Embed Size (px)
Citation preview

RNA primer–primase complexes serve as the signal forpolymerase recycling and Okazaki fragment initiationin T4 phage DNA replicationMichelle M. Spieringa, Philip Hanoiana, Swathi Gannavarama, and Stephen J. Benkovica,1
aDepartment of Chemistry, The Pennsylvania State University, University Park, PA 16802
Edited by Michael E. O’Donnell, Rockefeller University, New York, NY, and approved April 24, 2017 (received for review December 13, 2016)
The opposite strand polarity of duplex DNA necessitates that theleading strand is replicated continuously whereas the lagging strandis replicated in discrete segments known as Okazaki fragments. Thelagging-strand polymerase sometimes recycles to begin the synthesisof a new Okazaki fragment before finishing the previous fragment,creating a gap between the Okazaki fragments. The mechanism andsignal that initiate this behavior—that is, the signaling mechanism—
have not been definitively identified. We examined the role of RNAprimer–primase complexes left on the lagging ssDNA from primersynthesis in initiating early lagging-strand polymerase recycling. Weshow for the T4 bacteriophage DNA replication system that primer–primase complexes have a residence time similar to the timescaleof Okazaki fragment synthesis and the ability to block a holoenzymesynthesizing DNA and stimulate the dissociation of the holoenzymeto trigger polymerase recycling. The collision with primer–primasecomplexes triggering the early termination of Okazaki fragment syn-thesis has distinct advantages over those previously proposed be-cause this signal requires no transmission to the lagging-strandpolymerase through protein or DNA interactions, the mechanismfor rapid dissociation of the holoenzyme is always collision, and nounique characteristics need to be assigned to either identical poly-merase in the replisome. We have modeled repeated cycles of Oka-zaki fragment initiation using a collision with a completed Okazakifragment or primer–primase complexes as the recycling mechanism.The results reproduce experimental data, providing insights intoevents related to Okazaki fragment initiation and the overall func-tioning of DNA replisomes.
DNA replication | T4 bacteriophage | Okazaki fragment initiation |poymerase recycling | stochastic modeling
During DNA replication, the opposite polarity of the two DNAstrands necessitates that the leading DNA strand is copied
continuously whereas the lagging DNA strand is copied in discretesegments known as Okazaki fragments. The initiation of each newOkazaki fragment involves the release of the current Okazakifragment by the lagging-strand holoenzyme, the synthesis of aprimer by primase, and the handoff of the primer to the recycledlagging-strand polymerase. Two mechanisms have been proposedto describe how and when these multiple events of Okazaki frag-ment initiation occur to maintain the coordinated replication of theleading and lagging DNA strands within a replisome, and evidencesuggests that both mechanisms operate in the replisomes studiedthus far [T4 (1), T7 (2), and Escherichia coli (3)]. In the collisionmechanism, the lagging-strand holoenzyme is triggered to recyclewhenever it collides with the previous Okazaki fragment (4, 5),possibly due to the absence of ssDNA template downstream of thepolymerase (6, 7). In the signaling mechanism, the lagging-strandholoenzyme is triggered to stop the synthesis of the current Oka-zaki fragment short of the previous fragment, leaving a gap. Thisearly recycling of the polymerase is believed to result from one ormore events related to repetitive lagging-strand DNA synthesis,specifically the association of primase with the replisome (3, 8, 9),crosstalk between primase subunits within the replisome (10),
primer synthesis (11), the loading of a clamp onto a primer by aclamp loader (1, 12), the release of a priming loop (12), or thetension-induced dissociation of the lagging-strand holoenzyme(13). However, the mechanism and the trigger underlying the sig-naling mechanism have not been definitively identified.The primase from T4 bacteriophage uses two mechanisms to
synthesize RNA primers (14). The first mechanism involves thedisassembly of a primase subunit from the replisome with its newlysynthesized RNA primer; the result is the formation of a primer–primase complex on the lagging DNA strand. In the secondmechanism, a small ssDNA loop is formed between the helicaseand primase within the replisome; the result is that primase remainswith the replisome, and the naked RNA primer is available for theclamp loader to load a clamp protein. The percentage of primersused for Okazaki fragment synthesis compared with the totalnumber of primers synthesized is low; in ensemble rolling circlestudies on the T4 system, only between 20% and 40% are used toinitiate Okazaki fragments (1). Therefore, many unused primerscould build up ahead of the holoenzyme on the lagging DNAstrand in the form of primer–primase complexes. This study in-vestigates the possibility that these primer–primase complexes serveas blocks on the lagging strand and correspond to the “signal” thatcauses the lagging-strand polymerase to recycle.
Results and DiscussionModel System. In these studies, it was necessary to use a small DNAsubstrate modeling the essence of the much larger lagging strand
Significance
The lagging-strand polymerase sometimes begins the synthesisof a new Okazaki fragment prior to finishing the previousfragment, creating a gap between the Okazaki fragments. Themechanism and signal that initiate this behavior have not beendefinitively identified. We demonstrate that primer–primasecomplexes left on the lagging ssDNA from primer synthesis havea residence time similar to the timescale of Okazaki fragmentsynthesis and the ability to block holoenzymes synthesizing DNAand stimulate the dissociation of the holoenzyme, triggeringpolymerase recycling. The collision with primer–primase com-plexes triggering early polymerase recycling has distinct advan-tages over previously proposed “signals.” Modeling of repeatedcycles of Okazaki fragment initiation reproduced experimentaldata, providing insights into Okazaki fragment initiation andoverall functioning of DNA replisomes.
