Measurements made between 2-5 h after the beginning of the 16 h photoperiod. Means of 8—12replicates with standard errors. Significantly different from controls, *P <0-05, **P <0-01,
f Measurement unreliable because of partial degeneration of leaf tissue.
601—.aa.•ac(0Q
A.
•+• Flooded, WRSD-•- Flooded, WSD-o- Control, WSD
o
B.
FloodedControl
0 L •-
2 4 6Time /days
2 4 6Time/days
Fio. 2. Effect of flooding the soil on pea plants, A. Water saturation deficit (WSD) and waterresaturation deficit (WRSD). B. Leakage of ions from leaves floated on distilled-deionized water for 5 h.
Means of 6 replicates and standard errors. Leaf position 6.
(3) Positive and negative messagesUsing water content and ion leakage as markers we assessed how far desiccation injury
could be attributed to injurious substances received from the waterlogged roots or soil(positive messages) or to deficiencies brought about by anaerobic incapacitation of the roots
at University of W
innipeg on August 25, 2014
http://jxb.oxfordjournals.org/D
ownloaded from
498 Jackson and Kowalewska—Foliar Desiccation in Flooded Pea Plants
(negative messages). Chlorophyll content was also measured since chlorosis often accom-panied desiccation or preceded it.
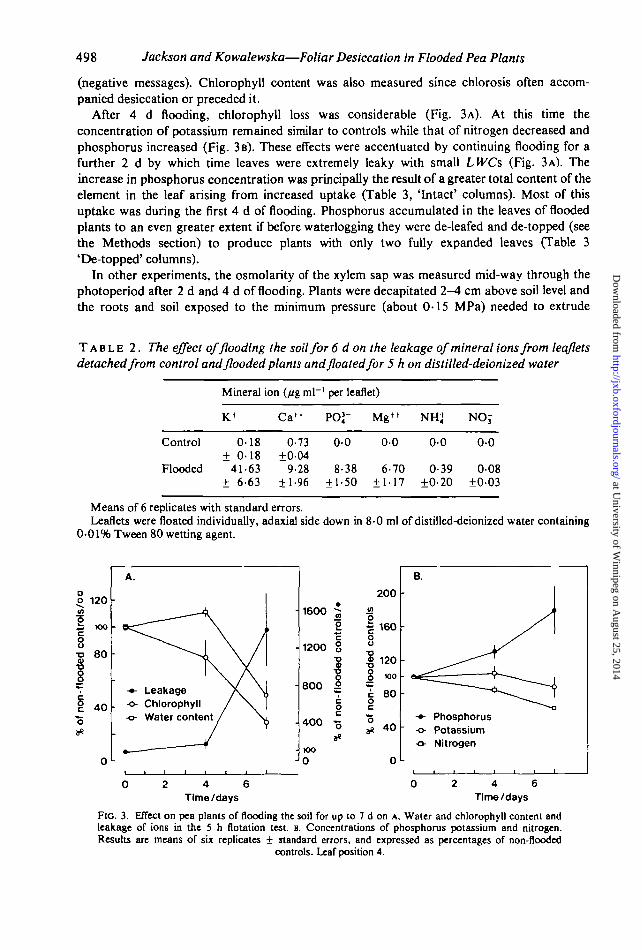
After 4 d flooding, chlorophyll loss was considerable (Fig. 3A). At this time theconcentration of potassium remained similar to controls while that of nitrogen decreased andphosphorus increased (Fig. 3 B). These effects were accentuated by continuing flooding for afurther 2 d by which time leaves were extremely leaky with small LWCs (Fig. 3A). Theincrease in phosphorus concentration was principally the result of a greater total content of theelement in the leaf arising from increased uptake (Table 3, 'Intact' columns). Most of thisuptake was during the first 4 d of flooding. Phosphorus accumulated in the leaves of floodedplants to an even greater extent if before waterlogging they were de-leafed and de-topped (seethe Methods section) to produce plants with only two fully expanded leaves (Table 3'De-topped' columns).
In other experiments, the osmolarity of the xylem sap was measured mid-way through thephotoperiod after 2 d and 4 d of flooding. Plants were decapitated 2-4 cm above soil level andthe roots and soil exposed to the minimum pressure (about 0-15 MPa) needed to extrude
T A B L E 2. The effect of flooding the soil for 6 d on the leakage of mineral ions from leafletsdetached from control and flooded plants and floated for 5 hon distilled-deionized water
Mineral ion (jig ml"1 per leaflet)
Ca+ POJ- Mg++ NH+ NO7
Control
Flooded
0-18+ 0-18
41-63+ 6-63
0-73±0-04
9-28+ 1-96
0 0 0 0 0 0 0-0
8-38 6-70 0-39 0-08±1-50 ±1-17 ±0-20 ±0-03
Means of 6 replicates with standard errors.Leaflets were floated individually, adaxial side down in 8-0 ml of distilled-deionized water containing
0-01% Tween 80 wetting agent.
