Embed Size (px)
Citation preview

HEAD & FACE MEDICINE
Feller et al. Head & Face Medicine 2014, 10:8http://www.head-face-med.com/content/10/1/8
REVIEW Open Access
Melanin: the biophysiology of oral melanocytesand physiological oral pigmentationLiviu Feller1,3*, Aubrey Masilana1, Razia AG Khammissa1, Mario Altini2, Yusuf Jadwat1 and Johan Lemmer1
Abstract
The presence of melanocytes in the oral epithelium is a well-established fact, but their physiological functions arenot well defined. Melanin provides protection from environmental stressors such as ultraviolet radiation andreactive oxygen species; and melanocytes function as stress-sensors having the capacity both to react to and toproduce a variety of microenvironmental cytokines and growth factors, modulating immune, inflammatory andantibacterial responses. Melanocytes also act as neuroendocrine cells producing local neurotransmitters includingacetylcholine, catecholamines and opioids, and hormones of the melanocortin system such as proopiomelanocortin,adrenocorticotropic hormone and α-melanocyte stimulating hormone, that participate in intracellular and inintercellular signalling pathways, thus contributing to tissue homeostasis.There is a wide range of normal variation in melanin pigmentation of the oral mucosa. In general, darker skinnedpersons more frequently have oral melanin pigmentation than light-skinned persons. Variations in oral physiologicalpigmentation are genetically determined unless associated with some underlying disease.In this article, we discuss some aspects of the biophysiology of oral melanocytes, of the functions of melanin, and ofphysiological oral pigmentation.
Keywords: Melanin, Oral melanocytes, Pigmentation, Melanocyte stem cell, Keratinocyte, Melanosome, Pigmenttransfer
IntroductionWhile under physiological conditions the number of mela-nocytes in the oral epithelium is the same regardless ofracial/ethnic origin [1], the colour of oral mucosa variesbetween persons and is determined by several factorsincluding the number and melanogenic activity of the me-lanocytes in the basal cell layer of the epithelium, differ-ences in number, size, and distribution of melanosomes,differences in the type of melanins, and the masking effectof heavily keratinized epithelium [2-4]. The variations inmelanin colourisation of the oral mucosa will be deter-mined against a background of the degree of vascula-rization of the tissues and by the level of haemoglobin inthe blood [2].Melanocytes containing melanin are present in the
basal cell layer of the epithelium even at those oral
* Correspondence: [email protected] of Periodontology and Oral Medicine, University of Limpopo(Medunsa Campus), Pretoria, South Africa3Department Periodontology and Oral Medicine, Box D26 School ofDentistry, Medunsa 0204, South AfricaFull list of author information is available at the end of the article
© 2014 Feller et al.; licensee BioMed Central LCommons Attribution License (http://creativecreproduction in any medium, provided the orDedication waiver (http://creativecommons.orunless otherwise stated.
mucosal sites that show no visible signs of melanin pig-mentation [5-7]. Oral melanocytes may or may not pro-duce melanin, but as in the case of the skin, the amountof melanin they produce is genetically determined [2,6].There are substantial variations in the degree of melaninpigmentation between persons of different racial/ethnicgroups and between persons of the same racial/ethnicgroup, and these variations are normal [2,8,9]. Phy-siological/racial melanin pigmentation of the oral mu-cosa is common in black persons [7], and is morefrequent in darker skinned whites (Caucasians) than inlighter skinned whites [10]. The oral mucosal melaninpigmentation may be patchy or uniform and most com-monly affects the gingiva [9].Melanocytes produce melanin in membrane-bound or-
ganelles termed melanosomes. Melanosomes have all theproteins and enzymes necessary for melanin biosynthesis,for maintaining the structure of the melanosome, and forthe maturation of the immature pre-melanosome into amature melanosome producing melanin [11,12]. As themelanosomes mature intracellularly, they are transported
td. This is an Open Access article distributed under the terms of the Creativeommons.org/licenses/by/2.0), which permits unrestricted use, distribution, andiginal work is properly credited. The Creative Commons Public Domaing/publicdomain/zero/1.0/) applies to the data made available in this article,

Feller et al. Head & Face Medicine 2014, 10:8 Page 2 of 7http://www.head-face-med.com/content/10/1/8
via microtubuli to the surfaces of the elongated dendritesof the melanocytes whence they are ultimately transferredto the keratinocytes in the ‘keratinocyte melanin unit’[11,12]. Within the keratinocytes, the melanin is preferen-tially localised within the nuclei forming protective bar-riers known as supranuclear ‘caps’ oriented in such a waythat they shield the nuclear DNA from ultraviolet (UV)radiation [13].Non-physiological alterations in melanin pigmentation
of the oral mucosa are related to genetic, metabolic, endo-crine, chemical or physical factors, to infective agents andto inflammatory or neoplastic processes [6,9]. It has beenreported that in about 30% of cases, oral mucosal mela-noma develops at sites of hyperpigmentation [2,14]. How-ever, the nature of the hyperpigmented pre-melanoma isobscure.Most of our knowledge about the regulation and biology
of oral melanocytes is derived from data obtained fromresearch on epidermal melanocytes which are histologi-cally and ultra-structurally similar [15,16]. However, whenextrapolating data from epidermal melanocytes, one needsto bear in mind that as a rule, oral mucosal melanocytesunder physiological circumstances are said to be less me-tabolically active [6]. Nevertheless, in some cases oralmelanocytes may be inherently metabolically active, andin other cases an increase in metabolic activity may be inresponse to environmental triggers such as hormones,inflammation or injury [6].The aim of this short review is to discuss some aspects
of the biology and physiology of oral melanocytes, thefunction of melanin and critically to evaluate the con-cept of intraoral physiological pigmentation.