Author contributions: M.M.S., S.G., and S.J.B. designed research; M.M.S. and S.G. per-formed research; M.M.S. and P.H. designed the stochastic modeling; P.H. wrote the codefor the stochastic modeling; M.M.S., P.H., and S.J.B. analyzed data; and M.M.S., P.H., andS.J.B. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1620459114/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1620459114 PNAS Early Edition | 1 of 6
BIOCH
EMISTR
Y

(Fig. S1). The substrate had template DNA with a 5′-GCT-3′priming recognition site and an annealed DNA oligonucleotide toserve as the Okazaki fragment being extended by the holoenzyme.The ends of the substrate were blocked with biotin–streptavidin toprevent the holoenzyme from sliding off the ends of the DNA,which would impact the measured rate of holoenzyme dissociation.Either an annealed DNA oligonucleotide to mimic the previousOkazaki fragment in the collision mechanism or a primer–primasecomplex to evaluate its capacity to initiate premature recycling inthe signaling mechanism were bound downstream of the holoen-zyme on this substrate. Due to the stochastic nature of primersynthesis by the T4 primase at priming recognition sites and theextremely low priming rate in the absence of helicase, an exoge-nous primer was used in these experiments instead of a primersynthesized in situ by primase. We used T7 RNA polymerase andtranscription runoff to generate the physiological 5-mer RNA primerincluding its 5′-triphosphate moiety of the T4 primase (Fig. S2). Herewe designate pppRNA and HO-RNA as this 5-mer RNA primerwith a 5′-triphosphate group and 5′-hydroxyl group, respectively.
Residence Time of pppRNA–Primase Complexes on ssDNA. Primer–primase complexes must have a residence time on the laggingstrand similar to the timescale of Okazaki fragment synthesis toblock the holoenzyme during the synthesis of an Okazaki fragmentand lead to its recycling. The fluorescence of a dye on the DNAsubstrate positioned 3 nt downstream from the priming recognitionsite (i.e., the primer–primase complex binding site) was found to besensitive to the presence or absence of a primer–primase complexon the DNA (Fig. S1A). This environmental sensitivity of the flu-orophore allowed us to study the kinetics of primer–primasecomplex dissociation from ssDNA by monitoring changes in thefluorescence of the dye as a function of time.We preassembled in a stopped-flow instrument the primer–
primase complex on the fluorescently labeled substrate and mixedit with unlabeled ssDNA trap to prevent rebinding. Extended timecourses of the fluorescence signal displayed three distinct phases(Fig. S3A). Two of these phases displayed decreases in the fluo-rescence signal as a function of time, and control experimentsdemonstrated these phases to be related to photobleaching of thedye (Fig. S3B) and fluorescence quenching of the DNA substratewhen mixed with a large excess of ssDNA trap (Fig. S3 C and D).The third phase, which was only observed in the presence of pri-mase or primer–primase complexes, displayed an increase in thefluorescence signal over time and was assigned to the dissociationof primase or primer–primase complexes from the fluorescentlylabeled substrate.Kinetic traces displaying only the relevant exponential term for
the dissociation phase using preformed complexes of primase alone,DNA primer–primase, or RNA primer lacking a 5′ triphosphatemoiety (HO-RNA)–primase bound to the fluorescently labeledsubstrate show very rapid increases in fluorescence (Fig. 1A) with anaverage rate constant of kdissociation = 7.3 ± 0.4 s−1 (Table S1). Theseresults indicate that the DNA or HO-RNA exogenous primers donot bind to primase subunits with high affinity and suggest that theobserved changes in fluorescence reflect primase protein alonedissociating from the fluorescently labeled substrate in each of theseexperiments. The correspondingly short half-life of 0.095 ± 0.005 sindicates that primase protein alone on the lagging ssDNA cannotserve to initiate lagging-strand holoenzyme recycling.In contrast, a physiological pppRNA primer formed a stable
complex with primase subunits on the fluorescently labeled sub-strate (Fig. 1A), demonstrating the importance of the 5′-tri-phosphate moiety in the formation of a pppRNA–primasecomplex. The dissociation rate of a pppRNA–primase complexfrom the DNA substrate was constant, kdissociation = 0.028 ±0.001 s−1 (t1/2 = 24.8 ± 0.9 s) (Table S1), whereas the amplitude ofthe fluorescence increase was dependent on the concentration ofthe pppRNA–primase complex formed. Kinetic traces displaying
only the relevant exponential term for the dissociation phase showan increase in amplitude up to a 1:1 ratio of pppRNA–primasecomplex to DNA substrate (Fig. 1B and Fig. S3E), suggesting astoichiometry of one primase subunit in every pppRNA–primasecomplex, which dissociates in a first-order kinetic process.Previously, we measured the lifetime of in situ pppRNA–primase
complexes on ssDNA to be 4.2 ± 0.3 s while under pressure from areannealing DNA hairpin in a magnetic tweezers assay (14), and weexpect an even longer lifetime for the complex in the absence of areannealing DNA hairpin. Our measured half-life of 24.8 ± 0.9 s,equal to a lifetime of 36 ± 1 s, for the pppRNA–primase complexon ssDNA is in good agreement with that expectation. At an av-erage nucleotide incorporation rate of 150 nt/s for the lagging-strand holoenzyme, it would take 6–13 s to synthesize an Okazakifragment of 1,000–2,000 bp in length. Therefore, a pppRNA–pri-mase complex with a half-life of 24.8 ± 0.9 s on ssDNA would besufficiently stable to create blocks on the lagging strand for a ho-loenzyme synthesizing Okazaki fragments.
pppRNA–Primase Complexes Block DNA Synthesis by Holoenzymes.For a primer–primase complex to cause the holoenzyme to re-cycle, the complex must be capable of blocking the progress of a
Fig. 1. Stability of pppRNA–primase complex to serve as a signal. Traces dis-play only the relevant exponential term for the primer–primase complex dis-sociation phase extracted from the entire time traces that include phasesrelated to fluorescence quenching due to mixing with ssDNA trap and photo-bleaching. (A) Time trace of pppRNA–primase complex (black trace) dissociationdisplays a sufficient half-life to serve as a signal for holoenzyme recycling. (In-set) Time traces of primase alone (red trace), DNA primer–primase (blue trace),and HO-RNA–primase (green trace) complex dissociation display a short half-life. (B) Time traces of pppRNA–primase complex dissociation over a series ofcomplex concentrations demonstrate a stoichiometery of one primer and pri-mase subunit per complex.
