Embed Size (px)
Citation preview

Bacterial Diversity in the Rhizosphere of
AVP1 Transgenic Cotton (Gossypium hirsutum
L.) and Wheat (Triticum aestivum L.)
Muhammad Arshad
2016
Department of Biotechnology
Pakistan Institute of Engineering & Applied Sciences
Nilore-45650 Islamabad, Pakistan


Reviewers and Examiners
Foreign Reviewers
1. Dr. Dittmar Hahn
Department of Biology, Texas State University,
601 University Drive San Marcos,
Fax +1 (512) 245 8713
Telephone: +1 (512) 245 3372
E-mail Address: [email protected]
2. Dr. Philippe Normad
Microbial Ecology Laboratory, UMR CNRS 5557, F-69622 Villeurbanne
Cedex Telephone: 33 (0)4-7243-1377
E-mail Address: [email protected]
University of Arkansas,
3. Dr. Katharina Pawlowski
Stockholm University
Mailing Address: SE-106 91 Stockholm, Sweden
Telephone (With Country Code): +46 8 16 37 72
E-mail Address: [email protected]
Thesis Examiners
1. Dr. Asghari Bano,
Department of Biosciences university of Wah cant
Telephone # 03129654341
E-mail Address: [email protected]
2. Dr. Muhammad Arshad
Department of Botany, PMAS AAU, Murree Road, Rawalpindi
Telephone: 051-9062207
E-mail Address: [email protected]
3. Dr. Amer Jamil,
Molecular Biochemistry Lab, Dept. of Chemistry and Biochemistry,
University of Agriculture Faisalabad
Telephone: 41-9201104
E-mail Address: [email protected]
Head of the Department (Name): Prof. Dr. Shahid Mansoor, S.I.
Signature with date: _____________________

Thesis Submission Approval
This is to certify that the work contained in this thesis entitled Bacterial Diversity in
the Rhizosphere of AVP1 Transgenic Cotton (Gossypium hirsutum L.) and Wheat
(Triticum aestivum L.), was carried out by Muhammad Arshad, and in my opinion,
it is fully adequate, in scope and quality, for the degree of M. Phil leading to Ph.D.
Furthermore, it is hereby approved for submission for review and thesis defense.
Supervisor: _____________________
Name: Dr. Muhammad Sajjad Mirza
Date: 27 December, 2016
Place: NIBGE, Faisalabad
Co-Supervisor: __________________
Name: Dr. Shaheen Asad
Date: 27 December, 2016
Place: NIBGE, Faisalabad
Head, Department of Biotechnology: ___________________
Name: Dr. Shahid Mansoor (S.I)
Date: 27 December, 2016
Place: NIBGE, Faisalabad

Bacterial Diversity in the Rhizosphere of
AVP1 Transgenic Cotton (Gossypium hirsutum
L.) and Wheat (Triticum aestivum L.)
Muhammad Arshad
Submitted in partial fulfillment of the requirements
for the degree of Ph.D.
2016
Department of Biotechnology
Pakistan Institute of Engineering and Applied Sciences
Nilore-45650 Islamabad, Pakistan

ii
Dedications
To
My Parents
&
My innocent kids
Muhammad and Anaya

iii
Acknowledgements
Nothing is deserving of worship except “ALMIGHTY ALLAH”, all praises for Him,
Who is the entire source of all knowledge and wisdom endowed to mankind. He guides
the way and gives me courage to complete this work. I offer my humblest gratitude
from deep sense of heart to the Holy Prophet, MUHAMMAD (Peace be Upon Him)
Who is, forever source of guidance and knowledge for humanity.
I am very grateful to my PhD supervisor Dr. Muhammad Sajjad Mirza,
Deputy Chief Scientist, NIBGE Faisalabad for his professional and technical guidance,
scientific discussions and suggestions, keen interest in completion of this task and moral
support during whole period of research and compilation of thesis. I also pay thanks to
my foreign supervisor Professor Dr. Johan Leaveau at Plant Pathology Department
University of California Davis CA, USA for his kind and technical support and valuable
contribution during my visit to the host lab for six month fellowship. I will appreciate
Mr. Gurdeep Rastogi, my senior colleague and Mr. Jan Tech, lab in charge at
Pathology Lab, University of California Davis CA, USA.
I would also like to appreciate and acknowledge the efforts of Dr. Shahid
Mansoor (S.I.), Director (NIBGE), and Dr. Suhail Hameed (Exe. Director NIBGE)
for maintaining the honor of this institute among other research organizations of
Pakistan. I would like to acknowledge Dr. Shaheen Asad (co-superviser) and Dr.
Nasir A saeed Principle Scientists at NIBGE, Faisalabad, for providing all plant
material. I would like to appreciate Mr. Muhammad Arshad Senior Scientist at NIBGE
and Mr. Masood Anwar for their cooperation during this course of study.
I am also indebted to my lab colleagues Dr. Muther Mansoor Qaisrani, Dr.
Muhammad Tahir and Mr. Ahmad Zaheer for their help in learning research
techniques and theoretical discussions. I am also thankful to Mr. Muhammad Ahmad
and Muhammad Imran technicians at Microbial Ecology Lab, NIBGE, Faisalabad,
for the kind help in conducting lab and field experiments. The help from Dr. Farooq

iv
Latif (DCS) and Dr. Ghulam Rasul (PS), at NIBGE, Faisalabad for analysis of
organic acids and phytohormones on HPLC is thankfully acknowledged.
Special thanks are due to my parents who waited a long time. I am especially
thankful to my wife Aisha Arshad who suffered my long absence at home brought up
my beloved kids Muhammad and Anaya with full care and provided me the spiritual
and moral support during this long period of study, research work and in thesis writing.
Many friends have helped me stay sane through these difficult years.
Particularly, I am thankful to Dr. Atif Iqbal, Dr. Asif Habib Dr. Ikram Anwar and
Sohail Mehmood Kareemi. Their support and care helped me to overcome setbacks
and stay focused on my graduate study. I greatly value their friendship and I deeply
appreciate their belief in me. I have no words to pay sincerest thanks to my friends for
their help, encouragement and great friendship that made easier for me to overcome
difficulties in all hard times.
At the end, I would like to acknowledge Higher Education Commission,
Pakistan, for providing me funds for my doctoral research in Pakistan and University
of California, Davis CA USA. Without this financial support I might not be able to
focus on my research.
Muhammad Arshad

v
Declaration of Originality
I hereby declare that the work accomplished in this thesis is the result of my own
research carried out in Soil & Environmental Biotechnology Division (NIBGE). This
thesis has not been published previously nor does it contain any material from the
published resources that can be considered as the violation of international copyright
law. Furthermore, I also declare that I am aware of the terms ‘copyright’ and
‘plagiarism’, and if any copyright violation was found out in this work I will be held
responsible of the consequences of any such violation.
__________________
(Muhammad Arshad)
27 December, 2016
NIBGE, Faisalabad.

vi
Copyrights Statement
The entire contents of this thesis entitled Bacterial Diversity in the Rhizosphere of
AVP1 Transgenic Cotton (Gossypium hirsutum L.) and Wheat (Triticum aestivum
L.) by Muhammad Arshad are an intellectual property of Pakistan Institute of
Engineering & Applied Sciences (PIEAS). No portion of the thesis should be
reproduced without obtaining explicit permission from PIEAS.

vii
Table of Contents
Dedications .................................................................................................................... ii
Acknowledgements ...................................................................................................... iii
Declaration of Originality .............................................................................................. v
Copyrights Statement .................................................................................................... vi
Table of Contents ......................................................................................................... vii
List of Figures ............................................................................................................... xi
List of Tables .............................................................................................................. xiv
Abstract ...................................................................................................................... xvii
List of Publications and Patents .................................................................................. xix
List of Abbreviations and Symbols.............................................................................. xx
1. Introduction ................................................................................................................ 1
1.1 Genetically Modified Crops .................................................................................. 1
1.2 Use of AVP1 Gene to Develop Transgenic Plants ................................................ 2
1.3 Bacterial Diversity in Rhizosphere of Genetically Modified Plants ..................... 4
1.4 Plant Growth Promoting Rhizobacteria (PGPR) .................................................. 5
1.4.1 Mode of Action of PGPR .......................................................................... 5
1.4.2 Nitrogen Fixation ...................................................................................... 6
1.4.3 Biological Nitrogen Fixation (BNF) ......................................................... 8
1.4.4 Diversity of Diazotrophic Bacteria ........................................................... 9
1.4.5 The Domain Archea .................................................................................. 9
1.4.6 Phosphorus Mineralization by Microbes for Plant Growth Promotion .. 11
1.4.7 Phytohormone Production by PGPR for Plant Growth Promotion ........ 12
1.4.8 PGPR as Biofertilizers ............................................................................ 14
1.5 Effect of Transgenic Plants in Rhizosphere Environment .................................. 15
1.5.1 Effect of Transgenic Plants on Soil Microorganisms ............................. 15
1.6 Diversity of Culturable and Non-Culturable Bacteria in the Rhizosphere ......... 18
1.6.1 16S rRNA Gene as a Tool for Studying Diversity of Culturable and Non-
Culturable Bacteria .......................................................................................... 18
1.6.2 Bacterial Diversity by Pyrosequencing Analysis of 16S rRNA Gene .... 19
1.6.3 Functional Genes for Bacterial Identification and Detection ................. 23

viii
1.6.4 nifH Metagenomics: A Tool to Study the Diversity of Diazotrophic
Bacteria ............................................................................................................ 23
1.6.5 Real Time PCR: A Gene Quantification Approach to Study the
Abundance of nif H and 16s rRNA Gene ........................................................ 24
2. Materials and Methods ............................................................................................. 27
2.1 Isolation of Bacteria from the Rhizosphere of Cotton and Wheat ...................... 27
2.1.1 Isolation of Diazotrophic Bacteria by Enrichment Culture Technique .. 25
2.2 Morphological Characterization of Bacteria ....................................................... 28
2.2.1 Colony and Cell Morphology ................................................................. 28
2.2.2 Culture Preservation................................................................................ 28
2.3 Phosphorus Solubilization .................................................................................. 28
2.3.1 Qualitative Assay for Phosphorus Solubilization by Bacteria ................ 28
2.3.2 Quantitative Estimation of Phosphate Solubilization by Bacteria .......... 28
2.3.3 Extraction and Quantification of Organic Acids Produced By Bacteria in
Pikovskaya Medium......................................................................................... 29
2.4 Indole Acetic Acid Production by Bacterial Isolates .......................................... 31
2.4.1 Colorimetric Estimation of IAA by Salkowski's Reaction (Spot Test) .. 31
2.4.2 Quantification of IAA Production .......................................................... 30
2.5 Identification of Bacterial Isolates by 16R rRNA Gene Sequence Analysis ...... 31
2.5.1 DNA Extraction from Pure Cultures of Bacterial Isolates.................... 31
2.5.2 Identification of Bacterial Isolates .......................................................... 31
2.6 Plant Inoculation Experiments ............................................................................ 32
2.6.1 Soil Analysis and Plant Material............................................................. 32
2.6.2 Bacterial Inoculum Preparation .............................................................. 32
2.6.3 Quick Screening of Bacterial Isolates in Sterilized Sand ....................... 32
2.6.4 Bacterial Inoculation of Cotton and Wheat Plants Grown In Earthen Pots
.......................................................................................................................... 33
2.6.5 Bacterial Inoculation of Wheat Plants Grown in Micro-Plots ................ 33
2.6.6 Statistical Analysis .................................................................................. 34
2.7 Estimation of Bacterial Population ..................................................................... 34
2.7.1 Bacterial Population by Counting Colony Forming Units (cfu/g of soil)
.......................................................................................................................... 34
2.7.2 Bacterial Population by Counting Most Probable Number (MPN) ........ 34
2.7.3 Real Time PCR ....................................................................................... 34
2.8 Extraction and Quantification of Root Exudates from the Rhizosphere ............. 35
2.9 Diversity of Diazotrophic Bacteria in the Rhizosphere of Transgenic and Non-
transgenic Plants of Cotton and Wheat .................................................................. 36

ix
2.9.1 PCR Amplification of nifH Gene from Soil DNA .................................. 36
2.9.2 Cloning of nifH Gene and Sequencing Reactions................................... 36
2.9.3 Phylogenetic Analysis ............................................................................. 37
2.10 Bacterial Diversity in the Rhizosphere of AVP1 Transgenic Cotton and Wheat
by Pyrosequencing Analysis .................................................................................. 37
2.10.1 16S rRNA Gene Amplification for Pyrosequencing ............................ 37
2.10.2 Analysis of the Pyrosequencing Data ................................................... 37
3. Results ...................................................................................................................... 39
3.1 Isolation of Bacteria from the Rhizosphere of AVP1 Transgenic Cotton ........... 39
3.2 Isolation of Bacteria from the Rhizosphere of AVP1 Transgenic Wheat ........... 39
3.3 Identification of Bacterial Isolates by 16S rRNA Gene Sequence Analysis ...... 42
3.4 Quantification of IAA Production by Bacterial Isolates ..................................... 51
3.5 Phosphate Solubilization ..................................................................................... 54
3.5.1 Qualitative Assay for Phosphate Solubilization by Bacterial Strains ..... 54
3.5.2 Quantitative Assay for Phosphate Solubilization by Bacterial Strains ... 54
3.6 Quantification of Organic Acid Production by Bacteria in Pikovskaya Medium
Used for Studying Phosphate Solubilization ......................................................... 55
3.7 Bacterial Inoculation of Cotton Plants ................................................................ 60
3.7.1 Experiment 1 (year 2009) ....................................................................... 60
3.7.2 Experiment 2 (Year 2010) .................................................................... 64
3.7.3 Experiment 3 (year 2011) ....................................................................... 67
3.8 Bacterial Inoculation of Wheat Plants ................................................................ 71
3.8.1 Experiment 1 (year 2009) ....................................................................... 71
3.8.2 Experiment 2 (year 2011-2012) .............................................................. 75
3.8.3 Experiment 3 (2012-2013) ...................................................................... 78
3.9 Bacterial Population ............................................................................................ 81
3.9.1 Real Time PCR Quantification of 16S rRNA and nif H genes from
Rhizospheric Soil ............................................................................................. 84
3.9.2 Detection of Root Exudates in the Rhizosphere of AVP1 Transgenic
Cotton and Wheat ............................................................................................ 86
3.10 Diversity of Diazotrophic Bacteria Determined by PCR Amplification of
Partial nifH gene from Soil DNA ........................................................................ 89
3.11 Bacterial Diversity by Pyrosequencing of 16S rRNA Gene. .......................... 105
3.11.1 16S rRNA Gene Sequences, Processing and Taxonomic Analysis .... 107
3.11.2 Bacterial Diversity in Cotton Rhizosphere ......................................... 107
3.11.3 Abundance of Bacterial Classes in Cotton Rhizosphere Soil ............. 107
3.11.4 Abundance of Bacterial Genera in the Rhizosphere of Cotton ........... 108

x
3.12 Bacterial Diversity in Wheat Rhizosphere ...................................................... 118
3.12.1 Abundance of Bacterial Classes in Wheat Rhizosphere Soil .............. 118
3.12.2 Abundance of Bacterial Genera in the Rhizosphere Wheat ................ 118
4. Discussion .............................................................................................................. 128
4.1 Conclusion and Future Perspectives ................................................................. 144
5. References .............................................................................................................. 145

xi
List of Figures
Figure 1-1 The role of intracellular plant growth promoting rhizobacteria (iPGPR)
and extracellular plant growth promoting rhizobacteria (ePGPR) in soil
ecosystem. .............................................................................................. 6
Figure 1-2 A sketch of nitrogen cycle showing the conversion of atmospheric
nitrogen into available forms. ................................................................ 7
Figure 1-3 Schematic representation of 16S rRNA gene annotated with variable
regions (V1 to V9) of 16S rRNA ......................................................... 21
Figure 1-4 16S rRNA gene with three distinct variable regions and primers ....... 21
Figure 1-5 Schematic representation of progress of enzymatic reaction in
pyrosequencing .................................................................................... 22
Figure 3-6 Isolation of bacteria on nutrient agar medium by serial dilution method
.............................................................................................................. 39
Figure 3-7 Genomic DNA extracted from bacterial isolates. ................................ 43
Figure 3-8 16S rRNA gene amplified from bacterial isolates. .............................. 43
Figure 3-9 Phylogenetic tree showing the phylogenetic relationship of different
strains of genus Bacillus and Paenibacillus. ........................................ 46
Figure 3-10 Phylogenetic tree showing the phylogenetic relationship of the
Brevibacillus strains. .......................................................................... 47
Figure 3-11 Phylogenetic tree showing the phylogenetic relationship of the
Arthrobacter strain ............................................................................... 48
Figure 3-13 Phylogenetic tree showing the phylogenetic relationship of Azospirillum
strain ..................................................................................................... 50
Figure 3-16 Plate assay for detection of phosphorus solubilization by bacterial
isolates on Pikovskaya medium supplemented with insoluble tri-calcium
phosphate (TCP) .................................................................................. 55
Figure 3-17: Organic acid production (µg/mL) by bacterial isolates in pure culture.
Bacterial cultures were grown for two weeks in Pikovskaya medium
containing insoluble tri-calcium phosphate. ........................................ 59
Figure 3-18 Bacterial inoculation experiments on cotton plants conducted in
different years in growth room. ........................................................... 60
Figure 3-19 Effect of bacterial inoculation on growth of cotton plants (transgenic and
non-transgenic) .................................................................................... 61
Figure 3-20 Effect of bacterial inoculation on root and shoot dry weights of
transgenic and non-transgenic plants ................................................... 63
Figure 3-21 AVP1 transgenic cotton plants grown under controlled conditions in
earthen pots .......................................................................................... 65

xii
Figure 3-22 Effect of bacterial inoculation on shoot dry weight, and yield (lint+seed)
of transgenic and non-transgenic plants ............................................... 67
Figure 3-23 Bacterial inoculation of AVP1 transgenic and non–transgenic cotton
grown under controlled conditions in earthen pots .............................. 68
Figure 3-24 Effect of bacterial inoculation on root dry weight, and yield (lint+seed)
of transgenic and non-transgenic plants ............................................... 70
Figure 3-25 Bacterial inoculation experiments on wheat plants conducted in different
years ..................................................................................................... 71
Figure 3-26 Effect of bacterial inoculation on growth of AVP1 transgenic wheat
plants grown in jars filled with sterilized sand .................................... 72
Figure 3-27 Effect of bacterial inoculation on shoot dry weight, and root dry weight
of transgenic and non-transgenic wheat plants .................................... 74
Figure 3-28 Effect of bacterial inoculation on growth of AVP1 transgenic and non-
transgenic wheat grown in micro-plots under natural conditions. ....... 75
Figure 3-29 transgenic and non-transgenic wheat plants grown in micro-plots ...... 77
Figure 3-30 Effect of bacterial inoculation on growth of AVP1 transgenic and non-
transgenic wheat grown in micro-plots under natural conditions. ....... 78
Figure 3-31 Effect of bacterial inoculation on straw weight, and grain weight of
transgenic and non-transgenic wheat plants ........................................ 80
Figure 3-32 Bacterial population (log cfu/g soil) on nutrient agar in the rhizosphere
of transgenic and non-transgenic cotton at 30, 60 and 90 days after
sowing (DAS) ...................................................................................... 82
Figure 3-33 Bacterial population (log cfu/g soil) on nutrient agar in the rhizosphere
of transgenic and non-transgenic wheat at 30, 60 and 90 days after
sowing (DAS) ...................................................................................... 82
Figure 3-34 Bacterial population of diazotrophs (log MPN/g soil in NFM) in the
rhizosphere of transgenic and non-transgenic cotton at 30, 60 and 90
days after sowing (DAS) ...................................................................... 83
Figure 3-35 Bacterial population diazotrophs (log MPN/g soil in NFM) in the
rhizosphere of transgenic and non-transgenic wheat at 30, 60 and 90
days after sowing (DAS) ...................................................................... 83
Figure 3-36 Real time quantification of 16S rRNA and nifH gene from rhizosphere
of AVP1 transgenic cotton and wheat .................................................. 86
Figure 3-37 Organic acid production in rhizosphere of AVP1 transgenic and non-
transgenic cotton and wheat. ................................................................ 88
Figure 3-38 Amplification of nifH gene from soil DNA extracted from AVP1
transgenic and non-transgenic cotton (A) and wheat (B) .................... 90
Figure 3-39 transgenic (A) and non-transgenic cotton (B). ..................................... 92
Figure 3-40 Distribution of diazotrophic bacteria in the rhizosphere of AVP1
transgenic (A) and non-transgenic wheat (B) ...................................... 98
Figure 3-41 Phylogenic tree constructed from nifH gene sequences retrieved from
AVP1 transgenic and non-transgenic cotton ...................................... 103

xiii
Figure 3-42 Phylogenic tree constructed from nifH gene sequences of AVP1
transgenic and non-transgenic wheat ................................................. 104
Figure 3-43 DNA extracted from the rhizosphere soil of cotton and wheat ........ 105
Figure 3-44 PCR amplification of 16S rRNA gene from the rhizosphere of AVP1-
transgenic and non-transgenic cotton (A) and wheat (B) using barcoded
primers ............................................................................................... 106
Figure 3-45 Abundance of bacterial phyla in the rhizosphere of AVP1 transgenic and
non-transgenic cotton. ........................................................................ 112
Figure 3-46 16S rRNA sequences belonging to different bacterial classes reterieved
from the rhizosphere of AVP1 transgenic and non-transgenic cotton 113
Figure 3-47 Bacterial genera detected in the rhizosphere of AVP1 transgenic and
non-transgenic cotton ......................................................................... 115
Figure 3-48 Abundance of important PGPR genera in the rhizosphere of AVP1
transgenic and non-transgenic cotton rhizosphere ............................. 117
Figure 3-49 Abundance of 16S rRNA sequences of different bacterial phyla in the
rhizosphere of AVP1 transgenic and non-transgenic wheat ............... 121
Figure 3-50 Abundance of 16S rRNA sequences belonging to different bacterial
classes dominant in wheat rhizosphere of AVP1 transgenic and non-
transgenic wheat................................................................................. 122
Figure 3-51 Bacterial genera detected only in the rhizosphere of AVP1 transgenic
and non-transgenic wheat .................................................................. 124
Figure 3-52 Abundance and comparison of PGPR detected in AVP1 transgenic and
non-transgenic rhizosphere of wheat. ................................................ 126

xiv
List of Tables
Table 1.1 Global area of biotech crops in mega countries in 2014 ........................ 3
Table 1.2 Categorization of PGPR on the bases of their action mechanism ......... 6
Table 1.3 Important nitrogen fixing bacteria residing in rhizosphere of different
crops plants .......................................................................................... 10
Table 1.4 Effect of transgenic plants on structure and functions of soil
microorganisms and their communities ............................................... 16
Table 1.5 Sequences of PCR primers for amplification of 16S rRNA gene ........ 22
Table 2.6 Primer sequence with titanium adopter sequence ................................ 38
Table 2.7 Soil samples with barcodes name and sequences ................................ 38
Table 3.8 Colony and cell morphology of the bacterial strains isolated from the
rhizosphere of AVP1 transgenic and non-transgenic cotton ................ 40
Table 3.9 Colony and cell morphology of the bacterial strains isolated from the
rhizosphere of AVP1 transgenic and non-transgenic wheat ................. 41
Table 3.10 Identification of bacterial isolates from rhizosphere of AVP1 transgenic
and non-transgenic cotton by 16S rRNA gene sequence analysis ....... 44
Table 3.11 Identification of bacterial isolates from rhizosphere of AVP1 transgenic
and non-transgenic wheat rhizosphere by 16S rRNA gene sequence
analysis ................................................................................................. 45
Table 3.12 Quantification of IAA produced by bacterial strains isolated from cotton
.............................................................................................................. 53
Table 3.13 Quantification of IAA produced by bacterial strains isolated from wheat
.............................................................................................................. 54
Table 3.14 Quantification of P solublization by bacterial isolates from cotton ..... 56
Table 3.15 Quantification of P solubilization by bacterial isolates from wheat .... 56
Table 3.16 Quantification of organic acid production (µg/mL) by bacterial isolates
in the growth medium used for P-solubilization .................................. 58
Table 3.17 Effect of bacterial inoculation on growth of AVP1 transgenic (A) and
non-transgenic cotton (B) grown in sterilized sand under controlled
conditions (year 2009) ......................................................................... 62
Table 3.18 Effect of bacterial inoculation on growth of AVP1 transgenic (A) and
non-transgenic cotton plants (B) grown in earthen pots under controlled
conditions (Year 2010). ....................................................................... 66
Table 3.19 Effect of bacterial inoculation on growth of AVP1 (A) transgenic and
non-transgenic cotton (B) grown in earthen pots under controlled
conditions ............................................................................................. 69

xv
Table 3.20 Effect of bacterial inoculation on growth of AVP1 transgenic (A) and
non-transgenic wheat (B) grown in sterilized sand under controlled
conditions ............................................................................................. 73
Table 3.21 Effect of PGPR strains on yield and growth parameters of transgenic (A)
and non-transgenic wheat (B) grown in micro-plots during 2011-2012
.............................................................................................................. 76
Table 3.22 Effect of PGPR strains on yield and growth parameters of transgenic (A)
and non-transgenic wheat (B) grown in micro plots during 2012-2013
.............................................................................................................. 79
Table 3.23 Relative gene abundance (copy number) of bacterial 16S rRNA and nif
H genes in the rhizospheric soil revealed by real time PCR ................ 85
Table 3.24 Detection of organic acids produced* as root exudates in rhizosphere of
AVP1 transgenic and non-transgenic cotton and wheat ....................... 87
Table 3.25 Diversity of diazotrophic bacterial sequences in the rhizosphere of AVP1
transgenic and non-transgenic cotton................................................... 91
Table 3.26 Identification of culturable diazotrophic bacterial sequences in the
rhizosphere of AVP1 transgenic cotton ................................................ 93
Table 3.27 dentification of culturable diazotrophic bacteria detected in the
rhizosphere of non-transgenic cotton ................................................... 94
Table 3.28 List of uncultured diazotrophic bacteria from AVP1 transgenic cotton
rhizosphere ........................................................................................... 95
Table 3.29 List of uncultured diazotrophic bacterial sequences from non-transgenic
cotton rhizosphere ................................................................................ 96
Table 3.30 Diversity of diazotrophic bacteria in the rhizosphere of AVP1 transgenic
and non-transgenic wheat .................................................................... 97
Table 3.31 Identification of culturable diazotrophic bacterial sequences in the
rhizosphere of AVP1 transgenic wheat ................................................ 99
Table 3.32 Identification of culturable diazotrophic bacteria detected in the
rhizosphere of non-transgenic wheat ................................................. 100
Table 3.33 List of uncultured diazotrophic bacteria from AVP1 transgenic wheat
rhizosphere ......................................................................................... 101
Table 3.34 List of uncultured diazotrophic bacteria from AVP1 transgenic wheat
rhizosphere ......................................................................................... 102
Table 3.35 16S rRNA sequences retrieved from rhizosphere of AVP1 transgenic
cotton and wheat with non-transgenic control ................................... 109
Table 3.36 Abundance of 16S rRNA sequences belonging to different phyla in the
rhizosphere of AVP1 transgenic and non-transgenic cotton .............. 108
Table 3.37 Abundance of 16S rRNA sequences of different bacterial genera (Top
50 genera) retrieved from transgenic and non-transgenic cotton ....... 113
Table 3.38 Bacterial genera detected only in the rhizosphere of transgenic cotton
............................................................................................................ 116

xvi
Table 3.39 Sequences of important PGPR genera detect in the rhizosphere of AVP1
transgenic and non-transgenic cotton................................................. 116
Table 3.40 Abundance of 16S rRNA sequences of different bacterial phyla in the
rhizosphere of AVP1 transgenic and non-transgenic wheat ............... 120
Table 3.41 Abundance of 16S rRNA sequences belonging to different bacterial
genera (Top 50 genera) retrieved from the rhizosphere of AVP1
transgenic and non-transgenic wheat ................................................. 122
Table 3.42 Bacterial genera detected only in the rhizosphere of transgenic wheat
............................................................................................................ 125
Table 3.43 Sequences of important PGPR genera detect in the rhizosphere of AVP1
transgenic and non-transgenic wheat ................................................. 125

xvii
Abstract
Present study was conducted to compare diversity of bacteria in the rhizosphere of
AVP1 transgenic cotton and AVP1 transgenic wheat with non-transgenic plants of both
the crops. Over-expression of the H+pyrophosphatase (H+PPase) AVP1 results in salt
and water stress tolerance. For studying the diversity of culturable bacteria, 12 strains
were isolated from cotton and 14 strains were purified from wheat and identified by
16S rRNA gene sequence analysis. After initial screening of the isolates on the bases
of phytohormone production and phosphate solubilization in pure culture as well as
plant growth promotion in short term experiments in sand culture, the efficient strains
were used as inoculants for plants grown in non-sterilized soil. Risk assessment studies
indicated no significant difference of bacterial populations in the rhizosphere of
transgenic and non-transgenic plants as determined by log cfu/g soil, MPN and copy
number of 16S rRNA and nifH genes. However, bacterial populations were variable at
different plant growth stages of both cotton and wheat. Using soil DNA, diversity of
diazotrophs and rhizospheric communities was assessed by sequence analysis of PCR-
amplified nifH gene and 16S rRNA genes, respectively. nifH sequences belonging to
well-known diazotrophic genera i.e Anabaena, Azospirillum, Bradyrhizobium and
Pseudomonas were abundant and common in the rhizosphere of AVP1 transgenic and
non-transgenic plants of cotton and wheat. A fraction of uncultured diazotrophs were
also detected in the rhizosphere of cotton and wheat. From the rhizosphere of cotton
(transgenic and non-transgenic) total 190249 sequences of 16S rRNA gene were
retrieved by pyrosequencing analysis which indicated 127747 sequences of bacteria,
8128 sequences of Archaea and 22964 sequences of unclassified bacteria. All the 19
bacterial phyla detected on the basis of 16S rRNA gene sequencing were represented
in the rhizosphere of both transgenic and non-transgenic cotton plants. From wheat
rhizosphere total 156282 sequences were obtained by pyrosequencing analysis of 16S
rRNA gene which indicated 128006 sequences of bacteria, 7928 sequences of Archea
and 21568 sequences of unclassified bacteria. All the 18 bacterial phyla detected on the
basis of 16S rRNA gene sequencing were represented in the rhizosphere of both the

xviii
transgenic and the non-transgenic wheat. In the present study comparison of the number
of sequences retrieved from transgenic and non-transgenic plants of cotton and wheat
indicated that all major bacterial groups (phyla) were represented in the rhizosphere of
both type of plants (transgenic and non-transgenic) and point to safe use of transgenic
plants.

xix
List of Publications and Patents
Journal Publications
Muhammad Arshad, Muhammad Arshad, Johan Leveaue, Shaheen Asad, Asma
Imran, Muhammad Sajjad Mirza. 2015. Culturable Bacterial Population in
the Rhizosphere of AVP1 Transgenic and non-transgenic Cotton and
Growth Promotion by Inoculated Strains of Arthrobacter, Azospirillum and
Brevibacillus. (JCR 2014) (ISSN: 1018-7081)

xx
List of Abbreviations and Symbols
ºC Degree centigrade
µL Micro litre
µm Micro meter
10X 10 times
ABA
ACC-
deaminase
Abscisic acid
1-aminocyclopropane-1-carboxylate deaminase
ANOVA Analysis of variance
ARA Acetylene reduction assay
ATP Adenosine triphosphate
AVP1 Vacuolar proton-pyrophosphatase from Arabidopsis
BLAST Basic local alignment search tool
BNF Biological nitrogen fixation
Bt cotton Bacillus thuringiensis transgenic cotton
cfu Colony forming units
cm Centimeter
CRD
DAP
Completely randomized design
Di-ammonium phosphate
DAS Days after sowing
GA Gluconic acid
GFP Green fluorescent protein
GMOs Genetically modified organisms
GMPs Genetically modified plants
HCN Hydrogen cyanide
HPLC High performance liquid chromatography
IAA Indole-3-acitic acid
LB Luria Bertani

xxi
MPN Most probable number
N Nitrogen
15N Isotope of nitrogen with atomic
N2 Atmospheric nitrogen
NCBI National Center for Biotechnology Information
NFM Nitrogen free malate
NTC Non-transgenic cotton
NTW Non-transgenic wheat
P Phosphorus
PGPR Plant growth promoting rhizobacteria
PSB Phosphate solubilizing bacteria
RCBD Randomized complete block design
TC Transgenic cotton
TCP Tri-calcium phosphate
TW Transgenic wheat

1. Introduction
1.1 Genetically Modified Crops
Genetic modification of plants, microbes and animals to incorporate useful traits is a
powerful technology for the development of sustainable agricultural systems.
Genetically modified plants (GMPs) with a wide variety of traits have been developed.
Most GMPs developed to date can be grouped into eight main categories: (i) resistance
to herbicides, (ii) resistance to pests, (iii) resistance to pathogens, (iv) resistance to
environmental stress, (v) altered root exudates, (vi) plants with altered composition,
(vii) ability to produce pharmaceutical or industrial compounds and (viii) elimination
of pollutants [1]. The first genetically modified plant was produced in 1982, using an
antibiotic-resistant tobacco plants [2]. The first field trials of genetically engineered
plants occurred in France and the USA in 1986, when herbicide resistant tobacco plants
were engineered [3]. In 1987, Plant Genetic Systems (Ghent, Belgium) was the first
company to develop genetically engineered (tobacco) plants with insect tolerance by
expressing genes encoding for insecticidal proteins from Bacillus thuringiensis (Bt) [4].
The first genetically modified crop approved for sale in the U.S., in 1994, was
the Flavr Savr Tomato, which was modified for its longer shelf life. GM technology has
addressed some of the most serious concerns of world agriculture and GM technology
can be applied to some of the specific problems of agriculture, indicating the potential
for benefits. Biotechnology has revolutionized crop improvement by producing GM
crops with enhanced availability and utilization of important traits. Transgenic crops
containing insect-resistance genes from Bacillus thuringiensis have made it possible to
reduce significantly the amount of insecticide applied on cotton [4]. Other insecticidal
proteins have been discovered including lectins, protease inhibitors, antibodies, wasp
and spider toxins, microbial insecticides and insect peptide hormones [3].
One of the major technologies that led to the “Green Revolution” was the
development of high-yielding semi-dwarf wheat varieties. Crops have been developed
that have an inbuilt resistance to biotic and abiotic stress i.e rice yellow mottle virus
(RYMV) devastates rice in Africa by destroying the majority of the crop. “Genetic

1. Introduction
2
immunization” was done by creating transgenic rice plants that were resistant to RYMV
[98]. Numerous other examples could be given i.e blight resistant potatoes and rice
bacterial leaf blight, plants modified to overproduce citric acid in roots and provide
better tolerance to aluminum in acid soils. The transgenic rice exhibiting an increased
production of beta carotene as a precursor to vitamin A and may be a useful tool to help
treat the problem of vitamin A deficiency in young children living in the tropics. Plant
genetic engineering technology is now being widely used for “biopharming”, or
production of pharmaceuticals in plants. (Text on crop plants with foreign has also been
given in Table 1.4).
1n 2010, 8 insect resistant Bt. and 1 hybrid cotton varieties were officially
approved by government of Pakistan. In Pakistan 2014 was the fifth year of
commercialization of Bt. crop with the area 2.9 million hectares. With the increase in
land area under transgenic crops (Table 1.1) concerns have been raised over the
potential detrimental effects of genetically modified plants on human health,
environment and non-target organisms including soil microbial communities. The
major concerns are the possibility of creating invasive plant species, the unintended
consequences of transgene flow to indigenous plants and microorganisms and
development of ‘super’ pests [6].
1.2 Use of AVP1 Gene to Develop Transgenic Plants
Salinity limits the plant growth affecting 20% of the world’s irrigated lands [7].The
harmful effects of salt on plants are a result of (i) water deficit that results from the
relatively high solute concentrations in soil and (ii) excessive sodium (Na+)
concentrations in the cytoplasm. Excessive Na+ in the cytoplasm changes ion ratios that
disturbs critical biochemical processes and also increases plasma membrane injury [9,
10]. Accumulation of compatible solutes and reduction of sodium ions in the cytoplasm
are two common mechanisms in plants to deal with the injury. Plants reduce excessive
Na+ in the cytoplasm by (i) excluding Na+ from cells using the Na+/H+ antiporter located
in the plasma membrane; and by pumping Na+ into vacuoles using Na+/H+ antiporter
located in the tonoplast [11].

1. Introduction
3
Table 1.1 Global area of biotech crops in mega countries in 2014 [8].
The compartmentalization of Na+ into vacuoles provides an efficient mechanism for
avoiding the toxic effects of Na+ in the cytosol. The transport of Na+ into vacuoles is
mediated by vacuolar Na+ /H+ antiporters that are driven by the electrochemical
gradient of protons. The proton-motive force generated by the vacuolar ATPase (V-
ATPase) and vacuolar pyrophosphatase (V-PPase) can drive secondary transporters,
such as the Na+/H+ antiporter and Ca2+/H+ antiporter, as well as organic acids, sugars,
and other compound transporters to maintain cell turgor [12]. The vacuolar H+-PPase
is a single subunit protein located in the vacuolar membrane [13]. It pumps H+ from the
cytoplasm into vacuoles with P Pi-dependent H+ transport activity. Theoretically, over-
expression of H+-PPase should enhance the ability to form the pH gradient between the
cytoplasm and vacuoles, resulting in a stronger proton-motive force for the Na+/H+
antiporter, Ca2+/H+ antiporter, and other secondary transporters. The accumulation of
cations, such as Na+, in vacuoles could increase the osmotic pressure of plants, while
reducing the toxic effects of these cations [14].
The transcription and translation of AVP1 and P-ATPases (Arabidopsis H+-
ATPase’s, AHAs), normally expressed in roots showed that low Pi increases transcript
and protein abundance of AVP1 and P-ATPase in Arabidopsis [14, 15]. Another
Rank Country Area (million
hectares)
Biotech Crops
1. USA 73.1 Maize, soybean, cotton, canola,
sugar beet, alfalfa, papaya, squash
2. Brazil 42.2 Soybean, maize, cotton
3. Argentina 24.3 Soybean, maize, cotton
4. Canada 11.6 Canola, maize, soybean, sugar beet
5. India 11.6 Cotton
6. China 3.9 Cotton, papaya, poplar, tomato,
sweet pepper
7. Paraguay 3.9 Soybean, maize, cotton
8. Pakistan 2.9 Cotton
9. South Africa 2.7 Maize, soybean, cotton
10. Uruguay 1.6 Soybean, maize

1. Introduction
4
phenotype of AVP1 i.e AtAVP1OX plants exhibited enhanced growth, enhanced
rhizosphere acidification, larger shoot formation and Pi uptake when grown on solid
Pi-deficient medium. Roots of different lines i.e AtAVP1OX, LeAVP1DOX and
OsAVP1DOX have higher K+ contents and thus exude greater amounts of organic acids
when compared with control plants. Transgenic tomatoes over-expressing the E229D
gain-of-function mutant (AVP1D) of the Arabidopsis H+-pyrophosphatase
(LeAVP1DOX) develop more robust root systems and are resistant to imposed soil
water deficits [16].
Over-expression of the H+pyrophosphatase (H+PPase) AVP1 resulted in salt and
water stress tolerant Arabidopsis plants [14]. The tolerance was initially explained by
an enhanced uptake of ions into their vacuoles. Presumably, the greater AVP1 activity
in vacuolar membranes provides increased vacuolar H+ to drive the secondary active
uptake of toxic (i.e. sodium) and nontoxic ions into the vacuole. The resulting decline
in vacuolar osmotic potential may trigger water uptake, permitting plants to survive
under conditions of low soil water potentials [15]. Significantly, further
characterization of these AVP1 overexpressing plants revealed an enhancement of their
root development, with obvious implications for their ability to withstand drought [16].
These results suggest that the H+PPase AVP1 is a potential target for genetic
engineering of root systems in agriculturally important crop plants.
1.3 Bacterial Diversity in Rhizosphere of Genetically
Modified Plants
Rhizosphere is the rooting zone of plants and includes the roots, soil attached to the
roots, and the adjacent soil under the influence of the roots [17]. Microorganisms are
also considered as important component of the rhizosphere and contribute to ecological
fitness of their host plant. Soil microbes are involved in important process that might
occur in the rhizosphere including plant growth promotion, plant protection,
pathogenesis, production of antibiotics, cycling of carbon, nitrogen and sulfur [18, 19].
Bacteria represent one of the three domains in the phylogenetic tree of life comprised
of Archaea, Bacteria and Eukarya [20]. Bacterial diversity generally refers to the
genetic diversity which is the amount and distribution of genetic information within the

1. Introduction
5
bacterial communities. Total number of species present (species richness) and
distribution of individuals among the species (evenness) are the functions of diversity.
1.4 Plant Growth Promoting Rhizobacteria (PGPR)
The term ‘rhizobacteria’ is used to describe the soil bacteria (PGPR) that competitively
colonize plants and stimulate growth by utilizing plant beneficial traits and by reducing
plant diseases [21]. PGPR constitute the key part of rhizosphere biota by successfully
establishing in the rhizosphere due to their adaptability in a wide variety of
environments, faster growth, and their ability to metabolize a wide range of compounds
[22]. Most rhizospheric bacteria establish an inoffensive interaction with the host plants
exhibiting no visible effect on the growth and overall physiology of the host [23]. In
negative interactions, the phytopathogenic rhizobacteria produce phytotoxic substances
such as hydrogen cyanide or ethylene and negatively influence the growth and
physiology of the plants. PGPRs on the other hand exert a positive effect on plant
growth by diverse mechanisms such as solubilization of nutrients, nitrogen fixation,
and production of phytohormones [24-26]. PGPR can be further classified into
extracellular plant growth promoting rhizobacteria (ePGPR), present in the rhizosphere,
on the rhizoplane or in the spaces between the cells of root cortex and intracellular plant
growth promoting rhizobacteria (iPGPR) generally located inside the specialized
nodular structures of root cells [27]. A large number of PGPR like Azospirillum,
Azotobacter, Bacillus, Enterobacter, Pseudomonas, Klebsiella and Paenibacillus
have been isolated from rhizosphere of various crops and their plant growth promoting
traits have been studied [28-32].
1.4.1 Mode of Action of PGPR
PGPR promote plant growth directly by utilizing mechanisms like biological nitrogen
fixation, phytohormone production e.g auxins, mineral solubilization or indirectly by
employing mechanisms basically related to biocontrol and include antibiotic
production, siderophores production to chelate available Fe in the rhizosphere,
synthesis of extracellular enzymes to hydrolyze the cell wall of fungal pathogens and
competition for niches within the rhizosphere [33, 34]. On the bases of their action
mechanism application of PGPR can be generalized into three broad categories i.e.
Biofertilizers, Biopesticides and Phytostimulators (Table 1.2).

1. Introduction
6
Table 1.2 Categorization of PGPR on the bases of their action mechanism
Figure 1-1 The role of intracellular plant growth promoting rhizobacteria
(iPGPR) and extracellular plant growth promoting rhizobacteria (ePGPR) in soil
ecosystem.
1.4.2 Nitrogen Fixation
Nitrogen (N) is one of the major plant nutrients, required for cellular synthesis of vital
biomolecules like enzymes, proteins, nucleic acids (DNA and RNA) and chlorophyll
[38]. More than 78% of nitrogen is present in the atmosphere in the gaseous N2 form
which is unavailable to the plants. Plants utilize only fixed forms of nitrogen e.g
ammonium (NH4+) and nitrate (NO3
-) for growth. In the biogeochemical nitrogen cycle
PGPR category Mechanism of action Reference
Biofertilizers Biological nitrogen fixation. Solubilization
of Phosphorus. Production of plant growth
regulators e.g (IAA).
[32, 35]
Biopesticides Promote plant growth indirectly by
controlling growth of phyto-pathogens.
Production of antibiotics, siderophores,
HCN, hydrolytic enzymes acquired and
induced systemic resistance.
[32,35]
Phytostimulators Production of phytohormones such as
indole acetic acid, gibberellic acid,
cytokinins and ethylene
[36, 37]

1. Introduction
7
the conversion of atmospheric N2 into NH4+ ammonium ions is driven by nitrogen
fixation process, which in turn is converted into nitrate (NO3-) through nitrification
process and finally returns to the atmosphere in gaseous nitrogen oxides and nitrogen
gas by denitrification process (Figure 1-2). The maintenance and replenishing of fixed
N as ammonium is essentially required for the formation of N containing compounds
in the living cells of all life forms.
The conversion of the atmospheric nitrogen into available forms takes place by
(i) Industrial nitrogen fixation at high temperature and pressure to produce chemical N
fertilizers. (ii) Conversion of N2 into oxides of nitrogen in the atmosphere by natural
lightening (iii) Biological nitrogen fixation (BNF) i.e the conversion of N2 to NH4+ by
diazotrophic prokaryotes.
Figure 1-2 A sketch of nitrogen cycle showing the conversion of atmospheric
nitrogen into available forms.