The origin of oral melanocytesMelanocytes are melanin-producing cells originating fromthe neural crest [17]. During development melanocytestem cells migrate from the neural crest to the skin and tomucous membranes. Active melanocytes are present inthe stria vascularis of the cochlea, in the leptomeninges, inthe substantia nigra and locus coerulus of the brain, andin the heart, where they play a number of as yet ill-definedroles [13,18].Melanocyte stem cells have the capacity for self-renewal
and for differentiation and thus can maintain the popula-tion of mature melanocytes. In the epidermis, the melano-cyte stem cells reside in the bulge region of hair follicles[19,20], but the niche in which they reside in the oral mu-cosa is unknown.Epidermal melanocyte stem cells give rise to transient-
amplifying melanocyte precursors that settle without thestem cell niche where they differentiate into mature mela-nin producing melanocytes [19,21-24]. Stem cell factor(SCF) and its tyrosine kinase receptor C-kit signalling path-ways are critical for epidermal melanocyte development
during embryogenesis [17,20], notch signalling pathwaysare essential in the maintenance of adult melanocyte stemcells, and thus for melanocyte homeostasis [20], endothelin1 plays a role in the differentiation of melanocyte pre-cursors, and micropthalmia-associated transcription factor(MITF) with its cAMP response element play a critical rolein melanogenesis [25].In the skin, migration of precursor melanocytes from
the dermis to their final destination in the basal cell layerof the epithelium is mediated by c-Kit/SCF, endothelin 1and 3, hepatocyte growth factor (HGF) and basic fibro-blast growth factor (bFGF). These precursor dermal mela-nocytes, as they pass through the basement membrane,express E-cadherin which later facilitates intercellularcommunication with neighbouring keratinocytes in thebasal cell layer of the epithelium [1]. Sometimes, mela-nocyte precursors on their way to the epithelium may be-come arrested in the lamina propria/dermis, and as theyhave the capacity to produce melanin, if they are aggre-gated they will form nevi [26]. The functional activity ofmelanocytes, both those in the basal cell layer of the epi-thelium and those that may have become arrested in thelamina propria is influenced by signals from the neigh-bouring fibroblasts [13,27,28].
The keratinocyte-melanocyte unitMature melanocytes are elongated dendritic cells resid-ing in the basal cell layer of the epithelium. They containall the proteins required for melanin biosynthesis andfor the structural maturation of melanosomes [12], in-cluding tryosinase (TYR), tyrosinase-related proteins-1(TRP-1) and TRP-2, gp 100, and melanoma antigen re-cognizable by T lymphocytes (MART-1) [13,29-31].In the basal cell layer of the epithelium the ratio of
melanocytes to keratinocytes ranges from 1:10 [30] to1:15 [6]. Melanocytes and keratinocytes form epidermalmelanin units, each unit consists of one melanocyte anda group of about 36 neighbouring keratinocytes. Themelanosomes produced by melanocytes are disseminatedvia a network of melanocytic dendritic processes to thekeratinocytes of the epidermal melanin unit [25,32]. Itappears that keratinocytes have some control over theprocess of dendritic melanosome transfer since the capa-city of keratinocytes to phagocytose the melanosomes isinfluenced by the degree of activation of the protease-activated receptor 2 (PAR-2) on the surface of keratino-cytes [33].It has been suggested that through biological mediators
present in the melanosomes transferred to keratinocytes,melanocytes can influence the functional activities of kerati-nocytes [13], and keratinocytes via an array of paracrine-like biological mediators that they secrete, have the capacityto regulate melanocyte melanogenesis [13,25,34]. Thesemediators include α-melanocyte stimulating hormone

Feller et al. Head & Face Medicine 2014, 10:8 Page 3 of 7http://www.head-face-med.com/content/10/1/8
(α-MSH), adrenocorticotropin hormone (ACTH), β-endorphin, bFGF, endothelins, HGF and SCF [35].However, such a mechanistic description is an over-
simplification as the ratio of keratinocytes to mela-nocytes is different at different stages of growth anddevelopment, and in adulthood the ratio is variablebeing determined by biological mediators secreted in thelocal microenvironment [33]. Furthermore, the nume-rical density of melanocytes in the epithelium varies indifferent parts of the skin or oral mucosa, and betweenthe same skin or mucosa sites in different persons, re-gardless of their racial/ethnic origin [36]. It appears thatLangerhans cells in the epithelium, and fibroblasts in thesubepithelial connective tissue play an important rolein maintaining the functional activity of the epidermalmelanin unit [30,31,33].The adherence of melanocytes to keratinocytes is by
means of tight junctions where they co-express E-cadherincell adhesion molecules and by gap junctions [33]. E-cadherin supresses melanocyte proliferation, but a switchfrom E-cadherin to N-cadherin owing to metabolically ortraumatically triggered events in the microenvironmentobviates the melanocytes from growth suppression.Melanocytes expressing N-cadherin can freely proli-ferate, migrate and self-aggregate in nests, and exhibitlongevity [32].
