Embed Size (px)
Citation preview

165
Journal of Oceanography, Vol. 57, pp. 165 to 175, 2001
Keywords:⋅ Dinophysis fortii,⋅ diarrhetic shellfishpoisoning,
⋅ Sanriku,⋅ Okkirai Bay,⋅ Otsuchi Bay,⋅ internal tidal wave,⋅ synechococcoidcyanobacteria,
⋅ cryptomonad.
* Corresponding author. E-mail: [email protected]
Copyright © The Oceanographic Society of Japan.
Recent Occurrences of Dinophysis fortii (Dinophyceae)in the Okkirai Bay, Sanriku, Northern Japan, and RelatedEnvironmental Factors
KAZUHIKO KOIKE1*, HIROTAKA OTOBE2, MINORU TAKAGI3, TADAHISA YOSHIDA1,TAKEHIKO OGATA1 and TAKASHI ISHIMARU4
1School of Fisheries Sciences, Kitasato University, Sanriku, Kesen, Iwate 022-0101, Japan2Otsuchi Marine Research Center, Ocean Research Institute, The University of Tokyo, Otsuchi, Kamiheii, Iwate 028-1102, Japan3Iwate Prefectural Fisheries Technology Center, Heita, Kamaishi, Iwate 026-0001, Japan4Department of Ocean Sciences, Tokyo University of Fisheries, Konan, Minato-ku, Tokyo 108-8477, Japan
(Received 15 November 1999; in revised form 13 October 2000; accepted 13 October 2000)
Occurrence of Dinophysis fortii, a causative organism of diarrhetic shellfish poison-ing, in the Okkirai Bay, Sanriku was surveyed in 1995, 1996, 1998 and 1999. In eachyear, its major occurrence was detected from the late May or early June and contin-ued until the late June or early July. Seawater temperature, salinity and nutrientsmeasurements suggested that inflows of offshore water into the bay played key roleon the first major occurrence of D. fortii. With an analysis of continuous temperaturedata in the Otsuchi Bay which locates north of the Okkirai Bay, this influent wasconsidered to be intermittent inflow of the offshore water by internal tidal waveswhich propagated from north to south. First occurrence peak of D. fortii was syn-chronous with phycobilin containing microalgae, synechococcoid cyanobacteria andcryptomonad, in all years. In vivo fluorescence measurement of D. fortii cells in 1995and 96 showed that the cells in these microalgal-rich water contained more phycobi-lin pigment than those in the microalgal-poor water. The result may support a hy-pothesis that D. fortii acquires phycobilin by an uptake of these microalgae. After thefirst major occurrence in the bay, D. fortii sometimes occurred in rather inshore wa-ters where showed elevated ammonium level possibly due to increased heterotrophicactivity. Together with another finding that D. fortii is mixotrophic, it could be as-sumed that the environment being suited to heterotrophic nutrition also stimulatesD. fortii growth in the bay.
tal parameters. Fukuyo (1997) reviewed that D. fortiimainly occurred along coasts in these areas where aredominated by either Tsushima, Tsugaru or Oyashio cur-rents, the main currents in the Tohoku and Hokkaido re-gions of the northern Japan. In 1995, 1996, 1998, and1999, we have carried out field surveys on the occurrenceof D. fortii in the Okkirai Bay of the Sanriku coast, wherebeing affected by DSP toxins. In this paper, we describethe survey results and discuss the pattern of occurrencewith affective simultaneous environmental parameters,such as seawater temperature, salinity and nutrients.
In addition to these factors, we also discuss the rela-tion between possible nutrition forms of D. fortii and en-vironmental factors. The cell density of phycobilin-con-taining algae is one of them. Although Dinophysis spp.
1. IntroductionSince Yasumoto et al. (1980) first found toxins for
diarrhetic shellfish poisoning (DSP) in Dinophysis fortiiPavillard, this species has been recognized as the mainorganism responsible for DSP toxin contamination inshellfish in the northern Japan. Intensive field surveys onthis species have been carried out in various coastal ar-eas of the northern Japan (e.g., Osaka, 1985; Igarashi,1986; Iwasaki, 1986). These surveys have elucidated theperiods of occurrence and some associating environmen-

166 K. Koike et al.
have not been successfully cultured, there have been anumber of observations on wild cells; the plastid of pho-tosynthetic Dinophysis is similar to that of cryptomonadin its pigment and structural profile (e.g., Schnepf andElbrächter, 1988); the pigment concentration and the plas-tid morphology are variable (Fukuyo, 1997); and D. fortiihas the ability to take up cryptomonad cells (Ishimaru etal., 1988). From these observations, it could be assumedthat the plastid of D. fortii might be derived from tempo-rally acquired microalgae, by a process termed“kleptoplastidy (Lasen, 1992; Laval-Peuto, 1992; Schnepfand Elbrächter, 1992)”. The validity of this hypothesishas not yet been adequately documented with experimen-tal or field results. Therefore, we measured the occurrenceof phycobilin-containing microalgae, cryptomonad andcyanobacteria that occur simultaneously with D. fortii inthe bay. The relationship between these algae and D. fortiiwill be discussed together with a result of the phycobilincontents of D. fortii.
2. Materials and MethodsWeekly (April to December) or monthly (January to
March) samplings were conducted at point A (12 m depth)in 1995 and at point B (26 m depth) in 1996, 1998, and1999, in the Okkirai Bay, Sanriku, Iwate, Japan (Fig. 1).No sampling was conducted in 1997. Samples were takenbetween 10:00 and 11:00 am. Water samples were col-lected with a Van Dorn sampler at surface, 5, and 10 mdepths in 1995 and at surface, 5, 10, 15, and 20 m depthsin the other years.