2 of 6 | www.pnas.org/cgi/doi/10.1073/pnas.1620459114 Spiering et al.
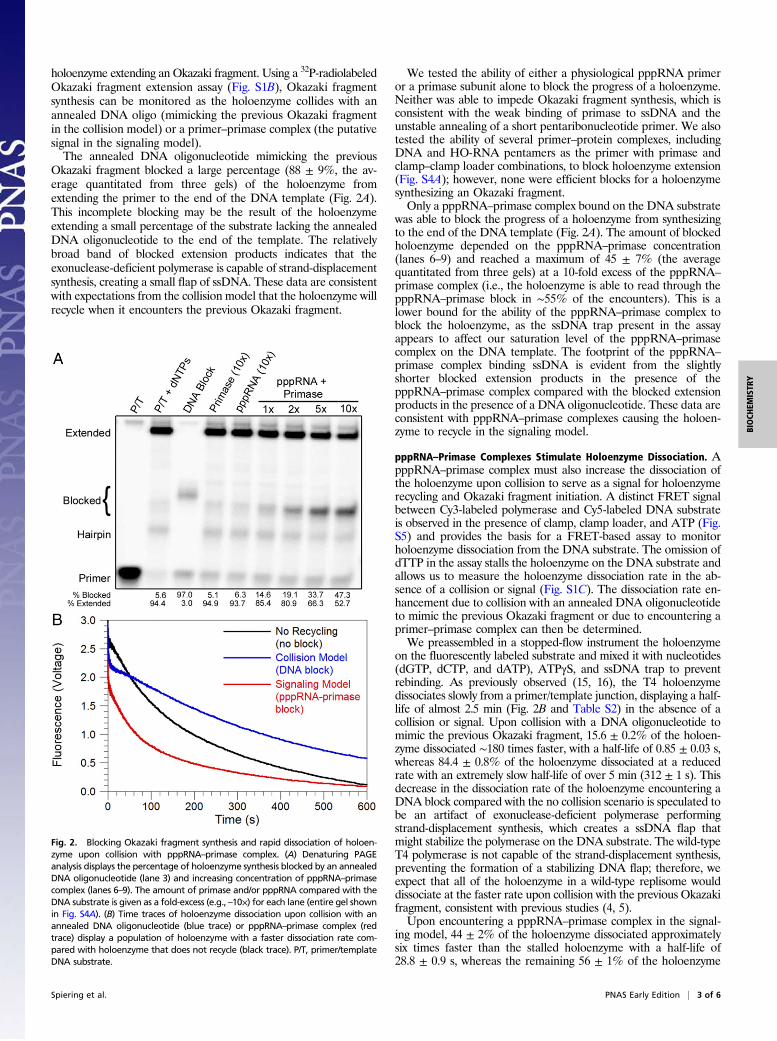
holoenzyme extending an Okazaki fragment. Using a 32P-radiolabeledOkazaki fragment extension assay (Fig. S1B), Okazaki fragmentsynthesis can be monitored as the holoenzyme collides with anannealed DNA oligo (mimicking the previous Okazaki fragmentin the collision model) or a primer–primase complex (the putativesignal in the signaling model).The annealed DNA oligonucleotide mimicking the previous
Okazaki fragment blocked a large percentage (88 ± 9%, the av-erage quantitated from three gels) of the holoenzyme fromextending the primer to the end of the DNA template (Fig. 2A).This incomplete blocking may be the result of the holoenzymeextending a small percentage of the substrate lacking the annealedDNA oligonucleotide to the end of the template. The relativelybroad band of blocked extension products indicates that theexonuclease-deficient polymerase is capable of strand-displacementsynthesis, creating a small flap of ssDNA. These data are consistentwith expectations from the collision model that the holoenzyme willrecycle when it encounters the previous Okazaki fragment.
We tested the ability of either a physiological pppRNA primeror a primase subunit alone to block the progress of a holoenzyme.Neither was able to impede Okazaki fragment synthesis, which isconsistent with the weak binding of primase to ssDNA and theunstable annealing of a short pentaribonucleotide primer. We alsotested the ability of several primer–protein complexes, includingDNA and HO-RNA pentamers as the primer with primase andclamp–clamp loader combinations, to block holoenzyme extension(Fig. S4A); however, none were efficient blocks for a holoenzymesynthesizing an Okazaki fragment.Only a pppRNA–primase complex bound on the DNA substrate
was able to block the progress of a holoenzyme from synthesizingto the end of the DNA template (Fig. 2A). The amount of blockedholoenzyme depended on the pppRNA–primase concentration(lanes 6–9) and reached a maximum of 45 ± 7% (the averagequantitated from three gels) at a 10-fold excess of the pppRNA–
primase complex (i.e., the holoenzyme is able to read through thepppRNA–primase block in ∼55% of the encounters). This is alower bound for the ability of the pppRNA–primase complex toblock the holoenzyme, as the ssDNA trap present in the assayappears to affect our saturation level of the pppRNA–primasecomplex on the DNA template. The footprint of the pppRNA–
primase complex binding ssDNA is evident from the slightlyshorter blocked extension products in the presence of thepppRNA–primase complex compared with the blocked extensionproducts in the presence of a DNA oligonucleotide. These data areconsistent with pppRNA–primase complexes causing the holoen-zyme to recycle in the signaling model.