The function of oral melanocytesThe functions of melanocytes are not fully understood,but it is clear that the melanin that they produce deter-mines the colour of skin, hair and eyes [37], and pro-vides protection from stressors such as UV radiation,reactive oxygen species (ROS) and free radicals in theenvironment. Melanins also have the capacity to seques-ter metal ions and to bind certain drugs and organicmolecules [32,38].The colour of skin and probably of any pigmented part
of the oral mucosa is genetically determined by thenumber and size of the melanosomes and the type ofmelanin (eumelanin, pheomelanin) that they produce.Environmental factors have only a modifying influenceon skin colour, though on an evolutionary scale this in-fluence can have a more profound effect. Melanosomesvary in size and contain both eumelanin and pheomela-nin. More, larger eumelanin impart a dark colour to theskin [33]. Small, few pheomelanin-containing melano-somes are associated with light skin, and the spectrumof size and number of melanosomes and the balance ofeumelanin to pheomelanin within the melanosomes, willdetermine all the other colour variations [32].As melanin synthesis is an oxygen dependent process,
paradoxically, it also generates ROS that may accumu-late in the melanocytes and cause DNA damage, and infact UV radiation exaggerates the production of ROS
during the biosynthesis of melanin and more particularlypheomelanin [39]. Thus, melanin possesses both antioxi-dant and ROS-dependent cytotoxic properties [18].Quinones and semiquinones which are intermediates
of melanogenesis are toxic or mutagenic with the po-tential to cause cytogenetic instability [40]; and L-dopa,another intermediate of melanogenesis, has the capacityto inhibit the production of proinflammatory cytokinesby T lymphocytes and monocytes, thus downregulatingimmune and inflammatory responses [38,40].Melanins produced by melanocytes residing in the basal
cell layer of the gingival epithelium have the capacity toneutralize ROS generated by dentogingival plaque-in-duced inflammation in the periodontal microenvironment[16]. Interestingly, a recent report shows that markers ofgingival inflammation are reduced in subjects with pig-mented gingiva compared to subjects with non-pigmentedgingiva, despite comparable dentogingival plaque levels inboth groups of subjects. However, one needs to bearin mind that the sulcular and junctional epithelium, incontrast to the keratinized epithelium usually does notharbour any melanocytes [16].Melanosomes contain lysosomal enzymes including
α-mannosidase, acid phosphatase, β-N acetylglycosamini-dase, β-galactosidase, and acid lipase that can degrade bac-teria [41]. Melanin itself can neutralise bacteria-derivedenzymes and toxins, and since it has strong binding pro-perties, it can also act as a physical barrier against mi-croorganisms [41]. Furthermore, melanocytes can act asantigen presenting cells, can stimulate T-cell proliferation,and can phagocytose microorganisms [32,41]. Thus, mela-nocytes and their products have the capacity to inhibitproliferation of bacterial and fungal microorganisms [41].As keratinocytes ascend through the cell layers of the
epithelium and are shed, their melanosomal membranesundergo degradation with release of melanin ‘dust’ whichbecomes entangled in the keratin filaments of the des-quamating surface cells. This melanin dust inactivatespathogenic chemicals, microbial toxins and other biologic-ally active molecules [18]. Therefore, melanocytes andmelanins may be viewed as an integral part of the innateimmune system with a role in neutralising the products ofsuperficial bacterial and fungal infective agents [41].It is possible that from an evolutionary point of view,
the primary role of melanocytes is not to produce mela-nin, for melanin does not confer a selective advantage tothe organisms, but that melanin production is only asecondary specialization so melanocytes must have othermore important functions to perform [13]. Melanocytesin skin, and perhaps in oral mucosa express genes en-coding corticotropin releasing factor (CRF), proopiome-lanocortin (POMC), ACTH, β endorphins, α-MSH andmelanocortin-1 receptor (MC1R). These elements of theskin melanocortin system have the capacity to neutralize