A.
•*- Leakage-o- Chlorophyll-o- Water content
1600
1200
co400 ~
1000
4 6Time /days
? 8800 2 =
200
160
120
80
40
0
B.
-
-
•
- • -
' 0-
-0-
1
-a
PhosphorusPotassiumNitrogen
• 1 1 1 1 • 1
2 4 6Time/days
FIG. 3. Effect on pea plants of flooding the soil for up to 7 d on A. Water and chlorophyll content andleakage of ions in the 5 h flotation test. B. Concentrations of phosphorus potassium and nitrogen.Results are means of six replicates ± standard errors, and expressed as percentages of non-flooded
controls. Leaf position 4.
at University of W
innipeg on August 25, 2014
http://jxb.oxfordjournals.org/D
ownloaded from
Jackson and Kowalewska—Foliar Desiccation in Flooded Pea Plants 499
T A B L E 3. Effect of flooding intact or de-topped pea plants on the phosphorus content anduptake of leaflets and their dry weight
Time fromstart offlooding(d)
04
ControlFlooded
7ControlFlooded
Phosphorus (jig, per leaflet)
Total content
Intact
205 ± 12
193+ 13384 ± 39***
202 + 11385 ± 29***
De-topped
232 ± 15
256 + 19733 ± 50***
256+ 12622 ± 64***
Uptake from
Intact
0
-12 ± 13179 ± 36***
- 3 + 7180 ±25***
dayO
De-topped
0
24 +501 ±
24 +390 ±
1945***
958***
Leafletdry wt(mg)
Intact
23 ± 1
23 ± 120 ± 1
26+ 118±0-9***
De-topped
27 ± 2
35 + 234 ± 2
3 7 + 227 ± 2***
Means of 12 replicates and their standard errors.*** Significantly different from controls P = <0001.Results are for single leaflets from leaf positions 4 and 5.
T A B L E 4. Effect of flooding intact pea plants at the five- to six-leaf stage on the osmolarityand water potential of the xylem sap expressed from the cut stump immediately after decap-itation 2—4 cm above the soil surface
Time (h) Osmolarity (mOsm) Water potential (MPa)
Control Flooded Control Flooded
0 27-6 ±4-2 — 0-067 ±0-010 —48 23-8 ±1-1 57-2 ±10-3* 0-058 ± 0-002 0-139 ±0-03*96 26-8 ±2-2 84-5 ± 13-4** 0-065 ± 0-006 0-206 ±0-03**
Means of five replicates and standard errors.Significantly different from controls, *P <0-05, **P <0-01.Conversion factor, 1-0 MPa = 410-5 mOsm at 20 °C.
50—100 fi\ of sap from the cut stump. Sap osmolarity was up to three times greater in floodedplants than in controls, thereby decreasing the xylem water potential by 0-08-0-14 MPa(Table 4).
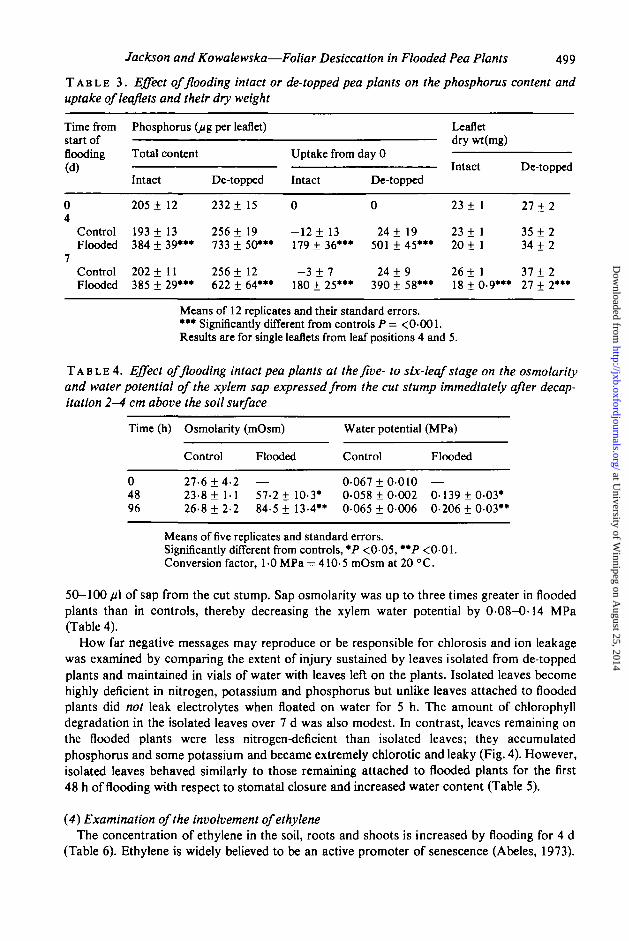
How far negative messages may reproduce or be responsible for chlorosis and ion leakagewas examined by comparing the extent of injury sustained by leaves isolated from de-toppedplants and maintained in vials of water with leaves left on the plants. Isolated leaves becomehighly deficient in nitrogen, potassium and phosphorus but unlike leaves attached to floodedplants did not leak electrolytes when floated on water for 5 h. The amount of chlorophylldegradation in the isolated leaves over 7 d was also modest. In contrast, leaves remaining onthe flooded plants were less nitrogen-deficient than isolated leaves; they accumulatedphosphorus and some potassium and became extremely chlorotic and leaky (Fig. 4). However,isolated leaves behaved similarly to those remaining attached to flooded plants for the first48 h of flooding with respect to stomatal closure and increased water content (Table 5).