The analysis of each sample was as follows. One literof the water sample was concentrated with a membrane
filter (5.0 µm pore size; 47 mm in diameter; AdvantecCo., Japan) without vacuuming to a volume of 5 mL(modified from a method of Iizuka, 1979), and the con-centrated sample was fixed with glutaraldehyde at the fi-nal concentration of 2.5%. The number of Dinophysis spp.cell per liter of the sample water was determined by count-ing 1/5 of the volume of the concentrated sample under amicroscope; thus, the detectable lower limit was 5cells·L–1. For estimating the biomass of synechococcoidcyanobacteria and cryptomonad, the sample water sievedthrough 20 µm mesh was fixed with glutaraldehyde at2.5% final concentration. The particles in the filtrate weretrapped onto a polycarbonate membrane filter (0.2 µmpore size; 13 mm in diameter; Advantec Co., Japan) bygentle vacuuming (<100 mm Hg). Then the filter wasmounted on a slide glass and covered with one drop ofnon-fluoresced immersion oil (Olympus Co., Japan) anda cover slip. They were observed under BH2-RFCepifluorescence microscope (Olympus Co., Japan)equipped with filter setting as follow; 455–490 nm exci-tation filter, 500 nm beam splitter. Detailed procedure isdescribed in Booth (1993). Detectable limit of themicroalgae in this method is ca. 20 cells·mL–1. The analy-sis of the nutrients (NO3-N, NO2-N, NH4-N, PO4-P) wereperformed according to the methods developed byValderrama (1995). Vertical profiles of seawater tempera-ture and salinity were measured with an AST-200 STD(Alec Electronics Co., Japan). Since time series data ofseawater temperature was not available in the Okkirai Bay,the data set of the Otsuchi Bay (Otobe et al., 1999) whichlocates 31 km north of the Okkirai Bay (Fig. 1) was usedfor analyzing the relation to the occurrence of D. fortii.
Fig. 1. Map of sampling points.
Occurrence of Dinophysis fortii in the Okkirai Bay 167
In 1995 and 1996, D. fortii cells collected by verti-cal hauling of 20 µm plankton net (from 20 m depth tothe surface) in the each sampling were subjected to meas-urement of relative in vivo fluorescence intensity of phy-cobilin pigment. The measurement was conducted on tenindividual cells using a P-1 micro photometer (Nikon Co.,Japan) equipped to the BH2-RFC epifluorescence micro-scope (Olympus Co., Japan) with ×40 objective lens. Thefilters setting was as follow; 530–545 nm excitation fil-ter, 570 nm beam splitter, 620 ± 15 nm barrier filter. Thisfilter combination was determined by testing the enoughdetection level of in vivo fluorescence intensity from thephycobilin pigment of cryptomonad culture. Sensitivityof the micro photometer on the each measurement werestandardized using a fluorescence-color acrylic plate in-stead of a slide glass. The cells were mounted on non-fluoresced slide glass and cover slip prior to the meas-urement.
3. Results
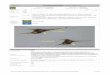
3.1 Occurrence of D. fortii in the Okkirai Bay and thesimultaneous factors of temperature, salinity and nu-trientsFigures 2 to 5 show vertical profiles and transitions
of D. fortii occurrence (A), temperature (B), salinity (C)and nutrients (D–G). The samplings were conductedweekly (April to December) or monthly (January toMarch) in the surveyed years, but only the results duringlate April through late July are presented in figures dueto no detectable D. fortii occurred on the other monthsexcept a few cases (5 August in 1996 and 25 Septemberin 1998: mentioned in Table 1). Table 1 summarized theperiods of D. fortii occurrence more than the detectablecell density (�5 cells·L–1) and the period of more than20 cells·L–1 which is sufficient level for toxin contami-nation in shellfish according to Igarashi (1985), and their
Fig. 2. Vertical profiles and transitions of D. fortii occurrence (A), seawater temperature (B), salinity (C), nutrients (D–G), andphycobilin containing microalgae (H, I) in 1995.
168 K. Koike et al.
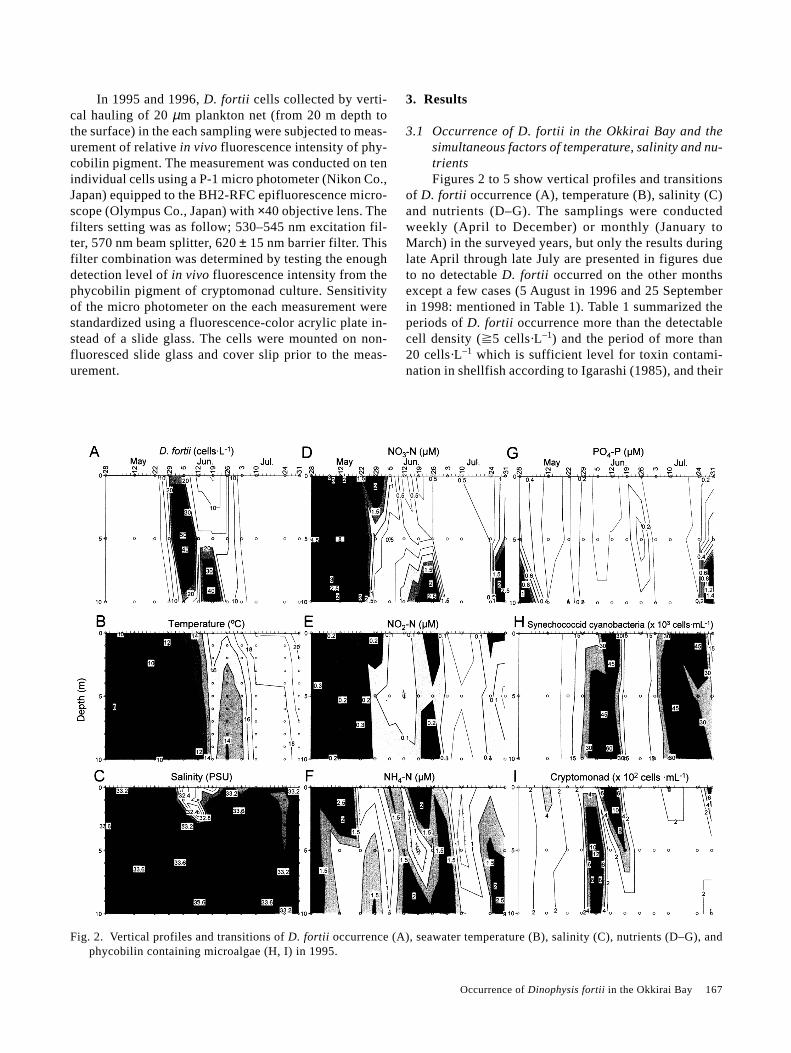
Fig. 3. Same as Fig. 2 but in 1996.
temperature ranges. In each year, the cell density morethan 20 cells·L–1 was observed from late May or earlyJune and continued until late June or early July. Tempera-ture for the detectable occurrence was varied from 7.2°Cand 22.5°C, and for the density of more than 20 cells·L–1
was in the range between 10.1 and 15.3°C. The specieswas not detected at a salinity less than 32.9 and the celldensities more than 20 cells·L–1 were detected at salinitymore than 33.2 with a single exception at the surface on
25 June 1996 (described in below). D. fortii occurred inextremely high cell densities in 1999, compared with theprevious years.