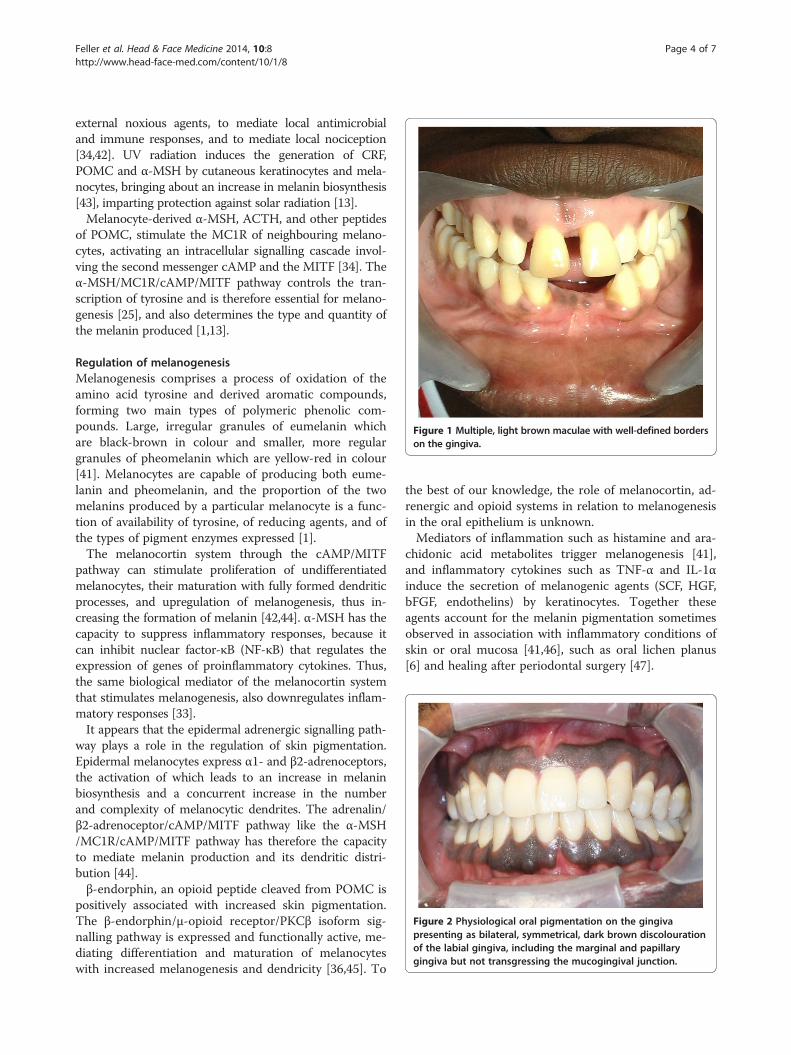
Figure 1 Multiple, light brown maculae with well-defined borderson the gingiva.
Figure 2 Physiological oral pigmentation on the gingivapresenting as bilateral, symmetrical, dark brown discolourationof the labial gingiva, including the marginal and papillarygingiva but not transgressing the mucogingival junction.
Feller et al. Head & Face Medicine 2014, 10:8 Page 4 of 7http://www.head-face-med.com/content/10/1/8
external noxious agents, to mediate local antimicrobialand immune responses, and to mediate local nociception[34,42]. UV radiation induces the generation of CRF,POMC and α-MSH by cutaneous keratinocytes and mela-nocytes, bringing about an increase in melanin biosynthesis[43], imparting protection against solar radiation [13].Melanocyte-derived α-MSH, ACTH, and other peptides
of POMC, stimulate the MC1R of neighbouring melano-cytes, activating an intracellular signalling cascade invol-ving the second messenger cAMP and the MITF [34]. Theα-MSH/MC1R/cAMP/MITF pathway controls the tran-scription of tyrosine and is therefore essential for melano-genesis [25], and also determines the type and quantity ofthe melanin produced [1,13].
Regulation of melanogenesisMelanogenesis comprises a process of oxidation of theamino acid tyrosine and derived aromatic compounds,forming two main types of polymeric phenolic com-pounds. Large, irregular granules of eumelanin whichare black-brown in colour and smaller, more regulargranules of pheomelanin which are yellow-red in colour[41]. Melanocytes are capable of producing both eume-lanin and pheomelanin, and the proportion of the twomelanins produced by a particular melanocyte is a func-tion of availability of tyrosine, of reducing agents, and ofthe types of pigment enzymes expressed [1].The melanocortin system through the cAMP/MITF
pathway can stimulate proliferation of undifferentiatedmelanocytes, their maturation with fully formed dendriticprocesses, and upregulation of melanogenesis, thus in-creasing the formation of melanin [42,44]. α-MSH has thecapacity to suppress inflammatory responses, because itcan inhibit nuclear factor-κB (NF-κB) that regulates theexpression of genes of proinflammatory cytokines. Thus,the same biological mediator of the melanocortin systemthat stimulates melanogenesis, also downregulates inflam-matory responses [33].It appears that the epidermal adrenergic signalling path-
way plays a role in the regulation of skin pigmentation.Epidermal melanocytes express α1- and β2-adrenoceptors,the activation of which leads to an increase in melaninbiosynthesis and a concurrent increase in the numberand complexity of melanocytic dendrites. The adrenalin/β2-adrenoceptor/cAMP/MITF pathway like the α-MSH/MC1R/cAMP/MITF pathway has therefore the capacityto mediate melanin production and its dendritic distri-bution [44].β-endorphin, an opioid peptide cleaved from POMC is
positively associated with increased skin pigmentation.The β-endorphin/μ-opioid receptor/PKCβ isoform sig-nalling pathway is expressed and functionally active, me-diating differentiation and maturation of melanocyteswith increased melanogenesis and dendricity [36,45]. To
the best of our knowledge, the role of melanocortin, ad-renergic and opioid systems in relation to melanogenesisin the oral epithelium is unknown.Mediators of inflammation such as histamine and ara-
chidonic acid metabolites trigger melanogenesis [41],and inflammatory cytokines such as TNF-α and IL-1αinduce the secretion of melanogenic agents (SCF, HGF,bFGF, endothelins) by keratinocytes. Together theseagents account for the melanin pigmentation sometimesobserved in association with inflammatory conditions ofskin or oral mucosa [41,46], such as oral lichen planus[6] and healing after periodontal surgery [47].