(4) Examination of the involvement ofethyleneThe concentration of ethylene in the soil, roots and shoots is increased by flooding for 4 d
(Table 6). Ethylene is widely believed to be an active promoter of senescence (Abeles, 1973).
at University of W
innipeg on August 25, 2014
http://jxb.oxfordjournals.org/D
ownloaded from
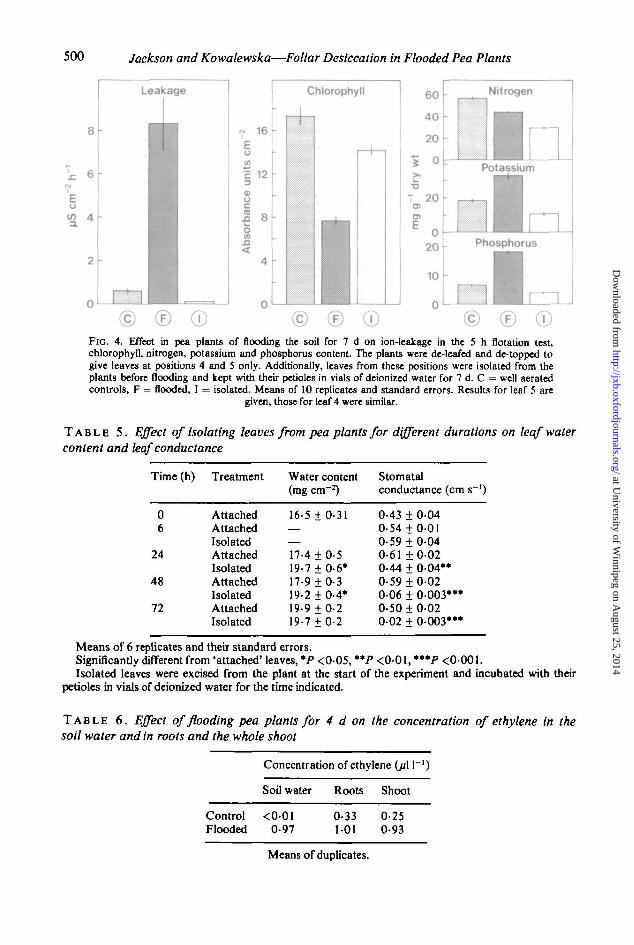
500 Jackson and Kowalewska—Foliar Desiccation in Flooded Pea Plants
u(/) 4
Leakage
© ©FIG. 4. Effect in pea plants of flooding the soil for 7 d on ion-leakage in the 5 h flotation test,chlorophyll, nitrogen, potassium and phosphorus content The plants were de-leafed and de-topped togive leaves at positions 4 and 5 only. Additionally, leaves from these positions were isolated from theplants before flooding and kept with their petioles in vials of deionized water for 7 d. C = well aeratedcontrols, F = flooded, I = isolated. Means of 10 replicates and standard errors. Results for leaf 5 are
given, those for leaf 4 were similar.
T A B L E 5. Ejfect of isolating leaves from pea plants for different durations on leaf watercontent and leaf conductance
Time (h)
06
24
48
72
Treatment
AttachedAttachedIsolatedAttachedIsolatedAttachedIsolatedAttachedIsolated
Water content(mg cm"2)
16-5 ±0-31——17-4 ± 0-519-7 + 0-6*17-9 ±0-319-2+ 0-4*19-9 + 0-219-7 + 0-2
Stomatalconductance (cm s~')
0-43 + 0-040-54 + 0-010-59 ±0-040-61 +0-020-44 + 0-04"0-59 + 0-020-06 + 0-003***0-50 + 0-020-02 + 0-003*"
Means of 6 replicates and their standard errors.Significantly different from 'attached' leaves, *P <0-05, **P <0-01, ***P <0-001.Isolated leaves were excised from the plant at the start of the experiment and incubated with their
petioles in vials of deionized water for the time indicated.