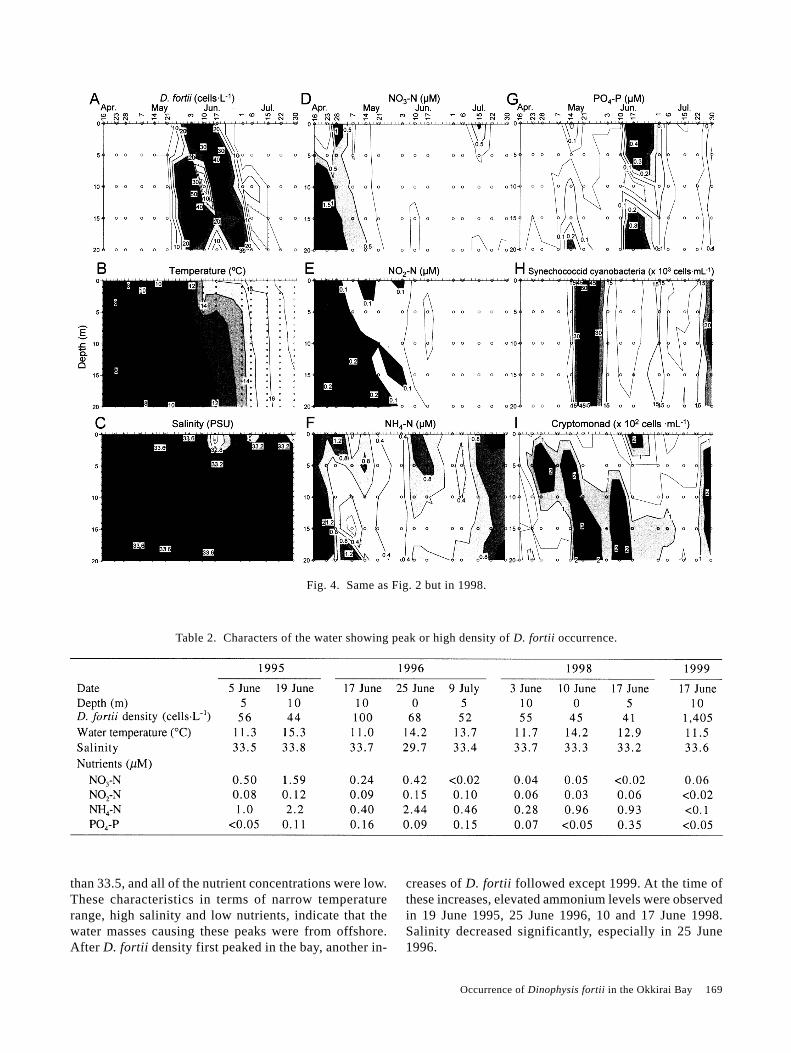
Table 2 summarizes characters of the water showingpeaks or in which D. fortii occurred in high cell densi-ties. When the cell densities first peaked in the bay (5June 1995, 17 June 1996, 3 June 1998 and 17 June 1999),the water temperature was in a narrow range between11.0–11.7°C. The salinities at these peaks were high, more
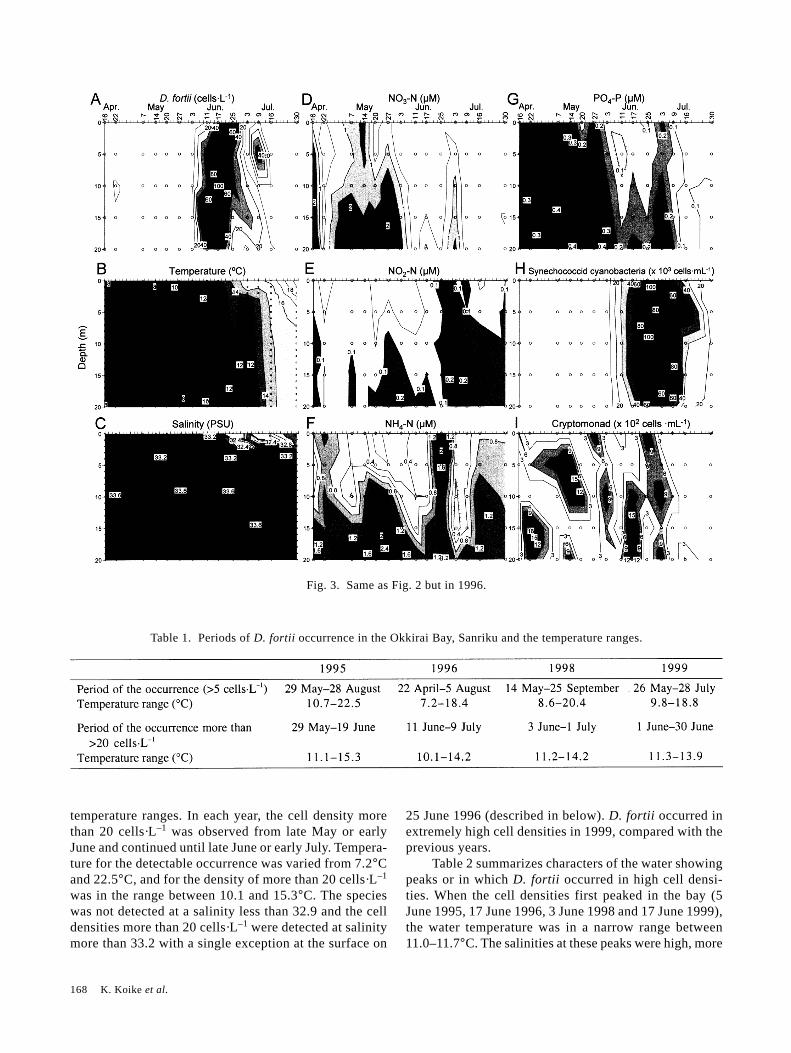
Table 1. Periods of D. fortii occurrence in the Okkirai Bay, Sanriku and the temperature ranges.