1. Introduction
8
1.4.3 Biological Nitrogen Fixation (BNF)
In biological nitrogen fixation process, gaseous nitrogen from the atmosphere is
reduced to ammonia, by the enzyme nitrogenase [39]. The process of molecular
nitrogen fixation is found in phylogenetically diverse groups of prokaryotic organisms,
the bacteria and archea, including aerobic, microaerophilic, facultative, and strictly
anaerobic microorganisms [40]. This biochemical reaction of dinitrogen conversion
into ammonium is highly energy expensive and it requires a significant amount of
reducing power, along with energy from ATP [41].
An enzyme complex called ‘nitrogenase’ found in prokaryotes catalyzes the
conversion of atmospheric dinitrogen (N2) in biological fixation process. Nitrogenase
enzyme is highly conserved in its role and structure among diverse prokaryotes [42].
The nitrogenase system is composed of two subunits of metallo-proteins. The subunit I
or dinitrogenase is larger component which performs nitrogen reduction and also
referred as MoFe-protein or component I. The molecular weight of componentI is 220
to 250 kD. Subunit II or component II, is the smaller component and known as
dinitrogenase reductase with molecular weight of 60-70 kD. Dinitrogenase reductase
pairs ATP hydrolysis to inter-protein electron transport and is composed of two similar
subunits encoded by nifH gene. Among nitrogen fixers, nif genes differ in their
structural organization. For example in gamma and alpha proteobacteria nifHDK
operon is responsible for transcription. In slow growing symbiotic associations nifH
and nifDK are separate operons by which transcription is associated [42].
NifH genes have been highly conserved through evolution [43]. This great
conservation of nifH genes provides a valuable molecular tools to examine
phylogenetic distributions and biological nitrogen fixation in the environment [44,
45]. The nifH gene has one of the largest non-ribosomal database sequences of diverse
culturable as well as uncultivated microorganisms from the environment [42]. The
nifH gene provides phylogenetic uniqueness that allows construction of trees of
relatedness for diazotrophic organisms. In order to analyze biological processes in
specific ecosystems without cultivation, nifH gene markers have been employed. nifH
N2 + 8e- + 16 ATP + 16 H2O 2NH3 + H2 +16 ADP + 16 Pi + 8H+
Nitrogenase

1. Introduction
9
genes have been amplified by PCR from environmental samples as well as from pure
cultures [45-47].
1.4.4 Diversity of Diazotrophic Bacteria
Several diazotrophic bacteria occur as free-living bacteria in the environment while
others (e.g. Rhizobium, Frankia) can induce root-nodules on legumes and non-
legumes and live as symbiotic entophytes. The common example of symbiotic
relationship between bacteria and plant is Rhizobium-legume symbiosis. A special
structure called ‘nodule’ is induced on roots of legumes (chickpea, lentil and other
plants) by rhizobia. Free-living diazotrophic bacteria have been isolated from the
rhizosphere, rhizoplane and interior of the roots of grasses, cereals and food crops like
wheat, rice, maize and sugarcane [48-50]. Free-living nitrogen fixing bacteria are
known to colonize rhizosphere of important crops and belonged to different genera
(Table 1.3).
1.4.5 The Domain Archea
Archaea are the third domain in the phylogenetic tree of life alongside Bacteria and
Eucarya [51, 52] and were considered as an assemblage of extremophilic organisms
without a major role in the earth ecosystems. The numbers of known taxa within the
Archaea are expanding and include diazotrophs as well. The Archaea are distributed
over two main phylogenetic branches based on 16S rRNA sequence comparisons, the
Euryarchaeota and the Crenarchaeota [52]. The Euryarchaeota contain the
methanogens, the halophiles, and some extreme thermophiles, while the Crenarchaeota
contain most of the extreme thermophiles. Within the Archaea, nitrogen fixation has
been found only in the methanogenic Euryarchaeota, however, within the methanogens,
nitrogen fixation is widespread [53]. M. thermolithotrophicus is the only organism
demonstrated to fix nitrogen at 60°C or above. In the Methanobacteriales, nitrogen
fixation has been demonstrated for Methanobacterium bryantii [54]. Diazotrophic
growth, 15N2 incorporation, or acetylene reduction has been reported for a number of
methanogens. The discovery of Archaea in oceanic plankton has triggered a huge
number of studies [55, 56]. These studies presented that Archaea are abundant and
diverse group of microorganisms in whole biosphere with a significant impact on
nutrient cycling [57]. Evidence for autotrophic growth of some phylotypes of

1. Introduction
10
Thaumarchaeota has been provided in soil [58]. Cultivation of novel archaeal strains
and culture-independent techniques in molecular biology have played an instrumental
role for recognizing and characterizing novel archaeal metabolisms and for estimating
their environmental impact that archaea are important players in carbon and nitrogen
cycling.
Table 1.3 Important nitrogen fixing bacteria residing in rhizosphere of different
crops plants
Genus PGPR activity Host References
IAA N2
fixation
P
solubil
ization
Azospirillum √ √ wheat, rice, maize,
sugarcane and other
graminous plants
[59]
Acetobacter
√ Sugarcane, sugarcane,
cotton wood
[60]
Acinetobacter √ sugarcane, cowpea [61]
Achromobacter √ √ sugarcane [62]
Azotobacter √ √ √ rice [63]
Agrobacterium √ √ sugarcane [64]
Alcaligenes √ wetland rice [65]
Azoarcus √ kallar grass [66]
Bacillus √ √ rice,cowpea, mangrove [67]
Burkholderia √ √ rice, sugarcane, grasses [68]
Enterobacter √ √ √ rice, sugarcane, grasses [69]
Paenibacillus √ √ rice,cowpea, mangrove
[70]
Pseudomonas √ √ √ wheat, sugarcane,
grasses, mangrove
[64]
Rhizobium √ √ legumes, peas, cow pea, [71]
Zoogloea √ √ √ kallar grass [66]

1. Introduction
11
1.4.6 Phosphorus Mineralization by Microbes for Plant Growth
Promotion
Phosphorus (P) is the second most important element after nitrogen that plant requires.
This element being structural component of the nucleic acids, proteins and
phospholipids is involved in important biological processes such as cell division,
photosynthesis, biological oxidations and transfer of energy and nutrient uptake by
plants [72]. Primarily soil phosphorus originates from weathering of soil minerals such
as apatite. Addition of P in soil also occurs from fertilizer application, agricultural
waste, plant residues, or bio-solids. Orthophosphate ions (HPO4-2 and H2 PO4 -) are
produced when apatite breaks down, organic residues are decomposed, or fertilizer P
sources are dissolved.
All phosphorus taken up by plants comes from phosphorus dissolved in the soil
solution. The amount of soluble phosphorus in the soil solution is very low, as most of
phosphorus is insoluble and thus unavailable to plants. The type of P-bearing minerals
that form in soil is highly dependent on soil pH. Soluble P, originating from any source,
reacts very strongly with Fe and Al to form insoluble Fe and Al phosphates in acidic
soils and with Ca to form insoluble Ca phosphates in alkaline soils. Recent interest
generated in finding P solubilizing microorganisms that solubilize the phosphate
present in soil is mainly due to the rising costs of phosphorus fertilizers. Therefore,
phosphate solubilizing rhizospheric bacteria offer a very attractive opportunity to
increase the bioavailability of phosphorus to plants [73]. Several reports have examined
the ability of different bacterial species to solubilize insoluble inorganic phosphate
compounds [74].
Plants and microorganisms use phosphatase enzyme to mineralize (hydrolyze)
organic P for uptake. Increased activity of phosphatases occurs in response to P
deficiency as part of P starvation responses [75]. Solubilization of phosphate due to the
production of organic acids by microbes is considered a major mechanism for P
solubilization [76]. The mineral phosphate solubilizing bacteria use root exudates e.g.
sugars as carbon source to form organic acids. A number of organic acids like malic,
glyoxalic, succinic, fumaric, tartaric, alpha keto butyric, oxalic, citric, 2-ketogluconic
and gluconic acid have been detected in cell-free supernatant growth medium of
bacteria [77]. The amount and type of the organic acids produced varies with the

1. Introduction
12
microorganism. Organic acids produced by phosphate solubilizing bacteria are used to
lower pH of the medium. Due to existence of equilibrium between anions and protons,
the protons are consumed in the dissolution of the phosphorus [78].
Chelation of cations has also been implicated in phosphate solubilizing.
Chelation involves the formation of two or more coordinate bonds between an anionic
or polar molecule and a cation, resulting in a ring structure complex [79]. Organic acid
anions, with oxygen containing hydroxyl and carboxyl groups, have the ability to form
stable complexes with cations such as Al3+, Ca2+, Fe2+ and Fe3+ and, that are often
bound with phosphate in poorly solubilized forms [80].
1.4.7 Phytohormone Production by PGPR for Plant Growth
Promotion
Phytohormone production is a well-established phenomenon that contributes to
plant growth promotion by PGPR [32, 81]. Phytohormones are small singling
molecules used by the plants for their growth and development in variable
developmental and environmental conditions. Plant growth promoting abilities of
PGPR are often related to the production of these growth regulators [82]. Several
bacterial genera have been reported for the production of phytohormones (IAA)
including Azospirillum, Acetobacter, Achromobacter, Azotobacter, Bacillus,
Enterobacter, Pseudomonas, Rhizobium, and Xanthomonas [83, 84]. Phenyl acetic acid
(PAA) is an auxin-like molecule, derived from amino acid metabolism and is known
for its weak auxin activity. Based on its aromatic structure, it was speculated that PAA
might be derived from phenylalanine. Azospirillum have been reported for the
production phytohormones, like indole-3-acetic acid and more specifically PAA [57].
PAA could only be identified in supernatant extracts from Azospirillum brasilense
cultures grown in LB medium PAA has been demonstrated to display growth-inhibitory
activity towards Gram-negative bacteria (including P. syringae and E. coli), Gram-
positive bacteria (including Bacillus subtilis and Staphylococcus aureus)
[364,365,367]. A high degree of similarity between IAA biosynthesis pathways in
plants and bacteria has been noticed and its role in plant-microbe interaction has been
studied. Tryptophan has been identified as a main precursor for IAA biosynthesis
pathways in bacteria. Five different pathways for biosynthesis of IAA have been
identified.

1. Introduction
13
(i) Indole-3-acetamide (IAM) pathway
(ii) Indole-3-pyruvate (IPyA)
(i) Tryptamine (TAM) pathway
(ii) Tryptophan side-chain oxidase (TSO)
(iii) Indole-3-acetonitrile (IAN)
The best characterized pathway in bacteria is IAM pathway with two step
reactions. In first step tryptophan-2-monooxygenase (iaaM) converts trptophan into
IAM. In the second step IAM hydrolase encoded by iaaH converts IAM into indole-3-
acetic acid (1AA). The genes iaaM and iaaH have been cloned and characterized from
various bacteria, such as Agrobacterium, Bradyrhizobium, Pseudomonas, Pantoea and
Rhizobium [85, 86]. Indole-3-pyruvate (IPyA pathway) pathway has been identified in
a number of bacterial genera including Azospirillum, Bradyrhizobium, Cyanobacteria
and Rhizobium. The first step is transamination of tryptopan into indole-3-pyruvate. In
the next step indole-3- acetic acid is decarboxylated into indole-3-acetaldehyde. In the
last step this aldehyde intermediate is oxidized into indole-3-acetic acid [87].
Insertional inactivation of the pathway resulted in a lower IAA production e.g. up to
90% reduction in Azospirillum brasilense [88] indicating the importance of the IPyA
pathway in auxin production. In bacteria, the tryptamine (TAM) pathway has been
identified in Bacillus by identification of tryptophan decarboxylase activity [88] and in
Azospirillum by detection of the conversion of exogenous tryptamine to IAA [89]. A
bacterial tryptophan-independent pathway could be demonstrated in Azospirillum
brasilense by feeding experiments with labeled precursors. This pathway is
predominant in case no tryptophan is supplied to the medium [88].
IAA-mediated ethylene production could increase root biomass, root hair
number and consequently the root surface area. Involvement of PGPR formulated
cytokinins, showed increase in root initiation, cell division, cell enlargement and
increase in root surface area of crop plants through enhanced formation of lateral and
adventitious roots [36, 37]. Although ethylene is essential for normal growth and
development in plants, at high concentration it can be harmful as it induces defoliation
and other cellular processes that may lead to reduced crop performance. Thus,
rhizobacteria assist in diminishing the accumulation of ethylene levels and re-establish

1. Introduction
14
a healthy root system needed to cope with environmental stress. The primary
mechanism includes the destruction of ethylene via enzyme ACC deaminase.
Rhizosphere bacteria such as Achromobacter, Azospirillum, Bacillus, Enterobacter,
Pseudomonas and Rhizobium have been reported with ACC deaminase activity [22,
27]. IAA-mediated ethylene production could increase root biomass, root hair number
and consequently the root surface area of PGPR inoculated tomato plants [17].
Involvement of PGPR formulated cytokinins have also been observed in root initiation,
cell division, cell enlargement and increase in root surface area of crop plants [36,37]
1.4.8 PGPR as Biofertilizers
Application of PGPR strains as Biofertilizers is increasing due to high price of chemical
fertilizers which are being used extensively in agricultural system. PGPR have been
continuously used to enhance the plant growth, seed emergence and overall yield of
crops in different agro-ecosystems. Bio-inoculant application of nitrogen fixing
bacteria such as Azospirillum, Azotobacter, Acinetobacter, Bacillus, Burkholderia,
Enterobacter and Pseudomonas resulted in increased plant growth and yield of various
crops [24].
Inoculation of rice varieties with Pseudomonas strain K1 showed an increase in
shoot biomass and grain yield over that of non-inoculated control plants [66]. Growth
responses of wheat after the inoculation with rhizobacteria suggested that the growth of
wheat basically depends on a number of factors like plant genotype, nature of PGPR
inoculants as well as environmental conditions [90]. A balanced nutrition of various
crops such as sorghum, barley, black gram, soybean and wheat can be achieved by
inoculation of these plants with diazotrophic and p-solubilizing bacteria in combination
rather than single microbe inoculation [91, 92]. Co-inoculation of Enterobacter and
rhizobia resulted in improved growth and yield of chickpea [93, 94]. Co-inoculation of
Bradyrhizobium with plant growth promoting rhizobacteria (PGPR) enhanced the
nodulation and root and shoot growth in mung bean [95].
The use of microbial preparations for increasing crop production has become
common practice in many countries including Pakistan. Several types of biofertilizers
are being produced commercially by public research organizations as well as private
sector. NIBGE is providing biofertilizer (Bio-power) for almost all major crops

1. Introduction
15
including wheat, rice, sugarcane, maize and leguminous crops. This product is based
on different consortia of beneficial bacteria and primarily contains a combination of
nitrogen fixers, P-solubilizer and IAA producers.
1.5 Effect of Transgenic Plants in Rhizosphere Environment
In countries where GMO technology has been accepted, the land area planted for
commercial production of transgenic plants is increasing every year [96]. The effects
of transgenic plants on the soil and ecosystem function should be carefully evaluated
before the release of any transgenic plant variety. Biosafety studies on plants should
include the study of their effects on soil organisms [97]. Plants are known to have a
profound effect on the abundance, diversity and activity of soil microorganisms living
in close proximity with their roots in a soil zone defined as the rhizosphere [98].
Bacteria inhabiting the rhizosphere, also referred as rhizobacteria, are responsible for
numerous functions including nutrient cycling and decomposition, which can
significantly influence vegetation dynamics [99]. Among these, plant growth-
promoting rhizobacteria represent one of the best-characterized functional group of
rhizobacteria known for playing a significant role in plant health and plant development
[100, 101]. As it can be assumed that any significant impact of plant genetic
transformation might alter these fundamental microbial processes, rhizobacteria have
been defined as good indicator organisms and have been studied to assess the general
impact of GMPs on the soil environment [102].
1.5.1 Effect of Transgenic Plants on Soil Microorganisms
Different transgenes, with expression of novel plant proteins, can potentially alter
diversity and abundance of rhizobacteria by direct release of novel proteins into the
rhizosphere through plant root exudation or enhance production and release [103].
Some unintentional effects on specific pests or pathogens may be carefully investigated.
Similarly risk to alter non-targeted rhizobacteria may be evaluated [104]. It has been
reported that Bt-recombinant DNA and expressed Cry proteins were released in soils
through root exudation and plant tissue decomposition where they remained intact and
chemically active for extended periods of time [105, 106]. T4 lysozyme a common
transgene protein, is not only present in root exudates but it maintains biological activity
after entering the soil [107]. Root system architecture, composition of root exudates
and their quantity, and ability of soil nutrient utilization are crops or cultivar-related

1. Introduction
16
factors which can be used as determinant in plant and rhizobacterial interactions [108].
Different studies presenting the effects of transgenic plants on soil micro-organisms
have been summarized in Table 1.4.
Table 1.4 Effect of transgenic plants on structure and functions of soil
microorganisms and their communities
Plant Transgenic trait Effect on soil biota Reference
Cotton Insect resistance
(cry1ac)
(CrylAc and CpTI )
Significant stimulation in
growth of culturable bacteria
and fungi with change in
substrate utilization.
No apparent impact on
microorganism populations in
rhizosphere soil
[105]
[109]
Wheat Pathogen
Resistance
(Root rot resistance
Chromosome S-615)
Variation in cultivable
rhizospheric community
[110]
Rice Insect resistance
(Cry1Ab)
(Cry1Ca)
No persistent effect on soil
enzymatic activities
No obvious adverse effects on
the growth of Chlorella
pyrenoidosa.
[111]
(i) Insect resistance
(cry1Ab)
No significant differences in
earthworm, micro-rthropods,
nematodes and protozoan
May affect AMF under
different environmental
conditions
Significant changes occurred
in the abundances (revealed
by qPCR) of ammonia-
oxidizing bacterial and
archaeal communities
[111]
[112]
[113]
[114]
(ii)Herbicide
resistance
(pat)
No effect of transgenic maize
was observed on genetic
diversity of bacterial
communities in rhizospheric
samples.
[115]
Maize

1. Introduction
17
Potato (i) Insect resistance
(Invertebrate pest
control conA and
GNA lections)
Altered CLPP pattern of
microbial community in
transgenic rhizosphere
[116]
(ii) Expressing the
phage T4 lysozyme
gene
Transgenic potato plant
roots showed high
bactericidal activity against
Bacillus subtilis adsorbed
artificially on potato roots
as compared to non-
transgenic plants
[107]
(iii) T4 lysozyme
producing plant lines
DL4 and DL5
No difference in growth of
bacterial communities was
observed between the
rhizosphere of transgenic
potato and non-transgenic
potato varieties.
[117]
Soybean Herbicide resistance
(Glufosinate tolerant
EPSPS)
Incidence of Fusarium
(soilborne pathogen) on
transgenic soybean roots was
greater within 1 week after
the application of
glyphosate as compared to
non-transgenic isoline.
[118]
Alfalfa Organic acid
expression
(a nodule-enhanced
malate
dehydrogenase)
Qualitative changes in the
abundance of bacterial
phylogenetic groups between
rhizosphere soils of
transgenic and
untransformed alfalfa.
[119]
Tobacco Expression of
proteinase inhibitor I
(cryIIIA. Bacillus
thuringiensis
var.tenebrionis (Bt.)
Numbers of collembella
colonies associated with
transgenic tobacco litter are
less as compared with non-
transgenic litter. Whereas
nematode population is high
in transgenic litter as
[120]
Rape (i)Herbicide
resistance
(Glufosinate
tolerant) (pat)
Rhizosphere and root interior
microbial populations
associated with a transgenic
canola have altered CLPP and
fatty acid methyl ester
(FAME) profiles compared to
[122]

1. Introduction
18
the profiles of a non-
transgenic counterpart.
(ii)Herbicide
resistance
(pat)
From root interior and rhizosphere
fewer Arthrobacter, Bacillus,
Micrococcus and Variovorax
isolates, and more
Flavobacterium and
Pseudomonas isolates were found
in the root interior of Quest
compared to Excel or Parkland.
The bacterial root-endophytic
community of the transgenic
cultivar, Quest exhibited a lower
diversity compared to Excel or
Parkland.
[123]
Arabidopsis
thaliana
No effect on E. coli populations [124]
1.6 Diversity of Culturable and Non-Culturable Bacteria in
the Rhizosphere
Biosphere is dominated by microorganisms but only 0.1-10% microorganisms are
culturable while the vast majority remains uncultured [125]. Microbial diversity and
community structure cannot be described precisely without having the information
about non-cultured microorganisms in a particular environment [126]. Cell-
independent molecular approach for studying the microbial population and community
structure inhabiting in environmental samples on the basis of 16S rRNA gene sequence
analysis has explored a new perspective in microbial ecology [47, 127]. These
“metagenomics” studies are based on DNA isolated from environment and harbor the
genome of the entire population in the environment “Metagenome”. These studies help
in understanding of the community structure and the metabolic potential of a
community [125, 128]. These metagenomic studies helped in the detection and
identification of genes involved in production of antibiotics, anticancer agents,
industrial enzymes and the enzymes involved in bio-degradation of heavy metals [129,
130]. Soil microbial diversity and community structure depends upon various factors
i.e soil pH, temperature, moisture contents, nature and amount of root exudates, crop
rotations, soil nutrient status and agricultural practices [131-134].

1. Introduction
19
1.6.1 16S rRNA Gene as a Tool for Studying Diversity of Culturable
and Non-Culturable Bacteria
Bacteria are usually identified using phenotypic techniques that are mainly based on
production of enzymes and metabolism of carbohydrates. A number of biochemical and
morphological methods have been employed for identification of bacteria [135]. These
methodologies are based on chromogenic enzymatic reactions and are available in
commercial kits (BioLog Inc, Hayward, CA, USA; API20E and API ZYM systems by
Vitek, Inc, St. Lois, MO, USA). Bacterial identification on the bases of intrinsic
antibiotic resistance [136, 137], fluorescent antibody techniques, polyacrylamide gel
electrophoresis of total protein [138] and fatty acid analysis [139], have been used.
DNA based genotypic methods DNA-DNA hybridization, DNA-RNA hybridization,
use of random or specific primers in PCR, RFLP (Restriction Fragment Length
Polymorphism) have also been used successfully for identification of bacteria.
Ribosomes are important component of protein synthesis machinery in all living cells
including bacteria. A bacterial ribosome is composed of multiple ribosomal proteins
and three ribosomal RNAs i.e 23S rRNA, 16S rRNA and 5S rRNA. The genes that
encode for RNAs are organized in the genome as rrn operon and multiple rrn operons
in a bacterial genome.
The first bacterial 16S rDNA containing 1524 nucleotides (GenBank accession
no.J01859) was sequenced in 1972 by Ehresmann and his colleagues for Escherichia
coli [140]. The nucleotide sequences among various bacteria are highly conserved, the
conservation and divergence reflect bacterial evolution and each bacterial species has
its unique 16S rDNA sequences [141]. With the invention of polymerase chain reaction
(PCR) technology, 16S rDNA sequencing became a tool for bacterial phylogeny
studies. Partial or complete 16S rDNA can be amplified by PCR. Conserved regions of
16S rDNA allow design of highly conserved primers for nearly universal amplification
of most bacterial species [106, 142]. The nucleotide sequences of the amplicons are
determined, compared with a database, yield homology matches and consequent
identification of a particular bacterium. It is the variable regions of 16S rDNA that give
discriminatory power (Figure 1-3). The longer sequences reads give more accurate
identification, however, at least even 200 bp may yield meaningful results. Universal
PCR primers are chosen as complementary to the conserved regions most commonly

1. Introduction
20
near the ends of the gene. The sequence of the variable region in between is used for
the comparative taxonomy [143] and these primers are used to amplify the 16S rRNA
gene (Table 1.5). Presently universal primer pairs are available for amplification of
partial or full length amplification of 16S rRNA gene [144-146]. There are multiple
public and private databases available, such as GenBank, Ribosomal Database Project
(RDP).
1.6.2 Bacterial Diversity by Pyrosequencing Analysis of 16S rRNA
Gene
Traditional molecular techniques like DNA finger printing, cloning, and Sanger’s
sequencing cover only a single aspect either bacterial communities in number of soils
or bacterial diversity in few soil samples. The less tiresome and most efficient technique
to study the bacterial community structure and composition on the basis of 16S rRNA
sequence analysis in soil is molecular pyrosequencing. This new technique enables us
to study the depth as well as width of the soil samples [129, 147]. Pyrosequencing
provides a huge amount of parallel sequences obtained from a single DNA as compared
to traditional methodology like cloning. Pyrosequencing is a bioluminometric DNA
sequencing technique based on sequencing by synthesis [148]. This technique relies on
the real-time detection of inorganic pyrophosphate (PPi) released on successful
incorporation of nucleotides during DNA synthesis (Figure 1-5).

1. Introduction
21
Figure 1-3 Schematic representation of 16S rRNA gene annotated with variable
regions (V1 to V9) of 16S rRNA
Figure 1-4 16S rRNA gene with three distinct variable regions and primers [149].

1. Introduction
22
Table 1.5 Sequences of PCR primers for amplification of 16S rRNA gene*
Primer name Sequence (5'-3') Reference
27F AGAGAGTTTGATCCTGGCTCAG [150]
1492R ATTAGATACCCNGGTAG [151]
PA AGACTTTGATCCTGCTCAG [144]
PH AAGGAGGTGATCCAGCCGCA [144]
338F ACTCCTACGGGAGGCAGCAG [152]
533R CCAGCAGCCGCGGTAAT [153]
787F AGGATTAGATACCCTGGTA [154]
530F TAAAACTYAAAKGAATTGACGGG [155]
355R ACTGCTGCSYCCCGTAGGAGT CT [155]
1100R YAA CGA GCG CAA CCC [156]
1100R GGGTTNCGNTCGTTG [157]
*http://bioinfo.unice.fr/454/454_analyses_of_diversity.html
PPi is immediately converted to adenosine triphosphate (ATP) by ATP sulfurylase, and
the level of generated ATP is sensed by luciferase producing photons. Unused ATP and
deoxynucleotide are degraded by the nucleotide degrading enzyme apyrase. The
presence or absence of PPi, and therefore the incorporation or non-incorporation of each
nucleotide added, is ultimately assessed on the basis of whether or not photons are
detected [158].
Figure 1-5 Schematic representation of progress of enzymatic reaction in
pyrosequencing
DNA template with hybridized primer and four enzymes involved in pyrosequncing are
added to a well of a micro-titer plate. The four nucleotides are added step wise, and
incorporation is followed using the enzyme ATP sulfurylase and luciferase. The
unincorporated nucleotides of each addition are continuously degraded by apyrase
allowing addition of subsequent nucleotide.

1. Introduction
23
1.6.3 Functional Genes for Bacterial Identification and Detection
Functional genes with conserved nucleotide sequences have a supportive role in
bacterial identification and phylogenetic grouping of bacteria. For identification and
phylogenetic studies of diazotrophic bacteria, nifH is the most commonly used gene
after 16S rRNA. This gene encodes for components of nitrogenase enzyme responsible
for nitrogen fixation. The amplification and sequencing of this gene helps in grouping
of nitrogen fixing bacteria [159-162]. Some PGPR also possess 1-aminocyclopropane-
1-carboxylic acid (ACC) deaminase gene that controls the plant ethylene level and
results in enlargement of root system. Amplification of ACC deaminase gene in PCR
is also used as supportive tool for identification and grouping of PGPR [163, 164]. PCR
amplification and sequence analysis of genes like gdh (encodes membrane glucose
dehydrogenase) and pqqE (a gene having a role in P-solubilization) have also been used
to determine relatedness of specific bacterial groups [165].
1.6.4 nifH Metagenomics: A Tool to Study the Diversity of
Diazotrophic Bacteria
The nifH gene, that encodes the iron protein subunit of nitrogenase, is often used as a
biomarker gene to assess the diversity of nitrogen fixing organisms in various
environments. This gene is conserved among diazotrophs, and it provides a good
phylogenetic correlation between nif H and the 16S rRNA gene [45, 166]. Bacterial
diversity on the basis of nifH gene amplification and sequencing of soil DNA has been
investigated from rhizosphere of mangrove forest of China [167] amazon rain forest
[168], oak-hornbeam forest (Chor-bush Forest) near Cologne, Germany [169], soils
from France and Senegal [170], Douglas fir forest in USA [171] and rice rhizosphere
at Kyushu University Farm, Japan [172].
Polymerase chain reaction (PCR) of the nifH gene combined with cloning and
sequencing gives information on the diazotrophic composition in an environment.
Numerous PCR primers targeting the nifH genes have been designed with different
range of specificity, from “universal” covering different diazotrophic taxa [170, 173,
174]. Using the nifH gene as a molecular marker in natural environments provides
evidence for potential nitrogen fixation [175]. nifH gene is the most thoroughly studied
among the genes of the nif operon, with an extensive collection of sequences in the data

1. Introduction
24
bank obtained from both cultured and uncultivated microorganisms from multiple
environments [172, 176, 177]. Cultivation-independent studies using clone library
analyses [177], denaturing gradient gel electrophoresis (DGGE) [178] and terminal or
complete restriction fragment length polymorphism (RFLP) methods [170] have
already shown the great diversity of nifH genes in natural environments. Despite this
extent of knowledge, little is known about the assemblage of nitrogen fixing
communities in the complex soil environment [179].
1.6.5 Real Time PCR: A Gene Quantification Approach to Study the
Abundance of nif H and 16s rRNA Gene
Real-time PCR (Quantitative PCR or Q-PCR) is widely used in microbial ecology, to
determine gene abundance or transcript numbers present in specific environmental
samples. Target specificity of any Q-PCR assay is determined by the selected primers
that allow the quantification of taxonomic or functional gene markers present in a mixed
community. Q-PCR is a robust, sensitive, and highly reproducible method to
quantitatively track phylogenetic and functional gene changes under varying
environmental and experimental conditions. Q-PCR provides data sets that describe the
abundance of specific bacteria or genes to compare other quantitative environmental
data sets. The importance of Q-PCR is increasing in microbial ecology as it provides
understanding about the roles and contributions of particular microbial and functional
groups within ecosystem functioning [180]. Real-time PCR monitors amplicons
production during each PCR cycle using a fluorescent probe that emits fluorescence
during the reaction as an indicator of the extent of amplification of the target [181]. The
fluorescent signal is directly proportional to the amplification of the target (number of
new copies made). The equipment monitors the signal’s increase until it reaches the
exponential level, which determines the threshold cycle. By definition, the threshold
cycle is the first cycle in which there is a significant increase in fluorescence above the
background or a specified threshold value. Through threshold cycle measurement,
comparative quantification analysis allows the determination of the concentration of
target amplicons. The higher the starting DNA copy number, the quicker (fewer cycles)
the amplicons threshold is reached. SYBR Green I, a dye that intercalates non-
specifically to all double- stranded DNA, is a good and simple choice for real-time
PCR. Standardized in house protocols allow confirmation of the desired product

1. Introduction
25
(amplicons) from primer-dimer or non-specific amplification by melting curve
analyses. The melting curve, generated by the real-time PCR equipment, is a plot of
fluorescence versus temperature, when performed with an intercalating dye such as
SYBR Green I. The melting temperature of a specific DNA sequence, which is a
function of the number of base pairs and % GC content, is defined as the temperature
at which 50% of the DNA is in double-stranded form and 50% is in single-stranded
form. The need to quantify microbial populations is a pressing one in many areas of
microbial ecology. The first applications of Q-PCR in microbial ecology were reported
in 2000 to target 16S rRNA genes [182, 183] determined the spatial and temporal
quantitative differences in the distributions of Synechococcus, Prochlorococcus and
archaea in marine waters [184] quantified archaeal 16S rRNA gene numbers within
samples from a deep sea hydrothermal vent effluent, hot spring water and from
freshwater sediments. By targeting highly conserved regions of the 16S rRNA gene, Q-
PCR assays have been designed to quantify ‘total’ bacterial (and/or archaeal) numbers
while targeting of taxa-specific sequences within hypervariable regions within the gene
enables quantification of sequences. Q-PCR has also been applied to quantify
functional genes within the environment. By targeting functional genes that encode
enzymes in key metabolic or catabolic pathways and genetic potential for a particular
microbial function within a particular environment can be assessed. To understand
microbial functioning in the environment it is essential to know what genes are present
and the diversity of these genes to determine their abundance and distribution within
the environment. Quantification of functional genes involved in ammonia oxidation
[185], nitrate reduction and denitrification [186], sulphate reduction [187],
methanogenesis [188] and methane oxidation [189] have been investigated.
Pakistan falls into arid and semi-arid climatic conditions and agriculture in the
country is mainly dependent on the scanty and unpredictable rainfall. Therefor
irrigation water is considered as the most important limiting factor for crop production
(368, 369). High salt concentrations in soil are also major factor contributing to low
crop production as significant portion of agricultural lands has been wasted due to
salinity. Different approaches have been used for crop improvement which include the
development of transgenic plants. Transgenic crops e.g AVP1 transgene, engineered to
tolerate drought and salinity could significantly increase yields for many developing
countries. However legal issues related to the release of transgenes for cultivation is in

1. Introduction
26
the country have to be resolved that transgene is “Biosafe”. Biosafty of the AVP1
transgenic cotton and wheat has to be investigated and the present study was designed
to study any effects of transgenes on indigenous bacterial populations.
The main objective of the present study was the risk assessment of soil
environment by comparing the diversity of PGPR from rhizosphere of transgenic and
non-transgenic plants (cotton and wheat). We isolated and identified plant growth
promoting rhizobacteria from the rhizosphere and after characterization, plant growth
promotion ability of bacterial strains was assessed by inoculating transgenic as well as
non-transgenic plants of wheat and cotton. The bacterial strains that showed positive
growth promotion were considered as biofertilizer of cotton and wheat plants,
respectively. Bacterial population in the rhizosphere of transgenic and non-transgenic
plants (cotton and wheat) was studied by cfu/g and MPN count on different growth
media and by real time quantification of nifH and 16S rRNA gene directly amplified
from soil DNA. Moreover, bacterial diversity in the rhizosphere of transgenic as well
as non-transgenic plants was investigated by 16S rRNA and nifH sequence analysis
using culture-independent techniques.

2. Materials and Methods
2.1 Isolation of Bacteria from the Rhizosphere of Cotton and
Wheat
Soil samples were collected from rhizosphere of transgenic and non-transgenic cotton
and wheat plants that were grown separately in green house under controlled conditions.
The representative soil samples along with some fine roots were collected and stored at
4°C and used for further studies. These rhizosphere soil samples were used for
preparation of 10X serial dilutions up to 10-5 in saline solution (0.89% NaCl). From
each dilution (10-3 to10-5), 100µL were spread on nutrient agar plates [190] and
Pikovskaya agar plates [191]. The plates were incubated at 30°C for 24 hours to 72
hours. Bacterial colonies were counted on the bases of difference in their morphology.
From Pikovskaya agar plates the colonies with clear halo zone were considered positive
for phosphorus solubilization. Pure bacterial colonies were obtained by continuous
streaking on fresh plates containing respective growth medium.
2.1.1 Isolation of Diazotrophic Bacteria by Enrichment Culture
Technique
Fresh root pieces (8-10 cm long) with adhering soil were added to 1 mL semi-solid
NFM medium in 1.5 ml eppendorf tubes [192]. After two days, 50 µL from each tube
were transferred to new eppendorf tubes containing fresh semi-solid NFM. This
procedure was repeated five to six times. Each time bacterial growth was observed
under microscope. A full loop with growth medium from eppendorf was streaked on
nutrient agar plates. These bacteria were further purified by repeated sub-culturing on
nutrient agar.

2. Materials and Methods
28
2.2 Morphological Characterization of Bacteria
2.2.1 Colony and Cell Morphology
Single colonies from purified bacterial culture were transferred to nutrient agar broth.
A single colony was streaked on fresh agar plate and incubated for 24 hours. Colony
morphology (i.e. color, shape, margins) was observed under light microscope
(Labophot-2 Nikon, Japan). Bacterial cell morphology and motility were studied by
taking a drop of bacterial suspension on a glass slide by mixing a drop of sterilized
deionized water and adjusting a cover slip on it. Cell morphology and shape were
recorded under the microscope (Microscope, LaboPhot 2 Nikon, Japan)
2.2.2 Culture Preservation
Purified bacterial isolates were grown in LB broth at 28oC for 48hours and preserved
in glycerol (20%) at -80°C.
2.3 Phosphorus Solubilization
2.3.1 Qualitative Assay for Phosphorus Solubilization by Bacteria
Bacterial isolates selected from Pikovskaya medium were grown in LB broth medium
at 30oC for 48hours with shaking (300rpm). From this culture 50µL were spotted on
Pikovskaya agar plates [192]. These plates were sealed with para-film and incubated at
30°C for 2 weeks. Bacterial colonies with clear halo zone formation were selected and
considered as positive strains for phosphate solubilization.
2.3.2 Quantitative Estimation of Phosphate Solubilization by Bacteria
Quantitative estimation of phosphorus solubilization by bacteria was done by
molybdate blue color method [193]. Bacterial isolates were grown in 100 mL conical
flask containing 50 mL of Pikovskaya broth medium (pH 7), supplemented with tri-
calcium phosphate (5 g.L-1) as insoluble P source [191]. These cultures were grown at
30°C on a shaker (150 rpm) for 15 days. Bacterial growth was transferred to 50 mL
clean falcon tubes. Cell-free supernatant was harvested by centrifugation at 5000 rpm
for 10 minutes (Biofuge Primo, thermoelectron Corporation, Germany). The
supernatant was filtered through 0.45 µm filter (Orange Scientific GyroDisc CA-PC,
Belgium) and O.D was recorded at 882 nm on a spectrophotometer (Camspec M350
double beam UV visible, UK). Solubilized phosphate was determined by preparing

2. Materials and Methods
29
standard curve of known concentration of KH2PO4 (2, 4, 6, 8, 10, 12 ppm) using Regent
A, and Reagent B.
For preparation of regent A, 12.0 g of ammonium paramolybdate [(NH4)
Mo4O2.4H2O] was dissolved in 250 mL of distilled water. Potassium antimony tartrate
solution was prepared separately by dissolving 0.290 g of Potassium antimony tartrate
in 100 mL of distilled water. Both solutions were added to 1.0 L of 5N H2SO4, mixed
thoroughly and 2L volume was made by adding distilled water. Reagent A was stored
in Pyrex glass volumetric flask in dark. For preparation of reagent B, 1.056 g of ascorbic
acid was dissolved in 200 mL of Reagent A. Reagent B was prepared fresh every time
when required.
For preparation of stock solution, 0.439 of KH2PO4 was dissolved in 500
mL of distilled water, thoroughly shaken in volumetric flask and diluted to 1.0 L by
distilled water. Five drops of toluene were added to stop microbial activity. This stock
solution containing 100 ppm soluble phosphorous was diluted to make 2, 4, 6, 8, 10,
12, 14, 16 and 20 ppm solution of soluble P for spectrophotometer analysis. Two mL
of these solutions and 4.0 mL of reagent B were mixed and diluted with distilled water
up to 25 mL. Blue color developed within approximately 10 minutes. A blank solution
was also prepared by adding 4.0 mL of reagent B into 21 mL of distilled water. Optical
density of these solutions was measured on spectrophotometer (Camspec M350-Double
Beam UV-Visible Spectrophotometer, 39 UK) at 882 nm.
A graph was plotted between optical density and concentration (ppm) of
standard solution. Same procedure was repeated for filtered cell-free supernatants of
bacterial cultures for measuring P solubilization.
2.3.3 Extraction and Quantification of Organic Acids Produced By
Bacteria in Pikovskaya Medium
Organic acids produced by bacterial cultures in Pikovskaya medium were extracted by
mixing the cell-free supernatant with equal volume of ethyl acetate. Ethyl acetate was
evaporated to dryness. Organic acids were re-suspended in 1.5 mL of methanol and
filtered through 0.2 µm filters (Orange Scientific Gyro Disc CA-PC, Belgium). The
samples were analyzed on HPLC PERKIN ELMER series 200 with 20 µL auto-
sampler PE NELSON 900 series interface, PE NELSON 600 series link and PERKIN
ELMER NCI 900 Network Chromatography interface using Diode-array detector at

2. Materials and Methods
30
210 nm and their UV spectra (190-400 nm), Microgaurd cation-H Precolumn. Aminex
HPx-87H analytical column was used for separation. Sulfuric acid (0.001 N) was used
as mobile phase with flow rate of 0.3 mL/minute. Solutions (100 µg/mL) of acetic acid,
citric acid, malic acid, succinic acid, gluconic acid, lactic acid and oxalic acid were used
as standard. Peak area and retention time of samples were compared with standards for
quantification of organic acids [79].
2.4 Indole Acetic Acid Production by Bacterial Isolates
2.4.1 Colorimetric Estimation of IAA by Salkowski's Reaction (Spot
Test)
To determine indole acetic acid (IAA) production, bacterial cultures were grown at
30°C in LB medium supplemented with tryptophan (100 mg/L). After incubation for
five days, cultures were centrifuged at 5,000 rpm for 10 minutes to obtain cell-free
supernatant and pH was adjusted to 2.8 with HCl (1N). Salkowski’s reagent (1.0 mL
of 0.5M FeCl3, 30 mL of conc. H2SO4 and 50 mL dH2O) was prepared for qualitative
analysis of IAA. Different standards of IAA (1000, 100 and 10 µg/mL) were prepared.
Equal volumes of cell-free supernatant, and Salkowski’s reagent (e.g. 250 µL of each)
were mixed. Development of pink color indicated the IAA producing ability of bacterial
strain.
2.4.2 Quantification of IAA Production
For quantification of indol-3-acetic acid (IAA) production, cell-free culture medium
was obtained by centrifugation at 5000 rpm for 15 minutes. After adjusting 2.8 pH with
HCl (IN) extraction was done with equal volumes of ethyl acetate [194]. This mixture
was evaporated to dryness and re-suspended in 1 mL of ethanol. The samples were
analyzed on HPLC (Varian Pro star) using UV detector and C -18 column. Methanol:
acetic acid: water (30:1:70 v/v/v) was used as a mobile phase at the rate of 1.0 mL/min.
Pure indole -3-acetic acid (1000 µg/mL) was used as standard. Computer software
(Varian) was used to compare the retention time and peak area.

2. Materials and Methods
31
2.5 Identification of Bacterial Isolates by 16R rRNA Gene
Sequence Analysis
2.5.1 DNA Extraction from Pure Cultures of Bacterial Isolates
For identification of bacterial isolates by PCR amplified 16S rRNA gene sequence
analysis, DNA from pure bacterial cultures was extracted by CTAB method with minor
modifications [195]. Single bacterial colony from nutrient agar plates was inoculated
to nutrient broth (5 mL) grown for 24 hours at 30°C with continuous shaking on shaker
150 rpm (Kuhner shaker, Switzerland). Cultures were centrifuged at 10,000 rpm for 2
minutes to pellet down bacterial cells. These cell pellets were re-suspended in 567 µL
TE buffer (Tris, EDTA, pH 8). Bacterial cell lysis was done by adding 30µL of 10%
(w/v) SDS (sodium dodycyle thiosulphate), followed by incubation at 37°C for one
hour. After incubation, 100 μL NaCl 5M and 80 μL of CTAB [Cetyl trimethyl
ammonium bromide; 10% CTAB/0.7 M NaCl] were added, mixed thoroughly and kept
at 65oC for 10 minutes. The lysate was centrifuged at 10,000 rpm for 10 minutes. The
supernatant was extracted twice with 780 µL chloroform/isoamyl alcohol (24:1). It was
followed by extraction with 780 µL of phenol/chloroform/isomyl alcohol (25:24:1).
The supernatant was incubated at -20°C for 30 min, after adding 20 µL of Na acetate
(3M, pH 5.2) and 1mL of absolute ethanol. DNA was then precipitated by
centrifugation at 13,000 rpm for 20 minutes. Washing of DNA pallet was done with
70% ethanol before drying under vacuum. The DNA pellet was dissolved in 100 µL
double distilled de- ionized water and stored at -20° C for further use.
2.5.2 Identification of Bacterial Isolates
16S rRNA gene of bacterial isolates from cotton and wheat was amplified by PCR using
conserve primers PA and PH [144]. PCR amplification was performed in a total volume
of 25 µL, containing 1 µL template DNA of 40 ng/µL concentration, 2.5 μL 10X Taq
polymerase buffer, 0.5 μL 10 mM dNTPs, 2 μL of 25 mM MgCl2, 1 μM each of primer
and 0.2 units of Taq DNA polymerase. PCR was performed in a thermal cycler
(Eppendorf, Germany). Conditions for PCR were 95°C for 5 minutes, followed by 30
cycles (95°C for 1 minute, 52°C for 1 minute, 72°C for 3 minutes) and final extension
at 72°C for 10 minutes. PCR reactions were analyzed by 1% (w/v) agarose gel
electrophoresis in 1X TAE buffer and visualized under UV light after staining with
ethidium bromide (0.2-0.5 μg/mL). The gel was run at 50V for one hour with 1kb DNA

2. Materials and Methods
32
ladder (Fermentas, Germany) as a size marker. The desired bands were eluted from the
gel. PCR products were purified with QIA quick PCR purification kit (QIAGEN, USA)
according to the manufacturer instructions and sent for sequencing to Macrogen Korea.
The obtained sequences of 16S rRNA were trimmed (Bio-Edit 7.1) and identified
through BLAST search at NCBI (National Centre for Biotechnology Information)
(www.ncbi.nlm.nhi.gov). These sequences were deposited to GenBank EMBL and
accession numbers were obtained. The sequences that showed closely related homology
and few others sequences including type strains of related genera were downloaded as
FASTA file format. These sequences were subjected to multiple alignment by
CLUSTAL W [196] and phylogenetic analysis (NJ) outlined by [197] was performed
using software MEGA6. The bootstrap replicate (BS) values of > 50% or greater
represent well supported nodes and thus only those were retained.
2.6 Plant Inoculation Experiments
2.6.1 Soil Analysis and Plant Material
AVP1 transgenic and non-transgenic cotton plants were provided by Gene
Transformation Lab, NIBGE. The soil (sandy loam soil, total N: 0.007%, available N:
0.0044%, available P: 1.85ppm, pH: 8.4, EC: 3.15 m/S, organic matter 0.006%) used
in all inoculation studies was collected from experimental fields of NIBGE.
2.6.2 Bacterial Inoculum Preparation
For inoculation of plants (cotton and wheat) bacterial cultures were grown in LB agar
broth (50 ml) at 30ºC for 48h and centrifuged at 4000 for 10 min to get the cell pellet.
The cell pellet was washed with 0.8% saline solution and re-suspended in 100 ml saline.
One ml of bacterial culture (≃109 cfu/ml) was applied directly near to the root system.
2.6.3 Quick Screening of Bacterial Isolates in Sterilized Sand
For quick screening of the bacterial isolates for plant growth promotion, cotton and
wheat seedlings were grown in sterilized sand for 40 days. The seeds were surface
sterilized with sodium hypo-chloride for 5 min and then washed with sterilized water.
Plastic jars (200 cm3) were used to grow seedlings and three seedlings of cotton and
five seedlings of wheat were maintained in each jar with five replicates and were kept
in growth room under controlled environment at 12 -25°C with average daylight of 10-

2. Materials and Methods
33
12 hours. One mL of Hoagland solution (1/8 strength) was provided twice a week as
nutrient source.
2.6.4 Bacterial Inoculation of Cotton and Wheat Plants Grown In
Earthen Pots
Seeds of cotton (transgenic and non-transgenic, variety Coker) and wheat plants
(transgenic and non-transgenic, variety Sehar 2006) were surface-sterilized with 0.1%
(NaOCl) for 5 minutes, followed by washing in sterilized distilled water. The seeds
were grown in earthen pots (radius 24cm, 32cm depth) containing non-sterilized field
soil and watered at alternated days. The plants were kept in green house under
controlled conditions of light, temperature and humidity (28° C, photoperiod of 16/8 h
light/dark, with light flux density approximately 1600 lux and 65% relative humidity).
Three plants of cotton and five plants of wheat were maintained in each pot with three
replicates. To each pot 1.8 g each of urea and DAP (equivalent to the
recommended dose for cotton and wheat ) was applied. Plants were harvested at
maturity (~180 days) and data of different agronomic parameters was recorded. For
measuring the cumulative root length, plant roots were separated and washed in water
and spread on a transparent polyethylene sheet. The sheet with roots was put on the
desktop scanner which scanned the roots and created a computer image of the roots.
The root length were measured on the P-IV IBM computer and scanner by using root
image analysis programme (Washington State University Research Foundation
programme, Washington state university, USA).
2.6.5 Bacterial Inoculation of Wheat Plants Grown in Micro-Plots
After the pot experiment the isolates were inoculated to wheat plants in micro-plots
(1.2m x 1.2m) in net house with natural conditions of temperature and moisture. Non-
transgenic plants of same variety were grown as a control. Experiments were laid out
in Randomized Complete Block Design (RCBD) with three replicates. The plant to
plant distance was kept 18 cm and row to row 36 cm distance was maintained. Nitrogen
as urea, phosphorus as single super phosphate (SSP) and potassium as KCl @ 80% of
recommended fertilizer doses (recommended doses for NPK are 120, 100 and 70 kg
ha-1, respectively) was applied to plants. Seed inoculation was done as described in
previous section. Plant dry weight was measured after oven drying at 65o C till constant
weight and grain yield was determined after harvesting.