pppRNA–Primase Complexes Stimulate Holoenzyme Dissociation. ApppRNA–primase complex must also increase the dissociation ofthe holoenzyme upon collision to serve as a signal for holoenzymerecycling and Okazaki fragment initiation. A distinct FRET signalbetween Cy3-labeled polymerase and Cy5-labeled DNA substrateis observed in the presence of clamp, clamp loader, and ATP (Fig.S5) and provides the basis for a FRET-based assay to monitorholoenzyme dissociation from the DNA substrate. The omission ofdTTP in the assay stalls the holoenzyme on the DNA substrate andallows us to measure the holoenzyme dissociation rate in the ab-sence of a collision or signal (Fig. S1C). The dissociation rate en-hancement due to collision with an annealed DNA oligonucleotideto mimic the previous Okazaki fragment or due to encountering aprimer–primase complex can then be determined.We preassembled in a stopped-flow instrument the holoenzyme
on the fluorescently labeled substrate and mixed it with nucleotides(dGTP, dCTP, and dATP), ATPγS, and ssDNA trap to preventrebinding. As previously observed (15, 16), the T4 holoenzymedissociates slowly from a primer/template junction, displaying a half-life of almost 2.5 min (Fig. 2B and Table S2) in the absence of acollision or signal. Upon collision with a DNA oligonucleotide tomimic the previous Okazaki fragment, 15.6 ± 0.2% of the holoen-zyme dissociated ∼180 times faster, with a half-life of 0.85 ± 0.03 s,whereas 84.4 ± 0.8% of the holoenzyme dissociated at a reducedrate with an extremely slow half-life of over 5 min (312 ± 1 s). Thisdecrease in the dissociation rate of the holoenzyme encountering aDNA block compared with the no collision scenario is speculated tobe an artifact of exonuclease-deficient polymerase performingstrand-displacement synthesis, which creates a ssDNA flap thatmight stabilize the polymerase on the DNA substrate. The wild-typeT4 polymerase is not capable of the strand-displacement synthesis,preventing the formation of a stabilizing DNA flap; therefore, weexpect that all of the holoenzyme in a wild-type replisome woulddissociate at the faster rate upon collision with the previous Okazakifragment, consistent with previous studies (4, 5).Upon encountering a pppRNA–primase complex in the signal-
ing model, 44 ± 2% of the holoenzyme dissociated approximatelysix times faster than the stalled holoenzyme with a half-life of28.8 ± 0.9 s, whereas the remaining 56 ± 1% of the holoenzyme
Fig. 2. Blocking Okazaki fragment synthesis and rapid dissociation of holoen-zyme upon collision with pppRNA–primase complex. (A) Denaturing PAGEanalysis displays the percentage of holoenzyme synthesis blocked by an annealedDNA oligonucleotide (lane 3) and increasing concentration of pppRNA–primasecomplex (lanes 6–9). The amount of primase and/or pppRNA compared with theDNA substrate is given as a fold-excess (e.g., –10×) for each lane (entire gel shownin Fig. S4A). (B) Time traces of holoenzyme dissociation upon collision with anannealed DNA oligonucleotide (blue trace) or pppRNA–primase complex (redtrace) display a population of holoenzyme with a faster dissociation rate com-pared with holoenzyme that does not recycle (black trace). P/T, primer/templateDNA substrate.
Spiering et al. PNAS Early Edition | 3 of 6
BIOCH
EMISTR
Y
dissociated with a half-life of 2.87 min (172 ± 9 s). The formerpopulation of holoenzyme (∼45%) is the same percentage of ho-loenzyme that was blocked by a pppRNA–primase complex in theOkazaki fragment extension assays (45 ± 7%). These data suggestthat this population of holoenzyme has been blocked by thepppRNA–primase complex and dissociated from the substrate.The dissociation rate of the latter ∼56% of the holoenzyme issimilar to the rate observed in the stalled holoenzyme case and isthe same percentage of holoenzyme that extended a primer to theend of the template in the Okazaki fragment extension assayscontaining pppRNA–primase complex (55 ± 7%). Taken together,these data suggest that this population of holoenzyme bypassedthe pppRNA–primase complex. In the context of a completeT4 replisome, holoenzyme dissociation due to collision with apppRNA–primase complex would be expected to entail sub-sequent recycling and initiation of a new Okazaki fragment—thatis, the signaling model. Although the dissociation rate of a holo-enzyme encountering a pppRNA–primase complex does not matchthe dissociation rate of a holoenzyme encountering an Okazakifragment (0.0241 ± 0.0007 s−1 vs. 0.82 ± 0.03 s−1) and appears tobe too slow to support coupled leading- and lagging-strand syn-thesis, other proteins and/or protein–protein interactions in thereplisome, especially the dimerization between the leading- andlagging-strand polymerases (17), may accelerate the lagging-strandpolymerase dissociation rate to a physiologically relevant level ina complete replisome.
