Embed Size (px)
Citation preview

Vol. 132, No. 2, 1985
October 30, 1985
BIOCHEMICAL AN0 BIOPHYSICAL RESEARCH COMMUNICATIONS
Pages 598-604
RAT LIVER HMG COA REDUCTASE, A GLYCOPROTEIN OF THE ENDOPLASMIC RETICULUM, IS IN EQUILIBRIUM
BETWEEN MONOMERIC AND DIMERIC FORMS
Diego Haro, Pedro F. Marrero, and Faust0 G. Hegardt
Department of Biochemistry, University of Barcelona School of Pharmacy, Barcelona 08028 Spain
Received September 3, 1985
Incubation of rat hepatocytes with [35 I S methionine in pulse and pulse- chase experiments followed by immunoprecipitation of the HMG CoA reductase and SDS-PAGE results in two labelled polypeptides of 104 and 180 Kdaltons. These two polypeptides have half lives of 80 and 46 minutes respectively When hepatocytes are incubated with mevalonolactone, and a pulse of [ 3%jme- thionine is given, the rate of synthesis of bot the 180 and 104 Kd peptides is strongly diminished. After treatment of the 2 1 35S labelled immunoprecipi- tates with endoglycosidase H, the 180 Kd reductase splits into two labelled peptides of 110 and 97 Kd. We suggest that in addition to the 104 Kd reducta se, the endoplasmic reticulum contains the dimer of two reductases linked by a carbohydrate chain. The equilibrium monomer-dimer probably regulates the rate of degradation of reductase. @ 1985 Academic Press, Inc.
3-Hydroxy 3 methylglutaryl CoA reductase (EC 1.1.1.34) the major reguk-
tory enzyme in the cholesterol biosynthetic patway is also responsible for
the biosynthesis of isoprenoid compounds such as isopentenyl tRNA, ubiquinone
and dolichol (1). Hence, the efficient regulation of HMG CoA reductase is very
important for a wide range of cellular functions.
HMG CoA reductase is a transmembrane protein that resides in the endo-
plasmic reticulum of the animal cell. Recently, the nucleotide sequence of a
cloned cDNA for hamster HMG CoA reductase has been established and the amino
acid sequence of this enzyme was deduced (2). This sequence showed that the
NH2 terminal of one third of total reductase is hydrophobic and contains 7
membrane spanning regions. The COOH terminal domain contains the catalytycally
active centre and projects into the cytoplasm (3).
This model involves that the physiological state of reductase in the
E.R. is only formed by a monomer of 97 Kd. In this paper we present evidence
Abbreviations used : PMSF, phenyl methyl sulphonyl fluoride; BSA, bovine serum albumin; TEMED, N,N,N',N'-tetramethylethylenediamine; Endo H, endo-D-N- acetylglucosaminidase H; SDS, sodium dodecyl sulphate; DTT, dithiothreitol; PAGE, polyacrylamide gel electrophoresis; HMG CoA reductase or reductase, 3- Hydroxy 3 methylglutaryl CoA reductase; DMSO, dimethylsulfoxide; PPO, 2,5-di- phenyloxazole.
0006-291X/85 $1.50 Copyright 0 1985 by Academic Press, Inc. All rights of reproduction in any form reserved. 598

Vol. 132, No. 2, 1985 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
that in rat hepatocytes, the monomeric form of reductase is in equilibrium
with a dimeric form also bound to the endoplasmic reticulum. The two monomers
are covalently linked by a carbohidrate chain composed by N-acetyl glucosamire
residues. The evidence of the dimer of 180 Kd comes from experiments involving
incorporation of c35 3 S methionine to the nascent molecule of reductase and imnu - noprecipitation with antireductase antibodies. When the dimeric form is incuba - ted with endoglycosidase H, the immunoprecipitable polypeptide of 180 Kd di-
sappears, producing two Mr bands of 110 and 97 Kd. These bands could represew
the reductase linked to carbohydrate chains of different length.
Since the monomeric and dimeric forms have different half lives (80 and
46 min respectively), we conclude that the dimerization- dedimerization reac-
tions may control the degradation of reductase and the cholesterol biosynthe-
sis as well.