2. Materials and Methods
34
2.6.6 Statistical Analysis
Data collected from pot and micro-plot experiments were analyzed statistically by
analysis of variance [198] technique, using the statistix (version 8.1) software. One way
ANOVA was applied in pot experiment data as well as in micro plots experiments.
Least significant difference test (Fisher’s LSD) at 5% probability was used to compare
the differences among treatments means.
2.7 Estimation of Bacterial Population
2.7.1 Bacterial Population by Counting Colony Forming Units (cfu/g
of soil)
For estimation of bacterial populations, soil samples from rhizosphere of transgenic and
non-transgenic plants of cotton grown during 2012 and from wheat experiments
conducted during 2011-12, were collected. These soil samples were collected at
different plant growth stages i.e 30, 60 and 90 days after sowing (DAS). Plant growth
conditions like fertilizers, irrigation, growth conditions have been mentioned above (see
2.6.4). Plants were uprooted carefully and soil adhering to the roots was collected.
Rhizosphere soil (1.0 g) was added to 9 mL saline solution (0.89%) and used for
preparation of 10X serial dilutions up to 10-5. From each dilution, 100 µl soil
suspension was spread on nutrient agar plates. The agar plates were incubated at 30°C
for 2-6 days and colony forming units (cfu/g) were counted.
2.7.2 Bacterial Population by Counting Most Probable Number (MPN)
Diazotrophic bacteria were estimated by the most probable number (MPN) method.
Collected soil samples (cotton and wheat rhizosphere) were suspended and serially
diluted in sterile water. Aliquots (100 µL) of each dilution was inoculated to eppendorf
tubes (5 replicates) containing semi-solid NFM (100µL) and incubated 28°C for 15
days. After incubation bacterial growth was observed under microscope and growth
positive eppendorf tubes were used for determination of MPN [194].

2. Materials and Methods
35
2.7.3 Real Time PCR
DNA Extraction from Soil Rhizosphere
Soil samples were collected from the rhizosphere of AVP1 transgenic cotton and wheat
plants as well as non-transgenic controls plants, grown during 2012 and 2011-12,
respectively. Three plants from each pot were uprooted, soil attached to the roots was
separated and composite sample was prepared by mixing 5g of soil from each treatment.
Total 8 soil samples were prepared for DNA isolation. Two sub samples (0.5g) from
this composite sample were used for DNA extraction by Fast DNA Spin kit for Soil by
FastPrep® instrument (MP Biomedicals, USA). Genomic DNA extracted from soil was
used for further studies (nifH and pyrosequencing analysis)
Quantification of 16S rRNA and nifH Genes from Soil DNA by Real Time PCR
Quantification of 16S rRNA and nifH gene in soil DNA was carried out in a Bio-Rad
CFX96 real-time PCR system (Bio-Rad Laborat ories, Hercu les CA). For 16S rRNA
gene amplification primers (534f/783r) and for nifH gene amplification (polF/polR)
were used. The 10 μL reaction volume contained 10 ng of DNA template, 5μL of
SsoFast™ EvaGreen® Supermix (Bio-Rad) and 1 μL of each primer (12.5 µM stock).
Real-time PCR reaction conditions included an initial 3 minute enzyme activation at 95
°C, followed by 40 cycles of 5S denaturation at 95 °C and 5S elongation at 53°C for
amplification of 16S rRNA gene with (534f/783r) and 54°C for nifH gene (polF/ploR).
For standard preparations, PCR products of strain Pseudomonas syrangae for 16S
rRNA and Rhizobium for nifH standards were used. PCR products were purified by gel
electrophoresis and quantified in a NanoDrop 2000 spectrophotometer (Thermo Fisher
Scienti fic Inc. Wilmington, DE). Target copy numbers were estimated from soil
samples. It was done by using standard curves that were generated from 10-fold
dilutions of PCR products in triplicate. The absolute copy numbers in the standards
were calculated based on DNA concentrations and size of PCR products. For
calculation 660 g mol−1 average molecular mass of a double-stranded DNA molecule
was considered [199]. All real-time PCR reactions on soil samples were performed in
duplicate and mean values were estimated for each DNA sample. Melt-curve analysis
of the PCR products was performed at the end of each real-time run. No DNA template
controls were included in every run.

2. Materials and Methods
36
2.8 Extraction and Quantification of Root Exudates from the
Rhizosphere
Rhizosphere soil samples were collected from AVP1 transgenic cotton (2012) and
wheat (2011-12) rhizosphere. Falcon tubes containing 30 ml of sterilized distilled water
were weighed, then soil attached with the roots was added in these tubes. Weight of
these tubes (soil+roots attached) was noted. The tubes with samples were placed on a
shaker at 200 rpm for 30 minutes and then centrifuged at 5000 rpm for 10 minutes.
Supernatants were collected into new Falcon tubes. This supernatant was concentrated
up to 1.5 mL in a concentrator (eppendorf Concentrator 5301, Germany) and filtered
through 0.2 µm filter (Orange Scientific GyroDisc CA -PC, Belgium). Samples were
analyzed on HPLC PERKIN ELMER with the same conditions that were used for
detection of organic acid from pure cultures [79].
2.9 Diversity of Diazotrophic Bacteria in the Rhizosphere of
Transgenic and Non-transgenic Plants of Cotton and
Wheat
2.9.1 PCR Amplification of nifH Gene from Soil DNA
PCR amplification of nifH gene was done from soil DNA extracted from cotton and
wheat rhizosphere (described in 2.7.3). Purified DNA was obtained by incubating DNA
with RNaseA (Promega Corporation, Madison, WI) at 37° for 10 minutes with final
concentration 100 µg/mL. A 360 bp fragment of the nifH gene was amplified by using
a pair of primers nif H F and nif H R [200]. Reaction mixture was prepared in 25µL
volume containing 12.5µL GoTaq®Green Master Mix (Promega, WI), 2 μL of template
DNA, and 1μL of each forward and reverse primer from 12.5 μM stocks. PCR
conditions consisted of an initial denaturation at 95 °C for 3 minutes, followed by 35
cycles of 94 °C for 45 seconds, 53 °C for 60 seconds, and 72 °C for 60 seconds, and a
final elongation at 72 °C for 10 minutes. PCR products were purified using High Pure
PCR Cleanup Micro Kit (Roche Applied Science, USA). Required amplified product
was confirmed by visualizing on 1.2% agarose gel with 1 kb marker.
2.9.2 Cloning of nifH Gene and Sequencing Reactions
PCR products were cloned using the TOPO TA Cloning kit (Invitrogen Corporation,
Carlsbad, CA) according to manufacturer’s instructions. Clone libraries were

2. Materials and Methods
37
constructed in vector pCR2.1-TOPO and positive clones were selected (randomly 30
clones from each library) and sequencing was performed using the BigDye Terminator
v3.1 Cycle sequencing kit (Applied Biosystems, Rotkreuz, Switzerland) with M13
forward primer. Sequencing was done on ABIPRISM 3700 DNA Analyzer (Applied
Biosystems).
2.9.3 Phylogenetic Analysis
To determine the distribution of nifH and 16S rRNA gene among transgenic cotton and
wheat rhizosphere soil, all sequencing data were blast searched at NCBI data bank.
Sequence alignment was done on CLUSTER X with all strains from this study and
other closely related sequences derived from GenBank data base [201]. To analyze all
these sequences Maximum likelihood (ML) was adopted as described by [202].
Bootstrap value of 50 or greater was kept as representative [203].
2.10 Bacterial Diversity in the Rhizosphere of AVP1
Transgenic Cotton and Wheat by Pyrosequencing
Analysis
2.10.1 16S rRNA Gene Amplification for Pyrosequencing
DNA extracted from soil samples (Table 7) was used in a PCR with primers PYRO799f
and 1492r [204] to amplify bacterial 16S rRNA gene sequences. ‘PYRO799f ’ is a
derivative of 799f [153] containing a 16S rRNA gene conserved region, a unique
barcode and a binding site for the pyrosequencing primer (Table 6). Each PCR reaction
was carried out in a 50µL reaction volume containing 50 to 100 ng of template DNA,
25 picomoles each of primer PYRO799f and 1492r, 0.1 µM of each dNTP, 1x Ex Taq
PCR buffer (Tak ara Bio Inc, Mountain view CA), and 1.5 units of high fidelity TaKaRa
Ex Taq enzyme (Takara Bio Inc). PCR conditions were as follows: denaturation at
95°C for 5 minutes, 30 cycles at 95°C for 45 seconds, 55°C for 45 seconds, and 72°C
for 2 minutes, followed by a 10 minutes elongations at 72°C. PCR reactions were run
on a 1% agarose gel and bacterial amplicons with the expected size of 0.7 kb were
recovered using a Qiaquick Gel Extraction Kit (Qiagen, Valencia, CA).
Pyrosequencing was performed on these amplicons using standard titanium chemistry.
Sequences were processed through the custom length and quality filters at CAGE and
binned by barcode. In order to minimize the effects of sequencing errors, reads that
were of atypical length (<200 or >650 bp) or had ≥ 1 nucleotide mismatch in the primer

2. Materials and Methods
38
or barcode sequence were excluded from the analysis. Only reads with an average
quality threshold value of ≥ 20 were considered to be qualified for further analysis.
2.10.2 Analysis of the Pyrosequencing Data
From each data set representing a single sample on the bases of attached barcode DNA,
sequences were picked up randomly as follows. Original FASTA file with all sequences
were opened in a text file. All eight treatments were separated on the bases of different
attached barcodes and considered as Sequence ID file in the FASTA SEQUENCE
SELECTION module at the Ribosomal Database Project's (RDP’s) Pyrosequencing
Pipeline (http://pyro.cme.msu.edu). These FATSA files were used as input into RDP
Classifier [205]. Using an 80% confidence threshold, the resulting hierarchy was
downloaded as a text file and imported into Microsoft Excel for quantification of the
contribution of individual taxa (phylum, class, order, family, or genus) to the total
population. From Microsoft Excel staked bar graphs were constructed to compare
diversity among the treatments on phylum and genera level. Separate stake bar graphs
was drawn among the resident and rare genera among the treatments.

2. Materials and Methods
39
Table 2.6 Primer sequence with titanium adopter sequence
Name Titanium A Adaptor Barcode Primer sequence
454 Primer A: CCATCTCATCCCTGC
GTGTCTCCGACTCAG
Barcode PYRO799F:AACMGG
ATTAGATACCCKG
454 Primer B: CCTATCCCCTGTGTG
CCTTGGCAGTCTCAG
No barcode 1492R:TACGGYTACC
TTGTTACGACTT
Table 2.7 Soil samples with barcodes name and sequences
No. Soil sample Barcode Name Barcode Sequence
1 TC1 TFA35 GTCAACTG
2 NTC1 TFA36 GTGTCACA
3 TC2 TFA37 TAATCGCG
4 NTC2 TFA38 TACCGCTT
5 TW1 TFA39 TAGGATCC
6 NTW1 TFA40 TCACACAG
7 TW2 TFA41 TCCAAGCA
8 NTW2 TFA42 TCGAGTTG

3. Results
3.1 Isolation of Bacteria from the Rhizosphere of AVP1
Transgenic Cotton
Soil samples were collected from the rhizosphere of transgenic and non-transgenic
cotton grown in green house during the year 2011. A total of 12 bacterial isolates were
purified on nutrient agar plates (Table 3.8). Among these, seven isolates were purified
from AVP1 transgenic cotton plants and five were isolated from non-transgenic cotton
plants.
3.2 Isolation of Bacteria from the Rhizosphere of AVP1
Transgenic Wheat
AVP1 transgenic wheat plants along with non-transgenic wheat plants were grown in
micro-plots in a net house under natural conditions during 2010-11. A total of 14
isolates were purified from rhizospheric soil on nutrient agar plates (Table 3.9). Among
these, nine bacterial isolates were purified from AVP1 transgenic wheat and five isolates
from non-transgenic wheat.
Figure 3-6 Isolation of bacteria on nutrient agar medium by serial dilution
method

3. Results
41
Table 3.8 Colony and cell morphology of the bacterial strains isolated from the
rhizosphere of AVP1 transgenic and non-transgenic cotton
Isolate Host Colony
morphology
Cell morphology
1. NTW1 Non-transgenic
Cotton
Small, yellow
irregular margins
Short rods
2. NTC-2NF Non-transgenic
Cotton
Small, yellow
smooth margins
Motile short rods
3. NTC-7 Non-transgenic
Cotton
Pinkish white,
small wrinkled
margins
Plump rods with empty
space
4. NTC-4 Non-transgenic
Cotton
Off white, large,
irregular margins
Medium size rods
5. AC Non-transgenic
Cotton
White, circular,
flat, undulated
margins
Medium size motile
rods
6. D-4
Transgenic
Cotton
Yellow, medium
size, smooth
margins
Very motile short rods
7. B-alpha
Transgenic
Cotton
Pure white, small,
round margins
Medium sized rods
with few shorter cells
8. TN4-3NF Transgenic
Cotton
Irregular, milky
white , gummy
Thin short rods
9. A5 Transgenic
Cotton
Medium size,
transparent
irregular shape
Large cells motile
10. Azo-BM31 Transgenic
Cotton
Pink, small,
smooth margins
Small rods, highly
motile
11. CC Transgenic
Cotton
Yellowish, small
convex surface
Small rods
12. DC Transgenic
Cotton
White, large
Thin rods, slightly
motile

3. Results
42
Table 3.9 Colony and cell morphology of the bacterial strains isolated from the
rhizosphere of AVP1 transgenic and non-transgenic wheat
Isolate Host Colony morphology Cell morphology
1. NTC-1NF Non-transgenic
wheat
Medium size, cream color
round margins
Very short, motile rods
2. NTC-11 Non-transgenic
wheat
Light brown color,
wrinkled margins
Short rods, slightly
motile
3. WP1 Non-transgenic
wheat
White large colonies,
irregular, wrinkled
margins,
Thin rods, motile
4. WP3 Non-transgenic
wheat
Brown, large, smooth
margins
Small cells, vey motile
5. WP8 Non-transgenic
wheat
White, medium size,
wrinkled margins
Thin rods, slightly
motile
6. A6 Transgenic
wheat
Large colonies, transparent,
wrinkled margins
Medium size cells,
joined together to form
long rods
7. WN1 Transgenic
wheat
Light yellow, small Very short, motile rods
8. WN2 Transgenic
wheat
Off white, very small Short rods, motile
9. WT2 Transgenic
wheat
Small size, brown,
transparent, smooth
margins
Very short, motile rods
10. WP2 Transgenic
wheat
Medium size, white,
smooth margins
Shiny, thin rods
11. NFM-2 Transgenic
wheat
Small size, off-white,
smooth margins
Motile rods
12. AZ Transgenic
wheat
Medium size, light yellow,
smooth margins
Small cells, motile
13.Azo-BM31 Transgenic
wheat
Pinkish white, smooth
margins, small size
Short rods, motile
14. WC Transgenic
wheat
Off white, small size, dry
irregular margins
Motile short rods

3. Results
43
3.3 Identification of Bacterial Isolates by 16S rRNA Gene
Sequence Analysis
Genomic DNA was extracted from pure cultures for the amplification of 16S rRNA
gene using conserved primers (Figure 3-7). Bacterial identification by 16S rRNA
sequence analysis indicated that bacterial strains isolated from the rhizosphere of AVP1
transgenic and non-transgenic cotton belonged to seven genera i.e Agrobacterium,
Arthrobacter, Azospirillum, Bacillus (7,strains), Brevibacillus, Pseudomonas, and
Rhizobium (Table 3.10). The isolates from AVP1 transgenic and non-transgenic wheat
rhizosphere belonged to genera Achromobacter (2 strains), Actinobacteria, Advenella,
Alcaligenese, Arthrobacter, Bacillus (4 strains), Bervibacterium, Pseudomonas (2
strains) and Rhizobium (Table 3.11).
Phylogenetic trees were constructed using 16S rRNA gene sequences of the
bacterial isolates along with related sequences in the NCBI data base. Phylogenetic
trees were constructed by using 16S rRNA gene sequences of bacterial isolates from
AVP1 transgenic cotton and wheat and also from representative non-transgenic.
Phylogenetic relationships of different strains of genus Bacillus (strain WP8 and WP2
isolated from AVP1 transgenic wheat) and Paenibacillus (NTC-7 isolated from non-
transgenic cotton) were studied on the basis of 16S rRNA gene sequences (Figure 3-
11). Three major clusters were observed in this tree. Bacillus sp. strain WP8 and WP3
clustered with other Bacillus sp. taken from the Gene Bank. Paenibacillus sp. strain
NTC-7 clustered with Paenibacillus edaphicus. Bacterial isolate Achromobacter strain
AZ clustered with other Achromobacter sp. and uncultured Achromobacter (Figure 3-
14). Phylogenetic analysis of different isolates belonging to genus Pseudomonas
(Pseudomonas strain D4 isolated from AVP1 transgenic cotton and WP3 isolated from
transgenic wheat) showed three major clusters (Figure 3-15). Pseudomonas strain WP3
clustered with Pseudomonas straminea and Pseudomonas sp. strain D4 established
cluster with other Pseudomonas strains reported from a variety of environments.

3. Results
44
Figure 3-7 Genomic DNA extracted from bacterial isolates.
Lane 1, 1kb DNA ladder; Lane 2, isolate Wp3; Lane 3, isolate Bα; Lane 4, isolate D4;
Lane 5, isolate WN1.
Figure 3-8 16S rRNA gene amplified from bacterial isolates.
Lane 1, 1kb ladder; Lane 2, -ve control; Lane 3, isolate Wp3; Lane 4, isolate Bα; Lane
5, isolate D4; Lane 6, isolate WN1.
1 2 3 4 5 6
1500b
p bp
250 bp
1000 bp 16S rDNA (PCR product)
3
000 bp
1 2 3 4 5
1000 bp Genomic DNA extracted
from bacterial pure cultures
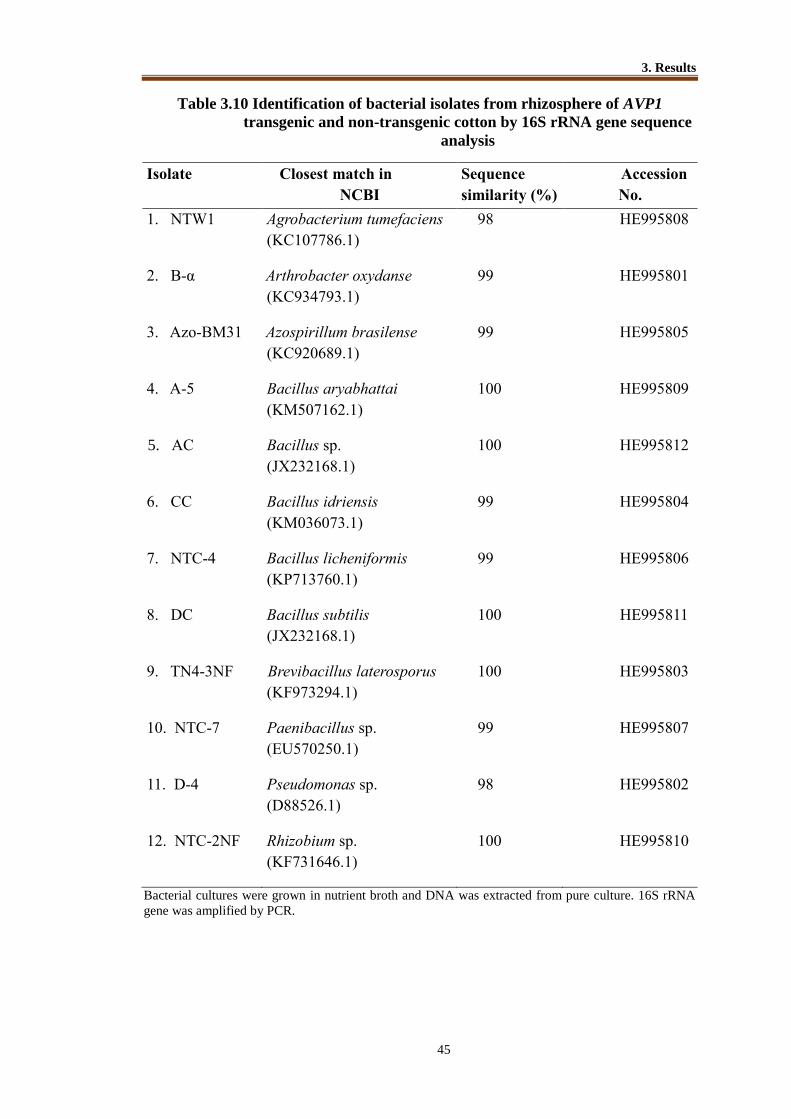
3. Results
45
Table 3.10 Identification of bacterial isolates from rhizosphere of AVP1
transgenic and non-transgenic cotton by 16S rRNA gene sequence
analysis
Isolate Closest match in
NCBI
Sequence
similarity (%)
Accession
No.
1. NTW1 Agrobacterium tumefaciens
(KC107786.1)
98 HE995808
2. B-α Arthrobacter oxydanse
(KC934793.1)
99 HE995801
3. Azo-BM31 Azospirillum brasilense
(KC920689.1)
99 HE995805
4. A-5 Bacillus aryabhattai
(KM507162.1)
100 HE995809
5. AC Bacillus sp.
(JX232168.1)
100 HE995812
6. CC Bacillus idriensis
(KM036073.1)
99 HE995804
7. NTC-4 Bacillus licheniformis
(KP713760.1)
99 HE995806
8. DC Bacillus subtilis
(JX232168.1)
100 HE995811
9. TN4-3NF Brevibacillus laterosporus
(KF973294.1)
100 HE995803
10. NTC-7 Paenibacillus sp.
(EU570250.1)
99 HE995807
11. D-4 Pseudomonas sp.
(D88526.1)
98 HE995802
12. NTC-2NF Rhizobium sp.
(KF731646.1)
100 HE995810
Bacterial cultures were grown in nutrient broth and DNA was extracted from pure culture. 16S rRNA
gene was amplified by PCR.

3. Results
46
Table 3.11 Identification of bacterial isolates from rhizosphere of AVP1
transgenic and non-transgenic wheat rhizosphere by 16S rRNA gene
sequence analysis
Isolate Closest match in NCBI Sequence
similarity
(%)
Accession
No
1. A6 Achromobacter sp. (HQ448952.1) 97 HE995801
2. AZ Achromobacter sp. (KM461115.1) 100 HE995802
3. NTC-1NF Advenella sp. (KM191133) 100 HE995812
4. WC Alcaligenes sp. (LM655389.1) 100 HE995804
5. NTC-11 Arthrobacter sp. (EU135627.1) 100 HE995805
6. Azo-BM30 Azospirillum sp. (NR118484.1) 100 HE995809
7. WP1 Bacillus safensis (LC015558.1) 100 HE995806
8. WP2 Bacillus pumilus (KM924441.1) 100 HE995807
9. WP8 Bacillus pumilus (KP224308) 100 HE995808
10. NFM-2 Brevibacterium sp. (HQ622520.2) 100 HE995811
11. WN1 Microbacterium sp. (KP301095.1) 99 HE995803
12. WT2 Pseudomonas putida (AM131104.1) 100 HE995813
13. WP3 Pseudomonas putida (KP313537) 100 HE995814
14. WN2 Rhizobium sp. (GU060510.1) 100 HE995815
Bacterial cultures were grown in nutrient broth and DNA was extracted from pure culture. 16S
rRNA gene was amplified by PCR

3. Results
47
Figure 3-9 Phylogenetic tree showing the phylogenetic relationship of different
strains of genus Bacillus and Paenibacillus.
Phylogenetic tree (Neighbor-Joining method) showing the phylogenetic relationship of
different strains of genus Bacillus (strain WP8 and WP3 isolated from AVP1 transgenic
wheat) and Paenibacillus (NTC-7 isolated from non-transgenic cotton) on the basis of
16S rRNA gene sequences. Amplified 16S rRNA gene fragment from the isolated
strains of Bacillus sp. strain WP8, WP3, and Paenibacillus sp. strain NTC-7 ( ) were
sequenced and BLAST searched through NCBI database. Closely related obtained from
NCBI databank and sequence of type strains ( ) were used and aligned using
CLUSTAR W. Distances were computed using the Jukes-Cantor method. The bootstrap
replicates (BS) values of 50% or greater represent well supported nodes and thus only
those were retained. E. coli (HM194886) was taken as an out group.
E coil (HM194886)

3. Results
48
Figure 3-10 Phylogenetic tree showing the phylogenetic relationship of the
Brevibacillus strains.
Phylogenetic tree (Neighbor-Joining method) showing the phylogenetic relationship of
the Brevibacillus strain TN4-3NF ( ) based on the sequences of the 16S rRNA gene.
Amplified 16S rRNA gene fragment from the isolated strain Brevibacillus strain TN4-
3NF (transgenic cotton) was sequenced and BLAST searched through NCBI database.
16S rRNA gene sequences from the current study along with those of the closely related
sequences obtained from NCBI databank and sequence of type strains ( ) were used
and aligned using CLUSTAR W. Distances were computed using the Jukes-Cantor
method. Numbers above the nodes represent maximum likelihood bootstrap support
above 50%.

3. Results
49
Figure 3-11 Phylogenetic tree showing the phylogenetic relationship of the
Arthrobacter strain
Phylogenetic tree (Neighbor-Joining method) showing the phylogenetic relationship of
the Arthrobacter strain B-α ( ) based on the sequences of the 16S rRNA gene.
Amplified 16S rRNA gene fragment from the isolated strains Arthrobacter strain B-α
was sequenced and BLAST searched through NCBI database. Closely related
sequences and sequence of type strains ( ) were obtained from NCBI databank and
aligned using CLUSTAR W. Distances were computed using the Jukes-Cantor method.
The bootstrap replicates (BS) values of 50% or greater represent well supported nodes.

3. Results
50
Figure 3-12 Phylogenetic tree showing the phylogenetic relationship of genus
Agrobacterium and genus Rhizobium
Phylogenetic tree (Neighbor-Joining method) showing the phylogenetic relationship of
genus Agrobacterium (strain NTW1) and genus Rhizobium (strain NTC-2NF isolated
from non-transgenic cotton) on the basis of 16S rRNA gene sequences. Amplified 16S
rRNA gene fragment from the isolated strains Agrobacterium strain NTW1 and
Rhizobium strain NTC-2NF ( ) were sequenced and BLAST searched through NCBI
database. Closely related sequences and sequence of type strains ( ) were obtained
from NCBI databank and aligned using CLUSTAR W. Distances were computed using
the Jukes-Cantor method. The bootstrap replicates (BS) values of 50% or greater
represent well supported nodes. E. coli (HM194886) was taken as out group.
.

3. Results
51
Figure 3-13 Phylogenetic tree showing the phylogenetic relationship of
Azospirillum strain
Phylogenetic tree (Neighbor-Joining method) showing the phylogenetic relationship of
Azospirillum strain BM31 isolated from AVP1 transgenic cotton on the basis of 16S
rRNA gene sequences. Amplified 16S rRNA gene fragment from the isolated strain of
Azospirillum strain BM31 ( ) were sequenced and BLAST searched through NCBI
database. Closely related sequences and sequences of type strain ( ) were downloaded
and aligned using CLUSTAR W. Distances were computed using the Jukes-Cantor
method. The bootstrap replicates (BS) values of 50% or greater represent well
supported nodes. E. coli (HM194886) was taken as out group.

3. Results
52
Figure 3-14 Phylogenetic tree showing the phylogenetic relationship of genus
Achromobacter
Phylogenetic tree (Neighbor-Joining method) showing the phylogenetic relationship of
genus Achromobacter (strain AZ isolated from AVP1 transgenic wheat) on the basis of
16S rRNA gene sequences. Amplified 16S rRNA gene fragment from the isolated strain
Achromobacter sp. strain AZ sequenced ( ) and BLAST searched through NCBI
database. Closely related sequences and sequences of type strain ( ) were downloaded
and aligned using CLUSTAR W and distances were computed using the Jukes-Cantor
method. The bootstrap replicates (BS) values of 50% or greater represent well
supported nodes. E. coli (HM194886) was taken as out group.
(HM19486)

3. Results
53
Figure 3-15 Phylogenetic tree showing the phylogenetic relationship of genus
Pseudomonas
Phylogenetic tree (Neighbor-Joining method) showing the phylogenetic relationship of
genus Pseudomonas (strain D4 isolated from AVP1 transgenic cotton and strain WP3
isolated from AVP1 transgenic wheat) on the basis of 16S rRNA gene sequences.
Amplified 16S rRNA gene fragment from the isolated strains Pseudomonas sp. strain
D4 and WP3 were sequenced and BLAST searched through NCBI database Closely
related sequences and sequences of type strain ( ) were downloaded and aligned using
CLUSTAR W and distances were computed using the Jukes-Cantor method. The
bootstrap replicates (BS) values of 70% or greater represent well supported nodes. E.
coli (HM194886) was taken as out group. The bootstrap replicates (BS) values of 50%
or greater represent well supported nodes. Pseudomonas stutzeri (U26416.1) was taken
as out group.

3. Results
54
3.4 Quantification of IAA Production by Bacterial Isolates
Phytohormone (IAA) production by bacterial isolates was determined on HPLC by
using standard methods. The isolates obtained from rhizosphere of cotton and wheat
were grown in LB broth medium supplemented with tryptophan as a precursor for IAA
biosynthesis. Among the bacterial isolates from cotton, maximum amount of IAA
(29.01 µg/mL) was determined in the growth medium of Bacillus sp. strain NTC-4,
followed by Agrobacterium strain NTW1 which produced 22.54 µg/mL of IAA in the
medium (Table 3.12). Bacillus sp. strain AC, DC and Bacillus sp. strain CC showed
relatively less production of IAA. Among the bacterial isolates from wheat rhizosphere,
maximum IAA production (19.88 µg/mL) was observed in the growth medium of
Arthrobacter sp. strain NTC-11 (19.6 µg/mL), followed by Brevibacterium sp. strain
NFM-2 (15.70 µg/mL) and Achromobacter sp. strain A6 (11.01 µg/mL) (Table 3.13)
Table 3.12 Quantification of IAA produced by bacterial strains isolated from
cotton
No. Bacterial strains IAA production*
(µg/mL)
1. Agrobacterium sp. strain NTW1 22.54 ± 3.3
2. Arthrobacter sp. strain Bα 10.06 ± 2.5
3. Azospirillum sp. strain Azo-BM31 12.3 ± 0.4
4. Bacillus sp. strain NTC-4 29.01 ± 5.5
5. Bacillus sp. strain AC 2.12 ± 0.2
6. Bacillus sp. strain CC 1.31 ± 0.2
7. Bacillus sp. strain A5 6.04 ± 0.1
8. Bacillus sp. strain DC 2.9 ± 0.1
9. Brevibacillus sp. strain TN4-3NF 11.25 ± 2.8
10. Paenibacillus sp. strain NTC-7 12.06 ± 1.2
11. Pseudomonas sp. strain D4 15.68 ± 1.4
12. Rhizobium sp. strain NTC-2NF 8.86 ± 2.2
*Bacterial cultures were grown for two weeks in LB medium containing tryptophan as
precursor of IAA. The values given are an average of 3 replicates.

3. Results
55
Table 3.13 Quantification of IAA produced by bacterial strains isolated from
wheat
No. Bacterial Strains IAA production* ( µg/mL)
1. Achromobacter sp. strain A6 11.21±1.43
2. Achromobacter sp. strain AZ 5.71±0.8
3. Alkaligense sp. strain WC 0.64±0.26
4. Arthrobacter sp. strain NTC-11 19.88±3.2
5. Advenella sp. strain NTC-1NF 7.86±1.2
6. Azospirillum sp. strain BM30 9 ±0.1
7. Bacillus sp. strain WP1 1.02±0.2
8. Bacillus sp. strain WP2 10.01±0.3
9. Bacillus sp. strain WP8 0.41±0.1
10. Brevibacterium sp. strain NFM-2 15.70±0.14
11. Microbacterium sp. strain WN1 4.64±0.6
12. Pseudomonas sp. strain WP3 9.18±1.25
13. Pseudomonas sp. strain WT2 8.01±2.11
14. Rhizobium sp. strain WN2 6.42±0.4 *Bacterial cultures were grown for two weeks in LB medium containing tryptophan as
precursor of IAA. The values given are an average of 3 replicates.
3.5 Phosphate Solubilization
3.5.1 Qualitative Assay for Phosphate Solubilization by Bacterial
Strains
Phosphate solubilization by bacterial isolates was studied on Pikovskaya agar plates
containing insoluble tri-calcium phosphate (TCP). Clear halo zone formation around
bacterial colonies indicated phosphate solubilization by bacteria in pure culture (Figure
3-16). These halo zones were observed around bacterial colonies after one week
incubation.
3.5.2 Quantitative Assay for Phosphate Solubilization by Bacterial
Strains
Bacterial strains that showed growth or clear halo zone on Pikovskaya agar plates were
selected for quantification of phosphate solubilization by spectrophotometric method.
Among the bacterial isolates from cotton rhizosphere, maximum P-solubilization
activity was observed in Arthrobacter sp. Bα (46.02 µg/mL), followed by Pseudomonas
sp. D4 (42.05 µg/mL) whereas minimum P-solubilization activity was recorded among
Bacillus sp. strain DC (1.32 µg/mL) and Bacillus sp. strain CC (4.52 µg/mL) (Table
3.14). Among the bacterial isolates from wheat rhizosphere, maximum P-solubilization
activity (122 µg/mL) was showed by Pseudomonas sp. strain WP3, followed by

3. Results
56
Achromobacter sp. strain A6 (33.5 µg/mL). Brevibacterium strain NFM2, and Bacillus
strains EC and WP1 did not show any P-solubilization activity (Table 3.15).
Figure 3-16 Plate assay for detection of phosphorus solubilization by bacterial
isolates on Pikovskaya medium supplemented with insoluble tri-
calcium phosphate (TCP)
NT.N4- NTCB-α
Halo-Halo-
WP1
WP8 Halo-
zone

3. Results
57
Table 3.14 Quantification of P solublization by bacterial isolates from cotton
Serial# Bacterial Strains Available P* (µg/mL)
1. Agrobacterium sp. strain NTW1 1.96 ± 0.4
2. Arthrobacter sp. strain B-α 46.02 ± 5.2
3. Bacillus sp. strain A-5 12.34 ± 0.6
4. Bacillus sp. strain AC 5.01 ± 0.4
5. Azospirillum sp. strain Azo-BM31 41.01 ± 1.7
6. Bacillus sp. strain CC 4.52 ± 1.2
7. Bacillus sp. strain NTC-4 9.08 ± 2.2
8. Bacillus sp. strain DC 1.32 ± 2.2
9. Brevibacillus sp. strain TN4-3NF 33.50 ± 1.2
10. Paenibacillus sp. strain NTC-7 14.08 ± 3.4
11. Pseudomonas sp. strain D4 42.05 ± 4.6
12. Rhizobium sp. strain NTC-2NF 6.94 ± 0.4
Table 3.15 Quantification of P solubilization by bacterial isolates from wheat
Serial #. Bacterial Strains Available P* (µg/mL)
1. Achromobacter sp. strain A6 33.5 ± 4.1
2. Achromobacter sp. strain AZ 7.4 ± 4.2
3. Alkaligense sp. strain WC 5.6 ± 0.5
4. Arthrobacter sp. strain NTC-11 27.0 ± 6.5
5. Advenella sp. strain NTC-1NF 2.5 ± 0.2
6. Azospirillum sp. strain Azo-BM30 27.1 ± 1.9
7. Bacillus sp. strain WP2 17.5 ± 1.4
8. Bacillus sp. strain WP8 22.0 ± 4.6
9. Microbacterium sp. strain WN1 26.0 ± 3.5
10. Pseudomonas sp. strain WP3 112.0 ± 3.7
11. Pseudomonas sp. strain WT2 17.0 ± 5.4
12. Rhizobium sp. strain WN2 26.2 ± 1.8
*Bacterial cultures were grown for two weeks in Pikovskaya growth medium (pH 7)
containing insoluble tri-calcium phosphate as insoluble phosphorus source. The values
given are an average of 3 replicates. No P solubilization was detected in pure culture of
Brevibacterium sp. strain NFM-2, and Bacillus strains EC and WP1.

3. Results
58
3.6 Quantification of Organic Acid Production by Bacteria
in Pikovskaya Medium Used for Studying Phosphate
Solubilization
Production of organic acids like acetic acid, citric acid, gluconic acid, lactic acid, malic
acid, oxalic acid and succinic acid by bacterial cultures was detected on HPLC in
Pikovskaya medium supplemented with sucrose as carbon source. Among these acids
acetic acid, citric acid, gluconic acid, lactic acid, malic acid, oxalic acid and succinic
acid were detected in the growth medium (Table 3.16). Among the tested strains,
Pseudomonas strain WP3 isolated from AVP1 transgenic wheat showed maximum
amount of acetic acid (38.21 µg/mL) production. Among the isolates from AVP1
transgenic cotton tested in this study Pseudomonas strain D4 showed maximum amount
of citric acid (13.87 µg/mL) production. Lactic acid and succinic acid were detected
relatively in low amounts. Oxalic acid was detected only in the pure cultures of
Paenibacillus strain NTC-7(1.02 µg/mL) and Bacillus strain WP8 (0.02 µg/mL) in low
amounts. Azospirillum strains BM31, Brevibacillus strain TN4-3NF isolated from
AVP1 transgenic cotton and Paenibacillus strain NTC-7 isolated from non-transgenic
cotton showed relatively higher amounts of all organic acids.

3. Results
59
Table 3.16 Quantification of organic acid production (µg/mL) by bacterial
isolates in the growth medium used for P-solubilization
*Organic acids by bacteria in Pikovskaya medium were quantified on HPLC and given
values (µg/mL) in the Table are an average of 3 replicates with standard deviation
*ND= Not detected.
Bacterial strains Acetic
acid
Citric
acid
Malic
acid
Lactic
Acid
Gluconic
Acid
Succinic
Acid
Achromobacter strain A6 12.54±1.2 4.3±2.01 ND* 2.31±1.9 ND* ND*
Agrobacterium strain NTW1 18.8±4.1 0.6±0.04 0.2±0.06 ND* 09±0.12 0.21±0.1
Arthrobacter strain B-α 19.05±3.8 7.0±1.2 ND* 0.3±0.03 0.1±0.04 0.21±0.02
Azospirillum strain BM31 18.02±08 6.5±1.5 2.08±09 2.5±1.1 4.56±0.1 0.9±0.12
Azospirillum strain BM30 8.74±0.8 5.6±0.8 ND* 3.1± 1.7 2.45±0.89 1.78±0.4
Bacillus strain A-5 5.1±1.4 8.3±2.8 ND* 0.2±0.12 ND* 1.4±0.02
Bacillus strain NTC-4 7.05±1.9 4.3±1.6 ND* 0.3±0.10 ND* ND*
Bacillus strain WP8 32.85±8 9.5±3.6 1.2±0.5 2.6±1.52 3.41±2.0 ND*
Brevibacillus strain TN4-
3NF
14.84±5.4 6.5±1.5 0.02±0.08 0.4±0.20 0.6±0.02 2.24±0.35
Microbacterium strain WN1 12.04±2.5 4.6±2.3 0.07±0.02 0.65±0.5 1.45±1.2 0.03±0.05
Paenibacillus strain NTC-7 14.56±4.3 7.9± 1.2 3.1±1.07 4.5±1.8 1.2±0.89 1.35±0.04
Pseudomonas strain D4 14.25±4.0 13.8±2.4 ND* 3.2±2.31 ND* ND*
Pseudomonas strain WP3 38.21±8 12.4±3.5 ND* 4.6±2.46 2.1±0.05 ND*
Pseudomonas strain WT2 11.0±2.6 3.6±1.27 1.2±0.12 0.71±0.8 1.25±1.25 0.31±0.02
Rhizobium strain WN2 9.65±0.8 5.8±0.95 ND* 2.2± 1.9 2.35±0.45 ND*

3. Results
60
Figure 3-17 Organic acid production (µg/mL) by bacterial isolates in pure
culture. Bacterial cultures were grown for two weeks in Pikovskaya medium
containing insoluble tri-calcium phosphate.
NTW1=Agrobacterium strain NTW1; B-α=Arthrobacter strain B-α; A5=Bacillus
strain A-5; NTC4=Bacillus strain NTC-4; TN4-3NF=Brevibacillus strain TN4-3NF;
NTC7=Paenibacillus strain NTC-7; D4=Pseudomonas strain; WN1= Microbacterium
strain WN1; WP3=Pseudomonas strain WP3; WP8=Bacillus strain WP8;
A6=Achromobacter strain A6; WT2= Pseudomonas strain WT2; WN2= Rhizobium
strain WN2; BM31= Azospirillum strain BM31; BM30= Azospirillum strain BM30

3. Results
61
3.7 Bacterial Inoculation of Cotton Plants
A series of experiments (Figure 3-18) during different years were conducted to evaluate
the effect of PGPR cotton plants. For quick screening of bacterial isolates, seedlings
were grown in small plastic jars filled with sterilized sand. These experiments were
performed with AVP1 transgenic cotton along with non-transgenic plants as control.
Transgenic and non-transgenic plants used in these experiments belonged to the same
event and generation. Transgenic plants were confirmed by PCR amplification of the
AVP1 gene by Gene Transformation Group, NIBGE and PCR positive plants were used
for data collection.
Figure 3-18 Bacterial inoculation experiments on cotton plants conducted in
different years in growth room.
3.7.1 Experiment 1 (year 2009)
Effect of Bacterial Inoculation on Growth of AVP1 Transgenic and Non–Transgenic
Cotton Grown in Sterilized Sand
For quick screening of bacterial strains for their growth promotion ability, a short term
experiment was conducted in sterile sand under controlled conditions (Figure 3-19). In
the present study 8 bacterial isolates selected on the bases of high IAA production and
phosphate solubilizing ability, were used as single-strain inoculants for AVP1
transgenic and non-transgenic cotton. These strains were tested for their growth
promotion potential in short-term experiments using sterilized sand. Sub-sets of
bacterial strains showing significant improvement in growth of cotton plants in sand
culture were applied as inoculum in pot experiment using field soil. Bacterial isolates
Agrobacterium strain NTW1, Arthrobacter strain Bα, Azospirillum strain BM31,
Bacillus strain NTC-4, Bacillus strain A5, Brevibacillus strain TN4-3NF, Paenibacillus
Cotton plant inoculation experiments
2009
2011
2012
Sand Culture (Sterilized sand)
Pot Experiment (Non-sterilized soil)
Pot Experiment (Non-sterilized soil)

3. Results
62
strain TNC-7 and Pseudomonas strain D4 were tested (Table 3.17). Plants were
harvested after 40 days of sowing and data on cumulative root length, root dry weight
and shoot dry weight were recorded. A significant (P≤ 0.05) improvement in growth
parameters of inoculated transgenic plants was recorded over non-inoculated control
plants (Table 3.17A; Figure 3-20). Among the transgenic plants, inoculated strains
showed significant improvement of cumulative root length, root dry weight and shoot
dry weight over non-inoculated control. Maximum increase in root length, root dry
weight and shoot dry weight of transgenic cotton plants was recorded in the treatments
inoculated with Arthrobacter strain Bα, Bacillus strain NTC-4, Brevibacillus strain
TN4-3NF and Pseudomonas strain D4. Maximum increase in root length (24%) and
root dry weight (22.1%) of transgenic plants was shown by inoculation of Arthrobacter
strain Bα. No significant effect was observed on the shoot dry weight and root dry
weight of transgenic plants inoculated with Agrobacterium strain NTW1. Inoculation
of non-transgenic plants with the same set of bacterial strains showed that there was no
significant effect of inoculation on the root dry weight of non-transgenic plants. A
significant (P≤ 0.05) impact on cumulative root length and shoot dry weight was
recorded among non-transgenic plants inoculated with Arthrobacter strain Bα,
Azospirillum strain BM31, Brevibacillus strain TN4-3NF and Pseudomonas strain D4.
Maximum increase in root length (24%) was recorded on inoculation of Azospirillum
strain BM31 and maximum shoot dry weight (26.7%) was shown by inoculation of
Pseudomonas strain D4.
Figure 3-19 Effect of bacterial inoculation on growth of cotton plants (transgenic
and non-transgenic)
Inoculated transgenic cotton plants Non-inoculated

3. Results
63
Table 3.17 Effect of bacterial inoculation on growth of AVP1 transgenic (A) and
non-transgenic cotton (B) grown in sterilized sand under controlled
conditions (year 2009)
A
Transgenic cotton
Treatment Cumulative root
length (cm)
Root dry
weight (g)
Shoot dry weight (g)
Control (Non-inoculated) 21.79 C 0.529C 2.098 D
Agrobacterium strain NTW1 24.63 B 0.532C 2.160 CD
Arthrobacter strain Bα 27.05 A 0.657A 2.448 B
Azospirillum strain BM31 23.65 BC 0.582B 2.474 B
Bacillus strain NTC-4 26.95A 0.687A 2.484 B
Bacillus strain A5 23.65 BC 0.620B 2.244 C
Brevibacillus strain TN4-3NF 26.20 AB 0.623B 2.528 B
Paenibacillus strain TNC-7 25.08 AB 0.624B 2.110 D
Pseudomonas strain D4 26.84 A 0.631B 2.652 A
LSD(P≤ 0.05) 2.83 0.071 0.24
B
Non-transgenic cotton
Treatment Cumulative root
length (cm)
Root dry
weight (g)
Shoot dry weight
(g)
Control (Non-inoculated) 20.20C 0.50 2.30D
Agrobacterium strain NTW1 25.30AB 0.49 2.24D
Arthrobacter strain Bα 25.90AB 0.51 2.89AB
Azospirillum strain BM31 26.26A 0.67 2.87AB
Bacillus strain NTC-4 25.00AB 0.67 2.60C
Bacillus strain.A5 22.15BC 0.63 2.32D
Brevibacillus strain TN4-3NF 23.10ABC 0.48 2.75BC
Paenibacillus strain TNC-7 22.00BC 0.51 2.34D
Pseudomonas strain D4 23.60ABC 0.53 2.94A
LSD(P≤ 0.05) 1.12 NS 0.31
Values in same column sharing the same letter do not differ significantly (P≤ 0.05) according
to Fisher’s LSD, (n=3). Three seeds were grown in plastic jars containing sterilized sand and
1.0 mL bacterial cultures (≃ 109cfu/mL) were inoculated to seedlings after emergence. Three
seedlings were maintained in each jar. Values are an average of five replications.
B

3. Results
64
Figure 3-20 Effect of bacterial inoculation on root and shoot dry weights of
transgenic and non-transgenic plants Control = non-inoculated, T= AVP1 transgenic cotton; NT= Non-transgenic cotton;
NTW1= Inoculated with Agrobacterium strain NTW1; Bα= Inoculated with
Arthrobacter strain Bα; NTC-4= Inoculated with Bacillus strain NTC-4; A5=
Inoculated with Bacillus strain A5; TN4-3NF = Inoculated with Brevibacillus strain
TN4-3NF; TNC-7= Inoculated with Paenibacillus strain TNC-7; D4 = Inoculated with
Pseudomonas strain D4
0
0.5
1
1.5
2
2.5
3
Wei
gh
t (g
)
Treatment
T.Root Dry weight (g) NT.Root Dry weight (g)
T.Shoot Dry weight (g) NT.Shoot Dry weight (g)

3. Results
65
3.7.2 Experiment 2 (Year 2010)
Effect of Bacterial Inoculation on Growth of AVP1 Transgenic and Non–Transgenic
Cotton Grown Under Controlled Conditions in Earthen Pots
Promising bacterial strains showing plant growth promotion in short term experiments
in sand culture were selected and used as inoculum for cotton plants grown in earthen
pots. Pots were filled with non-sterilized soil collected from NIBGE experimental field
area. Arthrobacter strain Bα, Azospirillum strain BM31, Bacillus strain NTC-4,
Brevibacillus strain TN4-3NF and Pseudomonas strain D4 were used as single strain
inocula for transgenic and non-transgenic cotton plants (Table 3.18)
Among the inoculated plants, transgenic plants inoculated with Pseudomonas
strain D4 showed an increase in shoot dry weight (13.7%), root dry weight (8.7%), root
length (28.5%), and yield (11.19%), over non-inoculated control. Moreover,
inoculation of transgenic plants with Brevibacillus strain TN4-3NF also showed the
improvement of shoot dry weight (12.9%), root dry weight (8.3%) and root length
(18.5%) (Table 18A). Inoculation of bacterial strains showed non-significant effect on
cotton boll production (no. of bolls). Maximum increase in yield of transgenic cotton
plant was recorded on inoculation with Brevibacillus strain TN4-3NF (11.2%) and
Pseudomonas strain D4 (9.6%).
Non-transgenic plants inoculated with Arthrobacter strain Bα, Pseudomonas
strain D4, and Bacillus strain NTC-4, showed increase in shoot dry weight which was
13%, 14.0% and 13.8.6%, respectively. Non-transgenic plants showed non-significant
effect of inoculated strains on yield except Pseudomonas strain D4 and Arthrobacter
strain Bα. Inoculation with Pseudomonas strain D4 and Arthrobacter strain Bα resulted
in 22 % and 23% increase in yield of non-transgenic plants, respectively (Table 3.18B).