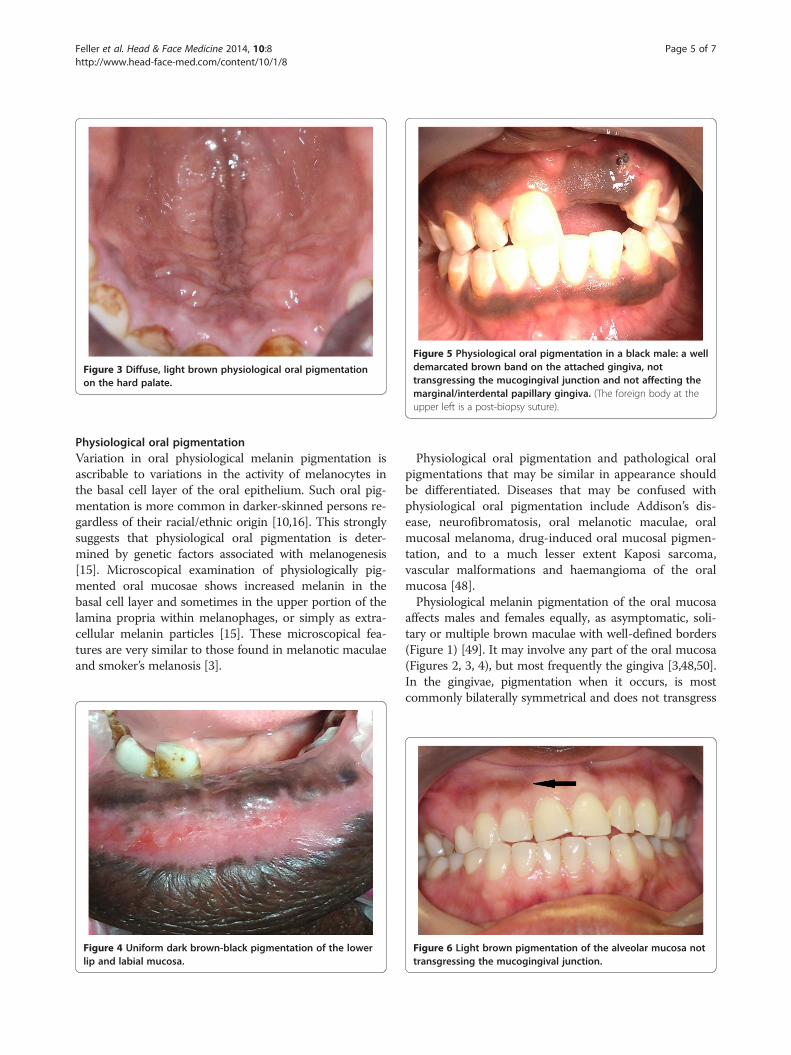
Figure 5 Physiological oral pigmentation in a black male: a welldemarcated brown band on the attached gingiva, nottransgressing the mucogingival junction and not affecting themarginal/interdental papillary gingiva. (The foreign body at theupper left is a post-biopsy suture).
Figure 3 Diffuse, light brown physiological oral pigmentationon the hard palate.
Feller et al. Head & Face Medicine 2014, 10:8 Page 5 of 7http://www.head-face-med.com/content/10/1/8
Physiological oral pigmentationVariation in oral physiological melanin pigmentation isascribable to variations in the activity of melanocytes inthe basal cell layer of the oral epithelium. Such oral pig-mentation is more common in darker-skinned persons re-gardless of their racial/ethnic origin [10,16]. This stronglysuggests that physiological oral pigmentation is deter-mined by genetic factors associated with melanogenesis[15]. Microscopical examination of physiologically pig-mented oral mucosae shows increased melanin in thebasal cell layer and sometimes in the upper portion of thelamina propria within melanophages, or simply as extra-cellular melanin particles [15]. These microscopical fea-tures are very similar to those found in melanotic maculaeand smoker’s melanosis [3].
Figure 4 Uniform dark brown-black pigmentation of the lowerlip and labial mucosa.
Physiological oral pigmentation and pathological oralpigmentations that may be similar in appearance shouldbe differentiated. Diseases that may be confused withphysiological oral pigmentation include Addison’s dis-ease, neurofibromatosis, oral melanotic maculae, oralmucosal melanoma, drug-induced oral mucosal pigmen-tation, and to a much lesser extent Kaposi sarcoma,vascular malformations and haemangioma of the oralmucosa [48].Physiological melanin pigmentation of the oral mucosa
affects males and females equally, as asymptomatic, soli-tary or multiple brown maculae with well-defined borders(Figure 1) [49]. It may involve any part of the oral mucosa(Figures 2, 3, 4), but most frequently the gingiva [3,48,50].In the gingivae, pigmentation when it occurs, is mostcommonly bilaterally symmetrical and does not transgress
Figure 6 Light brown pigmentation of the alveolar mucosa nottransgressing the mucogingival junction.