MATERIALS AND METHODS
Chemicals were obtained from the following sources: [35S]methionine (1100 Ci/mmol) from Amersham; formalin fixed Staphylococcus aureus (pansorbir$ from Calbiochem; leupeptin, aprotinin, antipain, PMSF, BSA(A-7906) and Endo H from Sigma; collagenase from Cooper Biomedicals Ltd; SDS, acrylamide, bisacry lamide, TEMED and ammonium persulfate from Bio-Rad. All other chemicals were- of the highest quality from commercial sources.
Cells. Hepatocytes were prepared after six hours darkness, from male Wistar rats fed at libitum and housed under 12 h light-dark cycles, basically as described by Seglen (4). Liver perfusion was initiated with Hank's Ca'+free 5 mM glucose buffer. After perfusion with 350 ml, collagenase was added to a final concentration of 0.5 mg/ml and BSA to a final concentration of 15 mg/ml. The recirculating perfusion with collagenase was carried out at 60 ml/min for 12-15 min. The liver was then removed and gently dispersed in Hank's Ca++&ee buffer with 15 mq/ml BSA and 5 mM glucose. The resulting cell suspension was washed three times and finally resuspended in Hank's buffer supplemented with amino acids and cofactors as described by Eagle (51, without methionine, plus 5 mM glucose, 1.5% BSA, and 20 mM Tricine pH 7.4. The cells were counted in a Neubauer haemocytometer. Immediately after isolation, 90% of the hepatocytes were viable as estimated by tripan blue exclusion test. The apparent viabili- ty of the cells incubated in suspension remained excellent for several hours. Cells (5 x 106) were incubated in a final volume of 1.5 ml of medium supple- mented with [35S]methionine (100 PCi; 1100 Ci/mmol).
HMG CoA reductase antibodies. Antibodies were raised in rabbits against purified enzyme (52 Kd) from rats feed 5% cholestiramine. The antibody and controly globulins were purified by three succesive ammonium sulfate precipi - tations at 40% saturation.
Immunoprecipitation of radiolabelled HMG CoA reductase. Radiolabelled hepatocytes with ["Slmethionine were pelleted and solubilized in 1 ml of buffer S (1% triton x-100, 0.5% deoxicholate, 0.1% SDS, 5 mM EGTA, 5 mM EDTA, 0.1 M NaCl, 100 )&I leupeptin, IO nM antipain, 15 pq/ml aprotinin, 2 mM PMSF, 0.01 phosphate buffer pH 7.5) at 4 OC. After centrifuging at 100.000 xq for 60 min, one ml of the supernatant was incubated for two hours at 4 OC with 7 globulin (6 ,ul for 106 cells) and then 5 pl 10% pansorbin was added for each 1 pl Yglobulin (30 min at 4 OC). After centrifuging the immune complexg were washed three times with buffer S, incubated for 45 min at room temperatu re and dispersed in 75 pl of buffer B (15% SDS, 10% Mercaptoethanol, in 62.5- mM Tris pH 6.8). After centrifuging at 12,000 xq for 5 min the supernatant was separated from pansorbin.

Vol. 132, No. 2. 1985 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Treatment with endo-D-N-acetylglucosaminidase H. After the final wash, the immuno complexes were eluted from the pansorbin by incubation with 20 P of buffer A (2% SDS, 30 mM DTT, 100 mM Tris pH 6.8) for 45 min at room tempera- ture. Then, 0.05 units of endoglycosidase H in 45 mM sodium citrate buffer, pH 5.5 / 0.2% SDS / 1 mM DTT / 100 pM leupeptin / 10 pM antipain / 15 ug/ml apro- tinin / 0.2 mM PMSF, were added to the immunoprecipitates and incubated over- night at 24 OC. 15 ul of buffer B were added to the samples prior to electro- phoresis.
Electrophoresis. 10 pl of 0.05% bromophenol blue in 50% glycerol were added to the samples, and then heated at 100 OC for 5 min and applied to poly- acrylamide slab gels-(7.5%) containing SDS acording to Laemmli (6). Stacking gel contained 5% polyacrylamide. When indicated urea was added to the gel up to a concentration of 8 M. All gels were processed for fluorography using DMSO- PPO and exposed to Mafe RP-Xl-ray film at -74 OC.