3. Results
66
Figure 3-21 AVP1 transgenic cotton plants grown under controlled conditions in
earthen pots
Non-
inoculated
transgenic plant
Inoculated transgenic plant Inoculated transgenic plant
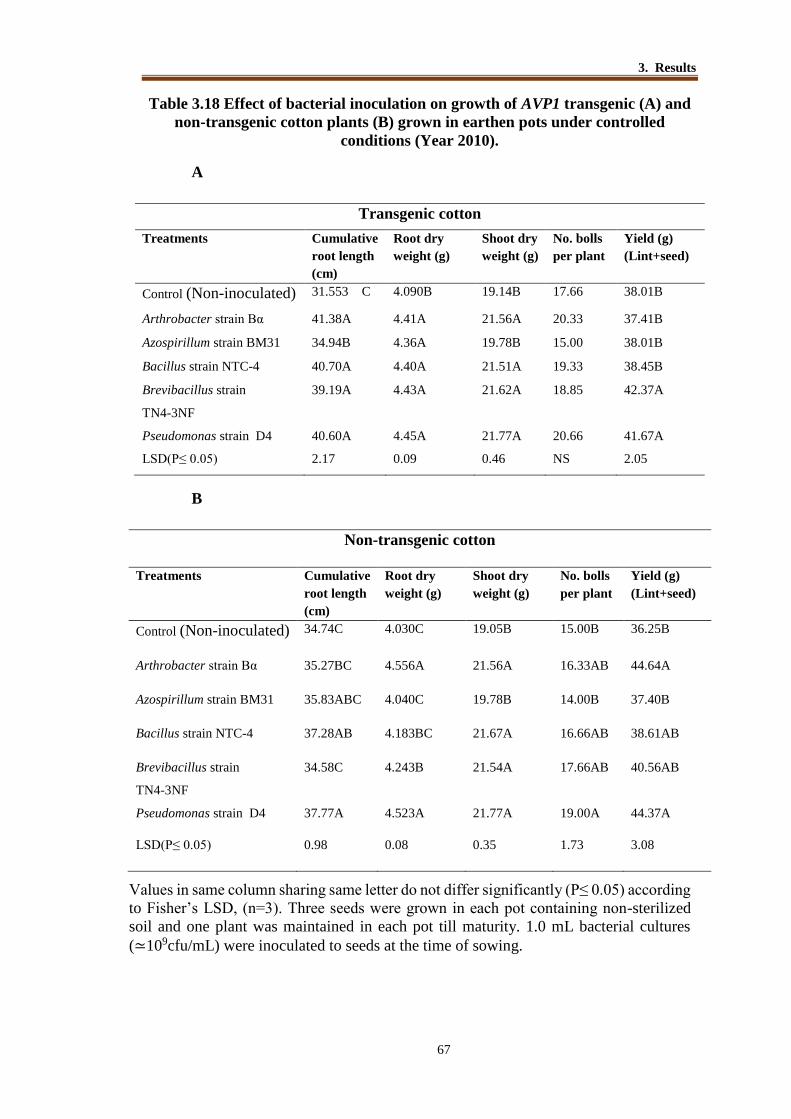
3. Results
67
Table 3.18 Effect of bacterial inoculation on growth of AVP1 transgenic (A) and
non-transgenic cotton plants (B) grown in earthen pots under controlled
conditions (Year 2010).
A
Transgenic cotton
Treatments Cumulative
root length
(cm)
Root dry
weight (g)
Shoot dry
weight (g)
No. bolls
per plant
Yield (g)
(Lint+seed)
Control (Non-inoculated) 31.553 C 4.090B 19.14B 17.66 38.01B
Arthrobacter strain Bα 41.38A 4.41A 21.56A 20.33 37.41B
Azospirillum strain BM31 34.94B 4.36A 19.78B 15.00 38.01B
Bacillus strain NTC-4 40.70A 4.40A 21.51A 19.33 38.45B
Brevibacillus strain
TN4-3NF
39.19A 4.43A 21.62A 18.85 42.37A
Pseudomonas strain D4 40.60A 4.45A 21.77A 20.66 41.67A
LSD(P≤ 0.05) 2.17 0.09 0.46 NS 2.05
B
Non-transgenic cotton
Treatments Cumulative
root length
(cm)
Root dry
weight (g)
Shoot dry
weight (g)
No. bolls
per plant
Yield (g)
(Lint+seed)
Control (Non-inoculated) 34.74C 4.030C 19.05B 15.00B 36.25B
Arthrobacter strain Bα 35.27BC 4.556A 21.56A 16.33AB 44.64A
Azospirillum strain BM31 35.83ABC 4.040C 19.78B 14.00B 37.40B
Bacillus strain NTC-4 37.28AB 4.183BC 21.67A 16.66AB 38.61AB
Brevibacillus strain
TN4-3NF
34.58C 4.243B 21.54A 17.66AB 40.56AB
Pseudomonas strain D4 37.77A 4.523A 21.77A 19.00A 44.37A
LSD(P≤ 0.05) 0.98 0.08 0.35 1.73 3.08
Values in same column sharing same letter do not differ significantly (P≤ 0.05) according
to Fisher’s LSD, (n=3). Three seeds were grown in each pot containing non-sterilized
soil and one plant was maintained in each pot till maturity. 1.0 mL bacterial cultures
(≃109cfu/mL) were inoculated to seeds at the time of sowing.

3. Results
68
Figure 3-22 Effect of bacterial inoculation on shoot dry weight, and yield
(lint+seed) of transgenic and non-transgenic plants
T=AVP1 transgenic plants; NT= non-transgenic plants; C = Non-inoculated control.
Bα = Inoculated with Arthrobacter strain Bα, BM 31= Inoculated with Azospirillum
brasilense strain BM31, NTC-4= Inoculated with Bacillus strain NTC-4, TN4.3NF=
Inoculated with Brevibacillus strains TN4-3NF, D4= Inoculated with Pseudomonas
strain D4
3.7.3 Experiment 3 (year 2011)
Effect of Bacterial Inoculation on Growth of AVP1 Transgenic and Non–Transgenic
Cotton Grown In Earthen Pots under Controlled Conditions
A pot experiment was conducted to study the effect of PGPR strains that were tested as
inoculum previously i.e year 2010. Bacterial strains Arthrobacter strain Bα,
Azospirillum strain BM31, Bacillus strain NTC-4, Brevibacillus strain TN4-3NF,
Pseudomonas strain D4 were used for inoculation of transgenic and non-transgenic
cotton plants. Inoculation with Arthrobacter strain Bα resulted in increased cumulative
root length (7.4%), shoot dry weight (8.9%), root dry weight (21.6%), and yield
(24.5%) of transgenic cotton plants over non-inoculated control. In this experiment
inoculation with Pseudomonas strain D4 resulted in increased cumulative root length
(7.9%), shoot dry weight (11.5%), root dry weight (21.8%) and yield (23%) of
transgenic cotton plants over control (Table 3.19A). Brevibacillus strain TN4-3NF also
showed a significant impact on cumulative root length (7.9%) and root dry weight
(21%) of inoculated plants as compared to non-inoculated control. Bacillus strain NTC-
4 did not show any significant impact on the growth of transgenic plants. The same
10
20
30
40
50
C Bα BM31 NTC-4 TN4-
3NF
D4
Wei
gh
t (g
)
Treatment
T.Shoot Dry Weight (g) NT.Shoot Dry Weight (g)
T.Yield (lint +seed) NT.Yield (lint +seed)

3. Results
69
isolates were applied to non-transgenic plants to study their performance as inoculants
(Table 19B). Inoculation of non-transgenic plants with Brevibacillus strain TN4-3NF
and Pseudomonas strain D4 resulted in the improvement of all growth parameters
studied. Two inoculated strains i.e Bacillus strain NTC-4 and Azospirillum strain BM31
did not show any improvement of plant growth (yield).
Figure 3-23 Bacterial inoculation of AVP1 transgenic and non–transgenic cotton
grown under controlled conditions in earthen pots
AVP1transgenic and non-transgenic cotton under controlled conditions
A
Non-transgenic plants AVP1 transgenic plants
AVP1 transgenic plants inoculated with different bacterial strains AVP1 transgenic
non-inoculated
B
Non-inoculated
Inoculated
Arthrobacter
Bα
Inoculated
Azospirillum
BM31
Inoculated
Pseudomonas
D4
Inoculated
Brevibacillus
TN4-3NF

3. Results
70
Table 3.19 Effect of bacterial inoculation on growth of AVP1 (A) transgenic and
non-transgenic cotton (B) grown in earthen pots under controlled conditions
A
Transgenic cotton
Treatment
Cumulative
root length
(cm)
Root
dry wt.
(g)
Shoot
dry wt.
(g)
Yield
(Lint +
seed)
Control (Non-inoculated) 53.50D 8.04B 22.00C 9.24C
Arthrobacter strain Bα 57.40A 9.78A 23.96AB 11.81AB
Azospirillum strain BM31 55.73B 8.31B 23.59AB 10.98AB
Bacillus strain NTC-4 54.41C 8.16B 22.26BC 10.57BC
Brevibacillus strain TN4-
3NF 56.35AB 9.74A 22.43BC 9.51C
Pseudomonas strain D4 57.76A 9.80A 24.54A 11.95A
LSD(P≤ 0.05) 0.69 1.4 0.92 0.40
B
Non-transgenic
Treatment Cumulative
root length
(cm)
Root dry
wt. (g)
Shoot
dry
wt.(g)
Yield
(Lint +
seed)
Control (Non-inoculated) 45.00D 7.04C 20.07D 10.66B
Arthrobacter strain Bα 48.33DC 7.88B 24.08A
B 12.10A
Azospirillum strain BM31 49.30C 7.61B 23.02B
C 11.68AB
Bacillus strain NTC-4 52.33BC 7.15C 22.59C 10.68B
Brevibacillus strain TN4-
3NF 54.43A 8.62A 24.98A 12.28A
Pseudomonas strain D4 55.10A 7.85B 22.32C 12.54A LSD(P≤ 0.05) 3.81 0.43 0.52 1.33
Values in same column sharing the same letter do not differ significantly (P≤ 0.05)
according to Fisher’s LSD, (n=3). Five seeds were grown in each pot containing
non-sterilized soil and three plants were maintained till maturity. 1.0 mL bacterial
cultures (≃109cfu/mL) were inoculated to seedlings after emergence. Three
seedlings were maintained in each pot with five replicates.

3. Results
71
Figure 3-24 Effect of bacterial inoculation on root dry weight, and yield
(lint+seed) of transgenic and non-transgenic plants T=AVP1 transgenic plants; NT= non-transgenic plants; C = Non-inoculated control.
Bα = Inoculated with Arthrobacter strain Bα, BM 31= Inoculated with Azospirillum
strain BM31, NTC-4= Inoculated with Bacillus strain NTC-4, TN4.3NF= Inoculated
with Brevibacillus strains TN4-3NF, D4= Inoculated with Pseudomonas strain D4.
0
5
10
15
20
25
Weig
ht
(g)
Treatment
T.Yield (lint +seed) NT.Yield (lint +seed)
T.Shoot Dry Weight (g) NT.Shoot Dry Weight (g)

3. Results
72
3.8 Bacterial Inoculation of Wheat Plants
A series of experiments during different years were conducted to evaluate the effect of
PGPR bacterial strains on wheat plants (Figure 3-25). For quick screening of bacterial
isolates, wheat plants were grown in small plastic jars filled with sterilized sand.
Efficient PGPR strains were selected to be used as inoculum in micro-plots under
natural conditions. These experiments were performed with AVP1 transgenic wheat
along with non-transgenic plants as control. Transgenic and non-transgenic plants used
in these experiments belonged to the same event and generation. Transgenic plants were
confirmed by PCR amplification of the AVP1 gene by Gene Transformation Lab
(NIBGE). Transgenic plants were confirmed by PCR amplification of the AVP1 gene.
All data on plant growth parameters were collected from PCR positive plants.
Figure 3-25 Bacterial inoculation experiments on wheat plants conducted in
different years
3.8.1 Experiment 1 (year 2009)
Effect of Bacterial Inoculation on Growth of AVP1 Transgenic and Non–Transgenic
Wheat Seeds Grown in Sterilized Sand under Controlled Conditions
Bacterial isolates from the rhizosphere of AVP1 transgenic and non-transgenic wheat
were inoculated to wheat seedlings grown in plastic jars filled with sterilized sand. The
plants were harvested 40 days after sowing. Bacterial isolates Achromobacter strain
A6, Arthrobacter strain NTC-11, Azospirillum strain BM30, Bacillus strain WP2,
Bacillus strain WP8, Brevibacterium strain NFM-2, Microbacterium strain WN1, and
Pseudomonas strain WP3 and were used as single strain inoculant for transgenic and
non-transgenic wheat plants (Table 3.20). Inoculation of AVP1 transgenic plants with
PGPR resulted in the growth improvement of wheat plants as indicated by an increase
in cumulative root length and dry weight of shoots and roots (Table 3.20A). Among the
Wheat plant inoculation experiments
2009
2010-11
2011-12
Sand Culture (Sterilized sand)
Micro-plot Experiment (Non-sterilized soil)
Micro-plot Experiment (Non-sterilized soil)

3. Results
73
tested strains, five strains i.e Achromobacter strain A6, Arthrobacter strain NTC-11,
Azospirillum strain BM30, Microbacterium strain WN1 and Pseudomonas strain WP3
showed an increase in all the growth parameters of transgenic plants over non-
inoculated control. Maximum increase in cumulative root length (22.1%), shoot dry
weight (10.5%) and root dry weight (23 %) over control plants was recorded in the
plants inoculated with Pseudomonas strain WP3. Inoculation with Arthrobacter strain
NTC-11 and Microbacterium strain WN1 resulted an increase (18.1% and 23 %,
respectively) in root dry weight over non-inoculated plants. The inoculation with
Azospirillum strain BM30 increased cumulative root length of plant (17.4 %) and
Achromobacter strain A6 increased root dry weight (21.5%) of AVP1 transgenic plants
over control. Among the bacterial inoculum tested for non-transgenic wheat plants, no
significant improvement of cumulative root length was recorded. Maximum increase in
shoot dry weight was recorded over control plants inoculated with Achromobacter
strain A6, Arthrobacter strain NTC-11 and Pseudomonas strain WP3. Maximum
increase (6%) in root dry weight of non-transgenic plants was recorded in plants
inoculated with Pseudomonas strain WP3 (Table 3.20B).
Figure 3-26 Effect of bacterial inoculation on growth of AVP1 transgenic wheat
plants grown in jars filled with sterilized sand
Table 3.20 Effect of bacterial inoculation on growth of AVP1 transgenic (A) and
non-transgenic wheat (B) grown in sterilized sand under controlled conditions
A

3. Results
74
B
Non–transgenic wheat
Treatment Cumulative
root length
Root dry
weight (g)
Shoot dry
weight (g)
Control (Non-inoculated) 24.64 0.70CD 2.54DE
Achromobacter strain A6 26.14 0.71CD 2.82A
Arthrobacter strain NTC-11 26.04 0.82AB 2.80AB
Azospirillum strain BM30 23.04 0.72CD 2.81AB
Bacillus strain WP2 25.44 0.75BC 2.72BC
Bacillus strain WP8 23.24 0.70CD 2.57DE
Brevibacterium sp. strain NFM-2 22.64 0.81AB 2.53E
Microbacterium strain WN1 22.44 0.72CD 2.73AB
Pseudomonas strain WP3 26.13 0.88A 2.81AB
LSD(P≤ 0.05) N.S 0.083 0.14
Transgenic wheat
Treatment
Cumulative
root length
(cm)
Root dry
weight (g)
Shoot dry
weight (g)
Control (Non-inoculated) 22.91D 0.65B 2.56B
Achromobacter strain A6 26.89ABC 0.79A 2.81A
Arthrobacter strain NTC-11 27.97A 0.77A 2.82A
Azospirillum strain BM30 26.95ABC 0.68B 2.57B
Bacillus strain WP2 22.85D 0.68B 2.62B
Bacillus strain WP8 26.52ABC 0.63B 2.65B
Brevibacterium sp. strain NFM-2 25.06CD 0.72AB 2.63B
Microbacterium strain WN1 27.91AB 0.64B 2.75A
Pseudomonas strain WP3 27.99A 0.80A 2.83A
LSD(P≤ 0.05) 2.88 0.092 0.14 Values in a column sharing same letter do not differ significantly (P≤ 0.05) according
to Fisher’s LSD, (n=3). Five seeds of wheat were grown in plastic jars containing
sterilized sand and 1.0 mL bacterial cultures (≃ 109cfu/mL) were inoculated to
seedlings after emergence. Three seedlings were maintained in each jar with five
replicates.
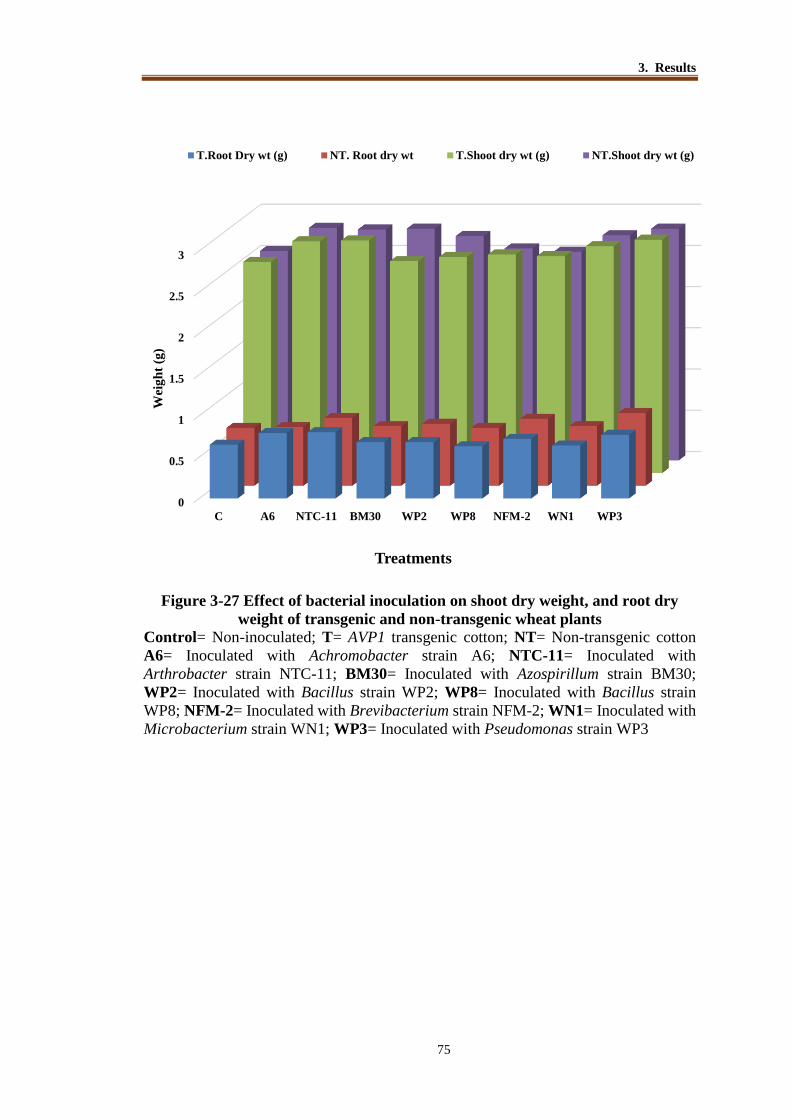
3. Results
75
Figure 3-27 Effect of bacterial inoculation on shoot dry weight, and root dry
weight of transgenic and non-transgenic wheat plants
Control= Non-inoculated; T= AVP1 transgenic cotton; NT= Non-transgenic cotton
A6= Inoculated with Achromobacter strain A6; NTC-11= Inoculated with
Arthrobacter strain NTC-11; BM30= Inoculated with Azospirillum strain BM30;
WP2= Inoculated with Bacillus strain WP2; WP8= Inoculated with Bacillus strain
WP8; NFM-2= Inoculated with Brevibacterium strain NFM-2; WN1= Inoculated with
Microbacterium strain WN1; WP3= Inoculated with Pseudomonas strain WP3
0
0.5
1
1.5
2
2.5
3
C A6 NTC-11 BM30 WP2 WP8 NFM-2 WN1 WP3
Wei
gh
t (g
)
Treatments
T.Root Dry wt (g) NT. Root dry wt T.Shoot dry wt (g) NT.Shoot dry wt (g)

3. Results
76
3.8.2 Experiment 2 (year 2011-2012)
Effect of bacterial inoculation on AVP1 transgenic and non-transgenic wheat grown
in micro-plots
Bacterial strains which showed efficient plant growth promotion in sand culture were
selected for use as inoculum of wheat in micro-plots. To evaluate the performance of
bacterial strains on growth of AVP1 transgenic and non-transgenic wheat, micro-plot
experiments were designed in a net house of NIBGE under natural (light and
temperature) conditions (Figure 3-28). In this experiment, Achromobacter strain A6,
Arthrobacter strain NTC-11, Microbacterium strain WNI, Azospirillum strain BM30
and Pseudomonas strain WP3 were used as single-strain inoculum. Plants were
harvested at maturity and data on different plant growth parameters (cumulative root
length, root dry weight, straw and grain weight) were recorded. Data analysis showed
a significant (<0.05) effect of inoculated strains on both transgenic and non-transgenic
plants as compared to non-inoculated control plants. Transgenic plants inoculated with
Arthrobacter strain NTC-11, Azospirillum strain BM30 and Pseudomonas strain WP3
showed significant (<0.05) increase in root length and root dry weight. Inoculation of
Arthrobacter strain NTC-11 showed maximum increase in the straw dry weight (8.1%)
over control and maximum increase in grain weight (8.2%) over control was noted in
the plants inoculated with Pseudomonas strain WP3. Inoculation of non-transgenic
wheat plant with Arthrobacter strain NTC-11, resulted in a maximum increase of
cumulative root length, straw weight and grain weight. All inoculated strains showed
significant improvement of straw weight and grain weight (Table 3.21).
Figure 3-28 Effect of bacterial inoculation on growth of AVP1 transgenic and
non-transgenic wheat grown in micro-plots under natural conditions.

3. Results
77
Table 3.21 Effect of PGPR strains on yield and growth parameters of transgenic
(A) and non-transgenic wheat (B) grown in micro-plots during 2011-2012
A
Transgenic wheat
Treatment Cumulative
root length (cm)
Root dry
weight(g)
Straw
weight (g)
Grain
weight (g)
Control (Non-inoculated) 22.05C 1.45C 656.67E 565.00C
Achromobacter strain A6 23.15BC 1.48C 677.00C 563.67C
Arthrobacter strain NTC-11 27.15A 1.67A 705.33A 594.33B
Microbacterium strain WNI 25.98AB 1.63B 669.67D 530.67D
Azospirillum strain BM30 27.16A 1.68A 692.00B 600.33B
Pseudomonas strain WP3 27.37A 1.67A 676.67C 611.67A
LSD(P≤ 0.05) 3.6 0.02 6.74 8.64
B
Transgenic wheat
Treatment Cumulative root
length (cm)
Root dry
weight(g)
Straw
weight (g)
Grain
weight (g)
Control (Non-inoculated) 22.66B 1.48C 636.0C 539.33 C
Achromobacter strain A6 23.83B 1.67 A 666.6B 562.00 B
Arthrobacter strain NTC-11 27.02 A 1.43C 687.6A 585.67 A
Microbacterium strain WNI 25.43AB 1.66AB 668.0B 556.00 B
Azospirillum strain BM30 24.06B 1.55BC 665.6B 558.67 B
Pseudomonas strain WP3 23.40 B 1.54C 659.0B 560.33 B
LSD(P≤ 0.05) 22.83 0.12 11.66 7.27
Wheat seeds were sown in Micro-plots (1.2mx1.2m) maintaining RXR distance 23 cm
and PXP distance15cm. Values are an average of 3 replications.

3. Results
78
Figure 3-29 Effect of bacterial inoculation on straw weight, and grain weight of
transgenic and non-transgenic wheat plants grown in micro-plots
C= Non-inoculated; T=AVP1 transgenic wheat, NT= Non-transgenic wheat, A6=
Inoculated with Achromobacter strain A6; NTC-11= Inoculated with Arthrobacter
strain NTC-11; WN1= Inoculated with Microbacterium strain WN1; BM30=
Inoculated with Azospirillum strain BM30; WP3= Inoculated with Pseudomonas strain
WP3
0
100
200
300
400
500
600
700
800
C A6 NTC-11 WN1 BM30 WP3
Wei
gh
t (g
)
Treatments
T.Grain weight (g) NT.Grain weight (g) T.Straw weight (g) NT.Straw weight (g)

3. Results
79
3.8.3 Experiment 3 (2012-2013)
Effect of Bacterial Inoculation on AVP1 Transgenic and Non-Transgenic Wheat
Grown in Micro-Plots
The experiment was conducted in cemented micro plots (1.2m X 1.2m) to evaluate the
effects of bacterial inoculation on AVP1 transgenic and non-transgenic wheat (Figure
3-30). The bacterial inocula included Achromobacter strain A6, Arthrobacter strain
NTC-11, Pseudomonas strain WP3, Azospirillum lipoferum strain BM30 and
Microbacterium strain WNI. Among the inoculated treatments of transgenic plants,
inoculation with four strains i.e Achromobacter strain A6, Arthrobacter strain NTC-11,
Pseudomonas strain WP3 and Azospirillum lipoferum strain BM30 resulted in a
significant increase in cumulative root length. Maximum increase in straw weight (12%
over control) grain weight (9%over control) was recorded in plants inoculated with
Pseudomonas strain WP3. (Table 3.22A). Inoculation with three strains (Arthrobacter
strain NTC-11, Azospirillum strain BM30 and Pseudomonas strain WP3) showed a
significant increase in root dry weight of non-transgenic plants. Maximum increase in
straw weight was recorded in the plants inoculated with Achromobacter strain A6,
Azospirillum strain BM30, Pseudomonas strain WP3 and Microbacterium strain WNI.
Inoculation of non-transgenic plants with Arthrobacter strain NTC-11 and
Microbacterium strain WNI resulted in a maximum increase in grain dry weight which
was 18.9% and 17.7%, respectively over non-inoculant control (Table 3.22B).
Figure 3-30 Effect of bacterial inoculation on growth of AVP1 transgenic and
non-transgenic wheat grown in micro-plots under natural conditions.

3. Results
80
Table 3.22. Effect of PGPR strains on yield and growth parameters of transgenic
(A) and non-transgenic wheat (B) grown in micro plots during 2012-2013
A
Transgenic wheat
Treatment Cumulative
root length
(cm)
Root dry
weight(g)
Straw
weight
(g)
Grain
weight (g)
Control (Non-inoculated) 20.06B 2.21C 555.6C 404.6D
Achromobacter strain A6 23.28A 2.47AB 556.6 C 415.0C
Arthrobacter strain NTC-11 23.33A 2.58A 659.6A 461.6A
Microbacterium strain WNI 20.66B 2.63A 566.6C 417.0C
Azospirillum strain BM30 22.00A 2.39B 614.6B 413.6C
Pseudomonas strain WP3 23.21A 2.49AB 635.0B 450.3B
LSD(P≤ 0.05) 2.68 0.15 20.7 8.35
B
Non-transgenic wheat
Treatment Cumulative root
length (cm)
Root dry
weight
(g)
Straw
weight
(g)
Grain
weight(g)
Control (Non-inoculated) 19.09C 2.36C 529.6B 389.3C
Achromobacter strain A6 21.33AB 2.56AB 536.6B 412.0B
Arthrobacter strain NTC-11 22.66A 2.60A 603.3A 435.6A
Microbacterium strain WNI 19.33BC 2.43BC 625.0A 406.0B
Azospirillum strain BM30 21.00AB 2.66A 638.3A 410.3B
Pseudomonas strain WP3 22.43AB 2.64A 629.0A 408.6B
LSD(P≤ 0.05) 0.26 0.14 37.56 7.27
Wheat seeds were sown in micro-plots (1.2mx1.2m) maintaining RXR distance 23 cm
and PXP distance15cm. Values are an average of 3 replications.

3. Results
81
Figure 3-31 Effect of bacterial inoculation on straw weight, and grain weight of
transgenic and non-transgenic wheat plants
T.C= Transgenic wheat non-inoculated, NTC= Non-transgenic wheat non-inoculated;
A6= Inoculated with Achromobacter A6; NTC-11=Inoculated with Arthrobacter strain
NTC-11; WN1=Inoculated with Microbacterium strain WNI; BM30=Inoculated with
Azospirillum strain BM30. WP3=Inoculated with Pseudomonas strainWP3
T.Grain…
NT.Grain…
T.Straw…
NT.Straw…
0
100
200
300
400
500
600
700
C A6 NTC-11 WN1 BM30 WP3
Wei
gh
t (g
)
Treatments
T.Grain weight (g) NT.Grain weight(g)
T.Straw weight (g) NT.Straw weight (g)

3. Results
82
3.9 Bacterial Population
Bacterial population (log cfu/g of soil) in the rhizosphere of transgenic and non-
transgenic plants of cotton and wheat was estimated on growth media at 30, 60 and 90
days after sowing (DAS). Soil samples were collected from the rhizosphere of cotton
and wheat plants (transgenic and non-transgenic) grown during 2012 and 2011-2012,
respectively. General bacterial population (cfu) was estimated by serial dilution method
on nutrient agar plates and diazotrophic bacterial population was estimated by MPN
(most probable number) using NFM (semi-solid) medium.
Data showed that bacterial populations (cfu/g soil) were not statistically
different in the rhizosphere of transgenic and non-transgenic plants of cotton and wheat,
however a shift in bacterial population was recorded at different growth stages. In
cotton rhizosphere, maximum bacterial population (5.63 log cfu/g soil) was recorded at
90 DAS among the transgenic cotton plants as compared to 60 and 30 DAS. From wheat
rhizosphere, maximum bacterial population (6.42 log cfu/g soil) was recorded at 60
DAS among transgenic wheat plants as compared to 30 and 90 DAS.
Diazotrophic bacterial population (MPN) did not show any significant
difference in cotton rhizosphere (transgenic and non-transgenic). However, in wheat
rhizosphere, a shift in bacterial population was recorded at 30 and 60 DAS. In wheat
rhizosphere, maximum bacterial population i.e 5.3 log MPN/g soil in transgenic wheat
and 4.8 log MPN/g soil were recorded in non-transgenic wheat at 60DAS.
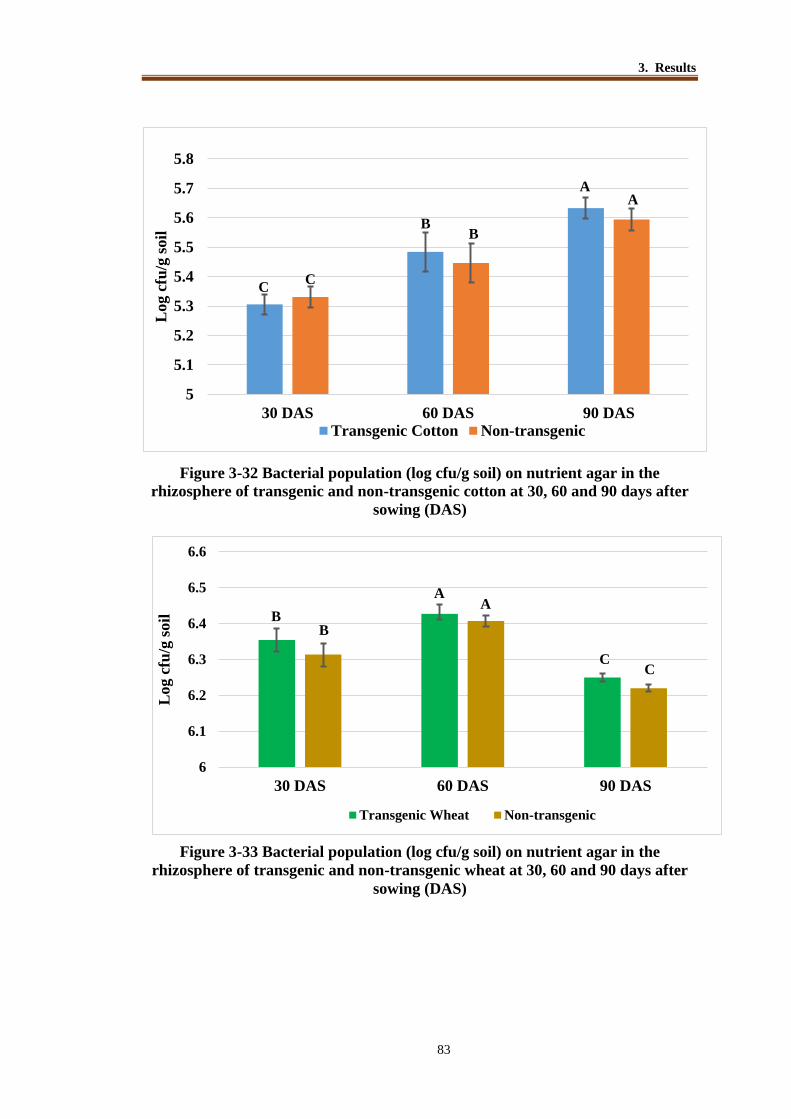
3. Results
83
Figure 3-32 Bacterial population (log cfu/g soil) on nutrient agar in the
rhizosphere of transgenic and non-transgenic cotton at 30, 60 and 90 days after
sowing (DAS)
Figure 3-33 Bacterial population (log cfu/g soil) on nutrient agar in the
rhizosphere of transgenic and non-transgenic wheat at 30, 60 and 90 days after
sowing (DAS)
5
5.1
5.2
5.3
5.4
5.5
5.6
5.7
5.8
30 DAS 60 DAS 90 DAS
Log
cfu
/g s
oil
Transgenic Cotton Non-transgenic
CC
BB
AA
6
6.1
6.2
6.3
6.4
6.5
6.6
30 DAS 60 DAS 90 DAS
Log c
fu/g
soil
Transgenic Wheat Non-transgenic
BB
AA
CC

3. Results
84
Figure 3-34 Bacterial population of diazotrophs (log MPN/g soil in NFM) in the
rhizosphere of transgenic and non-transgenic cotton at 30, 60 and 90 days after
sowing (DAS)
Figure 3-35 Bacterial population diazotrophs (log MPN/g soil in NFM) in the
rhizosphere of transgenic and non-transgenic wheat at 30, 60 and 90 days after
sowing (DAS)
3.0
4.0
5.0
6.0
7.0
8.0
30.DAS 60.DAS 90.DAS
MP
N /
g o
f so
il (
log
va
lues
)
Transgenic cotton Non-transgenic
AA
0
1
2
3
4
5
6
7
8
30 DAS 60 DAS 90 DAS
Transgenic wheat Non-transgenic
A
A A
C C
A

3. Results
85
3.9.1 Real Time PCR Quantification of 16S rRNA and nif H genes
from Rhizospheric Soil
Abundance of 16S rRNA and nif H genes was determined by real time PCR from the
rhizosphere soil samples collected at different growth stages (35 and 90 DAS). The soil
samples were collected in two replicates from rhizosphere of cotton (transgenic and
non-transgenic) plants during the year 2012 and from wheat (transgenic and non-
transgenic) plants grown during 2011-12. These samples were divided into two group’s
i.e Group A and Group B. Each sample was given a specific ID that represents its source
i.e TC for transgenic cotton, NTC for non-transgenic cotton whereas ‘TW’ for
transgenic wheat and ‘NTW’ for non-transgenic wheat.
In Group A, that belonged to cotton rhizospheric soil samples collected during
the year 2011, maximum copy number log 6.39 copies/g soil of 16S rRNA gene was
recovered in sample TC (transgenic cotton) collected at 90 DAS, followed by log 6.36
copies/g soil in sample NTC (non-transgenic cotton) collected at 90 DAS. A relatively
lower copy number 16S rRNA gene was recorded in samples which were collected at
35 DAS. nifH gene abundance data showed that population of diazotrophic bacteria was
maximum i.e log 5.85 copies/g soil in non-transgenic cotton soil sample (NTC) at 35
DAS and it was minimum in transgenic cotton rhizosphere at 35 DAS (Table 3.23).
The group B, belonged to rhizospheric soil samples from wheat rhizosphere
grown during 2009-10. Highest copy number (log 6.79 copies/g soil) of 16S rRNA gene
was estimated in non-transgenic wheat at 90 DAS whereas relatively low copy number
(log 6.54.79 copies/g soil) was observed in transgenic wheat rhizosphere at 90 DAS.
Population of diazotrophic bacteria indicated by copy number of nif H, was highest
(5.84 copies/g soil) in transgenic wheat at 35 DAS and slightly low in non-transgenic
wheat at 35 DAS. A low copy number of nifH was observed in both transgenic and non-
transgenic wheat at 90 DAS (Table 3.23).

3. Results
86
Table 3.23 Relative gene abundance (copy number) of bacterial 16S rRNA and
nif H genes in the rhizospheric soil revealed by real time PCR
Cotton
Samples
Group. A*
Sample.ID Sampling
time
16S rRNA gene
copies/ g of soil
(log values)
nif H gene
copies/ g of soil
(log values)
T-C 35 DAS 6.04 ± 0.30 5.66 ± 1.72
NT-C 35 DAS 6.22 ± 1.02 5.85 ± 2.90
T-C 90 DAS 6.39 ± 0.99 5.41 ± 1.70
NT-C 90 DAS 6.367 ± 2.1 5.39 ± 2.68
Wheat
Group. B**
Sample.ID Sampling
time
16S rRNA gene
copies/ g of soil
(log values)
nif H gene
copies/ g of soil
(log values)
T-W 35 DAS 6.54 ± 0.48 5.94 ± 0.98
NT-W 35 DAS 6.63 ± 0.87 5.83 ± 1.60
T-W 90 DAS 6.62 ± 2.14 5.26 ± 1.32
NT-W 90 DAS 6.79 ± 1.51 5.16 ± 2.61
*: Group A includes soil samples from AVP1 transgenic (TC) and non-transgenic cotton
(NT-C) rhizosphere, collected at different days after sowing (DAS) during 2011.
**:Group B includes soil samples from AVP1 transgenic(TW) and non-transgenic
wheat (NT-W) rhizosphere collected at different days after sowing (DAS) during 2009-
10. Given values are an average of three replicates with standard deviation

3. Results
87
Figure 3-36 Real time quantification of 16S rRNA and nifH gene from
rhizosphere of AVP1 transgenic cotton and wheat
TC= AVP1 transgenic cotton; NTC= Non-transgenic cotton, TW= AVP1 transgenic
wheat; NTW=Non-transgenic wheat
3.9.2 Detection of Root Exudates in the Rhizosphere of AVP1
Transgenic Cotton and Wheat
Organic acids (acetic acid, citric acid, malic acid and oxalic acid) produced as root
exudates in the rhizosphere of AVP1 transgenic cotton and wheat were also
investigated. The soil samples were collected in two replicates from rhizosphere of
cotton (transgenic and non-transgenic) plants during the year 2012 and from wheat
(transgenic and non-transgenic) plants grown during 2011-12. Maximum amount of
acetic acid (8.89 µg/mL) was detected in transgenic cotton rhizosphere. Malic acid was
detected in smaller amounts in all soil samples. A difference was observed in the
production of oxalic acid that was relatively higher in transgenic cotton rhizosphere
(6.72 µg/mL) whereas in non-transgenic cotton rhizosphere it was present relatively in
low amounts (3.84 µg/mL). Over all transgenic rhizosphere soil showed a relatively
higher amounts of organic acid production as compared to non-transgenic rhizosphere.

3. Results
88
Table 3.24. Detection of organic acids produced* as root exudates in rhizosphere
of AVP1 transgenic and non-transgenic cotton and wheat
A
Treatment Acetic acid Oxalic Acid Citric acid Malic acid
1. Transgenic
cotton
8.89 ± 1.30A 6.72 ± 2.31A 2.06 ± 0.71A 0.73± .05A
2. Non-transgenic
cotton
8.15 ± 1.12B 3.84 ± 1.20B 1.99 ± 0.45B 0.71± 0.04A
LSD (P ≤ 0.05) 0.03 0.02 0.01 N.S
B
Treatment Acetic acid Oxalic Acid Citric acid Malic acid
3. Transgenic
wheat
8.62 ± 1.60A 5.43 ± 2.2A 2.18 ± 0.95A 0.94± 0.01A
4. Non-transgenic
wheat
8.25 ± 2.14B 3.27 ± 1.5B 2.16 ± 0.20A 0.98± 0.02B
LSD (P ≤ 0.05) 0.04 0.02 NS 0.02
*Cotton (2012) and wheat (2011-12) plants were uprooted with adhering soil in three
replicates. Roots along with attached soil were washed to extract the roots exudates.
Extract was analyzed on HPLC to determine the organic acids present in exudates.
Values are an average of three replicates with standard deviation.

3. Results
89
Figure 3-37 Organic acid production in rhizosphere of AVP1 transgenic and non-
transgenic cotton and wheat
T.C= Transgenic cotton rhizosphere soil; NT.C= Non-transgenic cotton rhizosphere
soil; T.W= Transgenic wheat rhizosphere soil; NT.W= Non-transgenic wheat
rhizosphere soil.
Malic acid
Citric acid
Oxalic Acid
Acetic acid
0
1
2
3
4
5
6
7
8
9
TC NTC TW NTW
Org
an
ic a
cid
(µ
g/m
L)
Treatments
Malic acid
Citric acid
Oxalic Acid
Acetic acid

3. Results
90
3.10 Diversity of Diazotrophic Bacteria Determined by PCR
Amplification of Partial nifH gene from Soil DNA
Diversity of diazotrophic bacteria from the rhizosphere of AVP1 transgenic cotton and
wheat was determined by PCR amplification of partial nifH gene (~360bp) from DNA
directly extracted from the rhizosphere soil. Four clone libraries (TC, NTC, TW, NTW)
of nifH gene were constructed and 50 clones from each library were selected randomly
and sequenced. On the bases of sequence length and sequence quality 159 clones
(TC=41/50, NTC=39/50, TW=38/50, NTW=34/50) provided enough sequence
information required for comparison at NCBI databank. In all four clone libraries,
sequence related to non-culturable diazotrophic bacteria were high constituting about
78% in TC, 74% in NTC, 78% in TW, and 79% in NTW (Figure 3-39, 3-40).
From the clone library ‘TC’ belonging to diazotrophic bacteria in the
rhizosphere of AVP1 transgenic cotton plants, 41 readable sequences were obtained
which showed 82% of library coverage. Among 41 readable sequences, 32 sequences
(82%) showed homology with non-culturable diazotrophic bacterial sequences and 9
sequences (22%) with culturable diazotrophic bacteria in the databank. Among
culturable diazotrophic bacteria 7% sequences showed homology with Anabaena, 2%
with Azoarcus sp., 2% with Azospirillum, 5% with Azotobacter chroococcum, 2% with
Bradyrhizobium japonicum and 2 % with Pseudomonas sp. (Figure 3-39)
Library ‘NTC’ contained sequences of diazotrophic bacteria retrieved from
non-transgenic cotton rhizosphere. In this library 39 readable sequences were obtained
with library coverage of 78%. Among these readable sequences, 29 sequences (74%)
showed homology with non-culturable diazotrophic bacteria, 10 sequences (26%)
showed homology with culturable diazotrophic bacteria in the databank (Figure 3-39).
Among culturable diazotrophic bacteria, 5% sequences showed homology with
Anabaena, 3% with Azohydromonas, 3% with Azospira restricta, 3% with Azospirillum
brasilence, 8% with Bradyrhizobium japonicum, 3 % with Pseudomonas sp. and 3%
with Zoogloea oryzae. (Table 3.25)
Library ‘TW’ contained sequences from rhizosphere of AVP1 transgenic wheat.
In this library there were 38 readable sequences with library coverage 78%. Among
readable sequences 29 sequences (76%) showed homology with non-culturable
diazotrophic bacteria and 9 sequences (24%) with culturable diazotrophic bacteria.