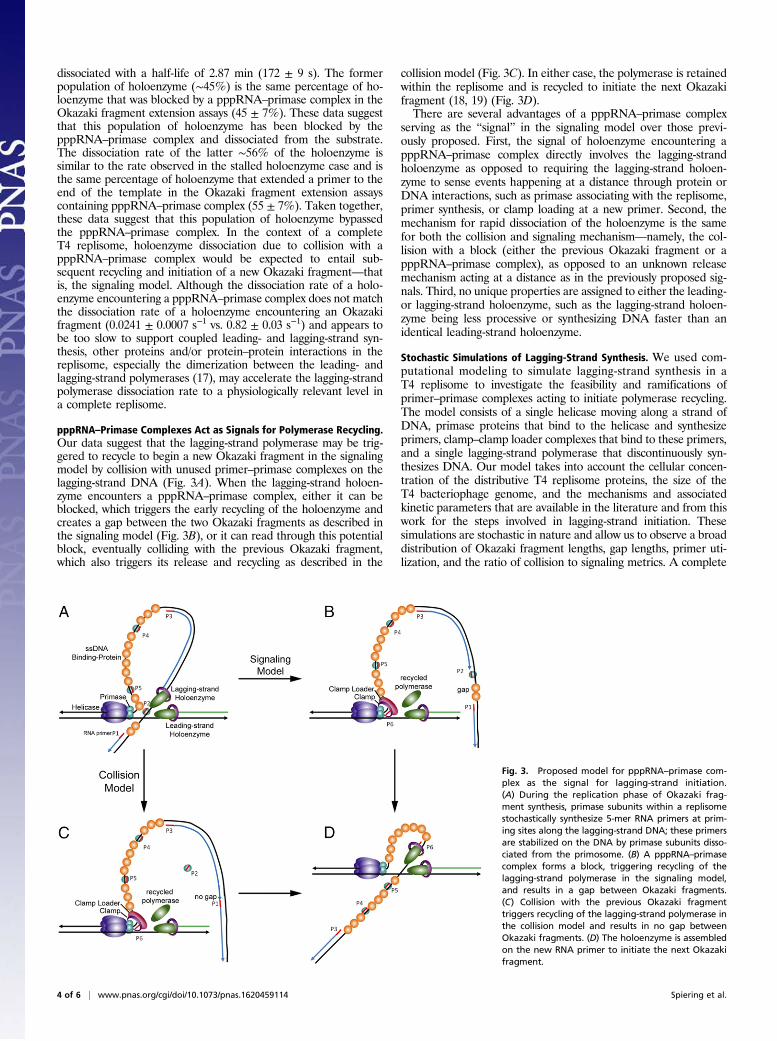
pppRNA–Primase Complexes Act as Signals for Polymerase Recycling.Our data suggest that the lagging-strand polymerase may be trig-gered to recycle to begin a new Okazaki fragment in the signalingmodel by collision with unused primer–primase complexes on thelagging-strand DNA (Fig. 3A). When the lagging-strand holoen-zyme encounters a pppRNA–primase complex, either it can beblocked, which triggers the early recycling of the holoenzyme andcreates a gap between the two Okazaki fragments as described inthe signaling model (Fig. 3B), or it can read through this potentialblock, eventually colliding with the previous Okazaki fragment,which also triggers its release and recycling as described in the
collision model (Fig. 3C). In either case, the polymerase is retainedwithin the replisome and is recycled to initiate the next Okazakifragment (18, 19) (Fig. 3D).There are several advantages of a pppRNA–primase complex
serving as the “signal” in the signaling model over those previ-ously proposed. First, the signal of holoenzyme encountering apppRNA–primase complex directly involves the lagging-strandholoenzyme as opposed to requiring the lagging-strand holoen-zyme to sense events happening at a distance through protein orDNA interactions, such as primase associating with the replisome,primer synthesis, or clamp loading at a new primer. Second, themechanism for rapid dissociation of the holoenzyme is the samefor both the collision and signaling mechanism—namely, the col-lision with a block (either the previous Okazaki fragment or apppRNA–primase complex), as opposed to an unknown releasemechanism acting at a distance as in the previously proposed sig-nals. Third, no unique properties are assigned to either the leading-or lagging-strand holoenzyme, such as the lagging-strand holoen-zyme being less processive or synthesizing DNA faster than anidentical leading-strand holoenzyme.
Stochastic Simulations of Lagging-Strand Synthesis. We used com-putational modeling to simulate lagging-strand synthesis in aT4 replisome to investigate the feasibility and ramifications ofprimer–primase complexes acting to initiate polymerase recycling.The model consists of a single helicase moving along a strand ofDNA, primase proteins that bind to the helicase and synthesizeprimers, clamp–clamp loader complexes that bind to these primers,and a single lagging-strand polymerase that discontinuously syn-thesizes DNA. Our model takes into account the cellular concen-tration of the distributive T4 replisome proteins, the size of theT4 bacteriophage genome, and the mechanisms and associatedkinetic parameters that are available in the literature and from thiswork for the steps involved in lagging-strand initiation. Thesesimulations are stochastic in nature and allow us to observe a broaddistribution of Okazaki fragment lengths, gap lengths, primer uti-lization, and the ratio of collision to signaling metrics. A complete
Fig. 3. Proposed model for pppRNA–primase com-plex as the signal for lagging-strand initiation.(A) During the replication phase of Okazaki frag-ment synthesis, primase subunits within a replisomestochastically synthesize 5-mer RNA primers at prim-ing sites along the lagging-strand DNA; these primersare stabilized on the DNA by primase subunits disso-ciated from the primosome. (B) A pppRNA–primasecomplex forms a block, triggering recycling of thelagging-strand polymerase in the signaling model,and results in a gap between Okazaki fragments.(C) Collision with the previous Okazaki fragmenttriggers recycling of the lagging-strand polymerase inthe collision model and results in no gap betweenOkazaki fragments. (D) The holoenzyme is assembledon the new RNA primer to initiate the next Okazakifragment.
4 of 6 | www.pnas.org/cgi/doi/10.1073/pnas.1620459114 Spiering et al.