Occurrence of Dinophysis fortii in the Okkirai Bay 169
than 33.5, and all of the nutrient concentrations were low.These characteristics in terms of narrow temperaturerange, high salinity and low nutrients, indicate that thewater masses causing these peaks were from offshore.After D. fortii density first peaked in the bay, another in-
creases of D. fortii followed except 1999. At the time ofthese increases, elevated ammonium levels were observedin 19 June 1995, 25 June 1996, 10 and 17 June 1998.Salinity decreased significantly, especially in 25 June1996.
Fig. 4. Same as Fig. 2 but in 1998.
Table 2. Characters of the water showing peak or high density of D. fortii occurrence.

170 K. Koike et al.
Fig. 5. Same as Fig. 2 but in 1999.
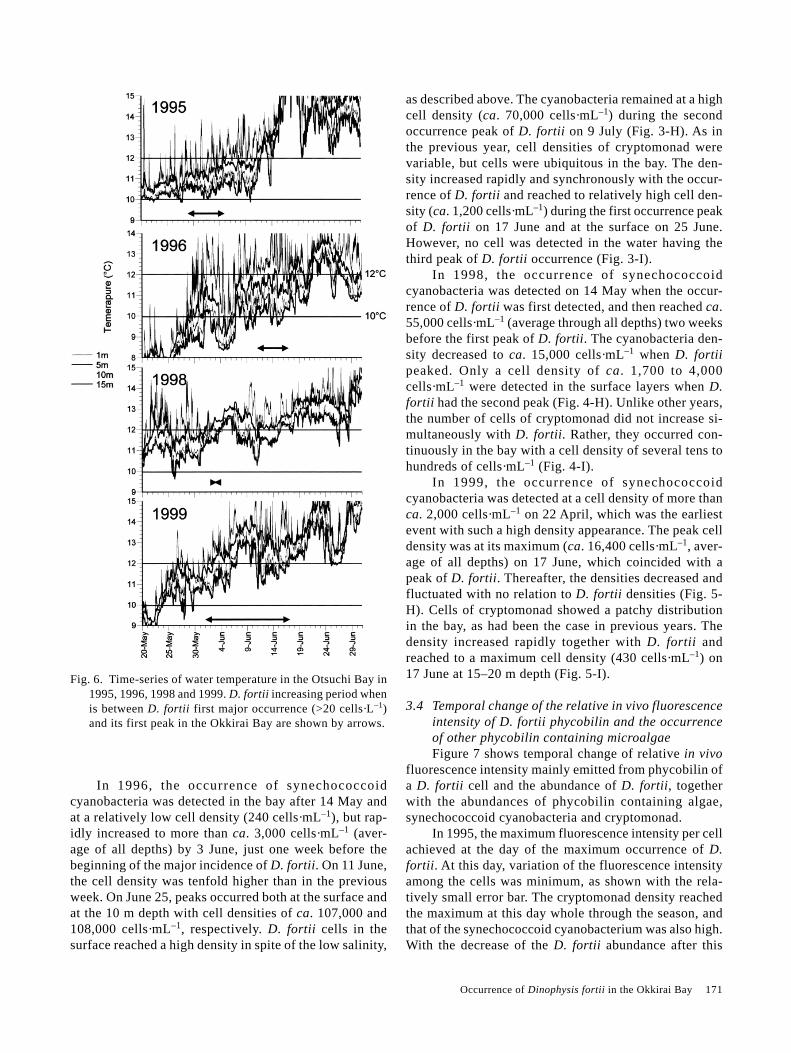
3.2 Relation between D. fortii occurrence and water tem-perature in the Otsuchi BayFigure 6 shows the time series of water temperatures
at 1, 5, 10 and 15 m depth in the Otsuchi Bay from lateMay to late June and the increasing period of D. fortiifrom the first major occurrence (>20 cells·L–1) to its firstpeak in the Okkirai Bay (arrows). The diurnal change ofthe temperature affected by the solar radiation appearedin the one-meter layer. From late May, water tempera-tures below 5 m layer gradually increased and thus strati-fied. Corresponding to the development of the stratifica-tion, the periodical fluctuations of the bottom layer watertemperature (10 m and 15 m) became larger. It seemedthat the periodical change was occurred by the internalwaves.
As mentioned above, occurrence of D. fortii morethan 20 cells·L–1 started at the temperature ca. 10°C andthe first peak appeared at ca. 11–12°C. During these pe-riods, it was frequently found that the temperature at 10m and/or 15 m layers changed in the range of 10–12°Cwith diurnal or semi-diurnal cycle.
3.3 Relation between D. fortii occurrence and phycobi-lin-containing microalgaeIn 1995, occurrence of synechococcoid cyanobacteria
first exceeded the detectable level (20 cells·mL–1) on 22May, one week before the detectable level of D. fortiioccurrence, and the average cell density for all sampleddepths was ca. 10,500 cells·mL–1. On the day of the firstincidence peak of D. fortii, the cell density reached ca.38,000 cells·mL–1. After this period, the cell densityseemed to modulate; the occurrence pattern had no par-ticular relationship to the D. fortii occurrence pattern. Celldensities had decreased to ca. 15,300 cells·mL–1 by thesecond occurrence peak of D. fortii. The cell density inthe bay remained higher than ca. 20,000 cells·mL–1 afterthis period (Fig. 2-H). Cryptomonad seemed to be patchybut constantly detected in the bay. The density increasedrapidly and synchronously with the occurrence of D. fortiiand reached its maximum cell density (1,220 cells·mL–1)in the same sample having the first occurrence peak forD. fortii. However, no cell was detected during the sec-ond incidence peak of D. fortii (Fig. 2-I).
Occurrence of Dinophysis fortii in the Okkirai Bay 171
In 1996, the occurrence of synechococcoidcyanobacteria was detected in the bay after 14 May andat a relatively low cell density (240 cells·mL–1), but rap-idly increased to more than ca. 3,000 cells·mL–1 (aver-age of all depths) by 3 June, just one week before thebeginning of the major incidence of D. fortii. On 11 June,the cell density was tenfold higher than in the previousweek. On June 25, peaks occurred both at the surface andat the 10 m depth with cell densities of ca. 107,000 and108,000 cells·mL–1, respectively. D. fortii cells in thesurface reached a high density in spite of the low salinity,
as described above. The cyanobacteria remained at a highcell density (ca. 70,000 cells·mL–1) during the secondoccurrence peak of D. fortii on 9 July (Fig. 3-H). As inthe previous year, cell densities of cryptomonad werevariable, but cells were ubiquitous in the bay. The den-sity increased rapidly and synchronously with the occur-rence of D. fortii and reached to relatively high cell den-sity (ca. 1,200 cells·mL–1) during the first occurrence peakof D. fortii on 17 June and at the surface on 25 June.However, no cell was detected in the water having thethird peak of D. fortii occurrence (Fig. 3-I).