3. Results
91
Among these culturable bacteria 10% sequences showed homology with
Agrobacterium, 3% with Azospirillum, 5% Bradyrhizobium, 3% with Pseudomonas,
and 3% Rhizobium.
Library ‘NTW’ contained sequences from the rhizosphere of non-transgenic
wheat. This library contained 34 readable sequences with 68% library coverage. In
these readable sequences 27sequences (79%) showed homology with non-culturable
diazotrophic bacteria and 7 sequences (21%) showed homology with culturable
diazotrophic bacteria. Among culturable diazotrophic bacteria 3% sequences showed
homology with Agrobacterium, 3% with Azospirillum, 12% with Pseudomonas and 3%
with Zoogloea. (Table 3.30)
Figure 3-38 Amplification of nifH gene from soil DNA extracted from AVP1
transgenic and non-transgenic cotton (A) and wheat (B)
Figure A: Lane 1 1kb marker, Lane 2 -Ve control, Lane 3 transgenic cotton (TC1), Lane
4 non-transgenic cotton (NTC1), Lane 5 transgenic cotton (TC2), Lane 6 non-
transgenic cotton (NTC2).
Figure B: Lane 1 1kb marker, Lane 2 -Ve control, Lane 3 transgenic wheat (TW1) ,
Lane 4 non-transgenic wheat (NTW1), Lane 5 transgenic wheat (TW2), Lane 6 non-
transgenic wheat (NTW2),

3. Results
92
Table 3.25 Diversity of diazotrophic bacterial sequences in the rhizosphere of
AVP1 transgenic and non-transgenic cotton
Description AVP1 transgenic cotton
(TC library)
Non-transgenic
cotton
(NTC library)
A. Total clone sequences 41 39
B. (i) Non-culturable diazotrophs 32 (78%) 29 (74%)
(ii) Culturable diazotrophs 9 (22%) 10 (26%)
C. (i) Anabaena sp. 3 (7%) 2 (5%)
(ii) Azoarcus sp. 1 (2%) 0
(iii) Azohydromonas australica 0 1 (3%)
(iv) Azospira restricta 0 1 (3%)
(iv) Azospirillum zeae 1 (3%) 1 (2%)
(iv) Azotobacter chroococcum 1 (3%) 0
(iv) Bradyrhizobium japonicum 1 (2%) 3 (8%)
(iv) Pseudomonas stutzri 1 (3%) 1 (3%)
(iv) Zoogloea oryzae 0 1 (2%)
TC’ and ‘NTC’ nifH gene libraries constructed from amplified nifH gene from
rhizosphere of AVP1 transgenic and non-transgenic cotton, respectively.

3. Results
93
Figure 3-39 Distribution of diazotrophic bacterial sequences in the rhizosphere
of AVP1 transgenic (A) and non-transgenic cotton (B).

3. Results
94
Table 3.26 Identification of culturable diazotrophic bacterial sequences in the
rhizosphere of AVP1 transgenic cotton
No.
Sequence IDa
& Accession
No.
Closest match
in NCBI
Sequence
similarity
(%)
Origin References
1 P1- 18
(LN736030.1)
Anabaena sp.
(HM063719.1)
99 Rice paddies
(India)
Un-published
2 P1- 15
(LN736031.1)
Anabaena sp.
(AJ716235.1)
92 [206]
3 P3-13
(LN736032.1)
Anabaena sp.
(JN1624601.1)
97 [207]
4 P1- 14
(LN736033.1)
Azospirillum zea
(JN162478.1)
94 Zea mays
(India)
[207]
5 P1- 22
(LN736034.1)
Azotobacter
chroococcum
(EF634050.1)
97 Agricultural
soil
(Italy)
unpublished
6 P3- 3
(LN736035.1)
Azoarcus
(AY6010541)
86 Kallar grass
(China)
[208]
7 P3- 4
(LN736036.1)
Bradyrhizobium
Japonicum
(HQ3356861)
87 Radish
paddy soil
(Nether
land)
[209]
8 P1-6
(LN736037.1)
Pseudomonas
stutzri
(CP000304.1)
73 [211]
Identification by NCBI BLASTN of nif H sequences obtained from the rhizosphere of
AVP1 transgenic cotton grown in controlled condition.
a:Sequence ID represents the clones obtained from nifH gene library ‘’TC’’

3. Results
95
Table 3.27 Identification of culturable diazotrophic bacteria detected in the
rhizosphere of non-transgenic cotton
No. Sequence IDa
& Accession
No.
Closest match in
NCBI
Sequence
similarity
(%)
Originc References
1 P2- 11
(LN736038.1)
Azohydromonas
australica
(JN162460.1)
99 Saline alkaline
soil India
[210]
2 P2- 3
(LN736039.1)
Azospirillum
brasilense
(EU048106.1)
99 Aquatic
terrestrial
environments.
Brazil
[211]
3 P2- 7
(LN736040.1)
Pseudomonas sp.
(HM063793.1)
89 Pulp and paper
wastewater
China
Unpublished
4 P2-4
(LN736041.1)
Zoogloea oryzae
(HQ335686.1)
89 Rice paddy soil
in Tokyo
[209]
5 P2-14
(LN736042.1)
Azospira restricta
(HQ190167.1)
89 Ground water
China
Unpublished
6 P3- 13
(LN736043.1)
Azoarcus,
(AY6010541)
86 [177]
7 P2-26
(LN736044.1)
Bradyrhizobium
japonicum
(EF583593.1)
93
Radish paddy
soils
Sweden
[212]
8 P2-23
(LN736045.1)
Bradyrhizobium
japonicum
(GQ289562.1)
92
Unpublished
china
9 P4-11
(LN736046.1)
Bradyrhizobium
japonicum
(GQ289582.1)
92 Unpublished
china
10 P4-20
(LN736047.1)
Anabaena sp.
(L04598.1)
93 Variety
environment
china
Unpublished
11 P4-7
(LN736048.1)
Anabaena sp.
(L04499.1)
96 [213]
a:Sequence ID represents the clones obtained from nifH gene library ‘’NTC’’

3. Results
96
Table 3.28 List of uncultured diazotrophic bacteria from AVP1 transgenic cotton
rhizosphere
# Clone Acc. No Source*
1 P3-5 KH744001.1
saline alkaline soil
2 P3-30 KH744002.1
3 P3-20 KH744003.1
4 P3-12 KH744004.1
5 P3-14 KH 744005.1
6 P3-16 KH 744006.1 cotton rhizospheric soil
7 P1-11 KH 744007.1
8 P1-29 KH 744008.1 soils
9 P1-1 KH 744009.1
10 P1-20 KH 744010.1
11 P3-6 KH 744011.1 paddy soil from China
12 P3-15. KH 744012.1 Marine sediments
13 P1-30 KH 744013.1 Oryza-root tissues
14 P3-19 KH 744014.1 Tea plants
15 P3-29 KH 744015.1 Bioremediation soil
16 P3-7 KH 744016.1
17 P3-21 KH 744017.1 Forest soil
18 P1-2 KH 744018.1 Thick mudflat sediment.
19 P1-23. KH 744019.1 Soybeans .
20 P3-23 KH 744020.1 Leaf surface
21 P3-24 KH 744021.1
22 P1-25 KH 744022.1 Indian Punjab soil
23 P3-9 KH 744023.1 California soil
24 P1-26 KH 744024.1 Rhizosphere
25 P1-19 KH 744025.1 Sea water
26 P3-1 KH 744026.1
27 P3-8 KH 744027.1
28 P1-16 KH 744028.1
29 P1-28 KH 744029.1 Microbial mat of salted soil
30 P1-5 KH 744030.1 Salt marsh elevation
31 P1-21 KH 744031.1 Chesapeake bay
*Uncultured diazotrophic source already reported in databank

3. Results
97
Table 3.29 List of uncultured diazotrophic bacterial sequences from non-
transgenic cotton rhizosphere
# Clone Acc. No Source*
1 P4-9 KH744032.1 cotton rhizosphere
2 P2-1 KH744033.1
3 P4-9 KH744034.1
4 P2-2 KH744035.1 alkaline soil sorghum
5 P4-28 KH744036.1
6 P4-2 KH744037.1
7 P4-15 KH744038.1
8 P4-12 KH744039.1 salt marsh soils
9 P4-22 KH744040.1 soil
10 P2-12 KH744041.1
11 P2-8 KH744042.1
alkaline soils
12 P4-17 KH744043.1
13 P2-28 KH744044.1
14 P4-25 KH744045.1
15 P2-16 KH744046.1 rice filed
16 P4-13 KH744047.1 paddy soils
17 P4-8 KH744048.1
18 P2-6 KH744049.1 roots of Oryza sative
19 P2-22 KH744050.1
20 P4-1 KH744051.1
Kollumerwaard bulk soil
21 P4-16 KH744052.1
22 P4-19 KH744053.1
23 P4-18 KH744054.1
24 P2-5 KH744055.1 Baltic sea
25 P2-9 KH744056.1 Turf grass soil
26 P4-30 KH744057.1 Forest soil
27 P2-15 KH744058.1 Salted pond microbial mat
28 P4-10 KH744059.1 Cotton rhizosphere
29 P2-20 KH744060.1
*Uncultured diazotrophic source already reported in databank

3. Results
98
Table 3.30 Diversity of diazotrophic bacteria in the rhizosphere of AVP1
transgenic and non-transgenic wheat
Description AVP1 transgenic
Wheat (TCW)
Non-transgenic Wheat
(NTW)
A. Total clone sequences 38 (79%) 34 (76%)
B. (i) Non-culturable diazotrophs 29 (76%) 27 (79%)
(ii) Culturable diazotrophs 9 (24%) 7 (21%)
C. (i) Agrobacterium tumefaciens
4 (8%) 1 (3%)
(ii) Azospirillum sp.
1 (3%) 1 (3%)
(iii) Bradyrhizobium japonicum
2 (5%) 0
(iv) Pseudomonas sp.
1 (3%) 4 (12%)
(v) Rhizobium sp.
1 (3%) 0
(vi) Zoogloea sp.
0 1 (3%)
TCW’ and ‘NTW’ nifH gene libraries constructed from amplified nif H gene from
rhizosphere of AVP1 transgenic and non-transgenic wheat, respectively.

3. Results
99
Figure 3-40 Distribution of diazotrophic bacteria in the rhizosphere of AVP1
transgenic (A) and non-transgenic wheat (B)
A
B

3. Results
100
Table 3.31 Identification of culturable diazotrophic bacterial sequences in the
rhizosphere of AVP1 transgenic wheat
No.
Sequence ID
& Accession
No.
Closest match in
NCBI
Sequence
similarity
(%)
Origin References
1 P7-26
(LM794549.1)
Rhizobium
gallicum
(EF634041.1)
99 "agricultural
soil" Italy
Un-published
2 P7-27
(LM794550.1)
Pseudomonas sp.
(FJ822997.1)
98 sugarcane
rhizosphere
from china
Un-published
3 P7-29
(LM794551.1)
Agrobacterium
tumefaciens
(FJ822995.1)
98 sugarcane
rhizosphere
from china
[214]
4 P5-22
(LM794552.1)
Azospirillum spp.
(GU256447.1)
95 china Un-published
5 P7-3
(LM794553.1)
Pseudomonas sp.
(FJ822997.1)
95 sugarcane in
china
Un-published
6 P5-28
(LM794554.1)
Azospirillum
brasilense
(GQ161230.1)
94 cereal crops
grown in
Greece
[215]
7 P5-6
(LM794555.1)
Azospirillum
brasilense
(GQ161230.1)
93 China [216]
8 P5-14
(LM794556.1)
Bradyrhizobium
japonicum
(GQ289577.1)
90 "reddish paddy
soil
[217]
9 P5-12
(LM794557.1)
Bradyrhizobium
japonicum
(GQ289567.1)
86 "reddish paddy
soil
Un-published
Identification by NCBI BLASTN of nif H sequences obtained from the rhizosphere of
AVP1 transgenic and non-transgenic wheat grown in controlled condition.
Sequence ID represents the clones obtained from nifH gene library ‘’TW’’
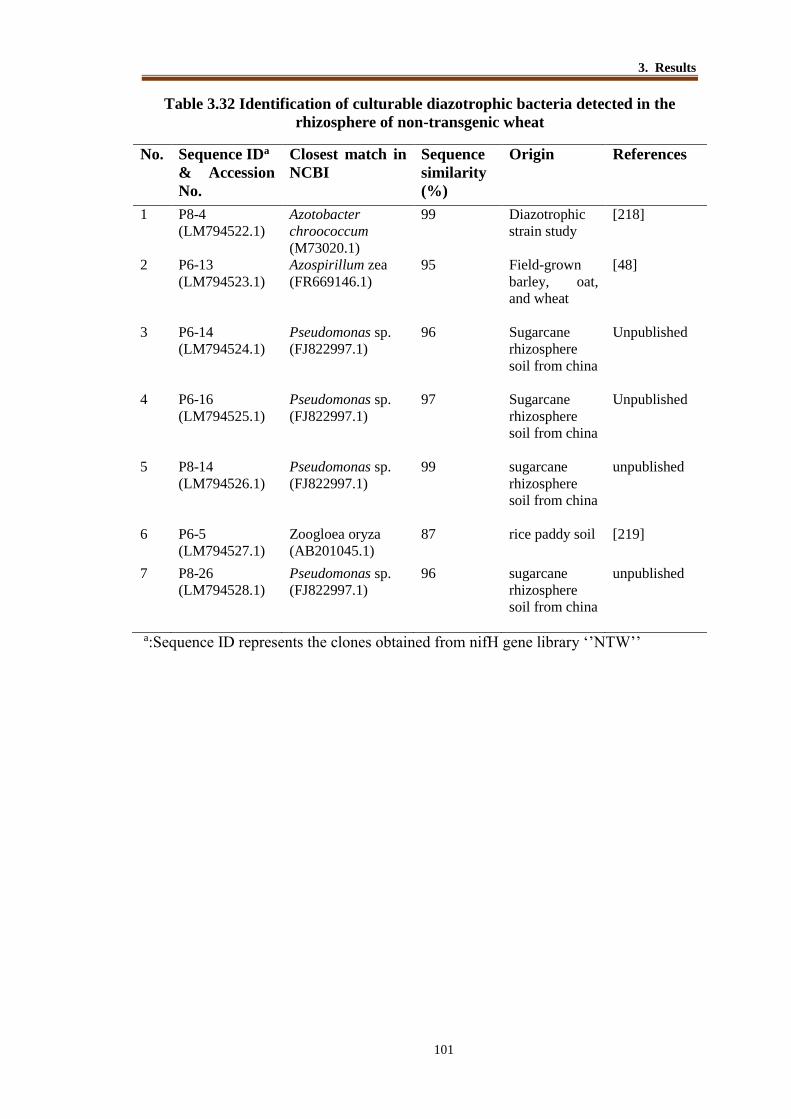
3. Results
101
Table 3.32 Identification of culturable diazotrophic bacteria detected in the
rhizosphere of non-transgenic wheat
No. Sequence IDa
& Accession
No.
Closest match in
NCBI
Sequence
similarity
(%)
Origin References
1 P8-4
(LM794522.1)
Azotobacter
chroococcum
(M73020.1)
99 Diazotrophic
strain study
[218]
2 P6-13
(LM794523.1)
Azospirillum zea
(FR669146.1)
95 Field-grown
barley, oat,
and wheat
[48]
3 P6-14
(LM794524.1)
Pseudomonas sp.
(FJ822997.1)
96 Sugarcane
rhizosphere
soil from china
Unpublished
4 P6-16
(LM794525.1)
Pseudomonas sp.
(FJ822997.1)
97 Sugarcane
rhizosphere
soil from china
Unpublished
5 P8-14
(LM794526.1)
Pseudomonas sp.
(FJ822997.1)
99 sugarcane
rhizosphere
soil from china
unpublished
6 P6-5
(LM794527.1)
Zoogloea oryza
(AB201045.1)
87 rice paddy soil
[219]
7 P8-26
(LM794528.1)
Pseudomonas sp.
(FJ822997.1)
96 sugarcane
rhizosphere
soil from china
unpublished
a:Sequence ID represents the clones obtained from nifH gene library ‘’NTW’’

3. Results
102
Table 3.33: List of uncultured diazotrophic bacteria from AVP1 transgenic
wheat rhizosphere
No. Clone Acc. No Source*
1 P5-3 LM794520.1 Roots of rice
paddy soil china 2 P7-24 LM794521.1
3 P5-23 LM794523.1
4 P7-22 LM794524.1 Agricultural soil Switzerland
5 P7-7 LM794526.1 Root and stem of field-
grown maize
6 P7-25 LM794527.1 "Saline-alkaline soil"
7 P7-1 LM794528.1
8 P5-4 LM794530.1 Free living diazotroph of soil
9 P7-14 LM794531.1 "Saline-alkaline soil"
10 P5-16 LM794533.1 "Kollumerwaard bulk soil"
soil neatherland 11 P7-20 LM794534.1
12 P5-26 LM794535.1
13 P7-13 LM794537.1
14 P7-19 LM794538.1
15 P5-9 LM794540.1
16 P5-25 LM794541.1
17 P7-2 LM794542.1
18 P7-17 LM794544.1
19 P7-21 LM794545.1
20 P5-7 LM794547.1
21 P5-17 LM794548.1
22 P7-6 LM794549.1
23 P5-15 LM794551.1 Free living diazotroph of soil
24 P5-27 LM794552.1 Montipora flabellata
25 P5-30 LM794554.1 Microbial mat on sandy
intertidal beach
26 P7-9 LM794555.1
27 P7-30 LM794556.1
28 P5-8 LM794558.1 Colombian Amazon Region
29 P5-1 LM794559.1 Malaysian soil
30 P5-19 LM794561.1 Mudflat mesocosms
31 P5-20 LM794562.1 "hot spring"
*Uncultured diazotrophic source already reported in databank

3. Results
103
Table 3.34 List of uncultured diazotrophic bacteria from AVP1 transgenic wheat
rhizosphere
Clone Acc.No Source*
1 P6-12 LM794563.1 Paddy soil in china with long term organic
management 2 P6-3 LM794565.1
3 P8-29 LM794566.1
4 P8-1 LM794568.1 Lelystad bulk soil" The Netherlands
5 P8-9 LM794569.1 Kollumerwaard bulk soil"
6 P8-2 LM794570.1
7 P8-7 LM794572.1
8 P6-23 LM794573.1
9 P6-7 LM794575.1
10 P8-16 LM794576.1
11 P8-28 LM794577.1
12 P8-3 LM794579.1 Falls Lake, North Carolina"
13 P8-17 LM794580.1 Rice roots
14 P8-5 LM794582.1
15 P6-21 LM794583.1
16 P6-20 LM794584.1 Rhizospheric soil of sorghum"
17 P8-10 LM794586.1 Bioremediated soil at Zhongyuan oil field
18 P6-18 LM794587.1 Turfgrass established soil"
19 P6-24 LM794589.1 Rice roots
20 P8-19 LM794590.1 Chesapeake Bay, Station 2, surface"
21 P6-1 LM794591.1
rhizospheric soil of sorghum"
22 P6-22 LM794593.1 Tea plantation soil
23 P8-8 LM794594.1 Sandy intertidal beach
24 P6-8 LM794596.1
25 P6-2 LM794597.1 Soil Mexico
26 P6-25 LM794598.1 Soil
27 P8-25 LM794600.1 Thick mudflat sediment"
*Uncultured diazotrophic source already reported in databank

3. Results
104
Figure 3-41 Phylogenic tree constructed from nifH gene sequences retrieved
from AVP1 transgenic and non-transgenic cotton
Group I
Group II
Group IV
Group V
Group III
Sequences from AVP1
transgenic Cotton
Sequences from non-
transgenic cotton

3. Results
105
Figure 3-42 Phylogenic tree constructed from nifH gene sequences of AVP1
transgenic and non-transgenic wheat
Group I
Group II
Group IV
Group V
Group III
Sequences from AVP1 transgenic wheat
Sequences from non-transgenic wheat

3. Results
106
Genomic DNA from soil
3.11 Bacterial Diversity Revealed by Pyrosequencing of 16S
rRNA Gene Amplified from Soil DNA
Diversity of culturable and uncultured bacteria from the rhizosphere of AVP1 transgenic
and non-transgenic cotton and wheat plants was investigated by amplification and
sequence analysis of 16S rRNA gene from soil DNA. Rhizosphere soil samples were
collected from AVP1 transgenic cotton and wheat along with non-transgenic plants as
their control from inoculation experiments conducted during 2012 (cotton) and 2010-
12 (wheat), respectively
Soil DNA was extracted from 4 rhizosphere soil samples i.e rhizospheric soil
from AVP1 transgenic cotton, non-transgenic cotton, AVP1 transgenic wheat and non-
transgenic wheat (Figure 43). DNA was extracted in two replicates of each rhizosphere
soil sample and total 8 samples were processed. 16S rRNA gene was amplified from
these 8 soil DNA samples (Figure 44) by using a forward primer ‘454 Primer A’ (799F
with an adapter sequence and specific barcode sequence) with 8 different barcodes for
all 8 soil DNA samples. ‘454PrimerB’ (1492R with a specific adaptor sequence) were
used as a reverse primer
Figure 3-43 DNA extracted from the rhizosphere soil of cotton and wheat
Lane 1, 1kb marker; Lane 2, soil DNA from transgenic cotton rhizosphere;
Lane 3, soil DNA from non-transgenic cotton rhizosphere; Lane 4, soil DNA from
transgenic wheat rhizosphere; Lane 5, soil DNA from non-transgenic wheat rhizosphere
1 2 3 4 5
3000 bp
1000 bp

3. Results
107
Figure 3-44 PCR amplification of 16S rRNA gene from the rhizosphere of
AVP1-transgenic and non-transgenic cotton (A) and wheat (B) using barcoded
A: Lane 1, 1kb marker; Lane 2, +Ve control; Lane 3, -Ve control, Lane 4, transgenic
cotton (TC1) Lane 5, non-transgenic cotton (NTC1), Lane 6, transgenic cotton (TC2),
Lane 7, non-transgenic cotton (NTC2)
B: Lane 1, 1kb marker; Lane 2, +Ve control; Lane 3, -Ve control, Lane 4, transgenic
wheat (TW1) Lane 5, non-transgenic wheat (NTW1), Lane 6, transgenic wheat (TW2),
Lane 7, non-transgenic wheat (NTW2)
3.11.1 Barcoded Pyrosequencing of 16S rRNA Gene, DNA Sequence
Processing and Taxonomic Analysis
From 8 soil DNA samples, 16S rRNA gene sequences covering V5, V6, and V7 were
amplified with PCR and pyrosequencing of these barcoded amplicons was performed.
The obtained sequences were analyzed through the Ribosomal Database Project (RDP)
pyrosequencing pipeline (http://pyro.cme.msu.edu). A total of 346531 roots/sequences
were obtained from all soil samples, with an average of 43316±14500 roots per sample
(Table 3.31). All these roots/sequences according to the individual sample treatment
were assigned hierarchy view at 80% confidence threshold. Collectively, 255753
sequences of 16S rRNA gene were related to bacteria, 31137±2840 sequences related
to Archaea, 44532 sequences belonged to unclassified bacteria (Table 3.35)
3.11.2 Bacterial Diversity in Cotton Rhizosphere
Among the retrieved sequences 127747 sequences of 16S rRNA were related to
bacteria, 8128 were related to Archaea and 22964 belonged to unclassified bacteria.
Among these sequences on an average 47242±3690 sequences were detected in the
1 2 3 4 5
1 2 3 4 5
700
700 PCR product
(Partial 16S rRNA gene)
A
B
PCR product (16S rRNA gene)
700 bp
700 bp

3. Results
108
rhizosphere of AVP1 transgenic cotton and about 47882±3198 sequences were detected
from non-transgenic cotton rhizosphere. All the 19 bacterial phyla detected on the basis
of 16S rRNA gene sequencing were represented in the rhizosphere of both transgenic
and non-transgenic cotton plants. In the rhizosphere of AVP1 transgenic cotton plants,
sequences of Proteobacteria were most abundant (16188 ± 3282 sequences), followed
by Crenarchaeota (14205 ± 2551 sequences) and Firmicutes (4103 ± 1820 sequences).
Sequences of the same three phyla were also abundant in the rhizosphere of non-
transgenic plants as 14914 ± 2527 sequences belonged to Proteobacteria, followed by
17390 ± 782 sequences of Crenarchaeota and 5651 ± 3885 sequences related to
Firmicutes (Table 3.36). Further analysis of retrieved 16S rRNA sequences indicated
occurrence bacterial genera in the rhizosphere of transgenic and non-transgenic cotton.
3.11.3 Abundance of Bacterial Classes in Cotton Rhizosphere Soil
16S rRNA sequences belonging to bacterial classes were found in cotton rhizosphere
soil. Among these classes on an average 42±4 were detected in transgenic cotton
rhizosphere and 38±07 classes were detected in non-transgenic rhizosphere soil.
Majority of classes were common in both the soils (transgenic and non-transgenic
rhizosphere). Results showed that in AVP1 transgenic cotton rhizosphere sequences
belonged to bacterial class Thermoprotei were most abundant (36%), followed by
Alpha-proteobacteria (16%), Beta-proteobacteria ( 6%), Gamma-proteobacteria (11%),
Actinobacteria (10%), Bacilli (10%), Delta-proteobacteria (5%), Sphingobacteria(4%),
Acidobacteria (1%) and Clostridia (1%). In non-transgenic cotton rhizosphere
sequences belonging to bacterial class ‘Thermoprotei’ were also most abundant (43%),
followed by Alphaproteobacteria (13%), Betaproteobacteria (7%),
Gammaproteobacteria (11%), Actinobacteria (4%), Bacilli (13%), Delta-proteobacteria
(4%), Sphingobacteria (3%), Acidobacteria GP3(1%) and Clostridia (1%)(Figure 3-46)
3.11.4 Abundance of Bacterial Genera in the Rhizosphere of Cotton
Data analysis at genus level showed that in transgenic cotton rhizosphere bacterial
sequences related to 344±5 genera were found and sequences related to 321±12 genera
were detected in non-transgenic cotton rhizosphere soil. Among these, sequences of
240 genera were commonly detected in transgenic and non-transgenic rhizosphere. This
indicated that majority of detected genera were common in both root systems i.e
transgenic and non-transgenic cotton rhizosphere (Table 3.37). 16S rRNA sequences

3. Results
109
related to 60 genera were found only in the rhizosphere of transgenic cotton (Table
3.38) and 16S rRNA sequences related to 33 genera were found only in the rhizosphere
of non-transgenic cotton. Bacterial genera that have been reported as PGPR were also
detected in the rhizosphere of transgenic and non-transgenic wheat (Table 3.39).
Comparison of the number of sequences retrieved from transgenic and non-transgenic
plants belonging to specific groups indicated quantitative differences but all major
bacterial groups were present in the rhizosphere of both type of plants.

3. Results
110
Table 3.35 16S rRNA sequences retrieved from rhizosphere of AVP1 transgenic
cotton and wheat with non-transgenic control
*Numbers in each row represent the abundance of sequences at different hierarchal
levels of bacterial domain mentioned in each column
a =TC1, transgenic cotton. b=NTC1, non-transgenic cotton, (collected during 2012) c=TC2, transgenic cotton, d =NTC2 non-transgenic cotton (collected during 2012) e=TW1 transgenic wheat f=NTW1 non-transgenic wheat (collected during 2011-12) g=TW2 transgenic wheat h= NTW2 non-transgenic wheat (collected during 2011-12)
Treatment Roots Unclassif
ied roots
Bacterial
sequences
Unclassified
bacterial
sequences
No. of
phyla
Total
genera
1. TC1a
(2012) 49852 47 34938 5887 19 348
2. NTC1b
(2012) 50144 64 32158 5415 23 330
3. TC2c
(2012) 44633 49 33734 6838 21 340
4.NTC2d
(2012) 45620 66 26917 4824 20 312
Total (cotton) 190249 226 127747 22964 99 1330
5. TW1e
(2011-12) 45318 62 34353 6027 21 310
6. NTW1f
(2011-12) 40456 31 33640 5108 20 315
7. TW2g
(2011-12) 36637 30 31654 6272 18 276
8. NTW2h
(2011-12) 33871 25 28359 4161 20 298
Total (wheat) 156282 148 128006 21568 79 1199

3. Results
111
Table 3.36 Abundance of 16S rRNA sequences belonging to different phyla in the
rhizosphere of AVP1 transgenic and non-transgenic cotton
No. Phylum Sequences retrieved from
AVP1 transgenic cotton
rhizosphere
Sequences retrieved
from non-transgenic
cotton rhizosphere
1 Proteobacteria 16188 ± 3282 14914 ± 2527
2 Crenarchaeota 14205 ± 2551 17390 ± 782
3 Firmicutes 4103 ± 1820 5651 ± 3885
4 Unclassified bacteria 5887 ± 1590 5415 ± 1027
5 Actinobacteria 3819 ± 1156 1783 ± 161
6 Bacteroidetes 2349 ± 632 1844 ± 397
7 Acidobacteria 1036 ± 475 824 ± 84
8 Planctomycetes 692 ± 842 5140 ± 7172
9 Chloroflexi 572 ± 340 259 ± 9
10 Euryarchaeota 440 ± 283 3787 ± 4646
11 Verrucomicrobia 379 ± 62 404 ± 97
12 Nitrospira 352 ± 12 218 ± 245
13 Armatimonadetes 267.5 ± 303 50 ± 18
14 Gemmatimonadetes 247 ± 77 259 ± 37
15 Deinococcus thermus 128 ± 72 94 ± 10
16 Chlamydiae 76 ± 14 72 ± 6
17 Incertaesedis 42 ± 6 122 ± 158
18 Chlorobi 5 ± 3 2 ± 2
19 Elusimicrobia 2 ± 1 61 ± 83
DNA extracted from rhizosphere soil AVP1 transgenic and non-transgenic cotton was
used for pyrosequencing analysis of 16S rRNA gene. Given numbers are average of 2
replicates.

3. Results
112
Figure 3-45 Abundance of bacterial phyla in the rhizosphere of AVP1 transgenic
and non-transgenic cotton.
38191783
2349
1844
1420517390
41035651
1618814914
5887 5415
0
5000
10000
15000
20000
25000
30000
35000
40000
45000
50000
55000
AVP1 transgenic
Cotton
Non-transgic cotton
No
. o
f P
hy
la
unclassified_Bacteria
Verrucomicrobia
TM7
Proteobacteria
Planctomycetes
Nitrospira
Gemmatimonadetes
Firmicutes
Euryarchaeota
Elusimicrobia
Deinococcus-Thermus
Crenarchaeota
Chloroflexi
Chlorobi
Chlamydiae
Bacteroidetes
Armatimonadetes
Actinobacteria
Acidobacteria
Proteobacteria
Crenarchaeota
Fermicutes
Unclassified Bacteria

3. Results
113
Table 3.37 Abundance of 16S rRNA sequences of different bacterial genera (Top
50 genera) retrieved from transgenic and non-transgenic cotton
No. Genera Sequences detected
in the rhizosphere
of transgenic cotton
Sequences detected in
the rhizosphere of non-
transgenic cotton
1 Bacillus 2221 3043
2 Steroidobacter 600 435
3 Cellvibrio 426 629
4 Lysobacter 410 95
5 Incertae sedis 388 442
6 Solirubrobacter 372 196
7 Niastella 363 264
8 Pseudoxanthomonas 360 224
9 Nitrospira 355 388
10 Microvirga 333 345
11 Gemmatimonas 302 233
12 Ohtaekwangia 298 186
13 Pseudomonas 291 439
14 Ensifer 254 185
Figure 3-46 16S rRNA sequences belonging to different bacterial classes reterieved
from the rhizosphere of AVP1 transgenic and non-transgenic cotton

3. Results
114
15 Paenibacillus 230 291
16 Arthrobacter 201 106
17 Devosia 184 95
18 Nitrosospira 180 204
19 Sphingomonas 180 188
20 Cystobacter 171 56
21 Tumebacillus 165 153
22 Dongia 157 125
23 Truepera 156 84
24 Acidobacteria 140 175
25 Streptomyces 139 46
26 Flavobacterium 124 56
27 Rubrobacter 115 58
28 Flavisolibacter 108 126
29 Nocardioides 104 22
30 Pontibacter 102 78
31 Chitinophaga 101 3
32 Amaricoccus 100 41
33 Marmoricola 97 50
34 Acidovorax 93 11
35 Skermanella 90 94
36 Rhizobium 80 132
37 Syntrophobacter 79 108
38 Altererythrobacter 78 49
39 Bryobacter 76 91
40 Arenimonas 72 226
41 Legionella 72 49
42 Rubellimicrobium 71 63
43 Phenylobacterium 66 57
44 Aquicella 64 39
45 Mesorhizobium 55 45
46 Hydrogenophaga 54 83
47 Ammoniphilus 53 112
48 Vasilyevaea 53 21
49 Armatimonadetes 49 58
50 Serpens 48 75

3. Results
115
Figure 3-47 Bacterial genera detected in the rhizosphere of AVP1 transgenic and
non-transgenic cotton
Table 3.38 Bacterial genera detected only in the rhizosphere of transgenic cotton

3. Results
116
Table 3.39: Sequences of important PGPR genera detect in the rhizosphere of
AVP1 transgenic and non-transgenic cotton
Genera Transgenic Cotton Non-Transgenic Cotton
Bacillus 2221 3043
Pseudoxanthomonas 360 224
Pseudomonas 291 439
Paenibacillus 230 291
Arthrobacter 201 106
Flavobacterium 124 56
Rhizobium 80 132
Mesorhizobium 55 45
Bradyrhizobium 48 48
Brevundimonas 40 16
Microbacterium 22 9
Azohydromonas 15 34
Azoarcus 5 4
Burkholderia 1 2
No. Bacterial genera Sequences detected in the
rhizosphere of AVP1
transgenic cotton
1 Pseudofulvimonas 31
2 Serratia 22
3 Azospirillum 14
4 Rhizobacter 13
5 Cellulosimicrobium 4
6 Croceicoccus 4
7 Gemmata 4
8 Microlunatus 4
9 Oxobacter 4
10 Armatimonadetes 3

3. Results
117
Figure 3-48 Abundance of important PGPR genera in the rhizosphere of AVP1
transgenic and non-transgenic cotton rhizosphere
2221
360
291230 201
8055 40
5
3043
224
439
291
106132
45 16 40
100
200
300
400
500
600
700
800
900
1000
Ge
ne
ra A
bu
nd
an
ce
Transgenic Cotton
Non-Transgenic Cotton

3. Results
118
3.12 Bacterial Diversity in Wheat Rhizosphere
From wheat rhizosphere (transgenic and non-transgenic) total 156282 sequences were
obtained by pyrosequencing analysis of 16S rRNA gene. Among these sequences
128006 sequences were related to bacteria, 7928 were Archeal sequences, and 21568
sequences belonged to unclassified bacteria. Among these sequences, on an average
40977±6138 sequences were detected in the rhizosphere of AVP1 transgenic wheat and
about 37163±4656 sequences from non-transgenic wheat rhizosphere. All the 18
bacterial phyla detected on the basis of 16S rRNA gene sequencing were represented
in the rhizosphere of transgenic and non-transgenic wheat. In the rhizosphere of AVP1
transgenic wheat Proteobacteria were most abundant (14327±3653 sequences),
followed by Crenarchaeota (9938±2921) and Firmicutes (6957±1724) sequences.
Sequences of Proteobacteria (13180±2784), Crenarchaeota (6580±1436) and
Firmicutes (8328±1257) were also abundant in the rhizosphere of non-transgenic wheat
as Crenarchaeota (Table 3.40).
3.12.1 Abundance of Bacterial Classes in Wheat Rhizosphere Soil
Sequences belonging to bacterial classes were found in wheat rhizosphere soil. Among
these sequences related to 44±2 bacterial classes were found in transgenic wheat
rhizosphere and sequences related to 42±0.7 bacterial classes were found in non-
transgenic wheat rhizosphere soil. Sequences belonging to majority of the classes
(94.84%) were common in both the soils. In AVP1 transgenic wheat, Thermoprotei
were most abundant (30%), followed by Alpha-proteobacteria (12%), Beta-
proteobacteria (7%), Gamma-proteobacteria (8%), Actinobacteria (5%), Bacilli (18%),
Delta-proteobacteria (10%), Sphingobacteria (5%), Acidobacteria GP3 (3%) and
Clostridia (2%). In non-transgenic wheat rhizosphere sequences of bacterial class
‘thermoprotei’ were also most abundant (21%), followed by Alpha-proteobacteria
(14%), Beta-proteobacteria (8%), Gamma-proteobacteria (8%), Actinobacteria (8%),
Bacilli (24%), Delta-proteobacteria (9%), Sphingobacteria (4%), Acidobacteria
GP3(3%) and Clostridia (1%) (Figure.3-50)
3.12.2 Abundance of Bacterial Genera in the Rhizosphere Wheat
In the present study 16S rRNA sequences related to 293±24 genera were found in the
rhizosphere of AVP1 transgenic wheat and in the rhizosphere of non-transgenic wheat
sequences related to 306±12 genera were detected. Among these 233±24 genera were

3. Results
119
common in the rhizosphere of both transgenic and non-transgenic wheat (Table 3.41).
Sequences of few genera were found only in the rhizosphere of AVP1 transgenic wheat
(Table 3.42). Bacterial genera that have been reported as PGPR were also detected in
the rhizosphere of transgenic and non-transgenic wheat (Table 3.43). Comparison of
the number of sequences retrieved from transgenic and non-transgenic plants belonging
to specific groups indicated quantitative differences but all major bacterial groups were
present in the rhizosphere of both type of plants.

3 Results
120
Table 3.40 Abundance of 16S rRNA sequences of different bacterial phyla in the
rhizosphere of AVP1 transgenic and non-transgenic wheat
No. Phylum Sequences retrieved
from AVP1 transgenic
Wheat rhizosphere
Sequences retrieved
from non-transgenic
wheat rhizosphere
1 Proteobacteria 14327±3653 13180±2784
2 Crenarchaeota 9938±2921 6580±1436
3 Firmicutes 6957±1724 8328±1257
4 Unclassified bacteria 6027±1203 5108±445
5 Bacteroidetes 1997±1005 1790±489
6 Acidobacteria 1820±989 1489±750
7 Actinobacteria 1640±945 2469±125
8 Euryarchaeota 904±435 198±57
9 Verrucomicrobia 569±253 504±165
10 Nitrospira 357±125 187±23
11 Chloroflexi 243±15 209±15
12 Chlamydiae 110±25 32±06
13 Gemmatimonadetes 96±17 116±25
14 Planctomycetes 64±05 55±04
15 Armatimonadetes 41±18 37±17
16 Incertaesedis 37±11 29±05
17 Deinococcus-Thermus 32±09 60±08
18 Chlorobi 27±07 38±14
DNA extracted from rhizosphere soil AVP1 transgenic and non-transgenic wheat was
used for pyrosequencing analysis of 16S rRNA gene. Given numbers are average of 2
replicates.
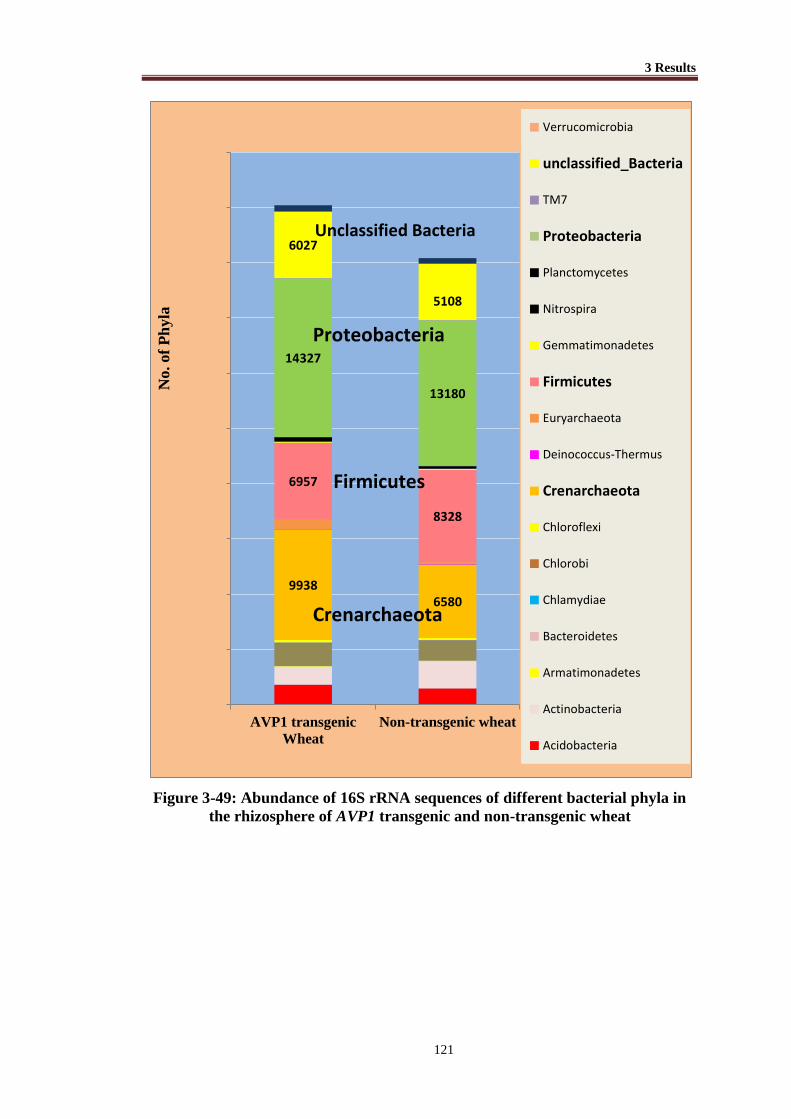
3 Results
121
Figure 3-49: Abundance of 16S rRNA sequences of different bacterial phyla in
the rhizosphere of AVP1 transgenic and non-transgenic wheat
99386580
6957
8328
14327
13180
6027
5108
0
5000
10000
15000
20000
25000
30000
35000
40000
45000
50000
AVP1 transgenic
Wheat
Non-transgenic wheat
No. of
Ph
yla
Verrucomicrobia
unclassified_Bacteria
TM7
Proteobacteria
Planctomycetes
Nitrospira
Gemmatimonadetes
Firmicutes
Euryarchaeota
Deinococcus-Thermus
Crenarchaeota
Chloroflexi
Chlorobi
Chlamydiae
Bacteroidetes
Armatimonadetes
Actinobacteria
Acidobacteria
7
00
Proteobacteria
Crenarchaeota
Firmicutes
Unclassified Bacteria

3 Results
122
Figure 3-50 Abundance of 16S rRNA sequences belonging to different bacterial
classes dominant in wheat rhizosphere of AVP1 transgenic and non-transgenic
wheat
Table 3.41 Abundance of 16S rRNA sequences belonging to different bacterial
genera (Top 50 genera) retrieved from the rhizosphere of AVP1 transgenic and
non-transgenic wheat
No. Genera Sequences detected
in the rhizosphere
of transgenic wheat
Sequences detected in
the rhizosphere of
non-transgenic wheat
1 Bacillus 3207 4738
2 Steroidobacter 794 761
3 Paenibacillus 490 418
4 Methanosarcina 458 40
5 Azotobacter 417 370
6 Nitrospira 344 181
7 Ohtaekwangia 338 295
8 Bryobacter 322 267
9 Sphingomonas 282 141
10 Flavisolibacter 281 262
11 Skermanella 235 318
12 Dongia 207 126
13 Microvirga 180 212
14 Niastella 176 226
15 Methanobacterium 174 44
16 Clostridium sensu stricto 153 85
17 Mesorhizobium 133 132
18 Armatimonadetes 114 45
19 Arthrobacter 113 93
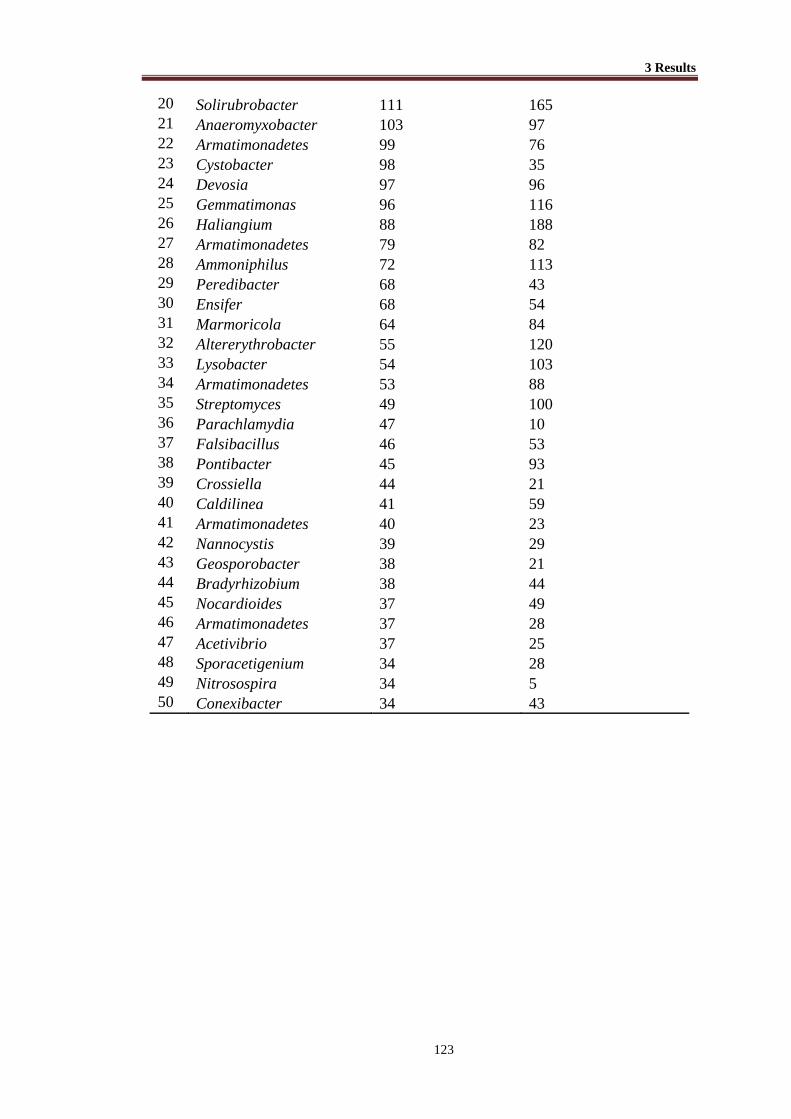
3 Results
123
20 Solirubrobacter 111 165
21 Anaeromyxobacter 103 97
22 Armatimonadetes 99 76
23 Cystobacter 98 35
24 Devosia 97 96
25 Gemmatimonas 96 116
26 Haliangium 88 188
27 Armatimonadetes 79 82
28 Ammoniphilus 72 113
29 Peredibacter 68 43
30 Ensifer 68 54
31 Marmoricola 64 84
32 Altererythrobacter 55 120
33 Lysobacter 54 103
34 Armatimonadetes 53 88
35 Streptomyces 49 100
36 Parachlamydia 47 10
37 Falsibacillus 46 53
38 Pontibacter 45 93
39 Crossiella 44 21
40 Caldilinea 41 59
41 Armatimonadetes 40 23
42 Nannocystis 39 29
43 Geosporobacter 38 21
44 Bradyrhizobium 38 44
45 Nocardioides 37 49
46 Armatimonadetes 37 28
47 Acetivibrio 37 25
48 Sporacetigenium 34 28
49 Nitrosospira 34 5
50 Conexibacter 34 43