RESULTS
To study synthesis of HMG CoA reductase, rat hepatocytes were pulse la-
belled with f5S]methionine, then the solubilized cell extracts were incubated
with antireductase yglobulin and later precipitated with pansorbin. To know
the amount of Yglobulin necessary for immunoprecipitation, we incubated cell
extracts with increasing amounts of antireductase 7 globulin and measured the
['"I S radioactivity that had precipitated. Results show that 6 pl of antibodies
fully immunoprecipitate the reductase present in lo6 cells.
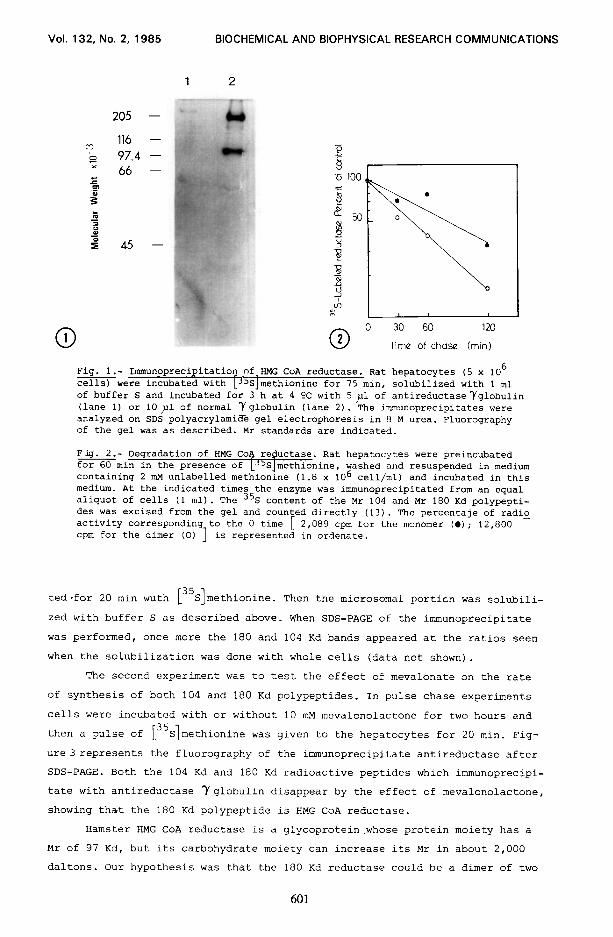
Figure 1 shows an autoradiogram of the immunoprecipitates after urea/SDS
gel electrophoresis. In addition to the 104 Kd reported by different authors
(7, 8, 91, a major band at 180 Kd is seen. Trace bands were also seen in the Mr
region of 60 and 50 Kd. By densitometric scanning, the 180 Kd band represents
73 % of total radioactivity of the gel, while 104 Kd is about 25%. The 180 Kd
had been previously reported by Chin et al (7) in UT1 cells appearing in less
than 7%. According to these authors the 180 Kd band was an aggregate of the 97
Kd reductase and was enriched when urea was omitted from the pansorbin elution
buffer. In our case, polyacrylamide gel electrophoresis with or without urea
did not change the ratio of radioactivity between the 180 and 104 Kd bands.
In order to decide whether or not the 180 Kd and 104 Kd polypeptides
could be different molecular entities, we established a protocol in with hepa-
tocytes were labelled with [35 I S methionine for one hour and then cells were pe
lleted and chased in 1 ml of medium containing 2 mM unlabelled methionine with -
out [35S]methionine. Figure 2 represents the turnover of rat liver 35 S labelled
reductase of the 104 and 180 Kd polypeptides. Half lives of 104 Kd and 180 Kd
reductase are 80 min and 46 min respectively. This data suggest that dimeriza-
tion produces an accelerated rate of degradation of reductase, this fact
suggesting a mechanism of control of enzyme activity.