In 1998, the occurrence of synechococcoidcyanobacteria was detected on 14 May when the occur-rence of D. fortii was first detected, and then reached ca.55,000 cells·mL–1 (average through all depths) two weeksbefore the first peak of D. fortii. The cyanobacteria den-sity decreased to ca. 15,000 cells·mL–1 when D. fortiipeaked. Only a cell density of ca. 1,700 to 4,000cells·mL–1 were detected in the surface layers when D.fortii had the second peak (Fig. 4-H). Unlike other years,the number of cells of cryptomonad did not increase si-multaneously with D. fortii. Rather, they occurred con-tinuously in the bay with a cell density of several tens tohundreds of cells·mL–1 (Fig. 4-I).
In 1999, the occurrence of synechococcoidcyanobacteria was detected at a cell density of more thanca. 2,000 cells·mL–1 on 22 April, which was the earliestevent with such a high density appearance. The peak celldensity was at its maximum (ca. 16,400 cells·mL–1, aver-age of all depths) on 17 June, which coincided with apeak of D. fortii. Thereafter, the densities decreased andfluctuated with no relation to D. fortii densities (Fig. 5-H). Cells of cryptomonad showed a patchy distributionin the bay, as had been the case in previous years. Thedensity increased rapidly together with D. fortii andreached to a maximum cell density (430 cells·mL–1) on17 June at 15–20 m depth (Fig. 5-I).
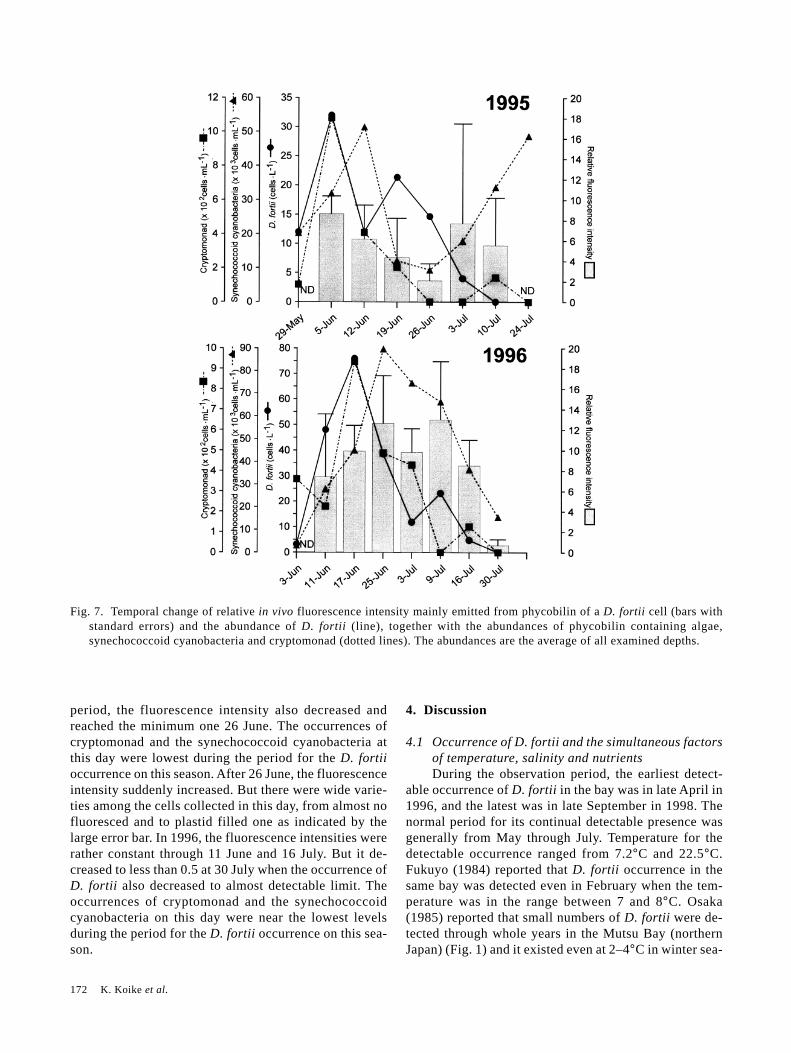
3.4 Temporal change of the relative in vivo fluorescenceintensity of D. fortii phycobilin and the occurrenceof other phycobilin containing microalgaeFigure 7 shows temporal change of relative in vivo
fluorescence intensity mainly emitted from phycobilin ofa D. fortii cell and the abundance of D. fortii, togetherwith the abundances of phycobilin containing algae,synechococcoid cyanobacteria and cryptomonad.
In 1995, the maximum fluorescence intensity per cellachieved at the day of the maximum occurrence of D.fortii. At this day, variation of the fluorescence intensityamong the cells was minimum, as shown with the rela-tively small error bar. The cryptomonad density reachedthe maximum at this day whole through the season, andthat of the synechococcoid cyanobacterium was also high.With the decrease of the D. fortii abundance after this
Fig. 6. Time-series of water temperature in the Otsuchi Bay in1995, 1996, 1998 and 1999. D. fortii increasing period whenis between D. fortii first major occurrence (>20 cells·L–1)and its first peak in the Okkirai Bay are shown by arrows.
172 K. Koike et al.
Fig. 7. Temporal change of relative in vivo fluorescence intensity mainly emitted from phycobilin of a D. fortii cell (bars withstandard errors) and the abundance of D. fortii (line), together with the abundances of phycobilin containing algae,synechococcoid cyanobacteria and cryptomonad (dotted lines). The abundances are the average of all examined depths.
period, the fluorescence intensity also decreased andreached the minimum one 26 June. The occurrences ofcryptomonad and the synechococcoid cyanobacteria atthis day were lowest during the period for the D. fortiioccurrence on this season. After 26 June, the fluorescenceintensity suddenly increased. But there were wide varie-ties among the cells collected in this day, from almost nofluoresced and to plastid filled one as indicated by thelarge error bar. In 1996, the fluorescence intensities wererather constant through 11 June and 16 July. But it de-creased to less than 0.5 at 30 July when the occurrence ofD. fortii also decreased to almost detectable limit. Theoccurrences of cryptomonad and the synechococcoidcyanobacteria on this day were near the lowest levelsduring the period for the D. fortii occurrence on this sea-son.