3 Results
124
Figure 3-51 Bacterial genera detected only in the rhizosphere of AVP1 transgenic
and non-transgenic wheat

3 Results
125
Table 3.42 Bacterial genera detected only in the rhizosphere of transgenic wheat
Genera
16S rRNA sequences belonging to
transgenic wheat
1 Nocardioides 37
2 Cellulosilyticum 6
3 Acrocarpospora 4
4 Chondromyces 4
5 Rickettsia 4
6 Coxiella 3
7 Desertibacter 3
8 Methano- brevibacter 3
9 Aeribacillus 2
10 Amnibacterium 2
Table 3.43 Sequences of important PGPR genera detect in the rhizosphere of
AVP1 transgenic and non-transgenic wheat
Genera Transgenic Cotton Non-Transgenic Cotton
Arthrobacter 113 93
Aspromonas 1 1
Azoarcus 16 30
Azospirillum 14 21
Azotobacter 417 370
Azotobacter 417 370
Bacillus 3207 4738
Bradyrhizobium 38 44
Brevibacillus 18 7
Brevibacillus 18 7
Mesorhizobium 133 132
Paenibacillus 490 418
Paenibacillus 490 418
Pseudomonas 12 14
Rhizobium 16 37
*genera that have been reported as PGPR of different crops

3 Results
126
Figure 3-52 Abundance and comparison of PGPR detected in AVP1 transgenic
and non-transgenic rhizosphere of wheat.
4
338
3
207
490
417
133 113
38 18 16 16 14 12
418370
13293
447
30 37 21 14
0
100
200
300
400
500
600
700
800
900
1000P
GP
R A
bu
nd
ance
PGPR of Wheat Rhizosphere
AVP1-transgenic Wheat
Non-Transgenic Wheat

4. Discussion
For the development of sustainable agricultural systems, genetic engineering offers
genetic modifications of crop plants to incorporate useful traits. There is an increase in
the adoptability and cultivation of genetically modified crops [220]. However the use
of genetically modified crops requires well defined risk assessment prior to adoption in
any agricultural system [221]. These risks include the impact of genetically modified
plants on soil-associated microbial communities as plants are known to have a profound
effect on the abundance, diversity and activity of soil microorganisms living in close
proximity with their roots in a soil zone called the rhizosphere [222].
Present study was conducted to evaluate the diversity of bacteria in the
rhizosphere of AVP1 transgenic cotton and AVP1 transgenic wheat as well as non-
transgenic plants of both the crops. Studies conducted include determination of
bacterial diversity by isolation of bacteria from rhizosphere of transgenic and non-
transgenic plants, impact of PGPR strains on growth of cotton and wheat plants,
estimation of bacterial population in the rhizosphere of transgenic and non-transgenic
plants (cotton & wheat) at different plant growth stages on bacterial growth medium as
well as by real time PCR. Furthermore diversity of diazotrophic bacteria was
investigated by PCR amplification, cloning and sequencing of nifH gene using soil
DNA. Finally bacterial diversity was determined by pyrosequencing analysis of 16S
rRNA gene from soil DNA.
Bacteria were isolated from AVP1 transgenic cotton and AVP1 transgenic wheat
along with their non-transgenic control plants. On different growth media total 26
isolates were obtained which included 12 isolates from cotton and 14 isolates from
wheat rhizosphere. These isolates were identified on the bases of 16S rRNA gene
comparison with closely related sequences reported in the databank (NCBI). Bacterial
strains belonging to genera Arthrobacter (strain Bα), Azospirillum (strain BM31),
Bacillus (strains A5, CC and DC), Brevibacillus (strain TN4-3NF) and Pseudomonas
(strain D4) were isolated from AVP1 transgenic cotton rhizosphere and Agrobacterium
(strain NTW1), Bacillus (strains NTC-7, NTC-4 and AC) and Rhizobium (strain NTC-

4. Discussion
128
2NF) from non-transgenic cotton. Bacterial strains belonging to genera Bacillus were
most common in both transgenic and non-transgenic cotton rhizosphere. The bacterial
isolates from AVP1 transgenic wheat rhizosphere belonged to genera Achromobacter
(strains A6 and AZ), Alcaligenese (strain WC), Bacillus (strain WP2), Brevibacterium
(strain NFM-2) and Pseudomonas (strain WT2). Bacterial strains belonging to genera
Arthrobacter (strain NTC-11), Bacillus (strain WP2), Advenella (strain NTC-1NF) and
Pseudomonas (strain WP3) were isolated only from non-transgenic wheat. Bacterial
strains belonging to genera Achromobacter, Bacillus and Pseudomonas were common
in both AVP1 transgenic and non-transgenic wheat rhizosphere.
Bacterial strains isolated and identified in the present study have been found in
soil, mostly as residents of plant roots. The Achromobacter strains
(family Alcaligenaceae) have been isolated from wheat and maize [223, 224].
Colonization of Achromobacter species in sunflower rhizosphere under drought
conditions conferred tolerance to water stress [225]. Agrobacterium has been found as
soil bacterium [175] and in association with the roots of cotton and many other crop
plants [226]. Isolation and identification of different bacterial strains belonging to genus
Arthrobacter has been reported from the rhizosphere of various crops [67].
Arthrobacter species are known to have a considerable ability to survive during severe
drought conditions. A stress tolerant phosphate solubilizer Arthrobacter strain has been
isolated from tomato rhizosphere [227, 228]. From cotton rhizosphere of the Indian soil,
the strains belonging to genera Azospirillum, Arthrobacter and Bacillus have been
isolated [229]. Isolation of Azospirillum strains has often been reported from cereal
crops but isolations from cotton have also been made [230]. Isolation of Azospirillum
strains from the rhizosphere of wheat, rice and sugarcane has been reported previously
from the same soil as used in the present study [25, 223, 231]. Bacterial strains related
to genera Bacillus colonize the cotton root zone and affect the plant growth by adopting
different mechanisms [109, 232]. The present study indicated that the strains of Bacillus
are colonize in the rhizosphere of AVP1 transgenic and non-transgenic plants of cotton
and wheat more frequently as compared to the other rhizospheric bacteria. Occurrence
of Bacillus in the rhizosphere of cotton has been reported in Pakistani soil. Several
studies proved the presence and abundance of Bacillus genera in the rhizosphere of
various crops [233, 234]. Studies on bacterial diversity of transgenic tobacco showed
that Bacillus were commonly present in transgenic and non-transgenic rhizosphere

4. Discussion
129
[235]. Brevibacillus has been established as a new genus by reclassification of the
Bacillus brevis group of species in the genus Bacillus [236]. Brevibacillus strains
present in soil are known to play role in bioremediation and biocontrol in cotton
rhizosphere [237]. Presence of bacterial strains belonging to genus Paenibacillus has
been shown in the rhizosphere of cotton [238, 239]. High efficiency in host root
colonization of plant rhizosphere by Pseudomonads have been reported resulting in
improved crop yield [240, 241]. Advenella have been isolated from tea and rice
rhizosphere as a phytase producing bacteria for their plant growth promoting activities
[242]. Rhizobia are known generally to invade the root systems of legumes to form
nodules but as free-living bacteria are also an integral part of rhizosphere biota
exhibiting successful rhizosphere colonization [243, 244].
Production of IAA by the bacterial isolates purified from cotton and wheat was
investigated in pure culture. It has been found that not only plants but bacteria and fungi
are also able to synthesize IAA [245]. IAA synthesized by bacteria play a major role in
root growth, cell elongation, tissue differentiation, plant growth promotion and
responses to light and gravity [246]. In the present study, IAA production was detected
in the pure culture of Achromobacter strain A6, Agrobacterium strain NTW1,
Arthrobacter NTC-11, Azospirillum strains BM31, Bacillus strain A5 and NTC-7,
Pseudomonas strain D4 and WT2. Achromobacter strains isolated from different crops
like wheat, brassica, maize, and canola have been shown to produce IAA [247-249].
Agrobacterium can induce root formation in some plants due to production of natural
plant growth promoting substances (IAA) [250]. Arthrobacter strains have been
reported for IAA production under stress conditions [251]. Many strains of
Azospirillum produce IAA in considerable amounts that favors the plant growth [252].
Studies on Pseudomonas sp. have suggested that these bacteria are able to synthesize
IAA [253]. The treatment of seeds or cuttings with Agrobacterium, Pseudomonas and
many other bacterial genera induced root formation of some plants because of natural
auxin production by bacteria [254]. Moreover, IAA can also be a signaling molecule in
bacteria and therefore can have a direct effect on bacterial physiology [255]. It has been
estimated that more than 80% of bacteria isolated from the rhizosphere can produce
plant growth regulator IAA [75, 243].
Phosphate solubilization by bacteria is an important mechanism utilized for
plant growth promotion [256]. In the present study, phosphate solubilization activity of

4. Discussion
130
bacterial isolates was studied in Pikovskaya medium supplemented with tri-calcium
rock phosphate as sole source of phosphorus. Among the tested strains, efficient
phosphorus solubilization was shown by Achromobacter strains A6, Arthrobacter
strains Bα and NTC-11, Azospirillum strain BM31, Bacillus strain WP8, and
Pseudomonas strains WP3 and D4. Phosphate solubilization activity has been reported
in several genera of PGPR including Arthrobacter, Achromobacter, Bacillus, and
Pseudomonas [22, 223, 257]. Agrobacterium and some other genera isolated from the
temperate countries have the ability to solubilize phosphorous [258]. Phosphate
solubilization ability of different strains of Arthrobacter has been reported from
subtropical soils [75]. Arthrobacter strains as stress tolerant phosphate solubilizing
rhizobacteria have been isolated from tomato rhizosphere soil [259]. Pseudomonas
strains enhanced productivity in rice crop by phosphate solubilization [260, 261].
Rhizospheric bacteria secrete organic acids like acetic acid, citric acid and
gluconic acid to lower the pH of medium that affects the phosphorous solubilization
ability of bacteria [262, 263]. Similar studies showed the involvement of organic acids
produced by different PGPR in mobilization of P in the rhizosphere [32, 264]. In the
present study Achromobacter strain A6, Agrobacterium strain NTW1, Azospirillum
strain BM31, Bacillus strains (AC, CC, and DC), Brevibacillus strain TN4-3NF,
Paenibacillus NTC-7 and Rhizobium strain NTC-2NF showed production of organic
acids in pure culture. Among the detected acids, acetic acid was detected relatively in
higher amounts as compared to the other acids. Production of acetic acid by different
PGPR strains from the same soil has already been reported [257]. Maximum acetic acid
production was shown by Pseudomonas strain WP3, followed by Bacillus strain WP8,
Arthrobacter strain Bα and Azospirillum strain BM31. Similar studies indicated that
Arthrobacter, Azospirillum and Bacillus produce organic acids in pure culture for
phosphate solubilization [75, 265].
Use of PGPR with the aim of improving nutrient availability and plant growth
promotion has been studied during past couple of decades [266, 267]. In the present
study 8 bacterial isolates showing high IAA and phosphorus solubilization ability were
used as single-strain inoculants for AVP1 transgenic and non-transgenic cotton. For
quick screening in short-term experiments (40 days duration), bacterial isolates were
tested as inoculants for plants in sterilized sand and the tested strains included
Agrobacterium strain NTW1, Arthrobacter strain Bα, Azospirillum strain BM31,

4. Discussion
131
Bacillus strain NTC-4, Bacillus strain A5, Brevibacillus strain TN4-3NF, Paenibacillus
strain TNC-7 and Pseudomonas strain D4.The inoculated transgenic and non-
transgenic plants showed significant improvement of most of the growth parameters
recorded in the study. Maximum increase in root length (24%) and root dry weight
(22.1%) of transgenic plants was shown by inoculation of Arthrobacter strain Bα.
Maximum increase in root length (24%) of non-transgenic plants was recorded on
inoculation of Azospirillum strain BM31. No significant effect was observed on the
shoot dry weight and root dry weight of transgenic and non-transgenic plants inoculated
with Agrobacterium strain NTW1 and Paenibacillus strain TNC-7. Therefore,
Arthrobacter strain Bα, Azospirillum strain BM31, Bacillus strain NTC-4,
Brevibacillus strain TN4-3NF, and Pseudomonas strain D4, were selected for further
inoculation of cotton plants (transgenic and non-transgenic) in non-sterilized soil.
These selected strains were used as inoculum in pot experiment during 2011
and experiment was repeated again during 2012. During 2011, maximum increase in
yield of transgenic cotton plant was recorded on inoculation with Brevibacillus strain
TN4-3NF (11.2%) and Pseudomonas strain D4 (9.6%) over non-inoculated control.
Non-transgenic plants did not show any significant effect on yield except Pseudomonas
strain D4 and Arthrobacter strain Bα which showed 22 % and 23% increase,
respectively in yield of non-transgenic plants. During 2012, inoculation of transgenic
plants with Arthrobacter strain Bα and Pseudomonas strain D4 resulted in increased
cumulative root length (7.4%, 7.9%), shoot dry weight (8.9%, 11.5%), root dry weight
(21.6%, 21.8%), and yield (24.58%, 23%) of transgenic cotton plants over control.
Inoculation of non-transgenic plants with Brevibacillus strain TN4-3NF and
Pseudomonas strain D4 resulted in the improvement of all growth parameters studied.
Arthrobacter strain Bα, Azospirillum strain BM31, Bacillus strain NTC-4 Brevibacillus
strain TN4-3NF and Pseudomonas strain D4 were efficient in growth promotion of
transgenic plants as well as non-transgenic plants and therefore, qualified as effective
PGPR strains of cotton. Bacterial strains belonging to these genera have been reported
to be the integral parts of rhizosphere biota exhibiting successful rhizosphere
colonization and beneficial effect on growth of crop plants [81, 83, 268, 269].
Arthrobacter are extremely common in soils and often constitute more than one-half of
the total bacterial population [270]. It has been reported that co-inoculation of plants
with Arthrobacter sp. Bacillus and Pseudomonas could alleviate the adverse effects of

4. Discussion
132
soil salinity [271]. The Azospirillum genus is able to colonize more than a hundred plant
species and known to significantly improve plant growth, development, and
productivity under agronomic conditions [272]. Increase in the growth of root hairs and
number of lateral roots of cotton has been observed in greenhouse conditions [273].
Azospirillum inoculation increased plant height, dry weight and nitrogen content by
increasing N uptake in cotton [274]. It has been reported that in cotton root association,
Azospirillum is capable of producing antibacterial and antifungal compounds,
siderophores and growth regulating substances [275]. Species of Bacillus are common
inhabitants among the resident microflora of various species of plants including cotton,
where they play an important role in plant protection and growth promotion [276, 277].
In another study cotton and some other crops raised with inoculation of commercial
product of Bacillus were effective against soil borne pathogens [278]. Growth
promotion of transgenic (Bt) cotton due to inoculation with Bacillus and Arthrobacter
strains has been reported recently [279]. Members of the genus Brevibacillus have
received considerable attention as potential inoculants due to their ability to survive
under stressful conditions by endospore formation [279, 280]. Brevibacillus spp.
isolated from sugarcane rhizosphere have shown plant growth-promoting potential in
in vitro experiments [281] and also on germination of eggplant and pepper grown under
organic amendments and in greenhouse conditions [282]. Pseudomonas spp. are
ubiquitous bacteria in agricultural soils and have many traits that make them well suited
as PGPR. Specific strains of the Pseudomonas group used as seed inoculants on crop
plants (potato, radish, sunflower and sugar beet) caused statistically significant yield
improvement in field tests [283]. It has been reported that inoculation of cotton plants
with Pseudomonas spp. resulted in improved growth parameters as has been observed
in the present study [284].
For wheat inoculation studies bacterial strains were tested as single-strain
inoculants in sterilized sand for quick screening in short-term experiments (40 days
duration) Achromobacter strain A6, Arthrobacter strain NTC-11, Azospirillum strain
BM30, Bacillus strain WP2, Bacillus strain WP8, Brevibacterium strain NFM-2,
Microbacterium strain WN1, and Pseudomonas strain WP3 were used as single strain
inoculants for transgenic and non-transgenic wheat plants. Among the tested strains,
five strains i.e Achromobacter strain A6, Arthrobacter strain NTC-11, Azospirillum
strain BM30, Microbacterium strain WN1 and Pseudomonas strain WP3 showed an

4. Discussion
133
increase in all the growth parameters of transgenic plants. Maximum increase in
cumulative root length (22.1%), shoot dry weight (10.5%) and root dry weight (23 %)
was recorded in the plants inoculated with Pseudomonas strain WP3 over non-
inoculated control plants. Among the bacterial inocula tested for non-transgenic wheat
plants, no significant improvement of cumulative root length was recorded. Maximum
increase in shoot dry weight was recorded over control plants inoculated with
Achromobacter strain A6 Arthrobacter strain NTC-11 and Pseudomonas strain WP3.
Efficient bacterial strains (Achromobacter strain A6, Arthrobacter strain NTC-11,
Microbacterium strain WNI, Azospirillum strain BM30 and Pseudomonas strain WP3)
were studied in two experiments conducted during 2010-11 and 2011-12 in micro-plots
under natural conditions. Data analysis of both experiments showed a significant effect
of inoculated strains on both transgenic and non-transgenic plants as compared to non-
inoculated control plants. However, Arthrobacter strain NTC-11, Pseudomonas strain
WP3 and Azospirillum strain BM30 were more efficient strains compared with
Achromobacter strain A6, and Microbacterium strain WNI. During 2010-11,
inoculation of Arthrobacter strain NTC-11 showed maximum increase in the straw dry
weight (8.1 %) and maximum increase in grain weight (8.2 %) was noted in the plants
inoculated with Pseudomonas strain WP3. All inoculated strains showed significant
improvement of straw weight and grain weight of wheat grown in micro-plots. In the
experiments Achromobacter strain A6, Arthrobacter strain NTC-11, Azospirillum
strain BM30 and Pseudomonas strain WP3 were identified as efficient PGPR strains
for wheat. The bacterial inoculants tested in this study have frequently been used to
promote growth of wheat and other crop plants [109, 285]. Arthrobacter spp.,
Achromobacter spp., and Bacillus spp., were previously isolated from rhizosphere of
wheat and reported as dinitrogen fixers [286]. Isolation and characterization of
Achromobacter strains from wheat and maize has been reported [223, 248]. Inoculation
of Achromobacter sp. has been reported to confer tolerance to salt and water stresses to
tomato, sunflower and pepper [60, 287, 288]. The inoculation of maize seeds with the
strains of Achromobacter and Arthrobacter caused a significant increase in the shoot
and root dry matter of different cultivars [289]. Co-inoculation of wheat with
Arthrobacter and Bacillus strains could alleviate the adverse effects of soil salinity on
wheat growth [228]. Seed inoculations of wheat with Azospirillum and Bacillus
significantly increased the grain and straw yield [290]. Beneficial effects of
Azospirillum on wheat yield in field and greenhouse conditions have been reported [79,

4. Discussion
134
257, 291]. In another study, Azospirillum strains obtained from wheat roots showed a
consistent positive effect on the yield of different wheat cultivars [292]. Recently
growth promotion of wheat by phosphate solubilizing Azospirillum has been reported
from Pakistan [257]. Microbacterium has been reported to occur as endophyte of
different crop plants playing important ecological roles [293, 294]. Pseudomonas,
Bacillus and Microbacterium strains increased root growth of wheat when applied as
inoculum [295]. There are many reports on the occurrence of Pseudomonas spp. in the
rhizosphere of wheat [123, 296-298].
The use of genetically modified or transgenic plants has a great potential for
sustainable agriculture. However an increase in the adoptability and cultivation of
genetically modified crops demands well-defined risk assessment [160]. These risk
assessment studies are essentially required because some transgenic plants (insect
resistant) are known to change rhizosphere environment through root exudates, which
consequently affects the growth of microorganisms in the rhizosphere [299]. In the
present study bacterial populations in the rhizosphere of AVP1 transgenic and non-
transgenic cotton and wheat were compared at different growth stages. Bacterial
population (cfu/g soil) were estimated on nutrient agar medium at three crop
developmental stages i.e 30, 60, and 90 DAS of cotton and wheat. In the present study
no significant difference was observed in bacterial population in the rhizosphere of
transgenic and non-transgenic rhizosphere soils at all three growth stages. Similar to
these results, no difference was observed in growth of bacterial communities between
the rhizosphere of transgenic potato and non-transgenic potato varieties [117]. Also, no
effect of transgenic maize has been observed on diversity of culturable bacterial
communities in rhizospheric soil samples [115]. Studies on culturable bacterial
population showed that B. thuringiensis toxins present in transgenic corn (Zea mays)
rhizosphere did not change culturable microbial communities [300]. The diversity of
rhizosphere bacteria of transgenic, herbicide-resistant corn was not different from that
of the corresponding non-transgenic variety [301].
In the present study bacterial population varied significantly with the crop
development stage. In cotton rhizosphere maximum bacterial populations i.e 5.63 (log
cfu/g soil) and 5.58 (log cfu/g soil) were recorded at 90 DAS among transgenic and
non-transgenic plants, respectively. In the rhizosphere of wheat, maximum bacterial
populations i.e 6.42 (log cfu/g soil) and 6.40 (log cfu/g soil) were recorded at 60 DAS

4. Discussion
135
in transgenic and non-transgenic plants, respectively and relatively low cfu/g soil were
detected at 30 DAS and 90 DAS. Population of diazotrophic bacteria in the rhizosphere
of transgenic and non-transgenic plants of cotton and wheat was observed in semi-solid
nitrogen-free malate (NFM) medium. Data showed that there was no statistically
significant difference among diazotrophic population (MPN) in the rhizosphere of
transgenic and non-transgenic plants of cotton and wheat at different stages except in
wheat rhizosphere at 90 DAS which was relatively higher compared with other growth
stages. This difference in bacterial population may be the effect of qualitative and
quantitative difference of substrates released by plant roots by exudation at different
developmental stages [302].
In the present study, real time PCR was used to quantify microbial
populations in the rhizosphere of cotton and wheat. Abundance of 16S rRNA and nifH
genes in rhizosphere was determined by real time PCR from the soil samples collected
from AVP1 transgenic and non-transgenic plants at different plant developmental stages
(35 DAS and 90 DAS). In cotton rhizospheric soil samples, more copy number of 16S
rRNA gene were recovered in AVP1 transgenic cotton (log 6.39 copies/g of soil), at 90
DAS and in non-transgenic cotton log 6.36 copies/g of soil of 16S rRNA gene at 90
DAS than at 35 DAS. These results showed that bacterial population was low at early
stage of cotton (35 DAS) and it increased with the crop development and was highest
at 90 DAS. Abundance of nifH gene was determined to estimate population of
diazotrophic bacteria in the rhizosphere. In the rhizosphere of transgenic and non-
transgenic cotton log 5.66 and log 5.85 nifH copies/g of soil were detected, respectively.
At 90 DAS, nifH sequences detected in the rhizosphere of transgenic and non-
transgenic cotton plants were log 5.41 and log 5.39 nifH copies/g of soil, respectively.
In the rhizosphere of transgenic and non-transgenic wheat relatively higher copy
number of 16S rRNA and nifH genes were detected at 35 DAS compared with 90 DAS.
Rhizosphere microbial communities are known to be affected by several factors
including plant growth stages [303]. Previous studies indicated that the rhizosphere
population decreases as a plant matures [304], where as other study showed that
microbial diversity increases with plant age [302]. In our study generally similar or very
close copy number of 16S rRNA and nifH gene in transgenic and non-transgenic plants
showed that the shift in bacterial population is an independent function of plant without
any influence of foreign gene (AVP1) incorporated into transgenic plants. A significant

4. Discussion
136
but not persistent change in the abundance of bacterial and archaeal communities
determined by real time PCR has been observed [305]. It has been reported that that the
changes in the microbial community structure associated with genetically modified
plants were temporary and did not persist into the next season [122]. Another study
showed that the abundance and diversity of nitrogen-fixing bacteria tended to increase
with duration of organic management but the highest number of nifH gene copies was
observed in the rhizosphere and bulk soil of 5 years organic management [306].
Organic acids have been hypothesized to perform many functions in soil
including root nutrient acquisition, mineral weathering, microbial chemotaxis and
metal detoxification. Organic acids as root exudates are released into the rhizosphere
as a result of rhizodeposition and play a major role in the maintenance of root-soil and
root-microbe contacts [79, 307]. Information on the content and composition of organic
acids and sugars in exudates of various crops is limited and has a very fragmented
character. A study on tomato plant exudates showed that citric acid was the major
organic acid in root exudates. Malic acid and succinic acid were also detected however,
their levels were dependent on the plant age. The percentage of malic acid decreased
between the seedling and root stage, whereas the opposite occurred with succinic acid,
which became a major organic acid between growth stages of tomato plant. In another
study cucumber and sweet pepper rhizosphere citric acid, malic acid and succinic acid
were present in substantial amounts, although the levels and timing was different among
various crops [121].
In the present study organic acids (acetic acid, citric acid, malic acid, lactic acid,
gluconic acid and succinic acid) produced as root exudates in soil solution collected
from rhizosphere of transgenic and non-transgenic plants of cotton and wheat were
assayed. To detect a particular acid from concentrated soil solution standards of six
acids were used (production of these six acids were also assayed from pure culture of
different bacterial strains). However, in soil solution only acetic acid, citric acid, malic
acid and lactic acids were detected in comparable amounts. Gluconic acid and succinic
acid were also detected in soil solution but there concentration were quite low (0.0002-
0.0004 μg/mL). Among the organic acids detected in soil solution, acetic acid was
detected in relatively higher amounts, followed by oxalic acid and citric acid.
Production of different organic acids (Acetic acid, Oxalic Acid, Citric acid, Malic acid)
were analyzed statistically that showed significant difference of organic acids

4. Discussion
137
production in soil solution of AVP1 transgenic and non-transgenic plants (cotton and
wheat). Production of Acetic acid, Oxalic Acid and Citric acid was significant among
AVP1 transgenic cotton plants plans as compared to non-transgenic plants. However
production of Malic acid was non-significant among transgenic and non-transgenic
cotton plants. AVP1 transgenic wheat plants showed significant production of Acetic
acid, Oxalic Acid and Malic acid as compared to non-transgenic plants. Citric acid
production was non-significant among transgenic and non-transgenic wheat plants. In
the present study, maximum amount of acetic acid (8.89 µg/mL) was detected in the
soil solution of transgenic cotton plants. Oxalic acid was the second dominant acid
detected in soil solution and relatively higher amounts were detected in the soil solution
of transgenic cotton and wheat plants as compared to non-transgenic plants soil
solutions. The presence of organic acids in the rhizosphere of different crops has been
reported previously [307, 308] and it has been found that rate of root exudates released
by the plant roots influence microbial biota and activity in rhizosphere [309].
Culture-independent molecular methods are being applied for the assessment of
diazotrophic diversity by amplifying, cloning and sequencing of the nifH gene from
environmental DNA samples [310, 311]. Diazotrophic diversity on the basis of nifH
gene amplification and sequencing of soil DNA from rhizosphere of mangrove forest
of China [312], oak-hornbeam forest (Chorbush Forest) Germany [169], soil from
France and Senegal [170], Douglas fir forest in USA [313] and rice rhizosphere at
Kyushu University Farm, Japan [172] has been reported. In the present study diversity
of diazotrophic bacteria in rhizosphere of AVP1 transgenic cotton and wheat was
explored by sequence analysis of nifH gene from soil DNA. Data analysis showed that
sequences related to Anabaena, Azospirillum, Bradyrhizobium and Pseudomonas were
common in AVP1 transgenic and non-transgenic cotton whereas nifH sequences of
Zoogloea, Azohydromonas and Azospira were detected only in non-transgenic cotton
rhizosphere soil samples. Common genera in AVP1 transgenic and non-transgenic
wheat were Agrobacterium, Azospirillum, Bradyrhizobium and Pseudomonas, however
sequences related to Zoogloea were detected only in non-transgenic wheat rhizosphere.
These results indicated that diverse diazotrophs community exist in the rhizosphere of
transgenic and non-transgenic plants that can be isolated and used as inoculants to
enhance the crop productivity [314]. In the present study representatives of bacterial
genera Agrobacterium, Azospirillum, Pseudomonas and Rhizobium were present

4. Discussion
138
among isolates and no isolate representing Azoarcus, Bradyrhizobium and Zoogloea
was obtained. Strains of Azospirillum; Azotobacter, Bacillus, Pseudomonas, Rhizobium
and Zoogloea and some other genera have been reported as non-symbiotic diazotrophs
of crop farming systems in Pakistan and elsewhere [25, 49, 315, 316]. The genus
Azospira was described by Reinhold-Hurek & Hurek [159] to accommodate a lineage
of nitrogen-fixing bacteria phenotypically differentiated from other strains originally
assigned to the genus Azoarcus [317]. The genus Azospira includes a single validly
published species, Azospira oryzae, strains of which have been isolated from surface-
sterilized roots of Kallar grass collected from Pakistan as well as from resting stages
(sclerotia) of a basidiomycete [318].
Our results showed that in all four clone libraries constructed from transgenic
cotton (TC), non-transgenic cotton (NTC), transgenic wheat (TW), non-transgenic
wheat (NTW), sequences related to non-cultured diazotrophic bacteria were abundant
and constituted about 78% in TC, 74% in NTC, 78% in TW, and 79% in NTW of total
detected sequences. Sequences of uncultured diazotrophic bacteria detected in the
present study have already been reported from cotton rhizosphere, alkaline soils, and
mostly were from agricultural soils (unpublished results reported at NCBI data bank).
Many authors have already described sequences corresponding to diverse unidentified
diazotrophs [45, 170]. These non-cultivated diazotrophs may be the dominant nitrogen-
fixing organisms in soil systems [319]. In our study 14 uncultured diazotrophic
sequences showed homology with uncultured sequences from saline alkaline soils of
The Netherlands, 5 uncultured sequences showed homology with the sequences that
have been reported from cotton rhizosphere of India. Uncultured bacterial nifH
sequences showing homology with sequences reported from paddy soils of china,
agricultural soil Switzerland, root of sugarcane from Indian Punjab, leaf surface of
tropical plants and from soil of California USA were also detected. Some sequences of
uncultured diazotrophic bacteria from AVP1 transgenic and non-transgenic wheat
rhizosphere showed homology with diazotrophic communities of Dutch soil which
have been extensively studied for seasonal variation in the diversity and abundance of
diazotrophic communities [209]. Other uncultured sequences showed homology with
uncultured database (NCBI) sequences from sugarcane rhizosphere of China,
rhizosphere of cereal crops of Greece and Agricultural soil of Italy. In the present study
majority of the retrieved sequences of culturable as well as non-culturable bacteria were

4. Discussion
139
commonly present in the transgenic and non-transgenic plants pointing to no significant
effect of transgenic plants on diazotrophic communities. Persistence of nitrogen fixing
bacteria without any effect of transgenic has also been reported [320].
In the present study, pyrosequencing analysis of 16S rRNA gene directly
amplified from rhizosphere soil DNA of AVP1 transgenic cotton and wheat along with
non-transgenic plants were carried out to compare bacterial diversity. Sequences related
to 19 bacterial phyla were detected in AVP1 transgenic and non-transgenic cotton and
sequences related to 18 bacterial phyla were detected in AVP1 transgenic and non-
transgenic wheat. In cotton and wheat root system, Actinobacteria, Acidobacteria,
Bacteroidetes, Crenarchaeota, Proteobacteria and Firmicutes were most dominant
among the detected phyla. The detection of these phyla has been reported through
metagenomic studies in different soils [321, 322]. Data analysis showed that in cotton
rhizosphere, sequences related to Actinobacteria, Acidobacteria, Proteobacteria and
Firmicutes were more abundant in transgenic plants except Crenarchaeota which were
dominant among non-transgenic plants. However in wheat rhizosphere, sequences
related to Acidobacteria, Bacteroidetes, Crenarchaeota and Proteobacteria were
relatively more abundant in transgenic wheat rhizosphere and sequences related to
Actinobacteria and Firmicutes were dominant in the rhizosphere of non-transgenic
plants. Major phyla detected in present study have been reported in a variety of
environments. Proteobacteria phylum is metabolically versatile and genetically
diverse, comprising the largest fraction of the bacterial community in soil ecosystems
including the rhizosphere [323]. Dominant populations of Actinobacteria have been
reported on maize roots in a tropical soil [324]. It was also found that members of the
Bacteroidetes group constituted dominant populations in the rhizosphere of maize and
canola. Members of these bacterial groups are capable of degrading complex
macromolecules, thus contributing to the turnover of carbon, nitrogen and phosphorus
[325]. Further analysis of the sequences showed that a total number of 80±05 and 88±06
bacterial classes were found in cotton and wheat rhizosphere, respectively. In our study
bacterial classes alpha, beta and gamma Proteobacteria belonging to phylum
Proteobacteria were most abundant, followed by Thermoprotei and Bacilli. The
members of the classes of Proteobacteria have been well -known for their role in
agriculture, possessing several plant growth promoting mechanisms including
phytohormones, siderophore, phosphate solubilization and nitrogen fixation [326,

4. Discussion
140
327]. Dominant members of Proteobacteria classes in our study were in accordance
with other studies on bacterial diversity in maize rhizosphere [328]. Second abundant
bacterial class was Thermoprotei, belonging to phylum Crenarchaeota.
The Crenarchaeota have been classified as a phylum of the Archaea kingdom. Initially,
the Crenarchaeota were thought to be sulfur-dependent extremophiles but recent
studies have indicated that Crenarchaeota may be the most abundant archaea in the
marine environment. Until recently all cultured Crenarchaeota had been considered as
“thermophilic” or “hyper-thermophilic” organisms, some of which have the ability to
grow at up to 113 °C. These Archea have been detected in agricultural soil in various
studies [329, 330].
Data analysis at genus level showed that in transgenic cotton rhizosphere
bacterial sequences related to 344±5 genera were found and sequences related to
321±12 genera were detected in non-transgenic cotton rhizosphere soil. Among these,
sequences of 240 genera were commonly detected in transgenic and non-transgenic
rhizosphere. This indicated that majority of detected genera were common in both root
systems i.e transgenic and non-transgenic cotton rhizosphere. Data showed that 16S
rRNA sequences of Bacillus were the most abundant in transgenic (2221 sequences)
and non-transgenic cotton (3043 sequences), followed by Steroidobacter, Cellvibrio,
Lysobacter and Rubrobacter. Steroidobacter genus belonging to the order Rhizobiales
has been reported as an agar-degrading bacterium recently isolated from soil collected
from a vegetable cropping field [331]. Cellvibrio, a genus of Pseudomonadaceae, is a
nitrogen-fixing bacterium isolated from the rhizosphere soils treated with manure
[332]. The genus Lysobacter is grouped in the family Xanthomonadaceae, belonging
to the Gamma-Proteobacteria, found in soil and water habitats [333]. Several members
of Lysobacter are known as biological control agents against soil borne phyto-
pathogens such as Rhizoctonia solani, Thielaviopsis basicola [334]. Studies have
suggested that beneficial bacteria belonging to Lysobacter probably play an important
role for soil suppressiveness and plant growth [335, 336]. Our data showed that the
relative abundance of Lysobacter was is much higher in the rhizosphere of transgenic
cotton. 16S rRNA gene sequences related to Rubrobacter spp. have been recovered
from moderate, terrestrial environments [337, 338]. The type strain of Niastella has
been isolated from soil cultivated with Korean ginseng. Sequences related to bacterial
genera known for PGPR like Bradyrhizobium, Pseudomonas, Rhizobium,

4. Discussion
141
Streptomyces, Serratia, and Azospirillum were frequently detected in the present study
[252, 339].
In this study, 16S rRNA sequences related to 60 genera were detected only in
AVP1 transgenic cotton rhizosphere. Pseudofulvimonas, Serratia, Azospirillum, and
Rhizobacter were most abundant genera detected only in the rhizosphere of transgenic
cotton. 16S rRNA sequences related to 33 genera were found only in the rhizosphere
of non-transgenic cotton. Among them the most abundant were Brachybacterium,
Glycomyces, Actinomycetospora, Aeromicrobium, and Achromobacter.
Brachybacterium is able to degrade anthracene very efficiently and could serve as better
candidate for bioremediation [340]. It has also been isolated from oil contaminated soils
[341]. Glycomyces, Actinomycetospora and Aeromicrobium belonging to
actinomycetes, have been isolated from soil in different regions of the world [342].
Aeromicrobium strains have been isolated from an alkaline soil [343]. Actinobacteria
are ideal candidates for developing microbial inoculants for use in agriculture
production system as phosphate solubilizers [342]. Actinomycetes are more widespread
and important in soil [344].The genus Fulvimonas was described by Mergaert et al.
[345] with a single species Fulvimonas soli which has been isolated from soil [345].
Members of the Actinobacteria, Rubrobacter and xylanophilus (phylum
Actinobacteria) are widespread in soils throughout the world. Nitrosomonas are
ammonia-oxidizing bacteria belonging to the phylum Proteobacteria were detected and
have been reported for bioremidation. Microvirga, a root nodule symbiotic bacterium
was isolated from cowpea grown in semi-arid Brazil [346]. It has been isolated from
rice field soil in China [347]. Novel aerobic bacterium Gemmatimonas was isolated
from an anaerobic and aerobic sequential batch reactor for wastewater treatment [348].
Other genera detected in cotton rhizosphere include Pseudoxanthomonas, a bio-
surfactant producing bacterium with high emulsifying activity, Hydrogenophaga, is
being used in bioreduction of vanadium (V) in groundwater [349] and Skermanella
strains have been purified from a sand sample collected from the desert of Xinjiang,
China [350].
In the present study 16S rRNA sequences related to 293±24 genera were found
in the rhizosphere of AVP1 transgenic wheat and in the rhizosphere of non-transgenic
wheat sequences related to 306±12 genera were detected. Among these 233±24 genera
were common in the rhizosphere of both transgenic and non-transgenic wheat. Among

4. Discussion
142
the dominant bacterial genera detected maximum sequences of Bacillus were recorded.
Bacterial sequences related to 42 genera were found only in the rhizosphere of
transgenic wheat in which Cellulosilyticum and chondromyces were abundant. 16S
rRNA sequences related to 63 genera were found only in the rhizosphere of non-
transgenic wheat where Nocardioides and Nonomuraea genera were most abundant.
Cellulosilyticum has been detected from an anaerobic ethanol-producing cellulolytic
bacterial consortium from hot springs basin with agricultural residues and energy crops
[351]. The genuine habitat of the genus Chondromyces is soil, as long as the pH is
slightly acid to slightly alkaline but is frequently found on the dung of herbivorous
animals, on decaying plant material and on the bark of trees and occasionally they have
also been found on the surface of plant leaves [352]. The genus Nonomuraea is a rare
actinomycete taxon with a long taxonomic history. The genus is less known among the
rare actinomycete genera as its taxonomic position was revised several times. It can be
found in diverse ecological niches, while most of its member species were isolated from
soil samples [353]. Nocardioides sp. widely distributed in agricultural soils degrades a
range of herbicides [354]. A ‘strange’ genus detected from wheat rhizosphere was
Haliangium, belong to family Haliangiaceae and represents a unique myxobacterial
taxon occupying a novel and distinct phylogenetic cluster. To date, there is no valid
standing nomenclature to classify the monotypic genus Haliangium. So far, all
members of this taxon were isolated from marine environment and currently the only
known application is the production of novel biologically active compounds [355].
Falsibacillus has been isolated from coastal regions and from rhizosphere soil of a
medical plant [356]. Bryobacter aggregatus has been proposed to accommodate three
strains of slowly growing, chemo-organotrophic bacteria isolated from acidic
Sphagnum peat bogs [357]. Comparison of the number of sequences retrieved from
transgenic and non-transgenic plants belonging to specific groups indicated quantitative
differences but all major bacterial groups were present in the rhizosphere of both type
of plants. Pyrosequencing data generated in the present study showed that important
PGPR genera like Arthrobacter, Azopsirillum, Azoarcus, Bacillus, Paenibacillus,
Mesorhizobium, Bradyrhizobium, and Pseudomonas were detected in all soil samples.
However bacterial genera Azohydromonas, Brevundimonas, Burkholderia
Pseudoxanthomonas, Microbacterium and Rhizobium were present only with cotton
root system where as bacterial genera Azotobacter, Azospirillum, Brevibacillus and
Rhizobium were detected in wheat root system only. In the present study, bacterial

4. Discussion
143
sequences related to genera Azoarcus, Azohydromonas, Bradyrhizobium and
Mesorhizobium were frequently detected but representatives of these could not be
isolated from soil as pure culture. Therefore, potential of these genera for plant growth
promotion could not be explored. Similarly cloning and sequencing of nifH gene
amplified from soil showed the presence of well-known diazotrophic genera like
Azotobacter, Azoarcus, Bradyrhizobium and Zoogloea in the rhizosphere but could not
be isolated as pure culture. On the other hand, representative of the diazotrophic
bacterial genera Arthrobacter, Paenibacillus, Brevibacillus and Enterobacter were
could not detected by nifH gene sequencing, however pyrosequencing analysis of 16S
rRNA gene revealed the presence of these and many other important PGPR genera.
In the present study, bacterial sequences that showed homology with uncultured
bacteria were also detected in both nifH and 16S rRNA gene sequencing. Data showed
that nifH sequences of uncultured nitrogen-fixing bacteria retrieved by the culture-
independent approach (nifH gene cloning and sequencing) were more than > 70% of
the total detected sequences. Similarly pyrosequencing of 16S rRNA gene showed that
approximately 20% sequences were related to uncultured bacteria. Presence of
uncultured bacterial sequences have been reported from different environments
depending upon the technique used for detection [358, 359]. A fraction of uncultured
bacterial sequences from the rhizosphere of different potato cultivars has been reported
[360]. Presence of uncultured bacterial communities has also been shown in the
rhizosphere of healthy and diseased wheat plants [361]. Uncultured rhizobacterial
communities in glyphosate-tolerant cotton rhizosphere were variable among transgenic
and non-transgenic plants [362]. The uncultured bacteria detected in the present study
as well as other studies simply indicates that the laboratory culturing techniques
presently being used are unable to grow these bacteria due to the lack of critical
information about specific biology of micro-organisms. It is expected that many
‘uncultured’ bacteria will be isolated and used not only for plant growth promoting but
also for other useful purposes.
4.1 Conclusion and Future Perspectives
Based on estimation of bacterial populations and bacterial diversity assessed in the
present study, it was found that AVP1 gene did not adversely affect soil enzymatic
properties and soil micro-flora. The present study supports introduction of AVP1-

4. Discussion
144
transgenic wheat in agricultural systems in the country as no adverse effects were
observed on indigenous bacterial communities.
In the present study, bacterial sequences related to several well-known PGPR like
genera Azoarcus, Azotobacter, Acinetobacter and Enterobacter were detected in the
rhizosphere but the same could not be isolated from soil as pure culture. Future studies
may focus on isolation of these important genera using selective growth media and
optimum growth conditions and explored for their PGPR potential. Soil DNA-based
studies also indicated presence of bacterial like Steroidobacter, Streptomyces,
Micromonospora, Lysobacter, Rubrobacter, Pseudoxanthomonas Rhodococcus,
Fulvimonas and Nitrosomonas which have been reported for important biological
activities like bio-remediation of heavy metals, degradation of cholestrol and rubber,
production of pharmaceutical agents and antibiotic producing activity. Future studies
may be directed to isolate and utilize these bacteria for bio-remediation of contaminated
soils and for production of useful bio-molecules. Efficient PGPR identified in the
present study may be utilized as single-strain inocula or in different combinations in
extensive field trials before final recommendation for commercial biofertilizer
production for cotton and wheat.

5. References
[1] B. E. Tabashnik, Y. Carrière, T. J. Dennehy, S. Morin, M. S. Sisterson, R. T.
Roush, et al., "Insect resistance to transgenic Bt crops: lessons from the
laboratory and field," Journal of Economic Entomology, vol. 96, pp. 1031-1038.
[2] R. T. Fraley, S. G. Rogers, R. B. Horsch, P. R. Sanders, J. S. Flick, S. P. Adams,
et al., "Expression of bacterial genes in plant cells," Proceedings of the National
Academy of Sciences, vol. 80, pp. 4803-4807, 1983.
[3] C. James and A. F. Krattiger, "Global review of the field testing and
commercialization of transgenic plants: 1986 to 1995," ISAAA Briefs, 1996.
[4] M. Vaeck, A. Reynaerts, H. Höfte, S. Jansens, M. De Beuckeleer, C. Dean, et
al., "Transgenic plants protected from insect attack," Nature, vol. 328, pp. 33-
37, 1987.
[5] B. Martineau, "First fruit: The Creation of the Flvr Savr tomato and the birth
of genetically engineered food," ed: McGraw-Hill, New York, 2001.
[6] L. L. Wolfenbarger and P. R. Phifer, "The ecological risks and benefits of
genetically engineered plants," Science, vol. 290, pp. 2088-2093, 2000.
[7] J. Rhoades and J. Loveday, "Salinity in irrigated agriculture," Agronomy, pp.
1089-1142, 1990.
[8] W. Gruissem, "Genetically modified crops: the truth unveiled," Agriculture &
Food Security, vol. 4, p. 3, 2015.
[9] M. L. Dionisio-Sese and S. Tobita, "Antioxidant responses of rice seedlings to
salinity stress," Plant Science, vol. 135, pp. 1-9, 1998.
[10] F. J. Maathuis and A. Amtmann, "K+ nutrition and Na+ toxicity: the basis of
cellular K+/Na+ ratios," Annals of Botany, vol. 84, pp. 123-133, 1999.
[11] J.-K. Zhu, "Regulation of ion homeostasis under salt stress," Current opinion in
plant biology, vol. 6, pp. 441-445, 2003.
[12] L. Taiz, "The plant vacuole," Journal of Experimental Biology, vol. 172, pp.
113-122, 1992.

5. References
146
[13] T. Mimura, M. Kura-Hotta, T. Tsujimura, M. Ohnishi, M. Miura, Y. Okazaki,
et al., "Rapid increase of vacuolar volume in response to salt stress," Planta,
vol. 216, pp. 397-402, 2003.
[14] R. A. Gaxiola, J. Li, S. Undurraga, L. M. Dang, G. J. Allen, S. L. Alper, et al.,
"Drought-and salt-tolerant plants result from over-expression of the AVP1 H+-
pump," Proceedings of the National Academy of Sciences, vol. 98, pp. 11444-
11449, 2001.
[15] R. A. Gaxiola, G. R. Fink, and K. D. Hirschi, "Genetic manipulation of vacuolar
proton pumps and transporters," Plant Physiology, vol. 129, pp. 967-973, 2002.
[16] S. Park, J. Li, J. K. Pittman, G. A. Berkowitz, H. Yang, S. Undurraga, et al.,
"Up-regulation of a H+ pyrophosphatase (H+ PPase) as a strategy to engineer
drought-resistant crop plants," Proceedings of the National Academy of
Sciences of the United States of America, vol. 102, pp. 18830-18835, 2005.
[17] P. Bhattacharyya and D. Jha, "Plant growth-promoting rhizobacteria (PGPR):
Emergence in agriculture," World Journal of Microbiology and Biotechnology,
vol. 28, pp. 1327-1350, 2012.
[18] A. D. Kent and E. W. Triplett, "Microbial communities and their interactions in
soil and rhizosphere ecosystems," Annual Reviews in Microbiology, vol. 56, pp.
211-236, 2002.
[19] R. Porcel, A. Zamarreno, J. Garcia-Mina, and R. Aroca, "Involvement of plant
endogenous ABA in Bacillus megaterium PGPR activity in tomato plants,"
BMC Plant Biology, vol. 14, p. 36, 2014.
[20] N. Lane, "The unseen world: reflections on Leeuwenhoek (1677)‘Concerning
little animals’," Philosophical Transactions of the Royal Society of London B:
Biological Sciences, vol. 370, p. 20140344, 2015.
[21] J. Kloepper and M. Schroth, "Relationship of in vitro antibiosis of plant growth-
promoting rhizobacteria to plant growth and the displacement of root
microflora," Phytopathology, vol. 71, pp. 1020-1024, 1981.
[22] P. Bhattacharyya and D. Jha, "Plant growth-promoting rhizobacteria (PGPR):
emergence in agriculture," World Journal of Microbiology and Biotechnology,
vol. 28, pp. 1327 - 1350, 2012.
[23] G. A. Beattie, "Plant-associated bacteria: survey, molecular phylogeny,
genomics and recent advances," In Plant-Associated Bacteria, ed: Springer,
2006, pp. 1-56.