TO test whether the 180 Kd polypeptide was a reductase or not two new
experiments were done: the first one (indirect) was to isolate by centrifuging
at 105,000 x g microsomes of rat hepatocytes that had been previously incuba-
600
Vol. 132, No. 2, 1985 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
116 97.4 66 -
0 1 0 2 0 Time 30 of 60 chase 120
(min)
Fig. l.- Immunoprecipitation of.BMG CoA reductase. Rat hepatocytes (5 x lo6 cells) were incubated with -('S methionine for 75 min, solubilized with 1 ml of buffer S and incubated for 3 h at 4 OC with 5 ,nl of antireductaseYglobulin (lane 1) or 10~1 of normal Yglobulin (lane 2). The immunoprecipitates were analyzed on SDS polyacrylamide gel electrophoresis in 8 M urea. Fluorography of the gel was as described. Mr standards are indicated.
Fig. 2.- Degradation of HMG Co+ reductase. Rat hepatocytes were preincubated for 60 min in the presence of LJ5Slmethionine, washed and resuspended in medium containing 2 mM unlabelled methionine (1.8 x lo6 cell/ml) and incubated in this medium. At the indicated times the enzyme was immunoprecipitated from an equal aliquot of cells (1 ml). The 35S content of the Mr 104 and Mr 180 Kd polypepti- des was excised from the gel and counted directly (13). The percentaje of radio activity corresponding to the 0 time cpm for the dimer (0) ]
[ 2,089 cpm for the monomer (0); 12,800 - is represented in ordenate.
ted-for 20 min wuth [35S]methionine. Then the microsomal porticn was solubili-
zed with buffer S as described above. When SDS-PAGE of the immunoprecipitate
was performed, once more the 180 and 104 Kd bands appeared at the ratios seen
when the solubilization was done with whole cells (data not shown).
The second experiment was to test the effect of mevalonate on the rate
of synthesis of both 104 and 180 Kd polypeptides. In pulse chase experiments
cells were incubated with or without 10 mu mevalonolactone for two hours and
then a pulse of [35S]meth' ionine was given to the hepatocytes for 20 min. Fig-
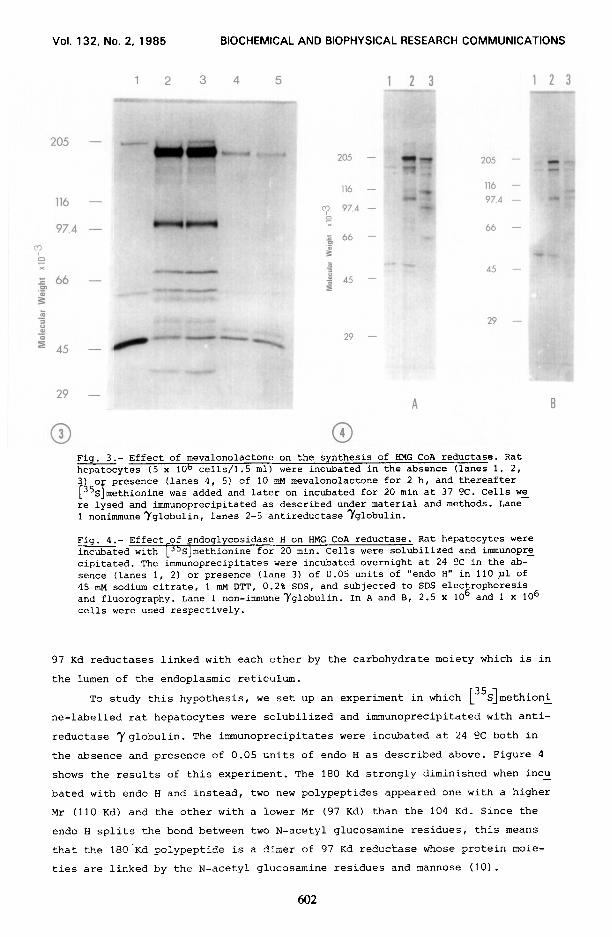
ure 3 represents the fluorography of the immunoprecipitate antireductase after
SDS-PAGE. Both the 104 Kd and 180 Kd radioactive peptides which immunoprecipi-
tate with antireductase Yglobulin disappear by the effect of mevalonolactone,
showing that the 180 Kd polypeptide is HMG CoA reductase.