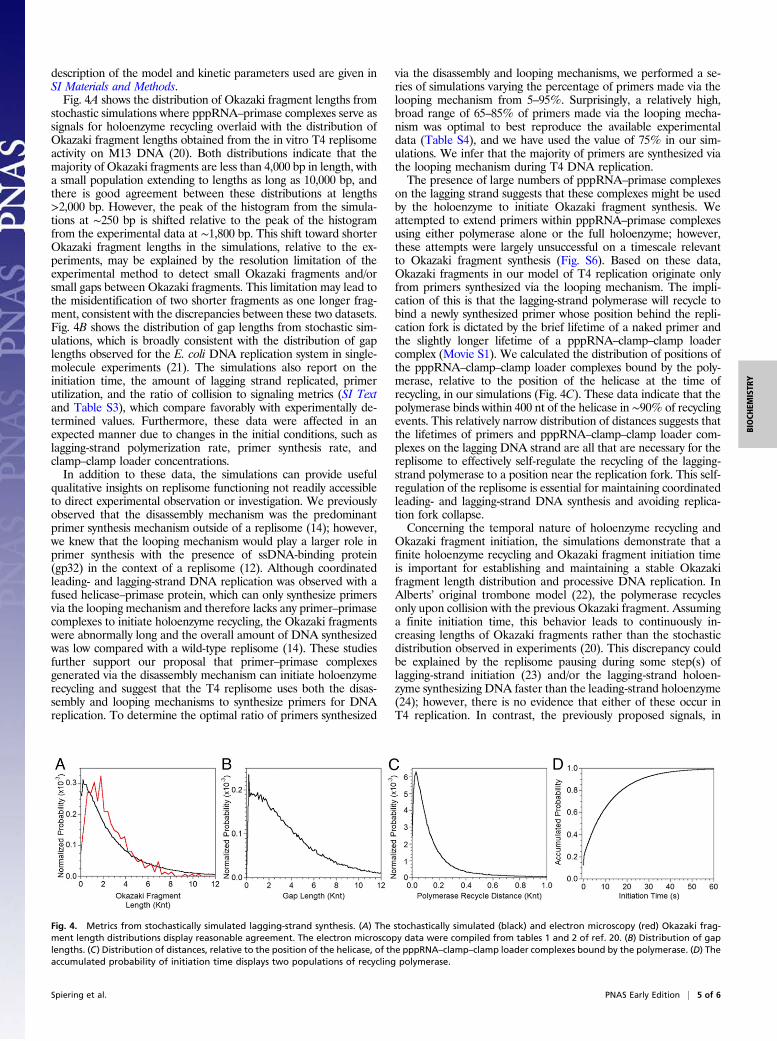
description of the model and kinetic parameters used are given inSI Materials and Methods.Fig. 4A shows the distribution of Okazaki fragment lengths from
stochastic simulations where pppRNA–primase complexes serve assignals for holoenzyme recycling overlaid with the distribution ofOkazaki fragment lengths obtained from the in vitro T4 replisomeactivity on M13 DNA (20). Both distributions indicate that themajority of Okazaki fragments are less than 4,000 bp in length, witha small population extending to lengths as long as 10,000 bp, andthere is good agreement between these distributions at lengths>2,000 bp. However, the peak of the histogram from the simula-tions at ∼250 bp is shifted relative to the peak of the histogramfrom the experimental data at ∼1,800 bp. This shift toward shorterOkazaki fragment lengths in the simulations, relative to the ex-periments, may be explained by the resolution limitation of theexperimental method to detect small Okazaki fragments and/orsmall gaps between Okazaki fragments. This limitation may lead tothe misidentification of two shorter fragments as one longer frag-ment, consistent with the discrepancies between these two datasets.Fig. 4B shows the distribution of gap lengths from stochastic sim-ulations, which is broadly consistent with the distribution of gaplengths observed for the E. coli DNA replication system in single-molecule experiments (21). The simulations also report on theinitiation time, the amount of lagging strand replicated, primerutilization, and the ratio of collision to signaling metrics (SI Textand Table S3), which compare favorably with experimentally de-termined values. Furthermore, these data were affected in anexpected manner due to changes in the initial conditions, such aslagging-strand polymerization rate, primer synthesis rate, andclamp–clamp loader concentrations.In addition to these data, the simulations can provide useful
qualitative insights on replisome functioning not readily accessibleto direct experimental observation or investigation. We previouslyobserved that the disassembly mechanism was the predominantprimer synthesis mechanism outside of a replisome (14); however,we knew that the looping mechanism would play a larger role inprimer synthesis with the presence of ssDNA-binding protein(gp32) in the context of a replisome (12). Although coordinatedleading- and lagging-strand DNA replication was observed with afused helicase–primase protein, which can only synthesize primersvia the looping mechanism and therefore lacks any primer–primasecomplexes to initiate holoenzyme recycling, the Okazaki fragmentswere abnormally long and the overall amount of DNA synthesizedwas low compared with a wild-type replisome (14). These studiesfurther support our proposal that primer–primase complexesgenerated via the disassembly mechanism can initiate holoenzymerecycling and suggest that the T4 replisome uses both the disas-sembly and looping mechanisms to synthesize primers for DNAreplication. To determine the optimal ratio of primers synthesized
via the disassembly and looping mechanisms, we performed a se-ries of simulations varying the percentage of primers made via thelooping mechanism from 5–95%. Surprisingly, a relatively high,broad range of 65–85% of primers made via the looping mecha-nism was optimal to best reproduce the available experimentaldata (Table S4), and we have used the value of 75% in our sim-ulations. We infer that the majority of primers are synthesized viathe looping mechanism during T4 DNA replication.The presence of large numbers of pppRNA–primase complexes
on the lagging strand suggests that these complexes might be usedby the holoenzyme to initiate Okazaki fragment synthesis. Weattempted to extend primers within pppRNA–primase complexesusing either polymerase alone or the full holoenzyme; however,these attempts were largely unsuccessful on a timescale relevantto Okazaki fragment synthesis (Fig. S6). Based on these data,Okazaki fragments in our model of T4 replication originate onlyfrom primers synthesized via the looping mechanism. The impli-cation of this is that the lagging-strand polymerase will recycle tobind a newly synthesized primer whose position behind the repli-cation fork is dictated by the brief lifetime of a naked primer andthe slightly longer lifetime of a pppRNA–clamp–clamp loadercomplex (Movie S1). We calculated the distribution of positions ofthe pppRNA–clamp–clamp loader complexes bound by the poly-merase, relative to the position of the helicase at the time ofrecycling, in our simulations (Fig. 4C). These data indicate that thepolymerase binds within 400 nt of the helicase in ∼90% of recyclingevents. This relatively narrow distribution of distances suggests thatthe lifetimes of primers and pppRNA–clamp–clamp loader com-plexes on the lagging DNA strand are all that are necessary for thereplisome to effectively self-regulate the recycling of the lagging-strand polymerase to a position near the replication fork. This self-regulation of the replisome is essential for maintaining coordinatedleading- and lagging-strand DNA synthesis and avoiding replica-tion fork collapse.Concerning the temporal nature of holoenzyme recycling and
Okazaki fragment initiation, the simulations demonstrate that afinite holoenzyme recycling and Okazaki fragment initiation timeis important for establishing and maintaining a stable Okazakifragment length distribution and processive DNA replication. InAlberts’ original trombone model (22), the polymerase recyclesonly upon collision with the previous Okazaki fragment. Assuminga finite initiation time, this behavior leads to continuously in-creasing lengths of Okazaki fragments rather than the stochasticdistribution observed in experiments (20). This discrepancy couldbe explained by the replisome pausing during some step(s) oflagging-strand initiation (23) and/or the lagging-strand holoen-zyme synthesizing DNA faster than the leading-strand holoenzyme(24); however, there is no evidence that either of these occur inT4 replication. In contrast, the previously proposed signals, in
Fig. 4. Metrics from stochastically simulated lagging-strand synthesis. (A) The stochastically simulated (black) and electron microscopy (red) Okazaki frag-ment length distributions display reasonable agreement. The electron microscopy data were compiled from tables 1 and 2 of ref. 20. (B) Distribution of gaplengths. (C) Distribution of distances, relative to the position of the helicase, of the pppRNA–clamp–clamp loader complexes bound by the polymerase. (D) Theaccumulated probability of initiation time displays two populations of recycling polymerase.