4. Discussion
4.1 Occurrence of D. fortii and the simultaneous factorsof temperature, salinity and nutrientsDuring the observation period, the earliest detect-
able occurrence of D. fortii in the bay was in late April in1996, and the latest was in late September in 1998. Thenormal period for its continual detectable presence wasgenerally from May through July. Temperature for thedetectable occurrence ranged from 7.2°C and 22.5°C.Fukuyo (1984) reported that D. fortii occurrence in thesame bay was detected even in February when the tem-perature was in the range between 7 and 8°C. Osaka(1985) reported that small numbers of D. fortii were de-tected through whole years in the Mutsu Bay (northernJapan) (Fig. 1) and it existed even at 2–4°C in winter sea-

Occurrence of Dinophysis fortii in the Okkirai Bay 173
son. Therefore, the cell under detectable levels (<5cells·L–1) may subsist in other seasons according to oursurvey.
D. fortii densities of more than 20 cells·L–1 occurredfrom late May or late June and continued until late Juneor early July. The temperature range when the cell den-sity exceeded 20 cells·L–1 was in a range between 10.1and 15.3°C, and the density dropped after the tempera-ture exceeded above ca. 15°C. Similar relation betweenthe water temperature range and the D. fortii occurrencewas also reported in the Kesennuma Bay (Fig. 1), on theSanriku coast (Hoshiai et al., 1996). The temperatureduring the first peak in the Okkirai Bay was in a verynarrow range between 11.0 and 11.7°C throughout theobservation period. The occurrence more than 20cells·L–1 may thus be estimated to begin at 10°C and reachto the peak at approximately 11–12°C.
With the relation to salinity and the nutrient, the cellswere not detected at salinities of less than 32.9 except forone case in 1996. The nutrient concentrations were al-ways low when D. fortii density first peaked. In 1999,the nutrient concentration was extremely low when D.fortii occurred in a high density.
From the results described above, it can be seen thatthe first peak of D. fortii cells occurs in a very narrowtemperature range, at high salinities, and in low nutrientconcentrations. These observations show that inflows ofoffshore water into the bay are important factor on thefirst major occurrence of D. fortii. In our monitoring re-sults of continuous water temperature in the Otsuchi Bayas shown in Fig. 6, the temperature at 10 and 15 m layerschanged periodically along with the stratification afterlate May. These temperature changes having a diurnal orsemi-diurnal cycle were clearly observed during the D.fortii increasing period in the Okkirai Bay, and the tem-perature of 10 and/or 15 m layers periodically droppedwithin the range of 10–12°C which is considered to beadequate to the major occurrence of D. fortii as describedabove. These temperature drops are considered to be in-termittent inflow of the offshore water by internal tidalwaves as indicated by Okazaki (1990). The inflow of off-shore water into the Okkirai Bay should have a time de-lay of about 45 hours after the event in the Otsuchi Bay.This is because the Okkirai Bay is located 31 km south ofthe Otsuchi Bay and the internal long period waves,closely relate to the intermittent vertical displacement ofthe offshore thermocline, were estimated to propagatesouthward at a speed of about 19 cm·sec–1 (Okazaki,1990). The above results suggest that the inflows of off-shore water by the internal tidal waves derived the firstmajor increase of D. fortii in the bay. It is still difficult toconfirm whether massive populations of D. fortii weretransported from the offshore region by the water move-ment, or the growths of the seed cells, subsisted in the
bay during winter till spring, were stimulated by the in-flows of offshore waters. Iwasaki (1986) already reportedthat D. fortii cells were transported from offshore regionby the internal wave movement in the Kashima-nada. Ourresult may support that the same mechanism is occurringalso in the Sanriku coast. Further offshore survey will beneeded together with the additional confirmation of theexistence of the seed cell in bay and its possible growthresponse after the inflow of offshore water.
There are observations on the field occurrence ofother photosynthetic species of Dinophysis. Significantabundances are frequently related to stratification of thewater column (e.g., Delmas et al., 1992; Reguera et al.,1995; Peperzak et al., 1996). Delmas et al. (1992) con-cluded stratification of sufficient magnitude (∆t > 5°C;temperature at the base of the upper mixed layer minusthat of the bottom layer) and duration (>2 weeks) are thefactors necessary for an increase in the number ofDinophysis cells. In this study, although the magnitudeof the difference is not as distinct as with their results,the major occurrences of D. fortii appeared below sig-nificant pycnocline. This was most noticeable when thetemperature was between 10 and 12°C. Then when sig-nificant differences between the surface and bottom tem-peratures (�2.6°C) were observed in 1996 and 1999, thedensity of D. fortii was high (�100 cells·L–1 at the maxi-mum). On the contrary, when the temperature differencewas small (<2.6°C) as in 1995 and 1998, the density waslow. These results also suggest that the effects of the in-ternal long period wave, which cause the stratification ofwater, and the internal tidal wave are important factorsfor the major occurrence of D. fortii in the bay.
4.2 Relation between possible nutrition forms of D. fortiiand other environmental factorsWe observed synchronous occurrences of
synechococcoid cyanobacteria and cryptomonad alongwith D. fortii. The same kind of phenomenon was con-firmed by Giacobbe et al. (1995) for Dinophysis saccu-lus with Synechococcus. Photosynthetic Dinophysis con-tain phycobilin (Geider and Gunter, 1989; Vesk et al.,1996; Hewes et al., 1998), which is typical photosyntheticpigment for cyanophyta, cryptophyta and rhodophyta, andtheir plastid structure is similar to that of cryptophyta(Schnepf and Elbrächter, 1988). However, no other tracesof cryptomonad organelles have been observed inDinophysis cell. Therefore, if the plastid is indeed an or-ganelle acquired from a cryptomonad, the association hasbeen suggested to be long established one (Lucas andVesk, 1990). On the contrary, Ishimaru et al. (1988) ob-served that D. fortii took up cryptomonad cells and main-tained the pigment for longer periods than those withoutaccess to cryptomonad cells. Therefore, it is a plausiblehypothesis that D. fortii temporally acquires its photo-

174 K. Koike et al.
synthetic pigments by uptake of cryptomonad plastids. Ifthis hypothesis is justifiable, the cyanobacteria (anothermicroalga besides cryptomonad that contains phycobilin)could also make a substantial contribution.