Feller et al. Head & Face Medicine 2014, 10:8 Page 6 of 7http://www.head-face-med.com/content/10/1/8
the mucogingival junction (Figure 2) [3,8,9,48,51], and insuch cases sometimes the free gingiva is not pigmented(Figure 5) [3,7,48]. Occasionally, the alveolar mucosa isaffected, and again the mucogingival junction is not trans-gressed (Figure 6). Oral pigmentation gradually appearsduring the first two decades of life [48] but affected sub-jects may be unaware of it [48,50].The extent and intensity of physiological oral pig-
mentation increases with age [52,53], concurrently withan increase in the number of oral melanocytes [15], per-haps because of the effects of potentially melanogenicstimuli such as recurrent minor functional injury, in-flammatory conditions, medication, or tobacco smokeare cumulative [3].The global frequency of physiological oral pigmenta-
tion in different racial/ethnic groups is unknown; but ithas been reported that about 95% of healthy black per-sons in the United States are affected; that pigmentationis more extensive in the anterior than in the posteriorportion of the mouth; and that when the gingiva isaffected the buccal/labial surfaces are more intenselypigmented than the lingual/palatal surfaces [7]. It hasbeen reported in one study from South Africa that oralpigmentation occurs in 98% of black persons [54].Eighty to a hundred percent of Australian Aborigines
have physiological oral pigmentation with equal genderdistribution [52]. By contrast Fry and Almeyda [55] havereported that only about 5% of white people in Englandhave physiological buccal mucosal pigmentation. Ingeneral, it appears that persons with more darkly pig-mented skin are more likely to have physiological oralpigmentation [7].
SummaryMelanocytes can adjust and respond to biological, physicaland chemical stimuli in their microenvironment, and canin turn generate physical and biochemical signals whichmay affect that microenvironment. Melanocytes have thecapacity to mediate antimicrobial and immune responsesand to act as neuroendocrine cells, and they producemelanin that provides protection from environmentalstressors such as UV radiation, reactive oxygen speciesand free radicals. Why certain persons are more affectedby physiological oral pigmentation is unknown, but cer-tainly it occurs predominantly in darker skinned persons.
ConsentWritten informed consent was obtained for the publica-tion of this report and any accompanying images.
Competing interestsThe authors declare that they have no competing interests.
Authors’ contributionsConcept of paper was devised by LF. LF, AM, RAGK, MA, YJ, and JL wrotethe manuscript. The manuscript was critically revised by LF, RAGK and JL.All authors read and approved the manuscript.
Author details1Department of Periodontology and Oral Medicine, University of Limpopo(Medunsa Campus), Pretoria, South Africa. 2Division of Anatomical Pathology,School of Pathology, University of the Witwatersrand, Johannesburg, SouthAfrica. 3Department Periodontology and Oral Medicine, Box D26 School ofDentistry, Medunsa 0204, South Africa.
Received: 9 January 2014 Accepted: 19 March 2014Published: 24 March 2014
References1. Lin JY, Fisher DE: Melanocyte biology and skin pigmentation. Nature 2007,
445:843–850.2. Pai A, Prasad S, Patil BA, Dyasanoor S, Hedge S: Oral pigmentation: case
report and review of malignant melanoma with flow charts for diagnosisand treatment. Gen Dent 2012, 60:410–416. quiz 417-418.
3. Meleti M, Vescovi P, Mooi WJ, van der Waal I: Pigmented lesions of theoral mucosa and perioral tissues: a flow-chart for the diagnosis andsome recommendations for the management. Oral Surg Oral Med OralPathol Oral Radiol Endod 2008, 105:606–616.
4. Squier CA, Kremer MJ: Biology of oral mucosa and esophagus. J NatlCancer Inst Monogr 2001, 29:7–15.
5. Barrett AW, Raja AM: The immunohistochemical identification of humanoral mucosal melanocytes. Arch Oral Biol 1997, 42:77–81.
6. Barrett AW, Scully C: Human oral mucosal melanocytes: a review. J OralPathol Med 1994, 23:97–103.
7. Dummett CO: Physiologic pigmentation of the oral and cutaneoustissues in the Negro. J Dent Res 1946, 25:421–432.
8. Chandra S, Keluskar V, Bagewadi A, Sah K: Extensive physiologic melaninpigmentation on the tongue: an unusual clinical presentation. ContempClin Dent 2010, 1:204–206.
9. McCarthy PL, Shlkar G: Distrubances in pigmentation. In Diseases of theOral Mucosa. Edited by McCarthy PL, Shlkar G. New York: McGraw-Hill;1964:228–235.
10. Amir E, Gorsky M, Buchner A, Sarnat H, Gat H: Physiologic pigmentation ofthe oral mucosa in Israeli children. Oral Surg Oral Med Oral Pathol 1991,71:396–398.