Spiering et al. PNAS Early Edition | 5 of 6
BIOCH
EMISTR
Y

particular those featuring primer synthesis or clamp loading asthe trigger, generally entail very short initiation times. In thesemodels, primer synthesis and clamp loading steps may occur whilethe lagging-strand polymerase is still synthesizing an Okazakifragment; this reduces the wait time for Okazaki fragment initia-tion considerably, assuming that intracomplex rebinding of poly-merase to a new clamp is very fast. If the initiation time is tooshort, the Okazaki fragment length continuously shortens becauseless lagging-strand DNA template is unwound by the helicase eachtime the polymerase is signaled to recycle. The initiation time forthe T7 DNA replication system was observed to follow a single-exponential dependence with a decay constant of 12.0 ± 0.4 s insingle-molecule experiments (2) and was subsequently shown to bedependent on the rate of primer release from the primase activesite (half-life of ∼7 s) (25). However, the initiation time for theE. coli DNA replication system has been estimated to be under0.1 s to sustain the physiological rate of DNA synthesis (7). Fig. 4Ddisplays the accumulated probability of initiation time from oursimulations. Two populations of initiation times can be seen: onewhere ∼25% of recycling polymerase binds to an already availablepppRNA–clamp–clamp loader complex in <1 s, and a secondwhere ∼75% of recycling polymerase must wait a median time of5.9 s for the formation of a pppRNA–clamp–clamp loader com-plex. Our simulations illustrate that the joint probabilities of syn-thesizing a primer, capturing this primer with clamp–clamp loader,and binding polymerase to this complex result in varying initiationtimes that naturally provide a mechanism for successive Okazakifragments to lengthen, whereas the signaling events provide amechanism for them to shorten (Movie S2). The interplay between
these two processes leads to a semirandom distribution of Okazakifragment lengths in our simulations, consistent with that observedexperimentally (20).
ConclusionsTaken together, our experimental data and simulations demon-strate for the T4 bacteriophage DNA replication system thatprimer–primase complexes left on the lagging ssDNA from primersynthesis form blocks for the lagging-strand holoenzyme and thatencounters with a pppRNA–primase complexes correspond to thesignal that causes the lagging-strand polymerase to recycle early,leaving a gap between Okazaki fragments as described in the sig-naling model. Data from previous studies that suggest a long-distance protein trigger for the signaling model are also consis-tent with polymerase recycling being triggered by an encounterwith a pppRNA–primase complex on the lagging strand as sug-gested here. It is possible that pppRNA–primase complexes couldsignal lagging-strand polymerase recycling in other replisomes thatuse an independent primase protein, such as E. coli (26) and eu-karyotes (27).
Materials and MethodsSI Materials and Methods details pppRNA primer and substrate synthesis,protein purification, stopped-flow fluorescence and FRET measurements, ra-dioactive assays, and stochastic modeling of lagging-strand synthesis; SI Textand figures are also available.
ACKNOWLEDGMENTS. This work was supported by National Institutes ofHealth Grant GM13306 (to S.J.B.).
1. Yang J, Nelson SW, Benkovic SJ (2006) The control mechanism for lagging strandpolymerase recycling during bacteriophage T4 DNA replication. Mol Cell 21:153–164.
2. Hamdan SM, Loparo JJ, Takahashi M, Richardson CC, van Oijen AM (2009) Dynamics ofDNA replication loops reveal temporal control of lagging-strand synthesis. Nature 457:336–339.
3. Li X, Marians KJ (2000) Two distinct triggers for cycling of the lagging strand poly-merase at the replication fork. J Biol Chem 275:34757–34765.
4. Hacker KJ, Alberts BM (1994) The rapid dissociation of the T4 DNA polymerase ho-loenzyme when stopped by a DNA hairpin helix. A model for polymerase releasefollowing the termination of each Okazaki fragment. J Biol Chem 269:24221–24228.
5. Carver TE, Jr, Sexton DJ, Benkovic SJ (1997) Dissociation of bacteriophage T4 DNApolymerase and its processivity clamp after completion of Okazaki fragment syn-thesis. Biochemistry 36:14409–14417.
6. Georgescu RE, et al. (2009) Mechanism of polymerase collision release from slidingclamps on the lagging strand. EMBO J 28:2981–2991.
7. Dohrmann PR, Manhart CM, Downey CD, McHenry CS (2011) The rate of polymeraserelease upon filling the gap between Okazaki fragments is inadequate to supportcycling during lagging strand synthesis. J Mol Biol 414:15–27.
8. Wu CA, Zechner EL, Reems JA, McHenry CS, Marians KJ (1992) Coordinated leading-and lagging-strand synthesis at the Escherichia coli DNA replication fork. V. Primaseaction regulates the cycle of Okazaki fragment synthesis. J Biol Chem 267:4074–4083.
9. Tougu K, Marians KJ (1996) The interaction between helicase and primase sets thereplication fork clock. J Biol Chem 271:21398–21405.
10. Corn JE, Pease PJ, Hura GL, Berger JM (2005) Crosstalk between primase subunits canact to regulate primer synthesis in trans. Mol Cell 20:391–401.
11. Yuan Q, McHenry CS (2014) Cycling of the E. coli lagging strand polymerase is trig-gered exclusively by the availability of a new primer at the replication fork. NucleicAcids Res 42:1747–1756.