T A B L E 6. Effect of flooding pea plants for 4 d on the concentration of ethylene in thesoil water and in roots and the whole shoot
Concentration of ethylene Oil 1~')
Soil water Roots Shoot
Control <001 0-33 0-25Flooded 0-97 101 0-93
Means of duplicates.
at University of W
innipeg on August 25, 2014
http://jxb.oxfordjournals.org/D
ownloaded from
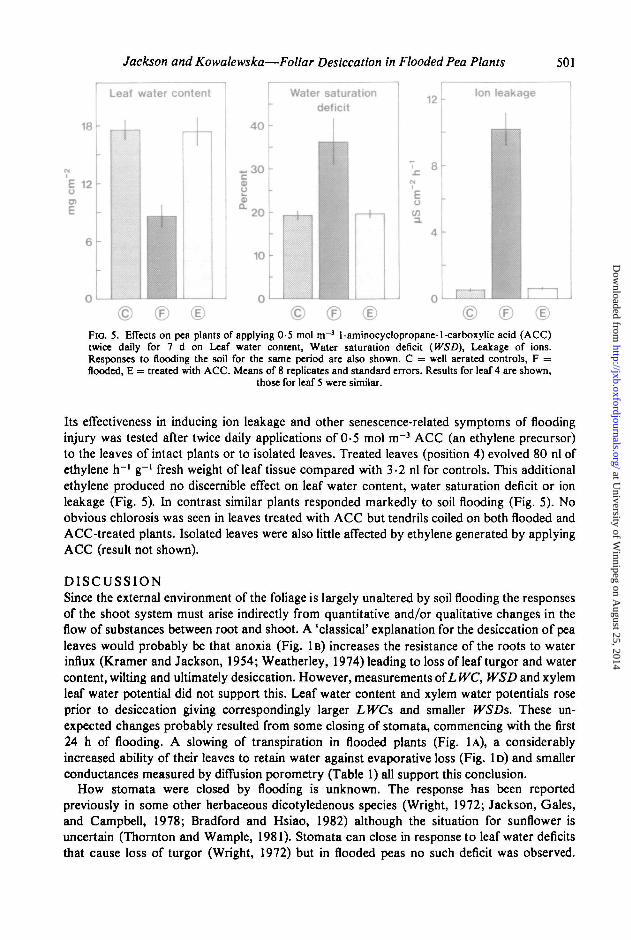
Jackson and Kowalewska—Foliar Desiccation in Flooded Pea Plants
12
501
40
20
10
Water saturationdeficit
© ©
u
© ©Fio. 5. Effects on pea plants of applying 0-5 mol m~3 1-aminocyclopropane-l-carboxylic acid (ACC)twice daily for 7 d on Leaf water content, Water saturation deficit (WSD), Leakage of ions.Responses to flooding the soil for the same period are also shown. C = well aerated controls, F =flooded, E = treated with ACC. Means of 8 replicates and standard errors. Results for leaf 4 are shown,
those for leaf 5 were similar.
Its effectiveness in inducing ion leakage and other senescence-related symptoms of floodinginjury was tested after twice daily applications of 0-5 mol m~3 ACC (an ethylene precursor)to the leaves of intact plants or to isolated leaves. Treated leaves (position 4) evolved 80 nl ofethylene h"1 g"1 fresh weight of leaf tissue compared with 3-2 nl for controls. This additionalethylene produced no discernible effect on leaf water content, water saturation deficit or ionleakage (Fig. 5). In contrast similar plants responded markedly to soil flooding (Fig. 5). Noobvious chlorosis was seen in leaves treated with ACC but tendrils coiled on both flooded andACC-treated plants. Isolated leaves were also little affected by ethylene generated by applyingACC (result not shown).
D I S C U S S I O NSince the external environment of the foliage is largely unaltered by soil flooding the responsesof the shoot system must arise indirectly from quantitative and/or qualitative changes in theflow of substances between root and shoot. A 'classical' explanation for the desiccation of pealeaves would probably be that anoxia (Fig. 1 B) increases the resistance of the roots to waterinflux (Kramer and Jackson, 1954; Weatherley, 1974) leading to loss of leaf turgor and watercontent, wilting and ultimately desiccation. However, measurements of L WC, WSD and xylemleaf water potential did not support this. Leaf water content and xylem water potentials roseprior to desiccation giving correspondingly larger LWCs and smaller WSDs. These un-expected changes probably resulted from some closing of stomata, commencing with the first24 h of flooding. A slowing of transpiration in flooded plants (Fig. 1A), a considerablyincreased ability of their leaves to retain water against evaporative loss (Fig. ID) and smallerconductances measured by diffusion porometry (Table 1) all support this conclusion.