Hamster HMG CoA reductase is a glycoprotein.whose protein moiety has a
Mr of 97 Kd, but its carbohydrate moiety can increase its Mr in about 2,000
daltons. Our hypothesis was that the 180 Kd reductase could be a dimer of two
601
Vol. 132, No. 2, 1985 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
205 -
116 -
97,4 -
m
0 x
205 -
5 X‘
ii A5 - B
205 -
116 -
97.4 -
66
A5
29
29 -
29 -
A
0 3 0 4
Fig. 3.- Effect of mevalonolactone on the synthesis of HMG CoA reductase. Rat hepatocytes (5 x 1Ob cells/l.5 ml) were incubated in the absence (lanes 1, 2, 3 o presence (lanes 4, 5) of 10 mu mevalonolactone for 2 h, and thereafter
+'i 1 S methionine was added and later on incubated for 20 min at 37 OC. Cells we re lysed and immunoprecipitated as described under material and methods. Lane 1 nonimmuneyglobulin, lanes 2-5 antireductase Yglobulin.
Fig. 4.- Effect-of _endoglycosidase H on HMG CoA reductase. Rat hepatocytes were incubated with LJ'S]methionine for 20 min. Cells were solubilized and immunopre cipitated. The &munoprecipitates were incubated overnight at 24 ?C in the ab- sence (lanes 1, 2) or presence (lane 3) of 0.05 units of "endo H" in 110 pl of 45 mu sodium citrate, 1 mM DTT, 0.2% SDS, and subjected to SDS electrophoresis and fluorography. Lane 1 non-immuneyglobulin. In A and B, 2.5 x lo6 and 1 x 106 cells were used respectively.
97 Kd reductases linked with each other by the carbohydrate moiety which is in
the lumen of the endoplasmic reticulum.
To study this hypothesis, we set up an experiment in which L3'S]methion&
ne-labelled rat hepatocytes were solubilized and immunoprecipitated with anti-
reductase 7 globulin. The immunoprecipitates were incubated at 24 9C both in
the absence and presence of 0.05 units of endo H as described above. Figure 4
shows the results of this experiment. The 180 Kd strongly diminished when incu -
bated with endo H and instead, two new polypeptides appeared one with a higher
Mr (110 Kd) and the other with a lower Mr (97 Kd) than the 104 Kd. Since the
endo H splits the bond between two N-acetyl glucosamine residues, this means
that the 18O.Kd polypeptide is a dimer of 97 Kd reductase whose protein moie-
ties are linked by the N-acetyl glucosamine residues and mannose (10).
602

Vol. 132, No. 2, 1985 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
DISCUSSION
Studies done by Goldstein and Brown group showed that the amino acid se-
quence of HMG COA reductase corresponds to a Mr of 97 Kd. This figure has to
be increased by approx 2,000 daltons due to the carbohydrate moiety which is
chiefly composed by N-acetyl glucosamine and mannose. These data coincide with
the 35 s material immunoprecipitated with antireductase antibodies after a poly - acrylamide gel electrophoresis (10). However, our studies on the rate of synthe -
sis of reductase in rat hepatocytes using [35 1 S methionine as a marker showed
that in addition to the 104 Kd band, another 180 Kd band appeared. This major
polypeptide is thought to be HMG CoA reductase under the following criteria :
i, it is immunoprecipitated by antireductase antobodies; ii, it is microsomal
in origin; iii, it is split by endo H into two polypeptides of analogous Mr
near 97 Kd; iiii, its rate of synthesis is diminished by mevalonolactone toge-
ther with the 97 Kd polypeptide. We agree with the results of Edwards et al
(11) who reported that mevalonate does not change the rate of synthesis of to-
tal protein in rat hepatocytes but decreases the rqductase synthesis rate. The
180 Kd band had been reported by Chin et al (7), they considered that it was
an aggregate that was enhanced by boiling SDS but dissociated by urea/SDS. The
SDS-PAGE of immunoprecipitated reductase in fig 1 was done in urea, the aggre-
gate is present in larger amounts than the 97 Kd reductase. In addition, the
different rate of degradation of 97 Kd and 180 Kd reductases, shows that they
are present in hepatocytes as different entities.
The different ratio 80%/20% corresponding to 180 vs 104 Kd reductase
bands with respect to different cellular lines (7), (9), (12), of about lo%/
90%, can be explained by the delipidization treatment done four days previous-
ly to the experiment. However, isolated rat hepatocytes being studied itnmedia-
tely after isolation represent the real physiological state of reductase in
rat liver. The different rate of degradation of the dimer with respect to the
monomer, near two fold, suggests a regulation mechanism of reductase and hen-
ce of cholesterol biosynthesis.