12. Nelson SW, Kumar R, Benkovic SJ (2008) RNA primer handoff in bacteriophage T4DNA replication: The role of single-stranded DNA-binding protein and polymeraseaccessory proteins. J Biol Chem 283:22838–22846.
13. Kurth I, Georgescu RE, O’Donnell ME (2013) A solution to release twisted DNA duringchromosome replication by coupled DNA polymerases. Nature 496:119–122.
14. Manosas M, Spiering MM, Zhuang Z, Benkovic SJ, Croquette V (2009) Coupling DNAunwinding activity with primer synthesis in the bacteriophage T4 primosome. NatChem Biol 5:904–912.
15. Kaboord BF, Benkovic SJ (1995) Accessory proteins function as matchmakers in theassembly of the T4 DNA polymerase holoenzyme. Curr Biol 5:149–157.
16. Perumal SK, Ren W, Lee TH, Benkovic SJ (2013) How a holoenzyme for DNA repli-cation is formed. Proc Natl Acad Sci USA 110:99–104.
17. Ishmael FT, Trakselis MA, Benkovic SJ (2003) Protein-protein interactions in the bac-teriophage T4 replisome. The leading strand holoenzyme is physically linked to thelagging strand holoenzyme and the primosome. J Biol Chem 278:3145–3152.
18. Kadyrov FA, Drake JW (2001) Conditional coupling of leading-strand and lagging-strandDNA synthesis at bacteriophage T4 replication forks. J Biol Chem 276:29559–29566.
19. Chen D, Yue H, Spiering MM, Benkovic SJ (2013) Insights into Okazaki fragmentsynthesis by the T4 replisome: The fate of lagging-strand holoenzyme componentsand their influence on Okazaki fragment size. J Biol Chem 288:20807–20816.
20. Chastain PD, 2nd, Makhov AM, Nossal NG, Griffith JD (2000) Analysis of the Okazakifragment distributions along single long DNAs replicated by the bacteriophageT4 proteins. Mol Cell 6:803–814.
21. Georgescu RE, Kurth I, O’Donnell ME (2011) Single-molecule studies reveal thefunction of a third polymerase in the replisome. Nat Struct Mol Biol 19:113–116.
22. Alberts BM, et al. (1983) Studies on DNA replication in the bacteriophage T4 in vitrosystem. Cold Spring Harb Symp Quant Biol 47:655–668.
23. Lee JB, et al. (2006) DNA primase acts as a molecular brake in DNA replication. Nature439:621–624.
24. Pandey M, et al. (2009) Coordinating DNA replication by means of priming loop anddifferential synthesis rate. Nature 462:940–943.
25. Hernandez AJ, Lee SJ, Richardson CC (2016) Primer release is the rate-limiting event in lagging-strand synthesis mediated by the T7 replisome. Proc Natl Acad Sci USA 113:5916–5921.
26. Yuzhakov A, Kelman Z, O’Donnell M (1999) Trading places on DNA–A three-pointswitch underlies primer handoff from primase to the replicative DNA polymerase. Cell96:153–163.
27. Yuzhakov A, Kelman Z, Hurwitz J, O’Donnell M (1999) Multiple competition reactions forRPA order the assembly of the DNA polymerase delta holoenzyme. EMBO J 18:6189–6199.
28. Valentine AM, Ishmael FT, Shier VK, Benkovic SJ (2001) A zinc ribbon protein in DNAreplication: Primer synthesis and macromolecular interactions by the bacteriophageT4 primase. Biochemistry 40:15074–15085.
29. Frey MW, Nossal NG, Capson TL, Benkovic SJ (1993) Construction and characterizationof a bacteriophage T4 DNA polymerase deficient in 3′→5′ exonuclease activity. ProcNatl Acad Sci USA 90:2579–2583.
30. Rush J, et al. (1989) The 44P subunit of the T4 DNA polymerase accessory proteincomplex catalyzes ATP hydrolysis. J Biol Chem 264:10943–10953.
31. Nossal NG (1979) DNA replication with bacteriophage T4 proteins. Purification of theproteins encoded by T4 genes 41, 45, 44, and 62 using a complementation assay. J BiolChem 254:6026–6031.
32. Burke RL, Munn M, Barry J, Alberts BM (1985) Purification and properties of thebacteriophage T4 gene 61 RNA priming protein. J Biol Chem 260:1711–1722.
33. Howorka S,Movileanu L, BrahaO, Bayley H (2001) Kinetics of duplex formation for individualDNA strands within a single protein nanopore. Proc Natl Acad Sci USA 98:12996–13001.
34. Sexton DJ, Kaboord BF, Berdis AJ, Carver TE, Benkovic SJ (1998) Dissecting the order ofbacteriophage T4 DNA polymerase holoenzyme assembly. Biochemistry 37:7749–7756.
35. Saito M, Matsumoto M (2008) SIMD-oriented fast mersenne twister: A 128-bit pseu-dorandom number generator. Monte Carlo and Quasi-Monte Carlo Methods 2006,eds Keller A, Heinrich S, Niederreiter H (Springer, Heidelberg), pp 607–622.
36. Humphrey W, Dalke A, Schulten K (1996) VMD: Visual molecular dynamics. J MolGraph 14:33–38, 27–28.
37. Berdis AJ, Benkovic SJ (1996) Role of adenosine 5′-triphosphate hydrolysis in the as-sembly of the bacteriophage T4 DNA replication holoenzyme complex. Biochemistry35:9253–9265.
6 of 6 | www.pnas.org/cgi/doi/10.1073/pnas.1620459114 Spiering et al.























![Nucleosid * DNA polymerase { ΙΙΙ, Ι } * Nuclease { endonuclease, exonuclease [ 5´,3´ exonuclease]} * DNA ligase * Primase](https://img.pdfslide.net/doc/110x75/56649cab5503460f9496ce53/nucleosid-dna-polymerase-nuclease-endonuclease-exonuclease.jpg)