How stomata were closed by flooding is unknown. The response has been reportedpreviously in some other herbaceous dicotyledenous species (Wright, 1972; Jackson, Gales,and Campbell, 1978; Bradford and Hsiao, 1982) although the situation for sunflower isuncertain (Thornton and Wample, 1981). Stomata can close in response to leaf water deficitsthat cause loss of turgor (Wright, 1972) but in flooded peas no such deficit was observed.
at University of W
innipeg on August 25, 2014
http://jxb.oxfordjournals.org/D
ownloaded from

502 Jackson and Kowalewska—Foliar Desiccation in Flooded Pea Plants
Closure must therefore have had some other cause, although an influence of transient waterdeficits during the first day of flooding cannot be entirely ruled out. No such deficit wasdetected in flooded tomatoes by Bradford and Hsiao (1982) although a small temporary fall inxylem leaf water potential was reported in this species by Jackson et al. (1978) after 6 h offlooding. Abscisic acid is a hormone active in closing stomata when applied to peas (Jackson,unpublished). Wright (1972) observed a five-fold increase in endogenous ABA after 5 dflooding of dwarf bean and Hall, Kapuya, Sivakumaran, and John (1977) found a temporaryincrease lasting 3 d in Viciafaba. Increases in this hormone may thus close stomata in flooded peaplants. A water deficit is the usual prerequisite for ABA concentrations to increase in thestomatal guard cells of environmentally stressed plants (Davies, Mansfield, and Wellbura,1980). There must, however, be a different cause if flooding does indeed increase ABA in theleaves of pea. Possible explanations include an importation of ABA from the flooded roots(positive message) or an accumulation of ABA that would normally have been exported to theroots (Hartung, 1976). We find that detached leaves maintained with their petioles in wateralso closed their stomata on the day following excision in the absence of water stress (Table 5)(Xu and Lou, 1980; Bradford and Hsiao, 1982). This supports the idea that stomata couldclose because anaerobic roots no longer supply the substances that normally maintainstomata open during the photoperiod (negative message). Such substances could well includegibberelh'n and cytokinin hormones since their application to flooded tomato plants partiallyinhibits closure (Gales and Jackson, unpublished) and consequently induces some wilting(Jackson and Campbell, 1979). Potassium ions are required for regulating the turgor ofstomatal guard cells and Regehr, Bazzaz, and Boggess (1975) speculated that potassiumdeficiency in flooded poplar may decrease stomatal apertures. This cannot be the mechanismin flooded peas because potassium in the leaves did not decrease (Fig. 3). An unexploredpossibility is that flooding increases the carbon dioxide content of leaves sufficiently to closestomata caused by the import of the gas from the soil or by injury to the photosyntheticapparatus (Bradford, 1982). Closing of stomata is probably a significant adaptive responsewithout which temporary increases in resistance to water flow through roots in the first hoursof waterlogging (Kramer and Jackson, 1954; Bradford and Hsiao, 1982) would favour wiltingand water stress that could prejudice recovery and survival. This view must be tempered,however, by the many reported examples linking stomatal closure with accelerated leafsenescence (Wardle, 1982 for a review). A more thorough examination of the hormonal,nutritional and water relationships of flooded plants that lead to stomatal closure and itsconsequences for the survival of flooding is clearly required.
Unexpectedly, after about 5 d of effective water conservation, leaves of flooded peasquickly lose water content and desiccate, although slow rates of transpiration and stomata]conductance ensure relatively gentle evaporative demand. It seems that leaf cells losetheir ability to absorb and retain water even under these mild conditions. This conclusion wasconfirmed by finding that after 6-7 d flooding, leaves had small water contents and largerWSDs that could not be rectified by a rehydration test in sealed vials. This shortfall,quantified as a WRSD (Fig. 2), coincided with the onset of leakiness to inorganic ions andlarger organic molecules, suggesting breakdown of the plasmalemma. We conclude that thisloss of semi-permeability causes a failure of osmotic control that ultimately leads todesiccation.
Some possible causes of membrane damage have been investigated. A loss of nitrogenoccurred before ion leakage (Fig. 3). Furthermore chlorophyll decreased before the leavesleaked ions (Fig. 4). Since nitrogen deficiency favours chlorosis (Drew, Sisworo, and Saker,1979) and chloroplast membranes are often the first to degenerate during senescence (Gahan,1982), a sequence of nitrogen loss leading to chloroplast injury that extends later to other
at University of W
innipeg on August 25, 2014
http://jxb.oxfordjournals.org/D
ownloaded from

Jackson and Kowalewska—Foliar Desiccation in Flooded Pea Plants 503
membrane systems is perhaps indicated. However, when nitrogen deficiency in excess of thatfound in flooded plants was induced by isolating leaves from the plant and incubating themwith their petioles in water, chlorosis was less severe than in leaves remaining on floodedplants. These isolated leaves also failed to leak ions in the 5 h-long flotation test (Fig. 4).