In the current field study, we found the synchronousoccurrence of synechococcoid cyanobacteria andcryptomonad along with D. fortii; synechococcoidcyanobacteria began to appear in high cell density justbefore the first occurrence of D. fortii and the cell den-sity of cryptomonad simultaneously increased with thefirst peak of D. fortii excepting the case in 1998. Thisdesignates that cells of D. fortii were always in the cir-cumstance where phycobilin-containing microalgae areavailable. Moreover, the cells of D. fortii in this circum-stance were filled with phycobilin pigment; they containedmore pigment than other cells in these microalgal-poorwater. These indicate that the circumstance sustaining themechanism mentioned in the hypothesis, where D. fortiitemporarilly acquires the pigment from these microalgae,is sufficiently evolving in the natural environment. Andthese may support the validity of the hypothesis, althoughthe acquisition mechanism of these microalge by pho-totrophic Dinophysis is still not a proven phenomenonand needs experimental confirmation.
Other than the contribution of cryptomonad andsynechococcoid cyanobacteria, are there any other possi-ble factors that regulate the growth of D. fortii in the bay?As mentioned above, the first occurrence peaks of D. fortiiin the bay are considered to be caused by offshore watersand internal tidal waves. However, there were other oc-currence peaks in inshore waters when the temperaturewas higher than 12°C and when the salinity was lowerthan 33.4, such as the cases of surface on 25 June 1996,10 June 1998, and 5–10 m on 17 June 1998. These peakswere on degradation periods of diatom blooms and in-volved increases in the ratio of NH4-N compared to othernutrients, probably due to increased heterotrophic activi-ties of bacteria and protists. Taylor et al. (1994) also re-ported that a dense occurrence of D. fortii was associatedwith elevated ammonium level on the British Columbiacoast. We already reported that D. fortii is a mixotrophicspecies and preys on eukaryote and possibly on bacteria(Koike et al., 2000). Thus, it could be assumed that theenvironment being suited to heterotrophic nutrition alsostimulates D. fortii growth in the bay.
5. SummaryOccurrence of Dinophysis fortii, a causative of
diarrhetic shellfish poisoning, in the Okkirai Bay locatedat Sanriku, Iwate, northern Japan was surveyed in 1995,1996, 1998 and 1999. Through the whole observationperiods, the occurrence of D. fortii in cell density exceed-ing 20 cells·L–1 was detected from late May or early Juneand continued until late June or early July. The tempera-
ture during these periods was in the range between 10.1and 15.3°C, and the first peak of the season always oc-curred in the very narrow temperature range between 11.0and 11.7°C. Consequently, the occurrence exceeding 20cells·L–1 was generally estimated to begin at 10°C andreach to the peak at 11–12°C. The species was not de-tected at salinity less than 32.9 with a few exceptions.Nutrient levels were always low during the first peak ofD. fortii. Prior to or during the D. fortii increasing periodin the Okkirai Bay, intermittent temperature drops hav-ing a diurnal or semi-diurnal cycle in the range of 10–12°C were observed at 10 and/or 15 m depths in theOtsuchi Bay locating at 31 km north from the OkkiraiBay. These temperature drops were considered to be in-termittent inflows of the offshore water by internal tidal.These results show that the inflows of offshore water bythe internal tidal waves derived the first major increaseof D. fortii in the Okkirai Bay. Other than the relation totemperature, salinity and nutrients, synchronous occur-rences of synechococcoid cyanobacteria and cryptomonadwere observed together with the first major occurrenceof D. fortii in the bay. The cells of D. fortii in this cir-cumstance were filled with phycobilin pigment compar-ing with other cells in the water where these microalgaewere scarce. This may partly support a hypothesis ofIshimaru et al. (1988) that D. fortii temporally acquiresthe pigment from phycobilin-containing microalgae forits own photosynthesis. After the first major occurrencein the bay, D. fortii sometimes occurred in rather inshorewaters where showed elevated ammonium level possiblydue to increased heterotrophic activity. Together withanother finding that D. fortii is mixotrophic (Koike et al.,2000), it could be assumed that the environment beingsuited to heterotrophic nutrition also stimulates D. fortiigrowth in the bay.
AcknowledgementsWe express our sincere thanks to Messrs. Hiroki
Noma, Takehito Kida, Tomoaki Inoue, Hajime Itoh,Ryuichi Kijima and Ms. Rie Sugiyama for collecting fielddata. Our thanks are also due to Dr. Katsushi Sekiguchi,Messrs. Yoshimasa Kaga and Shinnosuke Kaga of IwatePrefectural Fisheries Technology Center for their coop-eration on the field samplings, and Dr. Masaaki Kodama,Kitasato University, for his encouragement. This work waspartly supported by a grant-in-aid (No. 09760181) fromthe Ministry of Education, Science, Sports and Culture,and also by a grand-in-aid for Cooperative InternationalResearch Project on Marine and Coastal Environmentfrom Iwate Prefecture, Japan.
ReferencesBooth, B. C. (1993): Estimating cell concentration and biomass
of autotrophic plankton using microscopy. p. 199–206. In

Occurrence of Dinophysis fortii in the Okkirai Bay 175
Handbook of Methods in Aquatic Microbial Ecology, ed.by P. F. Kemp, B. F. Sherr, E. B. Sherr and J. J. Cole, LewisPublishers, Boca Raton.
Delmas, D., A. Herbland and S. Y. Maestrini (1992): Environ-mental conditions which lead to increase in cell density ofthe toxic dinoflagellates Dinophysis spp. in nutrient-rich andnutrient-poor waters of the French Atlantic coast. Mar. Ecol.Prog. Ser., 89, 53–61.
Fukuyo, Y. (1984): Okkirai Bay—with the occurrence ofdinoflagellates. p. 10–13. In Kaiyo-no-seibutsukatei, ed. byR. Marumo, Kouseisha Kouseikaku, Tokyo (in Japanese).
Fukuyo, Y. (1997): Dinophysis. p. 274–278. In Akasio-no-kagaku, 2nd ed., ed. by T. Okaichi, Kouseisha Kouseikaku,Tokyo (in Japanese).
Geider, R. J. and P. A. Gunter (1989): Evidence for the pres-ence of phycoerythrin in Dinophysis norvegica, a pinkdinoflagellate. Br. phycol. J., 24, 195–198.