5. References
147
[24] Y. Bashan and L. E. de-Bashan, "How the plant growth-promoting bacterium
Azospirillum promote plant growth-A critical assessment.," Advances in
Agronomy, vol. 108, pp. 77-136, 2010.
[25] M. Samina, M. S. Mirza, H. Jacqueline, B. Rene, P. Normand, B. Asghari, et
al., "Isolation and 16S rRNA sequence analysis of the beneficial bacteria from
the rhizosphere of rice," Canadian Journal of Microbiology, vol. 47, 2001.
[26] V. Sgroy, F. Cassán, O. Masciarelli, M. F. Del Papa, A. Lagares, and V. Luna,
"Isolation and characterization of endophytic plant growth-promoting (PGPB)
or stress homeostasis-regulating (PSHB) bacteria associated to the halophyte
Prosopis strombulifera," Applied Microbiology and Biotechnology, vol. 85, pp.
371-381, 2009.
[27] E. Gray and D. Smith, "Intracellular and extracellular PGPR: Commonalities
and distinctions in the plant-bacterium signalling processes," Soil Biology and
Biochemistry, vol. 37, pp. 395 - 412, 2005.
[28] D. P. K. Agrawal and S. Agrawal, "Characterization of Bacillus sp. strains
isolated from rhizosphere of tomato plants (Lycopersicon esculentum) for their
use as potential plant growth promoting rhizobacteria," International Jurnal of
Current. Microbiogy and. Applied Scices, vol. 2, pp. 406-417, 2013.
[29] E. J. Rubio, M. S. Montecchia, M. Tosi, F. D. Cassán, A. Perticari, and O. S.
Correa, "Genotypic Characterization of Azotobacteria isolated from
Argentinean soils and plant-growth-promoting traits of selected strains with
prospects for biofertilizer production," The Scientific World Journal, vol. 2013,
2013.
[30] K. A. Turk, A. P. Rees, J. P. Zehr, N. Pereira, P. Swift, R. Shelley, et al.,
"Nitrogen fixation and nitrogenase (nifH) expression in tropical waters of the
eastern North Atlantic," ISME Journal, vol. 5, pp. 1201-1212, Jul 2011.
[31] S. P. Saikia, D. Bora, A. Goswami, K. D. Mudoi, and A. Gogoi, "A review on
the role of Azospirillum in the yield improvement of non leguminous crops,"
African Journal of Microbiology Research, vol. 6, pp. 1085-1102, 2012.
[32] J. K. Vessey, "Plant growth promoting rhizobacteria as biofertilizers," Plant and
Soil, vol. 255, pp. 571-586, 2003/08/01 2003.
[33] J. W. Kloepper and C. J. Beauchamp, "A review of issues related to measuring
colonization of plant roots by bacteria," Canadian Journal of Microbiology, vol.
38, pp. 1219-1232, 1992.
[34] L. Van Loon, "Plant responses to plant growth-promoting rhizobacteria,"
European Journal of Plant Pathology, vol. 119, pp. 243-254, 2007.

5. References
148
[35] E. Somers, J. Vanderleyden, and M. Srinivasan, "Rhizosphere bacterial
signalling: a love parade beneath our feet," Critical Reviews in Microbiology,
vol. 30, pp. 205-240, 2004.
[36] B. Lugtenberg and F. Kamilova, "Plant-growth-promoting rhizobacteria,"
Annual Review of Microbiology, vol. 63, pp. 541-556, 2009.
[37] J. K. Vessey, "Plant growth promoting rhizobacteria as biofertilizers," Plant and
Soil, vol. 255, pp. 571-1157, 2003.
[38] O. Bockman, "Fertilizers and biological nitrogen fixation as sources of plant
nutrients: Perspectives for future agriculture," Plant and Soil, vol. 194, pp. 11-
25, 1997.
[39] M. Staal, S. Rabouille, and L. J. Stal, "On the role of oxygen for nitrogen
fixation in the marine cyanobacterium Trichodesmium sp.," Environmental
Microbiology, vol. 9, pp. 727-736, Mar 2007.
[40] D. T. Welsh, M. Bartoli, D. Nizzoli, G. Castaldelli, S. A. Riou, and P. Viaroli,
"Denitrification, nitrogen fixation, community primary productivity and
inorganic-N and oxygen fluxes in an intertidal Zostera noltii meadow," Marine
Ecology-Progress Series, vol. 208, pp. 65-77, 2000.
[41] C. M. Halbleib and P. W. Ludden, "Regulation of biological nitrogen fixation,"
Journal of Nutrition, vol. 130, pp. 1081-1084, May 2000.
[42] S. N. Dedysh, P. Ricke, and W. Liesack, "NifH and NifD phylogenies: an
evolutionary basis for understanding nitrogen fixation capabilities of
methanotrophic bacteria," Microbiology, vol. 150, pp. 1301-1313, May 2004.
[43] T. Ueda, Y. Suga, N. Yahiro, and T. Matsuguchi, "Remarkable N2-fixing
bacterial diversity detected in rice roots by molecular evolutionary analysis of
Nifh gene sequences," Journal of Bacteriology, vol. 177, pp. 1414-1417, Mar
1995.
[44] G. Steward, J. Zehr, R. Jellison, J. Montoya, and J. Hollibaugh, "Vertical
distribution of nitrogen-fixing phylotypes in a meromictic, hypersaline lake,"
Microbial Ecology, vol. 47, pp. 30-40, 2004.
[45] J. P. Zehr, B. D. Jenkins, S. M. Short, and G. F. Steward, "Nitrogenase gene
diversity and microbial community structure: A cross‐system comparison,"
Environmental Microbiology, vol. 5, pp. 539-554, 2003.
[46] P. Normand, R. Duran, X. Le Roux, C. Morris, and J.C. Poggiale, "Biodiversity
and microbial ecosystems functioning," In Environmental Microbiology:
Fundamentals and Applications, ed: Springer, 2015, pp. 261-291.

5. References
149
[47] J. Zhou, Z. He, Y. Yang, Y. Deng, S. G. Tringe, and L. Alvarez-Cohen, "High-
throughput metagenomic technologies for complex microbial community
analysis: Open and closed formats," mBio, vol. 6, pp. e 02288-14, 2015.
[48] A. Venieraki, M. Dimou, P. Pergalis, I. Kefalogianni, I. Chatzipavlidis, and P.
Katinakis, "The genetic diversity of culturable nitrogen-fixing bacteria in the
rhizosphere of wheat," Microbial Ecology, vol. 61, pp. 277-362, 2011.
[49] M. Mirza, S. Mehnaz, P. Normand, C. Prigent-Combaret, Y. Moënne-Loccoz,
R. Bally, et al., "Molecular characterization and PCR detection of a nitrogen-
fixing Pseudomonas strain promoting rice growth," Biology and Fertility of
Soils, vol. 43, pp. 163-170, 2006.
[50] K. A. Malik, R. Bilal, S. Mehnaz, G. Rasul, M. S. Mirza, and S. Ali,
"Association of nitrogen-fixing, plant-growth-promoting rhizobacteria (PGPR)
with kallar grass and rice," Plant and Soil, vol. 194, pp. 37-44, Jul 1997.
[51] C. R. Woese and G. E. Fox, "Phylogenetic structure of the prokaryotic domain:
the primary kingdoms," Proceedings of the National Academy of Sciences, vol.
74, pp. 5088-5090, 1977.
[52] C. R. Woese, O. Kandler, and M. L. Wheelis, "Towards a natural system of
organisms: proposal for the domains Archaea, Bacteria, and Eucarya,"
Proceedings of the National Academy of Sciences, vol. 87, pp. 4576-4579, 1990.
[53] W. B. Whitman, T. L. Bowen, and D. R. Boone, ". The methanogenic bacteria.
The prokaryotes: In a handbook on the biology of bacteria: ecophysiology,
isolation, identification, applications, vol. I., (Ed. 2), Springer 719-767, 1992.
[54] N. Belay, R. Sparling, B. S. Choi, M. Roberts, J. E. Roberts, and L. Daniels,
"Physiological and N-15-nmr analysis of molecular nitrogen-fixation by
Methanococcus-Thermolithotrophicus, Methanobacterium-Bryantii and
Methanospirillum-Hungatei," Biochimica et biophysica acta, vol. 971, pp. 233-
245, Oct 7 1988.
[55] E. F. DeLong, "Archaea in coastal marine environments," Proceedings of the
National Academy of Sciences, vol. 89, pp. 5685-5689, 1992.
[56] J. A. Fuhrman, "Novel major archaebacterial group from marine plankton,"
Nature, vol. 356, pp. 148-149, 1992.
[57] E. L. Madsen, "Identifying microorganisms responsible for ecologically
significant biogeochemical processes," Nature Reviews Microbiology, vol. 3,
pp. 439-446, 2005.

5. References
150
[58] P. Offre, J. I. Prosser, and G. W. Nicol, "Growth of ammonia‐oxidizing archaea
in soil microcosms is inhibited by acetylene," FEMS microbiology ecology, vol.
70, pp. 99-108, 2009.
[59] J. Mangmang, R. Deaker, and G. Rogers, "Effects of plant growth promoting
rhizobacteria on seed germination characteristics of tomato and lettuce,"
Journal of Tropical Crop Science, vol. 1(2), 2015.
[60] S. Shrivastava, D. Egamberdieva, and A. Varma, "Plant growth-promoting
rhizobacteria (PGPR) and medicinal plants: The State of the Art," In Plant
Growth Promoting Rhizobacteria (PGPR) and Medicinal Plants, ed: Springer,
2015, pp. 1-16.
[61] A. Alsohim, "Molecular and physiological characterization of plant growth
promoting rhizobacteria from rhizosphere soil in Al-Qassim, Saudi Arabia,"
Journal of Food, Agriculture & Environment 13.2:119, 2015.
[62] F. Xun, B. Xie, S. Liu, and C. Guo, "Effect of plant growth-promoting bacteria
(PGPR) and arbuscular mycorrhizal fungi (AMF) inoculation on oats in saline-
alkali soil contaminated by petroleum to enhance phytoremediation,"
Environmental Science and Pollution Research, vol. 22, pp. 598-608, 2015.
[63] R. Prasad, M. Kumar, and A. Varma, "Role of PGPR in soil fertility and plant
health," in Plant-Growth-Promoting Rhizobacteria (PGPR) and Medicinal
Plants, ed: Springer, 2015, pp. 247-260.
[64] H. Kaymak, F. Yarali, I. Guvenc, and M. F. Donmez, "The effect of inoculation
with plant growth rhizobacteria (PGPR) on root formation of mint (Mentha
piperita L.) cuttings," African journal of Biotechnology, vol. 7, 2008.
[65] M. Manjunath, R. Prasanna, P. Sharma, L. Nain, and R. Singh, "Developing
PGPR consortia using novel genera Providencia and Alcaligenes along with
cyanobacteria for wheat," Archives of Agronomy and Soil Science, vol. 57, pp.
873-887, 2011.
[66] K. Malik, R. Bilal, S. Mehnaz, G. Rasul, M. Mirza, and S. Ali, "Association of
nitrogen-fixing, plant-growth-promoting rhizobacteria (PGPR) with kallar grass
and rice," Plant and Soil, vol. 194, pp. 37-44, 1997.
[67] A. Kumar, A. Kumar, S. Devi, S. Patil, C. Payal, and S. Negi, "Isolation,
screening and characterization of bacteria from rhizospheric soils for different
plant growth promotion (PGP) activities: an in vitro study," Recent Research in
Science and Technology, vol. 4, 2012.
[68] S. Compant, J. Nowak, T. Coenye, C. Clément, and E. A. Barka, "Diversity and
occurrence of Burkholderia spp. in the natural environment," FEMS
Microbiology Reviews, vol. 32, pp. 607-626, 2008.

5. References
151
[69] A. Gupta, A. Saxena, M. Gopal, and K. Tilak, "Effect of plant growth promoting
rhizobacteria on competitive ability of introduced Bradyrhizobium sp.(Vigna)
for nodulation," Microbiological Research, vol. 153, pp. 113-117, 1998.
[70] V. Govindasamy, M. Senthilkumar, V. Magheshwaran, U. Kumar, P. Bose, V.
Sharma, et al., "Bacillus and Paenibacillus spp.: potential PGPR for
sustainable agriculture," In Plant growth and Health Promoting Bacteria, ed:
Springer, 2011, pp. 333-364.
[71] S. Kumar and K. B. Rao, "Biological nitrogen fixation: a review," IJALS, vol.
1, pp. 1-9, 2012.
[72] S. Poonguzhali, M. Madhaiyan, and T. Sa, "Isolation and identification of
phosphate solubilizing bacteria from chinese cabbage and their effect on growth
and phosphorus utilization of plants," Journal of Microbiology and
Biotechnology, vol. 18, pp. 773-7, Apr 2008.
[73] A. Vassilev, J.-P. Schwitzguébel, T. Thewys, D. van der Lelie, and J.
Vangronsveld, "The use of plants for remediation of metal-contaminated soils,"
The Scientific World Journal, vol. 4, pp. 9-34, 2004.
[74] V. Mohan and S. Menon, "Diversity status of beneficial microflora in saline
soils of Tamil Nadu and Pudhucherry in Southern India," Journal of Academia
and Industrial Research (JAIR), vol. 3, p. 384, 2015.
[75] Y. Chen, P. Rekha, A. Arun, F. Shen, W.-A. Lai, and C. Young, "Phosphate
solubilizing bacteria from subtropical soil and their tricalcium phosphate
solubilizing abilities," Applied Soil Ecology, vol. 34, pp. 33-41, 2006.
[76] M. Puente, Y. Bashan, C. Li, and V. Lebsky, "Microbial populations and
activities in the rhizoplane of rock‐weathering desert plants. I. Root colonization
and weathering of igneous rocks," Plant Biology, vol. 6, pp. 629-642, 2004.
[77] M. Rashid, S. Khalil, N. Ayub, S. Alam, and F. Latif, "Organic acids production
and phosphate solubilization by phosphate solubilizing microorganisms (PSM)
under in vitro conditions," Pakistan Journal of Biological Sciences, vol. 7, pp.
187-196, 2004.
[78] P. Delvasto, A. Valverde, A. Ballester, J. Igual, J. Muñoz, F. González, et al.,
"Characterization of brushite as a re-crystallization product formed during
bacterial solubilization of hydroxyapatite in batch cultures," Soil Biology and
Biochemistry, vol. 38, pp. 2645-2654, 2006.
[79] S. Scheffknecht, R. Mammerler, S. Steinkellner, and H. Vierheilig, "Root
exudates of mycorrhizal tomato plants exhibit a different effect on microconidia
germination of Fusarium oxysporum sp. lycopersici than root exudates from
non-mycorrhizal tomato plants," Mycorrhiza, vol. 16, pp. 365-370, 2006.

5. References
152
[80] P. Illmer and F. Schinner, "Solubilization of inorganic phosphates by
microorganisms isolated from forest soils," Soil Biology and Biochemistry, vol.
24, pp. 389-395, 1992.
[81] B. R. Glick, "The enhancement of plant growth by free-living bacteria,"
Canadian Journal of Microbiology, vol. 41, pp. 109-117, 1995/02/01 1995.
[82] B. R. Glick, "Plant growth-promoting bacteria: mechanisms and applications,"
Scientifica, vol. 2012, 2012.
[83] A. Idris, N. Labuschagne, and L. Korsten, "Efficacy of rhizobacteria for growth
promotion in sorghum under greenhouse conditions and selected modes of
action studies," The Journal of Agricultural Science, vol. 147, pp. 17-30, 2009.
[84] K. Tilak, N. Ranganayaki, K. Pal, R. De, A. Saxena, C. S. Nautiyal, et al.,
"Diversity of plant growth and soil health supporting bacteria," Current Science,
vol. 89, pp. 136-150, 2005.
[85] T. A. Clark, P. H. Dare, and M. E. Bruce, "Nitrogen fixation in an aerated
stabilization basin treating bleached kraft mill wastewater," Water Environment
Research, vol. 69, pp. 1039-1046, Jul-Aug 1997.
[86] M. Theunis, H. Kobayashi, W. J. Broughton, and E. Prinsen, "Flavonoids,
NodD1, NodD2, and nod-box NB15 modulate expression of the y4wEFG locus
that is required for indole-3-acetic acid synthesis in Rhizobium sp. strain
NGR234," Molecular Plant-Microbe Interactions, vol. 17, pp. 1153-1161,
2004.
[87] O. Steenhoudt and J. Vanderleyden, "Azospirillum, a free-living nitrogen-fixing
bacterium closely associated with grasses: genetic, biochemical and ecological
aspects," FEMS Microbiology Reviews, vol. 24, pp. 487-993, 2000.
[88] E. Prinsen, A. Costacurta, K. Michiels, J. Vanderleyden, and H. Van Onckelen,
"Azospirillum brasilense indole-3-acetic acid biosynthesis: evidence for a non-
tryptophan dependent pathway," Molecular Plant Microbe Interactions, vol. 6,
pp. 609-609, 1993.
[89] F. Cassán, R. Bottini, G. Schneider, and P. Piccoli, "Azospirillum brasilense and
Azospirillum lipoferum hydrolyze conjugates of GA20 and metabolize the
resultant aglycones to GA1 in seedlings of rice dwarf mutants," Plant
Physiology, vol. 125, pp. 2053-2061, 2001.
[90] A. Khalid, S. Tahir, M. Arshad, and Z. A. Zahir, "Relative efficiency of
rhizobacteria for auxin biosynthesis in rhizosphere and non-rhizosphere soils,"
Soil Research, vol. 42, pp. 921-926, 2005.

5. References
153
[91] A. Afzal and A. Bano, "Rhizobium and phosphate solubilizing bacteria improve
the yield and phosphorus uptake in wheat (Triticum aestivum L.)," International
Journal of Agriculture and Biology, vol. 10, pp. 85-88, 2008.
[92] A. Alagawadi and A. Gaur, "Inoculation of Azospirillum brasilense and
phosphate-solubilizing bacteria on yield of sorghum Sorghum bicolor L.
Moench in dry land," Tropical Agriculture, vol. 69, pp. 347-350, 1992.
[93] E. Elkoca, F. Kantar, and F. Sahin, "Influence of nitrogen fixing and phosphorus
solubilizing bacteria on the nodulation, plant growth, and yield of chickpea,"
Journal of Plant Nutrition, vol. 31, pp. 157-171, 2007.
[94] B. S. Mirza, M. S. Mirza, A. Bano, and K. A. Malik, "Co-inoculation of
chickpea with Rhizobium isolates from roots and nodules and phytohormone-
producing Enterobacter strains," Animal Production Science, vol. 47, pp. 1008-
1015, 2007.
[95] B. Shaharoona, M. Arshad, and Z. Zahir, "Effect of plant growth promoting
rhizobacteria containing ACC‐deaminase on maize (Zea mays L.) growth under
axenic conditions and on nodulation in mung bean (Vigna radiata L.)," Letters
in Applied Microbiology, vol. 42, pp. 155-159, 2006.
[96] G. S. Khush, "Genetically modified crops: the fastest adopted crop technology
in the history of modern agriculture," Agriculture and Food Security, vol. 1, p.
14, 2012.
[97] P. Jepson, B. Croft, and G. Pratt, "Test systems to determine the ecological risks
posed by toxin release from Bacillus thuringiensis genes in crop plants,"
Molecular Ecology, vol. 3, pp. 81-89, 1994.
[98] R. Pinton, Z. Varanini, and P. Nannipieri, The rhizosphere: In biochemistry and
organic substances at the soil-plant interface, CRC press, 2007, pp. 1-25.
[99] D. K. Angela and W. T. Eric, "Microbial commmunication and their interactions
in soil and rhizospere ecosystem " Annual Review of Microbiology, vol. 56,
2002.
[100] M. S. Khan, A. Zaidi, P. Wani, M. Ahemad, and M. Oves, "Functional diversity
among plant growth-promoting rhizobacteria: current status," In Microbial
Strategies for Crop Improvement, ed: Springer, 2009, pp. 105-132.
[101] Z. A. Siddiqui and M. S. Akhtar, "Synergistic effects of antagonistic fungi and
a plant growth promoting rhizobacterium, an arbuscular mycorrhizal fungus, or
composted cow manure on populations of Meloidogyne incognita and growth
of tomato," Biocontrol Science and Technology, vol. 18, pp. 279-290, 2008.

5. References
154
[102] K. E. Dunfield and J. J. Germida, "Impact of genetically modified crops on soil-
and plant-associated microbial communities," Journal of Environmental
Quality, vol. 33, pp. 806-815, 2004.
[103] M. Raubuch, K. Roose, K. Warnstorff, F. Wichern, and R. G. Joergensen,
"Respiration pattern and microbial use of field-grown transgenic Bt maize
residues," Soil Biology and Biochemistry, vol. 39, pp. 2380-2389, 2007.
[104] M. Mendelsohn, J. Kough, Z. Vaituzis, and K. Matthews, "Are Bt crops safe?,"
Nature Biotechnology, vol. 21, pp. 1003-1009, 2003.
[105] C. Palm, R. Seidler, D. Schaller, and K. Donegan, "Persistence in soil of
transgenic plant produced Bacillus thuringlensis var. kurstaki δ-endotoxin,"
Canadian Journal of Microbiology, vol. 42, pp. 1258-1262, 1996.
[106] G. Zwart, B. C. Crump, M. P. Kamst-van Agterveld, F. Hagen, and S.-K. Han,
"Typical freshwater bacteria: an analysis of available 16S rRNA gene sequences
from plankton of lakes and rivers," Aquatic Microbial Ecology, vol. 28, 2002.
[107] I. Ahrenholtz, K. Harms, J. de Vries, and W. Wackernagel, "Increased Killing
of Bacillus subtilison the Hair Roots of Transgenic T4 Lysozyme-Producing
Potatoes," Applied and Environmental Microbiology, vol. 66, pp. 1862-1865,
2000.
[108] M. J. Brimecombe, F. A. De Leij, and J. M. Lynch, "The effect of root exudates
on rhizosphere microbial populations," The Rhizosphere. Marcel Dekker, New
York, pp. 95-140, 2001.
[109] H. Zhang, L. Chen, S. Zhao, J. Ren, and X. Cao, "Knockout of itul Gene of
Bacillus subtilis S44 Strain and Impact of its," Plant Pathology Journal, vol.
13, pp. 125-132, 2014.
[110] J. Neal Jr, R. I. Larson, and T. Atkinson, "Changes in rhizosphere populations
of selected physiological groups of bacteria related to substitution of specific
pairs of chromosomes in spring wheat," Plant and Soil, vol. 39, pp. 209-212,
1973.
[111] W. Wei-xiang, Y. Qing-fu, M. Hang, D. Xue-jun, and J. Wen-ming, "Bt-
transgenic rice straw affects the culturable microbiota and dehydrogenase and
phosphatase activities in a flooded paddy soil," Soil Biology and Biochemistry,
vol. 36, pp. 289-295, 2004.
[112] T. E. Cheeke, H. Darby, T. N. Rosenstiel, J. D. Bever, and M. B. Cruzan, "Effect
of Bacillus thuringiensis (Bt) maize cultivation history on arbuscular
mycorrhizal fungal colonization, spore abundance and diversity, and plant
growth," Agriculture, Ecosystems & Environment, vol. 195, pp. 29-35, 2014.

5. References
155
[113] A. Fliebbach, M. Messmer, B. Nietlispach, V. Infante, and P. Mäder, "Effects
of conventionally bred and Bacillus thuringiensis (Bt) maize varieties on soil
microbial biomass and activity," Biology and Fertility of Soils, vol. 48, pp. 315-
324, 2012.
[114] S. R. Cotta, A. C. F. Dias, I. E. Marriel, F. D. Andreote, L. Seldin, and J. D. van
Elsas, "Different effects of transgenic maize and nontransgenic maize on
nitrogen-transforming archaea and bacteria in tropical soils," Applied and
environmental microbiology, vol. 80, pp. 6437-6445, 2014.
[115] S. Siciliano and J. Germida, "Taxonomic diversity of bacteria associated with
the roots of field‐grown transgenic Brassica napus cv. Quest, compared to the
non‐transgenic B. napus cv. Excel and B. rapa cv. Parkland," FEMS
Microbiology Ecology, vol. 29, pp. 263-272, 1999.
[116] B. Griffiths, I. Geoghegan, and W. Robertson, "Testing genetically engineered
potato, producing the lectins GNA and Con A, on non‐target soil organisms and
processes," Journal of Applied Ecology, vol. 37, pp. 159-170, 2000.
[117] H. Heuer, R. M. Kroppenstedt, J. Lottmann, G. Berg, and K. Smalla, "Effects
of T4 lysozyme release from transgenic potato roots on bacterial rhizosphere
communities are negligible relative to natural factors," Applied and
Environmental Microbiology, vol. 68, pp. 1325-1335, 2002.
[118] K. K. Rosenbaum, G. L. Miller, R. J. Kremer, and K. W. Bradley, "Interactions
between glyphosate, Fusarium infection of common waterhemp (Amaranthus
rudis), and soil microbial abundance and diversity in soil collections from
Missouri," Weed Science, vol. 62, pp. 71-82, 2014.
[119] M. Tesfaye, N. S. Dufault, M. R. Dornbusch, D. L. Allan, C. P. Vance, and D.
A. Samac, "Influence of enhanced malate dehydrogenase expression by alfalfa
on diversity of rhizobacteria and soil nutrient availability," Soil biology and
biochemistry, vol. 35, pp. 1103-1113, 2003.
[120] K. K. Donegan and R. J. Seidler, Effects of transgenic plants on soil and plant
microorganisms: US Environmental Protection Agency, National Health and
Environmental Effects Research Laboratory, Western Ecology Division, pp.
415-424, 1999.
[121] Y. Lv, H. Cai, J. Yu, J. Liu, Q. Liu, and C. Guo, "Biosafety assessment of GFP
transplastomic tobacco to rhizosphere microbial community," Ecotoxicology,
vol. 23, pp. 718-725, 2014.
[122] K. E. Dunfield and J. J. Germida, "Seasonal changes in the rhizosphere
microbial communities associated with field-grown genetically modified canola
(Brassica napus)," Applied and Environmental Microbiology, vol. 69, pp. 7310-
7318, 2003.

5. References
156
[123] J. Germida and S. Siciliano, "Taxonomic diversity of bacteria associated with
the roots of modern, recent and ancient wheat cultivars," Biology and Fertility
of Soils, vol. 33, pp. 410-415, 2001.
[124] K. Burris, A. Mentewab, S. Ripp, and C. N. Stewart, "An Arabidopsis thaliana
ABC transporter that confers kanamycin resistance in transgenic plants does not
endow resistance to Escherichia coli," Journal of Microbial Biotechnology, vol.
1, pp. 191-195, 2008.
[125] J. Handelsman, "Metagenomics: application of genomics to uncultured
microorganisms," Microbiology and molecular biology reviews : Microbiology
and Molecular Biology Reviews, vol. 68, pp. 669-754, 2004.
[126] D. S. Patrick and H. Jo, " Metagenomics for studying unculturable
microorganisms: cutting the Gordian knot.," Genome Biology, vol. 6, 2005.
[127] J. M. Kinross, A. W. Darzi, and J. K. Nicholson, "Gut microbiome-host
interactions in health and disease," Genome Medicine, vol. 3, p. 14, 2011.
[128] H. Nagayama, T. Sugawara, R. Endo, A. Ono, H. Kato, Y. Ohtsubo, et al.,
"Isolation of oxygenase genes for indigo-forming activity from an artificially
polluted soil metagenome by functional screening using Pseudomonas putida
strains as hosts," Applied Microbiology and Biotechnology, pp. 1-18, 2015.
[129] S. Mocali and A. Benedetti, "Exploring research frontiers in microbiology: the
challenge of metagenomics in soil microbiology," Reserch in Microbiology,
vol. 161, pp. 497-505, Jul-Aug 2010.
[130] S. G. Tringe, C. Von Mering, A. Kobayashi, A. A. Salamov, K. Chen, H. W.
Chang, et al., "Comparative metagenomics of microbial communities," Science,
vol. 308, pp. 554-557, 2005.
[131] V. Acosta-Martínez, J. Cotton, T. Gardner, J. Moore-Kucera, J. Zak, D. Wester,
et al., "Predominant bacterial and fungal assemblages in agricultural soils
during a record drought/heat wave and linkages to enzyme activities of
biogeochemical cycling," Applied Soil Ecology, vol. 84, pp. 69-82, 2014.
[132] R. Daniel, "The soil metagenome--a rich resource for the discovery of novel
natural products," Current Opinion in Biotechnology, vol. 15, pp. 199-403,
2004.
[133] D. Rolf, "The metagenomics of soil," Nature Reviews Microbiology, vol. 3,
2005.

5. References
157
[134] Y. Zhou, W. Liu, and H. Ye, "Effects of pesticides metolachlor and S-
metolachlor on soil microorganisms in aquisols. II. Soil respiration," Ying Yong
Sheng Tai Xue Bao, vol. 17, pp. 1305-9, Jul 2006.
[135] G. M. Garrity and J. G. Holt, "The road map to the manual," In Bergey’s
Manual® of Systematic Bacteriology, ed: Springer, 2001, pp. 119-166.
[136] J. Beynon, A. Ally, M. Cannon, F. Cannon, M. Jacobson, V. Cash, et al.,
"Comparative organization of nitrogen fixation-specific genes from
Azotobacter-Vinelandii and Klebsiella-Pneumoniae - DNA-Sequence of the
Nifusv Genes," Journal of Bacteriology, vol. 169, pp. 4024-4029, Sep 1987.
[137] G. P. Roberts, T. Macneil, D. Macneil, and W. J. Brill, "Regulation and
characterization of protein products coded by nif (Nitrogen-Fixation) genes of
Klebsiella-Pneumoniae," Journal of Bacteriology, vol. 136, pp. 267-279, 1978.
[138] B. Valdivia, M. Dughri, and P. Bottomley, "Antigenic and symbiotic
characterization of indigenous Rhizobium leguminosarum bv. tripolii recovered
from root nodules of Trifolium pratense L. sown into subterranean clover
pasture soils," Soil Biology and Biochemistry, vol. 20, pp. 267-274, 1988.
[139] R. W. Miller and J. C. Sirois, "Relative efficacy of different alfalfa cultivar-
Rhizobium meliloti strain combinations for symbiotic nitrogen-fixation,"
Applied and Environmental Microbiology, vol. 43, pp. 764-768, 1982.
[140] R. A. Zimmermann, A. Muto, P. Fellner, C. Ehresmann, and C. Branlant,
"Location of ribosomal protein binding sites on 16S ribosomal RNA,"
Proceedings of the National Academy of Sciences, vol. 69, pp. 1282-1286, 1972.
[141] E. Stackebrandt, B. Lewis, and C. Woese, "The phylogenetic structure of the
coryneform group of bacteria," Zentralblatt für Bakteriologie: I. Abt. Originale
C: Allgemeine, angewandte und ökologische Mikrobiologie, vol. 1, pp. 137-
149, 1980.
[142] K. Greisen, M. Loeffelholz, A. Purohit, and D. Leong, "PCR primers and probes
for the 16S rRNA gene of most species of pathogenic bacteria, including
bacteria found in cerebrospinal fluid," Journal of Clinical Microbiology, vol.
32, pp. 335-351, 1994.
[143] J. E. Clarridge, "Impact of 16S rRNA gene sequence analysis for identification
of bacteria on clinical microbiology and infectious diseases," Clinical
Microbiology Reviews, vol. 17, pp. 840-862, 2004.
[144] U. Edwards, T. Rogall, H. Blocker, M. Emde, and E. C. Bottger, "Isolation and
direct complete nucleotide determination of entire genes. Characterization of a
gene coding for 16S ribosomal RNA," Nucleic Acids Research, vol. 17, pp.
7843-53, Oct 11 1989.

5. References
158
[145] S. Weidner, A. Puhler, and H. Kuster, "Genomics insights into symbiotic
nitrogen fixation," Current Opinion in Biotechnology, vol. 14, pp. 200-205, Apr
2003.
[146] W. G. Weisburg, S. M. Barns, D. A. Pelletier, and D. J. Lane, "16S ribosomal
DNA amplification for phylogenetic study," Journal of bacteriology, vol. 173,
pp. 697-703, 1991.
[147] L. F. Roesch, R. R. Fulthorpe, A. Riva, G. Casella, A. K. Hadwin, A. D. Kent,
et al., "Pyrosequencing enumerates and contrasts soil microbial diversity," The
ISME Journal, vol. 1, pp. 283-290, 2007.
[148] M. Ronaghi, S. Shokralla, and B. Gharizadeh, "Pyrosequencing for discovery
and analysis of DNA sequence variations," Pharmacogenomics vol. 8, pp. 1437-
1441, 2007.
[149] O.-S. Kim, Y.-J. Cho, K. Lee, S.-H. Yoon, M. Kim, H. Na, et al., "Introducing
EzTaxon-e: a prokaryotic 16S rRNA gene sequence database with phylotypes
that represent uncultured species," International Journal of Systematic and
Evolutionary Microbiology, vol. 62, pp. 716-721, 2012.
[150] T. Z. DeSantis, E. L. Brodie, J. P. Moberg, I. X. Zubieta, Y. M. Piceno, and G.
L. Andersen, "High-density universal 16S rRNA microarray analysis reveals
broader diversity than typical clone library when sampling the environment,"
Microbial Ecology, vol. 53, pp. 371-383, 2007.
[151] L. F. W. Roesch, P. D. de Quadros, F. A. O. Camargo, and E. W. Triplett,
"Screening of diazotrophic bacteria Azopirillum spp. for nitrogen fixation and
auxin production in multiple field sites in southern Brazil," World Journal of
Microbiology & Biotechnology, vol. 23, pp. 1377-1383, Oct 2007.
[152] S. M. Huse, L. Dethlefsen, J. A. Huber, D. M. Welch, D. A. Relman, and M. L.
Sogin, "Exploring microbial diversity and taxonomy using SSU rRNA
hypervariable tag sequencing," PLOS Genetics, vol. 4, p. e1000255, 2008.
[153] G. Rastogi, J. J. Tech, G. L. Coaker, and J. H. Leveau, "A PCR-based toolbox
for the culture-independent quantification of total bacterial abundances in plant
environments," Journal of Microbiological Methods, vol. 83, pp. 127-132,
2010.
[154] H. E. Jakobsson, C. Jernberg, A. F. Andersson, M. Sjölund-Karlsson, J. K.
Jansson, and L. Engstrand, "Short-term antibiotic treatment has differing long-
term impacts on the human throat and gut microbiome," PLOS One, vol. 5, p.
e9836, 2010.
[155] S. Weidner, W. Arnold, and A. Puhler, "Diversity of uncultured microorganisms
associated with the seagrass Halophila stipulacea estimated by restriction

5. References
159
fragment length polymorphism analysis of PCR-amplified 16S rRNA genes,"
Applied and Environmental Microbiology, vol. 62, pp. 766-771, 1996.
[156] N. Sangwan, P. Lata, V. Dwivedi, A. Singh, N. Niharika, J. Kaur, et al.,
"Comparative metagenomic analysis of soil microbial communities across three
hexachlorocyclohexane contamination levels," PLOS One, vol. 7, pp. e46219,
2012.
[157] M. Kim, M. Morrison, and Z. Yu, "Evaluation of different partial 16S rRNA
gene sequence regions for phylogenetic analysis of microbiomes," Journal of
Microbiological Methods, vol. 84, pp. 81-87, 2011.
[158] H. Wu, W. Wu, Z. Chen, W. Wang, G. Zhou, T. Kajiyama, et al., "Highly
sensitive pyrosequencing based on the capture of free adenosine 5′
phosphosulfate with adenosine triphosphate sulfurylase," Analytical chemistry,
vol. 83, pp. 3600-3605, 2011.
[159] P. Gyaneshwar, E. K. James, N. Mathan, P. M. Reddy, B. Reinhold-Hurek, and
J. K. Ladha, "Endophytic colonization of rice by a diazotrophic strain of
Serratia marcescens," Journal of Bacteriology, vol. 183, pp. 2634-2645, 2001.
[160] L. Liu, Y. Wu, Y. X. He, N. Wu, G. Sun, L. Zhang, et al., "Effects of seasonal
snow cover on soil nitrogen transformation in alpine ecosystem: A review,"
Ying Yong Sheng Tai Xue Bao, vol. 22, pp. 2193-200, Aug 2011.
[161] F. Poly, L. Ranjard, S. Nazaret, F. Gourbière, and L. J. Monrozier, "Comparison
of nifH gene pools in soils and soil microenvironments with contrasting
properties," Applied and Environmental Microbiology, vol. 67, pp. 2255-2262,
2001.
[162] K. Zareen and L. D. Sharon, "Characterization of bacterial endophytes of sweet
potato plants," Plant and Soil, vol. 322, 2009.
[163] O. Babalola, "Beneficial bacteria of agricultural importance," Biotechnology
Letters, vol. 32, pp. 1559-1629, 2010.
[164] D. Blaha, H. Sanguin, P. Robe, R. Nalin, R. Bally, and Y. Moënne-Loccoz,
"Physical organization of phytobeneficial genes nifH and ipdC in the plant
growth-promoting rhizobacterium Azospirillum lipoferum 4VI," FEMS
Microbiology Letters, vol. 244, pp. 157-163, 2005.
[165] Y. B. Guo, J. Li, L. Li, F. Chen, W. Wu, J. Wang, et al., "Mutations that disrupt
either the pqq or the gdh gene of Rahnella aquatilis abolish the production of an
antibacterial substance and result in reduced biological control of grapevine
crown gall," Applied and Environmental Microbiology, vol. 75, pp. 6792-6803,
2009.

5. References
160
[166] C. C. Young, P. D. Rekha, W. A. Lai, and A. B. Arun, "Encapsulation of plant
growth-promoting bacteria in alginate beads enriched with humic acid,"
Biotechnology and Bioengineering, vol. 95, pp. 76-83, Sep 5 2006.
[167] B. Weng, X. Xie, J. Yang, J. Liu, H. Lu, and C. Yan, "Research on the nitrogen
cycle in rhizosphere of Kandelia obovata under ammonium and nitrate
addition," Marine Pollution Bulletin, vol. 76, pp. 227-240, 2013.
[168] B. S. Mirza and J. L. Rodrigues, "Development of a direct isolation procedure
for free-living diazotrophs under controlled hypoxic conditions," Applied and
Environmental Microbiology, vol. 78, pp. 5542-5549, 2012.
[169] C. Rösch, A. Mergel, and H. Bothe, "Biodiversity of denitrifying and
dinitrogen-fixing bacteria in an acid forest soil," Applied and Environmental
Microbiology, vol. 68, pp. 3818-3829, 2002.
[170] F. Poly, L. Ranjard, S. Nazaret, F. Gourbière, and L. Monrozier, "Comparison
of nifH gene pools in soils and soil microenvironments with contrasting
properties," Applied and Environmental Microbiology, vol. 67, pp. 2255-2317,
2001.
[171] F. Widmer, B. Shaffer, L. Porteous, and R. Seidler, "Analysis of nifH gene pool
complexity in soil and litter at a Douglas fir forest site in the Oregon cascade
mountain range," Applied and Environmental Microbiology, vol. 65, pp. 374-
454, 1999.
[172] T. Ueda, Y. Suga, N. Yahiro, and T. Matsuguchi, "Remarkable N2 fixing
bacterial diversity detected in rice roots by molecular evolutionary analysis of
nifH gene sequences," Journal of Bacteriology, vol. 177, pp. 1414-1417, 1995.
[173] M. Demba Diallo, B. Reinhold-Hurek, and T. Hurek, "Evaluation of PCR
primers for universal nifH gene targeting and for assessment of transcribed nifH
pools in roots of Oryza longistaminata with and without low nitrogen input,"
FEMS Microbiol Ecology, vol. 65, pp. 220-8, Aug 2008.
[174] J. P. Zehr, D. Harris, B. Dominic, and J. Salerno, "Structural analysis of the
Trichodesmium nitrogenase iron protein: implications for aerobic nitrogen
fixation activity," FEMS Microbiology Letters, vol. 153, pp. 303-309, Aug 15
1997.
[175] J. Young, L. Kuykendall, E. Martinez-Romero, A. Kerr, and H. Sawada, "A
revision of Rhizobium Frank 1889, with an emended description of the genus,
and the inclusion of all species of Agrobacterium Conn 1942 and Allorhizobium
undicola de Lajudie et al. 1998 as new combinations: Rhizobium radiobacter,
R. rhizogenes, R. rubi, R. undicola and R. vitis," International Journal of
Systematic and Evolutionary Microbiology, vol. 51, pp. 89-103, 2001.

5. References
161
[176] X. Y. Tan, T. Hurek, and B. Reinhold-Hurek, "Effect of N-fertilization, plant
genotype and environmental conditions on nifH gene pools in roots of rice,"
Environmental Microbiology, vol. 5, pp. 1009-1015, Oct 2003.
[177] Y. Zhang, D. Li, H. Wang, Q. Xiao, and X. Liu, "Molecular diversity of
nitrogen-fixing bacteria from the Tibetan Plateau, China," FEMS Microbiology
Letters, vol. 260, pp. 134-142, 2006.
[178] C. R. Lovell, Y. M. Piceno, J. M. Quattro, and C. E. Bagwell, "Molecular
analysis of diazotroph diversity in the rhizosphere of the smooth cordgrass,
Spartina alterniflora," Applied and Environmental Microbiology, vol. 66, pp.
3814-3822, 2000.
[179] J. A. Izquierdo and K. Nüsslein, "Distribution of extensive nifH gene diversity
across physical soil microenvironments," Microbial Ecology, vol. 51, pp. 441-
452, 2006.
[180] C. J. Smith and A. M. Osborn, "Advantages and limitations of quantitative PCR
(Q-PCR)-based approaches in microbial ecology," FEMS Microbiology
Ecology, vol. 67, pp. 6-20, 2009.
[181] L. G. Lee, C. R. Connell, and W. Bloch, "Allelic discrimination by nick-
translation PCR with fluorgenic probes," Nucleic Acids Research, vol. 21, pp.
3761-3766, 1993.
[182] C. Becker, A. Hammerle-Fickinger, I. Riedmaier, and M. Pfaffl, "mRNA and
microRNA quality control for RT-qPCR analysis," Methods, vol. 50, pp. 237-
243, 2010.
[183] J. C. Mar, Y. Kimura, K. Schroder, K. M. Irvine, Y. Hayashizaki, H. Suzuki, et
al., "Data-driven normalization strategies for high-throughput quantitative RT-
PCR," BMC Bioinformatics, vol. 10, pp. 110, 2009.
[184] M. T. Suzuki, L. T. Taylor, and E. F. DeLong, "Quantitative analysis of small-
subunit rRNA genes in mixed microbial populations via 5′-nuclease assays,"
Applied and Environmental Microbiology, vol. 66, pp. 4605-4614, 2000.
[185] A. Hermansson and P.-E. Lindgren, "Quantification of ammonia-oxidizing
bacteria in arable soil by real-time PCR," Applied and Environmental
Microbiology, vol. 67, pp. 972-976, 2001.
[186] L. Lopez-Bellido, R. J. Lopez-Bellido, R. Redondo, and J. Benitez, "Faba bean
nitrogen fixation in a wheat-based rotation under rainfed Mediterranean
conditions: Effect of tillage system," Field Crops Research, vol. 99, pp. 172-
172, Oct 30 2006.

5. References
162
[187] J. Leloup, A. Loy, N. J. Knab, C. Borowski, M. Wagner, and B. B. Jørgensen,
"Diversity and abundance of sulfate‐reducing microorganisms in the sulfate and
methane zones of a marine sediment, Black Sea," Environmental Microbiology,
vol. 9, pp. 131-142, 2007.
[188] Y. Guo, J. X. Liu, Y. Lu, W. Zhu, S. Denman, and C. McSweeney, "Effect of
tea saponin on methanogenesis, microbial community structure and expression
of mcrA gene, in cultures of rumen micro‐organisms," Letters in Applied
Microbiology, vol. 47, pp. 421-426, 2008.
[189] A. T. Bell, "The impact of nanoscience on heterogeneous catalysis," Science,
vol. 299, pp. 1688-1691, 2003.
[190] S. Lapage, J. E. Shelton, and T. Mitchell, "Chapter I media for the maintenance
and preservation of bacteria," Methods in microbiology, vol. 3, pp. 1-133, 1970.
[191] R. I. Pikovskaya, "Mobilization of phosphorus in soil in connection with vital
activity of some microbial species.," Micrbiology, vol. 17, pp. 362-370, 1948.
[192] Y. Okon, S. L. Albrecht, and R. H. Burris, "Methods for growing Azospirillum
lipoferum and for counting it in pure culture and in association with plants.,"
Applied Environmental Microbiology, vol. 33, pp. 85-8, Jan 1977.
[193] F. S. Watanabe and S. R. Olsen, "Test of an ascorbic acid method for
determining phosphorus in water and NaHCO3 extracts from soil.," Soil Science
Society of America Journal, vol. 29, pp. 677-678, 1965.
[194] T. M. Tien, M. H. Gaskins, and D. H. Hubbell, "Plant growth substances
produced by Azospirillum brasilense and their effect on the growth of pearl
millet (Pennisetum americanum L.)." Applied Environmental Microbiology,
vol. 37, pp. 1016-24, May 1979.
[195] F. M. Ausubel, "Molecular-genetics of symbiotic nitrogen-fixation," Cell, vol.
29, pp. 1-2, 1982.
[196] V. Thompson, "Associative nitrogen fixation, C4 photosynthesis, and the
evolution of spittlebugs (Hemiptera : Cercopidae) as major pests of neotropical
sugarcane and forage grasses," Bulletin of Entomological Research, vol. 94, pp.
189-200, Jun 2004.
[197] B. S. Mirza, A. Welsh, G. Rasul, J. P. Rieder, M. W. Paschke, and D. Hahn,
"Variation in Frankia populations of the Elaeagnus host infection group in
nodules of six host plant species after inoculation with soil," Microbial Ecology,
vol. 58, pp. 384-93, Aug 2009.