The above experiments indicate that pea leaves are not injured primarily by deficiencies inmaterials such as nitrogen normally provided by well-aerated roots (negative messages)because leaves isolated from such roots did not reproduce flooding injury. Ion leakage anddesiccation are more likely to be a consequence of the passage of injurious substances fromthe anoxic roots or soils (positive messages) to which they remain connected by the stele. Onepotential toxin is phosphorus. This major inorganic plant nutrient has been reported by othersto cause foliar necrosis following excessive accumulations after a period of phosphorusstarvation (Bhatti and Loneragan, 1970; Green, Ferguson, and Warder, 1973). Theconcentrations we measured in the leaves of flooded peas are abnormally large (Figs 3 and 4)and within the range of 12-50 fig g"1 dry weight reported to be associated with tissue damage(references in Bhatti and Loneragan, 1970). Necrosis and desiccation in pea leaves of floodedplants is most severe at the outer edges, where the phosphorus concentration may beparticularly high (Clarkson and Scattergood, 1982). The concentrating effect brought aboutby flooding is almost entirely the result of increased absorption by the leaves (Table 3). A lossin dry weight sustained during flooding for 7 d may also have raised the concentrations by asmall amount because the results are expressed on a dry weight basis (Table 3). This increaseduptake is surprising in view of the energy requiring nature of ordinary phosphate uptake byroots. The phosphorus may have its source in the degenerating root system itself.Accumulation of phosphorus is enhanced in flooded plants previously trimmed to only twoleaves (Table 3), presumably because of the resulting decrease in shoot size relative to that ofthe roots. Flooding does not increase phosphorus levels in monocots such as maize (Jacksonet al, 1981, but see Singh and Ghildyal, 1980) or barley (Drew and Sisworo, 1979), butphosphorus accumulation in other species and its potential as a positive message that injuresthe foliage of flooded plants deserves closer examination.
A second positive message that may accelerate desiccation is an increased daytimeosmolarity of the xylem sap (Table 4). The effect is probably the combined result ofdiminished dilution caused by slow transpiration and a release of solutes from anoxic rootsthat leak their cell contents (Grineva, 1961). In peas, a three-fold increase in osmolarity wasfound after 4 d flooding. The resulting lower xylem water potential would impose somerestriction on the movement of water into leaf cells although the significance of this is unclear.During the first 5 d of flooding stomatal closure more than compensated for this since theoverall leaf water potential rose sufficiently to permit L WCs to exceed that of control plants.
Ethanol is one of the solutes of xylem sap in flooded peas. Concentrations up to 2-1 molm~3 were found after 3 d flooding (Jackson et al, 1982). Ethanol is reputed to be highly toxicbut despite many suggestions in the literature that flooded plants die from ethanol poisoning,pharmacological tests show it to be innocuous at concentrations up to 100 times those presentin flooded peas (Jackson et al, 1982). Ethanol can therefore be eliminated as one of thepositive messages from anoxic roots that causes membrane injury to pea leaves.
Ethylene is another candidate that can probably be eliminated for similar reasons. The gas isoften said to promote leaf senescence (Abeles, 1973) although evidence in the literature canreveal rather weak activity (Burg, 1968, Table 2 and Fig. 5) or be indirect (Hallaway andOsborne, 1969). The additional ethylene in the shoots of flooded peas (Table 6) is probablyderived from ACC transported from anoxic roots as described previously for tomatoes (seeIntroduction for references). In that species, a 4—6-fold increase in ethylene production in thepetioles readily promoted the epinastic curvature typical of flooded tomato plants. However, a
at University of W
innipeg on August 25, 2014
http://jxb.oxfordjournals.org/D
ownloaded from

504 Jackson and Kowalewska—Foliar Desiccation in Flooded Pea Plants
25-fold increase in ethylene production in pea leaves obtained by applying ACC failed toreproduce symptoms of flooding injury such as ion leakage, decreased water content (Fig.5) or chlorosis. Isolated leaves were similarly unaffected, indicating that flooding is unlikelyto increase significantly the sensitivity of leaves to ethylene by suppressing the export ofinhibitors of ethylene action from the root system.
We conclude that desiccation in flooded pea plants is the result of membrane breakdownthat renders leaf cells osmotically incompetent. The effect seems to be a response to one ormore substances passing into the leaves from the flooded and anoxic roots (positive message).Ethylene and ethanol are insufficiently active to be responsible at least on their own but thepossibility of phosphate toxicity deserves examination in future work. Leaves becamedesiccated despite rapid stomatal closure, the cause of which is also unknown but it may beinduced by a negative message.
A C K N O W L E D G E M E N T SWe thank Dr. K. Gales and Miss Sarah M. Ayling for help in measuring stomatalconductances and xylem leaf water potentials, Mr. W. Downs and his colleagues in theChemistry and Electronics Section of the Letcombe Laboratory for analysing inorganicnutrients and Mrs Alice Izzard for excellent technical assistance.