11. Chi A, Valencia JC, Hu ZZ, Watabe H, Yamaguchi H, Mangini NJ, Huang H,Canfield VA, Cheng KC, Yang F, Abe R, Yamagishi S, Shabanowitz J,Hearing VJ, Wu C, Appella E, Hunt DF: Proteomic and bioinformaticcharacterization of the biogenesis and function of melanosomes.J Proteome Res 2006, 5:3135–3144.
12. Marks MS, Seabra MC: The melanosome: membrane dynamics in blackand white. Nat Rev Mol Cell Biol 2001, 2:738–748.
13. Plonka PM, Passeron T, Brenner M, Tobin DJ, Shibahara S, Thomas A, Slominski A,Kadekaro AL, Hershkovitz D, Peters E, Nordlund JJ, Abdel-Malek Z, Takeda K,Paus R, Ortonne JP, Hearing VJ, Schallreuter KU:What are melanocytes reallydoing all day long…? Exp Dermatol 2009, 18:799–819.
14. Mihajlovic M, Vlajkovic S, Jovanovic P, Stefanovic V: Primary mucosalmelanomas: a comprehensive review. Int J Clin Exp Pathol 2012,5:739–753.
15. Eisen D: Disorders of pigmentation in the oral cavity. Clin Dermatol 2000,18:579–587.
16. Nilima S, Vandana KL: Melanin: a scavenger in gingival inflammation.Indian J Dent Res 2011, 22:38–43.
17. Thomas AJ, Erickson CA: The making of a melanocyte: the specification ofmelanoblasts from the neural crest. Pigment Cell Melanoma Res 2008,21:598–610.
18. Wood JM, Jimbow K, Boissy RE, Slominski A, Plonka PM, Slawinski J,Wortsman J, Tosk J: What’s the use of generating melanin? Exp Dermatol1999, 8:153–164.
19. Nishimura EK, Jordan SA, Oshima H, Yoshida H, Osawa M, Moriyama M,Jackson IJ, Barrandon Y, Miyachi Y, Nishikawa S: Dominant role of theniche in melanocyte stem-cell fate determination. Nature 2002,416:854–860.

Feller et al. Head & Face Medicine 2014, 10:8 Page 7 of 7http://www.head-face-med.com/content/10/1/8
20. Kumano K, Masuda S, Sata M, Saito T, Lee SY, Sakata-Yanagimoto M, Tomita T,Iwatsubo T, Natsugari H, Kurokawa M, Ogawa S, Chiba S: Both Notch1 andNotch2 contribute to the regulation of melanocyte homeostasis.Pigment Cell Melanoma Res 2008, 21:70–78.
21. Cramer SF: Stem cells for epidermal melanocytes–a challenge forstudents of dermatopathology. Am J Dermatopathol 2009, 31:331–341.
22. Aubin-Houzelstein G, Djian-Zaouche J, Bernex F, Gadin S, Delmas V, Larue L,Panthier JJ: Melanoblasts’ proper location and timed differentiationdepend on Notch/RBP-J signaling in postnatal hair follicles. J InvestDermatol 2008, 128:2686–2695.
23. Shah KV, Chien AJ, Yee C, Moon RT: CTLA-4 is a direct target of Wnt/beta-catenin signaling and is expressed in human melanoma tumors.J Invest Dermatol 2008, 128:2870–2879.
24. Carlson JA, Murphy M, Slominski A, Vincent WL: Evidence of skin fieldcancerization. In Filed Cancerization: Basic Science and Clinical Applications.Edited by Dakubo GD E. Ontario, Canada: Nova Science Publishers;2011:317–370.
25. Schallreuter KU, Kothari S, Chavan B, Spencer JD: Regulation ofmelanogenesis–controversies and new concepts. Exp Dermatol 2008,17:395–404.
26. Neville BW, Damm DD, Allen CM, Bouquot JE: Acquired melanocytic nevus.In Oral and Maxillofacial Pathology. 3rd edition. Edited by Dolan J. St Louis,Missouri: Saunders Elsevier; 2009:382.
27. Yamaguchi Y, Passeron T, Watabe H, Yasumoto K, Rouzaud F, Hoashi T,Hearing VJ: The effects of dickkopf 1 on gene expression and Wntsignaling by melanocytes: mechanisms underlying its suppression ofmelanocyte function and proliferation. J Invest Dermatol 2007,127:1217–1225.
28. Yamaguchi Y, Itami S, Watabe H, Yasumoto K, Abdel-Malek ZA, Kubo T,Rouzaud F, Tanemura A, Yoshikawa K, Hearing VJ: Mesenchymal-epithelialinteractions in the skin: increased expression of dickkopf1 by palmoplantarfibroblasts inhibits melanocyte growth and differentiation. J Cell Biol 2004,165:275–285.
29. Paino F, Ricci G, De Rosa A, D’Aquino R, Laino L, Pirozzi G, Tirino V, Papaccio G:Ecto-mesenchymal stem cells from dental pulp are committed todifferentiate into active melanocytes. Eur Cell Mater 2010, 20:295–305.
30. Yamaguchi Y, Brenner M, Hearing VJ: The regulation of skin pigmentation.J Biol Chem 2007, 282:27557–27561.
31. Passeron T, Coelho SG, Miyamura Y, Takahashi K, Hearing VJ:Immunohistochemistry and in situ hybridization in the study of humanskin melanocytes. Exp Dermatol 2007, 16:162–170.