5. References
163
[198] J. Díaz-Lago, D. Stuthman, and K. Leonard, "Evaluation of components of
partial resistance to oat crown rust using digital image analysis," Plant Disease,
vol. 87, pp. 667-674, 2003.
[199] N. Fierer and J. Lennon, "The generation and maintenance of diversity in
microbial communities," American Journal of Botany, vol. 98, pp. 439-487,
2011.
[200] F. Poly, L. Monrozier, and R. Bally, "Improvement in the RFLP procedure for
studying the diversity of nifH genes in communities of nitrogen fixers in soil,"
Research in Microbiology, vol. 152, pp. 95-198, 2001.
[201] J. D. Thompson, T. J. Gibson, F. Plewniak, F. Jeanmougin, and D. G. Higgins,
"The CLUSTAL X windows interface: flexible strategies for multiple sequence
alignment aided by quality analysis tools.," Nucleic Acids Research, vol. 25, pp.
4876-82, Dec 15 1997.
[202] S. M. Babur, W. Allana, and H. Dittmar, "Growth of Frankia strains in leaf
litter-amended soil and the rhizosphere of a nonactinorhizal plant," FEMS
Microbiology Ecology, vol. 70, 2009.
[203] D. M. Hillis and J. J. Bull, "An empirical test of bootstrapping as a method for
assessing confidence in phylogenetic analysis," Systematic Biology, vol. 42, pp.
182-192, 1993.
[204] M. K. Chelius and J. E. Lepo, "Restriction fragment length polymorphism
analysis of PCR-amplified nifH sequences from wetland plant rhizosphere
communities," Environmental Technology, vol. 20, pp. 883-889, Aug 1999.
[205] C. Wang, P. F. Wang, and X. Hu, "Removal of COD (Cr) and nitrogen in
severely polluted river water by bank filtration," Environmental Technology,
vol. 28, pp. 649-57, Jun 2007.
[206] M. Héry, L. Philippot, E. Mériaux, F. Poly, X. Le Roux, and E. Navarro, "Nickel
mine spoils revegetation attempts: Effect of pioneer plants on two functional
bacterial communities involved in the N‐cycle," Environmental Microbiology,
vol. 7, pp. 486-498, 2005.
[207] M. Llirós Dupré and C. Borrego, Diversity, dynamics and activity of mesophilic
Archaea in stratified feshwater lakes. Implications in Biogeochemical Cycles:
Universitat de Girona, 2010.
[208] Y. Zheng, M. Dixon, and P. K. Saxena, "Growing environment and nutrient
availability affect the content of some phenolic compounds in Echinacea
Purpurea and Echinacea Angustifolia," Planta Medica, vol. 72, pp. 1407-14,
Dec 2006.

5. References
164
[209] M. C. P. e Silva, A. V. Semenov, J. D. van Elsas, and J. F. Salles, "Seasonal
variations in the diversity and abundance of diazotrophic communities across
soils," FEMS Microbiology Ecology, vol. 77, pp. 57-68, 2011.
[210] J. Keshri, A. Mishra, and B. Jha, "Microbial population index and community
structure in saline–alkaline soil using gene targeted metagenomics,"
Microbiological Research, vol. 168, pp. 165-173, 2013.
[211] M. R. Rodrigues Coelho, M. De Vos, N. P. Carneiro, I. E. Marriel, E. Paiva,
and L. Seldin, "Diversity of nifH gene pools in the rhizosphere of two cultivars
of sorghum (Sorghum bicolor) treated with contrasting levels of nitrogen
fertilizer," FEMS Microbiology Letters, vol. 279, pp. 15-22, 2008.
[212] I. Wartiainen, T. Eriksson, W. Zheng, and U. Rasmussen, "Variation in the
active diazotrophic community in rice paddy—nifH PCR-DGGE analysis of
rhizosphere and bulk soil," Applied Soil Ecology, vol. 39, pp. 65-75, 2008.
[213] H. Böhme, "Regulation of nitrogen fixation in heterocyst-forming
cyanobacteria," Trends in Plant Science, vol. 3, pp. 346-351, 1998.
[214] J. L. Hu, X. G. Lin, R. Yin, H. Y. Chu, H. Y. Zhang, J. H. Wang, et al.,
"Spatiotemporal evolvement of soil microbiological characteristics in upland
fields with different utilization duration in Cixi, Zhejiang Province," Ying Yong
Sheng Tai Xue Bao, vol. 19, pp. 1977-82, Sep 2008.
[215] A. K. Shukla, P. Vishwakarma, R. Singh, S. Upadhyay, and S. K. Dubey, "Bio-
filtration of trichloroethylene using diazotrophic bacterial community,"
Bioresource Technology, vol. 101, pp. 2126-2133, 2010.
[216] O. P. Shukla, U. N. Rai, and S. Dubey, "Involvement and interaction of
microbial communities in the transformation and stabilization of chromium
during the composting of tannery effluent treated biomass of Vallisneria spiralis
L," Bioresources Technology, vol. 100, pp. 2198-203, Apr 2009.
[217] C.-H. Xie and A. Yokota, "Zoogloea oryzae sp. nov., a nitrogen-fixing
bacterium isolated from rice paddy soil, and reclassification of the strain ATCC
19623 as Crabtreella saccharophila gen. nov., sp. nov," International Journal
of Systematic and Evolutionary Microbiology, vol. 56, pp. 619-624, 2006.
[218] I. R. Kennedy and Y. T. Tchan, "Biological nitrogen-fixation in non-
leguminous field crops - recent advances," Plant and Soil, vol. 141, pp. 93-118,
Mar 1992.
[219] C. H. Xie and A. Yokota, "Reclassification of Alcaligenes latus strains IAM
12599T and IAM 12664 and Pseudomonas saccharophila as Azohydromonas
lata gen. nov., comb. nov., Azohydromonas australica sp. nov. and Pelomonas

5. References
165
saccharophila gen. nov., comb. nov., respectively," International Journal of
Systimatics & Evolutionary Microbiology, vol. 55, pp. 2419-25, Nov 2005.
[220] B. M. Szabala, P. Osipowski, and S. Malepszy, "Transgenic crops: the present
state and new ways of genetic modification," Journal of Applied Genetics, vol.
55, pp. 287-294, 2014.
[221] W. Liu, Y. Luo, Y. Teng, Z. Li, and L. Q. Ma, "Bioremediation of oily sludge-
contaminated soil by stimulating indigenous microbes," Environmental
Geochemistry and Health, vol. 32, pp. 23-9, Feb 2010.
[222] R. Pinton, Z. Varanini, and P. Nannipieri, "The rhizosphere as a site of
biochemical interactions among soil components, plants, and microorganisms,"
The Rhizosphere. Marcel Dekker, New York, 2001 pp. 1-14.
[223] M. M. Qaisrani, M. S. Mirza, A. Zaheer, and K. A. Malik, "Isolation and
identification by 16s rrna sequence analysis of achromobacter, Azospirillum and
Rhodococcus strains from the rhizosphere of maize and screening for the
beneficial effect on plant growth," Pakistan Journal of Agricultural Sciences,
vol. 51, pp. 91-99, 2014.
[224] P. P. Reddy, "Potential role of PGPR in agriculture," In plant growth promoting
rhizobacteria for horticultural crop protection, ed: Springer, 2014, pp. 17-34.
[225] Y. Ma, M. Rajkumar, and H. Freitas, "Inoculation of plant growth promoting
bacterium Achromobacter xylosoxidans strain Ax10 for the improvement of
copper phytoextraction by Brassica juncea," Journal of Environmental
Management, vol. 90, pp. 831-837, 2009.
[226] P. Wang, F. Wang, M. Xu, and X. Xiao, "Molecular phylogeny of
methylotrophs in a deep-sea sediment from a tropical west Pacific Warm Pool,"
FEMS Microbiology & Ecology, vol. 47, pp. 77-84, 2004.
[227] S. Banerjee, R. Palit, C. Sengupta, and D. Standing, "Stress induced phosphate
solubilization by'Arthrobacter' sp. and 'Bacillus' sp. isolated from tomato
rhizosphere," Australian Journal of Crop Science, vol. 4, pp. 378, 2010.
[228] S. K. Upadhyay, J. S. Singh, A. K. Saxena, and D. P. Singh, "Impact of PGPR
inoculation on growth and antioxidant status of wheat under saline conditions,"
Plant Biology, vol. 14, pp. 605-611, 2012.
[229] B. Saharan and V. Nehra, "Plant growth promoting rhizobacteria: a critical
review," Life Sciences and Medicine Research, vol. 21, pp. 1-30, 2011.

5. References
166
[230] C. Prathibha, A. Alagawadi, and M. Sreenivasa, "Establishment of inoculated
organisms in rhizosphere and their influence on nutrient uptake and yield of
cotton," Karnataka Journal of Agricultural Sciences, vol. 8, 2012.
[231] S. Mehnaz, B. Weselowski, and G. Lazarovits, "Azospirillum zeae sp. nov., a
diazotrophic bacterium isolated from rhizosphere soil of Zea mays,"
International Journal Of Systematic And Evolutionary Microbiology, vol. 57,
2007.
[232] Q. Guo, W. Dong, S. Li, X. Lu, P. Wang, X. Zhang, et al., "Fengycin produced
by Bacillus subtilis NCD-2 plays a major role in biocontrol of cotton seedling
damping-off disease," Microbiological Research, vol. 169, pp. 533-540, 2014.
[233] K. U. Sudhir, P. S. Devendra, and S. Ratul, "Genetic diversity of plant growth
promoting rhizobacteria isolated from rhizospheric soil of wheat under saline
condition," Current Microbiology, vol. 59, pp. 317-320, 2009.
[234] S. Yasmin, F. Y. Hafeez, M. Schmid, and A. Hartmann, "Plant-beneficial
rhizobacteria for sustainable increased yield of cotton with reduced level of
chemical fertilizers," Pakistan Journal of Botany, vol. 45, pp. 655-662, 2013.
[235] W. Fernando, S. Nakkeeran, Y. Zhang, and S. Savchuk, "Biological control of
Sclerotinia sclerotiorum L. de Bary by Pseudomonas and Bacillus species on
canola petals," Crop Protection, vol. 26, pp. 100-107, 2007.
[236] O. Shida, H. Takagi, K. Kadowaki, and K. Komagata, "Proposal for two new
genera, Brevibacillus gen. nov. and Aneurinibacillus gen. nov," International
Journal of Systematic Bacteriology, vol. 46, pp. 939-946, 1996.
[237] R. Arya and A. K. Sharma, "Screening, isolation and characterization of
Brevibacillus borstelensis for the bioremediation of carbendazim." Journal of
Environmental Science and Sustainability, vol. 2, pp. 12 – 14, 2014
[238] X. H. Nguyen, K. W. Naing, Y. S. Lee, and K. Y. Kim, "Isolation of Butyl 2, 3‐Dihydroxybenzoate From Paenibacillus elgii HOA73 Against Fusarium
oxysporum sp. Lycopersici," Journal of Phytopathology, 2014.
[239] P. K. Pindi, T. Sultana, and P. K. Vootla, "Plant growth regulation of Bt-cotton
through Bacillus species," 3 Biotech, vol. 4, pp. 305-315, 2014.
[240] P. Joshi and A. Bhatt, "Diversity and function of plant growth promoting
rhizobacteria associated with wheat rhizosphere in North Himalayan Region,"
International Journal of Environmental Sciences, vol. 1, pp. 1135-1143, 2011.

5. References
167
[241] A. Khalid, M. Arshad, and Z. Zahir, "Screening plant growth‐promoting
rhizobacteria for improving growth and yield of wheat," Journal of Applied
Microbiology, vol. 96, pp. 473-480, 2004.
[242] P. Singh, V. Kumar, and S. Agrawal, "Evaluation of phytase producing bacteria
for their plant growth promoting activities," International Journal of
Microbiology, vol. 2014, 2014.
[243] B. R. Glick, “Some Techniques to Elaborate Plant–Microbe Interactions,” In
Beneficial Plant-Bacterial Interactions” ed: New York, Springer, 2015, pp 97-
122.
[244] B. Sarkar, A. K. Patra, and T. J. Purakayastha, "Transgenic Bt -cotton affects
enzyme activity and nutrient availability in a sub-tropical Inceptisol," Journal
of Agronomy and Crop Science, vol. 194, 2008.
[245] M. Arshad and W. Frankenberger Jr, "Microbial production of plant hormones,"
Plant and soil, vol. 133, pp. 1-8, 1991.
[246] O. Babalola, E. Osir, and A. Sanni, "Amplification of 1-amino-cyclopropane-1-
carboxylic (ACC) deaminase from plant growth promoting rhizobacteria in
Striga-infested soil," African Journal of Biotechnology, vol. 2, pp. 157-160,
2004.
[247] R. Farina, A. Beneduzi, A. Ambrosini, S. B. de Campos, B. B. Lisboa, V.
Wendisch, et al., "Diversity of plant growth-promoting rhizobacteria
communities associated with the stages of canola growth," Applied Soil
Ecology, vol. 55, pp. 44-52, 2012.
[248] P. Jha and A. Kumar, "Characterization of novel plant growth promoting
endophytic bacterium Achromobacter xylosoxidans from wheat plant,"
Microbial Ecology, vol. 58, pp. 179-88, Jul 2009.
[249] A. P. G. C. Marques, C. Pires, H. Moreira, A. O. S. S. Rangel, and P. M. L.
Castro, "Assessment of the plant growth promotion abilities of six bacterial
isolates using Zea mays as indicator plant," Soil Biology and Biochemistry, vol.
42, pp. 1229-1235, 2010.
[250] N. Singh, F. Chaudhary, and D. Patel, "Effectiveness of Azotobacter bio-
inoculant for wheat grown under dryland condition," Journal of Environmental
Biology, vol. 34, pp. 927-932, 2013.
[251] S. Banerjee, R. Palit, C. Sengupta, and D. Standing, "Stress induced phosphate
solubilization by Arthrobacter sp. and Bacillus sp. isolated from tomato
rhizosphere," Australian Journal of Crop Science, vol. 4, pp. 378-383, Aug
2010.

5. References
168
[252] Y. Bashan and L. E. De-Bashan, "Chapter Two-How the Plant Growth-
Promoting Bacterium Azospirillum Promotes Plant Growth: A Critical
Assessment," Advances in Agronomy, vol. 108, pp. 77-136, 2010.
[253] L. Halda-Alija, "Identification of indole-3-acetic acid producing freshwater
wetland rhizosphere bacteria associated with Juncus effusus L," Canadian
Journal of Microbiology, vol. 49, pp. 781-787, 2003.
[254] Y. Erturk, S. Ercisli, A. Haznedar, and R. Cakmakci, "Effects of plant growth
promoting rhizobacteria (PGPR) on rooting and root growth of kiwifruit
(Actinidia deliciosa) stem cuttings," Biological Research, vol. 43, pp. 91-98,
2010.
[255] S. Spaepen, J. Vanderleyden, and R. Remans, "Indole-3-acetic acid in microbial
and microorganism-plant signaling," FEMS Microbiol Reviews, vol. 31, pp.
425-48, Jul 2007.
[256] C. Speier, I. Vessey, and J. S. Valacich, "The effects of interruptions, task
complexity, and information presentation on computer‐supported decision‐making performance," Decision Sciences, vol. 34, pp. 771-797, 2003.
[257] M. Tahir, M. S. Mirza, A. Zaheer, M. R. Dimitrov, H. Smidt, and S. Hameed,
"Isolation and identification of phosphate solubilizer 'Azospirillum, Bacillus'
and 'Enterobacter' strains by 16SrRNA sequence analysis and their effect on
growth of wheat (Triticum aestivum L.)," Australian Journal of Crop Science,
vol. 7, p. 1284, 2013.
[258] J.-S. Jeon, S.-S. Lee, H.-Y. Kim, T.-S. Ahn, and H.-G. Song, "Plant growth
promotion in soil by some inoculated microorganisms," Journal Of
Microbiology Seoul , vol. 41, pp. 271-276, 2003.
[259] S. Banerjee, R. Palit, C. Sengupta, and D. Standing, "Stress induced phosphate
solubilization by Arthrobacter sp. and Bacillus sp. isolated from tomato
rhizosphere," Australian Journal of Crop Science, vol. 4, pp. 378-383, 2010.
[260] N. Gupta, D. Sahoo, and U. C. Basak, "Evaluation of in vitro solubilization
potential of phosphate solubilising Streptomyces isolated from phyllosphere of
Heritiera fomes (mangrove)," African Journal of Microbiology Research, vol.
4, pp. 136-142, Feb 4 2010.
[261] H. Rodrı́guez and R. Fraga, "Phosphate solubilizing bacteria and their role in
plant growth promotion," Biotechnology advances, vol. 17, pp. 319-339, 1999.
[262] G. Archana, A. Buch, and G. N. Kumar, "Pivotal role of organic acid secretion
by rhizobacteria in plant growth promotion," In Microorganisms in Sustainable
Agriculture and Biotechnology, ed: Springer, 2012, pp. 35-53.

5. References
169
[263] D. Mukherjee, J. Das, G. M. Sarkar, and S. C. Lahiri, "Studies on phosphate
solubilization by microbes and enrichment of soils with phosphates," Journal
of the Indian Chemical Society, vol. 72, pp. 767-770, Oct 1995.
[264] J.-M. Barea and A. E. Richardson, "Phosphate mobilisation by soil
microorganisms," In Principles of Plant-Microbe Interactions, ed: Springer,
2015, pp. 225-234.
[265] A. H. Goldstein, "Recent progress in understanding the molecular-genetics and
biochemistry of calcium-phosphate solubilization by gram-negative bacteria,"
Biological Agriculture & Horticulture, vol. 12, pp. 185-193, 1995.
[266] M. d. V. B. Figueiredo, L. Seldin, F. F. de Araujo, and R. d. L. R. Mariano,
"Plant growth promoting rhizobacteria: Fundamentals and applications," In
Plant growth and health promoting bacteria, ed: Springer, 2011, pp. 21-43.
[267] D. Paul and H. Lade, "Plant-growth-promoting rhizobacteria to improve crop
growth in saline soils: a review," Agronomy for Sustainable Development, vol.
34, pp. 737-752, 2014.
[268] H. C. Kaymak, I. Guvenc, F. Yarali, and M. F. Donmez, "The Effects of bio-
priming with PGPR on germination of radish (Raphanus sativus L.) seeds under
saline conditions," Turkish Journal of Agriculture and Forestry, vol. 33, pp.
173-179, 2009.
[269] A. Richardson, J. Lynch, P. Ryan, E. Delhaize, F. Smith, S. Smith, et al., "Plant
and microbial strategies to improve the phosphorus efficiency of agriculture,"
Plant and Soil, vol. 349, pp. 121-156, 2011.
[270] I. Cacciari and D. Lippi, "Arthrobacters: successful arid soil bacteria: a review,"
Arid Land Research and Management, vol. 1, pp. 1-30, 1987.
[271] M. Siddikee, P. Chauhan, R. Anandham, G.-H. Han, and T. Sa, "Isolation,
characterization, and use for plant growth promotion under salt stress, of ACC
deaminase-producing halotolerant bacteria derived from coastal soil," Journal
of Microbiology & Biotechnology, vol. 20, pp. 1577-1584, 2010.
[272] F. Cassán, J. Vanderleyden, and S. Spaepen, "Physiological and agronomical
aspects of phytohormone production by model plant-growth-promoting
rhizobacteria (PGPR) belonging to the genus Azospirillum," Journal of Plant
Growth Regulation, pp. 1-20, 2013.
[273] Y. Bashan, "Azospirillum-plant relationships: environmental and physiological
advances (1990-1996)," Canadian Journal of Microbiology, vol. 43, pp. 103-
121, 1997.

5. References
170
[274] M. Fayez and Z. Daw, "Effect of inoculation with different strains of
Azospirillum brasilense on cotton (Gossipium barbadense)," Biology and
Fertility of Soils, vol. 4, pp. 91-95, 1987.
[275] A. Pandey, E. Sharma, and L. M. S. Palni, "Influence of bacterial inoculation
on maize in upland farming systems of the Sikkim Himalaya," Soil Biology and
Biochemistry, vol. 30, pp. 379-384, 1998.
[276] G. Berg, "Plant-microbe interactions promoting plant growth and health:
perspectives for controlled use of microorganisms in agriculture," Applied
Microbiology & Biotechnology, vol. 84, pp. 11-8, Aug 2009.
[277] R. M. Lehman, C. A. Cambardella, D. E. Stott, V. Acosta-Martinez, D. K.
Manter, J. S. Buyer, et al., "Understanding and enhancing soil biological health:
the solution for reversing soil degradation," Sustainability, vol. 7, pp. 988-1027,
2015.
[278] K. Kavitha, S. Mathiyazhagan, V. Senthilvel, S. Nakkeeran, and G.
Chandrasekar, "Development of bioformulations of antagonistic bacteria for the
management of damping off of Chilli (Capsicum annuum L)," Archives Of
Phytopathology and Plant Protection, vol. 38, pp. 19-30, 2005.
[279] V. Kumar and R. Gera, "Isolation of a multi-trait plant growth promoting
Brevundimonas sp. and its effect on the growth of Bt-cotton," 3 Biotech, vol. 4,
pp. 97-101, 2014.
[280] D. Thuler, E. Floh, W. Handro, and H. Barbosa, "Plant growth regulators and
amino acids released by Azospirillum sp. in chemically defined media," Letters
in Applied Microbiology, vol. 37, pp. 174-178, 2003.
[281] T. d. l. M. Orberá, M. d. J. Serrat, and E. Ortega, "Potential applications of
Bacillus subtilis strain SR/B-16 for the control of phytopathogenic fungi in
economically relevant crops," Biotecnología Aplicada, vol. 31, pp. 13-17, 2014.
[282] S. Nápoles Vinent, M. Serrat Díaz, E. Ortega Delgado, H. Barbosa, and T.
OrberáRatón, "Effects of Brevibacillus borstelensis B65 on germination and
seedlings development of horticulture crops," Cultivos Tropicales, vol. 35, pp.
17-23, 2014.
[283] A. G. Vovides, Y. Bashan, J. A. Lopez-Portillo, and R. Guevara, "Nitrogen
fixation in preserved, reforested, naturally regenerated and impaired mangroves
as an indicator of functional restoration in mangroves in an arid region of
Mexico," Restoration Ecology, vol. 19, pp. 236-244, Mar 2011.
[284] L. Yao, Z. Wu, Y. Zheng, I. Kaleem, and C. Li, "Growth promotion and
protection against salt stress by Pseudomonas putida Rs-198 on cotton,"
European Journal of Soil Biology, vol. 46, pp. 49-54, 2010.

5. References
171
[285] M. Lucy, E. Reed, and B. Glick, "Applications of free living plant growth-
promoting rhizobacteria," Antonie van Leeuwenhoek, vol. 86, pp. 1-26, 2004.
[286] D. Sachdev, V. Agarwal, P. Verma, Y. Shouche, P. Dhakephalkar, and B.
Chopade, "Assessment of microbial biota associated with rhizosphere of wheat
(Triticum aestivum L.) during flowering stage and their plant growth promoting
traits," International Journal of Microbiology, vol. 61, pp. 154-168, 2009.
[287] C. Dimkpa, T. Weinand, and F. Asch, "Plant rhizobacteria interactions alleviate
abiotic stress conditions," Plant, Cell & Environment, vol. 32, pp. 1682-1694,
2009.
[288] G. Forchetti, O. Masciarelli, S. Alemano, D. Alvarez, and G. Abdala,
"Endophytic bacteria in sunflower (Helianthus annuus L.): isolation,
characterization, and production of jasmonates and abscisic acid in culture
medium," Applied Microbiology and Biotechnology, vol. 76, pp. 1145-1152,
2007.
[289] L. Arruda, A. Beneduzi, A. Martins, B. Lisboa, C. Lopes, F. Bertolo, et al.,
"Screening of rhizobacteria isolated from maize (Zea mays L.) in Rio Grande
do Sul State (South Brazil) and analysis of their potential to improve plant
growth," Applied Soil Ecology, vol. 63, pp. 15-22, 2013.
[290] O. Baris, F. Sahin, M. Turan, F. Orhan, and M. Gulluce, "Use of plant-growth-
promoting rhizobacteria (PGPR) seed inoculation as alternative fertilizer inputs
in wheat and barley production," Communications in Soil Science and Plant
Analysis, vol. 45, pp. 2457-2467, 2014.
[291] F. Pérez-Montaño, C. Alías-Villegas, R. Bellogín, P. Del Cerro, M. Espuny, I.
Jiménez-Guerrero, et al., "Plant growth promotion in cereal and leguminous
agricultural important plants: From microorganism capacities to crop
production," Microbiological Research, vol. 169, pp. 325-336, 2014.
[292] E. A. R. Cáceres, "Improved medium for isolation of Azospirillum spp," Applied
and Environmental Microbiology, vol. 44, p. 990, 1982.
[293] H. Marquez-Santacruz, R. Hernandez-Leon, M. Orozco-Mosqueda, I.
Velazquez-Sepulveda, and G. Santoyo, "Diversity of bacterial endophytes in
roots of Mexican husk tomato plants (Physalisixocarpa) and their detection in
the rhizosphere," Genetics and Molecular Research, vol. 9, pp. 2372-2380,
2010.
[294] R. P. Ryan, K. Germaine, A. Franks, D. J. Ryan, and D. N. Dowling, "Bacterial
endophytes: recent developments and applications," FEMS Microbiology
Letters, vol. 278, pp. 1-9, 2008.

5. References
172
[295] D. Egamberdieva, "Plant growth promoting properties of rhizobacteria isolated
from wheat and pea grown in loamy sand soil," Turkish Journal of Biology, vol.
32, pp. 9-15, 2008.
[296] B. Lugtenberg, "Life of Microbes in the Rhizosphere," In Principles of Plant-
Microbe Interactions, ed: Springer International Publishing, 2015, pp. 7-15.
[297] I. T. Paulsen, C. M. Press, J. Ravel, D. Y. Kobayashi, G. S. Myers, D. V.
Mavrodi, et al., "Complete genome sequence of the plant commensal
Pseudomonas fluorescens Pf-5," Nature Biotechnology, vol. 23, pp. 873-878,
2005.
[298] J. M. Whipps, "Microbial interactions and biocontrol in the rhizosphere,"
Journal of Experimental Botany, vol. 52, pp. 487-511, 2001.
[299] U. Mina, S. Khan, A. Choudhary, R. Choudhary, and P. Aggarwal, "An
approach for impact assessment of transgenic plants on soin
ecosystem," Applied Ecology and Environmental Research, vol. 6(3), pp. 1-19,
2007.
[300] M. Castaldini, A. Turrini, C. Sbrana, A. Benedetti, M. Marchionni, S. Mocali,
et al., "Impact of Bt corn on rhizospheric and soil eubacterial communities and
on beneficial mycorrhizal symbiosis in experimental microcosms," Applied and
Environmental Microbiology, vol. 71, pp. 6719-6729, 2005.
[301] S. Gyamfi, U. Pfeifer, M. Stierschneider, and A. Sessitsch, "Effects of
transgenic glufosinate-tolerant oilseed rape (Brassica napus) and the associated
herbicide application on eubacterial and Pseudomonas communities in the
rhizosphere," FEMS Microbiology Ecology, vol. 41, pp. 181-190, 2002.
[302] P. Nannipieri, J. Ascher, M. Ceccherini, L. Landi, G. Pietramellara, G. Renella,
et al., "Effects of root exudates in microbial diversity and activity in rhizosphere
soils," Molecular Mechanisms of Plant and Microbe Coexistence, vol. 2, pp.
339-365, 2008.
[303] W. Song, L. N. Yuan, L. Xiao, Z. Zhan, L. Y. Yang, and L. J. Jiang, "ALPase
activity and the distribution of phosphate solubilizing bacteria and the
relationship between them in sediments of Lake Taihu," Huan Jing Ke Xue, vol.
28, pp. 2355-60, Oct 2007.
[304] A. Kahnert, P. Mirleau, R. Wait, and M. A. Kertesz, "The LysR-type regulator
SftR is involved in soil survival and sulphate ester metabolism in Pseudomonas
putida," Environmental Microbiology, vol. 4, pp. 225-37, 2002.
[305] F. I. Parra-Cota, J. J. Peña-Cabriales, S. de los Santos-Villalobos, N. A.
Martínez-Gallardo, and J. P. Délano-Frier, "Burkholderia ambifaria and B.
caribensis Promote Growth and Increase Yield in Grain Amaranth (Amaranthus

5. References
173
cruentus and A. hypochondriacus) by Improving Plant Nitrogen Uptake," PLOS
One, vol. 9, p. e88094, 2014.
[306] Q. Wang, M. He, and Y. Wang, "Influence of combined pollution of antimony
and arsenic on culturable soil microbial populations and enzyme activities,"
Ecotoxicology, vol. 20, pp. 9-19, Jan 2011.
[307] D. V. Badri and J. M. Vivanco, "Regulation and function of root exudates,"
Plant, Cell and Environment, vol. 32, pp. 666-81, 2009.
[308] K. Jaeger, B. Dijkstra, and M. Reetz, "Bacterial biocatalysts: molecular biology,
three-dimensional structures, and biotechnological applications of lipases,"
Annual Reviews in Microbiology, vol. 53, pp. 315-351, 1999.
[309] D. Sachdev, H. Chaudhari, V. Kasture, D. Dhavale, and B. Chopade, "Isolation
and characterization of indole acetic acid (IAA) producing Klebsiella
pneumoniae strains from rhizosphere of wheat (Triticum aestivum) and their
effect on plant growth," Indian Journal of Experimental Biology, vol. 47, pp.
993-1993, 2009.
[310] M. M. Collavino, H. J. Tripp, I. E. Frank, M. L. Vidoz, P. A. Calderoli, M.
Donato, et al., "nifH pyrosequencing reveals the potential for location‐specific
soil chemistry to influence N2‐fixing community dynamics," Environmental
Microbiology, vol. 16, pp. 3211-3223, 2014.
[311] L. I. Falcon, E. Escobar-Briones, and D. Romero, "Nitrogen fixation patterns
displayed by cyanobacterial consortia in Alchichica crater-lake, Mexico,"
Hydrobiologia, vol. 467, pp. 71-78, Jan 2002.
[312] X. Meng, L. Wang, X. Long, Z. Liu, Z. Zhang, and R. Zed, "Influence of
nitrogen fertilization on diazotrophic communities in the rhizosphere of the
Jerusalem artichoke (Helianthus tuberosus L.)," Research in Microbiology, vol.
163, pp. 349-356, 2012.
[313] F. Widmer, B. Shaffer, L. Porteous, and R. Seidler, "Analysis of nifH gene pool
complexity in soil and litter at a Douglas fir forest site in the Oregon Cascade
Mountain Range," Applied and Environmental Microbiology, vol. 65, pp. 374-
380, 1999.
[314] N. Thaweenut, Y. Hachisuka, S. Ando, S. Yanagisawa, and T. Yoneyama, "Two
seasons' study on nifH gene expression and nitrogen fixation by diazotrophic
endophytes in sugarcane (Saccharum spp. hybrids): expression of nifH genes
similar to those of rhizobia," Plant and Soil, vol. 338, pp. 435-449, 2011.
[315] A. Kennedy, "Bacterial diversity in agroecosystems," Agriculture, Ecosystems
& Environment, vol. 74, pp.65-76. 1999.

5. References
174
[316] I. R. Kennedy, A. Choudhury, and M. L. Kecskés, "Non-symbiotic bacterial
diazotrophs in crop-farming systems: can their potential for plant growth
promotion be better exploited?," Soil Biology and Biochemistry, vol. 36, pp.
1229-1244, 2004.
[317] B. Reinhold-Hurek, T. Hurek, M. Gillis, B. Hoste, M. Vancanneyt, K. Kersters,
et al., "Azoarcus gen. nov., Nitrogen-Fixing Proteobacteria Associated with
Roots of Kallar Grass (Leptochloa fusca L.) Kunth), and Description of Two
Species, Azoarcus indigens sp. nov. and Azoarcus communis sp. nov,"
International Journal of Systematic Bacteriology, vol. 43, pp. 574-584, 1993.
[318] P. Zhang, J. Zheng, G. Pan, X. Zhang, L. Li, and R. Tippkotter, "Changes in
microbial community structure and function within particle size fractions of a
paddy soil under different long-term fertilization treatments from the Tai Lake
region, China," Colloids and Surface B: Biointerfaces, vol. 58, pp. 264-70,
2007.
[319] G. Y. Tan and W. K. Tan, "Interaction between Alfalfa Cultivars and Rhizobium
Strains for Nitrogen-Fixation," Theoretical and Applied Genetics, vol. 71, pp.
724-729, 1986.
[320] T. Coba de la Peña, F. J. Redondo, E. Manrique, M. M. Lucas, and J. J. Pueyo,
"Nitrogen fixation persists under conditions of salt stress in transgenic
Medicago truncatula plants expressing a cyanobacterial flavodoxin," Plant
Biotechnology Journal, vol. 8, pp. 954-965, 2010.
[321] V. Bhandari and R. S. Gupta, "Molecular signatures for the phylum (class)
Thermotogae and a proposal for its division into three orders (Thermotogales,
Kosmotogales ord. nov. and Petrotogales ord. nov.) containing four families
(Thermotogaceae, Fervidobacteriaceae fam. nov., Kosmotogaceae fam. nov.
and Petrotogaceae fam. nov.) and a new genus Pseudothermotoga gen. nov.
with five new combinations," Antonie van Leeuwenhoek, vol. 105, pp. 143-168,
2014.
[322] M. S. Strickland, C. Lauber, N. Fierer, and M. A. Bradford, "Testing the
functional significance of microbial community composition," Ecology, vol. 90,
pp. 441-51, Feb 2009.
[323] W. J. Xie, J. M. Zhou, and H. Y. Wang, "Influence of Cu2+, Cd2+ and
cypermethrin on microbial functional diversity in different fertilization soils,"
Huan Jing Ke Xue, vol. 29, pp. 2919-24, 2008.
[324] X. Li, J. Rui, Y. Mao, A. Yannarell, and R. Mackie, "Dynamics of the bacterial
community structure in the rhizosphere of a maize cultivar," Soil Biology and
Biochemistry, vol. 68, pp. 392-401, 2014.

5. References
175
[325] A. García‐Salamanca, M. A. Molina‐Henares, P. Dillewijn, J. Solano, P.
Pizarro‐Tobías, A. Roca, et al., "Bacterial diversity in the rhizosphere of maize
and the surrounding carbonate‐rich bulk soil," Microbial Biotechnology, vol. 6,
pp. 36-44, 2013.
[326] D. Perrig, M. L. Boiero, O. A. Masciarelli, C. Penna, O. A. Ruiz, F. D. Cassan,
et al., "Plant-growth-promoting compounds produced by two agronomically
important strains of Azospirillum brasilense, and implications for inoculant
formulation," Applied Microbiology & Biotechnology, vol. 75, pp. 1143-50,
2007.
[327] H. Antoun, C. J. Beauchamp, N. Goussard, R. Chabot, and R. Lalande,
"Potential of Rhizobium and Bradyrhizobium species as plant growth promoting
rhizobacteria on non-legumes: Effect on radishes (Raphanus sativus L.)," Plant
and Soil, vol. 204, pp. 57-67, 1998.
[328] M. K. Chelius and E. W. Triplett, "The diversity of archaea and bacteria in
association with the roots of Zea mays L," Microbial Ecology, vol. 41, pp. 252-
515, 2001.
[329] D. Fischer, M. Uksa, W. Tischler, T. Kautz, U. Köpke, and M. Schloter,
"Abundance of ammonia oxidizing microbes and denitrifiers in different soil
horizons of an agricultural soil in relation to the cultivated crops," Biology and
Fertility of Soils, pp. 1-4, 2013.
[330] S. Leininger, T. Urich, M. Schloter, L. Schwark, J. Qi, G. Nicol, et al., "Archaea
predominate among ammonia-oxidizing prokaryotes in soils," Nature, vol. 442,
pp. 806-809, 2006.
[331] M. Sakai, A. Hosoda, K. Ogura, and M. Ikenaga, "The growth of Steroidobacter
agariperforans sp. nov., a novel agar-degrading bacterium isolated from soil, is
enhanced by the diffusible metabolites produced by bacteria belonging to
rhizobiales," Microbes and Environments, vol. 29, pp. 89-95, 2014.
[332] C. Suarez, S. Ratering, I. Kramer, and S. Schnell, "Cellvibrio diazotrophicus sp.
nov., a nitrogen-fixing bacteria isolated from the rhizosphere of salt meadow
plants and emended description of the genus Cellvibrio," International Journal
of Systematic and Evolutionary Microbiology, vol. 64, pp. 481-486, 2014.
[333] p. Christensen and F. Cook, "Lysobacter, a new genus of nonfruiting, gliding
bacteria with a high base ratio," International Journal of Systematic
Bacteriology, vol. 28, pp. 367-393, 1978.
[334] T. S. Sullivan, M. B. McBride, and J. E. Thies, "Soil bacterial and archaeal
community composition reflects high spatial heterogeneity of pH, bioavailable
Zn, and Cu in a metalliferous peat soil," Soil Biology and Biochemistry, vol. 66,
pp. 102-109, 2013.

5. References
176
[335] A. C. Hayward, N. Fegan, M. Fegan, and G. Stirling, "Stenotrophomonas and
Lysobacter: ubiquitous plant‐associated gamma‐proteobacteria of developing
significance in applied microbiology," Journal of Applied Microbiology, vol.
108, pp. 756-770, 2010.
[336] R. Mendes, M. Kruijt, I. de Bruijn, E. Dekkers, M. van der Voort, J. H.
Schneider, et al., "Deciphering the rhizosphere microbiome for disease-
suppressive bacteria," Science, vol. 332, pp. 1097-1100, 2011.
[337] M. Davey, G. Holmes, and R. Johnstone, "High rates of nitrogen fixation
(acetylene reduction) on coral skeletons following bleaching mortality," Coral
Reefs, vol. 27, pp. 227-236, 2008.
[338] M. A. Furlong, D. R. Singleton, D. C. Coleman, and W. B. Whitman,
"Molecular and culture-based analyses of prokaryotic communities from an
agricultural soil and the burrows and casts of the earthworm Lumbricus
rubellus," Applied and Environmental Microbiology, vol. 68, pp. 1265-1279,
2002.
[339] O. Martínez-Viveros, M. Jorquera, D. Crowley, G. Gajardo, and M. Mora,
"Mechanisms and practical considerations involved in plant growth promotion
by rhizobacteria," Journal of Soil Science and Plant Nutrition, vol. 10, pp. 293-
319, 2010.
[340] M. K. Lily, A. Bahuguna, K. K. Bhatt, and K. Dangwal, "Degradation of
Anthracene by a novel strain Brachybacterium paraconglomeratum BMIT637C
(MTCC 9445)," International Journal of Environmental Sciences, vol. 3, pp.
1242-1252, 2013.
[341] X. Wang, Z. Zhang, D. Jin, L. Zhou, L. Wu, C. Li, et al., "Draft genome
sequence of Brachybacterium phenoliresistens strain W13A50, a halotolerant
hydrocarbon-degrading bacterium," Genome announcements, vol. 2, pp.
e00899-14, 2014.
[342] S. Saif, M. S. Khan, A. Zaidi, and E. Ahmad, "Role of Phosphate-Solubilizing
Actinomycetes in Plant Growth Promotion: Current Perspective," In Phosphate
Solubilizing Microorganisms, ed: Springer, 2014, pp. 137-156.
[343] J.-H. Yoon, C.-H. Lee, and T.-K. Oh, "Aeromicrobium alkaliterrae sp. nov.,
isolated from an alkaline soil, and emended description of the genus
Aeromicrobium," International journal of systematic and evolutionary
microbiology, vol. 55, pp. 2171-2175, 2005.
[344] S. Williams, F. Davies, C. Mayfield, and M. Khan, "Studies on the ecology of
actinomycetes in soil II. The pH requirements of streptomycetes from two acid
soils," Soil Biology and Biochemistry, vol. 3, pp. 187-195, 1971.

5. References
177
[345] J. Mergaert, "Thermomonas fusca sp. nov. and Thermomonas brevis sp. nov.,
two mesophilic species isolated from a denitrification reactor with poly
(caprolactone) plastic granules as fixed bed, and emended description of the
genus Thermomonas," International Journal of Systematic and Evolutionary
Microbiology, vol. 53, 2003.
[346] V. Radl, J. L. Simões-Araújo, J. Leite, S. R. Passos, L. M. V. Martins, G. R.
Xavier, et al., "Microvirga vignae sp. nov., a root nodule symbiotic bacterium
isolated from cowpea grown in semi-arid Brazil," International journal of
systematic and evolutionary microbiology, vol. 64, pp. 725-730, 2014.
[347] J. E. Zhang, A. X. Gao, H. Q. Xu, and M. Z. Luo, "Effects or maize/peanut
intercropping on rhizosphere soil microbes and nutrient contents," Ying Yong
Sheng Tai Xue Bao, vol. 20, pp. 1597-602, Jul 2009.
[348] Y. P. Zhang, E. L. Pohlmann, P. W. Ludden, and G. P. Roberts, "Regulation of
nitrogen fixation by multiple P-II homologs in the photosynthetic bacterium
Rhodospirillum rubrum," Symbiosis, vol. 35, pp. 85-100, 2003.
[349] X. Xu, S. Xia, L. Zhou, Z. Zhang, and B. E. Rittmann, "Bioreduction of
vanadium (V) in groundwater by autohydrogentrophic bacteria: Mechanisms
and microorganisms," Journal of Environmental Sciences, vol. 30 pp. 122-128,
2015.
[350] H. An, L. Zhang, Y. Tang, X. Luo, T. Sun, Y. Li, et al., "Skermanella
xinjiangensis sp. nov., isolated from the desert of Xinjiang, China,"
International Journal of Systematic and Evolutionary Microbiology, vol. 59, pp.
1531-1534, 2009.
[351] C. Zhao, Y. Deng, X. Wang, Q. Li, Y. Huang, and B. Liu, "Identification and
characterization of an anaerobic ethanol-producing cellulolytic bacterial
consortium in Great Basin hot spring with agricultural residues and energy
crops," Journal of Microbiology and Biotechnology, vol. 32, pp 55-77, 2014.
[352] G. Rückert, "Myxobacteria (Myxobacteriales) on leaf surfaces," Zeitschrift fur
Allgemeine Mikrobiologie, vol. 21, pp. 761-763, 1980.
[353] R. Sungthong and N. Nakaew, "The genus Nonomuraea: A review of a rare
actinomycete taxon for novel metabolites," Journal of Basic Microbiology,vol.
55, pp. 554-565, 2014.
[354] E. Topp, W. M. Mulbry, H. Zhu, S. M. Nour, and D. Cuppels, "Characterization
of s-triazine herbicide metabolism by a Nocardioides sp. isolated from
agricultural soils," Applied and Environmental Microbiology, vol. 66, pp. 3134-
3141, 2000.

5. References
178
[355] R. Fudou, Y. Jojima, T. Iizuka, and S. Yamanaka, "Haliangium ochraceum gen.
nov., sp. nov. and Haliangium tepidum sp. nov.: novel moderately halophilic
myxobacteria isolated from coastal saline environments," The Journal of
General and Applied Microbiology, vol. 48, pp. 109-116, 2002.
[356] B. Liu, G.-H. Liu, C. Sengonca, P. Schumann, J.-Y. Tang, M.-C. Chen, et al.,
"Bacillus wuyishanensis sp. nov., isolated from rhizosphere soil of a medical
plant, Prunella vulgaris, in Wuyi mountain of China," International Journal of
Systematic and Evolutionary Microbiology, 65(7), 2030-2035. 2014.
[357] I. S. Kulichevskaya, N. E. Suzina, W. Liesack, and S. N. Dedysh, "Bryobacter
aggregatus gen. nov., sp. nov., a peat-inhabiting, aerobic chemo-organotroph
from subdivision 3 of the Acidobacteria," International Journal of Systematic
and Evolutionary Microbiology, vol. 60, pp. 301-306, 2010.
[358] C. Muangchinda, S. Chavanich, V. Viyakarn, K. Watanabe, S. Imura, A.
Vangnai, et al., "Abundance and diversity of functional genes involved in the
degradation of aromatic hydrocarbons in Antarctic soils and sediments around
Syowa Station," Environmental Science and Pollution Research, pp. 1-11,
2014.
[359] K. Zengler, G. Toledo, M. Rappé, J. Elkins, E. J. Mathur, J. M. Short, et al.,
"Cultivating the uncultured," Proceedings of the National Academy of Sciences,
vol. 99, pp. 15681-15686, 2002.
[360] Ö. İnceoğlu, W. A. Al-Soud, J. F. Salles, A. V. Semenov, and J. D. van Elsas,
"Comparative analysis of bacterial communities in a potato field as determined
by pyrosequencing," PLOS One, vol. 6, p. e23321, 2011.
[361] C. Yin, S. Hulbert, K. Schroeder, O. Mavrodi, D. Mavrodi, A. Dhingra, et al.,
"Comparison of bacterial communities from inside and outside of Rhizoctonia
bare patches in wheat," Phytopathology, vol. 100, p. S142, 2010.
[362] J. Barriuso and R. P. Mellado, "Glyphosate affects the rhizobacterial
communities in glyphosate-tolerant cotton," Applied Soil Ecology, vol. 55, pp.
20-26, 2012.
[363] E. Somers, D. Ptacek, P. Gysegom, M. Srinivasan, and J. Vanderleyden,
"Azospirillum brasilense produces the auxin-like phenylacetic acid by using the
key enzyme for indole-3-acetic acid biosynthesis," Applied and environmental
microbiology, vol. 71, pp. 1803-1810, 2005.
[364] K. D. Burkhead, P. J. Slininger, and D. A. Schisler, "Biological control
bacterium Enterobacter cloacae S11: T: 07 (NRRL B-21050) produces the
antifungal compound phenylacetic acid in Sabouraud maltose broth culture,"
Soil Biology and Biochemistry, vol. 30, pp. 665-667, 1998.

5. References
179
[365] B. K. Hwang, S. W. Lim, B. S. Kim, J. Y. Lee, and S. S. Moon, "Isolation and
in vivo and in vitro antifungal activity of phenylacetic acid and sodium
phenylacetate from Streptomyces humidus," Applied and environmental
microbiology, vol. 67, pp. 3739-3745, 2001.
[366] Y. Kim, J.-Y. Cho, J.-H. Kuk, J.-H. Moon, J.-I. Cho, Y.-C. Kim, et al.,
"Identification and antimicrobial activity of phenylacetic acid produced by
Bacillus licheniformis isolated from fermented soybean," Current
Microbiology, vol. 48, pp. 312-317, 2004.
[367] I. Arnon, Agriculture in dry lands: in Principles and Practice ed: Elsevier, 2012,
vol. 26.
[368] S. Ayaz and S. Zeevar, "Balochistan Partnerships for Sustainable Development
Pakistan, IUCN " Climate Change and Coastal Districts of Balochistan-
Situation Analysis, Implications and Recommendations, 2012.
[369] P. Ahmad, M. Ashraf, M. Younis, X. Hu, A. Kumar, N. A. Akram, et al., "Role
of transgenic plants in agriculture and biopharming," Biotechnology Advances,
vol. 30, pp. 524-540, 2012.





