L I T E R A T U R E C I T E DABELES, F. B., 1973. Ethylene in plant biology. Academic Press, New York and London. Pp. 156-61.BARRS, H. D., 1968. Determination of water deficits in plant tissues. In Water deficits and plant growth.
Vol. I. Development, control and measurement. Ed. T. T. Kozlowski. Academic Press, New Yorkand London. Pp. 235-368.
BELFORD, R. K., CANNELL, R. Q., THOMSON, R. J., and DENNIS, C. W., 1980. Effects of waterlogging
at different stages of development on the growth and yield of peas (Pisum sativum L.). Journal ofthe Science of Food and Agriculture, 31,857-69.
BHATTI, A. S., and LONERAGAN, J. F., 1970. Phosphorus concentrations in wheat leaves in relation tophosphorus toxicity. Agronomy Journal, 62, 288-90.
BRADFORD, K. J., 1982. Regulation of shoot responses to root stress by ethylene, abscisic acid andcytoktnin. In Plant growth substances 1982. Ed. P. F. Wareing, Academic Press, London.Pp. 599-608.and HSIAO, T. C , 1982. Stomatal behaviour and water relations of waterlogged tomato plants.
Plant Physiology, 70,1508-13.and YANG, S. F., 1981. Physiological responses of plants to waterlogging. Horticultural Science,16, 25-30.
BURG, S. P., 1968. Ethylene, plant senescence and abscission. Plant Physiology, 43,1503-11.CANNELL, R. Q., GALES, K., SNAYDON, R. Q., and SUHAIL, B. A., 1979. Effects of short-term
waterlogging on the growth and yield of peas (Pisum sativum). Annals of Applied Biology, 93,327-35.and JACKSON, M. B., 1981. Alleviating aeration stresses. In Modifying the root environment to
reduce crop stress. Eds G. F. Arkin and H. M. Taylor. American Society of AgriculturalEngineers. St. Joseph. Pp. 141-92.
CLARKSON, D. T., and SCATTERGOOD, C. B., 1982. Growth and phosphate transport in barley andtomato plants during the development of, and recovery from, phosphate stress. Journal ofExperimental Botany, 33,865-75.
DAVIES, W. J., MANSFIELD, T. A., and WELLBURN, A. R., 1980. A role for abscisic acid in droughtendurance and drought avoidance. In Plant growth substances 1979. Ed. F. Skoog. Springer-Verlag, Berlin, Heidelberg, New York. Pp. 242-53.
DREW, M. C., and LYNCH, J. M., 1980. Soil anaerobiosis, micro-organisms, and root function. AnnualReviews of Phytopathology, 18,37—66.and SISWORO, E. J., 1979. The development of waterlogging damage in young barley plants in
relation to plant nutrient status and changes in soil properties. New Phytologist, 82,301-14.
at University of W
innipeg on August 25, 2014
http://jxb.oxfordjournals.org/D
ownloaded from

Jackson and Kowalewska—Foliar Desiccation in Flooded Pea Plants 505
and SAKER, L. R., 1979. Alleviation of waterlogging damage to young barley plants byapplication of nitrate and a synthetic cytokinin, and comparison between the effects ofwaterlogging, nitrogen deficiency and root excision. Ibid. 82,315-29.
GAHAN, P. B., 1982. Cytochemical and ultrastnictural changes in cell senescence and death. In Plantgrowth regulators in plant senescence, Monograph 8. Eds M. B. Jackson, B. Grout and I. A.Mackenzie. British Plant Growth Regulator Group, Wantage. Pp. 47-55.
GRABLE, A. R., 1966. Soil aeration and plant growth. Advances in Agronomy, 18,57-106.GREEN, D. G., FERGUSON, W. S., and WARDER, F. G., 1973. Accumulation of toxic levels of
phosphorus in the leaves of phosphorus-deficient barley. Canadian Journal of Plant Science, 53,241-6.
GRTNBVA, G. M., 1961. Excretion by plant roots during brief periods of anaerobiosis. Soviet PlantPhysiology, 8,549-52.
HALL, K. C , 1978. A gas chromatographic method for the determination of oxygen dissolved in waterusing an electron capture detector. Journal of Chromatographic Science, 16,311-13.