Giacobbe, M. G., F. Oliva, R. La Ferla, A. Puglisi, E. Crisafiand G. Maimone (1995): Potentially toxic dinoflagellatesin Mediterranean waters (Sicily) and related hydrobiologicalconditions. Aquat. Microb. Ecol., 9, 63–68.
Hewes, C. D., B. G. Mitchell, T. A. Moisan, M. Vernet and F.M. A. Reid (1998): The phycobilin signatures of chloroplastsfrom three dinoflagellate species: a microanalytical studyof Dinophysis caudata , D. fortii and D. acuminata(Dinophysiales, Dinophyceae). J. Phycol., 34, 945–951.
Hoshiai, G., M. Chiba and T. Onodera (1996): Kaidokuseibun·yudoku plankton tou monitoring jigyo houkokusho(Report of monitoring project on shellfish toxins and harm-ful plankton). Kesennuma Miyagi Prefectural FisheriesExperimental Station, 37 pp. (in Japanese).
Igarashi, T. (1985): Sanriku coast. p. 71–81. In ToxicDinoflagellate—Implication in Shellfish Poisoning, Bull.Jap. Assoc. Sci. Fish., Vol. 56, ed. by Y. Fukuyo, KouseishaKouseikaku, Tokyo (in Japanese).
Igarashi, T. (1986): Occurrence of Dinophysis fortii, adinoflagellate responsible for diarrhetic shellfish poison-ing at Kesennuma Bay. Bull. Tohoku Reg. Fish. Res. Lab.,48, 137–144 (in Japanese).
Iizuka, S. (1979): Biological prediction method for red tide. p.394–400. In Kaiyo Kankyo Tyousahou, Oceanographic So-ciety of Japan, Kouseisha Kouseikaku, Tokyo (in Japanese).
Ishimaru, T., H. Inoue, Y. Fukuyo, T. Ogata and M. Kodama(1988): Culture of Dinophysis fortii and D. acuminata withthe cryptomonad, Plagioselmis sp. p. 19–20. In Mycotoxinsand Phycotoxins, Special issue No. 1, ed. by K. Aibara, S.Kumagai, K. Ohtsubo and T. Yoshizawa, Jap. Ass.Mycotoxicol., Tokyo.
Iwasaki, J. (1986): The mechanisms of mass occurrence ofDinophysis fortii along the coast of Ibaraki prefecture. Bull.Tohoku Reg. Fish. Res. Lab., 48, 125–136 (in Japanese).
Koike, K., K. Koike, M. Takagi, T. Ogata and T. Ishimaru(2000): Evidence of phagotrophy in Dinophysis fortii(Dinophysiales, Dinophyceae), a dinoflagellate that causesdiarrhetic shellfish poisoning (DSP). Phycological Res., 48,
121–124.Lasen, J. (1992): Endocytobiotic consortia with dinoflagellate
hosts. p. 427–442. In Algae and Symbioses: Plants, Ani-mals, Fungi, Viruses, Interactions Explored, ed. by W.Reisser, Biopress, Bristol.
Laval-Peuto, M. (1992): Plastidic protozoa. p. 471–499. In Al-gae and Symbioses: Plants, Animals, Fungi, Viruses, Inter-actions Explored, ed. by W. Reisser, Biopress, Bristol.
Lucas, I. A. N. and M. Vesk (1990): The fine structure of twophotosynthetic species of Dinophysis (Dinophysiales,Dinophyceae). J. Phycol., 26, 345–357.
Okazaki, M. (1990): Internal tidal waves and internal long pe-riod waves in the Sanriku coastal seas, eastern coast ofnorthern Japan. La mer, 28, 5–29.
Osaka, Y. (1985): Mutsu Bay. p. 59–70. In ToxicDinoflagellate—Implication in Shellfish Poisoning, Bull.Jap. Assoc. Sci. Fish., Vol. 56, ed. by Y. Fukuyo, KouseishaKouseikaku, Tokyo (in Japanese).
Otobe, H., M. Ohtsuki, K. Morita, M. Kurosawa, Y. Iwama andK. Kashiwazaki (1999): Results of oceanographic and me-teorological observations at Otsuchi Bay. Otsuchi Mar. Sci.,24, 58–72.
Peperzak, L., G. J. Snoeijer, R. Dijkema, W. W. C. Gieskes, J.Joordens, J. C. H. Peeters, C. Schol, E. G. Vrieling and W.Zevenboom (1996): Development of a Dinophysis acumi-nata bloom in the River Rine Plume (North Sea). p. 273–276. In Harmful and Toxic Algal Blooms, ed. by T.Yasumoto, Y. Oshima and Y. Fukuyo, IOC/UNESCO, Paris.
Reguera, B., I. Bravo and S. Fraga (1995): Autoecology andsome life history stages of Dinophysis acuta Ehrenberg. J.Plank. Res., 17, 999–1015.
Schnepf, E. and M. Elbrächter (1988): Cryptophycean-like dou-ble membrane-bound chloroplast in the dinoflagellate,Dinophysis Ehrenb.: evolutionary, phylogenetic and toxi-cological implications. Botanica Acta, 101, 196–203.
Schnepf, E. and M. Elbrächter (1992): Nutritional strategies indinoflagellates: a review with emphasis on cell biologicalaspects. Europ. J. Protistol., 28, 3–24.
Taylor, F. J. R., R. Haigh and T. F. Sutherland (1994):Phytoplankton ecology of Sechelt Inlet, a fjord system onthe British Columbia coast. II. Potentially harmful species.Mar. Ecol. Prog. Ser., 103, 151–164.
Valderrama, J. C. (1995): Methods of nutrient analysis. p. 251–268. In Manual on Harmful Marine Microalgae, IOC Manu-als and Guides No. 33, ed. by G. M. Hallegraeff, D. M.Anderson and A. D. Cembella, UNESCO, Paris.
Vesk, M., T. P. Dibbayawan and P. A. Vesk (1996): Immunogoldlocalization of phycoerythrin in chloroplasts of Dinophysisacuminata and D. fortii (Dinophysiales, Dinophyta).Phycologia, 35, 234–238.
Yasumoto, T., Y. Oshima, W. Sugawara, Y. Fukuyo, H. Oguri,T. Igarashi and N. Fujita (1980): Identification of Dinophysisfortii as the causative organism of diarrhetic shellfish poi-soning. Nippon Suisan Gakkaishi, 46, 1405–1411.