The experiment made with endo H shows that the 180 Kd polypeptide can be
split into two monomers with similar Mr. Taking into account the fact that en-
do H splits the binding between two molecules of N-acetyl glucosamine, it
appears that the 180 Kd is present in the endoplasmic reticulum of rat hepato-
cytes and represents two monomers of reductase linked by covalent binding in
the carbohydrate moiety in the lumen of the reticulum. The different Mr of the
two subunits can be explained on the basis of their different amount of manno-
se.
In view of these experiments we propose a change in the model of reducta - se proposed by Liscum et al (3) as represented in Fig 5 : The monomer of reduc - tase is in equilibrium with a dimer composed of two monomers linked by the car -
603

Vol. 132, No. 2, 1985 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Fig. 5.- Model for the dimeric form of HMG CoA reductase in the membrane of the endoplasmic reticulum. The model of the monomer is taken from reference 3. Two monomers are linked by a carbohydrate chain composed of N-acetyl glucosamine and mannose residues.
bohydrate chain. Alternatively, the model could be extended to the formation
of a polymer of reductase cross-linked by one or more mannose oligosacharide
chains. However, our techniques of SDS-PAGE cannot allow us to prove this last
assertion.
ACKNOWLEDGMENTS
This work was supported by grants 3/602 (1981) and 3391-83 from Comisi6n
Asesora de Investigaci6n Cientffica y Tecnica and grant 85/1293 from Fondo de
Investigaciones Sanitarias de la Seguridad Social (Spain).
REFERENCES
1. 2.
3.
4. 5. 6. 7.
8.
9.
10.
11.
12.
13.
Brown, M.S. and Goldstein, J.L. (1980) J. Lipid Res. 21, 505-517, Chin, D.J., Gil, G., Russell, D.W., Liscum, L., Luskey, K.L., Basu, S.K., Okayama, H., Berg, P., Goldstein, J.L. and Brown, M.S. (1984) Nature (Land) 308, 613-617. Liscum, L., Finer-Moore, J., Stroud, R.M., Luskey, K.L., Brown, M.S. and Goldstein, J.L. (1985) J. Biol. Chem. 260, 522-530. Seglen, P.O. (1976) Methods Cell. Biol. 13, 29-83. Eagle, H. (1959) Science 130, 342. Laemmli, U.K. (1970) Nature (Land) 227, 680-685. Chin, D.J., Luskey, K.L., Faust,J.R., Mac Donald, R.J., Brown, M.S. and Goldstein, J.L. (1982) Proc. Natl. Acad. Sci. USA 79, 7704-77@8. Edwards, P.A., Lan, S.F. and Fogelman, A.M. (1983) J. Biol. Chem. 258, 10219-10222. Hardeman, E.C., Jenke, H.S. and Simoni, R.D. (1983) Proc. Natl.Acad.Sci. USA, 80, 1516-1520. Liscum, L., Cummings, R.D., Anderson, R.G.W., DeMartino, G.N., Goldstein, J.L. and Brown, M.S. (1983) Proc. Natl. Acad. Sci. USA 80, 7165-7169. Edwards, P.A., Lan, S.F., Tanaka, R.D. and Fogelman, A.M. (1983) J. Biol. Chem. 258, 7272-7275. Skalnik, D.G., Brown, D.A., Brown, P.C., Friedman, R.L., Hardeman, E-C., Schimke, R.T. and Simoni, R.D. (1985) J. Biol. Chem. 260, 1991-1994. Laskey, R.A. (1980) Methods Enzymol. 65, 363-371.
604



























![Designing Dimeric Lanthanide(III)-Containing Ionic liquids › ws › files › 158240242 › ...COMMUNICATION Designing Dimeric Lanthanide(III)-Containing Ionic liquids Éadaoin McCourt,[a]](https://img.pdfslide.net/doc/110x75/60b904bbc8cfbf6cfb110109/designing-dimeric-lanthanideiii-containing-ionic-liquids-a-ws-a-files-a.jpg)