32. Tolleson WH: Human melanocyte biology, toxicology, and pathology.J Environ Sci Health C Environ Carcinog Ecotoxicol Rev 2005, 23:105–161.
33. Nordlund JJ: The melanocyte and the epidermal melanin unit: anexpanded concept. Dermatol Clin 2007, 25:271–281. vii.
34. Slominski A, Wortsman J, Paus R, Elias PM, Tobin DJ, Feingold KR: Skin as anendocrine organ: implications for its function. Drug Discov Today Dis Mech2008, 5:137–144.
35. Hirobe T: Role of keratinocyte-derived factors involved in regulating theproliferation and differentiation of mammalian epidermal melanocytes.Pigment Cell Res 2005, 18:2–12.
36. Slominski A, Tobin DJ, Shibahara S, Wortsman J: Melanin pigmentation inmammalian skin and its hormonal regulation. Physiol Rev 2004,84:1155–1228.
37. Sturm RA: Molecular genetics of human pigmentation diversity. Hum MolGenet 2009, 18:R9–R17.
38. Plonka PM, Grabacka M: Melanin synthesis in microorganisms–biotechnological and medical aspects. Acta Biochim Pol 2006, 53:429–443.
39. Smit NP, van Nieuwpoort FA, Marrot L, Out C, Poorthuis B, van Pelt H,Meunier JR, Pavel S: Increased melanogenesis is a risk factor for oxidativeDNA damage–study on cultured melanocytes and atypical nevus cells.Photochem Photobiol 2008, 84:550–555.
40. Slominski A, Zbytek B, Slominski R: Inhibitors of melanogenesis increasetoxicity of cyclophosphamide and lymphocytes against melanoma cells.Int J Canc Suppl J Int Canc Suppl 2009, 124:1470–1477.
41. Mackintosh JA: The antimicrobial properties of melanocytes,melanosomes and melanin and the evolution of black skin. J Theor Biol2001, 211:101–113.
42. Peters A: The self-similarity of the melanocortin system. Endocrinology2005, 146:529–531.
43. Millington GW: Proopiomelanocortin (POMC): the cutaneous roles of itsmelanocortin products and receptors. Clin Exp Dermatol 2006, 31:407–412.
44. Grando SA, Pittelkow MR, Schallreuter KU: Adrenergic and cholinergiccontrol in the biology of epidermis: physiological and clinicalsignificance. J Invest Dermatol 2006, 123:1948–1965.
45. Kauser S, Schallreuter KU, Thody AJ, Gummer C, Tobin DJ: Regulation ofhuman epidermal melanocyte biology by beta-endorphin. J InvestDermatol 2003, 120:1073–1080.
46. Satomura K, Tokuyama R, Yamasaki Y, Yuasa T, Tatehara S, Ishimaru N,Hayashi Y, Nagayama M: Possible involvement of stem cell factor andendothelin-1 in the emergence of pigmented squamous cell carcinomain oral mucosa. J Oral Pathol Med 2007, 36:621–624.
47. Perlmutter S, Tal H: Repigmentation of the gingiva following surgicalinjury. J Periodontol 1986, 57:48–50.
48. Kauzman A, Pavone M, Blanas N, Bradley G: Pigmented lesions of the oralcavity: review, differential diagnosis, and case presentations. J Can DentAssoc 2004, 70:682–683.
49. Caldeira PC, de Sousa SF, Gomez RS, Silva TA: Diffuse pigmentation of theoral mucosa. Oral Surg Oral Med Oral Pathol Oral Radiol Endod 2010,110:550–553.
50. Lerman MA, Karimbux N, Guze KA, Woo SB: Pigmentation of the hardpalate. Oral Surg Oral Med Oral Pathol Oral Radiol Endod 2009, 107:8–12.
51. Gaeta GM, Satriano RA, Baroni A: Oral pigmented lesions. Clin Dermatol2002, 20:286–288.
52. Brown T: Oral pigmentation in the Aborigines of Kalumburu, North-WestAustralia. Arch Oral Biol 1964, 9:555–564.
53. Dummett CO, Barens G: Oromucosal pigmentation: an updated literaryreview. J Periodontol 1971, 42:726–736.
54. van Wyk CW: Mouth pigmentation patterns in a group of healthy SouthAfrican Bantu. S Afr Med J 1970, 44:177–180.
55. Fry L, Almeyda JR: The incidence of buccal pigmentation in caucasoidsand negroids in Britain. Br J Dermatol 1968, 80:244–247.
doi:10.1186/1746-160X-10-8Cite this article as: Feller et al.: Melanin: the biophysiology of oralmelanocytes and physiological oral pigmentation. Head & Face Medicine2014 10:8.
Submit your next manuscript to BioMed Centraland take full advantage of:
• Convenient online submission
• Thorough peer review
• No space constraints or color figure charges
• Immediate publication on acceptance
• Inclusion in PubMed, CAS, Scopus and Google Scholar
• Research which is freely available for redistribution
Submit your manuscript at www.biomedcentral.com/submit





