HALL, M. A., KAPUYA, J. A., SIVAKUMARAN, S., and JOHN, A., 1977. The role of ethylene in the
response of plants to stress. Pesticide Science, 8,217-23.HALLAWAY, M., and OSBORNE, D. J., 1969. Ethylene, a factor in defoliation induced by auxins,
Science, 163,1067-8.HARTUNG, W., 1976. Effect of water stress on transport of [2-14C] abscisic acid in intact plants of
Phaseolus coccineus L. Oecologia, 26,177-83.JACKSON, M. B., 1979a. Rapid injury to peas by soil waterlogging. Journal of the Science of Food and
Agriculture, 30,143-52.19796. Is the diageotropic tomato ethylene-deficient? Physiologiaplantarum, 46,347-51.and CAMPBELL, D. J., 1975. Movement of ethylene from roots to shoots, a factor in the responses
of tomato plants to waterlogged soil conditions. New Phytologist, 74, 397-406.1976. Waterlogging and petiole epinasty in tomato: The role of ethylene and low oxygen.
Ibid. 76,21-9.1979. The effects of benzyladenine and gibberellic acid on the responses of tomato plants to
anaerobic root environments and to ethylene. Ibid. 82, 331—40.DREW, M. C , and GIFFARD, S. C , 1981. Effects of applying ethylene to the root system of Zea
mays on growth and nutrient concentration in relation to flooding tolerance. Physiologiaplantarum, 52,23-8.
GALES, K., and CAMPBELL, D. J., 1978. Effect of waterlogged soil conditions on the productionof ethylene and on water relationships in tomato plants. Journal of Experimental Botany, 29,183-93.HERMAN, B., and GOODENOUGH, A., 1982. An examination of the importance of ethanol in
causing injury to flooded plants. Plant, Cell and Environment, 5, 163-72.KAWASE, M., 1974. Role of ethylene in induction of flooding damage in sunflower. Physiologia
plantarum, 31,29-38.KRAMER, P. J., and JACKSON, W. T., 1954. Causes of injury to flooded tobacco plants. Plant
Physiology, 29,241-5.PARKrNSON, K. J., and LEGG, B. J., 1972. A continuous flow porometer. Journal of Applied Ecology, 9,
669-75.REGEHR, D. L., BAZZAZ, F. A., and BOGGESS, W. R., 1975. Photosynthesis, transpiration and leaf
conductance ofPopulus deltoides in relation to flooding and drought. Photosynthetica, 9,52-61.SINGH, R., and GHILDYAL, B. P., 1980. Soil submergence effects on nutrient uptake, growth and yield
of five corn cultivars. Agronomy Journal, 72, 737—41.THORNTON, R. K., and WAMPLE, R. L., 1981. Changes of sunflower in response to water stress
conditions. Plant Physiology, 65, 7 (Suppl.).TROUGHT, M. C. T., and DREW, M. C , 1980. The development of waterlogging damage in young
wheat plants in anaerobic solution cultures. Journal of Experimental Botany, 31, 1573-85.UDENFRIEND, S., 1969. Fluorescence assay in biology and medicine, Vol. 2. Academic Press, New
York, London.WARDLE, K., 1982. Stomatal responses and foliar senescence. In Growth regulators in plant
senescence, Monograph 8. Eds M. B. Jackson, B. Grout and I. A. Mackenzie. British PlantGrowth Regulator Group, Wantage. Pp. 85-92.
WEATHERLEY, P. E., 1974, The hydraulic resistance of the root system. In Structure and function ofprimary root tissues. Ed. J. Kolek. Veda. Bratislava. P. 305.
at University of W
innipeg on August 25, 2014
http://jxb.oxfordjournals.org/D
ownloaded from

506 Jackson and Kowalewska—Foliar Desiccation in Flooded Pea Plants
WRIGHT, S. T. C , 1972. Physiological and biochemical responses to wilting and other stressconditions. In Crop processes in controlled environments. Eds A. R. Rees, K. E. Cockshull, D. W.Hand and R. G. Hurd. Academic Press, London, New York. Pp. 349—61.
Xu, X-D., and Lou, C-H., 1980. Presence of roots—a prerequisite for normal function of stomata onsweet potato leaves. Bulletin of the Bering Agricultural University, first issue. Pp. 37—45.(Chinese with English abstract.)
at University of W
innipeg on August 25, 2014
http://jxb.oxfordjournals.org/D
ownloaded from










![Stomatal Biology of CAM Plants1[CC-BY] - Plant … on Stomatal Biology Stomatal Biology of CAM Plants1[CC-BY] Jamie Males* and Howard Griffiths Department of Plant Sciences, University](https://img.pdfslide.net/doc/110x75/5ae808d37f8b9a08778f24b1/stomatal-biology-of-cam-plants1cc-by-plant-on-stomatal-biology-stomatal.jpg)



![Evolution of the Stomatal Regulation of Plant Water ...Update on Stomatal Evolution Evolution of the Stomatal Regulation of Plant Water Content[OPEN] Timothy J. Brodribb* and Scott](https://img.pdfslide.net/doc/110x75/5e87e202c27a1d71d24f112b/evolution-of-the-stomatal-regulation-of-plant-water-update-on-stomatal-evolution.jpg)














